Inventaire systématique des genres et espèces, représentants de la famille des acanthacées dans les districts de Tolagnaro et d’Amboasary-atsimo, Madagascar.par Andrianandrasana Balzac MBOLA VERSENE UNiversité d'Antananarivo - Doctorat en sciences de la vie et de l'environnement 2018 |

CONCLUSIONLa famille des Acanthaceae, principal objet de la présente thèse, possède environ 4000 espèces reparties sur 221 genres dans le monde tropical. Cette thèse a pour objectif de compléter les informations sur cette famille afin de pouvoir plus tard finaliser sa clé de détermination pour Madagascar. La zone d'étude est le Parc National d'Andohahela et ses environs, dans les districts de Tolagnaro et d'Amboasary-Atsimo. Cette zone a été choisie parce qu'elle est subdivisée en trois parties selon le paramètre `subdivision climatique', parties qui sont supposées comporter différentes espèces d'Acanthaceae. Ces domaines phytogéographiques correspondent en effet à trois conditions climatiques délimitées à partir de l'existence d'une faille pluviométrique au niveau de Ranopiso-Bevilany qui définit par conséquent trois habitats distincts de ces espèces à savoir la forêt humide de l'Est de la Parcelle 1 du Parc National d'Andohahela, une zone de transition à Dypsis decaryi au niveau de Ranopiso-Bevilany et les formations sèches de l'Ouest (forêt, fourré et savane). Les méthodes utilisées dans cette thèse sont surtout les inventaires de terrain des espèces d'Acanthaceae et leur étude morphologique en laboratoire qui sont les herbaria du Parc Botanique et Zoologique de Tsimbazaza, Antananarivo et de California Academy of Sciences de San Francisco. Les inventaires ont consisté à la collecte des spécimens et à l'étude de leur habitat. Pour cela, les paramètres écologiques relevés sont le facteur lumière traduit par l'ouverture du milieu, l'altitude et le type de sol. Vingt-neuf (29) espèces d'Acanthaceae, regroupées dans 17 genres, ont été collectés au cours de la présente étude. Leurs études morphologiques ont montré que ces Acanthaceae sont surtout des `hémicryptophytes' sauf Ruellia geay qui est un `nanophanérophyte', sont pour la plupart inermes (72%) et leur inflorescence est surtout de type épi. Une clé de détermination partielle a été faite en considérant ces espèces. Cinq (5) nouvelles espèces regroupées dans trois (3) genres ont été identifiées, à savoir Camarotea, Justicia et Hypoestes. Pour ce faire, le type biologique a été utilisé car c'est le premier caractère visible sur terrain et les conditions du paysage permettaient de le voir facilement. L'analyse des spécimens de l'herbarium du TAN, principal bénéficiaire des résultats produits ici permettra d'améliorer l'état des données sur les Acanthaceae afin d'atteindre l'objectif principal défini dans le cadre de cette thèse, à savoir la finalisation des différents tomes de cette clé. 143 Conclusion Treize (13) espèces sont visibles dans la partie humide, onze (11) au niveau de la zone de transition et sept (7) dans la partie sèche. Quelques espèces se retrouvent dans deux types d'habitat ; une seule espèce (1) est vue dans la zone humide et dans la partie sèche (Barleria kicthingi) et une seule espèce se rencontre dans les zones de transition et sèche (Ecbolium syringifolium). La première hypothèse énoncée dans le cadre de cette étude qui stipule que `les Acanthaceae sont des bons indicateurs climatiques' est vérifiée car chaque subdivision bioclimatique a ses propres espèces sauf pour les cas des deux espèces mentionnées ci-dessus. Vingt une (21) espèces se rencontrent en moyenne altitude, neuf (9) en haute, et vingt (20) en basse altitude. Quatre (4) espèces se développent exclusivement en moyenne altitude. Ce sont Hypoestes decaryana, H. strigata, H. sp.1 et Barleria decaryi. Trois espèces se trouvent dans quatre (4) types d'altitude différents, à savoir Asystasia gangetica, Barleria alluaudii, B. kitchingii et Phaulopsis imbricata. Contrairement aux données obtenues à partir des analyses des étiquettes des herbiers du TAN, les Acanthaceae ne se retrouvent pas seulement en milieu ouvert. En effet, seules huit (8) sur les vingt-neuf (29) espèces étudiées, soit 27% y poussent. Le reste se développe respectivement en milieu fermé avec neuf (9) espèces et treize (13) en milieu semi-fermé. La deuxième hypothèse qui stipule que `les espèces de la famille des Acanthaceae à Madagascar se développent en milieu ouvert', énoncée dans le cadre de cette étude n'est pas vérifiée car chaque type d'ouverture de milieu a ses propres espèces. Les différents types de sol du site de relevé ont leurs propres Acanthaceae sauf pour quatre espèces ; Ecbolium syringifolium est indifférente et pousse sur les trois types de sols ; Barleria her et B. kitchingii, Blepharis calcitrapa et B. maculata sont visibles sur deux types de sol, alluvionnaire et calcaire. La repartition globale montre bien une exclusivité car douze (12) espèces se trouvent sur sol alluvionnaire, deux (2) sur calcaire et quatorze (14) sur sol noir de forêt. Donc, les Acanthaceae à quelques exceptions près sont exigeants par rapport aux conditions édaphiques. Ces constats sont seulement valables dans le cas de la présente étude et les situations dans les autres régions de Madagascar doivent être étudiées à partir de plus vastes prospections. Les détails y relatives sont données plus haut en récommandations. Pouvoir finaliser la clé entière des Acanthaceae Malagasy, on peut suggérer en persperctive, les thèmes de recherches suivants : a. Réviser les Acanthaceae malagasy pour finaliser leurs classifications et arriver au niveau `espèce'. A titre d'exemple, les Justicieae possèdent plusieurs espèces qui se répartissent dans tout Madagascar, en particulier les cinq (5) nouvelles espèces (Justisia sp, Camarotea 144 Conclusion sp, Hypoestes sp.1, Hypoestes sp.2, Hypoestes sp.3) découvertes au cours de la présente étude ;
145 Références bibliographiques REFERENCES BIBLIOGRAPHIQUES
146 Références bibliographiques
147 Références bibliographiques 30.DANIEL, T.F., McDADE, L.A., MANKTELOW, M., and KIEL, C.A., 2008b. Toward a comprehensive understanding of phylogenetic relationships among lineages of Acanthaceae s.l. (Lamiales). American Journal of Botany. Vol. 95 (9) : 1136 - 1152.
148 Références bibliographiques
56.McDADE, L.A., DANIEL, T.F., KIEL, C.A., and VOLLESEN, K., 2005. Phylogenetic Relationships among Acantheae (Acanthaceae) : Major Lineages Present Contrasting Patterns of Molecular Evolution and Morphological Differentiation, The American Society of Plant Taxonomists, Systematic Botany, 30 (4) : 834 - 862. 57. McDADE, A.L., DANIEL, T.F. and CARRIE, A. K., 2008. Toward a comprehensive understanding of phylogenetic relationship among lineages of Acanthaceae S.L. (Lamiales). American Journal of Botany. Vol. 95, n°9 : 1136 - 1152. 149 Références bibliographiques
150 Références bibliographiques
151 Références bibliographiques WEBOGRAPHIE
152 Références bibliographiques 16. Reich, K. Esticos criticos sobre la Flore de Chile. 81e Familia. (consulté, fevrier 2017) http://images.mobot.org/efloras/FloraData/060/PDF/V05/ vol5-81 Acanthaceae .pdf
I Article PROCEDINGS OF THE CALIFORNIA ACADEMY OF SCIENCES Fourth Series Volume 58, No 8, pp, 121-131, 4 figs May 11, 2007 Anisotes (Acanthaceae) in Madagascar DANIEL1, T. F., MBOLA3, V. A. B., ALMEDA 1, F. et PHILLIPSON2, P., 1Department of Botany, California Academy of Sciences, 875 Howard Street, San Francisco, California 941-3 ; 2Missouri Botanical Garden, P.O. Box 299, St. Louis, MO, 63166-0299, U.S.A. and Muséum National d'Histoire Naturelle, Département Systématique et Evolution. Boîte postale 39, rue Cuvier 57, 75231 Paris, Cedex 05, France ; 3PNM-ANGAP, Parc National d'Andohahela, BP.73 Tolagnaro 624, Madagascar. balzacmbola2018@gmail.com Two species of Anisotes (Acanthaceae, Justicieae) are recognized from Madagascar. Anisotes madagascariensis is endemic to sandy, coastal regions of southern Madagascar. New morphological, distributional, and ecological data are presented for this species. A newly described species, A. divaricatus, is known only from calcareous, rocky substrates near Lake Tsimanampetsotsa in southwestern Madagascar. Major morphological distinctions between A. divaricatus and A. madagascariensis include : habit (divaricate branches vs. ascendant branches), length (5--11 vs. 14--45 mm long) and shape (obcordate to obdeltate vs. broadly ovate to elliptic) of leaf blades, corolla form (corolla tube : corolla 0.46--0.56 vs. 0.23--0.38; lower lip not spirally coiled vs. spirally coiled), and capsule pubescence (lacking trichomes vs. trichomes present). Images of flowers and pollen, morphological descriptions, and mapped distributions are provided for both species. Nineteen species occurring from tropical and southern Africa to Madagascar and tropical Arabia were recognized in the most recent revision of Anisotes (Baden 1981). The genus has traditionally been included in subfamily Acanthoideae, tribe Justicieae based on its 2-lipped, «imbricate» (presumably ascending cochlear aestivation in Anisotes and its relatives) corollas with bithecous stamens. Baden (1981) and others have noted that generic delimitation in Justicieae is problematic. Indeed, Anisotes does not appear well circumscribed in the tribe based on either macromorphological or palynological characters. No morphological synapomorphies can be identified for it, and it is reasonable to suppose that it is not necessarily monophyletic. Baden (1981) did not identify a probable closest relative for Anisotes, but he noted some morphological similarities (especially corollas) between Anisotes and Metarungia Baden (as Macrorungia C.B. Clarke). He distinguished them by the presence of elastically separating placenta (with attached retinacula) in capsules of the latter genus; placentas/retinacula remain attached to the inner surface of capsules in Anisotes. A single species of Anisotes has been reported from Madagascar, A.madagascariensis; however, Lindau (1898) described Symplectochilus madagascariensis Lindau from Madagascar, and Symplectochilus is now treated as congeneric with Anisotes. Baden (1981) indicated that the taxonomic position of S. madagascariensis «remains doubtful due to inadequate type material.» Indeed, the type (Antsiranana : Bay of Diego Suarez, forest, 1833, Goudot s.n. at G, photo!) of this name was collected in a different habitat far from the known distribution of A. madagascariensis, does not represent the same taxon as the type of A. madagascariensis, and does not necessarily pertain to the genus Anisotes. However, plants resembling Anisotes have been reported (e.g., Gautier 2002) in other regions of Madagascar ; II Article 122 PROCEDING OF THE CALIFORNIA ACADEMY OF SCIENCES Fourtth series, Volume 58, n°8 Benoist annotated specimens at P with names of several new species and new combinations for plants from Madagascar and the Comoros, but he did not publish them ; and several herbarium specimens of unknown taxa (e.g., Phillipson 1904 at CAS from Mahajanga, Bosser 5917 at TAN from Montagne d'Ambre) show affinities to Anisotes. This summary of Anisotes in Madagascar incorporates information not known to or reported by Baden (1981) for A. madagascariensis, such as : descriptions of capsules and seeds, fruiting phenology, detailed distributional data, chromosome number, and phylogenetic position based on molecular sequence data. In addition, a second distinctive species from southwestern Madagascar is newly described and compared to A. madagascariensis. Recent molecular phylogenetic studies that included Old World «justicioids» (McDade et al. 2000 ; Kiel et al. 2006) reveals that Anisotes is closely related to several species of Justicia from the Old World (e.g., J. adhatoda L., J. anagalloides (Nees) T. Anderson, and J. campylostemon (Nees) T. Anderson) and Duvernoia aconitiflora A. Meeuse. Based on limited sampling of Anisotes, Kiel et al. (2006) showed that the African species A. rogersii S. Moore is sister to J. adhatoda, and that this pair is sister to A. madagascariensis in a strongly supported clade. Additional molecular markers as well as more samples of Anisotes and Old World «justicioids» will be necessary to test monophyly for the 19 species treated by Baden (1981) and to resolve possible phylogenetic relationships among them, but there appears to be little doubt that A. madagascariensis is closely related to African species of Anisotes. Metarungia appears to be monophyletic (based on a sampling of two species ; Kiel et al. 2006) and basal to a larger clade that includes Anisotes. Daniel (2007) reported a chromosome count of n = 17 for Anisotes madagascariensis, the first number reported for the genus. Chromosome numbers of n =17 have been reported elsewhere among Old World «justicioids» in two species of Justicia (Daniel and Chuang 1998 ; Daniel 2000 ; Daniel et al., 2000), in Duvernoia aconitiflora (Daniel et al., 2000), and in Rungia repens Nees (Subramanian and Govindarajan 1980 ; however, most counts for this species are n = 10 : Narayanan 1951 ; Ellis 1962 ; Ranganath 1981 ; Krishnappa and Ranganath 1982 ; Saggoo 1983). Counts of n = 17 are well established in both J. betonica L. of section Betonica and J. adhatoda of section Vasica. The common chromosome number shared by A. madagascariensis, J. adhatoda, and D. aconitiflora undoubtedly reflects their close phylogenetic relationships as revealed by molecular markers (Kiel et al. 2006). In spite of counts of n = 17 in J. betonica and Rungia repens, neither the former species nor the latter genus appear especially closely related to the clade consisting of Anisotes and J. adhatoda based on molecular phylogenies (McDade et al. 2000 ; Kiel et al. 2006). The sole chromosome count for Metarungia, posited as a potentially close relative of Anisotes by Baden (1981), is n = 14 (Daniel et al. 2000). Muller et al. (1989) described and figured pollen of A. madagascariensis (from Humbert & Swingle 5295) as 3-colporate and 6-pseudocolpate. Baden (1981; based on Perrier de la Bâthie 9494) described it as 3-porate with a trema area having reticulate bands (i.e., 3-colporate, 6-pseudocolpate). Variation among other species of the genus (Baden 1981) includes pollen 2--4-porate (or 2-colporate) with the trema area having two or four rows of insulae rather than pseudocolpi. Baden (1981) used palynological differences to help delimit six sections of Anisotes. Pollen of the newly described species from Madagascar, A. divaricatus, resembles that of A. madagascariensis in size, aperture type and number, and exine microsculpturing (i.e., bireticulate). III Article Daniel et al., ANISOTES (ACANTHACEAE) IN MADAGASCAR 123 Anisotes Nees Type : A. trisulcus (Forssk.) Nees Himantochilus T. Anders. ex Benth. Type: H. sessiliflorus T. Anders. Symplectochilus Lindau Type: S. formosissimus (Klotzsch) Lindau Suffrutescent perennials, shrubs, or small trees. Leaves deciduous or evergreen, opposite, sessile to petiolate, margin entire to crenate. Inflorescences of axillary or terminal spikes, racemes, or thyrses ; bracts opposite; bracteoles present or absent; dichasia 1-3- flowered, sessile to pedunculate. Flowers sessile to pedicellate ; calyx 5-lobed; corolla reddish, purple, orange, or yellowish, tube #177; cylindric to ampliate distally, limb 2-labiate, upper lip convavo-convex, usually #177; arched, entire to 2-fid at apex, internally rugulate, lower lip deeply 3-lobed, straightforwardly splayed, recoiled, or spirally twisted, aestivation presumably ascending cochlear (lower lip outermost in bud, but lobes of lower lip either not evident or not overlapping in buds observed). Stamens 2, inserted near apex of corolla tube, exserted from corolla tube but not extending beyond upper lip (under which they are #177; enclosed), anthers 2-thecous, thecae parallel to #177; oblique, subequally to unequally inserted, #177; equal to unequal in size, basal appendage present or absent; staminodes absents ; pollen 2-, 3-, or 4-aperturate, colporate/pseudocolpate or porate (to colporate) in trema regions studded with circular insulae. Stigma 2-lobed or lobes obscure. Capsule stipitate, retinacula remaining attached to inner wall of mature capsule. Seeds 4 (or fewer by abortion), lenticular. Key to Species of Anisotes in Madagascar 1. Leaf blades broadly ovate to broadly elliptic to elliptic to ovate-elliptic, 14--45 mm long, 10-- 38 mm wide, 1.1--2.7 times longer than wide, rounded (to emarginate) at apex ; calyx 2--3 mm long, margin of lobes #177; densely ciliate ; corolla with the internal surface conspicuously lighter colored than the external surface, corolla tube : corolla = 0.23--0.38, corolla tube 9--15 mm long, upper lip 20--35 mm long and distally whitish to pinkish along the margin, lower lip spirally coiled, 18--28 mm long, lobes 12--14 mm long; stamens 26--33 mm long; capsule pubescent with flexuose to antrorse eglandular trichomes 0.05--0.1 mm long; growing on sandy flats and dunes. A. madagascariensis growing on limestone A. divaricatus Anisotes madagascariensis R. Ben. Bull. Soc. Bot. France 76:1036. 1929. TYPE : MADAGASCAR. Toliara: dunes, rives du Lac Mananpetsa [Tsimanampetsotsa], coté Mahafaly, June 1910, H. Perrier de la Bâthie 9494 (lectotype, designated by Baden in 1981: P!; isolectotype: P, digital image!). Figure 1. IV Article 124 PROCEDING OF THE CALIFORNIA ACADEMY OF SCIENCES Fourtth series, Volume 58, n°8 Shrubs to 2 m tall; older branches (#177; divaricate to) ascending usually at angles of 40-70°. Young stems quadrate-ridged, densely and evenly pubescent with (flexuose to) antrorse to antrorsely appressed eglandular trichomes 0.2-0.4 mm long, pubescence resulting in a grayish green coloration. Leaves coriaceous to subsucculent, petiolate, petioles to 12 mm long, blades
Figure 1. Flowers of Anisotes divaricatus (a--b) and A. madagascariensis (c). a. Almeda 9237, scale = 4 mm, photo by F. Almeda. b. Almeda 9237, scale = 5 mm, photo by H. Mally. c. Almeda 7705, scale = 8 mm, photo by F. Almeda. V Article Daniel et al., ANISOTES (ACANTHACEAE) IN MADAGASCAR 125 broadly ovate to broadly elliptic to elliptic to ovateelliptic, 14-45 mm long, 10-38 mm wide, 1.12.7 times longer than wide, acute at base, rounded (to emarginate) at apex, venation obscure, surfaces densely and evenly pubescent with cauline type trichomes, pubescence resulting in a grayish green coloration. Inflorescence of #177; congested, axillary (usually only in axils of distalmost 1-3 pairs of leaves) and/or terminal, pedunculate spikes, peduncles to 24 mm long, rachis densely pubescent like young stem, internodes usually very short or inconspicuous. Bracts opposite, all fertile, subtending a single flower, broadly triangular, 1-2 mm long, 1.5-2.2 m wide, abaxial surface densely pubescent with antrorse eglandular trichomes 0.1-0.2 mm long. Bracteoles absent. Flowers sessile (to subsessile), usually more than 2 per spike. Calyx 5-lobed, 2-3.5 mm long, tube ca. equal to lobes (i.e., slightly longer to slightly shorter than lobes), lobes ovate to triangular, 0.6-1.8 mm long, 0.9-1.1 mm wide, 0.30-0.69 times as long as calyx, abaxial surface nearly glabrous to sparsely pubescent (especially near apex) with antrorse to antrorsely appressed eglandular trichomes 0.05-0.1 mm long, margin #177; densely ciliate with flexuose to antrorse eglandular trichomes. Corolla maroon to dull red to pink-brown externally, pinkish to whitish internally, externally glabrous, (30-) 33-47 mm long, tube proximally cylindric, #177; gradually expanded distally, 9-15 mm long, 3-4 mm in diameter near midpoint, corolla tube : corolla = 0.23-0.38, upper lip 20-35 mm long, internally rugulate, entire at apex, distal portion with whitish to pinkish margin, lower lip spirally coiled, 18-28 mm long, lobes 12-14 mm long, 1.6-3.2 mm wide. Stamens 26-33 mm long, inserted near apex of corolla tube, exserted from mouth of corolla but not surpassing upper lip, filaments whitish, glabrous, thecae maroon, unequal in size (distal theca longer, 3.2-4 mm long, proximal theca shorter, 2.4-3.5 mm long), unequally inserted (overlapping by 1-1.6 mm), glabrous, lacking basal appendages (or proximal theca with an inconspicuous appendage to 0.1 mm long); pollen 62.5-74 ìm polar diameter (P), 32.5-42.5 ìm equatorial diameter (E), P:E = 1.52-1.90. Style 29-36 mm long, glabrous, stigma lobes obscure or 0.2 mm long. Nectar disk cupuliform and often visible between calyx lobes. Capsule 18-24 mm long, externally pubescent with flexuose to antrorse eglandular trichomes 0.05-0.1 mm long, stipe 9-11 mm long, head ellipsoid to obovoid. Seeds 4, discoid, (3.2-) 4.5-5 mm long, (3.6-) 4.1-5 mm wide, surfaces minutely papillose or rugose and shiny, margin #177; swollen and smooth. (n = 17, Daniel 2007). Floral Phenology. Flowering : throughout the year ; fruiting : May, August-October. Distribution and habitats. Endemic to southern Madagascar; plants occur on dunes and in sandy flats in regions of spiny forest (with Diderea madagascariensis and Euphorbia stenoclada) generally along the coast of the Mozambique Channel at elevations from sea level to 20 m (Fig. 2). Two apparently wild collections were made in coastal regions of the Indian Ocean near Ambovombe Androy, well to the east of Cap Sainte Maire and about 225 km disjunct from the nearest locale of the species at Itampolo on the west coast (Allorge 428 and Decary 2729--not seen, but cited among the syntypes of A. madagascariensis in the protologue). The coastal region between Itampolo and Ambovombe remains poorly collected, and it is possible that the range of the species is somewhat more extensive than presently known in coastal regions of southern Madagascar. Local names. «Berava,» «beravo» (Koechlin 25) ; «hazontsohy» (Ravelonanahary 3286-RN) ; «hazontsoy» (Ravelonanahary 2787-RN, 3926-RN, 4287-RN) ; «hazontsohy lahy» (Ravelonanahary 4159-RN) ; «voamany» (Koechlin 1) ; «voamamy» (Koechlin 2). Baden (1981) treated species of Anisotes in six sections based primarily on characters of the inflorescence and pollen. Anisotes madagascariensis was placed into section Spiciflori based on its inflorescence VI Article 126 PROCEDING OF THE CALIFORNIA ACADEMY OF SCIENCES Fourtth series, Volume 58, n°8 spikes, non-reticulate nerved bracts, «large» corolla: tube ratio, and 3-aperturate pollen with apertures flanked by pseudocolpi (Fig. 3). The only other species placed into this section by Baden (1981), A. formosissimus (Klotzsch) Milne-Redhead, occurs in Mozambique, Zimbabwe, and Malawi in southeastern Africa. It differs from A. madagascariensis by its longer spikes, bracts, and calyx; glabrate leaves; and bracteoles, which are present. Although Baden (1981) cited only 5 collections of this species, plants are rather frequent along the southwestern coast of Madagascar. Because plants often either persist or become established in disturbed habitats, this attractive plant is well suited for horticultural use in local landscaping. Cours 4643 differs from other collections by having spikes 2-flowered and flowers subsessile (i.e., borne on pedicels to 1 mm long); in other respects, it resembles most other collections. Although pollinators have not been documented for A. madagascariensis, Long-billed Green Sunbirds (Nectarinia notata) were observed visiting flowers of Daniel & Butterwick 6736 in November.
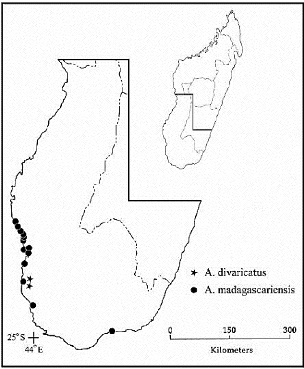
Figure 2. Map of southwestern Madagascar showing distributions of A. divaricatus and A. madagascariensis. Additional specimens examined. MADAGASCAR. Toliara : Ambovombe, chemin 100 m avant l'océan, 11 September 1992 (flr), L. Allorge 428 (P) ; just N of Itampolo on Mozambique Channel, 1 April 1996 (flr), F. Almeda 7705 (CAS) ; Itampolo, 13 May 1951 (flr, frt), J. Bosser 78 (P, TAN) ; Efoetsy, 12 May 1951 (flr), J. Bosser 116 (P), 126 (TAN); environs de Tulear, November 1956 (flr), J. Bosser 10598 (P, TAN) ; au nord du Fiherenana, 27 June 1958 (flr), R. Capuron I8623-SF (P) ; without locality, M. Cours 4643 (P) ; ca. 5 km S of Hotel Lakana Vezo in Ifaty toward Toliara (ca. 20 km N of Toliara along coast), ca. 0m, 5 November 1993 (flr), T. Daniel & M. Butterwick 6736 (CAS) ; ca. 28 km N of Toliara, just N of Ifaty along road to Ankaraobato, 23°6'S, VII Article Daniel et al., ANISOTES (ACANTHACEAE) IN MADAGASCAR 127 Homolle 1650 (P) ; environs de Tuléar, delta du Fiherenana, 6 August 1928 (flr, frt), H. Humbert & C. Swingle 5164 (P, TAN), 14--26 September 1924 (flr), H. Humbert et al. 2425 (P) ; du Lac Manampetsotsa au delta de La Linta (Cote Sud-Ouest), 1--10 m, 17--24 August 1928 (flr), H. Humbert & C. Swingle 5295 (P, UC); Anakao, 3 September 1967 (flr), B. Koechlin 1 (P) ; Manombo, Anjevo, 4 November 1967 (flr), B. Koechlin 2 (P) ; Songeritelo, 23 September 1967 (sterile), B. Koechlin 25 (P) ; environs de Tuléar, route de Manombo, 3 November 1960 (flr), J. Leandri & R. Jean de Dieu 3626 (P), 3634 (P) ; 21 km N of Ifaty on track from Manambo to Tsifota, near Fiserenamasay, 22°48'S, 43°18'E, 0 m, 4 August 1993 (flr), B. Lewis et al. 515 (TAN); Tsivonoa N., 11 September 1961 (flr), J. Peltier & M. Peltier 3384 (P, TAN) ; environs de Tulear, August 1919 (flr, frt), H. Perrier de la Bâthie 19811 (P) ; 20km N of Tulear, near resort of Mora Mora, Ifaty [«Italy»], 23°11'S, 43°07'E, 10 m, 7 May 1987 (flr), P. Phillipson 1776 (CAS, P, TAN) ; N of Itampolo, 24°41'S, 43°57'E, 10 October 1990 (flr), P. Phillipson et al. 3739 (CAS, P, TAN) ; Befanany, 2 August 1921 (flr), H. Poisson 277 (P) ; outskirts of village of Itampolo, 24°42'S, 43°57'E, 0 m, 9 February 1990 (flr), B. du Puy et al. MB686 (P, TAN) ; NW of Toliara, Forest of Mikea, ca. 13 km N of mouth of Manombo River, 22°53'S, 43°24'E, 10 m, 4 December 1993 (flr), D. du Puy et al. M691 (P, TAN) ; Tulear, Raich 1097 (TAN); District Tulear, Réserves Naturelles, 22 May 1951 (flr), L. de G. Ravelonanahary 2787-RN (P, TAN) ; District Betioky, Réserves Naturelles, 31 December 1951 (flr), L. de G. Ravelonanahary 3286-RN (P) ; District Betioky, Réserves Naturelles, L. de G. Ravelonanahary 3926-RN (P, TAN) ; District Betioky, Réserves Naturelles, July 1952 (flr), 4159-RN (P, TAN) ; District Tulear, Réserves Naturelles, 24 October 1952 (flr), L. de G. Ravelonanahary 4287-RN (P, TAN) ; 35 km N of Toliara along coast road, 23°04'S, 43°35'E, 20 m, 12 December 1988 (flr), G. Schatz & J. Miller 2471 (CAS) ; ca. 19 km NNW of Tulear on RN 9, ca. 2 km N of Songoritela toward Morombe, 0 m, 9 June 1991 (flr), 23°12'S, 43°37'E, J. Zarucchi et al. 7575 (TAN). Anisotes divaricatus TF Daniel, Mbola, Almeda, and Phillipson, sp. Nov. TYPE: MADAGASCAR. Toliara: Réserve du Lac Tsimanampetsotsa (No. 10), NW corner of reserve, 24°04'S, 43°46'E, escarpment of calcareous plateau, 50 m, 8 October 1990, P. Phillipson et al. 3721 (holotype: TAN!; isotypes: CAS!, P!). Figures 1, 4. Frutex usque ad 1 m, ramis vetustis #177; divaricatis. Folia petiolata, laminae subsucculentae, late ovatae vel subcirculares vel obcordatae vel obdeltatae vel oblatae, 5-11 X 4.5-11 mm, 0.7-1.3-plo longiores quam latiores, emarginatae vel truncatae ad apicem, costa solum manifesta (in pagina abaxiali). Spicae axillares, pedunculares. Bracteae triangulares vel late triangulares, 1-1.3 mm longae. Calyx 1.3-2.7 mm longus. Corolla rubra-purpurea intus, 27-36 mm longa. Capsula 13-20 mm longa, trichomatibus eglandulosis nullis. Semina 3.8-4.8 mm longa. Scrambling to clambering or arching shrub to 1 m; older branches #177; divaricate. Young stems quadrate-ridged [similar to subhexagonal but only small valleys at corners], densely and evenly pubescent with (flexuose to) antrorse to antrorsely appressed eglandular trichomes 0.2-0.4 mm long, resulting in a grayish cast. Leaves petiolate, petioles to 5 mm long, blades subsucculent, broadly obovate to subcircular to obcordate to obdeltate to oblate, 5-14 mm long, 4.5-14.3 mm wide, 0.7-1.3 times longer than wide, acute at base, emarginate to truncate at apex, venation obscure, midvein only evident on abaxial surface, surfaces and margin #177; densely and evenly pubescent with cauline type trichomes, resulting in a grayish cast. Inflorescence of #177; congested, axillary (in axils of distalmost 1-3 pairs of leaves), pedunculate spikes; peduncles 2-11 mm long, rachis densely to sparsely pubescent like young stems, internodes to 1 mm long. Bracts opposite, all fertile, each subtending a single flower, triangular to broadly triangular, 1-1.3 mm long, 0.8-1.2 mm wide, abaxial surface densely pubescent with (flexuose to) antrorse to antrorsely appressed eglandular VIII Article 128 PROCEDING OF THE CALIFORNIA ACADEMY OF SCIENCES Fourtth series, Volume 58, n°8 trichomes 0.1-0.2 mm long. Bracteoles absent. Flowers sessile to subsessile (i.e., pedicels to 0.3 mm long). Calyx 5-lobed, 1.3-2.7 mm long, tube shorter than lobes, lobes triangular to lanceolate to ovate to obovate, 1-1.7 mm long, 0.7-1 mm wide, 0.63-0.87 times as long as calyx, abaxial surface sparsely pubescent (especially distally) with antrorse to antrorsely appressed eglandular trichomes 0.05-0.1 mm long, margin eciliate to sparsely ciliate with flexuose to antrorse eglandular trichomes. Corolla #177; concolorous, dull purple-red (maroon), or #177; discolorous with the tube pink-brown (bronze colored) and the lobes of the lower lip maroon, externally mostly glabrous (occasionally with a few flexuose to retrorse eglandular trichomes to 0.2 mm long
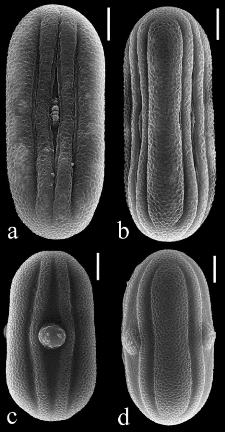
Figure 3. Pollen of Anisotes divaricatus (a, b; Almeda 9237) and A. madagascariensis (c,d; Daniel et al. 10422). a. apertural view. b. interapertural view. c. apertural view. d. interapertural view. Scale lines = 10 ìm. proximally),27-40 mm long, tube 14-20 mm long, narrow proximal portion subcylindric, 4-7 mm long, #177; abruptly expanded into a funnelform throat, throat 9-14 mm long, 2.8-4 mm I diameter near midpoint, corolla tube:corolla = 0.46-0.56, upper lip (11-) 14-18 mm long, internally rugulate, entire to emarginate at apex, lacking a pale colored margin distally, lower lip recurved to reflexed, 11-19 mm long, lobes linear, 8-12 mm long, 1-4 mm wide, central lobe larger than lateral lobes. Stamens 15-18.5 mm long, inserted near apex of corolla tube, exserted from mouth of corolla but usually not surpassing upper lip (or surpassing it by up to 1 mm), filaments cream, glabrous, thecae yellow and brown, unequal in size (distal larger, 2.4-3.3 mm long; proximal shorter, 2-2.8 mm long), unequally inserted (overlapping by 0.8-1.2 mm), parallel to slightly oblique, glabrous, lacking basal appendages or proximal theca with an inconspicuous basal appendage 0.05 mm long; pollen 72 ìm polar diameter (P), 28-30.5 ìm equatorial diameter (E), P:E= 2.4-2.6. Style 24-32 mm long, glabrous, stigma 0.2-0.3 mm long, lobes obscure. Nectar disk asymmetric, often with a lobe-like appendage to 0.6 mm long protruding between outermost (i.e., farthest from rachis) pair of calyx lobes. Capsule 13-20 mm long, surface #177; scurfy but lacking noticeable eglandular trichomes, stipe 5-8 mm long, head ovoid-ellipsoid to obovoid-ellipsoid with a slight medial constriction. Seeds 4, discoid, 3.8-4.8 mm long, 3.7-5.1 mm wide, surfaces minutely papillose or rugose, shiny, margin sometimes #177; swollen and smooth. IX Article Daniel et al., ANISOTES (ACANTHACEAE) IN MADAGASCAR 129 Phenology. Flowering : August-May; fruiting : August-October, April. Distribution and habitat. Endemic to southwestern Madagascar; plants occur on limestone rock (Mahafaly Plateau limestone) with spiny forest (dominated by Alluaudia comosa, Commiphora madagascariensis, Delonix floribunda, D. adansonioides, Adansonia rubrostipa, Euphorbia oncoclada, E. laroo, and Uncarina stellulifora) in the vicinity of Lake Tsimanampetsotsa at elevations from 22--50 m (Fig. 2).
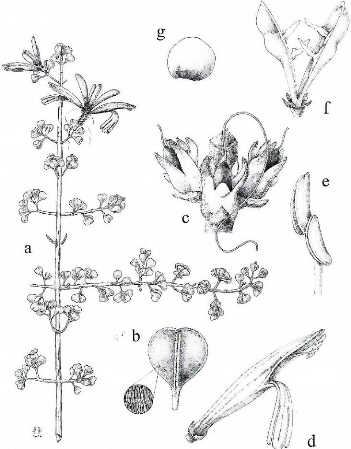
Figure 4. Anisotes divaricatus. a. Habit (Leandri & Saboureau 4441), ×0.5. b. Leaf (abaxial surface), ×2.1, with close-up of pubescence, ×16 (Leandri & Saboureau 4441). c. Inflorescence nodes with corollas removed (Bosser 1762), ×8. d. Flower (Bosser 15609 & Perrier de la Bâthie 19156), ×1.7. e. Distal portion of stamen (Perrier de la Bâthie 19156), ×7.8. f. Capsule (Perrier de la Bâthie 19156), ×3. g. Seed (Perrier de la Bâthie 19156), ×3.5. Drawn by Erin Hunter. X Article 130 PROCEDING OF THE CALIFORNIA ACADEMY OF SCIENCES Fourtth series, Volume 58, n°8 Anisotes divaricatus is both morphologically and ecologically distinct from A. madagascariensis. However, based on its spicate inflorescence, bracts lacking reticulate venation, and 3-aperturate pollen (Fig. 3), A. divaricatus would also appear to belong to section Spiciflori. Like A. madagascariensis and unlike A. formosissimus, the only other species of section Spiciflori, A. divaricatus lacks bracteoles. Indeed, the two Malagasy species are the sole representatives of the genus in which these structures are not present. Both Malagasy species occur in the vicinity of Lake Tsimanampetsotsa and the general collecting locales are sometimes identical for each (e.g., «Efoetsy» or «near Lake Tsimanampetsotsa»). In most such instances, however, plants of A. madagascariensis were noted to have been collected from maritime sands or dunes, and at lower elevations than those of A. divaricatus. A collection from a rocky forest on «Plateau Mahafaly» (Perrier de la Bâthie 9521 at P) contains a small branch that likely pertains to A. divaricatus. Like that species, it has round to oblate leaves on petioles up to 5 mm long and corollas with the tube:corolla=0.47. However, it differs from that species by its slightly longer vegetative trichomes (up to 0.5 mm long), larger leaves (up to 15 × 17 mm), smaller corollas (to 17 mm long) with the tube to 8 mm long and not or but barely expanded distally, and smaller anther thecae (1.3--1.5 mm long). Until additional material is available that resembles this collection or that links it to specimens noted herein, Perrier de la Bâthie 9521 is not included within the circumscription of A. divaricatus. Paratypes. MADAGASCAR. Toliara: NE sector of Tsimanampetsotsa National Park, 24°02.895'S, 43°45.138'E, ca. 22 m, 25 August 2006 (flr, frt), F. Almeda 9237 (CAS, TAN); Efoetsy, 12 May 1951 (flr), J. Bosser 187 (P) ; Lac Tsimanampetsotsa, R.N. 10, February 1962 (flr), J. Bosser 15,609 (P, TAN) ; Réserve 10, Lac Tsimanampetsotsa, February 1962 (flr), M. Keraudren 1403 (P); environs du Lac Tsimanampetsa, environs de l'aven de Mitoho, 23 November 1960 (flr), J. Leandri & P. Saboureau 4441 (P); Manampetsa, April 1933 (flr, frt), H. Perrier de la Bâthie 19156 (P); Réserve de Tsimanampetsotsa, SW corner of reserve, 24°11'S, 43°48'E, 50 m, 13 January 1989 (flr), P. Phillipson & S. Rabesihanaka 3169 (CAS, P, TAN). ACKNOWLEDGMENTS Field studies in 2003 were supported by a grant from the National Science Foundation (DEB 0108589) to L.A. McDade and T.F. Daniel. We thank De Ada and Henry Mally for photographing A. divaricatus at Lake Tsimanampetsotsa; Heritiana Ranarivelo, Lucinda McDade, and Randriambololona Tantely Nirina for assisting with field work and logistics in Madagascar; and Erin Hunter, the 2006 Fellows Intern in Biological Illustration at the California Academy of Sciences, for her exceptional drawing, which helped to relocate the new species at Lake Tsimanampetsotsa in 2006. We are grateful for courtesies extended by the following herbaria from which specimens were examined: CAS, P, TAN, and UC. We also thank MO for sending duplicate specimens of Anisotes and other Malagasy Acanthaceae to CAS. The staff at G kindly provided a digital image of the type of Symplectochilus madagascariensis. XI Article Daniel et al., ANISOTES (ACANTHACEAE) IN MADAGASCAR 131 LITERATURE CITED BADEN, C. 1981. The genus Anisotes (Acanthaceae), a taxonomic revision. Nordic Journal of Botany 1:623--664. BENOIST, R. 1929. Descriptions d'espèces nouvelles d'acanthacées de Madagascar. Bulletin de la Société Botanique de France 76:1031--1038. DANIEL, T.F. 2000. Additional chromosome numbers of American Acanthaceae. Systematic Botany 25:15-25. DANIEL, T.F. 2007. Chromosome numbers of miscellaneous Malagasy Acanthaceae. Brittonia 58: in press. DANIEL, T.F. AND T.I. CHUANG. 1998. Chromosome numbers of cultivated Acanthaceae and systematic implications. Pp. 309--330. In : P. Mathew and M. Sivadasan (eds.), Diversity and Taxonomy of Tropical Flowering Plants. Mentor Books, Calicut. DANIEL, T.F., K. BAKLWILL, AND M.-J. BALKWILL. 2000. Chromosome numbers of South African Acanthaceae. Proceedings of the California Academy of Sciences 52:143--158. ELLIS, J.L. 1962. Chromosome numbers in some members of Acanthaceae. Science & Culture 28:191--192. GAUTIER, L. 2002. Liste commentée des phanérogames de la Réserve Spéciale de Manongarivo, Madagascar. Boissiera 59 : 105--239. KIEL, C.A., L.A. MCDADE, AND T.F. DANIEL. 2006. The species-rich and biologically diverse «justicioids» (Acanthaceae : Justicieae) : phylogeny, biogeography, and morphological evolution. Botany 2006: Abstracts, Scientific Meeting, Abstract 622:284. KRISHNAPPA, D.G. AND R.M. RANGANATH. 1982. Acanthaceae. Pp. 364--365. In: A. Löve (ed.) IOPB chromosome number reports LXXV. Taxon 31:342--368. LINDAU, G. 1898. Einige neue acanthaceen. Annuaire du Conservatoire et du Jardin Botaniques de Genève 2:1--140. Mc.DADE, L. A., T.F. DANIEL, S.E. MASTA, AND K.M. RILEY. 2000. Phylogenetic relationships within the tribe Justicieae (Acanthaceae): evidence from molecular sequences, morphology, and cytology. Annals of the Missouri Botanical Garden 87:435-- 458. MULLER, J., M. SCHULLER, H. STRAKA, AND B. FRIEDRICH. 1989. Palynologia Madagassica et Mascarenica. Fam. 182 : Acanthaceae. Tropische und Subtropische Pflanzenwelt 67:138--187. NARAYANAN, C.R. 1951. Somatic chromosomes in the Acanthaceae. Journal of the Madras University 21:220--231. RANGANATH, R.M. 1981. Morphological and cytological studies in Acanthaceae. Ph.D. Thesis, Bangalore. [not seen, cited in : Monographs in Systematic Botany from the Missouri Botanical Garden 8:33. 1984] SAGGOO, M.I.S. 1983. Cytomorphological studies on plants of economic importance of Bicarpellatae from India. Ph.D. Thesis, Punjabi University, Patiala. [not seen, cited in: Monographs in Systematic Botany from the Missouri Botanical Garden 13:18. 1985] SUBRAMANIAN, D. AND T. GOVINDARAJAN. 1980. Cytotaxonomy of some species of Acanthaceae. Journal of Cytology and Genetics 15:90--92. Copyriths (c) 2007 by the California Academy of Sciences
XII Poster POSTER : Mbola, V. A. B., Daniel, T.F., Letsara, R., Ramavovolona, Faramalala, M.H., Distribution des représentants de la famille des Acanthaceae dans le Sud de Madagascar. AETFAT, 2010 Antananarivo.
XIII Annexe |
|