3.4. Coefficient de variation
L'évolution du coefficient de variation (CV final/CV
initial) parait moins importante chez les alevins venant de la condition tiroir
alimenté avec l'eau de forage (1,1#177;0,1). En revanche, elle semble
avoir été plus élevée chez les individus venant des
conditions claies sans substrat (1,2#177; 0,8) et chez ceux venant de la
condition tiroir alimenté avec l'eau de rivière (1,3#177;0,8).
Cependant l'analyse statistique n'a permis de détecter
aucune différence significative entre les moyennes.
3.5. Lipides et acides gras
? Lipides totaux
Pour les lipides totaux, une différence significative a
été détectée entre les larves de la condition
tiroir alimenté avec l'eau de forage (210#177;13 mg/g de poids sec) et
celles de la claie avec galets (193 #177;12 mg/g de poids sec) (p = 0,001333).
Aucune autre différence significative n'a été mis en
évidence entre les larves des autres conditions.
Aucune différence significative n'a pas non plus
été détectée entre les larves de toutes les
conditions en ce qui concerne les phospholipides (PL). En revanche, pour les
lipides neutres (LN), une différence significative a été
trouvée entre les individus de la condition tiroir alimenté avec
l'eau de forage (92 #177;8 mg/g de poids sec) et ceux de la condition claie
avec substrat (77#177; 7mg/g de poids sec) ou celle de la rivière Aisne
(p = 0,000324) (tableau 3).
? Acides gras saturés et
monoinsaturés
Les conditions d'incubation semblent influencer la
qualité énergétique des larves à l'émergence
au niveau de la fraction des LN. En effet, au niveau des acides gras
saturés, une différence significative a été
trouvée pour le C14: 0 entre les larves de la condition claie sans
substrat (2,3 mg/g poids sec) et celles de la condition claie avec galets (1,7
mg/g poids sec). Pour le C16:0, une différence significative a
été trouvée entre les larves de la condition tiroir
alimenté avec l'eau de forage (10,4 mg/g poids sec) et celle de la
condition claie avec galets (7 mg/g poids sec). Une différence
significative a été également détectée entre
les larves de la condition tiroir alimenté avec l'eau de forage (3,9
mg/g poids sec) et celles de la condition claie avec galets (3,2 mg/g poids sec
pour le C18: 0. Cette influence des conditions d'incubation sur la
qualité énergétique des larves à l'émergence
a été également observée pour les acides gras
monoinsaturés (AGMI) de cette fraction lipidique. Une différence
significative a été trouvée entre les larves de la
condition tiroir alimenté avec l'eau de forage (6,3#177;0,7 mg/g poids
sec) et les larves de la condition claie avec galets (4,1#177;0,7 mg/g poids
sec) pour le C16: 1 (n-7). Pour le C18: 1 (n-7), une différence
significative a été trouvée entre les individus de la
condition tiroir alimenté avec l'eau de forage (3,4#177;0,3 mg/g poids
sec) et ceux de la condition claie avec galets (2,6#177;0,5 mg/g poids sec).
Aucune différence significative n'a été
détectée pour le 0 : 1 (n-9) (tableau 4).
Dans la fraction des PL, aucune différence
significative n'a été trouvée pour les acides gras
saturés C14:0 et C16: 0. En revanche, une différence
significative a été mise en évidence entre les larves de
la condition tiroir alimenté avec l'eau de forage (3,5#177;0,4 mg/g
poids sec) et celles de la

Nombre de larves émergées
A
140
120
100
40
80
60
20
0
Radier 1 Radier 2 Radier 3

Nombre de larves émergées
B
40
90
80
70
60
50
30
20
10
0
Radier 1 Radier 2 Radier 3
Figure 10:Evolution de l'émergence dans
les rivières Aisne (A) et Samson (B)
20
condition claie avec galets (2,8#177;0,5 mg/g poids sec). Pour
les AGMI, C16: 1 (n-7), C18 : 1 (n-7) et C18: 1 (n-9), aucune différence
significative n'a été détectée entre les larves des
différentes conditions d'incubation. Par contre, une différence
significative a été trouvée entre les larves de la
condition claie sans substrat (0,19#177;0,03 mg/g poids sec) et les larves de
la condition claie avec galets (0,13#177;0,02 mg/g poids sec) pour le 0: 1(n-9)
(tableau 5).
? Acides gras (n-3 et n-6)
Le DHA (2:6(n-3)) est l'acide gras de la série n-3
présent en plus forte proportion dans les PL et les LN. La
qualité biochimique relative aux acides gras de la série n-3 et
n-6 des larves à l'émergence était peu différente
entre les différentes conditions d'incubation.
Toutefois pour la fraction LN, des différences
significatives ont été détectées pour des acides
gras importants. A ce titre, les larves de la condition tiroir alimenté
avec l'eau de forage sont significativement plus riches en acide arachidonique
(0: 4 (n-6)) (1,2#177;0,10 mg/g poids sec) que les individus de la condition
claie sans substrat (0,94#177;0,07 mg/g poids sec). La teneur en EPA (0: 5
(n-3)) est significativement plus élevée chez les larves issues
des conditions tiroir alimenté avec l'eau de forage (7,2#177;0,7 mg/g
poids sec). Les larves issues de cette variante d'incubation sont aussi
significativement plus riches en DHA (13,4#177;1,4 mg/g poids sec) que les
larves de toutes les autres conditions.
Pour la fraction PL, les larves de la condition claie sans
substrat sont significativement plus riches en acide arachidonique (1,2 mg/g
poids sec) que les larves de la condition rivière Samson (1mg/g poids
sec). L'EPA est significativement plus élevé chez les larves de
la condition tiroir alimenté avec l'eau de forage (4,7#177;0,3 mg/g
poids sec) que chez celles de la condition claie avec galets (4#177;0,2 mg/g
poids sec). Les larves de la condition claie sans substrat sont
significativement plus riches en DHA (16#177;0,7 mg/g poids sec) que celles de
la condition rivière Samson (14#177;0,4 mg/g poids sec) (tableau 6).

Taux de survie à l'émergence
(%)
120
100
40
80
60
20
0
b b
a
a
a a

Taux de survie post-émergence (%)
40
90
80
70
60
50
30
20
10
0
Claie
avec
galets
b
Claie
sans
substrat
c
Tiroir
avec eau
de
rivière
bc
Tiroir
avec eau
de
forage
a
21
Figure 11: Variation du taux de survie
à l'émergence (A) et après 19 jours post émergence
(B) en fonction des conditions d'incubation (moyenne + écart-type). (A):
N= 3, P= 0,37 pour les rivières ; (B): N= 3, P= 0,000615).
Poids des larves (mg)
160
140
120
100
40
80
60
20
0
a a a a
b
c

Longueu r des larves (mm)
30 b a c a a bc
25
20
15
10
0
5
Figure 12 : Variation du poids moyen (A) (N=
54, P= 3,6E-39) et de la longueur (B) (N= 54, P= 8,98E-30) des larves à
l'émergence en fonction des conditions d'incubation ( moyenne +
écart-type).
22

Gain de poids (%)
250
200
150
100
50
0
Claie
sans
substrat
Claie
avec
galets
Tiroir
avec eau
de rivière
Tiroir
avec eau
de forage

SGR ( %/ jou r)
4
0
7
6
5
3
2
1
claie sans substrat
Claie avec galets
Tiroir avec
eau de
rivière
Tiroir avec
eau de
forage
Figure 13 : Variation du gain de poids relatif
(A) (N= 3, P= 0,06) et du SGR (B) (N= 3, P= 0.69) des alevins
post-émergents ( moyenne + écart-type).

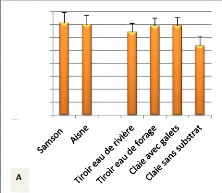
CV final / CV initial
0,5
2,5
1,5
0
2
1
Claie sans substrat
Claie avec galets
Tiroir
avec eau
de rivière
Tiroir
avec eau
de forage
Figure 14 : Evolution du coefficient de
variation du poids en fonction des conditions d'incubation ( moyenne +
écart-type). (N= 3, P= 0,89)
Tableau 3 : Composition en lipides corporels
(mg/g de poids sec) des larves à l'émergence en fonction du mode
d'incubation. Les moyennes sur une même ligne
accompagnées de lettres différentes sont significativement
différentes (P<0,05). Lorsqu'aucune lettre n'apparaît, il n'y a
pas de différence significative. N=6.
|
Fractions
lipidiques
|
Rivière Aisne
|
Rivière Samson
|
Claie avec
galets
|
Claie sans
substrat
|
Tiroir alimenté
avec eau de
forage
|
Tiroir alimenté
avec eau de
rivière
|
|
Lipides totaux
|
184,1#177;18,1ab
|
185,3#177;8,1ab
|
171,7#177;13,4b
|
192,7#177;12,2ab
|
210,2#177;12,7a
|
193,5#177;15,3ab
|
|
Phospholipides
|
43,0#177;1,3
|
39,9#177;1,1
|
42,7#177;6,5
|
45,2#177;3,0
|
43,7#177;1,9
|
43,4#177;0,9
|
|
Lipides neutres
|
73,5#177;11,5b
|
77,9#177;5,1ab
|
65,3#177;8,5b
|
77,4#177;6,5ab
|
91,9#177;8,4a
|
78,0#177;8,4ab
|
23
Tableau 5 : Composition en acides gras corporels
(mg/g de poids secs) des larves à l'émergence en fonction du mode
d'incubation, fraction LN. Les moyennes sur une même ligne
accompagnées de lettres différentes sont significativement
différentes (P<0,05). Lorsqu'aucune lettre n'apparaît, il n'y a
pas de différence significative. N=6. NQ : valeur présente mais
non quantifiable
|
Acides gras
|
Rivière Aisne
|
Rivière
Samson
|
Claie avec
galets
|
Claie sans
substrat
|
Tiroir alimenté avec eau de forage
|
Tiroir alimenté
avec eau de
rivière
|
|
Saturés
|
|
|
|
|
|
|
|
C14 :0
|
1,8#177;0,3ab
|
2,1#177;0,3ab
|
1,7#177;0,3b
|
2,3#177;0,2a
|
2,2#177;0,2ab
|
1,9#177;0,3ab
|
|
C16 :0
|
8,0#177;1,2ab
|
8,8#177;0,8a
|
7#177;0,8b
|
8,4#177;0,8ab
|
10,4#177;0,8a
|
8,4#177;0,9ab
|
|
C18:0
|
3,6#177;0,6ab
|
3,8#177;0,2ab
|
3,2#177;0,3b
|
3,2#177;0,5b
|
3,9#177;0,4a
|
3,6#177;0,5ab
|
|
0 :0
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
|
Monoinsaturés
|
|
|
|
|
|
|
|
C16 : 1(n-7)
|
4,7#177;0,8b
|
4,8#177;0,6b
|
4,1#177;0,7b
|
5,3#177;0,5ab
|
6,3#177;0,7a
|
5,0#177;0,6ab
|
|
C18: 1(n-7)
|
2,8#177;0,4ab
|
3,1#177;0,3ab
|
2,6#177;0,5b
|
3,1#177;0,4ab
|
3,4#177;0,3a
|
3,1#177;0,5ab
|
|
C18: 1(n-9)
|
19,3#177;3,2b
|
20,8#177;1,4ab
|
18,8#177;2,6b
|
21,7#177;1,7ab
|
24,9#177;2,4a
|
21,4#177;2,5a
|
|
0: 1(n-9)
|
0,47#177;0,01
|
0,53#177;0,01
|
0,41#177;0,07
|
0,52#177;0,06
|
0,46#177;0,04
|
0,49#177;0,09
|
|
n-3
|
|
|
|
|
|
|
|
C18 :3(n-3)
|
1,40#177;0,2b
|
1,50#177;0,2ab
|
1,20#177;0,2b
|
1,60#177;0,1ab
|
1,80#177;0,2a
|
1,50#177;0,2ab
|
|
C18 :4(n-3)
|
0,40#177;0,1ab
|
0,46#177;0,1ab
|
0,38#177;0,1b
|
0,50#177;0,1ab
|
0,51#177;0,1a
|
0,44#177;0,1ab
|
|
0 :5(n-3)
|
5,59#177;0,85b
|
5,62#177;0,63b
|
4,80#177;0,68b
|
5,17#177;0,52b
|
7,17#177;0,70a
|
5,86#177;0,47b
|
|
2 :5(n-3)
|
3,30#177;0,58ab
|
3,51#177;0,51ab
|
2,75#177;0,33b
|
3,13#177;0,22ab
|
3,68#177;0,38a
|
3,26#177;0,39ab
|
|
2 :6(n-3)
|
11,08#177;1,6b
|
10,5#177;1,2b
|
9,23#177;1,07b
|
10,03#177;0,94b
|
13,39#177;1,37a
|
11,36#177;1,08ab
|
|
n-6
|
|
|
|
|
|
|
|
C18 :2(n-6)
|
6,28#177;1,1b
|
7,2#177;0,9ab
|
5,80#177;0,86b
|
7,24#177;0,47ab
|
8,23#177;0,82a
|
6,81#177;0,83ab
|
|
C18 :3(n-6)
|
0,14#177;0,0b
|
0,16#177;0,02ab
|
0,15#177;0,04ab
|
0,19#177;0,04ab
|
0,21#177;0,04a
|
0,18#177;0,02ab
|
|
0 :2(n-6)
|
0,43#177;0,08
|
0,48#177;0,08
|
0,38#177;0,05
|
0,47#177;0,03
|
0,47#177;0,04
|
0,45#177;0,07
|
|
0 :3(n-6)
|
0,68#177;0,11
|
0,72#177;0,08
|
0,65#177;0,12
|
0,77#177;0,08
|
0,79#177;0,09
|
0,73#177;0,10
|
|
0 :4(n-6)
|
0,99#177;0,11b
|
1,00#177;0,08ab
|
0,91#177;0,11b
|
0,94#177;0,07b
|
1,16#177;0,10a
|
1,05#177;0,08ab
|
|
2 :4(n-6)
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
24
Tableau 6 : Composition en acides gras corporels
(mg/g de poids secs) des larves à l'émergence en fonction du mode
d'incubation, fraction PL. Les moyennes sur une même
ligne accompagnées de lettres différentes sont significativement
différentes (P<0,05). Lorsqu'aucune lettre n'apparaît, il n'y a
pas de différence significative N=6. NQ : valeur présente mais
non quantifiable
|
Acides gras
|
Rivière Aisne
|
Rivière
Samson
|
Claie avec galets
|
Claie sans substrat
|
Tiroir alimenté
avec eau de
forage
|
Tiroir alimenté
avec eau de
rivière
|
|
Saturés
|
|
|
|
|
|
|
|
C14 :0
|
0,4#177;0,0
|
0,3#177;0,0
|
0,4#177;0,3
|
0,4#177;0,0
|
0,4#177;0,0
|
0,4#177;0,0
|
|
C16 :0
|
9,4#177;0,3
|
9#177;0,1
|
9,6#177;1,4
|
9,4#177;0,7
|
9,9#177;0,4
|
9,7#177;0,2
|
|
C18:0
|
3#177;0,1ab
|
2,7#177;0,2b
|
2,8#177;0,5b
|
3,6#177;0,5a
|
3,5#177;0,4a
|
3,1#177;0,3ab
|
|
0 :0
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
|
Mono insaturés
|
|
|
|
|
|
|
|
C16 : 1(n-7)
|
0,7#177;0,1
|
0,6#177;0,1
|
0,6#177;0,1
|
0,7#177;0,1
|
0,6#177;0,1
|
0,7#177;0,1
|
|
C18: 1(n-7)
|
0,9#177;0,1
|
1#177;0,1
|
0,9#177;0,1
|
1#177;0,1
|
1#177;0,1
|
0,9#177;0,1
|
|
C18: 1(n-9)
|
4,7#177;0,1
|
4,4#177;0,2
|
5,3#177;2
|
4,8#177;0,2
|
4,8#177;0,2
|
4,7#177;0,1
|
|
0: 1(n-9)
|
0,16#177;0,03ab
|
0,13#177;0,03b
|
0,13#177;0,02b
|
0,19#177;0,03a
|
0,14#177;0,02ab
|
0,14#177;0,01ab
|
|
n-3
|
|
|
|
|
|
|
|
C18 :3(n-3)
|
0,18#177;0,08
|
0,13#177;0,01
|
0,14#177;0,05
|
0,12#177;0,01
|
0,13#177;0,01
|
0,15#177;0,02
|
|
C18 :4(n-3)
|
0,03#177;0,0
|
0,03#177;0,0
|
0,05#177;0,05
|
0,03#177;0,0
|
0,03#177;0,0
|
0,03#177;0,0
|
|
0 :5(n-3)
|
4,53#177;0,20ab
|
4,05#177;0,25b
|
3,98#177;0,22b
|
4,51#177;0,46ab
|
4,74#177;0,26a
|
4,45#177;0,13ab
|
|
2 :5(n-3)
|
1,52#177;0,11a
|
1,32#177;0,13ab
|
1,19#177;0,08b
|
1,57#177;0,16a
|
1,49#177;0,14a
|
1,38#177;0,10ab
|
|
2 :6(n-3)
|
14,50#177;0,42b`
|
13,63#177;0,41`
|
14,21#177;0,49
b`
|
15,76#177;0,72a
|
14,19#177;0,49b
|
15,06#177;0,34ab
|
|
n-6
|
|
|
|
|
|
|
|
C18 :2(n-6)
|
0,90#177;0,03
|
0,77#177;0,02
|
1,32#177;1,36
|
0,88#177;0,08
|
0,83#177;0,06
|
0,83#177;0,04
|
|
C18 :3(n-6)
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
NQ
|
|
0 :2(n-6)
|
0,13#177;0,01ab
|
0,11#177;0,02ab
|
0,10#177;0,02b
|
0,14#177;0,02a
|
0,12#177;0,01ab
|
0,12#177;0,01ab
|
|
0 :3(n-6)
|
0,025#177;0,02
|
0,21#177;0,01
|
0,23#177;0,05
|
0,25#177;0,02
|
0,24#177;0,02
|
0,24#177;0,02
|
|
0 :4(n-6)
|
1,02#177;0,02ab
|
0,94#177;0,04b
1,00#177;0,07ab
|
1,20#177;0,09a
|
1,05#177;0,06ab
|
1,07#177;0,03ab
|
|
2 :4(n-6)
|
NQ
|
NQ NQ
|
NQ
|
NQ
|
NQ
|
25
|
|


