1.4.- Système olfactif des poissons
Les organismes aquatiques évoluent dans un milieu
où les informations visuelles sont souvent limitées, alors que
les informations chimiques abondent. Cela est d'autant vrai pour la carpe
commune qui vit préférentiellement dans le fond. Les poissons
vivent alors dans un brouillon chimique complexe (Stecey et al.,
2002). Ainsi, ils développent plein d'aptitude pour survivre
grâce aux sens et au système chimiosensoriel (Stecey et al.,
2002 ; Charles & Sorensen 2008). L'olfaction est le sens reliant les
neurones aux axones dans le nerf olfactif (nerfs crâniens primaires)
(Charles & Sorensen, 2008). Elle est entre autre une fonction par
laquelle l'odorat reçoit l'impression des odeurs (Muller,
1845). Le système olfactif joue un rôle important pour la
plupart des animaux parce qu'il leur permet de communiquer efficacement et
d'interpréter les informations fournies par l'environnement
(Séverine, 2001). Comme chez les organismes terrestres,
l'olfaction et la gustation chez les animaux aquatiques sont habituellement
identifiées comme les systèmes chimiosensoriels. Leurs
rôles sont souvent confondus vu que leurs fonctions sont semblables.
Cependant, il existe bien des différences de structure et de fonction
entre elles (Stecey et al., 2002 ; Charles & Sorensen, 2008).

L'existence d'un sens olfactif chez les animaux aquatiques n'a
été reconnue que tardivement (Rider et Bauchaut, 1984).
Duméril, 1807 pensait que l'olfaction était liée
à des substances gazeuses. Weber cité par Rider et
Bauchaut, 1984 quant à lui considérait que seul le
goût a un sens chimique général chez les animaux
aquatiques. Ce n'est qu'au début du 20ème
siècle que l'existence d'un vrai sens olfactif a été
reconnu et accepté chez le poisson (Herrick, 1908 ; Parker, 1910).
Cependant, la controverse ne cessa qu'en 1929 quand Schering, 1929
cité par Rider et Bauchaut, (1984) mettait en
évidence le rôle de l'olfaction dans le retour des saumons aux
frayères. Ainsi, d'autres études ont mis en
évidence l'existence de la sensibilité chimique (olfaction et
goût) chez la plupart des poissons (Rider et Bauchaut, 1984).
Situé au-dessus de la bouche, les sacs nasaux de l'odorat jouent
plusieurs rôles important dans divers types de comportement, notamment
dans la recherche des aliments, les phénomènes de homing
chez les migrateurs, la reproduction, la défense contre les
prédateurs etc. (François et Saint-André,
2015).

« Conditionnement de carpes communes (Cyprinus
carpio) à la détection olfactive des composés
aromatiques de
décomposition de viande de porc par
imprégnation des oeufs et des larves »,
Ulg/Septembre 2015 3

Comme chez les vertébrés, le système
olfactif des poissons contient 3 composantes neuroanatomiques principales
(Stecey et al., 2002).

L'Epithélium olfactif
appelé aussi système olfactif
périphérique est entre autre constitué des ouvertures
(cavités nasales) de la face dorsale de la tête du poisson en
avant des yeux (Laberge et Hara, 2001). Il comprend 3 principales
classes de cellules neuronales réceptrices (Fig.1) (Hamdani and
Døving, 2006, Stecey et al., 2002): les neurones ciliés qui
possèdent de longues dendrites ; les neurones microvillositaires avec de
courtes dendrites et les neurones de la crypte qui sont sphériques et ne
possédant ni cils ni microvillosités (Hansen et al., 2003,,
Hansen and Finger, 2000, Zeiske et al, 2003 , Ichikawa et Ueda, 1977,
Thommesen, 1983 , Kermen et al., 2013). Autres que les neurones
sensoriels, il existe aussi dans l'épithélium olfactif d'autres
types de neurones non-sensoriels (Bazaes et al., 201 , Kermen et al.,
2013).
Le Bulbe olfactif est la structure du cerveau
des vertébrés qui reçoit la grande majorité des
neurones sensoriels olfactifs à travers les nerfs olfactifs. Ainsi,
comprendre les mécanismes neurophysiologiques qui régissent le
traitement des odeurs perçues dans le bulbe olfactif nécessite
une compréhension profonde de ses propriétés
physiologiques (Kermen et al., 2013).
Il est structurellement organisé en 4 couches
concentriques de cellules et comprend plus de 20.000 neurones (Friedrich et
al., 2009). De superficielle à profonde nous trouvons:
? une couche de fibres olfactives primaires formée par
les axones des neurones sensoriels olfactifs; (Sato et al., 2007)
? une couche glomérulaire contenant environ 140 modules
sphériques de glomérules (Braubach et al, 2012);
? la couche externe de cellules, constituée de cellules
mitrales riches en soma cellulaire (Fuller and Byrd, 2005, Fuller et al,
2006);
? la couche interne de cellules, contenant les corps
cellulaires de différents inter neurones, (Edwards et Michel, 2002,
Bundschuh et al, 2012). Les principales cellules du bulbe olfactif des
poissons sont mitrales glutamatergiques (Edwards et Michel, 2002).
Des expériences montrent que chez les poissons, les
acides aminés activent la partie ventrale du bulbe olfactif et les sels
biliaires la partie médiane (Hamdani & Doving, 2007).
D'autres expériences l'ont confirmé par des enregistrements
d'électrodes de surface chez les salmonidés (Thommesen, 1978
, Doving et al, 1980). Donc, il y a une structuration spatiale des
récepteurs olfactifs neuronaux en fonction de l'odeur perçue. Ce
qui traduira sans doute les différents types de comportement liés
à chaque type d'odeur dans le bulbe olfactif. Donc, contrairement
à ce qui se trouve dans l'épithélium olfactif, dans le
bulbe olfactif chaque classe de substances odorantes est
représentée par une zone glomérulaire distincte
(Kermen et al., 2013 , Hamdani & Doving, 2007).
Le pédoncule olfactif est
formé par la projection des neurones du second degré du bulbe
olfactif. Il se divise en 2 types en fonction de leur position anatomique : le
pédoncule olfactif médian (POM) et le pédoncule
olfactif latéral (POL) (Laberge et Hara, 2001 , Hamdani &
Doving, 2007). Le pédoncule olfactif médian (POM)
est divisé en 2 faisceaux distincts ; un faisceau latéral ou
dorso-latéral (DL) et un faisceau médial ou medio
ventral (MV) qui induisent respectivement, les réactions
comportementales de reproduction et des réactions d'alarmes. Le
pédoncule olfactif latéral (POL) quant à lui
induit des réactions comportementales liées à la nutrition
(Laberge et Hara, 2001 , Daghfous et al., 2012).

« Conditionnement de carpes communes (Cyprinus
carpio) à la détection olfactive des composés
aromatiques de
décomposition de viande de porc par
imprégnation des oeufs et des larves », Ulg/Septembre 2015
4
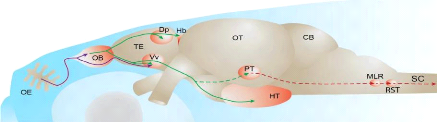

OE: épithélium olfactif ;
OB: bulbe olfactif ; TE:
télencéphale ; Dp: partie
postérieure dorsal du télencéphale ; Vv:
Noyau ventral du télencéphale ; HT:
hypothalamus
Figure 1: Système olfactif des poisons
(Kermen et al., 2013) 1.5.- La
décomposition d'un corps en milieu aquatique
En milieu aquatique, un corps se décompose
différemment qu'en milieu terrestre. Ainsi, la décomposition se
différent en temps et en étapes de décomposition en
fonction du milieu. Plusieurs facteurs conditionnent le temps et les type de
décomposition qu'un corps subit en milieu aquatique. Citons entre autre
: la température du milieu, la présence des microorganismes et
leur diversité spécifique, le teneur en sel de l'eau
(salée ou douce), la position du corps dans le milieu (en surface ou au
fond), l'état initial du corps (malade, blessé...). Ainsi,
distingue-t-on 5 grandes phases dans la décomposition d'un corps en
milieu aquatique qui sont : la phase de cadavre frais ; la phase de
putréfaction, la phase de fermentation ; la phase de desséchement
et la phase de squelette (Anderson & Hobischak, 2004 ; Ayers,
2010). Ainsi, dans un corps en décomposition notamment en phase de
putréfaction et fermentation, les différentes réactions
chimiques de décomposition (protéines du corps), sous l'effet des
microorganismes du milieu quand les conditions sont favorables
(température, salinité,...), libèrent des composés
chimiques servant souvent de moyens pour découvrir et vérifier
l'état de décomposition (phase) d'un corps. Parmi les produits,
nous trouvons des composés volatiles notamment des composés
phénoliques, aromatiques et surtout des polyamines biogènes.
| 

