d- Etude des lames minces
Vu la nature essentiellement marneuse des dépôts,
seul un petit nombre de lames minces (16 au total) a été
réalisé dans les niveaux les plus indurés, surtout dans le
Cénomanien terminal ou au tout début du Turonien. Il aurait pu
être intéressant d'indurer certains échantillons plus
tendres afin d'en étudier le microfaciès, mais cette technique
n'a pas pu être exploitée dans le cadre de notre étude.
Dans ce qui suit, une description succincte de chaque lame va
être proposée, illustrée de nombreux clichés
(planches 9, 10 et 11). Dans cette description, les classifications communes de
Folk (1962) et de Dunham (1962) seront couramment employées.
Ces lames, réalisées le plus souvent dans du
calcaire marneux, et plus rarement dans du calcaire franc (Cénomanien
moyen et Turonien inférieur) montrent des caractéristiques
communes : il s'agit toujours de calcaire à texture micritique, de
teinte souvent sombre (excepté dans le Turonien inférieur
où les teintes sont généralement plus claires). Le contenu
en microfossiles et les microfaciès sont cependant fort
différents d'une lame à l'autre.
PLANCHE 9 (page suivante) Vraconnien
Lame 1B (2 clichés)
Calcaire micritique sombre, de type
packstone à wackestone, à
calcisphères nombreuses, foraminifères
planctoniques globuleux (Hedbergella, Globigerinelloides) et rares
ostracodes. Des cubes de pyrite sont bien visibles en remplissage de certaines
formes (photo 1). De rares `lentilles' de matière organique sont
présentes.
Certaines parties de la lame montrent une succession
caractéristique de microbancs clairs, riches en calcisphères,
alternant avec des bancs sombres, contenant moins de calcisphères et
proportionnellement plus de foraminifères planctoniques (photo 2).
Cénomanien inférieur
Lame 15
Calcaire micritique sombre, de type
wackestone, à calcisphères, foraminifères
planctoniques globuleux, petits foraminifères benthiques, ostracodes et
fragments d'échinodermes. La présence d'oxydes de fer, et surtout
de glauconie est remarquable (cette dernière a d'ailleurs
été décrite précédemment dans un
échantillon contigu (n°16)).
Les foraminifères planctoniques reconnues sont les
suivants : Hedbergella simplex, H. delrioensis, Heterohelix sp.,
Globigerinelloides sp.
Cénomanien moyen Lame
46
Calcaire micritique, de type wackestone,
à nombreux débris de bivalves,
calcisphères et rares foraminifères, souvent très
recristallisés, donc impossibles à identifier.
Lame 62 (2 clichés)
Calcaire micritique, de type
wackestone à packstone, à nombreux
débris (bivalves, échinodermes), foraminifères
benthiques (miliolidés, Lenticulina sp....),
calcisphères, ostracodes et rares foraminifères planctoniques
(globuleux). Dans certaines parties de la lame, des traces de
bioturbation sont visibles (photo 2).
Lame 74 (aucune photo)
Calcaire micritique, de type
wackestone, à débris
d'organismes (bivalves, gastéropodes, échinodermes),
foraminifères benthiques et calcisphères. Cette lame est
globalement très proche de la précédente, excepté
une richesse fossilifère moindre. Des traces de bioturbation
sont également décelables.
|
|
Chap. III : Résultats et interprétations
|
|
Lame 1 B (1)
|
Lame 1 B (2)
|
|
|
|
Lame 15
|
|
Lame 46
|

Légende : B : foraminifère benthique, 0 :
ostracode, GI : glauconie, Hed : Hedbergella
sp.
Planche 9 : Lames minces (Vraconnien -- Cénomanien
moyen).
PLANCHE 10 (page suivante)
Cénomanien supérieur
Lame 95 : photo 1
Calcaire micritique sombre, de type
mudstone, à foraminifères planctoniques
globuleux (Hedbergella, Whiteinella, Heterohelix). La lame est
encombrée de nombreuses lentilles de matière organique
sombres.
Lame 99 : photo 2
Calcaire micritique, de type
mudstone, d'une couleur beaucoup plus claire que
l'échantillon précédent et visuellement très
différent (absence totale cette fois-ci de lentilles de MO). Les rares
microorganismes présents sont des calcisphères, des
foraminifères planctoniques globuleux (Whiteinella, Heterohelix
probables) et des benthiques calcaires. La présence
d'oxydes de fer est également marquante.
Cénomanien supérieur -- faciès
Bahloul4 Lame 103 (2 clichés)
Calcaire micritique sombre, de type
wackestone à packstone, très riche en microorganismes,
principalement des foraminifères planctoniques globuleux, mais
également en petits foraminifères benthiques, ostracodes,
débris d'échinodermes et rares filaments (= petits bivalves
fins). La présence de glauconie et de lentilles
de MO en grande quantité est également remarquable.
Les foraminifères planctoniques, pour la plupart d'une
grande taille exceptionnelle, comprennent Heterohelix sp.,
Hedbergella simplex, H.delrioensis, Globigerinelloides sp.
Lame 104 (2 clichés)
Calcaire micritique sombre, de type
wackestone, à foraminifères
planctoniques globuleux, également de grande taille :
Heterohelix sp., Hedbergella sp., Whiteinella sp., W.
paradubia. Des filaments, plutôt rares, sont également
présents, ainsi que des lentilles de MO en abondance.
En revanche, la glauconie, fréquente dans la lame
précédente, est totalement absente, de même que les
organismes benthiques.
Lame 95 Lame 99

Lame 103 (1)
Lame 104 (1) Lame 104 (2)

Op* nie.>
·
le
b
500 iam
·
·1111,..

Légende : B : Foraminifère benthique, H :
Heterohelix sp., Hed : Hedbergella sp., GI : Glauconie, W :
Whiteinella sp.
Planche 10 : Lames minces (Cénomanien
sup.).
PLANCHE 11 (page suivante) Lame 105 (2
clichés)
Calcaire micritique sombre, de type
wackestone, à foraminifères
planctoniques globuleux : Heterohelix sp. (majoritaires),
Hedbergella sp., Whiteinella sp.. Les traces de MO
sont abondantes (lentilles).
Remarque: cette lame a été
réalisée parallèlement à la stratification du fait
d'un délitage en fins
feuillets. Les informations déduites en sont donc
nécessairement amoindries. Lame 106 (2 clichés)
Calcaire micritique sombre, de type
mudstone à wackestone, à
foraminifères planctoniques globuleux (Hedbergella
sp., Whiteinella sp., rares Heterohelix sp.),
calcisphères, rares filaments et lentilles de MO
nombreuses. La richesse en microfossiles est nettement moins
élevée que dans la lame précédente. Des formes
assez mystérieuses sont également observées,
peut-être s'agit-il de fragments d'échinodermes (ophiures ??).
Turonien inférieur
Lames 110, 114, 118, 120, 122 (3 clichés)
Nous avons choisi de traiter ces différentes lames
ensemble car elles présentent un microfaciès très
proche.
Il s'agit de calcaire micritique, clair, de
type mudstone, à rares microorganismes,
souvent aux contours flous, donc très difficiles à
identifier. Sont présents des calcisphères, quelques ostracodes,
de très rares foraminifères planctoniques globuleux (dont des
Heterohelix) et d'aussi rares foraminifères benthiques.
Les oxydes de fer sont fréquents, en
taches ou en remplissage de microfractures (stylolithes). Ce genre de structure
est d'ailleurs commun (lame 120).
Des auréoles sombres, finement ciselées, entourent
parfois certains microorganismes (oxydes ? lame 118).
Lame 116: photo ci-dessous

Calcaire micritique, de type
mudstone, au sein duquel aucun microorganisme n'a pu
être observé. Le microfaciès, très différent
des lames précédentes, nous a conduit à décrire
cette lame séparément. On y observe en effet un litage
bien marqué (succession de lits sombres et d'autres lits
clairs), et une structure par endroit très perturbée, avec une
modification nette de l'orientation du litage (bioturbation ?).
Lame 105 (1) Lame 105 (2)

Légende :
H : Heterohelix sp., Hed :
Hedbergella sp., O : ostracode, Oph : ophiure (
?).
Planche 11 : Lames minces (Cénomanien sup.
--
Turonien inf.).


3. Interprétations
Nous aborderons successivement les aspects
biostratigraphiques, puis paléoenvironnementaux /
paléogéographiques. Suivra ensuite une description de la crise
cénomano-turonienne, ainsi que de la transition entre Vraconnien et
Cénomanien du début de coupe. La réponse à quelques
interrogations servira enfin de conclusion à ce chapitre.
a- Axe Biostratigraphie
Préciser la biostratigraphie est chose difficile quand
les déterminations sont incertaines et la documentation disponible trop
rare. L'extrême rareté des foraminifères planctoniques
carénés (et des ammonites) constitue également un handicap
pour une biostratigraphie précise, d'autant plus que les
spécimens observés ne sont pas toujours en excellent état
de conservation, donc parfois difficiles à déterminer
précisément.

,,,_
. ,-`' I -P,
te ft] m
_Q
Ç)
ir cr O
Tura nien inférieur
-
-
-
_
--
--
--
--
--
--
_r ri
-
-
Cénomanien moyen é supérieur
-
·
Cenomanien inférieur
Vraconnien
La figure ci-contre présente la répartition
|
106
|
|
des différentes espèces de foraminifères
|
104
|
|
planctoniques utiles pour la
|
100
|
|
biostratigraphie. Seuls certains
|
95
|
|
échantillons sont indiqués, ceux-ci ne
|
90
|
|
représentent qu'une petite partie du total
|
86
|
|
des échantillons (à titre d'exemple, seuls
|
80
|
|
14 échantillons sur 70 étudiés
contiennent des spécimens du genre
|
|
|
Rotalipora).
|
70
|
|
De plus, l'échelle verticale n'est pas respectée
(en réalité, l'épaisseur du
|
60
|
|
Cénomanien inférieur est largement
accrue).
|
50
|
|
La présence des espèces
appartenant aux genres Hedbergella,
Globigerinelloides
et Heterohelix n'est
pas précisée, ceux-ci
étant communs à
|
40
|
|
32
|
|
presque tous les niveaux.
|
28
|
|
24
|
|
20
|
|
14
|
|
11
|
|
2
|
Figure 15 : Foraminifères planctoniques --
Répartition
de quelques espèces-clés.
En référence à la biozonation du
Crétacé proposée par Robaszynski et Caron (1995) [fig.16],
il est donc possible de différencier, tant bien que mal, plusieurs zones
dans la coupe.
|
Périodes
|
Globotruncanidés
|
Hétérohélicidés
|
|
CONLACIEN
|
I
|
DICARINELLA concavata
^
|
nuttaifi
|
cane
|
s
|
PSEUDOTEXTULARIA
|
|
|
|
|
TURONIEN
|
S
|
|
HETEROHELIX
globulosa {=reussi)
|
|
n
|
|
M
|
MARE schneegansi
|
|
HFLV helvetica
|
'hely
|
|
I
|
|
WHIT. archaeocretacea
|
|
CENOMANIEN
|
S
|
|
ROTAL. cushmani
|
|
I ar
|
|
HE TEROHFLIX moremani
|
g
|
|
M -
|
ROTAL. reichefi
|
lcush
|
|
I
|
ROTAL.
globotruncartoides (= brotzeni)
|
r
|
|
ALBIEN
|
S
|
ROTAL. appenninica
|
A
gl.
|
|
ROTAL. ticinensis
|
ap.
--.%
|
|
m
|
|
ROTAL. subticinensis
-A
|
tic
|
|
TICIAt praeticinensis sb
|
ROTAL Rotalipora, Ticinelfa, VVHIT. Whiteineila,
HELV. Heivetoglobotruncana, MARG. Marginotruncana
Figure 16 : Biozonation du Crétacé moyen
à supérieur par les foraminifères
planctoniques.
(d'après Robaszynski et Caron, 1995)
Les différentes espèces observées
permettent de réaliser un découpage. La limite inférieure
du Cénomanien n'a pas pu être définie avec
précision, mais, en revanche, trois zones ont été
reconnues entre le début du Cénomanien et celui du Turonien :
- Cénomanien inférieur (zone à
Rotalipora brotzeni)
Sa limite inférieure (avec le Vraconnien) ne peut
être située précisément.
Cette zone comporte, en plus de nombreuses espèces des
genres Hedbergella et Globigerinelloides, les espèces
Favusella washitensis, Rotalipora appenninica, Heterohelix moremani et
plusieurs spécimens qui semblent bien appartenir à
l'espèce R. brotzeni.
C'est dans cette zone que la diversité
spécifique est la plus grande (fig.12), ce qui semble correspondre aux
dires de Hart et Ball (1986) qui parlent 'd'un accroissement rapide de la
diversité de la faune aux environs du Cénomanien moyen'.
Cénomanien moyen à supérieur (zone
à Rotalipora cushmani)
L'espèce index n'a pas été
identifiée avec certitude, mais l'entrée dans cette zone est
à supposer du fait de l'absence des espèces
précédentes (F.washitensis, R. appenninica et
R.brotzeni). Plusieurs spécimens, attribués au genre
Rotalipora, n'ont malheureusement pas pu être identifiés
avec plus de précisions à cause d'un trop mauvais état de
conservation. Les
derniers spécimens du genre Rotalipora sont
observés dans l'échantillon n°86, au niveau de la partie
finale du Cénomanien supérieur.
Par ailleurs, les espèces appartenant aux genres
Hedbergella, Heterohelix (H.moremani essentiellement) et
Globigerinelloides restent communes
- Partie finale du Cénomanien -- début du
Turonien (zone à Whiteinella archeocretacea)
Marquée par l'absence des spécimens du genre
Rotalipora et par l'apparition des Whiteinella (W. baltica, W.
paradubia et W. archeocretacea), cette zone est bien
caractérisée. Les genres Hedbergella et Heterohelix
sont toujours présents, ce dernier étant très
abondant à certains niveaux. En plus de l'espèce Heterohelix
moremani, bien représentée, s'ajoute l'espèce
Heterohelix globulosa.
L'oursin Heterodiadema libycum (observé dans les
bancs calcaires du début du Turonien) semble également être
caractéristique de cette période.
b- Axe Paléoenvironnement /
Paléogéographie
· Etude des microfossiles
:
En plus de la présence d'espèces ou
d'associations d'espèces particulières, qui peuvent indiquer des
conditions environnementales spécifiques, c'est également la
variation de la proportion des différents types de micro-organismes
(benthiques ou pélagiques, foraminifères ou ostracodes...) qu'il
est intéressant d'étudier.
La figure présentée à gauche montre ainsi
l'évolution de différents rapports selon le milieu
considéré.
A ces rapports pourrait être rajouté le rapport
[Globuleux/ Total des planctoniques], qui diminue théoriquement avec
l'approfondissement du milieu.
Figure 17 : Variations du contenu
micropaléontologique en domaine marin exprimées par
différents rapports (d'après Rey, 1983 in Andreu,
1991).
54
Domaine marin
|
|
margino- littoral
|
infra- littoral
|
circa- littoral
|
bathyal
|
abyssal/ hadal
|
|
|
-----7.---F---------
0
|
Planctonique/benthique
|
|
|
|
calcaires/agglutinés
|
|
|
|
---NN
|
..,.._______V 0
|
individus/espèces
|
|
|
|
0 foranninifèresios
|
racodes
|
|
|
®
|
|
|
|
_I--.
0 lisses/0 ornés
|
Les rapports (D, (D et a) concernent les
foraminifères
|
Globalement, les proportions :
> de foraminifères planctoniques
> de foraminifères à test agglutiné
(parmi les foraminifères benthiques)
> de foraminifères à morphologie
carénée (parmi les foraminifères planctoniques)
> d'ostracodes à carapace lisse
|
8.10.
|
augmentent avec
l'approfondissement du
milieu
|
|
Tandis que dans le même temps,
> la proportion d'ostracodes
(parmi l'ensemble des
microfossiles)
|
ca11>
|
diminue avec
l'approfondissement du
milieu
|
Plusieurs auteurs apportent même plus de
précisions, il semble ainsi que le régime planctonique domine
parmi les microorganismes à partir de la plate-forme distale. La
proportion de foraminifères planctoniques croît en effet
régulièrement depuis les milieux littoraux (Vivière, 1985
; Masse, 1988) :
- 30 à 40 % au niveau de la plate-forme externe,
- 40 à 50 % aux environs de la limite circalittoral /
bathyal,
- 50 à 70 % sur le talus.
Concernant les différences de morphologie des
foraminifères planctoniques (globuleux/caréné),
Groshény et Malartre (1997) précisent que les morphotypes
globuleux (considérées comme des équivalents anciens des
formes épineuses actuelles), qui correspondent à des formes
simples, avec cycle de reproduction court, se trouvent au niveau de la
plate-forme (ou en bordure) jusqu'à une profondeur de 100 à 200
m. Au contraire, les morphotypes carénés (pour leur part
équivalents aux formes non-épineuses actuelles), plus complexes,
et donc au cycle de reproduction plus long, ont besoin d'une grande tranche
d'eau, au large du talus.
Selon ces mêmes auteurs, l'augmentation de la proportion
de formes carénées au détriment des formes globuleuses
peut donc être expliquée par la mise en place d'un milieu de plus
en plus profond et ouvert. Un tel milieu favoriserait également
l'accroissement de la taille des individus et de la diversité
spécifique (Murray, 1976).
· Caractéristiques
générales et implications paléoenvironnementales
:
Connaissant ces généralités, il est
possible d'interpréter les variations observées en termes de
variation des paléoenvironnements. En ce qui nous concerne, et bien que
quelques caractéristiques générales (tab.3) puissent
être relevées, il paraît souhaitable de diviser la coupe en
plusieurs secteurs aux caractéristiques communes La variabilité
semble en effet importante.
|
Caractéristiques
diverses
(pétrographiques, minéralogiques,
géochimiques, paléontologiques)
|
|
Types de dépôts
|
Principalement marnes ou calcaires marneux
|
|
Couleur
|
Généralement sombre
|
|
Géochimie
|
COT
|
0,1 à 2 %
|
|
Minéralogie
|
Glauconie
|
5,7% des échantillons
|
|
Gypse
|
60 % des échantillons
|
|
Nodules ferrugineux / pyrite
|
45 % des échantillons
|
|
Fossiles
|
Bivalves, gastéropodes, échinodermes, rares
ammonites
|
|
Valeur des rapports déterminés
à l'issu du tri
|
|
Rapports
|
Moyenne
|
Variation
|
|
F/(F+O)
|
0.87
|
0.17 à 1
|
|
0 lisse / 0
|
0.95
|
0.56 à 1
|
|
P/(P+B)
|
0.54
|
0.02 à 1
|
|
PG/(PG+PC)
|
0.99
|
0.92 à 1
|
|
BA/(BA+BC)
|
0.65
|
0 à 1
|
Tableau 3 : Résumé des
caractéristiques générales de la coupe
étudiée.
Signification des sigles utilisés :
F : foraminifère, 0 : ostracode, P / PG / PC :
foraminifère planctonique, à morphologie globuleuse (G) ou
carénée (C), B / BA / BC : Foraminifère benthique,
à test agglutiné (A) ou calcaire (C).
Ce tableau nous permet de voir que, en moyenne :
les ostracodes sont peu abondants, et sont le plus souvent
à carapace lisse,
les foraminifères planctoniques représentent un peu
plus de la moitié du total des foraminifères,
les foraminifères planctoniques à morphologie
globuleuse dominent nettement par rapport à ceux à morphologie
carénée,
les foraminifères benthiques à test calcaire sont
relativement rares par rapport aux agglutinés,
les dépôts sont le plus souvent marneux et
contiennent fréquemment gypse, pyrite et/ou nodules ferrugineux,
le taux de COT est relativement faible, excepté dans les
niveaux de transition du Cénomanien au Turonien,
les fossiles observés comportent de nombreux bivalves, des
gastéropodes, des échinodermes et de rares ammonites.
Par conséquent,
· La granulométrie des dépôts
(majoritairement marneux ou calcaro-marneux), l'absence de fossiles
caractéristiques de milieux superficiels, la faible proportion
d'ostracodes ornés, ainsi que la dominance de foraminifères
benthiques à test agglutiné, plaident en faveur d'un
milieu de dépôt relativement profond, type plate-forme
distale.
· La proportion de foraminifères planctoniques
(environ 50%) ainsi que l'extrême rareté des foraminifères
planctoniques de type caréné permet d'exclure un milieu
plus profond type talus. La proportion généralement
élevée de foraminifères benthiques à test
agglutiné paraît cependant étonnante, de même que la
très grande rareté des foraminifères planctoniques
carénés.
· La présence fréquente de nodules
ferrugineux et de pyrite indique un milieu à tendance anoxique,
et même nettement anoxique à la limite
cénomanoturonienne (ce que confirment des valeurs relativement
élevés de COT, de l'ordre de 1 à 2 %).
En résumé, il faut envisager un milieu
de dépôt de type plate-forme externe (en limite du talus), une
profondeur de l'ordre de 200 m et une tendance à l'anoxie
fréquente.
· Comment
l'hétérogénéité des résultats
observée mène à une interprétation d'ordre
séquentiel... :
En effet, au vu d'une variabilité parfois grande, la
coupe a été subdivisée en plusieurs zones,
numérotées de 1 à 7 (fig. 18). Ces différentes
zones ont été distinguées surtout vis-à-vis de
leurs teneurs relatives en foraminifères planctoniques, ce qui est a
priori une bonne indication des variations de profondeur du milieu de
dépôt.
On comprend que le Cénomanien, loin d'être une
période homogène, (ce qui aurait d'ailleurs été
étonnant du fait de sa durée, plus de 4 Ma) semble être
caractérisé en réalité par des fluctuations
eustatiques bien marquées, que nous allons nous attacher à mettre
en évidence.
En plus de paramètres déjà
présentés précédemment (fig.12 et 14), la figure 18
donne deux autres indications : y sont indiqués les échantillons
contenant de la glauconie et ceux contenant des calcisphères.

1111 1111 1111 1111 1111
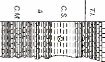
1 1111111111111111111111111111 1 1 1 1 11 1
1 1 1 1 11 1 1 1111 11 1 11 1 1 1 1 1 1 1 1 11
lei1111111111r111111111telll
11111111111111111k111111111111111III
1111111111111[11111111111111 111 1..!1 1
II I 1 1 11 1 1 1 11 11 1 11 1 1 111111 HI
en
11111111111111111111111111 11 1P-4 1 1 11 1 1 1 11 1 1 1 1 11 11
1 11 1 1 11r>111 111
1111111111111111111111111111 II 1 11 1 1 1 11 1 1 1 1
1 11 1 1 1 11111


A
Q
Q

O
ro
Co
Co
N
o
C)
0 0

·
·
·
0-
Pr
Figure 18 : Présentation des principaux rapports
de tri et des zones définies dans la coupe.
Les différentes zones définies sont donc
caractérisées comme suit :
· zones impaires :
proportion de foraminifères planctoniques
élevée à très élevée (plus de 80 %),
proportion d'ostracodes généralement très faible,
excepté dans un échantillon de la zone 7. Les ostracodes
rencontrés y sont presque tous de type lisse,
foraminifères benthiques de différents types :
domination des agglutinés (zones 1, 3) ou des calcaires (zones 5, 7),
foraminifères planctoniques carénés
quasi-absents,
glauconie présente uniquement dans la zone 7,
calcisphères abondantes en zones 1 et 7,
taux de CaCO3 variable, particulièrement faible en zones
3 et 5, plus élevé ailleurs, taux de COT variable :
généralement faible (0,1 à 0,4 %), il augmente dans
quelques niveaux lors de la transition avec le Turonien. Aucune augmentation
des valeurs n'est à noter dans les niveaux de la fin du Vraconnien,
malgré une couleur très sombre.
· zones paires :
proportion de foraminifères planctoniques plus faible (35
% en moyenne),
proportion d'ostracodes plus élevée, notamment
en fin de zone 4 et dans la zone 6. Cette dernière contient en outre une
proportion d'ostracodes ornés nettement plus importante,
variabilité des types de foraminifères benthiques
(calcaire/agglutiné), bien que les agglutinés dominent le plus
souvent,
foraminifères planctoniques carénés
détectables, mais toujours très rares, glauconie présente
en zones 2 (bien représentée) et 6 (rare),
calcisphères fréquentes,
taux de CaCO3 également très variable.
Par conséquent :
Les premières zones (impaires)
caractérisent un milieu profond, apparemment plus proche du talus que de
la plate-forme externe, tandis que les autres (paires) indiquent a priori un
milieu nettement moins profond, bien ancré sur la plate-forme
externe.
Ces résultats nous permettent de reconnaître la
succession de quatre cycles eustatiques, ce qui nous
amène à une interprétation d'ordre séquentiel. Les
zones définies correspondent en fait à des
séquences de dépôt.
Si on se réfère aux travaux de Vail et al.
(1977), ces séquences seraient donc des séquences de
3ème ordre (d'une durée de l'ordre du million
d'années), auxquelles se surimposerait une tendance transgressive
générale (séquences de grande ampleur, de l' et 2""
ordre). La figure 19 (page suivante) permet de détailler la succession
observée.
|
Alt.
(m)
|
Log
|
Niveau marin
|

Prismes Séq.
|
PHN
|
|
|
IV
|
|
IT
|
|
|
PBPF
|
|
|
PHN
|
|
|
IT
|
|
|
III
|
|
PBN
|
|
|
PBPF
|
|
|
PHN
|
|
|
IT
|
|
|
Il
|
|
|
PBN
|
|
|
PBPF
|
|
|
PHN
|
I
|

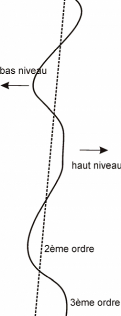
haut niveau
3ème ordre
V: Vraconnien, C.I./M./S. : Cénomanien
inférieur / moyen / supérieur, : Turonien
inférieur.
Figure 19 : Séquences de dépôt et
variations eustatiques au cours du Cénomanien.
PHN : Prismes de Haut Niveau, PBPF : Prisme de bordure de
plate-forme, IT : Intervalle transgressif, PBN : Prisme de bas
niveau.
Ces résultats se rapprochent de ceux obtenus par Herkat
(2004) sur des coupes situées dans les Aurès. L'auteur y
décrit en effet la succession de plusieurs mégaséquences
transgressives-régressives au cours du Crétacé
supérieur. La première d'entre elle, qui nous intéresse en
premier lieu, s'étend de l'Albien terminal au Cénomanien
supérieur et se compose, toujours selon Herkat (2004) de 4
séquences de 3ème ordre, constituées
d'alternances de marno-calcaires. Dans la partie centrale du bassin, l'ensemble
se termine par une surface durcie (`hard ground'), alors que dans la partie
distale les faciès pélagiques sont nettement mieux
représentés.
Comme dans ces travaux, notre étude
révèle donc la succession de 4 séquences au sein des
étages Albien terminal / Cénomanien, les formations
observées (essentiellement des marnes) semblant appartenir à la
partie distale du bassin (pas de calcaire à stromatolithes, de ride de
courant ou de trace de `hard ground'). Ce sont donc essentiellement les
variations observées dans les assemblages de microfossiles qui nous ont
conduit à proposer ce découpage.
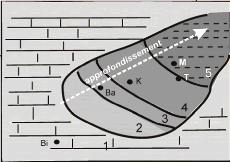
Par ailleurs, Herkat et Kechid-Benkherouf (2006) proposent une
reconstitution paléogéographique de la zone est-atlasique au
cénomano-turonien (fig. 20). Les auteurs y détaillent la
présence d'une paléo-rampe, subdivisée en
différents secteurs aux profondeurs croissantes vers l'est. Selon cette
carte, notre zone d'étude se situe dans un secteur de transition entre
rampe (id. plate-forme continentale) et bassin, ce qui correspond bien
à nos précédentes hypothèses.
36
35
/7-.111111MMZEM
-ariliMMMM
-.1
·01MMMM
MMM
//
M
·
·MMMMMMMUIP
Lmm
MMMMirr..- . .rolaMMMMMMMMIP"- MIMMMMMMMMM
OP- MMMMMMMMMMMMMMMMMMMMMM MMMMMMMMBEMMMMBMMMUIP-

MMMMMMMMMMMMUMMMMMMMIIIP-
MMMUMUMMMUMfit3iiMMMMMMP-
mmmumn FI tillf
·MEMM
MMIMMMIMIIIMIPMLIMMIUMMINZIMIMMIII
·""
MMMMMMIllierMdlIMIMMMMIIIIP'-
fflMMMir/AiiiiiMMMMIMMIlm"- MMZEZ
·MMMIMMM.P"--
IMMMMMMMM usP- IMMEMMMMMW IMMMM
·111
·""
Mffla
·MiMMIM10
·`'
IMMM1
·1
·M
·111"--

MM Iffl
=CP /KM
IMIZIGIM AMM
CIIMMiCliah, MMM
ei "le le lel a r .. Ang IL
.,5..,«
iMMIZ.111
'411M-MMIIMMEZEM.` AMIM%
smisammMMI -Iimow.-d=umilen
AmMIMMMIM

MWMMfflMMMM Wdellii=11=1W.IIMI s ./MM
OIMIMMMMale.a.M1h.111.111MIMEMZI_
iMM
·,<.M
IMMBIMEMBIEMBIrdi- manaiMIMM.,_ fflMMOY.IM
MOrAIMMU IM- MilEiIMZIMiSr
.IMMUlih.MM
-"MIMIMIC:157àà
A WAMMMILL.
-"IIIMISMIZIM
..IMÀfflMfliMM
IZOM
OMIMUMMMB=.1-ffln5MIMIIMMIegaZIMMMligirMiM
ime.nimmmummmmmmlIMIMIIF mmimmarezimm ÉMIIMINIONMBMIIMMOMM
Mainro
IMMWP1WWIMMIIMUMMMIEMMMe--M MMMMM:%à
·MMM
IMIMMMMMMMMMMMMMMMMMMMMMMMMMM
I MMM--M--MMMFM--MMMIMMUMMIMMMMMMMM
MMMMMMMMUUMMMUMEMMMMIMMMMMM



I II I1,
mua Plate-forma Mâlejammum

I MMMMMMMMIEMMMM3MMMMMMMMMM MMMMMMMMMMMMMMMMMMMMMMMMMM I
t M MM
rrrrrrrrrrrrrrrrrrrrr
MMMMMMMMMMMMMMMffluEMMIMMMMMMMMM
I- I - I -
- I
- -1 -
B
2 3 4 5
1
1 I I
6
7 8 0 9
Figure 20 : Reconstitution
paléogéographique du domaine est-atlasique au
cénomano-turonien (modifié
d'après Herkat et
Kechid-Benkherouf, 2006).
1 : Rampe proximale, 2 : Rampe médiane, 3 :
Rampe distale, 4 : Transition rampe-bassin,
5 : Bassin, 6 : Tell, 7 :
Failles, 8 : Failles supposées, 9 : Zone d'étude.
A : Carte
détaillée, B : Schéma synthétique.
| 

