3.1.2 La variable à expliquer, le CAU
calculéà la récolte
Dans un premier temps on observe la relation entre azote
apportéet azote absorbéà partir de l'ensemble des mesures
d'absorption d'azote réalisées dans les essais. Le coefficient de
corrélation linéaire entre les deux grandeurs atteint 0,79, ce
qui montre l'intensitéforte de la liaison. Une représentation
graphique permet de confirmer les hypothèses de
linéaritéposées en introduction (fig. 3.2), le CAU
correspondant àla pente de la relation pour les données d'un
essai et les modalités d'un stade d'apport.
Azote absorbé à la récolte en
fonction de l'azote apporté
0 50 100 150 200 250 300 350

Azote absorbé (kg N /ha)
250
200
150
100
50
0
l
l
l
r = (0,79)
l
l
l
l
l
l
modalités
1 0
2 40Z21
3 80Z21
4 (X-120)Z30
5 (X-80)Z30
6 (X-40)Z30
7 (X-120)Z30+40Z32
8 (X-120)Z30+80Z32
9 (X-120)Z30+40Z39
10 (X-120)Z30+80Z39
azote apporté (kg N /ha)
Figure 3.2 - Relation entre les quantités d'azote
apportées par l'engrais et les quantités d'azote
absorbées
Le CAU est calculépour chaque bloc, de chaque
modalité, de chaque essai, suite àà un apport d'engrais.
Sur l'ensemble des valeurs de CAU obtenues durant le cycle de culture,
certaines sont beaucoup trop fortes ou trop faibles pour être
réalistes, on borne les CAU entre 0% et 150%, éliminant ainsi 71
données, laissant 2050 calculs de CAU. L'évolution du CAU en
fonction du temps depuis le moment de l'apport peut être
représenté. La figure 3.3 permet d'apprécier cette
évolution en se basant sur quelques essais représentatifs. On
remarque que le CAU évolue rapidement, ce qui appuie
l'intérêt d'utiliser des variables relatives au moment de
l'apport. Le CAU a étésystématiquement
mesuréà la récolte, on se base donc dorénavant sur
le CAU Z92 : on veut prévoir l'efficacitéde
Évolution du CAU de 6 essais
0 500 1000 1500 2000
|
21
|
22
|
34
|
|
C
C
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C
|
|
C
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6
|
|
|
|
8
|
|
|
18
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
CAU
|
1,2 1,0 0,8 0,6 0,4 0,2 0,0
|
1,2
1,0
0,8

Modalités
C 2
3
4
5
6
7
8
9
10
0,6
0,4
0,2
0,0
19
0 500 1000 1500 2000 0 500 1000 1500 2000
Cumul de températures après apport (dj)
Figure 3.3 - Évolution du CAU moyen en cours de culture
de 6 essais
l'engrais au terme de la période d'absorption de la
culture.
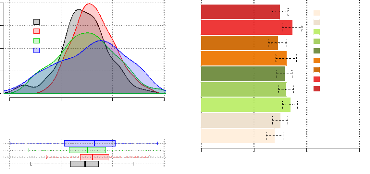
La distribution des 765 valeurs de CAU
déterminés à la récolte évolue selon les
stades d'apport (figure 3.4) : plus l'apport est tardif, plus la distribution
des valeurs semble variable. En considérant les connaissances actuelles
du CAU on veut i) rendre compte des variations entre CAU obtenus pour des
stades d'apports différents, ii) vérifier que le CAU est constant
pour les apports d'un même stade. Une analyse de variances est
réalisée sur l'ensemble des CAU mesurés àZ92. Elle
tend à expliquer le CAU par les modalités de fertilisation. Les
étapes de l'analyse amènent à considérer l'effet
aléatoire de l'essai et son interaction avec la
modalité(équation 3.1). Le test de l'effet du fertilisant fournit
une probabilitécritique 0,008, au seuil de risque : á =
5% on rejette l'hypothèse H0 ?m mm = 0. Les
tests des coefficients (avec pour contrainte >m2 = 0) mettent en
évidence une différence
significative des apports àZ30 ou àZ39, par rapport
aux apports à Z21 (représentés par la modalité2) :
des apports à des stades différents entraînent des
variations du CAU (tab 3.1). Ensuite, au regard
de la représentation graphique, les différences
entre modalités 7 et 8 (apport Z32) d'une part, 9 et 10 (apport Z39)
d'autre part, même si elles ne sont pas significatives, ne semblent pas
négligeables, les CAU entre modalités fertilisées au
même stade semblent variables. Or, l'apport d'azote étant au
maximum inférieur de 40 kg N ha-1
àla dose optimale X, on peut aisément
supposer que les doses apportées aient toujours étésous
optimales : en conséquence les CAU étaient attendus constants. Il
faudra donc chercher d'autres explications, notamment du côtéde la
pluviométrie ou de l'INN. Mais attention, bien que l'on perçoive
des différences entre modalités d'un même stade d'apport on
ne pourra pas les expliquer. En effet, les variables dont nous disposons ne
permettent pas de différencier les modalités d'un même
stade, puisque à un stade donnénous disposons :
- de VC relatives àun cumul de degrés jours
(équivalentes pour des apports réalisés le même
jour);
- d'INN relatifs àune mesure du témoin commun
aux modalités d'un même stade (deux modalités partageant le
même témoin sont décrites par les mêmes valeurs
d'INN);
- de pluviométries calculées pour une date
d'apport (équivalentes pour un apport le même jour).
Un modèle explicatif du CAU ne discriminera pas les
modalités d'un même stade avec le jeu de données dont nous
disposons. Autrement dit, la variabilitépour un même stade
d'apport ne pourra être expliquée. D'ailleurs, cette
variabilitéest importante en ordre de grandeur puisque pour un essai,
20
les CAU mesurés à la récolte, et pour les
modalités qui partagent le même stade d'apport, l'écart
type moyen est de 14,9%.
yiem = u + mm +
Ee + MEme + åiem
(3.1)
- Ee iid, : Ee ~
.V(0,óE);
- å iid, : å .V(0, ó);
- MEme iid, : MEme .V(0,
óME);
- Ee, MEme et
åmei indépendants deux à deux.
|
Distribution du CAU selon le stade
d'application
|
Moyennes aujustées de CAU en fonction de la
modalité de fertilisation
|
Densité
0,0 0,5 1,0 1,5 2,0
0,0 0,5 1,0 1,5
Stade d'application
Z21 Z30 Z32 Z39
l
CAU
10
4
2
9
8
7
6
5
3
0,0 0,5 1,0 1,5
CAU
40Z21
80Z21 (X-120)Z30 (X-80)Z30 (X-40)Z30 (X-120)Z30+ (X-120)Z30+
(X-120)Z30+ (X-120)Z30+
Figure 3.4 - Distribution du CAU selon les stades de
fertilisation et comparaison des CAU obtenus pour chaque
modalité(moyennes ajustées des CAU par modalitéde
fertilisation et leur intervalle de confiance)
Table 3.1 - Estimation des coefficients liés aux
modalités
|
Estimation
|
ó
|
test t
|
|
u
|
0,70
|
0,04
|
17,04
|
|
Modalité3
|
0,05
|
0,04
|
1,22
|
|
Modalité4
|
0,15
|
0,04
|
3,42
|
|
Modalité5
|
0,11
|
0,04
|
2,55
|
|
Modalité6
|
0,09
|
0,04
|
2,18
|
|
Modalité7
|
0,11
|
0,05
|
2,06
|
|
Modalité8
|
0,03
|
0,05
|
0,56
|
|
Modalité9
|
0,16
|
0,05
|
3,17
|
|
Modalité10
|
0,05
|
0,05
|
1,03
|
fréquence
0 10 30 50
3.1.3 Les variables explicatives 3.1.3.1 La vitesse de
croissance
Les calculs de cinétiques ont
étéautomatisés. Pour chaque modalitéde chaque essai
on dispose d'une expression de la cinétique de croissance à trois
paramètres (K, fi et À,
équation 2.5, p. 10). De manière générale, les
ajustements obtenus sont de bonne qualité(fig. 3.5). En effet, le biais
est environ nul et la Racine de l''Ecart Quadratique Moyen (REQM) est en
moyenne de 0,40 t/ha (en sachant que àla récolte, la biomasse
maximale mesurée était de : 14 t/ha). Cependant, les valeurs des
paramètres ajustés sont parfois très variables, notamment
K, qui correspond àla limite de production permise, qui atteint
des valeurs supérieures à30 t/ha : ces ajustements ne peuvent
être admis car hors du domaine de validitéagronomique. De
même, des valeurs de À très fortes sont
détectées; celles-ci sont corrélées àK
(r = 0,40) puisque pour atteindre un K hors norme il faut une
échelle de temps supérieure que peut représenter
À. Le retour sur les données montre que ces valeurs sont
associées à des ajustements de cinétiques
fragilisés par le manque de données acquises pendant la culture,
notamment entre Z39 et Z92. Au total, ce sont 17 ajustements qui sont
problématiques sur les 340 possibles.
fréquence
0 10 20 30 40 50 60
Distribution des REQM

0,0 0,2 0,4 0,6 0,8 1,0 REQM
Distribution du biais

-0,05 0,00 0,05 0,10 0,15
Biais
21
Figure 3.5 - Qualitédes ajustements des modèles
de croissance. Distribution de la Racine de l'Erreur Quadratique d'Estimation
et du biais (différence moyenne) entre estimations et valeurs
observées
Il est possible d'ajuster un modèle de croissance pour
chaque essai et chaque modalité(Le Souder et al., 2007). On s'attend
à ce que la différenciation en terme de VC n'apparaisse
qu'après l'apport. Autrement exprimé, avant la fertilisation,
rien ne distingue la modalitéde son témoin : leur régimes
de croissance sont analogues. Cependant, le niveau de fertilisation conditionne
la production de biomasse
à la récolte. De fait, le paramètre
K, qui représente la capacitélimite du milieu, varie
d'une modalitéà l'autre. Ces variations se ressentent
sensiblement sur les autres paramètres de l'expression de la
cinétique, il en résulte que les VC aux dates
d'apport différent en moyenne de 0,2 g MS
m-2dj-1 entre la
modalitéet son témoin, la différence maximale étant
de 0,45 g MS m-2dj-1.
La figure 3.7 fournit un exemple d'ajustement de cinétiques illustrant
ces écarts d'estimations de VC entre modalitéet témoin. On
est donc contraint d'utiliser les VC calculées sur les témoins,
en considérant un régime de croissance de la
modalitéanalogue àcelui de son témoin évitant ainsi
d'introduire une variabilité»artificielle».
En comparaison aux VC des expérimentations de Limaux
(1999), la gamme obtenue est plus grande : elle varie de 0,004 à1,81
g MS m-2dj-1 (moyenne
= 0,60, écart-type = 0,47) contre un intervalle de VC de 0,15
à0,91 g MS m-2dj-1
pour Limaux (1999) (moyenne = 0,42, écart type = 0,21), ceci
s'expliquant par l'ajout dans le protocole de CAU pour des apports tardifs pour
lesquelles
Estimation des paramètres

K

50
40
30
20
10
7 6 5 4 3 2
4000
3500
3000
2500
2000
1500

l
ë
â
22
Figure 3.6 - Estimations des paramètres des
cinétiques de croissance obtenus par régression non
( 1 - e-(t/ë)â)
linéaire, MS(t) = K
les VC sont encore plus fortes (Z32 et Z39) (figure 3.8). Le
graphique 3.8 illustre le lien fort qui existe entre VC et stade de
fertilisation, la VC est d'autant plus forte que le stade physiologique du
bléest avancé. Ceci est vrai parce que la fertilisation
intervient avant le point d'inflexion de la cinétique de croissance. La
VC, par sa construction dépendante des sommes de températures,
est donc porteuse pour partie de l'information liée au stade
physiologique du bléau moment de l'apport.
|
Cinétique labergement 2005
|
Vitesse de croissance
(agrandissement)
|

800 900 1100 1300
VC (gr de MS m-2dj-1)
0,0 0,5 1,0 1,5 2,0
Z21
0
40Z21
80Z21
(X-120)Z30 (X-80)Z30 (X-40)Z30 (X-120)Z30+40Z32 (X-120)Z30+80Z32
(X-120)Z30+40Z39 (X-120)Z30+80Z39
Z30
Z32
Z39
0 500 1500 2500
0 5 10 15 20
MS (t/ha)
Z92
Z21
Z30 Z32 Z39
Z65
Vitesse de croissance
0 500 1500 2500
Z21 Z30
Z32 Z39
Z65
Z92
0,0 0,5 1,0 1,5 2,0
VC (gr de MS m-2dj-1)
23
Degrés jour Degrés jour
Figure 3.7 - Ajustement de cinétiques de croissance.
Exemple des modèles de croissance ajustés pour les
modalités de l'essai de Labergement (21), conduit en 2005
Évolution de la VC au cours du temps, selon les
stades
VC à l'apport (gr de MS m-2dj-1)
0,0 0,5 1,0 1,5

l
l
Z21 Z30 Z32 Z39
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
0 500 1000 1500 2000
Somme de températures depuis le semis (dj)
Figure 3.8 - Évolution de la VC
24
3.1.3.2 État de nutrition
azotée
Mesures de teneurs en azote

Stade de fertilisation
Année
0 1 2 3 4 5
l
l
l
l
l
l
l
l
l
l
l
l
Teneur en azote %
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
l
données brutes
données brutes
à l'apport
teneur en azote critique carence en azote
0 5 10 15
Matière sèche t/ha
2004 2005 2006
N Ncrit
INN =
0,6 0,9 1,2
0,6 0,9 1,2
N Ncrit
INN =
Z21 Z30 Z32 Z39
Figure 3.9 - Teneurs en azotes mesurée et INN
calculés selon l'année ou le stade de fertilisation
La figure 3.9 représente les teneurs en azote
mesurées au cours de la culture en fonction des mesures de biomasses
correspondantes. Globalement, la majoritédes teneurs mesurées
révèlent une teneur en azote inférieure à la teneur
en azote critique. Toutefois le nombre de cas oùl'azote est en
excès augmente pour des valeurs de matières sèches plus
faibles, concernant donc les mesures les plus précoces. L'INN traduit
l'éloignement àla teneur en azote critique. Au moment de
l'apport, on calcule 440 valeurs d'INN. L'INN varie de 0,54 à1,26
(écart type : 0,17). Pour connaître l'implication des effets
aléatoires on réalise l'analyse des composantes de la variance du
modèle 3.2 (tab. 3.2). Le stade est ce qui induit le plus de
variabilitédans les mesures, devant l'essai. À eux deux ils
expliquent 62% de la variabilitéde l'INN.
INNiaes = u +
Aa +
Ss + Ee|a +
ASas + åiaes
(3.2)
|
|


