Chapitre III
Optimisation par
algorithmes
génétiques
3.1- Introduction.
Au fil des années, de nombreuses recherches ont
été effectuées et de nombreuses approches ont
été proposées pour effectuer le meilleur réglage et
coordination des PSSs.
Dans les systèmes de puissance, nous avons besoin
d'installer plusieurs PSSs. Traditionnellement, les paramètres de ces
PSSs sont réglés séquentiellement et
séparément (Abe et al., 1983; Arredondo, 1997; Yee
et al., 2004). Dans les stratégies de réglage
séquentiel, basées sur les méthodes de réglage
présentées dans le chapitre précédent, les PSSs
sont alors conçus pour amortir les oscillations des modes, mode par
mode. Par exemple, la méthode linéaire séquentielle
(Linear Sequential Method, LSM) est basée sur les résidus. Dans
cette méthode, les PSSs sont réglés d'une manière
progressive (Yee et al., 2004) :
- on calcule tout d'abord les résidus de la fonction de
transfert du système en boucle
ouverte. Puis un PSS est ajouté et réglé en
utilisant les informations de ces résidus.
- ensuite, un second PSS est introduit et réglé en
se basant sur les informations des
résidus du système avec le premier PSS en place.
- ce processus continue jusqu'à ce que le système
atteigne des caractéristiques de stabilité satisfaisantes.
Une autre méthode séquentielle basée sur
le placement des pôles du système est connue sous le nom :
positionnement séquentiel des valeurs propres (Sequential Eigenvalue
Assignment) (Fleming et al., 1981). Dans cette méthode, les
paramètres des PSSs sont représentés comme des variables
dans les équations caractéristiques du système. Ces
équations sont résolues itérativement jusqu'à ce
que les valeurs propres se trouvent placées aux endroits
déterminés préalablement. La solution représente
les paramètres des PSSs recherchés. Comme les "bonnes" valeurs
propres spécifiées sont souvent choisies par expertise, les
valeurs déterminées des paramètres des PSSs peuvent
être situées en dehors de gammes correspondantes de fonctionnement
(Hong et al., 1999). De ce fait, la méthode doit être
appliquée de façon itérative jusqu'à obtenir des
paramètres adéquats.
Bien que les méthodes de réglage
séquentiel soient simples et aient donné
généralement des résultats satisfaisants pour
l'amortissement des oscillations, ces méthodes ne peuvent assurer une
optimisation globale des PSSs et, ainsi, la stabilité globale du
système. Les aspects négatifs de ces méthodes sont
liés aux hypothèses restrictives et à la nature intuitive
du processus de réglage (Fleming et al., 1981). En outre, les
interactions dynamiques entre les différents modes ont une influence
significative sur le réglage des PSSs : le réglage d'un PSS pour
stabiliser un mode peut ainsi produire des effets opposés sur les autres
modes.
La littérature montre qu'à la place des
méthodes de réglage séquentiel, une optimisation
simultanée des PSSs du système entier peut être
utilisée (CIGRE, 1999). Les méthodes d'optimisation
simultanée peuvent permettre d'atteindre plusieurs objectifs :
- une stabilité globale.
- un bon fonctionnement du système de puissance avec un
amortissement satisfaisant pour diverses conditions de fonctionnement et
configurations du système.
- une minimisation des interactions antagonistes possibles entre
les PSSs.
Grâce à la rapidité du
développement de l'informatique, l'utilisation des outils et des
algorithmes d'optimisation devient de plus en plus aisée et efficace.
Cette révolution informatique a permis une ouverture vers des
méthodes synthétiques telles les méthodes d'optimisation
stochastiques. A la différence des méthodes analytiques dans
lesquelles on
cherche à trouver une solution théorique exacte
ou une bonne approximation numérique, les méthodes stochastiques
constituent une approche originale dans laquelle on cherche à trouver
des solutions satisfaisant aux mieux différents critères souvent
contradictoires. Elles peuvent aussi synthétiser des solutions nouvelles
et originales sans idées préconçues.
La résolution d'un problème d'optimisation
consiste à explorer un espace de recherche afin d'optimiser (maximiser
ou minimiser) une fonction donnée (une fonction objectif) sous certaines
contraintes. La complexité du problème, en taille ou en
structure, relative à l'espace de recherche et à la fonction
à optimiser conduit à développer diverses méthodes.
Ces méthodes peuvent être regroupées en deux
catégories : les méthodes déterministes (classiques) et
les méthodes non-déterministes (stochastiques).
1- Les méthodes déterministes
:
Qualifiées de classiques (telles la méthode du
gradient, les méthodes énumératives,...), elles
n'utilisent aucun concept stochastique. Une méthode déterministe
utilise donc toujours le même cheminement pour arriver à la
solution, et nous pouvons donc déterminer à l'avance les
étapes de la recherche. Ces méthodes sont limitées par
leur "faible" espace de recherche. Elles requièrent des
hypothèses sur la fonction objectif à optimiser, telles que
continuité et dérivabilité de la fonction en tout point du
domaine des solutions. Elles consistent généralement à se
focaliser sur un point unique de l'espace de recherche en le
déplaçant au cours de temps dans le but de trouver un extremum.
Ces méthodes sont généralement efficaces lorsque
l'évaluation de la fonction est très rapide ou lorsque sa forme
est connue à priori. Mais, lorsque la dimension du
problème ou l'espace de recherche est grand, ces méthodes peuvent
:
- avoir des temps de calcul déraisonnables ou
- boucler et revenir sans cesse au même point.
Enfin, un grand nombre de fonctions à optimiser ne sont
pas dérivables et souvent même pas continues. Par
conséquent, ces méthodes restent limitées à des
problèmes très précis.
2- Les méthodes non-déterministes
:
Ces méthodes, qualifiées de stochastiques, sont
bien adaptées aux problèmes importants et complexes (tels les
problèmes discrets) ou même à des problèmes ayant
des multifonctions objectif. Dans cette famille de méthodes, on parle
couramment d'heuristique et de métaheuristiques.
Une méthode heuristique est une
méthode d'optimisation adaptée à un problème
particulier. Parmi ces méthodes, figurent la méthode de
Monte-Carlo, le recuit simulé, la recherche tabou, la méthode de
descente, ... . Ces diverses méthodes travaillent toujours à
partir d'une solution initiale en essayant de l'améliorer au maximum en
fonction des contraintes du problème étudié. Ainsi, elles
ont toutes un point commun : à savoir leurs itérations reposent
sur l'amélioration d'une solution unique. Leurs évolutions sont
ainsi basées sur la notion de parcours : l'évolution vers une
solution optimale se fait en testant successivement une solution voisine de la
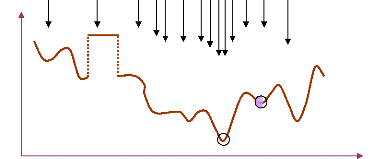
solution courante. La figure (38) illustre une représentation simple
d'une méthode d'optimisation à parcours (heuristique).
Une méthode métaheuristique
consiste en une stratégie de choix pouvant piloter une ou plusieurs
heuristiques de manière abstraite, sans faire appel à un
problème spécifique. Les méthodes métaheuristiques
sont ainsi des méthodes à population de solutions : à
chaque itération, elles manipulent un ensemble de solutions en
parallèle (Osman et al., 1996). Ces
méthodes sont aussi considérées comme des
méthodes d'optimisation globale : elles visent la
détermination de l'optimum global de la fonction objectif du
problème, en évitant le "piégeage" dans l'un de ses optima
locaux. Elles comblent ainsi le handicap des méthodes classiques et des
méthodes heuristiques en conduisant la recherche vers l'optimum global.
En outre, ces méthodes font usage de l'expérience
accumulée durant la recherche de l'optimum, pour mieux guider la suite
du processus de recherche. Elles permettent ainsi d'explorer et d'exploiter
l'espace de recherche plus efficacement. Une représentation
schématique simple des métaheuristiques est donnée
à la figure (39).
Dans cette représentation, les métaheuristiques
(MH) tentent de trouver l'optimum global (OG) d'un problème
d'optimisation (f(x)) difficile (avec par exemple des
discontinuités (DC)), sans être piégé par les optima
locaux (OL).

Figure 38. Représentation
simplifiée d'une approche heuristique.
MH
f(x)
DC
OL
OG
x
Figure 39. Représentation
simplifiée d'une approche métaheuristique.
Les algorithmes évolutionnaires regroupent une
multitude de métaheuristiques différentes. Cependant, tous ont un
point commun : ils sont basés sur les principes de la théorie
évolutionnaire. Ils simulent l'évolution naturelle des structures
individuelles afin de trouver une solution optimale. Dans chaque
génération, une nouvelle approximation de
solution optimale se produit par des processus de
sélection des individus selon leurs performances dans le domaine du
problème. Les individus sélectionnés vont être
reproduits en utilisant des opérateurs empruntés aux
génétiques naturelles. Ces processus mènent à
l'évolution de la population des individus les mieux adaptés
à leur environnement (Pohlheim, 2005).
Ces méthodes reçoivent de plus en plus
d'intérêt en raison de leurs capacités potentielles
à résoudre des problèmes complexes. Sous le concept
d'algorithmes évolutionnaires, on trouve plusieurs méthodes tels
les algorithmes génétiques, la programmation
évolutionnaire, les stratégies d'évolution et la
programmation génétique. Les étapes communes de ces
méthodes peuvent être résumées comme suit
(Bäck, 1996) :
- une initialisation aléatoire d'une population
d'individus (des points de l'espace de recherche).
- une coopération et une compétition parmi les
individus de la population concernée selon les principes Darwinistes et
génétiques de la survie du "meilleur" (sélection,
croisement et mutation).
- un nombre d'itérations (générations)
faisant évoluer la solution (l'individu) courante et finissant par
atteindre la solution optimale.
Une caractéristique importante pour les algorithmes
évolutionnaires est l'absence d'information sur le gradient pour
élaborer les solutions. Cette propriété donne aux
algorithmes évolutionnaires une flexibilité satisfaisante leur
permettant d'être utilisés aisément dans une grande
variété de problèmes tels les problèmes fortement
non-linéaires, les problèmes non-continus, ... . De plus, les
algorithmes évolutionnaires travaillent sur un ensemble de points (une
population) et non pas sur un seul point ; cela augmente la probabilité
de trouver la solution optimale. Comme ces algorithmes utilisent les principes
stochastiques, ils ne demandent aucune structure particulière pour le
problème à optimiser.
Grâce à leurs nombreux avantages, les algorithmes
évolutionnaires sont appliqués avec beaucoup de succès
dans le domaine industriel et l'ingénierie. De nombreux problèmes
liés aux systèmes de puissance sont résolus par les
algorithmes évolutionnaires telles les répartitions
économiques, la prévision de charges, la planification de
génération, la coordination des contrôleurs, les
études de sûreté des systèmes de puissance,... .
Les algorithmes génétiques, forment une des
principales classes des algorithmes évolutionnaires ; ils ont
suscité beaucoup d'enthousiasme depuis plusieurs années. Leur
efficacité pour produire des solutions de qualité dans un grand
nombre de problème d'optimisation est maintenant bien établie.
Ils représentent les méthodes d'optimisations les plus
utilisées.
3.2- Les Algorithmes Génétiques (AGs).
3.2.1- Introduction.
Comme nous l'avons déjà indiqué, les
algorithmes génétiques sont des méthodes de recherche
stochastiques de solutions basées sur des mécanismes s'inspirant
de processus naturels (sélection) et des phénomènes
génétiques (croisement et mutation). Les travaux de scientifiques
du XIXe siècle (Lamarck, Darwin, Mendel,...) et du
XXe siècle (Watson et Crick) ont permis de mettre en
évidence les principaux mécanismes à l'oeuvre au cours de
l'évolution des populations. Ces recherches ont débouché
sur l'idée que l'évolution repose
d'une part sur l'intervention du hasard au cours des
croisements et des mutations, et d'autre part, sur un processus de
sélection des individus les plus aptes à survivre dans un
environnement donné. C'est précisément cette notion de
hasard guidé par la sélection qui est à la base des
algorithmes génétiques.
Dans les années soixante et soixante-dix, le professeur
J. Holland à l'université de Michigan (Holland, 1975) a
développé, sur la base des travaux précédents, les
principes fondamentaux des algorithmes génétiques, son objectif
étant de concevoir des systèmes artificiels possédant des
propriétés similaires aux systèmes naturels. L'ouvrage de
Goldberg (Goldberg, 1989) a permis de mieux faire connaître les
algorithmes génétiques et il a marqué le début de
l'intérêt actuel pour ces techniques.
La théorie des algorithmes génétiques
utilise un vocabulaire similaire à celui de la génétique
naturelle. Cependant, les processus naturels auxquels elle fait
référence sont beaucoup plus complexes que les algorithmes
génétiques.
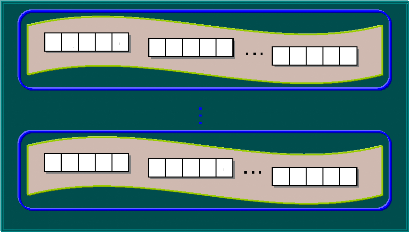
On définit ainsi un individu dans une population.
L'individu est représenté par un ou plusieurs chromosomes
constitués de gènes qui contiennent les caractères
héréditaires de l'individu. Les principes de sélection, de
croisement, de mutation utilisés s'appuient sur les processus naturels
du même nom (Barnier et al., 1999).
Pour un problème d'optimisation donné, un
individu représente un point de l'espace de recherche c.-à-d. une
solution potentielle. On lui associe la valeur du critère à
optimiser qui définit sa performance. Pour progresser d'une population
d'individus à une nouvelle population, on utilise
séquentiellement les processus de sélection, de croisement et de
mutation. La sélection a pour but de favoriser les meilleurs
éléments de la population pour le critère
considéré. Le croisement et la mutation ont pour but d'assurer
l'exploration et l'exploitation de l'espace de recherche. Par
conséquent, les individus les moins adaptés vont être
exclus et les meilleurs vont survire pour construire la nouvelle population.
Chaque itération de l'algorithme génétique
représente une génération. Au fur et à mesure de la
progression d'une génération à l'autre, la solution
trouvée va converger vers la solution optimale à atteindre
(Negnevitsky, 2002).
Généralement, nous pouvons dire qu'un algorithme
génétique dans sa forme générale nécessite
de préciser les points suivants :
- le codage des solutions et la génération d'une
population initiale.
- la fonction de performance pour calculer l'adaptation de chaque
individu de la population.
- le croisement des individus d'une population pour obtenir la
population de la génération suivante.
- l'opération de mutation des individus d'une population
afin d'éviter une convergence prématurée.
- les paramètres de réglage : taille de la
population, probabilités de croisement et de mutation, critère
d'arrêt.
Ces points vont être discutés en détail dans
la suite de ce chapitre, mais nous allons tout d'abord définir les
expressions utilisées dans la théorie des algorithmes
génétiques.
Dans un problème d'optimisation à n
variables, nous faisons correspondre un gène
à chaque variable cherchée. Chaque gène est
représenté par une chaîne de caractères choisis dans
un alphabet fini (souvent binaire). Les gènes s'enchaînent
ensemble "bout à bout" pour
construire un chromosome, chaque
chromosome représentant une solution potentielle sous une forme
codée. Ces chromosomes constituent les briques de base contenant les
caractéristiques héréditaires des individus. Un chromosome
(ou plusieurs) forme un individu qui
représente à son tour une solution potentielle dans l'espace de
recherche correspondant du problème. Etant donné que les AGs
travaillent sur un ensemble de points de l'espace de recherche, nous appelons
l'ensemble des points choisis (à savoir les individus) une
population. Au fur et à mesure des
générations (itérations), une population des
individus mieux adaptés va être créée.
La figure (40) représente les relations entre les
expressions génétiques d'un AG en codage binaire.

Population
0
1
1
1
Gène (1)
Gène (1)
Chromosome
Chromosome
0
0
0
1
0
0
0
1
Individu (Nind)
1
1
Gène (2)
Gène (2)
Individu (1)
1
1
0
0
0
1
0
1
Gène (Ngens)
Gène (Ngens)
0
1
0
1
1
1
1
1
Figure 40. Les niveaux d'organisation des
éléments d'un AG. 3.2.2- Codage et
initialisation.
La première étape de la construction d'un AG est
le choix du type de codage des paramètres du problème. La
façon de coder les solutions potentielles est un facteur
déterminant dans le succès d'un AG. Ainsi, plusieurs types de
codage sont possibles dans la littérature, tels les codages binaires,
Gray, réel,... . Le codage le plus populaire dans la
représentation d'un AG est le codage binaire {0, 1 } ; les solutions
sont codées selon des chaînes de bits de longueur fixe
(Chipperfield et al., 1994). La plupart des théories
liées aux AGs étaient élaborées en se basant sur le
concept de codage binaire proposé par J. Holland et son groupe (Holland,
1975). Les opérateurs de l'AG, croisement et mutation, sont en effet
plus faciles à mettre en oeuvre avec ce type de codage. En outre, le
codage binaire représente la méthode la plus facile et la mieux
adaptée de coder des éléments qu'ils soient réels,
entiers, booléens, ... (Mitchell, 1996). On parle dans ce cas de
génotype en ce qui concerne la représentation binaire
d'un individu et de phénotype pour ce qui est de sa valeur
réelle correspondante dans l'espace de recherche.
Une fois le choix du type de codage déterminé,
une population initiale doit être créée pour le
départ de l'AG. La population initiale a pour but de donner naissance
à des générations successives, mutées et
hybridées à partir de leurs parents. Le choix de la population
initiale influence fortement la rapidité et l'efficacité de l'AG.
Si la position de l'optimum dans l'espace de recherche est totalement inconnue,
il est naturel de générer aléatoirement des individus en
faisant des tirages uniformes dans chacun des domaines associés aux
composantes de l'espace de recherche, en veillant évidemment à ce
que les individus produits respectent les contraintes. Si par contre, des
informations à priori sur le problème sont disponibles,
il parait naturel de générer les individus dans un sous-domaine
particulier afin d'accélérer la convergence. Habituellement,
cette population initiale est générée d'une manière
aléatoire et directement dans sa représentation codée.
Par exemple pour créer une population binaire
de Nind individus dans lesquels chaque chromosome
(individu) est représenté par Ngen
gènes, il suffit simplement d'effectuer Nind ×
Ngen tirages de nombres aléatoires distribués
uniformément sur l'ensemble {0, 1 }, (Chipperfield et al., 1994).
La seconde étape dans la construction de l'AG est le
calcul de la performance (fitness) de chaque individu faisant partie
de la population. Pour ce faire nous devons en premier lieu décoder les
chromosomes (précisément les gènes de chaque chromosome)
en les convertissant en leurs valeurs réelles (numériques)
(Negnevitsky, 2002).
Considérons un problème d'optimisation de
n variables à optimiser, où l'espace de recherche de chaque
variable xj se trouve entre une limite inférieure
xmin,j et une limite supérieure xmax,j :
D = [xmin, j , x max,j]
avec j = 1, 2,..., n. On associe des points du domaine D
à
Sj chaînes de bits (Sj gènes) de
longueur lS. Ainsi, chaque chaîne Sj sera donc
composée de lS
éléments binaires : Sj =
(si)j ; i = 1, 2,..., lS, où si ? {0,1}. Toute
chaîne binaire Sj peut être
décodée en une valeur réelle xj en
utilisant les deux règles suivantes (Negnevitsky, 2002) :
- dans la première, nous convertissons les valeurs
binaires de chaque gène, en valeurs de base décimale selon la
règle suivante :
lS
à ( ) 2 (117)
( )
l i
S -
x j s i j
= ·
?=
i 1
- ensuite, nous calculons les valeurs réelles
correspondantes appartenant à l'espace de recherche donné, par la
règle suivante :
x x
max, min,
j j
-
fd x x x x
( ) à
= = + · lS (118)
j j j
min, -
2 1
Par exemple, supposons que nous cherchons à maximiser
une fonction f en fonction d'une variable réelle x
appartenant à l'espace de recherche D = [-1 , 2]. Soit
S une chaîne binaire représentant une solution possible
avec une longueur lS = 22 :
S = 1,1,0,0,1,0,1,0,0,0,0,1,1,0,1,1,1,1,1,1 ,1,1 }
{
En appliquant les relations (117) et (118), nous obtenons
respectivement la valeur entière de x (en base décimale)
et sa valeur réelle :
l S =22
à 2 1 2 1 2 0 2 1 2 3311359
= ? · = × + × + × + + × =
l i
- 21 20 19 0
S
x si L
i = 1

|
x = -
|
2 ( 1)
- -
1 3311359 22
+ · = ? -
x 1 . 368469 [ 1,2]
2 1
-
|
Après avoir générer la population
initiale, nous devons attribuer à chacun des individus une note qui
correspond à son adaptation au problème. Cette adaptation est la
mesure de la fonction de performance associée à la fonction
objectif du problème. Les notions de ces deux fonctions font l'objet du
paragraphe suivant.
3.2.3- Fonctions objectif et de performance. 3.2.3.1-
Fonction objectif.
A l'inverse des méthodes déterministes
d'optimisation, les algorithmes génétiques ne requièrent
pas d'hypothèse particulière sur la régularité de
la fonction à optimiser (objectif). Ainsi, les algorithmes
génétiques n'utilisent pas les dérivées successives
de la fonction objectif, ce qui rend leurs domaines d'application très
vaste. Aucune hypothèse sur la continuité n'est également
requise. Le peu d'hypothèses requises permet de traiter des
problèmes très complexes. La fonction objectif peut être le
résultat d'une simulation ou d'un modèle mathématique.
Généralement, la fonction objectif, d'un
problème quelconque d'optimisation à K contraintes (dit
problème contraint), peut être formulée comme suit :
Optimisation {Fobj(X) : X?
Dk} (119)
Avec :
? =
k k K
: 1, ,
L
?R R
et H D
: a
k
DK =
?
?? ? ? ?
? b k
?
??
(120)
?
?
??
Fobj(X) : la fonction objectif du
problème.
Hk : la fonction de contrainte.
D : l'ensemble des solutions potentielles du
problème.
Dk : l'espace des solutions réalisables
(c.-à-d. l'ensemble des solutions potentielles en respectant les
contraintes).
La fonction objectif peut être formulée d'un
ensemble de fonctions de dimension n. Elle est donnée d'une
façon générale comme suit : Fobj(X) =
{f1 ,f2,L, f i ,L, fn
}.
Lorsque i = 1 la fonction est dite monoobjectif,
autrement elle est multiobjectif.
Contrairement à l'optimisation monoobjectif, la
solution d'un problème d'optimisation multiobjectif est rarement unique.
Elle est constituée de différentes solutions, représentant
l'ensemble des meilleurs compromis vis-à-vis des
différents objectifs du problème.
Les méthodes existantes pour formuler une fonction
multiobjectif sont diverses. Nous allons expliquer brièvement ces
méthodes qui peuvent être classées en trois grandes
familles (Régnier, 2003) :
1- Les méthodes à
priori. Dans ces méthodes, on définit,
à priori, le compromis désiré entre les objectifs
avant de lancer la méthode de résolution. Nous trouvons dans
cette famille la plupart des méthodes scalaires telles la méthode
de pondération, la méthode des objectifs bornes, la
méthode du critère globale,... . Le principe de ces
méthodes se base sur la transformation du problème multiobjectif
en un problème monoobjectif, en pondérant l'ensemble des
fonctions objectif initiales.
2- Les méthodes
progressives. Dans ces méthodes, on affine le choix
du compromis entre les objectifs au cours de l'optimisation. Contrairement aux
méthodes à priori, ces méthodes ont
l'inconvénient de monopoliser l'attention de l'utilisateur tout au long
du processus d'optimisation. La méthode lexicographique, par exemple,
consiste à minimiser séquentiellement les différents
objectifs du problème. L'ordre de minimisation est fixé en
fonction des résultats séquentiels obtenus. La méthode
progresse alors par transformations successives du problème
d'optimisation.
3- Les méthodes a
posteriori. Dans ces méthodes, il n'est plus
nécessaire de modéliser les préférences entre les
objectifs avant l'optimisation. Ces méthodes se contentent de produire
un ensemble de solutions plutôt qu'un unique compromis. Nous pouvons
ensuite choisir a posteriori une solution du compromis.
3.2.3.2- Fonction de performance.
Chaque chromosome apporte une solution potentielle au
problème à optimiser. Néanmoins, ces solutions n'ont pas
toutes le même degré de pertinence. C'est à la fonction de
performance (fitness) de mesurer cette efficacité pour
permettre à l'AG de faire évoluer la population dans un sens
bénéfique en cherchant la solution meilleure. Autrement dit, la
fonction de performance, fp(X), doit pouvoir attribuer
à chaque individu un indicateur représentant sa pertinence pour
le problème que nous cherchons à résoudre. La performance
sera donc donnée par une fonction à valeurs positives
réelles. La construction d'une fonction de performance appropriée
est très importante pour obtenir un bon fonctionnement de l'AG.
Dans le cas du codage binaire, la fonction de performance doit
affecter une valeur positive au codage binaire correspondant
(phénotype) à chaque chaîne binaire
(génotype). Ainsi, elle permet de déterminer
l'efficacité de la solution présentée par le
génotype pour résoudre le problème.
La fonction de performance fp(X) est
généralement dérivée de la fonction objectif
Fobj(X) du problème. Elle est généralement
donnée par la relation suivante :
fp(X) = g(Fobj(X))
(121)
Où : g : représente la transformation de
la fonction objectif en performance relative.
Les AGs sont naturellement organisés pour
résoudre les problèmes de maximisation (c. -à- d. trouver
les valeurs positives maximales de la fonction objectif). Ainsi, pour les
problèmes de maximisation, la fonction de performance peut être
considérée comme la fonction objectif même. Le but de l'AG
est alors simplement de trouver la chaîne binaire qui maximise la
fonction de performance :
max(fp(X)) = min(Fobj(X))
(122)
Dans le cas de problèmes de minimisation, le
problème doit être modifié de sorte qu'il soit
équivalent à celui de maximisation. Ainsi, il nous faudra
modifier la fonction objectif de telle sorte que la fonction de performance
soit maximale :
max(fp(X)) = -min(Fobj(X))
(123)
max(fp(X)) = 1/min(Fobj(X))
(124)
max(fp(X)) = max(C-
Fobj(X)) ; (C : une grande constante positive)
(125)
Une fois que la performance de chaque individu dans la
population actuelle est évaluée, les mécanismes
évolutionnaires entrent en jeu pour explorer et exploiter le plus
largement possible l'espace de recherche et faire ainsi évoluer la
population de manière progressive. Les opérateurs de l'AG
cherchent à imiter ces mécanismes.
3.2.4- Sélection.
Pour déterminer les individus devant participer au
résultat optimal de l'AG, un opérateur sélection doit
être appliqué. Cet opérateur détermine la
capacité de chaque individu à persister dans la population et
à se reproduire. En règle générale, la
probabilité de survie d'un individu sera directement reliée
à sa performance relative au sein de la population. Cela traduit bien
l'idée de la sélection naturelle Darwiniste : les gènes
les plus performants ont la probabilité la plus élevée de
se reproduire dans la population, tandis que ceux qui ont une performance
relative plus faible auront tendance à disparaître.
Néanmoins, cette approche doit être appréhendée avec
précaution, puisqu'on souhaite transférer aux
générations suivantes les meilleures caractéristiques
génétiques et pas seulement les meilleurs individus.
En effet, un individu peut avoir certains gènes
présents dans la solution optimale tout en présentant une
performance médiocre : son élimination serait donc nuisible.
Cependant, les individus ayant une bonne performance sont toujours plus
susceptibles de posséder les bons gènes. Ainsi, une
caractéristique importante d'un opérateur de sélection
peut être définie en termes de pression sélective
exercée sur la population. Cette notion doit être prise en compte
d'une manière équilibrée. Si la pression sélective
est trop forte, cela revient à "écraser" toute forme de
diversité possible et la convergence de l'algorithme risque d'être
prématurée en conduisant vers un optimum local. Si l'on passe
à l'autre extrême, une pression sélective trop faible,
l'algorithme tend à se rapprocher d'une simple recherche
aléatoire avec une progression lente de la population (Duvigneau,
2006).
Il existe plusieurs méthodes pour représenter la
sélection. La méthode la plus couramment utilisée
proposée par Goldberg (Goldberg, 1989) est connue sous le nom de
sélection par roulette biaisée (Roulette Wheel). D'autres
méthodes de sélection sont aussi apparues dans la
littérature, la plus connue étant celle du tournoi (Tournament
Selection).
3.2.4.1- Sélection par la roulette
biaisée.
Dans cette méthode, chaque chromosome est copié
dans la nouvelle population avec une probabilité proportionnelle
à sa performance relative : on parle ainsi de sélection
proportionnelle à la performance. Dans un codage binaire, le principe de
cette méthode consiste à associer à chaque individu
Xi (d'une population de taille Nind) une
probabilité Rp,i proportionnelle à sa performance
fp(Xi). Cette probabilité (dite la fonction de performance
normalisée) peut être ainsi calculée comme le
taux de la performance du ième individu
pondéré par la somme des performances de toute la population :
R P , i
fp ( ) (126)
Nind
?=
i 1
X i
fp ( )
X i
On doit donc effectuer autant de tirages aléatoires
qu'il y a d'individus dans la population. Chaque individu est alors reproduit
avec la probabilité Rp ; certains individus
(notés les bons) auront plus de chance d'être reproduits, les
autres (notés mauvais) seront éliminés.
Dans la pratique, nous associons à chaque individu un
secteur (ou un segment) dont la surface (la longueur) est proportionnelle
à sa performance. Ces éléments sont ensuite
concaténés sur une base normalisée entre 0 et 1. Enfin,
nous reproduisons le principe de tirage aléatoire utilisé dans
les roulettes de casinos, d'où le nom de ce mécanisme de
sélection, avec une caractéristique linéaire
(sélection proportionnelle à la performance). Nous tirons alors
un nombre aléatoire, de distribution uniforme entre 0 et 1, et nous
déterminons l'individu sélectionné. Ainsi, les grands
secteurs (c.-à-d. les bons individus) auront plus de chance d'être
choisis que les petits secteurs. Les individus ayant une forte performance sont
donc privilégiés au détriment des individus moins forts,
tout en gardant la notion de tirage aléatoire. Ainsi, avec cette
méthode un même individu pourra être
sélectionné plusieurs fois. Mais une certaine diversité
est cependant maintenue, car même les individus les moins performants
conservent une chance d'être choisis. Pour garder la même taille de
la population initiale dans la population suivante, on doit lancer la roulette
autant de fois qu'il y a d'individus de la population.
Considérons un problème simple de maximisation dont
la fonction objectif est donnée par la forme suivante :
Fobj = 4· x · (1- x)
L'espace de recherche de la variable x est D = [0,1].
Nous traitons ce problème en décodant les points
de l'espace de recherche en chaînes binaires de longueur lS = 8,
et en initialisant une population de départ de 4 individus
(Nind = 4).
Pour appliquer la méthode de la roulette biaisée,
nous calculons en premier lieu, pour chaque individu, la fonction de
performance fp(Xi) et la probabilité correspondante
Rp,i.
Le tableau suivant résume le résultat pour chaque
élément :
|
N°
individu
|
Chromosome
Si
|
Valeurs
décodées Xi
(individus)
|
Performance
fp(Xi) =Fobj
=
4·x·(1-x)
|
Probabilité
Rp,i
= fp(Xi) / ?(
fp(Xi))
|
Intervalles
de
probabilité
|
|
1
|
1
|
0 1
|
1 1
|
0 1
|
0
|
0.7294
|
0.7895
|
0.29
|
[0 , 0.29]
|
|
2
|
1
|
1 0
|
1 1
|
1 1
|
0
|
0.8706
|
0.4507
|
0.18
|
[0.29 ,
|
0.47]
|
|
3
|
0
|
0 0
|
1 1
|
0 1
|
0
|
0.1020
|
0.3665
|
0.15
|
[0.47 ,
|
0.62]
|
|
4
|
0
|
1 1
|
0 1
|
1 0
|
0
|
0.423 5
|
0.9766
|
0.38
|
[0.62
|
, 1]
|
|
|
|
|
|
|
|
? fp = 2.5831
|
|
|
|
Tableau 1. Résultats de
l'évaluation des individus dans la population initiale.
Ensuite, nous associons à chaque intervalle de
probabilité un secteur équivalent de la roulette (c.-à-d.
un individu), comme le montre la figure (41) :
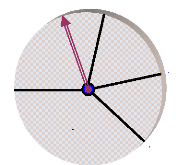
0.29
Rp1 Rp2
0.47
0
1
Rp3
Rp4
0.62
Figure 41. Sélection par roulette
biaisée.
Enfin, nous effectuons le tirage d'un nombre aléatoire
dans l'intervalle [0,1] et nous reproduisons ce tirage 4 fois. Nous
sélectionnons ainsi les individus en positionnant chaque nombre obtenu
par tirage dans le secteur équivalent de la roulette. Le tableau (2)
donne la valeur obtenue pour chaque tirage et les individus
sélectionnés correspondants.
|
N°
tirage
|
Valeur obtenue
|
Individus
sélectionnés
|
|
Chromosomes
équivalents
|
|
|
1
|
0.43
|
X2
|
Xá 1
|
=
|
1
|
1 0
|
1 1
|
1 1
|
0
|
|
2
|
0.89
|
X4
|
Xá 2
|
=
|
0
|
1 1
|
0 1
|
1 0
|
0
|
|
3
|
0.18
|
X1
|
Xá 3
|
=
|
1
|
0 1
|
1 1
|
0 1
|
0
|
|
4
|
0.75
|
X4
|
Xá 4
|
=
|
0
|
1 1
|
0 1
|
1 0
|
0
|
Tableau 2. Résultat de
sélection.
Nous trouvons que l'individu X3 est
éliminé de la population tandis que l'individu X4 est
reproduit deux fois.
3.2.4.2- Sélection par tournoi.
La méthode de la roulette, comme les autres
méthodes de sélection proportionnelle à la performance,
exige principalement deux étapes pour créer la population
intermédiaire. Premièrement, nous calculons la performance
moyenne de la population, ensuite nous calculons la probabilité de
sélection pour chaque individu. La méthode du tournoi est plus
directe, puisqu'elle ne demande pas le calcul de la performance moyenne de la
population (Mitchell, 1996).
Cette méthode consiste à simuler
Nind tournois, un individu étant
sélectionné à chaque fois. Un échantillon de
np individus (2 minimum) est prélevé au
hasard à chaque tournoi. Le
participant dans l'échantillon ayant la performance la
plus élevée sera alors adopté pour accéder à
la population intermédiaire. Etant donné que le choix des
échantillons des individus se fait aléatoirement, il est donc
tout à fait possible que certains individus participent à
plusieurs tournois. Il est aussi possible qu'un de ces individus gagne
plusieurs fois et il sera par conséquent copié autant de fois
dans la population intermédiaire. La pression sélective de cette
méthode est directement reliée à la taille de
l'échantillon (np). Plus la taille
np est grande, plus la pression est grande (Duvigneau,
2006). Cette méthode est caractérisée par une pression
sélective plus forte que celle de la méthode de la roulette
biaisée : pour qu'un individu peu performant puisse être
sélectionné, il faut que tous ses adversaires soient moins bons
que lui. Ainsi, cette méthode doit être privilégiée
dans le cas d'une population de grande taille.
Dans le cas où le nombre de participants dans chaque
tournoi est égal à deux (np = 2), la
sélection est dite par « tournoi binaire ». La
sélection d'un individu Xsélect,K par tournoi
binaire peut être faite de la façon suivante :
|
XsélectK
,
|
=
|
? ??
??
|
X Si fp X
( ) >
i i
X Sinon
j
|
fp X
( )
j
|
(127)
|
Où, K={1,2,L,Nind}.
i, j sont des nombres aléatoires
non-égaux, i, j ? {1,2,L, Nin d }.
3.2.4.3- Conclusion.
La méthode de la roulette reste la méthode de
sélection la plus utilisée car elle est relativement efficace.
Cependant, pour certains problèmes, elle est "un peu trop
aléatoire". C'est pour cela que pour certains problèmes, la
méthode du tournoi est plus préférée. Cette
dernière étant "moins aléatoire" que la roulette, elle
permet ainsi une plus grande fiabilité.
Disposant finalement d'une population d'individus
non-homogène, la diversité de la population doit être
entretenue au cours des générations afin de parcourir le plus
largement possible l'espace de recherche. Pour ce faire, les individus
sélectionnés (dits les parents) sont alors introduits dans le
bassin de reproduction (Mating Pool) où ils seront de nouveaux choisis
aléatoirement pour voir leurs chromosomes subir des transformations par
les opérateurs génétiques. Ces derniers sont les
opérateurs de croisement et de mutation (dits les opérateurs de
diversification et d'intensification du milieu).
3.2.5- Croisement.
Dans les AGs, le croisement est considéré comme
le principal opérateur pour produire des nouveaux chromosomes. Comme son
homologue dans la nature, le croisement produit de nouveaux individus en leur
transférant quelques parties de la matière
génétique de leurs parents. L'objectif du croisement est donc
d'enrichir la diversité de la population en manipulant la structure des
chromosomes (Chipperfield et al., 1994). Initialement, le croisement
associé au codage par chaînes de bits (codage binaire) est le
croisement à découpage de chromosomes.
Ainsi, dans le codage binaire, les individus, qui
résultent de la sélection, sont groupés de manière
aléatoire par paire définissant ainsi les parents. Ensuite,
chaque couple peut subir un croisement avec une probabilité
Pc donnée. Cette étape peut être
effectuée comme suit
Pour chaque couple, un nombre aléatoire P est
tiré dans l'intervalle [0,1] et comparé ensuite avec la
probabilité de croisement Pc :
- si P > Pc, le couple ne subit
pas de croissement et un clonage de chromosome aura
lieu. Les deux enfants produits sont ainsi une copie exacte de
leurs parents.
- si P < Pc, le croisement a
lieu et un échange des parties des chromosomes des parents
va produire deux enfants par couple de parents.
Après avoir tiré les couples qui vont être
"croisés", l'opérateur de croisement peut donc être
appliqué. Plusieurs types de croisement sont présentés
dans la littérature, tels : le croisement seul point, le croisement
multipoints, le croisement uniforme,... .
3.2.5.1- Croisement seul point.
Dans ce type de croisement, un point de croisement est choisi
aléatoirement pour le couple ; la position de ce point M est
définie par:
M? {1,2,L,l S -1} (128)
lS : la longueur de chromosome (nombre de bits dans le
chromosome).
Les deux parents seront ainsi divisés en deux segments
; tête et queue. Le segment tête du premier parent est
combiné avec le segment queue du deuxième parent : on obtient
ainsi le premier enfant. La combinaison entre le segment tête de
deuxième parent et le segment queue de premier parent produit le
deuxième enfant.
Pour mieux expliciter ce type de croisement, nous allons
l'appliquer à l'exemple présenté dans le paragraphe de la
sélection.
Considérons la population de quatre individus
déterminée par l'opération de sélection, nous
pouvons la diviser aléatoirement en deux couples qui peuvent être
par exemple : [(Xá1, Xá4), (Xá2,
Xá3)].
Considérons ensuite une probabilité de
croisement Pc = 50% (nous avons choisi une faible
probabilité de croisement étant donné la "petite" taille
de la population considérée). Cela signifie que nous pouvons
tirer seulement un seul couple au hasard et lui appliquer le croisement
chromosomique. Supposons que le hasard ait désigné le
deuxième couple (Xá2, Xá3) pour croisement, il
nous faut maintenant déterminer la position du point M de
croisement dans les parents. Cette position peut être tirée au
hasard ou choisie préalablement. Afin de rendre l'exemple plus parlant,
nous décidons d'appliquer ce croisement sur le milieu (M = 4)
du chromosome, comme le montre la figure (42).
Ainsi, l'application de l'opération de croisement engendre
deux nouveaux individus (les deux enfants Xâ 2 et
Xâ 3).
Le croisement en un seul point a l'avantage d'être
simple et facile à appliquer. De plus, ce type de croisement donne de
bons résultats dans des applications où certaines informations
importantes sur le problème sont déjà connues. Enfin, pour
des problèmes d'optimisation en temps réel ou des
problèmes ayant un grand nombre de variables, cette méthode peut
donner une convergence rapide vers une solution optimale.
Avant Après
Croissement Croissement
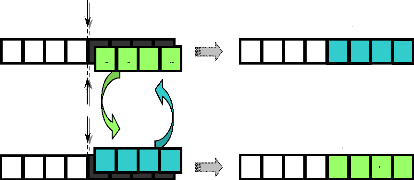
0
1
Parent Xá 2
Parent Xá 3
0
1
1
1
0
1
1
1
0
1
0
1
0
0
Enfant Xâ 2
0
1
1
0
1
0
1
0
Enfant Xâ 3
1
0
1
1
1
1
0
0

Figure 42. Croisement seul point.
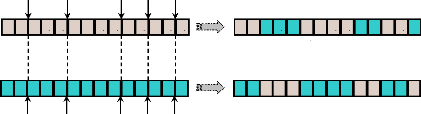
3.2.5.2- Croisement multipoints.
A la différence du croisement seul point, ce type de
croisement s'applique en plusieurs points (m points) et chaque
chromosome sera ainsi découpé en (m+ 1) segments. La
position de chaque point, Mi, se détermine
aléatoirement, avec :
Mi { l S } i m
? L - = L
1,2, , 1 ; 1,2, ,(129)
lS : la longueur de chromosome.
m : le nombre de points de croisement donné (Pour
m = 1, on retourne au cas du croisement seul point).
Ensuite, les bits entre deux points de croisement successifs
du premier parent vont être échangés avec les bits
correspondants du deuxième parent. La figure (43) montre un croisement
en cinq points (m = 5), les points de croisement Mi
étant {2, 5, 9, 11, 13 } :
M1 = 2 M2 = 5 M3 = 9 M4 = 11 M5 = 13
1110111
0
0
1
1
1
0
1
Enfant 1
1
01000001 1 0 0 0 0
Enfant 2
1 0 1 0 1 0 0 01 1 1 0 01
1 1 1 0 0 1 1 0 0 1 0 1 00
Parent 2
Parent 1
Figure 43. Croisement multipoints (m =
5).
La nature complexe du croisement multipoints permet d'obtenir
une bonne exploration de l'espace de recherche au détriment d'une
convergence rapide vers la bonne solution comme le ferait la première
méthode. Ainsi, cette méthode de croisement est beaucoup plus
robuste.
3.2.5.3- Croisement uniforme.
Dans le croisement multipoints, chaque segment du chromosome,
compris entre deux points de croisement successifs, peut contenir un ou
plusieurs bits. Le croisement uniforme généralise ce
schéma pour lier à chaque bit un point de croisement. Pour ce
faire, un chromosome masque, de même longueur que le chromosome des
parents, est créé : la valeur de ses bits étant
aléatoire. La parité de chaque bit dans le masque
détermine successivement le parent qui va donner ses bits à
l'enfant (Chipperfield et al., 1994). L'enfant
généré va copier le bit de son premier parent si le bit
correspondant de masque est `1', ou de son deuxième parent si le bit
correspondant de masque est `0'. Ce schéma ne fait générer
qu'un seul enfant. Pour générer le deuxième, un autre
chromosome masque est nécessaire. Les bits de ce dernier sont
créés par complémentation des bits du premier masque.
L'explication de cette méthode est décrite par l'exemple suivant
:

0
1
0
1
1
0
1
Parent 1
Enfant 1

1
1
1
1
1
1
0
0
0
1
0
1
1

0
1
1
1
Masque 1
Masque 2
|
1
|
0
|
1
|
0
|
1
|
1
|
0
|
0
|
1
|
0
|
0
|
1
|
|
0
|
0
|
1
|
1
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
1
|
Parent 2 Enfant 2
Figure 44. Croisement uniforme.
La nature complexe du croisement uniforme, comme pour le
croisement multipoints, améliore encore l'exploration de l'espace de
recherche. Avec le croisement uniforme, basé sur le transfert de
matière génétique d'un parent à un enfant par bit
de chromosome, on obtient un algorithme génétique beaucoup moins
dépendant du schéma de codage. Enfin, cette méthode peut
être plus précise dans l'identification d'optima globaux.
3.2.6- Mutation.
A la suite des opérateurs de sélection et de
croisement, on mime à nouveau un phénomène biologique,
celui de la mutation. Au niveau biologique, une mutation est une modification
de l'information génétique par dégradation ou substitution
locale de paire de base : ceci permet de produire une nouvelle structure
génétique. L'opérateur de mutation dans le cas des AGs
possède la propriété d'ergodicité du parcours de
l'espace de recherche : cette propriété indique que l'AG sera
susceptible d'atteindre tous les points de l'espace, sans pour autant les
parcourir tous dans le processus de résolution
(Bontemps, 1995). La séquence des opérations de sélection
et de croisement peut mener l'AG à "stagner" dans un ensemble de
solutions identiques. Dans de telles conditions, tous les chromosomes
deviennent identiques et, ainsi, la performance moyenne de la population ne
s'améliore plus. Dans ce cas, la mutation aide l'AG à
éviter la perte de diversité génétique et, par
conséquent, elle garantit que l'AG ne va pas être bloqué
dans un optimum local (Negnevitsky, 2002).
Le principe de la mutation consiste à modifier, avec
une probabilité Pm faible, certains bits des
chromosomes. Nous tirons tout d'abord pour chaque bit un nombre
aléatoire P dans l'intervalle [0,1]. Puis, nous le comparons
avec une probabilité de mutation Pm donnée
:
- si P > Pm, le bit ne subira pas
aucune modification.
- si P < Pm, la mutation est
appliquée au bit correspondant.
Ainsi, le bit choisi pour muter sera remplacé par une
valeur aléatoire, souvent proche de la valeur initiale. Dans le cas du
codage binaire, cette mutation s'effectue simplement en remplaçant le
bit `0' par `1' et vice versa.
Pour continuer la présentation de l'opérateur de
mutation, nous reprenons l'exemple précédent (paragraphe
croisement) et nous appliquons la mutation sur la population résultante
du croisement.
Rappelons les individus résultants :
Xâ 1 = 1 1 0 1 1 1 1 0
Xâ 2 = 0 1 1 0 1 0 1 0
Xâ 3 = 1 0 1 1 1 1 0 0
Xâ 4 = 0 1 1 0 1 1 0 0
Nous proposons arbitrairement une probabilité de
mutation Pm = 0.01. Pour déterminer les bits qui
subiront une mutation, nous tirons un nombre aléatoire compris entre 0
et 1 pour chaque bit et nous le comparons avec Pm. La
figure suivante montre les bits choisis pour la mutation et l'application de
cet opérateur :
Xfl 3

0
1
1
0
1
1
0
0
Xfl 4
Xã 3

0
1
1
0
1
1
0
0
Xã 4


0
1
1
1
1
1
1
0
0
1
1
1
1
1
0
1
0
1

1
0
Xfl 1
1
0
0
1
1
0
1
0
Xã 1
0
0
1
0
1
1
1
0

1
1
1
0
1
1
0
0
1
Xfl 2
1
0
1
1
1
1
1
0
Xã 2
Figure 45. Exemple de mutation.
Les individus résultants de la mutation,
(Xã1, Xã2,
Xã3, Xã4), vont former la
nouvelle population qui va remplacer la population initiale pour la
deuxième génération.
3.2.7- Création de la génération
suivante et critères d'arrêt.
Pour réaliser une boucle d'une itération de
l'AG, ce dernier doit d'abord regrouper les individus survivants après
mutation dans une nouvelle population. Ensuite, l'AG va calculer la performance
pour chaque nouvel individu. Enfin, si un critère d'arrêt de
l'algorithme n'est pas encore atteint, la nouvelle population doit alors
remplacer la population actuelle et une nouvelle boucle sera ainsi
lancée.
Puisque les AGs sont des méthodes de recherches
stochastiques, il est difficile de spécifier de façon rigoureuse
des critères de convergence. Par exemple, la performance d'une
population peut rester stable pour un certain nombre de
générations avant qu'un individu supérieur puisse
apparaître. Ainsi, l'application d'un critère d'arrêt
devient une vraie problématique. Une pratique commune est
d'arrêter l'AG après certain nombre de générations
et d'examiner ensuite la qualité de la solution trouvée par
rapport à la définition du problème. Un nombre typique de
générations peut aller de 50 jusqu'à 500
générations (Negnevitsky, 2002).
Par ailleurs, d'autres critères peuvent être
appliqués pour déterminer l'arrêt de l'AG tels que :
- l'amélioration de la solution ne dépasse plus un
certain seuil
- la fonction objectif du problème atteint une valeur
donnée
- le temps de calcul atteint une valeur
prédéterminée.
Nous terminons l'exemple que nous avons commencé de
traiter par le calcul de la performance de chaque nouvel individu :
|
N°
individu
|
Chromosome
Sãi
|
Valeurs
décodées
Xãi
|
Performance fp(Xãi)
=Fobj
|
|
1
|
0
|
1 1
|
1
|
1 1
|
1
|
0
|
0.4941
|
0.9999
|
|
2
|
0
|
0 1
|
0
|
1 1
|
1
|
0
|
0.1804
|
0.5914
|
|
3
|
1
|
0 1
|
1
|
1 1
|
1
|
0
|
0.745 1
|
0.7597
|
|
4
|
0
|
1 1
|
0
|
1 1
|
0
|
0
|
0.4235
|
0.9766
|
|
|
|
|
|
|
|
|
? fp = 3.3276
|
Tableau 3. Résultats de
l'évaluation des individus dans la nouvelle population.
En comparant le tableau (1) de l'évaluation de la
population initiale et le tableau (3) de
l'évaluation de la nouvelle population, nous pouvons tirer
les remarques suivantes :
- le meilleur individu (Xãj)
dans la nouvelle population est un individu nouveau (issu
d'une opération de mutation).
- ce nouvel individu présente une performance
supérieure à celle du meilleur individu de la population initiale
(fp(Xãj) = 0.9999 contre
fp(Xj) = 0.9766).
- l'individu X4 reste toujours présent dans la
nouvelle population (renommé Xã4).
- la nouvelle performance totale (et donc la performance moyenne)
est supérieures à la valeur de départ (?
fp(Xãi) = 3.3276 contre
? fp(Xi) = 2.5831).
- l'individu (Xã1),
associé à la performance maximale
fp(Xã1), peut être ainsi
considéré comme la solution optimale du problème.
Etant donné que le problème traité est
très simple, l'AG a pu atteindre une solution optimale dés la
première itération.
3.2.8- Compromis exploration et exploitation.
Généralement, la recherche d'une solution
optimale dans un espace complexe implique souvent un compromis entre deux
objectifs apparent contradictoires : l'exploration et l'exploitation.
L'exploration a pour objectif une recherche large des
régions nouvelles et inconnues de l'espace de recherche en
récoltant de l'information sur le problème à optimiser.
L'exploitation se propose d'utiliser l'information acquise aux points
déjà explorés pour définir et parcourir ainsi les
zones les plus intéressantes de l'espace de recherche. Une recherche
purement aléatoire (comme le fait la méthode de Monte-Carlo) est
bonne pour l'exploration mais mauvaise pour l'exploitation alors que la
recherche dans le voisinage (comme le fait la méthode de la descente)
est une bonne méthode d'exploitation mais une mauvaise méthode
d'exploration. La littérature montre que les algorithmes
génétiques peuvent réaliser un compromis
équilibré et raisonnable entre exploration et exploitation. Mais
il est important de bien doser l'usage de ces deux ingrédients afin que
l'exploration puisse identifier rapidement des régions de l'espace de
recherche qui contiennent des solutions de bonnes qualités, sans perdre
trop de temps à exploiter des régions moins prometteuses. Pour ce
faire, il faut ainsi ajuster les paramètres de réglage des
opérateurs génétiques, sachant que le croisement favorise
plus l'exploration tandis que la mutation favorise plus l'exploitation.
3.2.9- Paramètres de réglage de
l'AG.
Il y a principalement trois paramètres de base pour le
"fonctionnement" d'un AG : - le nombre d'individus dans la population
Nind (dit la taille de la population).
- la probabilité de croisement Pc.
- la probabilité de mutation Pm.
La réussite et la rapidité d'un AG
dépendent fortement des valeurs choisies pour ces paramètres. Le
bon réglage de ces paramètres est un problème parfois
délicat (Mitchell, 1996).
Nous discutons ci-dessous l'influence de chaque paramètre
et la gamme de valeurs qu'il peut prendre :
1- la taille de la population
Nind
Ce paramètre doit être judicieusement
réglé en fonction de la taille du problème.
Généralement, nous pouvons dire que si la taille de la population
est :
- trop faible, l'AG peut converger trop rapidement vers de
mauvaises solutions. - trop grande, le temps de calcul de l'AG peut
s'avérer très important.
En règle générale, plus la taille de la
population est grande, plus le nombre de solutions potentielles
évaluées est élevé. La littérature montre
que les meilleures valeurs de taille de population sont comprises entre 50 et
100 individus (Mitchell, 1996).
2- la probabilité de croisement
Pc
Comme nous l'avons dit, la probabilité de croisement
joue un rôle très important dans l'exploration de l'espace de
recherche du problème. En générale, plus la
probabilité de croisement est élevée, plus il y aura de
nouvelles structures apparaissant dans la nouvelle population. Ainsi, si la
probabilité de croisement est :
- trop élevée, les "bonnes" structures
apportées par la sélection risquent d'être détruites
trop vite.
- trop faible, la recherche de la solution optimale risque de
stagner.
Le taux habituel de la probabilité de croisement est
choisi entre 0.7 et 0.95.
3- la probabilité de mutation
Pm
La mutation, comme nous l'avons vu, est un opérateur
secondaire, mais elle reste très importante pour l'AG. Elle a pour
objectifs l'introduction de diversité dans la population et la meilleure
exploitation de l'espace de recherche. Ainsi, si la probabilité de
mutation est :
- trop élevé, la mutation rend la recherche
très aléatoire.
- trop faible, la recherche risque de stagner.
Le taux habituel de la probabilité de mutation est choisi
entre 0.001 et 0.05. 3.3- Conclusion.
Les algorithmes génétiques sont des
méthodes métaheuristiques d'optimisation globale basées
sur des concepts de génétique et de sélection naturelle.
Le composant principal des AGs est le gène qui se compose d'une
chaîne de caractères (souvent binaire). Les gènes
s'enchaînent et forment les chromosomes. Ces derniers forment les
individus dans l'espace de recherche. Ainsi, les AGs travaillent sur une
population d'individus, où chacun de ces derniers représente une
solution possible pour le problème donné. Dans chaque
itération de l'AG, la performance de chaque individu de la population
courante est calculée. Les opérateurs de
génétiques, sélection, croisement et mutation, sont
appliqués successivement pour créer une nouvelle population
jusqu'à l'approche rigoureuse de la solution optimale.
Les algorithmes génétiques offrent plusieurs
avantages :
- Ils ne demandent pas d'informations à priori ou
des propriétés particulières de la fonction objectif du
problème.
- Leurs performances par rapport aux algorithmes classiques sont
bien remarquées lorsque par exemple les espaces de recherches sont
importants.
- Outre leur facilité de programmation et de manipulation,
ils sont facilement adaptables à tout type de problème
d'optimisation.
- Ils peuvent être utilisés avec profit pour traiter
des problèmes n'étant pas optimisables efficacement par des
approches purement mathématiquement.
Par ailleurs, les algorithmes génétiques
présentent certaines limites :
- Le temps de calcul est souvent important : ils
nécessitent de nombreux calculs, en particulier au niveau de la fonction
objectif.
- Les paramètres de réglage (telles la taille de
la population, la probabilité de croisement, ...) sont parfois
difficiles à déterminer. Or le succès de
l'évolution en dépend et plusieurs essais sont donc
nécessaires.
- Ils ne garantissent pas toujours la découverte de
l'optimum global en un temps fini. En effet, lorsqu'une population
évolue, il se peut que certains individus occupant à un instant
une place importante au sein de cette population deviennent majoritaires. A ce
moment, il se peut que la population converge vers cet individu en
s'écartant ainsi d'individus plus intéressants et en
s'éloignant de l'individu vers lequel on devrait converger.
Nous pouvons conclure que l'efficacité des AGs
dépend d'un compromis entre deux objectifs contradictoires : la
rapidité et la précision. La rapidité est souvent
mesurée en nombre d'évaluations de la fonction objectif. Cette
dernière représente la plupart du temps la partie la plus
"gourmande" en temps de calcul. La précision se rapporte à la
distance entre l'optimum trouvé par l'AG et l'optimum réel, du
point de vue de la solution ou de la valeur. Bien souvent, un AG rapide est peu
précis, et inversement.
Dans le chapitre suivant, nous utilisons les principales
caractéristiques des AGs développés dans ce chapitre pour
une optimisation globale des PSSs.
| 

