II. 1. Formes isolées : les
tachyzoïtes
Les tachyzoïtes sont généralement
intracellulaires et peuvent être libérés lors de
l'éclatement des cellules contenant le parasite.5

Figure 1 : Eléments morphologiques typiques
de Toxoplasma, les tachyzoïtes5
La forme est en croissant de longueur 5 à 8
ìm et de largeur 3 à 5 ìm et d'extrémité
effilée. Ces éléments apparaissent au microscope à
contraste de phase avec un cytoplasme homogène et réfringent. Le
noyau est très net et il occupe une position centrale. Des
phénomènes de glissements aident à la mobilité des
tachyzoïtes mais ils ne possèdent pas d'organes locomoteurs. 5
II. 2. Formes groupées
Les formes groupées sont divisées en
trois parties : les pseudokystes, les kystes et les oocystes.
- Les pseudokystes
Les pseudokystes sont responsables de la forme
aiguë de la maladie. Ils sont en effet intracellulaires. Ils se logent
dans une vacuole parasitophore de la cellule hôte qui constitue la paroi
du pseudokyste. Ils peuvent mesurer entre 15 et 30 ìm. Leur
présence caractérise la phase proliférative de
l'infection.8

Figure 2 : Schéma et vue microscopique des
pseudokystes5
Les pseudokystes renferment 100 à 200
tachyzoïtes qui n'occupent pas la totalité de la cellule hôte
dont le noyau demeure net. Ils sont colorables par la fuchsine. Ces
pseudokystes n'ont qu'une durée momentanée, et libèrent
des tachyzoïtes qui envahissent d'autres cellules.?
- Les kystes
Egalement intracellulaires, ils sont
différés des pseudokystes par l'occupation quasi-totale de la
cellule parasitée dont le noyau est déformé, aplati et
réduit à une lame qui occupe la périphérie. Les
kystes sont plus volumineux que les pseudokystes. La forme est principalement
subspherique. Les kystes mesurent 60 à 100 ìm et causent une
déformation de la cellule hôte. ?

Figure 3 : Schéma et vue microscopique des
kystes 5
Les kystes contiennent des milliers de
bradyzoïtes en croissant dont le noyau occupe une position excentrique
à l'extrémité arrondie. La présence des kystes
correspond à la phase chronique de la maladie. La rupture de la cellule
qui les porte libère des kystes enveloppés. Les kystes se
transforment en pseudokystes quand l'immunité de l'hôte est
rompue. Ils sont le plus souvent localises dans le système
réticulo-histiocytaire (SRH). 5
- Les oocystes
C'est la forme parasitaire rencontrée dans
les cellules épithéliales de l'hôte définitif. Le
zygote est issu de la fécondation d'un gamète femelle par un
gamète mâle et qui reste enkyste dans la coque ovulaire. Un
éclatement des cellules épithéliales hôtes aident
à l'élimination des oocystes dans le milieu extérieur et
le mélange aux excréments. Leurs formes est subspheriques de
longueur de 10ìm et de largeur de 12ìm. Ils subissent la
sporogonie en milieu extérieur. La sporulation des oocystes renferme
deux sporocystes contenant chacun 4 sporozoïtes en virgule mesurant
7ìm de longueur et 1.5ìm. 5
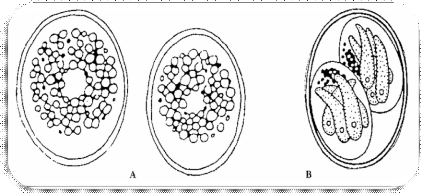
Figure 4 : Schéma des oocystes immatures (A)
et oocystes sporulés (B) de la T. gondii5
L'hôte définitif, le chat en
particulier, la sporogonie à la production d'un oocyste suite à
la fécondation des gamètes femelles qui a lieu dans l'estomac de
cet animal. Ce dernier est la seule espèce animale domestique à
héberger la reproduction sexuée du toxoplasme. L'oocyste est la
forme la plus résistante dans le milieu extérieur lors de
l'élimination avec les selles du chat.²
L'oocyste contient 8 sporozoïtes groupes en 2
sporocystes accoles dans le tube digestif du chat. Cet oocyste
représente l'aboutissement du cycle sexué chez le chat et
constitue la forme infectieuse metacyclique ou forme contaminant pour
l'homme.²
III. Etude biologique
III. 1. Modalités de la reproduction
Il existe une reproduction sexuée, trouvée chez
les chats et certains félidés sauvages ; c'est-à-dire
chez les hôtes définitifs, et reproduction asexuée chez
tous les hôtes, définitifs et intermédiaires.
III. 1. 1. La reproduction sexuée de T.
gondii
Elle ne peut avoir lieu que chez un hôte
définitif.
a) La gamètogonie
Au lieu de continuer à subir la multiplication
asexuée, certains schizontes vont se différencier en gamontes
puis en gamètes. Le moment de survenue de la gamètogonie pourrait
être influence par différents facteurs qui rendent les conditions
environnementales plus mauvaises : une surpopulation des formes
asexuées, le début de la mise en place de la réponse
immune ou bien l'initiation d'un traitement antitoxoplasmique. Elle se
déroule généralement dans l'iléon.°
La micro-gamétogenèse est à l' origine
des gamètes males (appelés microgamète). La
différenciation a lieu dans la vacuole parasitophore des
entérocytes de l'hôte. Il présente un appareil perforateur
apical, un volumineux noyau associéà une grande
mitochondrie.°
La macro-gamétogenèse est à
l' origine des gamètes femelles. Ses formes ovoïdes subspheriques
possèdent un noyau central, plusieurs micropores qui favorisent la
pénétration et donc la fécondation par un
microgamète.°
La pénétration du microgamète
dans la cellule hôte puis dabs le macrogamonte est permise par le
fonctionnement des flagelles et du perforatorium. La fusion de deux
noyaux haploïdes aboutit à la formation d'un zygote diploïde
et est suivie de la formation d'une paroi à cinq couches constituant
l'endokyste et l'ectokyste. L'oeuf est donc enkysté dans la cellule
hôte, au sein de l'épithélium intestinal : il porte
alors le nom d'oocyste et il est alors immature.°
b) La sporogonie
Elle aboutit, à partir d'un zygote
diploïde, à la formation de huit sporozoites haploïdes. La
sporulation est exogène (en milieu extérieur) et ne se
réalise que si les conditions environnementales sont
favorables :°
- Une humidité relative de 30% minimum avec un optimum
de 90% environ ;
- Une température de 10 à 30°C ;
- Une teneur en oxygène atmosphérique suffisante
pour la réalisation du métabolisme aérobie important.
Dans les conditions optimales, la sporulation
s'achève en un à cinq jours.
III. 1. 2. La reproduction
asexuée
a) Lors du cycle entero-épithélial
C'est un type de division qui ne se déroule qu'au cours
du cycle coccidien, dans les entérocytes et à l'intérieur
de la vacuole parasitophore : le noyau de la cellule
mèreappeléeschizonte se divise en nombreux noyaux-fils separes.
Cette multiplication entraine une dilatation de la vacuole parasitophore, la
compression des élémentscellulaires et enfin l'éclatement
de l'entérocyte qui libère alors de nombreux
schizozoïtes ; ces derniers vont alors parasiter d'autres
entérocytes et subir de nouvelles schizogonies.
b) Lors du cycle extra-intestinal
Dans la vésicule parasitophore de la cellule
hôte, il y a formation d'ébauches de cellules filles. Ce processus
est très rapide, c'est pourquoi les endodyozoïtes ainsi
formés ont été appelés par Frenkel, en 1973,
`'tachyzoïtes'' (tachus = rapide) : les
tachyzoïtes sont donc les formes de multiplications rapide du parasite.
Ce phénomène se poursuit à
l'intérieur de la vacuole de la cellule hôte. Des multiplications
successives aboutissent à la formation des pseudokystes dont la paroi
est fine. La membrane finit par éclater et les tachyzoïtes sont
libérées et peuvent alors parasiter d'autres cellules de
l'hôte (intermédiaire ou même définitifs).
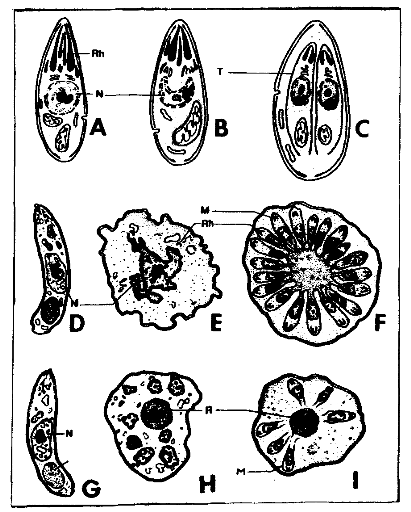
Figure 5 :Schémas des différents
types de multiplication asexuée : endodyogenie (A, B et C),
endopolygenie (D, E et F), merogonie (G, H et I) ; corps residuels (R),
schizozoïtes (M), noyaux (N), rhoptries (Rh) et tachyzoïtes
(T)²
III. 2. Autres fonctions biologiques
III. 2. 1. La locomotion
Les flux lymphatiques et surtout sanguins,
assurent la dissémination du parasite la position enterale. Le
péristaltisme gastro-intestinal assure la progression des oocystes
sporulés et des kystes ingérés vers l'intestin
grêle, et l'excrétion dans le milieuextérieur.?
De nombreux facteurs extérieurs vont
intervenir dans la dissémination de la T. gondii(le vent,
l'eau, les animaux, les engins et les vêtements humains,...).?
Aussi, le parasite est capable de petits
déplacements permettant de se rapprocher des cellules hôtes
grâce à la mobilisation de son cytosquelette interne très
développé et la mise en action du système de
pénétration (mécanique et enzymatique) dans la cellule
hôte. Enfin les flagelles des gamètes males jouent un rôle
important dans la fécondation du gamète femelle.?
III. 2. 2. La nutrition
La toxoplasmose est un parasite intracellulaire et cette
fonction est réaliséegrâceà des échanges
transmembranaires intenses. Les
élémentsnécessairesà la réalisation des
différentes fonctions biologiques du parasite sont présents dans
le cytoplasme de la cellule hôte. Le parasite utilise les réserves
glucidiques et l'oxygènetrouvé dans la cellule hôte pour la
réalisation du métabolisme respiratoire.
III. 2. 3. La protection : résistance
et sensibilité
La paroi des oocystes, trèsépaisse et
très peu perméableconfère aux sporozoïtes une grande
résistance face à divers facteurs mécaniques et
physico-chimiques de l'environnement nombreux désinfectants. En
revanche, il est sensible à la sècheresse et à la chaleur
(66°C pendant vingt minutes).
IV. Cycle biologique
IV. 1. Le cycle entero-épithélial ou
coccidien
Il n'a lieu que chez l'hôte définitif
(félidés) et peut se produire suite à l'ingestion des
kystes tissulaires. Le devenir du kyste ingéré : la paroi du
kyste est détruite par l'action simultanée des contractions
stomacales, du péristaltisme intestinal et des sucs digestifs (phase
d'excystement). Certains bradyzoïtespénètrent alors dans les
entérocytes de l'intestin grêle. En position intracellulaire, le
bradyzoïte devient schizonte : stade intermédiaire. Les
schizontes subissent alors la multiplication asexuée ; il en
résulte de nombreuses générationsconstituantes cinq types
morphologiques différents : les types A, B, C, D et E. Seulement
les types D et E subissent la reproduction sexuée. Apres la
fécondation, les zygotes s'enkystent et deviennent des oocystes
immatures ; ces derniers sont libérés dans le milieu
extérieur avec les selles de l'hôte. Lorsque les conditions
d'humidité, de température et d'oxygénation du milieu sont
favorables, la sporulation peut se produire.4
Une minorité de bradyzoïtes ne subit
pas le cycle entero-épithéliale mais pénètre
directement la lamina propriade l'intestin grêle puis passe dans
le sang et la lymphe.4
IV. 2. Le cycle extra-intestinal
Il a lieu aussi bien chez les
hôtesdéfinitifs que chez les hôtes intermédiaires et
résulte le plus souvent de l'ingestion d'oocystes sporules ou de kystes
tissulaires.4
Les sporozoïtes sont libérés
par l'action mécanique et enzymatique de la partie antérieure du
tube digestif, vont pénétrer dans la paroi de l'intestin et le
parasite subit une série de divisions. La multiplication cause
l'éclatement de la cellule hôte et les rends libres pour parasiter
les autres cellules.4
Une grande partie d'entre eux peut alors se
répandre dans les tissus extra-intestinaux via les circulations. Les
tachyzoïtes sont capables de se multiplier dans la plupart des types
cellulaires. Lorsqu'une réponse immune efficace est mise en place, la
multiplication tachyzoïtique ralentit et les pseudokystes se transforment
en kystes. Ces derniers, s'ils sont consommes par un nouvel hôte,
pourront initier un nouveau cycle selon l'hôte.4

Figure 6: Schema du cycle biologique de T.
gondii5
TROISIEME PARTIE:
ETUDE EPIDEMIOLOGIQUE DE LA TOXOPLASMOSE
I. Les populations atteintes
I. 1. Les hôtes définitifs
Chez les hôtes définitifs, T. gondii
peut effectuer le cycle entero-épithélial mais aussi le cycle
extra-intestinal. Le chat domestique (Feliscatus) n'est pas le seul
hôte définitif connu ; en effet, l'observation a
démontré l'excrétion d'oocystes chez d'autres
félidés comme : le chat sauvage, le puma, l'ocelot, le
margay, le jaguar, le lynx, le tigre, le lion,...°
I. 2. Les hôtes
intermédiaires
Contrairement aux précédents, les hôtes
intermédiaires ne peuvent subir que le cycle extra-intestinal. La
plupart des vertébrés homéothermes semblent
concernes : de nombreux mammifères (rongeurs, lagomorphes,
mustélidés, canidés, équidés,
bovidés, suidés, primates, homme) et de nombreux oiseaux. En
addition de nombreux vertébrés a sang froid (reptiles,
amphibiens, poissons).°
I. 3. Les hôtes
parénétiques
Il existe aussi des hôtes capables de
véhiculer les oocystes ou des kystes qui n'évoluent pas mais qui
peuvent être disperses, intacts, de cette manière. Ces derniers
sont les insectes carnassiers, coprophages, des lombrics, des mollusques... Ils
conservent le caractère infectant du parasite, et, s'ils sont consommes
par les hôtes sensibles, T. gondii peut poursuivre son cycle de
développement.°
II. Les sources du parasite
II. 1. Les réservoirs d'oocystes : les
fèces des hôtes définitifs
Ce n'est vrai que pendant la phase
d'excrétion. Seuls les félidés domestiques et certains
félidés sauvages comme le lion, le jaguar, le lynx, le tigre,
peuvent êtreexcréteurs.
Ainsi, les selles des chats infectées et
excréteurs peuvent contaminer :
- L'eau (mares, lacs, étangs, ruisseaux,
rivières...) ;
- Le sol (terre des jardins, terrains de jeu, bac à
sable, sol des prairies...) ;
- Les végétaux (fruits, légumes,
pâturages...).
| 

