Epidémiologie, diversité et pathogénicité de la cercosporiose noire à l'est de la rdcongopar DIEUMERCI ROOSEVELT Université évangélique en Afrique - Ingénieur Agronome 2017 |

Introduction
Cultivés dans le monde entier, les bananiers sont menacés par diverses maladies et ravageurs, dont les attaques les plus dangereuses sont celles des champignons (Mycosphaerella fijiensis Morelet), et Fusarium oxysporum f. ssp. Cubense, des virus (Banana Bunchy Top Virus (BBTV), les nématodes (Helicotylenchus mulicinetus, Pratylenchys speijeri n. ssp. Meloidogyne spp. et Radopholus similis), des bactéries (Xanthomonas campestris pv. musacearum, certaines lignées 2 pathogènes de Pseudomonas solanacearum) ou d'insectes (le charançon Cosmopolites sordidus) (Mourichon et al., 2000 ; Jones, 2000 et Onautshu et al., 2013). La Cercosporiose noire du bananier ou maladie des raies noires causée par un champignon ascomycète M. fijiensis, constitue la maladie la plus destructrice du bananier plantain (Onautshu et al., 2013), et elle s'étend à toutes les régions de culture (Meredith et Lawrence, 1970 et Wilson, 1987). La maladie des raies noires est plus disséminée dans le monde et évolue rapidement chez les variétés déjà sensibles à M. musicola (Onautshu et al., 2013). Ainsi dit, elle induirait des pertes des rendements de 76 % et la réduction de la qualité des fruits surtout pendant le second cycle de production, tandis que les effets combinés de la maladie, des ravageurs et du déclin de fertilité du sol réduirait le rendement de 93 % (Ramsey et al., 1990 ; Mobambo et al.,1996 ; Damme, 2008 ; Chillet et al., 2009 ; Daniells, 2009 et Kassi et al., 2014). Au Sud-Kivu, elle cause des dégâts plus importants chez le bananier et le bananier plantain. Elle attaque uniquement le feuillage et provoque la réduction de l'activité photosynthétique de la plante conduisant à la baisse des rendements (Byenda, 2015). Combiné à d'autres facteurs et maladies, notamment l'absence de semences de qualité, la sensibilité des variétés disponibles à certaines maladies telles que la cercosporiose et la rosette (Mobambo et al., 1996a ; Martinez et al., 2000 ; Chausse et al., 2012 et Bizimana et al., 2012), le flétrissement bactérien du bananier ou Banana Xanthomonas wilt (BXW), la fusariose, le bunchy top, le charançon du bananier, les fourmis prédatrices et les nématodes constituent un problème dans la production bananière (Tushemereirwe et al., 2003; Youdeowei, 2004 et Byenda, 2015). Beaucoup de travaux ont été effectués l'état phytosanitaire du bananier dans la région et au cours desquels plusieurs maladies ont été détectées (le bunchy top (BBTV), le wilt bactérien du bananier les cercosporioses ainsi que l'helminthosporiose (Byenda, 2015). Malgré ces travaux, peu d'études ont été orientées vers l'évaluation de la distribution de la cercosporiose noire et la caractérisation des isolats de M. fijiensis dans la région ainsi que fournir l'information sur la sensibilité des génotypes cultivés. La gestion durable de cette maladie, nécessite une évaluation épidémiologique de la cercosporiose noire dans la région ainsi qu'une caractérisation des isolats de l'agent pathogène qui sont en circulation. 3 Quelques questionnements sont à considérer : > Quel serait l'incidence et la sévérité de la cercosporiose noire le Walungu et Kabare ? > Quelles seraient les diversités des isolats de M. fijiensis dans le Sud-Kivu comparativement aux populations de M. fijiensis trouvées dans le monde ? > Quelle devrait être la pathogénicité de ces différents isolats de M. fijiensis sur quelques nouvelles variétés de bananier ?
4.1. Objectif général L'objectif général de ce travail est l'étude de la diversité de la cercosporiose noire du bananier dans le territoire de Walungu et Kabare en vue de proposer les stratégies de contrôle et de gestion de la maladie. 4.2. Objectifs spécifiques Spécifiquement notre travail a comme objectifs : > Déterminer l'incidence et la sévérité de la cercosporiose noire du bananier et bananier plantain dans les territoires de Kabare et Walungu. > Evaluer les caractéristiques phénotypiques des différents isolats de M. fijiensis retrouvés dans les deux territoires par rapport à ceux rapportés dans le monde. > Tester la pathogénicité des isolats de M. fijiensis identifiés au Sud-Kivu. 4 Chapitre 1. Généralités sur le bananier 1.1. Le bananier 1.1.1. Origine, description et classification Le bananier est une culture originaire de l'Asie du Sud-Est, où il est cultivé depuis près de 10000 ans (Raemaekers, 2001 ; Abadie et al., 2003 ; Kouassi, 2005 ; Lassois et al., 2009 ; Bizimana et al., 2012). La Malaisie ou l'Indonésie parait être le centre probable de diversification (Daniells et al., 2001 ; Lassois et al., 2009). Le bananier cultivé est une plante herbacée vivace, géante et monocotylédone appartenant à l'ordre des Zingibérales ou scitaminales et à la famille des Musaceae et au genre Musa (Lassois et al., 2009). Botaniquement, le genre Musa est divisé en espèces séminifères possédant des fruits non comestibles et en variétés à fruits charnus sans graines (fruits parthénocarpiques). Dans la section Eumusa, Musa acuminata (symbole de génome : A) et Musa balbisiana (symbole de génome : B) sont des espèces sauvages qui sont à l'origine des variétés cultivées (Abadie et al., 2003 ; Cirad-Flhor, 2003 ; Lassois et al., 2009). Les espèces sauvages non comestibles, aux fruits à graines, servent à d'autres fins que l'alimentation humaine (fibre, alimentation du bétail, etc.). Elles sont toutes diploïdes (AA et BB) (Abadie et al., 2003 ; Cirad-Flhor, 2003 ; Lassois et al., 2009). Tandis que les variétés cultivées à présent sont en général des clones triploïdes stériles et aspermes (AAB et ABB), provenant soit des croisements interspécifiques entre les deux espèces séminifères sauvages diploïdes principales dont Musa acuminata et M. balbisiana, soit de la seule M. acuminata (AAA) (Lassoudière, 2007 et Lassois et al., 2009). 1.1.2. Description morphologique a) L'appareil végétatif Le bananier est une plante monocotylédone, elle constitue une herbe géante et vivace chez laquelle le pseudo-tronc est constitué par l'emboitement des gaines foliaires (Champion, 1963). Le méristème terminal de la tige vraie souterraine appelé improprement « bulbe » émet les feuilles (Bakry et al., 1997). 5 Le bananier peut atteindre une hauteur allant de 1,5 à 6m en Afrique (Raemaekers, 2001). Un bananier adulte est constitué d'un cormus (partie souterraine ou vraie tige du bananier) avec racines et rejets, d'un pseudo-tronc ayant de feuilles et un régime des fruits (Raemaekers, 2001). Chez le bananier, il y a deux sortes de bourgeons ; le bourgeon apical et les bourgeons latéraux. Le bourgeon apical disparait lorsque le méristème apical se transforme en inflorescence, tandis que les bourgeons latéraux deviennent des rejets, qui constituent en réalité les branches du tronc principal. Etant donné que ces rejets deviennent des nouvelles plantes à fruits, il est considéré comme plante vivace (Raemaekers, 2001 et Dhed'a et al., 2011). Ce sont les premières rangées de fleurs, appelées mains, qui forment les régimes de fruits. Ces premières constituent les fleurs femelles (Lassois et al., 2009). Après les fleurs femelles, il y a deux à trois mains de fleurs neutres qui apparaissent avec toutes les pièces florales avortées, suivies par les mains de fleurs mâles composées d'ovaires réduits et d'étamines bien développées (Lassois et al., 2009). 1.1.3. Ecologie du bananier Les bananiers et bananiers plantains se rencontrent dans des zones agro-écologiques se situant entre 30° latitude Nord et Sud (Dhed'a et al., 2011 et Onautshu et al., 2013). a) Le climat Le bananier, est une culture exigeante en eau. Les racines n'absorbant aisément que le tiers de la tranche dite habituellement utile. En climats chauds et humides, les besoins de la plante sont couverts lorsque les précipitations atteignent 35 mm de pluie par semaine, soit en moyenne 2500 mm de pluie repartie au cours de l'année (Dhed'a et al., 2011 ; Onautshu et al., 2013). La grande particularité du bananier est qu'elle est une herbe très riche en eau, considéré dans son ensemble, il est composé à plus de 90% d'eau (Lassoudière, 2007). La température optimale est comprise entre 25 et 30°C, au-delà de 35 à 40°C et en deçà de 14°C, des anomalies surviennent, la croissance des feuilles est retardée (Swennen et Vuylsteke, 2001 ; Lassoudière, 2007 ; Dhed'a et al., 2011 et Onautshu et al., 2013). Le bananier supporte des fortes insolations si l'approvisionnement en eau est suffisant ; elle est une culture des jours longs facultatifs. Le seuil limite pour les heures d'insolation est de 1500 à 1800 heures, mais 6 2000 à 2400 heures sont favorables. Le manque de lumière agit donc sur la hauteur des plants et peut faire « filer » les rejets (Dhed'a et al., 2011 et Onautshu et al., 2013). Le vent intervient sur la culture par l'incidence de sa vitesse sur le feuillage par lacération de ce dernier jusqu'à la destruction des cultures (Lassoudière, 2007) ; des pertes sont dues par des vents violents, soit en brisant les feuilles aux pétioles, soit en cassant les pseudos troncs (Dhed'a et al., 2011). b) le sol Les sols pour l'implantation des bananeraies sont très variés dans le monde. Les sols profonds, limoneux et bien drainés conviennent mieux pour la culture du bananier ; le N, P, K, Ca et Mg sont les éléments indispensables pour un bon développement et une production élevée, ces éléments sont apportés sous forme d'engrais organique comme le fumier, le compost, les engrais biologiques et les déchets agroindustriels (Laprade et Ruiz, 1999 ; Raemaekers, 2001). Un pH variant entre 4,0 et 8,0 est nécessaire pour la bonne croissance et le bon développement des plantes (Raemaekers, 2001). La croissance et le rendement sont fonction de la fertilité des sols (Swennen et Vuylsteke, 2001). 1.2. La cercosporiose noire du bananier 1.2.1. Son origine et sa distribution La cercosporiose noire ou maladie des raies noires a été identifiée pour la première fois en 1963 sur la côte sud-est de Viti Levu (îles Fidji) (Mourichon et al., 1997). Un champignon ascomycète, est à l'origine de la maladie de cercosporiose noire (Onautshu et al., 2013). Elle est considérée comme la maladie la plus dévastatrice des bananiers (Lassoudière, 2007). Avec le temps, la cercosporiose jaune s'est vu rapidement remplacée par la cercosporiose noire en Asie et en Amérique centrale, en raison de sa virulence plus grande et du spectre d'hôtes plus large (Onautshu et al., 2013). En Afrique, la maladie des raies noires a été signalée pour la première fois en Zambie en 1973 (Onautshu et al., 2013). Elle a été signalée en R.D.C par Mourichon en 1986. A Kinshasa, des études de l'épidémiologie ont été réalisées (Mobambo, 2002) et à Kisangani (Onautshu, 2007) ont confirmé la présence de cette maladie en R.D.C. 7 1.2.2. Symptômes Il est souvent difficile de donner les différences entre les symptômes de la cercosporiose noire et celle de la cercosporiose jaune. Sur la face supérieure du limbe apparait le premier symptôme sous une forme de tiret jaune pâle pour la cercosporiose jaune ou marron foncé sur la face inférieure du limbe de 1 à 2mm de long pour la cercosporiose noire (Mourichon et al., 1997). Elle affecte plusieurs cultivars résistants à la cercosporiose jaune, tels ceux du sous-groupe des bananiers plantains (AAB). La diminution des rendements peut aller dans certains cas, à plus de 50 % (Mourichon et al., 1997).
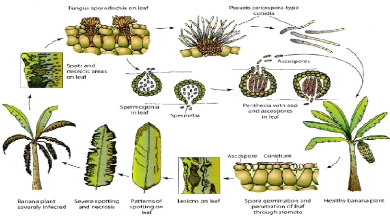
Fig. 1 : Stades de développement de la cercosporiose noire en champs d'après Fouré (1982). A= Stade 1 : Décolorations et ponctuations brunes de moins de 0,5 mm sur la surface inférieure de la feuille ; B = Stade 2 : Raies brunes rouilles inférieures à 4 mm et visible sur les deux faces ; C = Stade 3 : Raies allongées et élargies ; D = Stade 4 :Taches brun-noir elliptiques ; E = Stade5: Taches brun-noir entourées d'un halo jaune ; F = Stade 6 : Taches desséchées virant au gris avec en son centre des points noirs qui correspondent aux fructifications du pathogène (Onautshu et al., 2013). 1.2.3. Biologie, classification et diversité génétique de M. fijiensis Mycosphaerella fijiensis Morelet est l'agent causal de la maladie des raies noires du bananier. C'est un champignon qui appartient au Phylum des Ascomycota, dans la classe des Dothideomycètes, Sous-classe des Dothideomycetidées, Ordre des Capnodiales, Famille des Mycosphaerellaceae, Genre Mycosphaerella (Onautshu et al., 2013). Létude des populations pathogènes en utilisant de marqueurs génétiques montre l'existence d'une diversité génétique très importante notamment chez M. fijiensis (Carlier et al., 1994). Il existe deux caractéristiques biologiques qui doivent avoir un effet déterminant sur la structure de ces parasites : 8 ? la présence d'une forme parfaite, hétérothallique, très développée; ce facteur est connu pour être un facteur important de diversité; ? Les capacités de dispersion élevées, augmentant d'autant les flux de gènes entre populations (Mourichon, sd). Champignon hétérothallique, M.fijiensis se reproduit de façon sexuée et asexuée. La forme asexuée (anamorphe) est appelée Paracercospora fijiensis et celle sexuée (téléomorphe) est appelée Pseudocercospora fijiensis. Il a une croissance qui est lente en culture in vitro. Le diamètre de croissance d'une culture issue d'une seule conidie est presque 1cm après 38 jours d'incubation à 26°C (Meredith, 1970). Pour une bonne croissance, la température optimale est dans les intervalles de 24-28°C tandis que celle de la germination optimale est aux environs de 26°C. De 10 à 21 jours d'incubation, les colonies peuvent produire des conidies (Onautshu et al., 2013). L'observation au microscope montre que ce champignon présente un conidiophore plus ou moins septé (0 à 5 septa), droit ou courbé souvent renflé à la base (jusqu'à 8 um de diamètre), mesurant 16,5-62,5 um de long et 4 -7 um de large. Les conidies présentent une forme de massue, droites ou incurvées, pointues à l'apex, tronquées ou arrondies à la base, et peuvent avoir jusqu'à 10 cloisons (généralement 5 à 7 cloisons), mesurant 30-132 um de long et 2,5-5 um de large. Pour la production de la forme sexuée (téléomorphe), le champignon développe des spermogonies qui vont spécifier des spermaties (en forme de bâtonnet de 2,5-5 x 1,0-2,5 um de large). Il s'en suit la formation des périthèces (47-85 um) dans lesquels se spécifient les asques et les ascospores (11.5-16.5 x 2.5-5.0 um) (Onautshu et al., 2013).
Fig. 2 : Conidiophore et conidies (A) ; Spermogonie et spermaties (B) ; périthèce, asques et ascospores (C) de M. fijiensis (Onautshu et al., 2013). Génétiquement, la structure des populations de M. fijiensis provenant des différentes régions de la planète a été étudiée avec les marqueurs RFLP (polymorphisme de longueur des fragments 9 de restriction) ; ces études prouvent qu'il existe un niveau élevé de variabilité génétique entre les isolats de Philippines et de la Papouasie Nouvelle Guinée (Carlier et al.,1994 et Onautshu et al, 2013). 1.2.4. Etude épidémiologique de la cercosporiose noire Les ascospores et les conidies de M. fijiensis forment le potentiel d'inoculum qui est à la base des épidémies dangereuses. La propagation de M. fijiensis sur de longues distances est assurée par les ascospores qui sont chez cette espèce la principale source d'inoculum. Les différentes étapes de l'infection (pollution, germination, développement épiphylle, pénétration stomatique) sont sous le contrôle de conditions abiotiques (vent, rosée, pluie, évaporation, température) (Meredith et Laurence, 1970). 1.2.5. Cycle d'infection de M. fijiensis M. fijiensis a un cycle infectieux qui soit haplobionte et comprenant quatre phases: l'infection, l'incubation et latence correspondant au début de la colonisation des tissus, la sporulation suivie de la formation de propagules infectieuses, et la propagation de l'inoculum secondaire (Onautshu et al., 2013).
Fig. 3 : Le cycle infectieux de M. fijiensis (Carlier, 2010). Différemment des conidies, les ascospores sont constituées dans des pseudothèces se trouvant sur les feuilles vieilles des bananiers ou bien sur les feuilles mortes posées à même le sol. Le 10 mode de dissémination des ascospores est le vent et sont lancées violemment suite au desséchement du périthèce elles sont donc responsables de la dispersion à une longue distance. Mais les conidies sont disséminées par les pluies. En ce qui concerne les infections, les ascospores ou les conidies donnent le même type de taches et de développement subséquent de la maladie (Onautshu et al., 2013). Pour la germination des ascospores de M. fijiensis, une température variant 10 et 38°c, mais l'optimum de 27°C avec un minimum de 20°C (Perez, 1996). 1.2.6. Sensibilité des variétés Trois grands types de comportements vis-à-vis de la maladie caractérisent l'ensemble des variétés étudiées que l'on peut classer en trois catégories bien distinctes :
1.2.7. Moyens de lutte contre la MRN Pour la lutte contre la MRN, plusieurs techniques de lutte sont utilisées telles que la lutte biologique, la lutte chimique (les fongicides systémiques), les pratiques culturales, mais également l'utilisation des variétés améliorées. Pour une lutte intégrée de la cercosporiose noire, toutes ces méthodes sont utilisées simultanément dans différentes régions de la production des bananes (Mourichon et al., 1997, Orozco-Santos et al., 2011 et Onautshu et al., 2013). Il a été utilisé pour la détermination du positionnement global du milieu d'étude et son niveau d'élévation. 11 Chapitre 2. Milieu d'étude, matériels et méthodes 2.1. Milieu d'étude Le travail s'était déroulé dans les territoires de Walungu et Kabare. Ces deux territoires se situent dans le Sud-Kivu à l'Est de la R.D.Congo ; où une enquête a été réalisée et la récolte des feuilles présentant les symptômes de la MRN sur lesquelles les isolats de M. fijiensis ont été obtenus. La caractérisation des isolats de M. fijiensis s'était réalisée dans le laboratoire de biologie moléculaire de la Faculté des Sciences Agronomiques et Environnement à l'Université Évangélique en Afrique (U.E.A). Dans le territoire de Walungu, les groupements suivants ont été concernés par notre travail : Izege, Ikoma, Walungu centre, Mushinga, Nyangezi, Irhongo, Burhale et la collectivité de Kaziba. A Kabare les groupements suivants ont été considérés : Katana, Bushumba, Miti, Mudaka, Mumosho, Bugorhe, Mudusa et Chirunga. 2.2. Méthodes et matériels 2.2.1. Matériels
12
Les matériels non fongiques utilisés au laboratoire pour l'isolement et la caractérisation de M. fijiensis ont été, entre autre les matériels du laboratoire la boite de pétrie (dans laquelle le milieu de culture était coulé, c'est un bocal de conservation de la gélose), la balance de précision (a été utilisée dans la détermination de la quantité de réactifs à utiliser dans la composition du milieu de culture), le stérilisateur ou hôte à flux laminaire (a permis de travailler sans contamination), les éprouvettes, l'erlenmeyer (ballon à fond plat) (ont aidé pour le mélange des solutions), l'autocuiseur et l'autoclave (pour stériliser les récipients destinés à recevoir les milieux de culture et autres outils), les écouvillons (nécessaires pour l'écouvillonnage consistant à faire rouler délicatement les écouvillons sur les milieux sélectifs), l'incubateur (pour l'incubation à 25°C pendant 14 jours). ? Le milieu Pomme de terre Dextrose Agar (PDA) A constitué le milieu de culture de M. fijiensis dont la préparation est de Prendre 200g de pomme de terre bouilli puis filtré; ajouter 20g de dextrose et 20g d'agar dans un litre d'eau distillé et chauffé pendant quelques minutes pour dissoudre l'agar et en fin stériliser dans l'autoclave pendant 20 minutes à une température de 120°C sous une pression d'une à une et demi atmosphère. Le milieu est coulé dans des boites de pétri à raison d'au moins que toute la base de la boite soit couverte par le milieu. D'autres matériels utilisés sont : le para film (utilisé pour coller les boites de pétri. 2.3. Méthodes 2.3.1. Enquête Les travaux d'enquête et d'observation s'étaient déroulés pendant la période pluvieuse, du 21 décembre 2016 au 21 janvier 2017. 13 Une observation des bananiers malades suivie d'une enquête à un questionnaire fermé et d'une méthode quantitative ont été réalisées. Pour l'obtention des résultats escomptés, les personnes ayant des bananeraies ont été enquêtées et les différentes questions leur ont été adressées. Il était question de connaitre les différents cultivars (variétés) des bananiers rencontrés dans la région ; les maladies qui attaquent les bananiers, l'incidence et la sévérité de la cercosporiose noire du bananier à Walungu et Kabare, les cultivars sensibles et résistants à cette maladie ainsi que les moyens de lutte utilisés par les paysans. ? Echantillonnage Pour ce qui concerne l'échantillonnage, en vue de la détermination de l'incidence et de la sévérité de la maladie des raies noires, deux territoires ont été pris en compte ; Walungu et Kabare, avec huit groupements à Kabare et sept groupements à Walungu plus la collectivité de Kaziba. Deux champs ou bananeraies ont été pris dans chaque groupement en raison d'un champ par localité. Ensuite 20 plantes ont été choisies dans chaque champ de bananier selon la méthode de diagonale. Les feuilles présentant des symptômes de la cercosporiose noire du bananier ont été récoltées et en fin, ces feuilles ont été utilisées pour l'isolement de M. fijiensis. 160 personnes étaient enquêtées en raison de 10 personnes par groupement d'où 80 par territoire. L'enquête s'est déroulée du 21 décembre 2016 au 21 janvier 2017. 2.3.2. Evaluation de l'intensité de la cercosporiose noire du bananier L'intensité a été déterminée en fonction de deux paramètres qui sont l'incidence et la sévérité. L'incidence étant la proportion des unités malades ou organes d'une des parties analysées ; c'est le dégât causé par une maladie donnée. Cette incidence a été déterminée par la formule suivante : incidence= Nbre de plants malades * 100/Nbre de plants observés. La sévérité étant une évaluation qualitative du degré d'attaque (par exemple le pourcentage de la surface foliaire nécrosée). L'indice de sévérité mesure à un moment donné le taux de surface foliaire détruite par la maladie des raies noies sur un bananier (Craenen, 1998). 14 L'indice de sévérité (IS) de la maladie des raies noires a été calculée selon la formule suivante (Craenen, 1998) : Indice de Sévérité = ? ( N-1) T*100, où b = niveau de lésion des feuilles notées sur une échelle allant de 0 à 6 : 0 = pas de symptôme ; 1 = Moins de 1 % de surface foliaire touchée (uniquement stries et/ou au plus 10 lésions) ; 2 = 1 à 5 % de surface foliaire touchée ; 3 = 6 à 15 % de surface foliaire touchée ; 4 = 16 à 33 % de surface foliaire touchée ; 5 = 34 à 50 % de surface foliaire touchée ; b 6 = 51 à 100 % de surface foliaire touchée ; n = Nombre de feuilles présentant de lésion ; N = 7= nombre d'indices dans l'échelle de notation des lésions des feuilles ; T = nombre total de feuilles évaluées. 2.3.4. Isolement et caractérisation des isolats de M. fijiensis Pour la caractérisation des isolats, les feuilles présentant les symptômes de M. fijiensis provenant de Walungu et Kabare, ont été utilisées dans le laboratoire de biologie moléculaire de la Faculté des Sciences Agronomiques et Environnement de l'Université Evangélique en Afrique (UEA). Le milieu PDA (Pomme de terre Dextrose Agar) a été utilisé pour la culture des différents isolats de M. fijiensis. ? Isolement proprement dit Les isolats de M. fijiensis ont été obtenus par la technique de décharge des ascospores sur milieu gélosé (H2O Agar), qui ont été ensuite repiquées sur milieu Potato Dextrose Agar (PDA) (Carlier et al., 2003). Pour la mise à décharge, les morceaux de feuilles nécrosées de 1cm ont été pris, ensuite mis à tremper dans un litre d'eau de javel pendant cinq secondes afin de les humidifier. Les morceaux de feuilles ont été ensuite placés à l'intérieur des boites de Pétri contenant 20ml pour chacune. La face supérieure de la feuille était dirigée vers le haut, face inférieure au milieu de culture. Les boites ont été incubées à température ambiante pendant quatre jours, puis le repiquage sur PDA s'est effectué. Après repiquage, les boites étaient incubées à 25°C pendant 14 jours. L'observation au microscope a été réalisée pour l'identification des conidies 14 jours après. Les isolats obtenus ont été repiqués en boites de Pétrie afin de conserver la collection établie. La figure suivante illustre la procédure de l'isolement des populations de M. fijiensis. 15
Fig. 4 : les morceaux de feuilles avec symptômes de M. fijiensis dans la boite de pétrie ? Caractérisation phénotypique ou morphologique Les caractéristiques phénotypiques macroscopiques, visent à connaitre la coloration, la forme, l'aspect du mycélium et la vitesse de croissance des mycéliums des isolats en croissance sur milieu PDA pendant 10 jours. La croissance radiale fongique a été mesurée et calculée selon la formule suivante : avec, ri comme rayon spécifique, di le diamètre spécifique et do le diamètre de la rondelle fongique de départ (Onautshu et al., 2013). Les caractéristiques phénotypiques microscopiques ont concerné la forme et le nombre de cloisons (septa) des conidies et ont eu lieu 14 jours après incubation sur milieu PDA par la méthode de scotch au grossissement 40x, et ont été réalisées à partir des petits fragments de culture prélevés dans la boite de pétrie et montés entre lames et lamelles microscopiques avec une goutte du Gemsa concentré placée sur la lame puis le scotch a été placé sur la goutte de Gemsa. La face contenant les fragments de culture dirigée vers le haut ensuite, une autre goutte a été mise sur le scotch (sur la face dirigée vers le haut) et en fin, une lamelle y a été placée. 2.3.5. Tests de pathogénicité et évaluation de la résistance de certaines variétés à la MRN. Deux isolats de M. fijiensis dont M. fijiensisKIS-WAL et M. fijiensisBAR-KAT ont été utilisés pour le test de pathogénicité sur le bananier en fonction de nos deux territoires d'étude. Leur choix a été influencé par la vitesse de croissance mycélienne (la faible et la plus grande vitesse de croissance). L'activité s'était passée en serre, à une température de 25-27°C. Les variétés de bananier FHIA17 (à cuire/bière), NARITA4 et NARITA13 (à cuire) ont été utilisées pour ce test. L'essai s'est déroulé en serre avec un dispositif complètement aléatoire avec deux facteurs (isolats et variétés). Le dispositif expérimental est parenté à la figure suivante :
FHIA17 M. f.BAR-KAT T M. f.KIS-WAL NARITA4 M. f.BAR-KAT T M. f.KIS-WAL NARITA13 M. f.BAR-KAT T M. f.KIS-WAL FHIA17 M. f.BAR-KAT T M. f.KIS-WAL NARITA4 M. f.BAR-KAT T M. f.KIS-WAL NARITA13 M. f.BAR-KAT T M. f.KIS-WAL FHIA17 M. f.BAR-KAT T M. f.KIS-WAL NARITA4 M. f.BAR-KAT T M. f.KIS-WAL NARITA13 M. f.BAR-KAT M. f.KIS-WAL T 16 Légende : M. f.: M. fijiensis, T : Témoin. Fig. 5 : Dispositif expérimental en serre en block complétement aléatoire Le taux de développement (r) de la maladie a été déterminé en appliquant l'équation suivante : i e xi e xo r = ( Log - Log ; où t1= temps final, t0= temps initial, x1= indice de sévérité final ti-to i-xi i-xo) et x0= indice de sévérité initial (Castâno, 2002). Et la formule de l'indice de sévérité de Craenen, (1998) était ensuite utilisée, Indice de Sévérité = ?nb(1V-i) T * 100. Préparation de l'inoculum L'inoculum a été préparé en prenant les boites de pétrie contenant des mycéliums d'isolats M. fijiensisKIS-WAL et M. fijiensisBAR-KAT. Dix millilitres d'eau distillée y ont été ajoutés et les mycéliums ont été lentement grattés. Cette solution a ensuite été centrifugée pendant 20 minutes pour une homogénéisation. L'inoculation de la solution a été réalisée par pulvérisation sur les bananiers d'une quantité de 3ml. 17 La pathogénicité a été évaluée au 11ème et au 18ème jour après inoculation des bananiers. 2.4. Analyse des données Les données ont été encodées dans le logiciel Microsoft Excel 2010 ; les analyses des données quantitatives et qualitatives se sont faites par le logiciel STATISTIX 9.0. Les résultats en graphiques ont été générés par Microsoft Excel 2010. L'analyse de la variance a été utilisée pour l'incidence et l'indice de sévérité à Walungu et Kabare. La comparaison entre l'incidence et l'indice de sévérité à Walungu et Kabare a été effectuée par le test T de Tukey HSD. Au laboratoire, les données obtenues par observation au microscope et sur boite de pétrie pour la croissance des mycéliums ont été récoltées dans un carnet et encodées dans Excel 2010. 18 Chapitre 3. Présentation, interprétation et discussion des résultats 3.1. Présentation et interprétation des résultats 3.1.1. Objectif 1. Incidence et sévérité de la cercosporiose noire du bananier
Le tableau ci-dessous donne toutes les variétés des bananiers rencontrées dans les territoires de Walungu et celui de Kabare. 19 Tableau 1 : Les variétés/cultivars rencontrées à Walungu et Kabare
Le tableau ci-haut montre que dans le territoire de Kabare, toutes ces variétés y sont cultivées excepté la variété Mukombozi, de même qu'à Walungu on a toutes ces variétés sauf Poyo. Quel que soit le territoire, le Magizi est le cultivar le plus rencontré, suivi par le Kisamunyu, Kamera et Gros Michel. A Kabare le cultivar Ndundu est le moins rencontré tandis que Buganda est le moins cultivé à Walungu. D'autres cultivars sont cultivés dans les deux territoires presqu'avec les mêmes proportions. Les plantains et les cultivars à bière sont les plus cultivés dans cette partie de la province du Sud-Kivu (Magizi, Kisamunyu, Ndundu et Buganda). d) Les principales maladies des bananiers rencontrées dans le Walungu et Kabare. Le graphique ci-après montre les différentes maladies rencontrées sur les bananiers dans les territoires de Walungu et Kabare. 20
Wilt BBTV Cercosporiose Helminthosporiose Fusariose Kabare Walungu Maladies rencontrées en % 40 80 70 60 50 30 20 10 0 Fig. 6 : Les maladies rencontrées sur les bananiers dans le Walungu et Kabare La figure 6 montre que les maladies les plus rencontrées chez les bananiers dans les territoires de Walungu et Kabare sont principalement la fusariose du bananier, le flétrissement bactérien du bananier ou wilt bactérien du bananier, la cercosporiose noire du bananier ou maladie des raies noires (MRN), la Banana Bunchy Top Virus (BBTV) et l'helminthosporiose du bananier. Ce sont des maladies qui font plus de dégâts dans cette région, l'helminthosporiose ne cause que peu de dégâts à la culture du bananier. Dans les villages enquêtés, il y a eu toujours présence de ces cinq maladies dont le wilt bactérien et la fusariose sont les maladies les plus rencontrées sur les bananiers cultivés et cela beaucoup plus à Kabare qu'à de Walungu. Elles sont suivies par la maladie de la cercosporiose noire dans les deux territoires, mais cette dernière est plus rencontrée chez les bananiers à Walungu qu'à Kabare. La BBTV connu sous le nom de « Syndicat » est plus rencontrée à Walungu. e) Les principaux moyens de gestion et lutte contre les maladies chez les bananiers Le tableau suivant explique les différents moyens de gestion et de lutte contre les maladies du bananier. 21 Tableau 2 : Moyens de gestion des bananeraies
Légende : Dess= Dessouchage, Ster= Stérilisation, Sarcl= Sarclage, Tot= Total. Il ressort du tableau 2 que l'entretien constitue le moyen essentiel de gestion des bananeraies et de lutte contre les maladies à Walungu et Kabare. La majorité de nos enquêtés, les agriculteurs des bananiers font l'entretien de leurs bananeraies, cela est dû par le manque de la connaissance sur l'utilisation des produits phytosanitaires et le manque des moyens financiers. Dans les deux territoires, pour lutter contre les maladies, le dessouchage des plantes malades constitue la solution la plus efficace (52.5 % de nos enquêtés). A Walungu ; c'est beaucoup plus la stérilisation des outils ou instruments qui est utilisé pour l'entretien (58 % de nos enquêtés) alors qu'à Kabare c'est le dessouchage qui est plus pratiqué (77.5 % de nos enquêtés). De nos enquêtés, quel que soit le territoire, personne n'utilise les produits phytosanitaires (les pesticides) pour lutter contre les maladies des bananiers. Les causes principales de non utilisation des pesticides sont le manque des connaissances sur les produits phytosanitaires et le manque des moyens financiers pour s'en procurer. f) Susceptibilité des cultivars à la cercosporiose noire du bananier Le tableau suivant donne des renseignements sur les cultivars/variétés résistants et sensibles rencontrés à Walungu et Kabare selon l'appréciation des paysans. 22 Tableau 3 : Susceptibilité des cultivars à la cercosporiose noire du bananier quant aux paysans dans le Walungu et Kabare.
Légende : + : cultivars moyennement susceptible, ++ : susceptibles, +++ : très susceptibles ; - : moyennement resistant, - - : resistants, - - - : pas susceptible. Du tableau 3, il est à noter que le cultivar qui n'est susceptible à la maladie des raies noires est le Mukombozi ; après les enquêtes réalisées dans les territoires de Walungu et Kabare, les résultats montrent Mukombozi est le cultivar le plus résistant à la cercosporiose noire du bananier. Selon les explications de certains cultivateurs des bananiers dans les deux territoires, le nom « Mukombozi » dérive de la résistance du cultivar aux maladies notamment le wilt bactérien du bananier, la Banana Bunchy Top Virus (BBTV) et la maladie de la sigatoka noire. Les Malaya et Magizi sont les cultivars moyennement résistants, tandis que Ndundu est résistant à la cercosporiose noire du bananier, mais sa résistance est faible par rapport à Mukombozi. Les résultats montrent que le Kisamunyu et Barhabesha sont les cultivars susceptibles à la maladie et cela est justifié par le fait qu'ils sont des cultivars d'altitude. 23 g) L'incidence et la sévérité de la maladie des raies noires et analyse de la variance pour l'incidence dans les territoires. Le tableau ci-après illustre l?incidence et la sévérité de la MRN à Walungu et Kabare. Tableau 4 : Incidence et sévérité de la cercosporiose noire du bananier
Légende : A et B: moins sévère, C : sévère, D : moyennement sévère et E : très sévère. 24 Du tableau 4, il ressort que l'incidence de la maladie des raies noires est de 56.9 % à Walungu et Kabare. Elle est plus élevée à Walungu (72 %) qu'à Kabare (42 %). En ce qui concerne l'indice de sévérité, qui mesure à un moment donné le taux de surface foliaire détruite par la maladie sur un bananier. Cet indice est très élevé à Walungu avec 42.3% de surface foliaire des bananiers sont détruites par la maladie de la cercosporiose noire du bananier tandis qu'à Kabare il est de 35.2 %. Après analyse de la variance (ANOVA), il ressort que l'incidence et l'indice de sévérité de la maladie ont variée en fonction de deux territoires d'étude de façon significative (P-value ? 0.05, voire en annexe). La séparation de moyenne a montré une forte incidence à Walungu (71.9) avec un indice de sévérité de 42.3 % par rapport à Kabare (41.9) avec un indice de sévérité de 35.2 %. 3.1.2. Objectif 2 : Caractérisation phénotypique des isolats de M. fijiensis a) Dénomination des différents isolats identifiés dans le Walungu et Kabare. Le tableau qui suit donne les dénominations des isolats identifiés à Walungu et Kabare. Tableau 5 : Les dénominations des isolats rencontrés
Légende: BAR-KAT: Barhabesha Katana, GRO-BUSH: Gros-Michel-Bushumba, MUSH-MUM: Musheba-Mumosho, KAM-IZE: Kamera-Izege, NAK-IKO: Nakasimbu-Ikoma, KIS-WAL : Kisamunyu-Walungu centre. Le tableau 5 donne les différents isolats de M. fijiensis qui ont été identifiés dans le laboratoire de biologie moléculaire à la faculté des sciences agronomiques et environnement. Ces isolats concernent la variété sur laquelle les feuilles d'isolement ont été prélevées et le groupement d'origine des feuilles. 25 b) Nombre de cloisons des conidies des différents isolats obtenus et leurs ascospores. La figure qui suit donne les informations sur le nombre de cloisons ou segments des conidies des isolats de M. fijiensis.
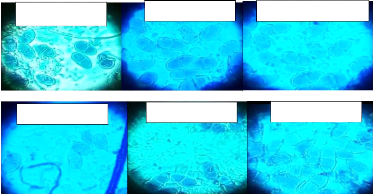
Nombre de cloisons des conidies 10 9 8 7 6 5 4 3 2 1 0 cloison minimal cloison maximal Les isolats identifiés à Walungu et Kabare Fig. 7 : Nombre de cloisons des conidies des isolats de M. fijiensis La figure suivante illustre la structure des ascospores des isolats de M. fijiensis.
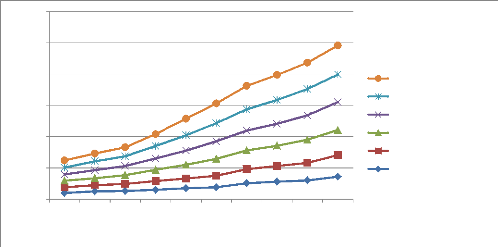
M. f. BAR-KAT M. f. KAM-IZE M. f. GRO-BUSH M. f. NAK-IKO M. f. KIS-WAL M. f. MUSH-MUM Fig. 8 : La structure des ascospores observées sur microscope optique agrandis 40x. La figure 8 montre comment les conidies de M. fijiensis sont cloisonnées et cela est fonction des différents isolats rencontrés à Walungu et Kabare. L?isolat M. fijiensisBAR-KAT présente des 26 conidies avec des cloisons allant de 3 à 8 cloisons, le M. fijiensisGRO-BUSH présente des cloisons allant de 4 à 9. Pour le M. fijiensisKIS-WAL, ses conidies ont marqué beaucoup plus de cloisons que tous les autres isolats (5 à 9 cloisons). Le M. fijiensisMUSH-MUM possède des cloisons de 3 à 6, c'est l'isolat avec moins de cloisons. Mais le M. fijiensisNAK-IKO, a des cloisons allant de 3 à 7. Et en fin, le M. fijiensisKAM-IZE, a des cloisons qui vont de 4 à 8. Les clés d'identification pour le M. fijiensis, prouvent que cette dernière a des conidies avec des cloisons allant de 1 à 10. La figure 11 montre que les ascospores de M. fijiensis sont constituées de deux cellules et ces derniers présentent une forme séparée. c) La croissance mycélienne de différents isolats de M. fijiensis La figure ci-après donne les longueurs et vitesses de croissance des différents isolats de M. fijiensis identifiés à Walungu et Kabare.
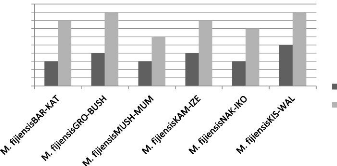
Croissance mycéliènne en mm 40 60 50 30 20 10 0 1 2 3 4 5 6 7 8 9 10 Jours de croissance M. fijiensisKIS-WAL M. fijiensisNAK-IKO M. fijiensisKAM-IZE M. fijiensisMUSH-MUM M. fijiensisGRO-BUSH M. fijiensisBAR-KAT Fig. 9 : La croissance mycélienne des différents isolats de M. fijiensis La figure 9 montre qu'après 10 jours, la croissance des mycéliums est arrivée à 9,3mm (M. fijiensisKIS-WAL, la croissance la plus élevée) en général. Mais cette croissance est fonction de chaque isolat. L'isolat M. fijiensisBAR-KAT est arrivé à 7,2mm et M. fijiensisGRO-BUSH présente la croissance la plus faible avec 6,9mm. D'autres isolats ont atteint presque une même longueur Légende : M. f. : M. fijiensis 27 mycélienne (8,8mm). Tout au début, cette croissance est un peu rapide et se ralentit petit à petit. La vitesse de croissance est fonction de chaque isolat c'est-à-dire ; pour l'isolat M. fijiensisKIS-WAL, sa vitesse de croissance est de 0,93mm/jour, pour l'isolat M. fijiensisBAR-KAT, elle est de 0,72mm/jour. L'isolat M. fijiensisGRO-BUSH enregistre la vitesse de croissance la plus faible et est de 0,69mm/jour. Tandis que les isolats M. fijiensisKAM-IZE et M. fijiensisNAK-IKO ont atteint une vitesse de croissance de 0,88mm/jour. Les populations de M. fijiensis ayant une croissance lente avec un maximum de 25mm pendant 45jours, ceci démontre que les isolats obtenus à Walungu et Kabare appartiendraient à M. fijiensis. d) La couleur des mycéliums de ces différents isolats de M. fijiensis Le tableau suivant montre les différentes caractéristiques macroscopiques des différents isolats de M. fijiensis qui ont été isolés dans les territoires de Walungu et celui de Kabare. Ce tableau donne des renseignements sur la couleur des mycéliums de différents isolats, la forme et la couleur du fond de la boite de pétrie. Tableau 6 : La couleur, la forme des mycéliums de différents isolats et la couleur du fond
28 La figure suivant illustre la couleur des mycéliums de M. fijiensis à la face inférieure de la boite de pétrie.
Fig. 10 : La coloration noire de M. fijiensis à la face inferieure de la boite de pétrie. Du tableau 6, il y ressort que les différents isolats de M. fijiensis obtenus dans les territoires de Walungu et Kabare sont différents par leur couleur, leur forme et sont tous identiques pour ce qui concerne la face inferieure ou le fond de la boite de pétrie. En observant la face inférieure de la boite de pétrie ou le fond de la boite de pétrie, on observe une coloration noirâtre ; ce qui caractérise les populations de M. fijiensis. Ainsi dit, l'isolat M. fijiensisBAR-KAT et M. fijiensisKIS-WAL possèdent une couleur blanc-verdâtre, l'isolat M. fijiensisGRO-BUSH présente une coloration blanc-rosâtre, l'isolat M. fijiensisMUSH-MUM et M. fijiensisNAK-IKO ont une coloration verdâtre ; tandis que l'isolat M. fijiensisKAM-IZE présente une coloration grisâtre. Toutes ces colorations font parties des caractéristiques principales des populations de M. fijiensis. Ces isolats présentent presque la même forme des mycéliums qui est la forme irrégulière légèrement bombée. C'est-à-dire que leur forme dans la boite de pétri n'est pas constante, elle varie d'un endroit à un autre. 3.1.3. Objectif 3 : Evaluation de la pathogénicité Il ressort des résultats que l'inoculation des bananiers par ces deux isolats a donné lieu à des symptômes typiques de la maladie des raies noires dans les conditions de la serre dont, les premiers signes visibles (face inferieure de la feuille) de la maladie sont des petits jaunes ou rouges qui sont apparues au onzième jour. Au 18ème jour, les petits points jaunes étaient devenus des tirets brun-foncés parallèles aux nervures secondaires. La figure suivante montre les symptômes de M. fijiensis en serre. 29
Fig. 11 : Symptômes de M. fijiensis en serre
La figure suivante explique le pouvoir pathogène de M. fijiensisBAR-KAT et M. fijiensisKIS-WAL sur les trois variétés utilisées. FHIA17 NARITA13 NARITA4 120
Fig.12 : Sensibilité de différentes variétés face à M. fijiensisKIS-WAL et M. fijiensisBAR-KAT. Il ressort de la figure 12 que l'isolat M. fijiensisKIS-WAL est plus virulent que l'isolat M. fijiensisBAR-KAT. Cela est dû par les conditions de développement des isolats dans les milieux, leur constitution génomique, mais également, il peut y avoir des mutations génétiques. Tous les plants de la variété NARITA13 et NARITA4 ont été infectés. M. fijiensisBAR-KAT a infecté plus NARITA4 (68% des plants) que NARITA13 (37% des plants). La variété FHIA17 a présenté une résistance face à ces deux isolats, il n'y avait pas de symptôme jusqu'au 18ème jour. b) L'indice de sévérité de la cercosporiose noire La figure ci-dessous montre comment l'indice de sévérité est différent suivant les variétés et le temps d'observation. 30
30 M. fijiensisKIS-WAL M. fijiensisBAR-KAT Indice de sévérité en % 25 20 15 10 0 5 11e jour 18e jour FHIA17 11e jour 18e jour NARITA13 11e jour 18e jour NARITA4 Fig. 13 : L'indice de sévérité de la MRN pour les différentes variétés La figure 13 explique que l'indice de sévérité le plus élevé était de M. fijiensisKIS-WAL sur NARITA13 (25%). Ce qui confirme que le M. fijiensisKIS-WAL est plus sévère que le M. fijiensisBAR-KAT. c) Le taux de développement de la maladie (en % par jour) Le tableau suivant donne le taux de développement de la MRN. Tableau 7. Le taux de développement de la maladie par jour
Il ressort du tableau 7 que le M. fijiensisBAR-KAT avait un taux de développement élevé sur NARITA13. Le M. fijiensisKIS-WAL a présenté un taux de développement élevé sur les deux cultivars, ce qui explique sa plus forte virulence. On constate que lorsque l'indice de sévérité augmente, le taux de développement augmente aussi. Le faible taux de développement de la maladie a été observé chez le M. fijiensisBAR-KAT sur NARITA4. 31 3.2. Discussion des résultats Eu égard aux résultats obtenus lors de l'étude de ce travail dont l'objectif était d'étudier la diversité de la cercosporiose noire du bananier dans les territoires de Walungu et Kabare dans le sens de mettre au point des stratégies pour lutter contre cette maladie causée par un champignon ascomycète Mycosphaerella fijiensis. Pour ce qui concerne les maladies rencontrées sur la culture du bananier dans les territoires de Walungu et Kabare, la fusariose du bananier et le wilt bactérien du bananier occupent la première position, suivie de la cercosporiose noire du bananier, l'helminthosporiose et la Banana Bunchy Top (BBTV). Nos résultats corroborent ceux trouvés par Ndungo (2008), Damme et al., (2013) et Byenda (2015) en ce qui concerne la maladie du wilt bactérien du bananier, tous ont déterminé son incidence et sa sévérité montrant que l'incidence était très élevée parce qu'il y n'a pas de variétés qui soient résistantes d'où la maladie est très sévère. Selon Bizimana et al., (2012), le wilt bactérien, la fusariose, la Banana Bunchy Top Virus et le cercosporiose noire du bananier sont les maladies plus redoutables pour la culture du bananier, ces résultats sont semblables à ceux que nous avons trouvé, parce que ce sont toujours les quatre maladies qui sont dominants dans notre région d'étude. Quant à la fusariose, elle prédomine dans la région de façon endémique par sa capacité à se maintenir dans le sol pendant plus de 30ans. Pour l'évaluation de la situation de la cercosporiose noire du bananier dans les territoires de Walungu et Kabare, cette maladie présentait une incidence qui est de 57% et une sévérité très élevée de façon générale. Mais dans certains groupements l'incidence arrivait jusqu'à 80%. L'incidence et la sévérité les plus élevées se trouvaient dans le territoire de Walungu que dans le territoire de Kabare, ceci peut être due par le fait la cercosporiose jaune du bananier était déjà signalée dans le territoire de Walungu (Zihalirhwa, 2011), or les variétés déjà attaqués par cette dernière sont très sensibles à la cercosporiose noire et attaque les bananiers de haute et basse altitude différemment de la Sigatoka jaune qui est rencontrée uniquement en basse altitude (Mourichon et al., 1997). D'après les résultats de Onautshu (2007) et Onautshu et al., (2013), dans la région de Kisangani, il montre que l'incidence de la maladie des raies noires varie entre 40 - 90%, ces résultats sont les mêmes que les nôtres. Mobambo et Naku (1993), dans la région du Haut-Zaïre, et De Lapeyre et al., (2010) ont déterminé l'incidence de cette maladie et ont trouvé que, cette dernière avait une forte agressivité. Les études de Mobambo et al., (1996), dans 32 la ville de Kinshasa a trouvé la même incidence de la sigatoka noire. Selon Mourichon et al., (1997), il a démontré que cette maladie présentait une gravité élevée (très agressif) et a également montré que la maladie présentait une incidence et une sévérité très élevée. La variété Mukombozi présente une certaine résistance à la cercosporiose noire du bananier, les autres variétés sont sensibles à cette maladie. D'où le nom de Mukombozi provient de la résistance qu'a cette variété face à la maladie des raies noires. Contrairement à nos résultats, Dhed'a et al., (2011) et Onautshu et al., (2013) dans la région de Kisangani, ont trouvé que la variété Yangambi km5 était très résistante à la cercosporiose noire. Pour la caractérisation morphologique des isolats obtenus à Walungu et Kabare, nos résultats montrent que la dénomination des isolats est faite suivant la variété sur laquelle la feuille présentant des symptômes a été tirée et le groupement d'origine. Les isolats suivants ont été obtenus : M. fijiensisBAR-KAT (Barhabesha-Katana), M. fijiensisGRO-BUSH (Gros-Michel-Bushumba), M. fijiensisMUSH-MUM (Musheba-Mumosho), M. fijiensisKAM-IZE (Kamera-Izege), M. fijiensisNAK-IKO (Nakasimbu-Ikoma) et M. fijiensisKIS-WAL (Kisamunyu-Walungu centre). Le nombre de cloisons des conidies pour les différents isolats obtenus varient entre 3 et 9. D'après les résultats de Tshidibi et al., (2013), le nombre de cloisons se situant dans les intervalles de 310. La moyenne pour le nombre de cloisons de conidies étant de 1 à 10 (CMI, 1974 ; Jones, 2000 ; Carlier et al., 2003 et Onautshu et al., 2013), ce qui a confirmé nos résultats. En ce qui concerne la vitesse de croissance mycélienne moyenne, elle était de 0,68mm/jour sur milieu PDA à température ambiante (25°C) après 10 jours. En comparaison avec les isolats de Tshidibi et al., (2013) la croissance mycélienne moyenne de LIT-TSH (1,38mm/jour) suivi de LIT-MAK (1,25mm/jour) alors que MAN-MAN (1,00mm/jour), LUX-MAK (1,00mm/jour), KIS-MAN (1,00mm/jour) après 14 jours, alors que les nôtres avaient une croissance de M. fijiensisKIS-WAL (0,93mm/jour), le M. fijiensisBAR-KAT (0,72mm/jour), le M. fijiensisGRO-BUSH (0,69mm/jour) et les M. fijiensisKAM-IZE et M. fijiensisNAK-IKO (0,88mm/jour). Contrairement à nos résultats ; Onautshu et al., (2013) a trouvé que les isolats plus lents avaient 0,19mm/jour et les plus rapides 0,36mm/jour sur PDA à 25°C, M. fijiensis étant un champignon à croissance lente (Tshidibi et al., 2013). Pour la couleur du mycélium à la face inférieure de la boite de pétrie, cette dernière est de couleur noire, qui est l'une des caractéristiques principales de M. fijiensis. Les résultats de 33 (Tshidibi et al., 2013 et Onautshu et al., 2013) confirment les nôtres selon lesquels, la présence de la couleur noire à la face inférieure du mycélium. La couleur du mycélium à la partie supérieure étant de blanc-verdâtre, blanc-rosâtre, verdâtre ou grisâtre selon l'isolat et leur forme étant irrégulière et légèrement bombée, ces mêmes résultats ont été trouvés par Tshidibi et al., (2013) et Onautshu et al., (2013). Eu égard à ces caractéristiques, nous pouvons confirmer l'hypothèse selon laquelle ces isolats appartiendraient à M. fijiensis. Pour le test de pathogénicité, les lésions macroscopiques sont apparues au 11ème jour après inoculation jusqu'au 18ème jour, les isolats M. fijiensisKIS-WAL et M. fijiensisBAR-KAT avaient un pouvoir pathogène face à deux cultivars NARITA4 et NARITA13, le M. fijiensisKIS-WAL était plus virulent que le M. fijiensisBAR-KAT. Cette virulence peut être expliquée par le fait qu'il existe plusieurs souches en fonction des zones de développement, de leur génétique et des mutations qui peuvent surgir au cours de leur développement. Le cultivar FHIA17 a présenté une certaine résistance à la cercosporiose noire du bananier. Les résultats trouvés par Rosales et al., (2010) et Kodjo et al., (sd), ont démontré que les FHIA étaient résistants à la cercosporiose noire. Par contre l'étude de Onautshu et al., (2013), dans la région de Kisangani où il a inoculé 20 isolats dans les mêmes conditions ; les premiers symptômes visibles sont observés à partir du 17ème jour et il est arrivé au stade nécrotique (stade 5) au 30-35ème jour. Or notre étude s'est limitée au deuxième stade. 34 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||