II. MATERIEL ET
METHODES
II.1. Site d'étude
Le foyer de la maladie du sommeil de Campo (2 O 20'N, 9 O
52'E) est situé à 80 Km de la ville de Kribi dans la
région du Sud Cameroun (figure 8). C'est un foyer hypo-endémique
de THA où aucune évolution épidémique de la maladie
n'y a été signalée depuis plusieurs années
(Melachio et al., 2011) . Le foyer de la THA de Campo se trouve le
long de la côte Atlantique et s'étend le long du fleuve Ntem qui
constitue la frontière entre le Cameroun et la Guinée
Equatoriale. Le climat est de type équatorial avec quatre saisons. La
végétation est constituée d'une forêt dense
équatoriale moins dégradé avec des terres agricoles, des
zones marécageuses et des marais le long de la côte (Simo et
al., 2014). Il s'agit d'une zone de forêt équatoriale avec un
réseau hydrographique dense, des zones marécageuses et des
mangroves. Cette zone est riche en faune sauvage et fait partie du parc
national de ?Campo Ma'an? (Njiokou et al., 2004a). L'agriculture, la
pêche et la chasse sont les principales activités menées
dans cette localité.
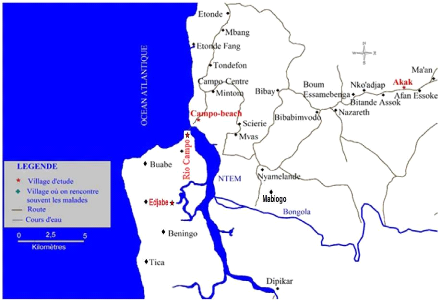
Figure 8 : Carte du Foyer de la THA de Campo
(Institut National de Cartographie Yaoundé Cameroun,
1976).
24
Caractérisation génétique de Glossina
pallicera pallicera circulant dans le foyer de la maladie du sommeil de
Campo du Sud forestier Camerounais rédigé par GOMSEU DJOUMSIE
Emmanuel Boris
II.2. Echantillonnage
Les glossines utilisées dans le cadre de cette
étude ont été capturées dans deux villages de Campo
(Akak, campo Beach) et deux villages limitrophes de la Guinée
Equatoriale (Edjabé et Rio campo) du 29 mars au 12 avril 2012 en
utilisant des pièges pyramidaux. Quelques glossines provenant de Fontem
(Région du Sud-Ouest, Cameroun) et Dodéo (Région de
l'Adamaoua, Cameroun) ont été utilisées pour des
comparaisons génétiques. Lors de la prospection entomologique,
des pièges pyramidaux (Gouteux et Lancien, 1986) ont été
installés dans des biotopes (les points d'eau, les puits et les points
ombragés, les bords des routes ou des pistes) favorables aux mouches
tsétsé. Les pièges étaient installés pendant
quatre jours consécutifs et les glossines étaient
collectées une fois par jour.
Une fois les mouches tsétsé collectées,
chacune d'elles a subi une identification morphologique pour déterminer
l'espèce et le sexe. Après cette identification, les pattes de
chaque glossine ont été enlevées et introduites dans des
microtubes contenant de l'alcool à 95° et la carasse
conservée dans de l'éthanol 95° pour d'autres études
ultérieures. Les microtubes ont été conservés sur
le terrain à la température ambiante. Une fois au laboratoire,
ces microtubes ont été conservés à -20°C
jusqu'à l'extraction d'ADN.
II.3. Extraction de l'ADN
L'ADN a été extrait de 6 pattes de glossines en
utilisant le CTAB 5% (Cetyl Trimethyl Ammonium Bromide) selon la méthode
décrite par Navajas et al. (1998). Brièvement, les tubes
contenant les pattes des glossines ont été sorties du
congélateur, puis laissés ouverts sur la paillasse toute la nuit
dans le but de faire évaporer l'alcool contenu dans les tubes.
Après l'évaporation complète de l'éthanol, 600 ul
de la préparation du CTAB (composition en annexe 1) ont
été ajoutés, puis les pattes ont été
broyées à l'aide des pistons. Le mélange obtenu a
été incubé pendant une heure au bain marie. Ensuite, 600
ul d'un mélange de chloroforme et d'alcool isoamylique (24/1, v/v) ont
été ajoutés dans chaque tube. Après une
homogénéisation lente pendant 10 minutes, l'ensemble a
été centrifugé à 8000 trs/min pendant 10 minutes.
La phase supérieure (phase aqueuse) contenant les acides
nucléiques a été transférée dans un nouveau
tube où 430 ul d'isopropanol ont été ajoutés afin
de précipiter les acides nucléiques. L'ensemble a
été homogénéisé puis incubé à
-20°C pendant une heure pour faire précipiter l'ADN. Une fois cette
incubation terminée, les tubes ont été centrifugés
à 13000 trs/min pendant 10 minutes et le surnageant a été
vidé en conservant le culot d'acides
25
Caractérisation génétique de Glossina
pallicera pallicera circulant dans le foyer de la maladie du sommeil de
Campo du Sud forestier Camerounais rédigé par GOMSEU DJOUMSIE
Emmanuel Boris
nucléiques (extrait d'ADN). Ce culot a
été lavé avec 900 ul d'éthanol 70°C et
centrifugé à 13000trs/min pendant 10 minutes. L'éthanol a
été entièrement vidé et les tubes contenant le
culot d'acides nucléiques ont été laissés ouverts
sur la paillasse pendant deux heures afin d'éliminer alcool
résiduel. Le culot a été suspendu dans 50 ul d'eau et
conservé à -20 °C pour les analyses moléculaires
ultérieures.
II.4. Analyses moléculaires II.4.1. Principe de la
PCR
La PCR est une technique moléculaire permettant
d'amplifier in vitro une région spécifique d'un acide
nucléique donné afin d'en obtenir une quantité
exponentielle de copies, suffisante pour être détectée et
étudiée. Au cours des réactions PCR, les produits obtenus
à la fin de chaque cycle servent de matrice pour le cycle suivant.
Chaque cycle comprend trois étapes:
? La dénaturation des doubles brins d'ADN à
température élevée (90-98°C).
? L'hybridation des amorces qui se fait à une
température définie selon la nature des amorces (cette
température varie généralement de 40 à 65°C).
Au cours de cette étape, les amorces qui sont des
oligonucléotidiques se lient, par appariement des bases
complémentaires au moyen des liaisons hydrogènes, à la
séquence cible à copier.
? L'élongation à 72°C, qui est la
température d'activité optimale de la Taq ADN polymérase.
Au cours de cette étape, les brins complémentaires d'ADN sont
synthétisés à partir des extrémités 3'OH
libre des amorces hybridées (Borde, 2006). La figure 9 ci-dessous
récapitule les différentes étapes de la PCR.
26
Caractérisation génétique de Glossina
pallicera pallicera circulant dans le foyer de la maladie du sommeil de
Campo du Sud forestier Camerounais rédigé par GOMSEU DJOUMSIE
Emmanuel Boris
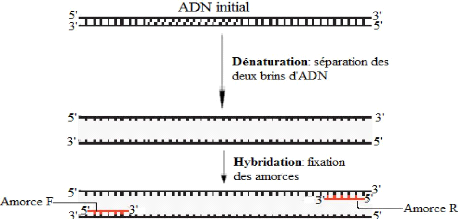

Figure 9: Différentes étapes de la PCR
(Jackson et al., 2006)
| 

