c-2 -- Les foraminifères
Abondantes et diversifiées, les faunes de
foraminifères peuvent apporter de nombreuses informations :
l'étude des foraminifères planctoniques est
généralement très utile pour les interprétations
stratigraphiques, au contraire des foraminifères benthiques qui, de part
leur longévité généralement importante, ne sont le
plus souvent que peu significatifs de ce point de vue (Benkherouf, 1988),
nous rechercherons également à reconstituer les
paléoenvironnements en nous basant sur l'ensemble des données
acquises.
L'évolution de tous les paramètres cités
(ratio benthique / planctonique, diversité, proportion des
différents types d'organismes...) est présentée dans les
figures 12 et 14.
· Ratio benthique
/planctonique
Un peu plus de la moitié des
foraminifères triés (54%) sont des foraminifères
planctoniques, ce qui est compatible avec le chiffre avancé par
Benkherouf (1988) dans la région du Dj. Dyr (pourcentage estimé
à 50 %). La variabilité est grande cependant, entre des
assemblages nettement dominés par des foraminifères planctoniques
(plus de 60 % voire même jusqu'à 100 % des organismes), des
assemblages mixtes et d'autres enfin où les populations benthiques sont
majoritaires.
Nous distinguerons donc 3 types d'assemblages :
type 1 : population planctonique majoritaire (> 42 % des
échantillons),
type 2 : population mixte (;---' 18 %),
type 3 : population benthique majoritaire (z 40 %).
· Evolution de la diversité
spécifique (fig.12(c))
Classée en trois catégories, faible (< ou =
à 5), moyenne (< ou = à 10) ou grande (> 15), la
diversité est très variable selon les niveaux.
Généralement très faible dans les assemblages de type 1
(planctoniques), elle peut être nettement plus élevée,
notamment dans les assemblages de types 2 (mixtes), dans lesquels
jusqu'à 20 espèces ont pu être reconnues.
Le Cénomanien inférieur
débute par des assemblages de type 1, à diversité
très réduite, mais rapidement la diversité
s'accroît, pour atteindre les valeurs les plus élevées de
l'ensemble du Cénomanien.
La transition avec le Cénomanien moyen
est marquée par des assemblages pauvres, auxquels
succèdent des assemblages plus riches (jusqu'à 12 espèces
reconnues), mais la grande diversité du Cénomanien
inférieur n'est jamais égalée.
Enfin, le Cénomanien supérieur,
et notamment la transition avec le Turonien, est globalement
très pauvre en espèces. Il n'est pas rare qu'une à deux
espèces soient ultra- dominantes (aussi bien d'ailleurs des
espèces planctoniques dans les assemblages de type 1 que des benthiques
dans ceux de type 3).
· Richesse microfossilistique apparente
(nb microorganismes / g trié) (fig.120)
Elle reflète essentiellement la richesse plus ou moins
grande en éléments fins (< 63 gm). La moyenne de 450 individus
par gramme cache mal une très grande variabilité, avec deux
extrêmes très éloignés (de 32 à près
de 3000 individus par g !). Dans la zone du Dj.
Dyr, Benkherouf (1988) évoque quant à elle une
richesse microfossilistique (réelle) particulièrement
basse, de seulement 40 individus par gramme en moyenne.
Plusieurs remarques peuvent cependant être faites :
les niveaux caractérisés par des assemblages de
type 1 (planctoniques) sont généralement ceux où la
richesse apparente est la plus élevée. La proportion en
éléments fins doit donc y être beaucoup plus importante,
au contraire, les niveaux les plus riches en organismes
benthiques (foraminifères mais aussi ostracodes) sont globalement
beaucoup plus pauvres en espèces. Le corollaire logique est que la
proportion en éléments fins y est nettement moindre,
il existe malgré tout quelques exceptions à ces
deux constatations, notamment pour certains niveaux du Cénomanien
terminal, où la richesse apparente est basse, malgré une
proportion élevée en organismes benthiques.
· Foraminifères planctoniques :
variation et caractérisation
Avec plus de 99 % des individus observés, les
espèces globuleuses dominent outrageusement les assemblages de
foraminifères planctoniques (fig.14). Les genres
Globigerinelloidesb et Hedbergella sont parmi les
plus fréquents, accompagnés, à certains niveaux, par de
très nombreux Heterohelixc (30 % environ en moyenne,
mais jusqu'à plus de 90 % dans les niveaux de transition avec le
Turonien). Les espèces du genre Whiteinella, également
présentes, sont plus rares, de même que les Favusella
(voir en figure 13 les clés de détermination des principaux
genres rencontrés).
Ont été reconnues les espèces :
- Hedbergella planispira (TAPPAN, 1940), H.
delrioensis (CARSEY, 1926), H.
simplex (MORROW, 1934) [rare],
- Heterohelix moremani (CUSHMAN, 1938) [espèce
fréquente, et parfois accompagnée d'autres types
d'hétérohélicidés, nettement plus globuleux],
- Favusella washitensisd (CARSEY, 1926)
[espèce rare, Cénomanien inférieur],
- Whiteinella archeocretacea (PESSAGNO, 1967), W.
baltica (DOUGLAS et RANKIN, 1969), W.paradubia (SIGAL, 1952b)
[Cénomanien supérieur].
D'autres espèces ont très certainement
échappé à notre diagnostic (présence de
pseudo-Schackoines ?) et beaucoup ne sont absolument pas identifiables du fait
d'une trop mauvaise conservation.
On l'aura compris, si les espèces globuleuses sont
tellement abondantes, en contrepartie, celles carénées sont
particulièrement rares 1 (à peine 1 %). Seulement
1/4 des échantillons triés ont
révélé la présence d'espèces
carénées (du genre Rotaliporaa), par ailleurs
jamais très nombreuses (2 à 3 spécimens en moyenne dans
chacun des échantillons concernés). Il s'avère
malheureusement (pour nous !) que ce sont essentiellement des espèces
carénées qui sont utilisées pour les déterminations
biostratigraphiques, dans la période considérée en tous
les cas (fig.9, 16).
Seules les espèces Rotalipora appenninica
(RENZ, 1936) et R. brotzeni (SIGAL, 1948) ont pu être mises
en évidence de façon formelle. Les autres spécimens, a
priori des rotalipores également, n'ont pas pu être
déterminés plus précisément, à cause d'un
état de conservation largement insuffisant. L'espèce R.
cushmani (MORROW, 1934) notamment n'a pas été
identifiée avec certitude.
1
Dans la région du Dj. Dyr, Benkherouf (1988) parle
quant à elle de 20 % d'espèces carénées.
|
Morphologie du test
|
Type d'ouverture
du test
|
Présence
d'une
carène
?
|
Familles
|
Genres
|
Illustration - Exemples
|
|
Bi-sérié
|
Terminale
|
non
|
Heterohelicidae
|
Heterohelix
|
Heterohelix moremani
|
|
Planispiral é
(enroulement selon
un
plan)
|
O-E
|
non
|
Planomalidae
|
Globigerinelloides
|
Globigerinelloides blowi
1
|
|
Trochospiralé
(enroulement selon
un axe)
|
0*-E*, presque P*
+
lèvre
|
non
|
Globotruncanidae
|
Hedbergella
|
. Hedbergella
delrioensis
. _
|
|
O-E, presque P
+ lèvre
+
ouvertures
supplémentaires sur le
côté ombilicale
|
oui
|
Globotruncanidae
|
Rotalipora
|
e-
- - i e Rotalipora
,
· '- -. reicheli
r ' -
)) ti'Èef
4,- . '
|
|
O-E
+ orticus
p
|
non
|
Globotruncanidae
|
Whiteinella
|
Whiteinella archeocretacea
.
|
* O = Ombilicale, E = extraombilicale, P =
périphérique
Figure 13 : Clés de détermination des
principaux genres de foraminifères planctoniques
rencontrés.
· Foraminifères
benthiques
Avec plus de 40 espèces reconnues, les populations de
foraminifères benthiques sont très variées, mais leur
répartition n'est pas homogène tout le long du profil. En effet,
entre les niveaux inférieurs du Cénomanien à population
benthique riche et diversifiée, et les niveaux supérieurs
où le benthos se réduit à de rares espèces
dominantes, la différence est grande.
Dans ce qui suit, nous différencierons deux grands
types de foraminifères benthiques, les calcaires (BC) et les
agglutinés2 (BA), dont le mode de formation est radicalement
différent. 60 % environ des foraminifères benthiques sont
de type agglutiné (avec une teneur en BA > 70 % pour plus de
la moitié des échantillons) [fig.14]. Les foraminifères de
type calcaire sont donc beaucoup moins fréquents, et ne dominent
réellement que dans 1/4 environ des échantillons
triés, essentiellement dans des niveaux du Cénomanien
inférieur.
Essai de classification des individus
rencontrés :
(basé sur les classifications de Loeblich et Tappan
(1987, 1992), et sur les modifications ultérieures proposées par
Lee et al. (2000) et Kaminsky (2004) pour ce qui concerne les
foraminifères agglutinés)
0 Parmi les foraminifères agglutinés,
on retrouve :
Ammobaculites sp. (CUSHMAN), Ammobaculites turonicus
(SAID et KENAWY), Ammodiscus cretaceus (REuSS), Ammomarginulina ? sp.
(LOEBLICH et TAPPAN), Dorothia oxycona (REuSS), Eggerella sp.
(CUSHMAN), Gaudryinoides (GEODAKCHAN), Textularia chapmani
(LALICKER), Textularia rioensis (CARSEY), Thomasinella punica
(SCHLUMBERGER), Tritaxia pyramidata (REuSS), Trochamminoides sp.
(CUSHMAN)...
Les foraminifères benthiques agglutinés sont
essentiellement composés de Lituolidés
(majoritairement du sous-ordre des Lituolina),
accompagnés, à certains niveaux, par des
Textularidés en grand nombre.
Classification des organismes identifiés :
Classe FORAMINIFEREA d'Orbigny, 1826 Sous-classe
TEXTULARIIA Mikhalevich, 1980
Ordre ASTRORBIZIDA Lankester, 1885
Sous-ordre AM1VIODISCINA Mikhalevich, 1980
Super-famille AMMODISCACEA Reuss, 1862
Famille AMMODISCIDAE Reuss, 1862
Sous-famille AMMODISCINAE Reuss, 1862
Genre Ammodiscus Reuss, 1862
Ammodiscus cretaceus (Reuss, 1845) p1.5.1
|
Ordre LITUOLIDA Lankester, 1885
|
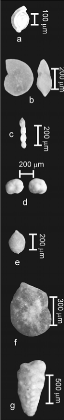
Photos a à e: benthiques calcaires
Photos f et g : benthiques agglutinés
|
|
2
Selon les auteurs, on trouvera agglutinés =
agglutinants = arénacés.
|
|
Sous-ordre LITUOLINA Lankester, 1885
Super-famille LITUOTUBACEA Loeblich et Tappan,
1984
Famille TROCHAMIVHNOIDAE Haynes et Nwabufo-Ene,
1998
Genre Trochamminoides Cushman, 1910
Trochamminoides sp. (Cushman, 1910) .p1.5.2 et 3 ?
Super-famille LITUOLACEA de Blainville, 1827
Famille LITUOLIDAE de Blainville, 1827
Sous-famille AMMOMARGINULINAE Podobina, 1978*
Genre
Ammobaculites Cushman, 1910
Ammobaculites sp. (Cushman, 1910) p1.5.4
Ammobaculites turonicus (Said et Kenawy, 1956)
..p1.5.5
Genre Ammomarginulina Wiesner, 1931
Ammomarginulina ? sp. (Wiesner, 1931) p1.5.6
Sous-ordre VERNEUILININA Mikhalevich et
Kaminski, 2004
Super-famille VERNEUILINACEA Cushman, 1911
Famille TRITAXIIDAE Plotnikova, 1979*
Genre Tritaxia Reuss, 1860
Tritaxia pyramidata (Reuss, 1863) p1.5.7
Famille REOPHACELLIDAE Mikhalevich et Kaminski,
2004
Sous-famille SPIROPLECTINATINAE Cushman, 1928
Genre Gaudryinoides Geodakchan, 1969*
Gaudryinoides sp. (Geodakchan, 1969)* p1.5.8
Ordre TEXTULARIIDA Delage et Herouard, 1896
Sous-ordre TEXTULARIINA Delage et Herouard,
1896
Super-famille EGGERELLACEA Cushman, 1937
Famille EGGERELLIDAE Cushman, 1937
Sous-famille DOROTHIINAE Balakhmatova, 1972*
Genre Dorothia Plummer, 1931
Dorothia oxycona (Reuss, 1860) p1.5.9
Sous-famille EGGERELLINAE Cushman, 1937
Genre Eggerella Cushman, 1935
Eggerella sp. (Cushman, 1935) .p1.5.14 ?
Super-famille TEXTULARIACEA Ehrenberg, 1838
Famille THoMASINELLIDAE Loeblich et Tappan,
1984
Genre Thomasinella Schlumberger, 1893
Thomasinella punica (Schlumberger, 1893) p1.5.12
Famille TEXTULARIIDAE Ehrenberg, 1838
Sous-famille TEXTULARIINAE Ehrenberg, 1838 Genre Textularia Defrance,
1824
Textularia chapmani (Lalicker, 1935) p1.5.10
Textularia rioensis (Carsey, 1926) p1.5.11
* ces différents genres, espèces ou familles sont
cités par Kaminsky (2004).

Planche 5 : Foraminifères benthiques
agglutinés.
1. Ammodiscus cretaceus
(REUSS), 2. et 3 ?.
Trochamminoides sp. (CUSHMAN), 4.
Ammobaculites sp. (CUSHMAN), 5.
Ammobaculites turonicus (SAID et
KENAWY), 6. Ammomarginulina ? sp.
(WIESNER), 7. Tritaxia pyramidata
(REUSS), 8. Gaudryinoides sp.
(GEODAKCHAN), 9. Dorothia oxycona
(REUSS), 10. Textularia chapmani
(LALICKER), 11. Textularia rioensis
(CARSEY), 12. Thomasinella punica (SCH
LU M BERGER), 13.indéterminé 1, 14. Eggerella ? sp.
(CUSHMAN).
(c) Les foraminifères benthiques calcaires
reconnus sont les suivants :
Cassidella sp.(KHAN) = Coryphostoma, Dentalina
sp. (RISSO), Gavelinella intermedia (BERTHELIN), Gavelinella
cenomanica (BROTZEN), Globulina prisca (REuSS), Gyroidinoides nitidus
(REuSS), Lagena apiculata (REuSS), Lenticulina rotulata (LAMARCK),
Nodosaria sp. (LAMARCK), Oolina sulcata (WALKER et JACOB),
Planularia complanata (REuSS), Quinqueloculina sp.(d'ORBIGNY),
Spiroloculina sp. (d'ORBIGNY), Tristix acutangulum (REuSS),
Tristix excavatum (REuSS), Valvulineria sp. (CUSHMAN).
Les organismes les plus fréquemment rencontrés sont
des Rotalidés, les autres genres et espèces
restant anecdotiques dans la plupart des niveaux.
Classification des organismes rencontrés :
Classe FORAMINIFEREA d'Orbigny, 1826
Ordre LAGENIDA Lankester, 1885
Super-famille NODOSARIACEA Ehrenberg, 1838
Famille NODOSARIIDAE Ehrenberg, 1838
Sous-famille NODOSARIINAE Ehrenberg, 1838
Genre Dentalina Risso, 1826
Dentalina sp. (Risso, 1826) p1.6.1
Genre Nodosaria Lamarck, 1812
Nodosaria sp. (Lamarck, 1812) p1.6.2 et 3
Genre Tristix Macfayden, 1941
Tristix acutangulum (Reuss, 1863) p1.6.7
Tristix excavatum (Reuss, 1863) p1.6.8
Famille LAGENIDAE Reuss, 1862
Sous-famille LAGENINAE Reuss, 1862
Genre Lagena Walker et Jacob, 1798
Lagena apiculata (Reuss, 1851) p1.6.9
Famille VAGINULINIDAE Ehrenberg, 1838
Sous-famille LENTICULININAE Chapman, Parr et Collins, 1934
Genre Lenticulina Lamarck, 1804
Lenticulina rotulata (Lamarck, 1804) p1.6.10
Sous-famille MARGINULININAE Wedekind, 1937
Genre Planularia Defrance, 1826
Planularia complanata (Reuss, 1845) p1.6.11
Super-famille POLYMORPHINACEA d'Orbigny, 1839
Famille POLYMORPHINIDAE d'Orbigny, 1839
Sous-famille POLYMORPHININAE d'Orbigny, 1839
Genre Globulina d'Orbigny, 1839
Globulina prisca (Reuss, 1863) p1.6.12
Famille ELLIPSOLAGENIDAE A. Silvestri, 1923
Sous-famille OOLININAE Loeblich et Tappan, 1961
Genre Oolina d'Orbigny, 1839
Oolina sulcata (Walker et Jacob, 1798) p1.6.13
Ordre MILIOLIDA Lankester, 1885
Sous-ordre MILIOLINA Delage et Hérouard,
1896
Super-famille MILIOLACEA Ehrenberg, 1839
Famille SPIROLOCULINIDAE Wiesner, 1920
Genre Quinqueloculina d'Orbigny, 1826
Quinqueloculina sp. (d'Orbigny, 1826) .p1.6.15
Genre Spiroloculina d'Orbigny, 1826
Spiroloculina sp. (d'Orbigny, 1826) p1.6.16
Ordre ROTALIIDA Lankester, 1885
Super-famille CHILOSTOMELLACEA Brady, 1881
Famille GAVELINELLIDAE Hofker, 1956 Genre Gavelinella Brotzen, 1942
Gavelinella intermedia (Berthelin, 1880) p1.7.1
Gavelinella cenomanica (Brotzen, 1945) p1.7.2
Genre Gyroidinoides Brotzen, 1942
Gyroidinoides nitidus (Reuss, 1845) .p1.7.3, 4 ( ?)
Genre Valvulineria Cushman, 1926
Valvulineria sp. (Cushman, 1926) p1.7.5
Super-famille BULIMINACEA Jones, 1875
Famille BOLIVINITIDAE Cushman, 1927
Sous-famille FURSENKOININAE Loeblich et Tappan, 1961 Genre
Fursenkoina (Loeblich et Tappan, 1961)
La détermination du genre observé est
particulièrement problématique, puisque sa dénomination
est variable selon les auteurs. On trouve ainsi :
Fursenkoina viscida (Khan) selon Holbourn et alii
(2001) p1.6.7
Cassidella viscida (Khan, 1950), selon Magniez-Jannin
(1975)
Coryphostoma? sp. selon Moullade et alii. (2008)
En plus de ces organismes, un certain nombre d'autres n'ont
pas pu être déterminés, dont des foraminifères, et
divers organismes (algues, calcisphères, mollusques ?? (etc)). Les
planches 5,6 et 7 en donnent plusieurs illustrations.
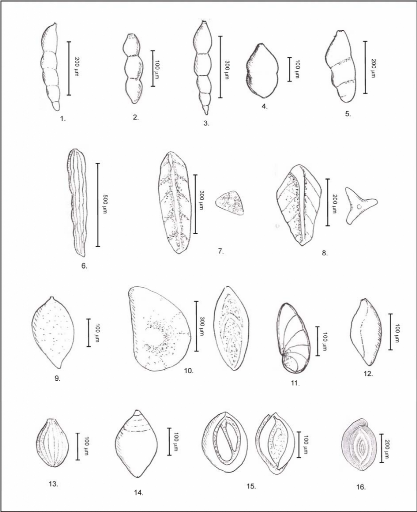
Planche 6 : Foraminifères benthiques
calcaires.
1. Dentalina sp. (RISSO), 2
et 3. Nodosaria sp.
(LAMARCK), 4.Pseudonosaria aff. Humilis?,
5.indéterminé 2, 6. Nodosaria aff.
Chapmani ?, 7. Tristix acutangulum
(REUSS), 8. Tristix excavatum (REUSS),
9. Lagena apiculata (REUSS), 10.
Lenticulina rotulata (LAMARCK), 11.
Planularia complanata (REUSS), 12.
Globulina prisca (REUSS), 13. Oolina
sulcata (WALKER et JACOB), 14.indéterminé 3, 15.
Quinqueloculina sp. (D'ORBIGNY), 16.
Spiroloculina sp. (D'ORBIGNY).

Planche 7 : Foraminifères benthiques calcaires
(suite).
1. Gavelinella intermedia (BERTHELIN), 2.
Gavelinella cenomanica (BROTZEN), 3.
Gyroidinoides nitidus (REUSS), 4.
Gyroidinoides sp. ?, 5. Valvulineria sp.
(CUSHMAN), 6.indéterminé, 7. Cassidella
(KHAN) = Cotyphostoma? sp.,
8.indéterminé 4 = Gubkinella ??,
9.indéterminé 5, 10.indéterminé 6,
11.indéterminé 7 = Bonetocardiella ??,
12.indéterminé 8, 13.indéterminé
9.
Le tableau qui suit détaille la répartition et
l'abondance des différents types de foraminifères benthiques
déjà cités ci-dessus (tab.2).
|
Organisme
|
Planche
|
Répartition
|
Abondance
|
|
Vrac. --
Cénomanien
inf.
|
Cénomanien
moy.
|
Cénomanien
sup.
|
|
Ammobaculites sp.
(CUSHMAN)
|
5.4
|
|
|
|
*
|
|
Ammobaculites
. 1 ,
turorucus (SAID et
KENAWY)
|
5.5
|
|
|
.(
|
*
|
|
Ammodiscus cretaceus 2(REUSS)
|
5.1
|
|
|
|
*
|
|
Ammomarginulina ?
sp. (LOEBLICH et TAPPAN)
|
5.6
|
|
|
|
***
|
|
Dorothia oxycona
(REuSS)
|
5.9
|
|
|
|
*
|
|
Eggerella ? sp.3
(CUSHMAN)
|
5.14
|
|
|
|
***
|
|
Gaudryinoides
(GEODAKCHAN)
|
5.8
|
|
|
|
*
|
|
Textularia chapmani
(LALICKER)
|
5.10
|
|
|
|
*
|
|
Textularia rioensis
(CARSEY)
|
5.11
|
|
|
|
**
|
|
Thomasinella punica4
(SCHLUMBERGER)
|
5.12
|
|
|
|
*
|
|
Tritaxia pyramidata
(REuSS)
|
5.7
|
|
|
|
*
|
|
Trochamminoides sp.
(CUSHMAN)
|
5.2, 3 ?
|
|
|
|
***
|
|
Cassidella
sp. . (KH AN) = Coryphostoma
|
7.7
|
|
|
|
**
|
|
Dentalina sp. (RISSO)
|
6.1
|
|
|
|
*
|
|
Gavelinella intermedia
(BERTHELIN)
|
7.1
|
|
|
|
**
|
|
Gavelinella cenomanica
(BROTZEN)
|
7.2
|
|
|
|
**
|
|
Globulina prisca
(REuSS)
|
6.12
|
|
|
|
*
|
|
Gyroidinoides nitidus (REUSS)
|
7.3
|
|
|
|
***
|
|
Lagena apiculata
|
6.9
|
|
|
|
**
|
|
(REuSS)
|
|
|
|
|
|
|
Lenticulina rotulata
(LAMARCK)
|
6.10
|
|
|
|
*
|
|
Nodosaria sp.
(LAMARCK)
|
6.2, 3
|
|
|
|
*
|
|
Oolina sulcata
(WALKER et JACOB)
|
6.13
|
|
|
|
*
|
|
Planularia complanata (REUSS)
|
6.11
|
|
|
|
*
|
|
Quinqueloculina sp.(d'ORBIGNY)
|
6.15
|
|
|
|
*
|
|
Spiroloculina sp. 5
(d'ORBIGNY)
|
6.16
|
.(
|
.(
|
.(
|
**
|
|
Tristix acutangulum
(REuSS)
|
6.7
|
|
|
|
*
|
|
Tristix excavatum
(REuSS)
|
6.8
|
.(
|
|
|
*
|
|
Valvulineria sp.
(CUSHMAN)
|
7.5
|
|
|
|
**
|
|
Indéterminé 1
|
5.13
|
|
|
|
*
|
|
Indéterminé 2
|
6.5
|
|
|
|
*
|
|
Indéterminé 3
|
6.14
|
|
|
|
*
|
|
Indéterminé 4
|
7.8
|
.(
|
|
|
*
|
|
Indéterminé 5 6
|
7.9
|
|
|
|
*
|
|
Indéterminé 6 6
|
7.10
|
|
|
|
*
|
|
Indéterminé 7
7
Bonetocardiella ??
|
7.11
|
|
|
|
*
|
|
Indéterminé 8
|
7.12
|
|
|
|
*
|
|
Indéterminé 9
8
|
7.13
|
|
|
|
*
|
Degré d'abondance : * rare ** commun ***
très fréquent
Tableau 2 : Détail de la répartition et de
l'abondance des foraminifères benthiques.
1 : Ammodiscus cretaceus est une
espèce rare. Seul un petit nombre de spécimens a
été observé dans le Cénomanien inf. (n°30) et
sup. (n°94).
2 : Ammobaculites turonicus a
été décrit uniquement à un niveau dans le
Cénomanien sup. (n°73).
3 : Le genre présenté comme
Eggerella (?) est particulièrement abondant à certains
niveaux du Cénomanien sup. Ses structures externes étant
particulièrement difficiles à distinguer, sa détermination
est difficile.
4 : L'espèce Thomasinella punica
n'est abondante qu'à deux niveaux du Cénomanien moyen
(n°52 et 54), et exceptionnelle ailleurs (n°36 du Cénomanien
inf.). Notez sa taille exceptionnellement grande pour un foraminifère
(plusieurs mm).
5 : Les foraminifères du genre
Spiroloculina constituent presque 100 % des formes observées
pour l'échantillon n°36 (également très riche en
ostracodes). Ailleurs, ils sont rares.
6 : Ces formes n'ont été
observées que dans de rares niveaux du Cénomanien moyen. Il ne
s'agit probablement pas de foraminifères (calcisphères (indet.5)
ou autre ?).
' : Cette forme, qui rappelle une calpionelle ( Albien) n'a
été observée que dans de rares niveaux du début du
Cénomanien inf. Il pourrait s'agit en réalité d'un type de
calcisphère, apparenté au genre Bonetocardiella
figuré par Robaszynski et al. (1993).
8 : Un seul échantillon du
Cénomanien moyen (n°52) a révélé de telles
formes, facilement repérables par leur grande taille, d'autant plus
qu'elles sont présentes en grand nombre au côté de
l'espèce Thomasinella punica. Là
encore, il ne s'agit probablement pas de foraminifères (mollusques
??).
La plupart des espèces décrites semble
caractériser des milieux de plate-forme externe ou de talus (Benkherouf,
1988). Aucune espèce franchement littorale3 n'a
été observée, pas plus que des espèces typiques de
milieux profonds.
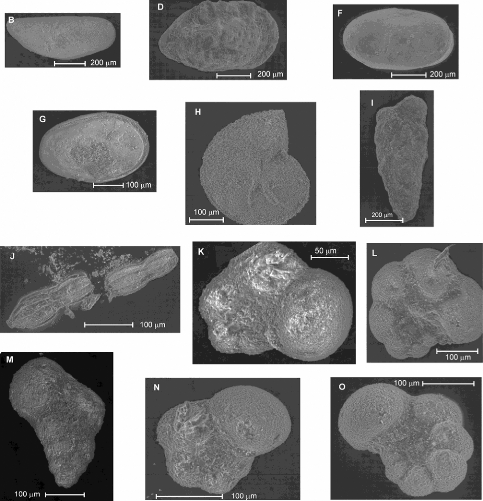
Page suivante, se trouve une planche (pl. 8) regroupant
quelques clichés de microfossiles pris au MEB, incluant des ostracodes
(photos A à G), des foraminifères benthiques, calcaires (H et J)
ou agglutinés (I) ; et d'autres planctoniques globuleux (K à O).
Ces quelques formes ne constituent qu'une faible proportion du total des formes
observées, et ont été sélectionnées
uniquement en rapport avec leur bon état de conservation.
Légende de la planche 8
:
A et B : Spécimens apparentés
à Paracypris sp.(SARS), vue latérale. C
: Procytherura ? sp. (WHATLEY), vue
latérale.
D : Spécimen appartenant à la
famille des Trachyleberididae, vue latérale.
E : Idem, vue dorsale.
F : Cytherella sp. (JONES),
vue latérale droite.
G : Cytherella sp. (JONES),
vue latérale gauche.
H : Lenticulina sp.
(LAMARCK).
I : Textularia sp.?
(DEFRANCE).
J : Nodosaria sp.
(LAMARCK).
K et L : Hedbergella sp.
(BRONNIMANN et BROWN)
M : Heterohelix moremani
(CUSHMAN).
N: Hedbergella sp. (BRONNIMANN
et BROWN)
m : Whiteinella sp.
(PESSAGNO)
3
au niveau du Dj. Dyr, Benkherouf (1988) a décrit deux
espèces considérées comme allochtones, à milieu de
vie plutôt
superficiel, à savoir Charentia cuvillieri et
Archaecyclus cenomaniana., Pour notre part, ces espèces n'ont
pas été observées.
G
|
B
|
e
|
w7
|
.. .- ... " ;" '-. , '' ' .
|
|
|
i_ ii 200um
|
|
1 .
|
Fele
|
1-.1 200um
1-1 200 gm
Planche 8 : Illustrations de quelques microfossiles au
MEB.
(clichés pris au laboratoire de Tautavel,
Université de Perpignan, France)
En figure 14 (page suivante) sont présentés divers
paramètres (obtenus à l'issu du tri des microfossiles), dont
certains méritent une explication :
0 Pourcentage de foraminifères benthiques (B),
planctoniques (P) et d'ostracodes (0).
(c) Proportion de foraminifères benthiques (B)
et de foraminifères benthiques de type agglutiné
(BA).
G Proportion de foraminifères planctoniques
(P) et de foraminifères planctoniques de type Heterohelix (PH)
:
Ce type de foraminifères globuleux domine parfois
à certains niveaux, notamment en toute fin
du Cénomanien
où ils constituent la presque totalité des foraminifères
planctoniques observés.
(c) Rapport (benthique agglutiné (BA) /total
des benthiques (B)) :
Comme en (c), la prédominance de ce type de
foraminifères parmi les organismes benthiques apparaît nettement
(plus de 65 % du total). Les foraminifères benthiques de type calcaire
ne dominent qu'à de rares niveaux.
G Rapport (planctonique globuleux (PG) /total des
planctoniques (P)) :
Ce type de foraminifères planctoniques domine très
nettement face à ceux de type caréné, extrêmement
rares.
(c) Rapport (planctonique/total des
foraminifères) :
L'évolution en dents de scie montre une
variabilité importante, avec des niveaux nettement dominés par
des foraminifères planctoniques (ceux-ci dominent nettement à 4
reprises au moins) et d'autres au contraire par des foraminifères
benthiques.
(c) Rapport (foraminifères total (F)/
ostracodes (0)) :
Généralement rares, les ostracodes
représentent parfois, comme on le voit, la majorité des
microorganismes observés.
Chap. III : Résultats et interprétations

rn
O
fn
o
2
o
O
0,
O O
|
113
|
|
P 1
,..
0 ,
il II
Il II
11 .I.
I
|
|
k Ili ill I.L I i I lui
,00,,,,Iimilpoliiii,[iiiiii,Fili
iiii, i ,,I[1111[111111[11111111111111111
Ill 111/I11[111111111111111D111[111
11111111[1111[1111 I 1 1111 1 1 r
111
|
l
:i
|
i,. l'el
il
II
'I!
|
ij
Ii
il
Il
I '
|
Hu
1
!
|
ii 1 II
, [Hum
HI
[ 11 1> 11111
r1111111
1111111
|
r
l'
1' 1111
|
itli$
ri
il g
I
1 I
lill
y.
'MN
|
|
|
l
|
!
|
|
|
_
_
_
_
_
|
|
|
_
_
-
-
_
|
|
|
|
_
_
|
|
to
>
|
|
\
|
|
|
_
_
-
_
|
|
|
|
|
|
_
|
|
|
|
|
|
|
|
|
_
|
|
_
|
_ É
in--.
_-
·.-__,_
|
-
-
|
|
|
|
|
_
_
|
1
|
..
-
-
|
|
_
_
_
_
_
_
|
----- ---_______._.----------- \ -_ 1
. 1
|
- _ - -
|
cs) O
1
0
w 0
-n
o
O
É 1 i l 3 e
Figure 14 : Log stratigraphique et paramètres de
tri (suite).
| 

