|

UNIVERSITE D'ANTANANARIVO
DOMAINE DES SCIENCES ET TECHNOLOGIES
ECOLE DOCTORALE EN SCIENCES DE LA VIE ET DE
L'ENVIRONNEMENT
THESE POUR L'OBTENTION DU DIPLOME DE DOCTORAT
EN
SCIENCES DE LA VIE ET DE L'ENVIRONNEMENT
SPECIALITE : SCIENCES DU
VEGETAL
INVENTAIRE SYSTEMATIQUE DES GENRES ET ESPECES,
REPRESENTANTS
DE LA FAMILLE DES ACANTHACEAE DANS LES
DISTRICTS DE TOLAGNARO ET
D'AMBOASARY-ATSIMO,
MADAGASCAR

Présentée par : MBOLA VERSENE Andrianandrasana
Balzac
soutenue publiquement, le 12 décembre 2018 devant le jury
composé de :
Président : Professeur RAMAVOVOLOLONA
Directeur de thèse : Professeur Miadana Harisoa
FARAMALALA
Rapporteur interne : Professeur Vonjison RAKOTOARIMANANA
Rapporteur externe : Professeur Samuel RAZANAKA
Examinateur : Professeur Bakolimalala RAKOUTH


UNIVERSITE D'ANTANANARIVO
DOMAINE DES SCIENCES ET TECHNOLOGIES
ECOLE DOCTORALE EN SCIENCES DE LA VIE ET DE
L'ENVIRONNEMENT
THESE POUR L'OBTENTION DU DIPLOME DE DOCTORAT
EN
SCIENCES DE LA VIE ET DE L'ENVIRONNEMENT
SPECIALITE : SCIENCES DU VEGETAL
INVENTAIRE SYSTEMATIQUE DES GENRES ET ESPECES,
REPRESENTANTS
DE LA FAMILLE DES ACANTHACEAE
DANS LES DISTRICTS DE TOLAGNARO
ET
D'AMBOASARY-ATSIMO, MADAGASCAR
Présentée par : MBOLA VERSENE Andrianandrasana
Balzac
soutenue publiquement, le 12 décembre 2018 devant le jury
composé de
Président : Professeur RAMAVOVOLOLONA
Directeur de thèse : Professeur Miadana Harisoa
FARAMALALA
Rapporteur interne : Professeur Vonjison RAKOTOARIMANANA
Rapporteur externe : Professeur Samuel RAZANAKA
Examinateur : Professeur Bakolimalala RAKOUTH

i
REMERCIEMENTS
La présente étude est le fruit de la
collaboration entre California Academy of Sciences de San Francisco (CAS),
Madagascar National Parks (MNP) et le Ministère de l'Ecologie, de
l'Environnement et des Forêts (MEEF) dans le cadre de l'évaluation
des ressources naturelles dans les aires protégées à
Madagascar.
- Mes vifs remerciements vont à Madame Le
Professeur RAMAVOVOLOLONA, Enseignant Chercheur et Responsable de
l'Equipe d'Accueil de la spécialité « Sciences du
Végétal » de l'Université d'Antananarivo et qui a
bien voulu faire partie du membre du jury et elle m'a aussi fait un grand
honneur d'en assurer la présidence.
- J'exprime ma vive reconnaissance à Madame
Le Professeur Miadana Harisoa FARAMALALA, Enseignant Chercheur
à l'Université d'Antananarivo, de bien vouloir accepté
d'être mon encadreur malgré ses occupations. Je suis très
reconnaissant pour les conseils et aussi le temps que vous m'avez
accordés.
- Monsieur Le Professeur Vonjison
RAKOTOARIMANANA, Enseignant Chercheur à
l'Université d'Antananarivo, pour avoir accepté d'être mon
rapporteur , mes vifs remerciements s'adressent à vous. Je suis
très honoré de vous avoir parmi le jury de ma
thèse.
- Madame Le Professeur Bakolimalala
RAKOUTH, Enseignant Chercheur à l'Université
d'Antananarivo, qui me fait l'honneur d'évaluer et d'examiner ce
travail. Merci infiniment pour le temps que vous avez alloué à la
lecture, la correction et l'amélioration de cette thèse ,
- Monsieur Le Professeur Samuel
RAZANAKA, Chercheur au Centre National de Recherche sur
l'Environnement (CNRE), pour avoir accepté d'évaluer mon travail
et siéger parmi les membres du jury , je vous adresse mes
sincères remerciements.
Je remercie tous mes professeurs depuis ma première
année d'études. Je suis très heureux de leur exprimer ma
gratitude et mes reconnaissances ,
Je remercie le California Academy of Sciences de San
Francisco représenté par Monsieur Franck ALMEDA,
PhD, Curateur Emérite du Centre de Recherche en Botanique de
ii
California Academy of Sciences, San Francisco Californie,
qui a accepté de financer ce travail de recherche, dans le cadre de la
collaboration citée ci-dessus.
Je remercie vivement Monsieur Daniel THOMAS,
PhD, Curateur Emérite du Centre de Recherche en Botanique de
California Academy of Sciences, San Francisco (CAS), Californie, qui a
accepté, malgré ses lourdes tâches et
responsabilités, de m'aider dans la réalisation de ce travail. Je
ne vous exprimerais jamais assez toute ma reconnaissance pour cette
thèse.
Mes remerciements s'adressent aussi à :
-Monsieur Guy Suzon RAMANGASON,
Directeur Général de Madagascar National Parks ; qu'il trouve
à travers ce document un outil de travail pour la gestion durable de ce
beau parc.
-L'équipe de California Academy of Sciences de San
Francisco et celle d'Antananarivo, en particulier à Messieurs
Heritiana RANARIVELO et Rokiman LETSARA qui m'ont
aidé du point de vue logistique lors des études sur le
terrain.
Le laboratoire de Botanique du Parc Botanique et
Zoologique de Tsimbazaza à Antananarivo et son équipe, pour les
précieuses aides qu'ils m'ont apportées.
-Monsieur Roger Lala ANDRIAMIARISOA,
qui a bien voulu assurer les dessins des plantes pour
l'illustration des descriptions, malgré ses lourdes taches.
-Monsieur Refela Ernest RAZAKANJOELINA
et Monsieur Rabenandrasana Pierrot MONJA,
Directeurs successifs du Parc National d'Andohahela et toute leur
équipe. Qu'ils trouvent à travers ce mémoire un outil pour
valoriser durablement les ressources naturelles au profit de
l'écotourisme et de l'éducation relative à
l'environnement.
Les communautés locales du district de Tolagnaro qui
m'ont accueilli dans leurs villages ; je leur exprime toute ma
gratitude.
Tous mes collègues et amis pour leur encouragement et
leurs conseils.
Toute ma famille qui m'a soutenu sans limite pour parvenir
à ce stade.
Enfin, je tiens à remercier tous ceux qui ont
contribué, de près ou de loin à la réalisation de
ce travail.
A toutes et à tous, merci !
iii
TABLE DES MATIERES
Remerciements ... i
Liste des tableaux vii
Liste des cartes vii
Liste des figures viii
Liste des photographies ix
Acronymes et abréviations xi
Glossaire ... xii
INTRODUCTION 1
I. CONTEXTE GENERAL ET MILIEU D'ETUDE 3
I.1. ETAT DES CONNAISSANCES SUR LES ACANTHACEAE 3
I.2. MILIEU D'ETUDE 4
I.2.1. Localisation géographique de la zone d'étude
5
I.2.2. Facteurs abiotiques 5
I.2.2.1. Géologie, géomorphologie et sol 5
I.2.2.2. Hydrographie 8
I.2.2.3. Climats 9
I.2.3. Facteurs biotiques 12
I.2.3.1. Végétation et flore 13
I.2.3.2. Faune 16
I.2.3.3. L'homme et ses activités 17
II. MATERIELS ET METHODES 22
II.1. FAMILLE DES ACANTHACEAE 22
II.1.1. Classification 22
II.1.2. Morphologie des Acanthaceae 23
II.2. MÉTHODES D'ETUDE 25
II.2.1. Travaux sur le terrain 25
II.2.2. Travaux en laboratoire 28
II.2.3. Traitement des données 29
III. RÉSULTATS ET INTERPRETATIONS 31
III.1. RESULTATS DES ENQUETES ETHNOBOTANIQUES 31
III.2. DESCRIPTION DES HABITATS DE LA ZONE D'ETUDE 32
III.2.1. Habitats de la partie humide 32
III.2.2. Habitats de la zone de transition 33
III.2.3. Habitats de la partie sèche 33
III.3. MORPHOLOGIE ET BIOLOGIE DES ESPECES ETUDIEES 34
III.3.1. Caractéristiques morphologiques des Acanthaceae
34
III.3.1.1. Type biologique 34
III.3.1.2. Appareil végétatif 36
III.3.1.3. Appareil reproducteur 40
III.3.2. Clé de détermination des genres 44
III.4. DESCRIPTIONS DES TAXONS 47
III.4.1. Asystasia Jussieu 47
III.4.1.1. Asystasia gangetica (L.) Anders ex Thwaites
49
III.4.2. Barleria Linné 52
III.4.2.1. Barleria alluaudii Benoist 52
III.4.2.2. Barleria decaryi Benoist 54
III.4.2.3. Barleria humbertii Benoist 57
III.4.2.4. Barleria her Benoist 60
III.4.2.5. Barleria kitchingii Baker 62
III.4.3. Blepharis Jussieu 65
III.4.3.1. Blepharis calcitrapa var. decaryi
Benoist. 66
III.4.3.2. Blepharis maculata Benoist 68
III.4.4. Brillantaisia Beauv. 71
III.4.4.1. Brillantaisia madagascariensis Anders. ex
Lindau 71
III.4.5. Camarotea Scott-Elliot 74
III.4.5.1. Camarotea sp. 74
III.4.6. Crossandra Salisb. 77
III.4.6.1. Crossandra longipes Benoist. 77
III.4.7. Dicliptera Jussieu 79
II.4.7.1. Dicliptera decaryi Benoist 80
III.4.8. Ecbolium Kurz 82
III.4.8.1. Ecbolium madagascariense Vollesen 82
III.4.8.2. Ecbolium syringifolium (Vahl) Vollesen 85
III.3.9. Forsythiospis Baker 87
III.3.9.1. Forsythiopsis vincoides Benoist 88
III.3.10. Hypoestes Solander ex Brown 90
III.3.10.1. Hypoestes decaryana Benoist 91
III.3.10.2. Hypoestes longituba Benoist 93
III.3.10.3. Hypoestes poissonii Phillipson 96
III.3.10.4. Hypoestes strigata 98
III.3.10.5. Hypoestes sp. 1 100
iv
v
III.3.10.6. Hypoestes sp. 2 102
III.3.10.7. Hypoestes sp. 3 105
III.3.11. Justicia Linnaeus 108
III.3.11.1. Justicia sp. 1 108
III.3.12. Lasiocladus Bojer ex Ness 110
III.3.12.1. Lasiocladus anthospermifolius (Bojer et
Ness) Benoist 111
III.3.13. Mendoncia Velloso in Vandelli 113
III.3.13.1. Mendoncia delphina Magnaghi 114
III.3.14. Neuracanthus Ness. 117
III.3.14.1. Neuracanthus richardianus (Nees) Boivini ex.
Benoist 117
III.3.15. Phaulopsis Willdenow 119
III.3.15.1. Phaulopsis imbricata ssp.
madagascariensis M. Manktelow 120
III.3.16. Ruellia Benoist 123
III.3.16.1. Ruellia geayi var. tomentosus
Benoist 123
III.3.17. Stenandriopsis Moore 125
III.3.17.1. Stenandriopsis carduacea Benoist 125
III.3.18. Les Acanthaceae de l'herbarium de Tsimbazaza 128
III.4. RELATIONS ENTRE LES ACANTHACEAE ET LES
FACTEURS
ECOLOGIQUES 129
III.4.1. Répartition des espèces selon le
paramètre `subdivision climatique' 129
III.4.2. Altitude 131
III.4.3. Ouverture du milieu 133
III.4.4. Sols 135
III.4.5. Relations entre les espèces et les
paramètres écologiques des sites de relevé 136
IV. DISCUSSIONS ET RECOMMANDATIONS 140
IV.1. DISCUSSIONS SUR LA METHODOLOGIE 140
IV.2. RARETE DE LA RECHERCHE SUR LES ACANTHACEAE A
MADAGASCAR 141
IV.3. APPLICATIONS 141
CONCLUSION 144
REFERENCES BIBLIOGRAPHIQUES 148
ARTICLE I
POSTER XII
ANNEXE 1 : QUESTIONNAIRE D'ENQUETE XIII
vi
ANNEXE 2 : REPARTITION DES ESPECES RENCONTREES EN
FONCTION DES
PARAMETRES DU MILIEU. XIV
vii
LISTE DES TABLEAUX
Tableau 1 : Températures et précipitations de la
zone d'étude 10
Tableau 2 : Températures et précipitations de
Tolagnaro 11
Tableau 3 : Températures et précipitations de
Behara moyenne pour l'anné 1981-2010. 12
Tableau 4 : Les classifications de la famille des Acanthaceae
22
Tableau 5 : Les utilisations des plantes par la population 31
Tableau 6 : Distribution des espèces par `type biologique'
35
Tableau 7 : Répartition des espèces par rapport
à la présence de poils 38
Tableau 8 : Répartition des espèces par rapport
à la présence des épines 40
Tableau 9 : Répartition des espèces par rapport aux
types d'inflorescence 41
Tableau 10 : Répartition des espèces par classe de
diamètre de graines 43
Tableau 11 : Liste des taxa collectés 48
Tableau 12 : Liste des espèces identifiées 128
Tableau 13 : Répartition des espèces selon la
`subdivision climatique' des sites de relevé 129
Tableau 14 : Indice de similitude de Jaccard des sites de
relevé pour le paramètre `subdivision
climatique' 131
Tableau 15 : Répartition des espèces en fonction de
l'altitude des sites de relevé 132
Tableau 16 : Représentation des indices de Jaccard pour le
paramètre `altitude' 133
Tableau 17 : Répartition des espèces en fonction du
paramètre `ouverture du milieu' des sites
de relevé 134
Tableau 18 : Indices de Jaccard des sites
de relevé pour le paramètre `ouverture du milieu' 134 Tableau 19
: Répartition des espèces par rapport au paramètre `type
de sol' des sites de
relevé 145
Tableau 20 : Indice de Jaccard des sites de relevé pour le
paramètre `type de sol' 136
LISTE DES CARTES
Carte 1 : Carte de localisation sur fond de
végétation 6
Carte 2 : Extrait du croquis géologique de la
région des Chaînes Anosyennes 7
Carte 3 : Extrait de la carte des réseaux hydrographiques
et principaux bassins fluviaux de
Madagascar (Chaperon et al. 1993, modifié par
Mbola, 2018) 9
Carte 4 : Les zones de collectes 26
Carte 5 : Répartition des Acanthaceae dans les districts
de Tolagnaro et d'Amboasary-
Atsimo 130
LISTE DES FIGURES
Figure 1 : Diagramme ombrothermique de Tolagnaro, selon
Gaussen 11
Figure 2 : Diagramme ombrothermique de Behara, selon Gaussen
12
Figure 3 : Représentation graphique des espèces
par type biologique. 34
Figure 4 : Répartition des espèces par type de
tige vue en section 37
Figure 5 : Répartition des espèces par la
présence ou non de poils 37
Figure 6 : Répartition des espèces par la
présence ou non d'épines 39
Figure 7 : Répartition des espèces par type
d'inflorescence 42
Figure 8 : Répartition des espèces par classe de
diamètre des graines 42
Figure 9 : Asystasia gangetica Anders et Thwaites
51
Figure 10 : Barleria alluaudii Benoist 54
Figure 11 : Barleria decaryi Benoist 56
Figure 12 : Barleria humbertii Benoist 59
Figure 13 : Barleria her Benoist 61
Figure 14 : Barleria kitcthingii Baker 64
Figure 15 : Blepharis calcitrapa Benoist var.
decaryi Benoist. 67
Figure 16 : Blepharis maculata Benoist. 70
Figure 17 : Brillantaisia madagascariensis Anders. ex
Lindau 73
Figure 18 : Camarotea sp. 76
Figure 19 : Crossandra longipes Benoist 78
Figure 20 : Dicliptera decaryi Benoist 81
Figure 21 : Ecbolium madagascariense Vollesen 84
Figure 22 : Ecbolium syringifolium (Vahl) Vollesen
86
Figure 23 : Forsythiopsis vincoides Benoist 89
Figure 24 : Hypoestes decaryana Benoist 92
Figure 25 : Hypoestes longituba Benoist 95
Figure 26 : Hypoestes poissonii Phillipson 97
Figure 27 : Hypoestes strigata 99
Figure 28 : Hypoestes sp. 1 102
Figure 29 : Hypoestes sp. 2 104
Figure 30 : Hypoestes sp. 3 106
Figure 31 : Justicia sp. 1 109
Figure 32 : Lasiocladus anthospermifolius Benoist.
112
Figure 33 : Mendoncia delphina Magnaghi. 116
Figure 34 : Neuracanthus richardianus Benoist 119
viii
Figure 35 : Phaulopsis imbricata ssp.
madagascariensis M. Manktelow 122
Figure 36 : Ruellia geayi var. tomentosus
Benoist 124
Figure 37 : Stenandriopsis carduacea R Benoist 127
Figure 38 : Répartition des espèces selon le
paramètre `subdivision climatique' des sites de
relevé 131
Figure 39 : Représentation graphique
de la répartition des espèces en fonction de l'altitude 132
Figure 40 : Répartition des espèces en fonction de
la variable `ouverture du milieu' 133
Figure 41 : Répartition des espèces en fonction du
type de sol 136
Figure 42 : Représentations géométriques des
espèces et des paramètres écologiques des sites
de relevé. 138
LISTE DES PHOTOGRAPHIES
Photo 1 : Forêt dense humide, au col de Manangotry, vue
d'Androangaloaka (C.R. Isaka
Ivondro) 14
Photo 2 : Fourré à Dypsis decaryi (C.R.
Ranopiso) 15
Photo 3 : Haut fourré à Alluaudia ascendens
(C.R. Ankariera Bevilany). 16
Photo 4 : Défrichement suivi de mise à feu dans la
vallée de Fanota, C.R. Isaka Ivondro (fôret
humide) 17
Photo 5 : Terrains de cultures sur les flancs des
collines (Village de Mahatsinjo, C.R. Ranopiso)
18
Photo 6 : Défrichement dans la localité de Bezaha,
C.R. Ambatoabo (fourré) 18
Photo 7 : Feux de renouvellement de pâturage au niveau des
savanes en moyenne altitude, zone
de Malio, C.R. Ranopiso 19
Photo 8 : Vente de viande de sanglier à Analamantsaky
(C.R. Ankariera Bevilany) 20
Photo 9 : Toiture de case faite en feuilles de Dypsis decaryi
et murs en planches d'Alluaudia
procera (Village d'Analamantsaky, C.R. Ankariera
Bevilany) 21
Photo 10 : Fragment de tige de Asystasia gangetica
Anders et Thwaites 69
Photo 11 : Fleur de Asystasia gangetica Anders et
Thwaites 69
Photo 12 : fragment de tige de Barleria alluaudii
Benoist 72
Photo 13 : Fleur de Barleria alluaudii Benoist 72
Photo 14 : Fragment de tige de Barleria decaryi Benoist
76
Photo 15 : Fleur de Barleria decaryi Benoist 76
Photo 16 : Fragment de tige de Barleria humbertii
Benoist 77
Photo 17 : Fleur de Barleria humbertii Benoist 77
Photo 18 : Fragment de tige et inflorescence de Barleria her
Benoist 81
Photo 19 : Fleur de Barleria her Benoist 81
ix
Photo 20 : Fragment de tige Barleria kitcthingii
Baker 82
Photo 21 : Fleur de Barleria kitcthingii Baker 82
Photo 22 : Fragment de tige Blepharis calcitrapa
Benoist 87
Photo 23 : Fleur de Blepharis calcitrapa Benoist
87
Photo 24 : Inflorescence de Brillantaisia madagascariensis
Anders. ex Lindau 91
Photo 25 : Fleur de Brillantaisia madagascariensis
Anders. ex Lindau 91
Photo 26 : Fragment de tige Camarotea sp. 94
Photo 27 : Fleur de Camarotea sp. 94
Photo 28 : Fragment de tige et inflorescence de Crossandra
longipes Benoists 98
Photo 29 : Fleur de Crossandra longipes Benoists
98
Photo 30 : Inflorescence de Dicliptera decaryi
Benoist 101
Photo 31 : Fleur de Dicliptera decaryi Benoist 101
Photo 32 : Ecbolium madagascariense Vollesen 102
Photo 33 : Fragment de tige et inflorescence de Ecbolium
syringifolium (Vahl) Vollesen 106
Photo 34 : Fleur de Ecbolium syringifolium (Vahl)
Vollesen 106
Photo 35 : Fragment de tige et inflorescence de
Forsythiopsis vincoides Benoist 109
Photo 36 : Fleur de Forsythiopsis vincoides Benoist
109
Photo 37 : Fragment de tige et inflorescence de Hypoestes
decaryana Benoist 112
Photo 38 : Fleur de Hypoestes decaryana Benoist
112
Photo 39 : Fragment de tige et inflorescence de Hypoestes
longituba Benoist 113
Photo 40 : Boutons floraux de Hypoestes longituba
Benoist 113
Photo 41 : Hypoestes poissonii Phillipson 115
Photo 42 : Tige de Hypoestes strigata 117
Photo 43 : Fleur de Hypoestes strigata 117
Photo 44 : Fragment de tige de Hypoestes sp. 1
121
Photo 45 : Fleur et fruit de Hypoestes sp. 1 121
Photo 46 : Fragment de tige et inflorescence de Hypoestes
sp. 2 122
Photo 47 : Fleur de Hypoestes sp. 2 122
Photo 48 : Fragment de tige et inflorescence de Hypoestes
sp. 3 126
Photo 49 : Inflorescence de Hypoestes sp. 3 126
Photo 50 : Fragment de tige et inflorescence de Justicia
sp. 1 129
Photo 51 : Inflorescence de Justicia sp. 1 129
Photo 52 : Fragment de tige et inflorescence de
Lasiocladus anthospermifolius Benoist. 132
Photo 53 : Fleur de Lasiocladus anthospermifolius
Benoist. 132
Photo 54 : Fragment de tige de Mendoncia delphina
Magnaghi. 134
x
xi
Photo 55 : Fruit de Mendoncia delphina Magnaghi.
134
Photo 56 : Fragment de tige et inflorescence de
Neuracanthus richardianus Benoist 138
Photo 57 : Inflorescence de Neuracanthus richardianus
Benoist 138
Photo 58 : Fragment de tige et inflorescence de Phaulopsis
imbricata ssp. madagascariensis
Manktelow 140
Photo 59 : Fleur de Phaulopsis imbricata ssp.
madagascariensis Manktelow 140
Photo 60 : Inflorescence de Ruellia geayi var.
tomentosus Benoist 144
Photo 61 : Fragment de tige et inflorescence de
Stenandriopsis carduacea R Benoist 145
xii
ACRONYMES ET ABREVIATIONS
ADEMA : Aérodrome DE MAdagascar
ANGAP : Association Nationale pour la Gestion des Aires
Protégées
APG : Angiosperm Phylogeny Group
BRHAMS : Botanical Recherch and Herbarium Management System
BVM : Code d'herbiers de MBOLA Balzac
CAS-MAD : California Academy of Sciences, antenne
Madagascar
CAS-USA : California Academy of Sciences, United States of
America
FOFIFA : FOibe FIkarohana ho amin'ny Fampandrosoana ny eny
Ambanivohitra
FAPBM : Fondation pour les Aires Protégées et la
Biodiversité de Madagascar
GPS : Global Positioning System
GPM : Groupe des Spécialistes des Plantes de
Madagascar
IPNI : The International Plant Names Index
LRK : Code d'herbiers de LETSARA Rockiman
MINENVEF : Ministère de l'Environnement et des Eaux et
Forêts
MEF : Ministère de l'Environnement et des
Forêts
MEFT : Ministère de l'Environnement, des Forêts
et du Tourisme
MNHN : Muséum National d'Histoire Naturelle de Paris
MNP : Madagascar National Parks
NAP : Nouvelle Aire Protégée
ONE : Office National pour l'Environnement
PBZT : Parc Botanique et Zoologique de Tsimbazaza
PHBM : Projet d'Aménagement du Haut Bassin de
Mandrare
PN : Parc National
PN AHL : Parc National d'Andohahela
QMM : Qit Madagascar Minerals
REE : Résultat de l'Evaluation Environnementale
RIP : Route d'Intérêt Provincial
RN : Route Nationale
RNI : Réserve Naturelle Intégrale
SAPM : Système des Aires Protégées de
Madagascar
SIG : Système d'Information Géographique
TAN : Herbarium du Parc Botanique et Zoologique de
Tsimbazaza
xiii
GLOSSAIRE
Acuminé : Extrémité d'une feuille se
terminant en pointe effilée (par un acumen).
Anisophylle : Lorsque deux feuilles alternes ou opposées
sont nettement différentes
dans la taille ou la forme.
Apiculé : Muni d'une courte pointe.
Collatéraux : Ovules insérés côte
à côte.
Cupule : Structure en forme de petite tasse (exemple disque
cupuliforme).
Cyme : Inflorescence définie où l'axe principal se
termine par une fleur, qui
fleurit en premier, et sa croissance s'arrête.
Cystholithe : Petite masse de cristaux d'un sel de calcium
présente dans les tissus de
certaines feuilles.
Exserte : Qui s'étend au-delà des organes
enveloppants (généralement des
périanthes).
Deltoïde : Le limbe est attaché par la pointe du
delta.
Hirsute : Se dit d'un organe muni de longs poils très
fournis.
Mutique : Dépourvu d'arête ou de mucron.
Obdeltoïde : En forme de delta renversé.
Obelliptique : Elliptique, la partie attachée est plus
grande que l'apex.
Pubescent : Couvert de poils courts, fins et mous, plus ou moins
frisés ou sinueux.
Silique : Fruit sec déhiscent typique des Brassicaceae et
des Acanthaceae.
Tomenteux : Couvert de poils nombreux, mous et
enchevêtrés, comme un feutre.
Villeux : Ayant de longs poils doux, souvent courbés, mais
non feutrés.
1
Introduction
INTRODUCTION
La dislocation de bloc de Gondwana au début du
Crétacé et l'isolement de Madagascar du continent africain depuis
80 millions d'années ont doté la grande île d'une
inestimable richesse en biodiversité. Madagascar est connu et reconnu
mondialement pour ses valeurs biologiques et écologiques par sa flore
unique qui est très riche car elle comporte 80 % à 90 %
d'espèces endémiques ; et encore ces valeurs ne concernent que
les Phanérogames et les Ptéridophytes. Les plantes vasculaires
Malagasy, actuellement objet de publications, constituent 80% des familles
existantes à Madagascar (Goodman, 2008). Les estimations du nombre
d'espèces Malagasy avancées par les chercheurs varient entre 8500
(White, 1983) et 14.000 (Goodman et Benstead, 2005, Phillipson et al.,
2006) groupées en 1730 genres, 243 familles et 64 Ordres actuellement
reconnus. A l'heure actuelle, des familles botaniques restent encore à
étudier et la famille des Acanthaceae en fait partie (Callmander et
al., 2014). Les premiers travaux décrivant les genres et
espèces Malagasy furent publiés depuis le début du
XIXe siècle. Ensuite, Grandidier et ses successeurs avaient
poursuivi les travaux. La flore de Madagascar n'a cessé d'intriguer les
naturalistes des siècles durant, et continue à susciter un
intérêt de plus en plus grand (Goodman, 2008).
La famille des Acanthaceae, objet de la présente
étude, est la 11e grande famille botanique mondiale en termes
d'espèces (Scotland et Vollesen, 2000). Ces plantes, parfois grimpantes
ou épiphytes, herbacées, arbustives ou rarement arbres sont
répandues dans toutes les régions du monde. On les rencontre
principalement en Inde, Malaisie, Afrique, Brésil et Amérique
centrale (
http://fr.wikipedia.org/wiki/Acanthaceae).
Cette famille présente environ 4000 espèces réparties sur
221 genres d'après Vollesen (1989) et sur 275 genres d'après
Daniel (1997). A Madagascar, la famille des Acanthaceae fait l'objet d'une
flore écrite par Benoist, sous la Direction de Humbert en 1967 dans la
série "Flore de Madagascar et des Comores" (Plantes vasculaires), la
182e famille. Cet auteur a prévu trois tomes pour cette
famille mais seul le tome I fut finalisé à cette époque.
Ce tome décrit les sous familles des Thunbergioideae, des Nelsonoideae
et des Acanthoideae, excepté la tribu des Justicieae de cette
dernière. Quelques espèces de cette tribu ont été
pourtant décrites (Justicia longispica, Hypoestes
capitata, Hypoestes decaryana, Dicliptera decaryi et
Justicia ankaratrensis (Benoist, 1920, 1929, 1947, 1949 et 1967).
Le Parc National d'Andohahela a été choisi pour
mener cette thèse car ces taxons s'y rencontrent dans des milieux
très diversifiés contrairement à la littérature.
Ainsi, le Parc National d'Andohahela et ses périphéries
constituées à l'Est par les vallées de Manampanihy et
d'Efaho et à l'Ouest par la vallée de la Mandrare sont alors la
zone d'étude considérée ici. Entre ces deux zones existe
une zone de transition pouvant correspondre au Domaine du Centre de Humbert
(Rajeriarison et Faramalala, 1999). Ce parc fait partie du réseau des
Parcs Nationaux de
2
Introduction
Madagascar géré par Madagascar National Parks.
C'est un massif montagneux à affleurements de roches granitiques
d'orientation Nord-Sud appelé globalement `Chaines Anosyennes' culminant
à plus de 1900m. Ce parc présente une richesse inestimable en
matière de diversité biologique dont fait partie les Acanthaceae
; et ces deux versants offrent des habitats propices à ces plantes.
L'objectif principal de la présente thèse est
alors d'apporter des éléments botaniques complémentaires
pour faire avancer les recherches sur les Acanthaceae et afin de pouvoir
publier plus tard les deux tomes restants de la flore. En effet, elle a pour
but de décrire les spécimens existants dans le Parc Botanique et
Zoologique de Tsimbazaza, dans le Centre California Academy of Sciences de San
Francisco et au Museum National d'Histoire Naturelle de Paris et ceux
collectées lors de cette recherche. Les travaux de laboratoire se sont
déroulés au Parc Botanique et Zoologique de Tsimbazaza (PBZT) et
au California Academy of Sciences (CAS). Les initiations en systématique
ont été faites au CAS et furent tout de suite appliquées
aux spécimens de PBZT.
Les objectifs spécifiques en sont :
- Mener l'inventaire des genres et espèces de la
famille des Acanthaceae dans le Parc National d'Andohahela et ses environs ;
- Faire un état de lieu de l'herbarium du Parc
Botanique et Zoologique de Tsimbazaza (TAN) en terme de représentants de
la famille des Acanthaceae et identifier quelques spécimens de cette
dernière;
- Proposer une clé partielle de détermination
par analyse morphologique des espèces etudées ;
- Elaborer une carte de distribution des espèces de la
zone étudiée ;
Les hypothèses de la présente thèse sont
formulées comme suit :
- H1 : Les différentes espèces de la famille des
Acanthaceae à Madagascar sont des bons indicateurs écologiques
;
- H2 : Les espèces de la famille des Acanthaceae
à Madagascar se développent en milieu
ouvert.
Le présent travail se divise en quatre parties :
- La première partie est consacrée au milieu
d'étude ;
- La deuxième porte sur les matériels et
méthodes ;
- La troisième présente les résultats et
leurs interprétations ;
- La quatrième et dernière partie traite les
discussions, les perspectives et les
recommandations ;
- Et enfin la conclusion.
CONTEXTE GENERAL ET MILIEU D'ETUDE
|
3
Contexte général et milieu
d'étude
I.1. ETAT DES CONNAISSANCES SUR LES ACANTHACEAE
La famille des Acanthaceae comporte dans le monde environ 4000
espèces réparties sur 221 genres d'après Vollesen (1989)
et 4300 espèces qui sont reparties dans 350 genres selon Koekemoer et
al. (2014). A Madagascar, cette famille est représentée
par 400 espèces reparties dans sept tribus et trente trois genres. Ces
valeurs ont été obtenues sur la base des collectes de
spécimen d'herbiers existants dans plusieurs pays aux herbaria de Kew,
de Missouri, de Paris, ... Parmi les travaux éffectués sur les
Acanthaceae, on peu citer : Bremekamp (1938, 1957) respectivement au Surinam et
en Nouvelle Guinée ; Leonard (1951) en Colombie ; Baden (1981) sur le
genre Anisotes en Afrique; Ezcurra, (1993) sur Ruellia en
Amérique du Sud ; Bennet et Scotland (2003) sur le genre
Strobilanthes à Java ; Wasshausen et Wood (2004) en Bolivie ;
Daniel et Figueiredo (2009) au Sao Tomé et Prýncipe.
Par ailleurs, des études sur les grains de pollens ont
été réalisées par Carine et Scotland (1998) sur le
genre Strobilanthes Blume dans le Sud de l'Inde et au Sri Lanka ;
Shendage et Yadav, (2009) sur Barleria L. de l'Inde ; Anisa et
al. (2015) sur Rhinacanthus Nees et Hypoestes Sol.
ex R. Br. de l'Yemen ; Nurul Aini et al. (2017) sur les Ruellioideae
dans le Péninsule de Malaisie. Depuis 1900, des botanistes ont
mené des recherches sur la famille des Acanthaceae un peu partout dans
le monde et cela a abouti à des bases pour la clé de
détermination. Cette famille a été étudiée
dans le monde entier, même au niveau des lieux reculés et
difficiles d'accès. Elle est caractérisée par des fleurs
zygomorphes et un calice à 4 à 5 lobes persistants, une corolle
gamopétale, un tube cylindrique ou ventriculé, lèvre
à 5 pièces inégales ; 4 ou 2 étamines fertiles avec
0,1,2 ou 3 staminodes ; ovaire supère, conique ou oblong-cylindrique,
bicarpellé, syncarpe, placentation axile ; fruit capsulaire bivalve et
2-loculaire, rarement drupacé (Perveen et Qaiser, 2010 ; Wood, 1997).
Clarke, depuis 1900, à Madagascar, a axé ses
recherches sur le genre Ecbolium, suivi de Benoist qui travaillait
depuis 1920 sur plusieurs genres et espèces de cette famille. Ce
dernier, en 1967, a fini la rédaction du tome I de la flore des
Acanthaceae. Cette flore présente trois sous familles (Thunbergioideae,
Nelsonioideae et Acanthoideae) avec 32 genres et 400 espèces qui se
rencontrent dans presque toutes les régions de Madagascar. D'une
manière générale, les étiquettes des herbiers
montrent que ces espèces sont surtout collectées au niveau des
aires protégées. C'est dans ce cadre que California Academy of
Sciences, à travers les programmes de recherche de Daniel (1999-2017), a
mis en place en 2005 un projet sur l'étude de la famille des Acanthaceae
Malagasy afin de contribuer à sa révision. Des résultats
préliminaires par genre peuvent déjà être
mentionnés :
4
Contexte général et milieu
d'étude
· Définition du nombre des chromosomes
d'Anisotes (Daniel, 2006) ;
· Détermination d'une nouvelle espèce, A.
divaricatus (Daniel et al., 2007) ;
· Délimitation des liens infragnériques entre
les Acanthaceae, les Acanthoideae et les Justicieae, Populina richardii
et Ecbolium (Daniel et al., 2008a) ;
· Détermination de la phylogénie
moléculaire des Nelsonioideae (Acanthaceae) et précision de la
phytogeographie des Elytraria (Wenk et Daniel, 2009) ;
· Détermination de nouvelles espèces
d'Anisotes de Madagascar (A. hygroscopicus, A.
perplexus, A. subcoriaceus, A. venosus. (Daniel et al.,
2013) ;
· Distinction de trois espèces de Mendoncia
à partir de Mendoncia cowanii (Moore) à savoir
Mendoncia kely, M. decaryi et M.
delphina selon une étude morphologique des spécimens de
l'herbarium de Tsimbazaza (Magnaghi et Daniel, 2017).
Parallèlement aux études de Daniel, T.F., des
collectes de spécimens d'espèces Malagasy ont été
faites par plusieurs auteurs. Ces collectes ont abouti aux résultats
suivants :
· Révisions de Megalochlamys et
d'Ecbolium en relation de travail avec Kew (Vollesen, 1989) et du
gerne Neuracanthus (Bidgood et Brummitt, 1998) ;
· Etude morphologique des pollens des Ruellia
d'Afrique et de Madagascar (Furness et Grant, 1996) ;
· Classification des Acanthaceae par Scotland et Vollesen,
2000 ;
· Inclusion du genre Avicennia L. (mangrove) dans
la famille des Acanthaceae (McDade et al., 2008) ;
· Etudes phylogénétiques des Acantheae
(McDade et al., 2005) et des Ruellieae (Tripp et al., 2013)
;
· Révision taxonomique de Eusiphon
longissimum Benoist qui est en fait une Ruellia L. et
précisée pour être Ruellia quatziticola Calmander
et Phillipson (Callmander et Phillipson, 2014).
I.2. MILIEU D'ETUDE
Les représentants de la famille des Acanthaceae
existent dans tout Madagascar (PBZT, 2017). La zone d'étude est choisie
car l'extrême Sud-Est de Madagascar a une particularité
bioclimatique intéressante pour déterminer le ou les facteurs
écologiques prépondérants pour leur répartition. Ce
site présente en effet trois types de bioclimats où la
végétation varie sur une petite distance et où plusieurs
auteurs (Eboroke, 1994, Koechlin et al., 1974, Rakotondrasoa,
2011,...) ont déjà mentionné l'existence d'une zone de
transition (formation à Dypsis decaryi) entre l'Est qui est
5
Contexte général et milieu
d'étude
humide et le Sud qui est sec. Phytogéographiquement, la
zone d'étude comporte de l'Est vers l'Ouest, une forêt dense
humide, la formation de transition à Dypsis decaryi, et enfin
à l'Ouest où se rencontrent un mélange de fourré
à Didieraceae et Euphorbia et de forêt sèche.
Le paragraphe suivant présente la localisation de la
zone d'étude, ses facteurs biotiques et abiotiques et enfin l'homme et
ses activités.
I.2.1. Localisation géographique de la zone
d'étude
La présente thèse a été
menée à l'extrême Sud-Est de Madagascar, dans le Parc
National d'Andohahela et ses environs immédiats. La carte 1 donne la
localisation de la zone d'étude sur fond de végétation
afin de situer tout de suite les différents habitats des Acanthaceae. Ce
parc se trouve à cheval entre les districts de Tolagnaro et celui
d'Amboasary-Atsimo, dans la région de l'Anosy, sous les
coordonnées géographiques comprises environ entre 120000m et
180000m Sud et entre 390000m et 450000m Est (Projection Laborde). Lors de son
classement en Réserve Naturelle Intégrale (RNIn°11) en 1939,
le Parc National d'Andohahela avait une superficie de 30 000ha ; puis en 1966,
après une rédelimitation, sa superficie est de 76.020ha
(Décret 66-242 du 1er juin 1966). Enfin, par le décret
97-1043 du 8 août 1999, son statut est changé en Parc National
sans modification de superficie (
https://www.parcs-madagascar.com/aire-protégée/parc-national-andohahela).
Il comporte toujours trois parcelles dont la première (Parcelle 1)
couvre 63.100ha, la deuxième (Parcelle 2), 12.420ha et la
dernière (Parcelle 3), 500ha (ANGAP, 2001).
La zone d'étude se divise en trois parties tenant
compte des trois parcelles du Parc National d'Andohahela qui se trouvent dans
des conditions bioclimatiques différentes, supposées influencer
la répartition des Acanthaceae ; ce sont la partie humide, la zone de
transition et la partie sèche. Dans la suite de la rédaction, ces
appellations seront conservées.
I.2.2. Facteurs abiotiques
I.2.2.1. Géologie, géomorphologie et
sol
La carte 2 présente un croquis de la géologie
simplifié de la zone d'étude. La partie humide de ce site
comprend la première parcelle du parc avec quatre sommets
dépassant 1900m d'altitude et son point culminant à 1972m
à Andohahela. Géologiquement, elle est formée par le
complexe de Fort-Dauphin qui est composé d'une couche granitique et de
la `charnockite' et par celui de Ranomafana à caractère
dominant de silico - alumineux et siliceux (Majmundar, 1961).
Contexte général et milieu
d'étude


7
Contexte général et milieu
d'étude

Carte 2 : Extrait du croquis géologique de la
région des Chaînes Anosyennes (De la Roche, 1958, modifié
par Mbola, 2017).
Le sol y est noir en général, argileux et peu
profond parsemé par des affleurements granitiques qui se rencontrent un
peu partout, même dans la forêt.
8
Contexte général et milieu
d'étude
Sa topographie est constituée par des escarpements
successifs limités par des falaises et des vallées
encaissées, en particulier la Manampanihy au Nord et
Andriambe-Efaho au Sud. La zone de trasition est formée d'un
plateau entrecoupé par une série de collines et de vallons dont
l'altitude varie de 120 à 1200m avec la parcelle 3. Elle montre des
affleurements de roches granitiques et des sols souvent érodés,
de lithosol sous le fourré à Didiereaceae au niveau de l'axe
Bevilany - Ambatoabo - Esomony en passant par Mahamavo.
Enfin, la partie sèche correspond réellement au
domaine du Sud de Humbert où se trouve la parcelle 2 et est
composée de deux blocs, le groupe de Tranomaro et celui du volcanisme de
l'Androy. Le premier est composé par les couches de Tsilamaha, d'Esira
et de Tranomaro comportant une série de collines et de vallons aux
altitudes variant de 120 à 1006m à Vohidagoro, son point
culminant (Petit, 1998). Ces différentes couches génèrent
un sol de types ferrugineux tropicaux, souvent érodé en sol
sableux limoneux. Le deuxième groupe, appelé aussi `massif
Androyen' est caractérisé par des carapaces de sols ferralitiques
de couleur roux, sur un soubassement cristallin (De la Roche, 1958).
I.2.2.2. Hydrographie
Une ligne de séparation des unités
hydrographiques (Carte 3) sépare la zone étudiée en deux
versants Est et Sud. Son versant Est correspond à la partie humide,
tandis que dans le Sud se trouve la zone de transition et la partie
sèche. L'Est est composé de deux sous-bassins versants, la
Manampanihy et l'Andriambe-Efaho. Le Sud comporte les bassins versants
d'Ambatoabo et de la Mandrare. Ce dernier est délimité par les
Chaînes Anosyennes (Beampingaratra) et le rebord Manambien de l'Ivakoany
(Chaperon et al., 1993).
Les principales rivières y sont la Manampanihy,
l'Andriambe-Efaho et la Mandrare, qui prennent leurs sources au niveau des
quatre bassins versants cités plus haut.
- La Manampanihy coule le long des chaînes
Anosyennes en direction Sud-Ouest / Nord-Est et se jette dans l'Océan
Indien au niveau de Manantenina. Son principal affluent est
Andohaela.
- L'Efaho coule du Nord au Sud entre les reliefs de
Tsitongambarika et Beampingaratra et rejoint l'Océan Indien au niveau
d'Ambinanibe.
- La Mandrare prend sa source dans le massif du
Beampingaratra, près du pic Trafonaomby, vers 1800 m d'altitude et se
dirige vers le Sud-Est et rejoint l'Océan Indien au sud d'Amboasary. Ses
principaux affluents sont Manambolo et Mananara.
- Les rivières Tsimelahy et Andeomara
prennent sources dans le sous bassin d'Ambatoabo. Ces deux cours d'eau se
jettent dans l'Océan Indien à Antsovelo/Amboasary-Atsimo.
9
Contexte général et milieu
d'étude

Carte 3 : Extrait de la carte des réseaux
hydrographiques et principaux bassins fluviaux de Madagascar (Chaperon et
al. 1993, modifié par Mbola, 2018)
I.2.2.3. Climats
La zone d'étude comporte des stations
météorologiques et des stations synoptiques
présentées en Tableau 1. Une station météorologique
donne beaucoup plus d'informations car elle présente un abri
météorologique avec plusieurs appareils pour la
température au sol, la température ambiante, la vitesse et
l'orientation du vent, la pluviométrie, l'insolation et les
rosées. Une station synoptique par contre ne mesure que les
températures et les précipitations.
L'étude du climat se résume ici à
l'étude de la température et des précipitations de la zone
d'étude pour pouvoir analyser les répartitions de la
végétation à travers les bioclimats. Ainsi, un diagramme
ombrothermique de Gaussen est utilisé pour représenter
graphiquement ces facteurs.
10
Contexte général et milieu
d'étude
Tableau 1 : Températures et précipitations de la
zone d'étude
|
Stations
météorologiques
|
Localisation
géographique
Système
Laborde (m)
|
Température
moyenne
annuelle
(°C)
|
Précipitation
moyenne
annuelle
(mm)
|
|
Ampasy Nahampoana
|
454.682 - 127.579
|
22°1
|
2190
|
|
Behara
|
394.373 - 129.904
|
24°5
|
520
|
|
Tolagnaro
|
455.727 - 120.900
|
23°0
|
2590
|
|
Ranomafana
|
452.839 - 172.451
|
22°0
|
1890
|
|
Stations synoptiques
|
|
|
|
|
Ampandrandava
|
314.839 - 230.085
|
24°0
|
920
|
|
Ambatoabo
|
422.134 - 124.980
|
24°2
|
250
|
|
Ankariera Bevilany
|
413.245 - 124.231
|
25°1
|
250
|
|
Maromby
|
413.108 - 196.231
|
15°8
|
1800
|
|
Lac Anony
|
404.963 - 109.959
|
24°5
|
230
|
(Source : MAEP, 2003, Paulian et al., 1973)
Les courbes des pluviosités moyennes mensuelles (P) et
les températures moyennes mensuelles (T) permettent de mettre en
évidence le nombre de mois secs de l'année d'une zone
donnée. Le principe en est basé sur la formule P = 2T où
les précipitations sont inférieures à deux fois la
température, définissant les zones à mois
écologiquement secs (Gaussen, 1955). Le tableau 2 et la figure 1
montrent le diagramme ombrothermique de Tolagnaro qui est
considérée ici comme la station type de la partie humide de la
zone d'étude, qui a un climat humide chaud (Humbert, 1955). La
température, atténuée par l'altitude de l'Anosy
continentale, y a une valeur moyenne de 23°C avec une amplitude thermique
journalière de 10°C. La précipitation annuelle
apportée par l'alizé est de 1500 à 2000mm et peut
atteindre 3000mm (Humbert, 1955 ; MAEP, 2003).
Les variations de la pluviosité et de la
température sur les versants occidentaux sont expliquées par
l'effet de Foehn engendré par la présence des reliefs. Les vents
venant de l'Est, en montant en altitude, se déchargent de la
quasi-totalité de leur humidité. Les effets de la
continentalité jouent également un rôle non
négligeable dans la répartition des régions thermiques
à travers la diminution de l'influence régulatrice et
modératrice des vents humides au fur et à mesure que l'on se
déplace vers l'Ouest.
Aucune station type météorologique ne se trouve
dans la zone de transition. Seule la station synoptique d'Ambatoabo
(thermomètre et éprouvette) y existe mais les archives de la
commune, responsable des relevés météorologiques y
relatifs ne permettent pas d'avoir des valeurs significatives pour dresser un
diagramme ombrothermique correspondant à cette zone. Pourtant, ces
données sont très importantes car une faille climatique existe
dans ce site particulier où deux saisons nettes sont mentionées,
la saison chaude, de novembre à avril et la fraîche, de mai
à octobre (Ratsivalaka-Randriamanga, 1987).
11
Contexte général et milieu
d'étude
Tableau 2 : Températures et précipitations de
Tolagnaro
|
Juil.
|
Aout
|
Sept.
|
Oct.
|
Nov.
|
Déc.
|
Janv.
|
Février
|
Mars
|
Avril
|
Mai
|
Juin
|
|
Températures (°C)
|
20°5
|
20°7
|
21°6
|
22°9
|
24°1
|
15°5
|
16°5
|
26°5
|
25°7
|
24°1
|
22°2
|
20°2
|
|
Précipitations (mm)
|
60,2
|
99,5
|
25
|
59,3
|
120,9
|
386,8
|
256,8
|
96,8
|
96,8
|
134,5
|
119,8
|
70,9
|
(Source : Humbert, 1955 ; MAEP, 2003)

Figure 1 : Diagramme ombrothermique de Tolagnaro, selon
Gaussen (Source : (Humbert, 1955 ; MAEP, 2003)
La partie sèche de la zone d'étude est soumise
à un climat chaud et sec, avec une température annuelle de
24°C (climat tropical sec). Le climat le plus sec y est observé. La
saison écologiquement sèche y dure plus de huit mois, d'Avril
à Novembre. Le tableau 3 présente les détails des moyennes
des témpératures et précipitations pour Behara entre 1981
et 2010.
Ceci est matérialisé par la
représentation du diagramme ombrothermique de la station type de Behara
(Figure 2). En résumé, les deux districts de la région
Anosy, Tolagnaro et Amboasary-Atsimo, montrent une diversité climatique
très accentuée. Les précipitations moyennes annuelles
12
Contexte général et milieu
d'étude
diminuent d'Est en Ouest et du Nord au Sud, occasionnant une
forte continentalité au Sud-Ouest de Madagascar.
Tableau 3 : Températures et précipitations de
Behara moyenne pour l'anné 1981-2010.
|
Juil.
|
Aout
|
Sept.
|
Oct.
|
Nov.
|
Déc.
|
Janv.
|
Fév.
|
Mars
|
Avril
|
Mai
|
Juin
|
|
Températures moyennes (°C)
|
21°
|
22°
|
24°6
|
25°9
|
29°9
|
30°2
|
31°2
|
31°
|
29°8
|
28°9
|
22°2
|
21°6
|
|
Précipitations moyennes (mm)
|
25,2
|
12,1
|
18,2
|
30,1
|
48,4
|
85,2
|
82,1
|
81,6
|
44,1
|
30,2
|
28,1
|
30,2
|
(Source : WWW.Maproom dgm, 2018)

Figure 2 : Diagramme ombrothermique de Behara, selon Gaussen
(Source : WWW.Maproom dgm, 2018)
I.2.3. Facteurs biotiques
Comme mentionné plus haut, la zone d'étude comporte
une forêt dense humide, une formation de transition à Dypsis
decaryi, un fourré à Didiereaceae et Euphorbia
mélangé avec de la forêt sèche. Ce paragraphe
va les décrire ainsi que la faune qui y habite et les activités
humaines.
13
Contexte général et milieu
d'étude
I.2.3.1. Végétation et flore
La zone d'étude présente plusieurs successions
de formations végétales. D'Est en Ouest, il est occupé par
la forêt dense humide (vallée de Manampanihy et celle d'Efaho), la
formation de transition de Ranopiso (caractérisée par la
présence du g. Dypsis decaryi) et le haut fourré
arboré à Didiereaceae et Euphorbia (à partir
d'Ankariera Bevilany en passant par Ambatoabo vers Behara) associé avec
de la forêt sèche (du côté de Tranomaro vers
Tsivory). La limite entre la forêt sèche et le fourré est
difficile à déterminer. Le passage en est très progressif
et la répartition de ces deux formations dépend pour une grande
partie des types de sol (Koechlin et al., 1974). L'extrême Sud
de Madagascar abrite un haut fourré arboré épineux,
xérophile, totalement atypique, avec les familles des Didiereaceae et
des Euphorbiaceae extrêmement variées et de nombreuses
espèces de plantes succulentes endémiques (Koechlin et
al., 1974). Les descriptions de la végétation
présentées dans ce travail de recherche sont basées sur
les travaux de Humbert et Cours-Darne (1965), de Paulian et al.
(1973), de Koechlin et al. (1974), de Goodman (2008), de Rajeriarison
et Faramalala (1999) et de Moat et Smith (2007).
? La partie humide de la zone d'étude est
composée par la forêt dense humide (Photo 1), pouvant atteindre
30m de hauteur et est constituée par plusieurs strates. C'est une
forêt hétérogène caractérisée par la
richesse des associations végétales dont la composition
floristique varie avec l'altitude. La végétation y est dense
jusqu'à 800m d'altitude. Au-delà, les arbres de grande taille
disparaissent pour laisser place à une brousse éricoïde
constituée par Erica sp., Dombeya seyrigii,
Psiadia dracaenifolia, Helichrysum fulvescens, Helichrysum
sp., Senecio vangaindrani, Kalanchoe
sp. et Xerophyta dasylirioides. Le
climat local de cette forêt est caractérisé par une
atmosphère très humide et fraîche et est favorable aux
épiphytes, aux nombreux parasites, aux saprophytes et aux lianes qui
prolifèrent au niveau de la strate supérieure (ANGAP, 2004).
? La strate supérieure est formée par des arbres
de hauteurs variables comme Canarium madagascariense, Albizzia gummifera,
Eugenia sp., Dilobeia thouarsii, Vepris macrophilla, Salvadoropsis
arenicola, Ocotea sp., Tambourissa religiosa, Pandanus sparganioides,
Weinmannia rutenbergii, Weinmannia sp., Oncostemon sp.,
Ocotea sp., Schefflera monophylla et Cuphocarpus
sp. et des lianes telles que Mendoncia
cowanii et Hagellaria indica.
14
Contexte général et milieu
d'étude

Photo 1 : Forêt dense humide, au col de Manangotry, vue
d'Androangaloaka (C.R. Isaka Ivondro) La strate moyenne (4-8m) montre des
représentants de la famille des Humbertiaceae, des Myristicaceae, des
Apocynaceae, des Tiliaceae, des Erythroxylaceae et des Ochnaceae et aussi par
les genres Gaertnera sp., Gymnosphaera sp., et par
les espèces Filicium longifolium, Plagioserphus stelechanthus,
Antidesma petiolare, Aphloia theiformis, Cyathea dregei, Gymnosphaera
articola et Arundinaria sp. La strate inférieure (0-4m),
lâche et discontinue, ne devient dense qu'en altitude, avec d'abondants
jeunes plants de Craspidospermum verticillatum, Dracaena bakeri,
Viguieranthus denginervus, Streptocarpus suffriticocus, Croton
sp. et diverses espèces
herbacées (Acanthaceae, Urticaceae, Fougères et Impatiens
sp., Vaccinium secundiflorum, Tachiadenus longifolius et
Cyperus tsaratananensis). A cause du climat, c'est l'habitat
préféré de beaucoup d'épiphytes (Rhipsalis
baccifera, Hymenophyllum sibthorpioides, Hymenophyllum
sp., Peperomia sp., Medinilla uncidens, Asplenium
friesiorum var. nerophilum, Asplenium
sp. et Polystichum coursii). Par
contre, on y trouve peu de lianes mais pas de mousses (Andriarimalala, 2014 ;
Paulian et al., 1973). C'est surtout cette strate inférieure
qui est l'habitat des Acanthaceae, objet de la présente étude.
? Dans la zone de transition de la zone d'étude se
trouve la formation à Dypsis decaryi (Photo 2), qui se localise
entre les isohyètes 800 et 1200mm (Ratsivalaka-Randriamanga, 1987).
15
Contexte général et milieu
d'étude

Photo 2 : Fourré à Dypsis decaryi (C.R.
Ranopiso)
Hautes de 8 à 12m, peu pénétrables, sans
strates et présentent de nombreuses formes d'adaptation à la
sécheresse comme la spinescence, même au niveau des fruits, la
réduction de la surface foliaire, le renflement de la tige, la
lactescence, la ramification et le port en zigzag, le développement de
feuilles charnues et grasses. Partout, la végétation comporte
deux strates car la strate inférieure n'existe pas ; le sol
apparaît souvent. Outre Dypsis decaryi, la strate
supérieure est constituée par Dalbergia greveana,
Commiphora marchandii, C. grandifolia, Euphorbia plagiantha, E.
enterophora, Croton mongue, Neobegeua mahafaliensis, Pachypodium rosulatum,
Albizzia fatipatiky, Terminalia seyrigii, Alluaudia
ascendens, A. dumosa, Tetrapterocarpon geayi, Euphorbia plagiantha et
E. enterophora. La strate moyenne, discontinue est composée par
beaucoup d'arbustes qui sont Barleria alluaudii, Cedrelopsis
grevei, Enterophora sp., Uncarina leandri, Megistostegium perrieri,
Pachypodium runtenbergianum, Croton sp., Physena
seseliflora, Aloe vahombe, A. divaricata, Rhigosum
madagasacariensis, Canthium sp., Chadsia grevei, Euphorbia
leucodendron, Commiphora marchandii, C. stellulata, Uncarina grandidieri et
Commiphora brevicalyx (Eboroke, 1994 ; Andriarimalala, 2014).
Des espaces couverts par des savanes arborées
(Tamarindus indica, Gymnosporia linearis, Poupartia
caffra) souvent d'origine anthropique se rencontrent dans cette zone.
? La zone sèche de la zone d'étude est couverte
par une forêt sèche associée à un haut fourré
arboré (Photo 3), par endroit, peu pénétrable,
composé d'arbres épineux ou d'arbustes ramifiés,
buissonnants, sans tapis herbacé (Koechlin et al., 1974).
La strate supérieure, avec une hauteur variable, présente
Adansonia za, Commiphora aprevalii, C. lamii, Alluaudia
16
Contexte général et milieu
d'étude
procera, A. acendens et Gyrocarpus americanus.
La strate moyenne y est caractérisée par la présence
des jeunes plants de la strate supérieure et des Crassulaceae, des
Euphorbiaceae, des Pachypodium rosulatum, Adenia olabeensis, Cyphostema
elephantropus, Diospyros aculeata, Evonymopsis longipes, Polycardia aquifolium,
P. lateralis, Rinorea spinosa, Ruellia sp., Pachypodium sp. Cette
strate est l'habitat privilégié des Acanthaceae. L'ensemble est
doté d'une couleur grisâtre dominante. Le fourré y est
entrecoupé par des savanes à Heteropogon contortus, Panicum
maximum, Eragrostis cilianensis ... (Bosser, 1969). Le long des cours
d'eaux apparaît une forêt galerie à Tamarindus indica
ou à Breonadia salicina de superficie très
réduite. Des clairières existent et sont composées par
Heteropogon contortus, Hyparrhenia rufa et Imperata cylindrica.
Elles sont parsemées par de rares Mascarenhasia
madagascariensis et Aphloia theaeiformis et quelques Acanthaceae
telles que Barleria
sp. et Ecbolium sp.

Photo 3 : Haut fourré à Alluaudia ascendens
(C.R. Ankariera Bevilany).
I.2.3.2. Faune
La forêt humide est l'habitat de faune endémique
telle que Newtonia fanovanae (Oiseau), Phelsuma antanosy,
Paragehyra gabriellae, Mantella haraldmeieri et Furcifer balteatus
(Reptiles) et quelques lémuriens, Eulemur colaris et
Daubentonia madagascariensis ou Aye aye (ANGAP, 2004).
Dans la formation à Dypsis decaryi et la
formation sèche se trouvent des lémuriens (Lemur catta,
Propithecus verreauxi, Microcebus griseorifus), différentes
espèces de reptiles telles que Astrochelys radiata, Oplurus
quadrimaculatus, Chalarodon madagascariensis ... et quelques
17
Contexte général et milieu
d'étude
oiseaux dont Terpsiphone mutata, Upupa epops, Drongo,
Monticola sp., Coua ruficeps, C. gigas, ...
(Rakotondrasoa, 2011).
I.2.3.3. L'homme et ses activités
La zone d'étude est habitée par les Antanosy,
les Androy et les Bara. Les Antanosy, en général sont
constitués de trois (3) clans, le clan Tambolo dans la vallée de
la Manampanihy, les Tavaratra dans la vallée d'Andriambe-Efaho et les
Tatsimo se trouve à l'extrême Sud. Ils sont tous à la fois
agro-éleveurs et exploitants forestiers. Les deux Communes rurales de
Maromby et d'Ambatoabo sont peuplées à la fois par les Antandroy,
les Antanosy et les Bara provenant de Betroka. Ces derniers ne sont pas
nombreux mais sont pourtant la cause du climat d'insécurité de
cette zone, surtout à propos de l'élevage de zébus.
a. Agriculture
La population pratique la culture du riz, du manioc, de patate
douce, de maïs, des arbres fruitiers, de caféiers et tout
récemment de la vanille. L'agriculture se développe dans deux
endroits différents, hors et en forêt. Les couvertures
forestières, les fourrés et les savanes sont
défrichés pour les transformer en champs de culture (Photos 4, 5
et 6). Le riz est cultivé sur les bas-fonds et sur les flancs de
collines profitant de l'abondance des eaux de ruissèlement mais
où existent aussi des arbres fruitiers et des cultures sèches.

Photo 4 : Défrichement suivi de mise à feu dans la
vallée de Fanota, C.R. Isaka Ivondro (fôret humide)
18
Contexte général et milieu
d'étude

Photo 5 : Terrains de cultures sur les flancs des collines
(Village de Mahatsinjo, C.R. Ranopiso)

Photo 6 : Défrichement dans la localité de Bezaha,
C.R. Ambatoabo (fourré)
La pratique de mise à feu fait partie de l'habitude des
populations partout dans la zone d'étude. Mais comme la gestion des feux
n'est pas encore une pratique prioritaire pour eux, ils se propagent et peuvent
provoquer des feux de forêt.
b. Elevage
L'élevage de zébu, de chèvre et de mouton
est pratiqué dans la zone d'étude. L'élevage extensif est
le mode le plus courant pour toutes les communautés. La pratique de feu
de renouvellement de pâturage (Photo 7) peut souvent provoquer des feux
de brousse surtout quand ceci n'est pas maîtrisé.
Contexte général et milieu
d'étude

feu
19
Photo 7 : Feux de renouvellement de pâturage au niveau
des savanes en moyenne altitude, zone de Malio, C.R. Ranopiso
Les Antanosy, pour qui la viande et l'élevage de
chèvre sont tabous, pratiquent l'élevage bovin et ils pratiquent
le pâturage forêstier. Par contre, les Bara et les Antandroy
élèvent en même temps les chèvres, les moutons et
les zébus en forêt sèche, en fourré mais aussi en
savane (Kulus, 2001).
c. Exploitations des ressources
naturelles
Les ressources forestières sont utilisées
à des fins individuelles surtout au quotidien. Toutes les formations
végétales sont pillées de leurs produits par le
prélèvement de bois d'oeuvre ou d'énergie, des plantes
médicinales et médico-magiques et de leurs animaux sauvages et
cela surtout à des fins commerciales ou à usage personnel. Des
collectes de tubercules, de fruits et de feuilles sont pratiquées par la
population en période de pénurie.
Deux types d'exploitations de ressources naturelles existent
dans la zone d'étude, à savoir les exploitations licites ou
règlementaires et les illicites. Légalement, la forêt dense
humide constitue surtout la source de matières pour la fabrication de
longrines, de planches, de bois carré, de goélettes, de charbon
de bois et est aussi le foyer de collecte illicite de bois de rose. Les graines
de Dilobeia thouarsii sont collectées par la population de la
partie humide pour en fabriquer de l'huile à usage domestique ou
à but commercial. Ces produits sont destinés aux centres urbains.
Les forêts sèches et les fourrés sont les lieux de collecte
illicite de tortues radiées qui sont vendues dans les villages et
même dans les centres urbains les plus proches. La population pratique
aussi le braconnage de sanglier (Photo 8), de lémuriens (Lemur catta
et Propithecus verreauxi verreauxi) et d'oiseaux (Coua
ruficeps et C. gigas....).
20
Contexte général et milieu
d'étude

Photo 8 : Vente de viande de sanglier à Analamantsaky
(C.R. Ankariera Bevilany)
Plusieurs ressources naturelles sont exploitées
illicitement par la population locale dans la zone d'étude.
Alluaudia procera est coupée pour la fabrication des planches
et Cedrelopsis grevei pour le charbon de bois. Les
tradipraticiens utilisent comme plantes médicinales, Cedrelopsis
grevei, Vanilla humblotii, V. madagascariensis, Neobeguea
mahafaliensis, Croton sp., Aloe divaricata,
Commiphora monstruosa, Fernandoa madagascariensis, ..., et
comme plantes à vertu médico-magique Cynodon dactylon,
Leptadenia madagascariensis, Marsdenia cordifolia, Usnea sp., Aloe
divaricata, A. vahombe, Adenia olaboensis, Cyphostema laza. Crossandra
longipes, Uncarina leandrii, Ravenala madagascariensis
sont prélevées et vendues toujours illicitement comme
plantes ornementales à partir des différents types de formations
végétales. Enfin, Cedrelopsis grevei,
Tetrapterocarpon geayi, Gyrocarpus americanus, Alluaudia
procera, Aloe suzanae et Dypsis decaryi sont
prélevées pour la construction de cases (Photo 9). Cette
dernière est très prisée par la population car elle est
à usage multiple et ses graines servent surtout d'aliment de subsistance
en période de disette.
21
Contexte général et milieu
d'étude

Photo 9 : Toiture de case faite en feuilles de Dypsis
decaryi et murs en planches d'Alluaudia procera (Village
d'Analamantsaky, C.R. Ankariera Bevilany)
d. Exploitations minières
La zone humide comporte une exploitation d'ilménite par
le Qit Madagascar Minerals (QMM) depuis plus de vingt ans, au niveau de la
Commune Rurale d'Ampasy Nahampoana. La zone sèche est à
potentialité importante en ressources minières dans les Communes
Rurales de Tranomaro et de Maromby. La première est un site
d'exploitation de l'uranothorianite et de micaschiste depuis plus d'une
cinquantaine d'années (Bazot, 1976). Les gisements de mica phlogopite,
situés dans les pyroxénites du sud de Madagascar, furent
découverts par Rossi en 1908 (Guillani et al., 2007). La
seconde, où se trouve Andranondambo est le terrain d'exploitation de
saphir depuis plus de trente ans.
22
Matériels et méthodes
II.1. FAMILLE DES ACANTHACEAE
La famille des Acanthaceae, objet de la présente
étude, est répandue dans les régions tropicales. Elle est
représentée par des espèces d'arbres, arbustives,
herbacées et lianescentes. Ces plantes ont en général de
belles petites fleurs zygomorphes, vivement colorées.
II.1.1. Classification
Les trois différentes classifications de la famille des
Acanthaceae qui existent dans la littérature sont données
ci-dessous (Tableau 4).
Tableau 4 : Les classifications de la famille des Acanthaceae
|
Selon de Candolle (1819)
|
Selon Hennig (1950)
|
Selon `Angiosperm Phylogeny Group' (APG III, 2009)
|
|
Classis prima :
|
Clade : Angiospermes
|
Règne : Plantae, Végétal
|
|
Dicotyledoneæ
|
Clade : Dicotylédones vrais
|
Sous Règne : Tracheobionta,
|
|
Subclassis III :
|
Clade : Astéridées
|
Plantes vasculaires
|
|
Corollifloræ
|
|
|
|
Ordo CXLV : Acanthaceæ.
|
Clade : Lamiidées
Ordre : Acanthales Lindl.
|
Division : Magnoliophyta, Angiosperme, Plantes à fleurs
|
|
(1833)
Famille : Acanthaceae Juss.
|
Classe : Magnoliopsida, Dicotylédones
|
|
(1789)
|
Sous Classe : Asteridae
|
|
|
Ordre : Lamiales
|
|
|
Famille : Acanthaceae.
|
La famille des Acanthaceae est composée de trois (3) trois
familles, Sept (7) tribus, 33 genres et 400espèces. En 1967, Benoist
avait proposé la classification suivante pour les Acanthaceae de
Madagascar :
1 - La sous famille des Thunbergioidae, à
tribu unique, celle des Thunbergieae avec trois genres
Mendoncia (3 espèces), Thunbergia (8 espèces)
et Pseudocalyx, monospécifique qui est Pseudocalyx
saccatus.
2 - La sous famille des Nelsonioideae est
formée par la seule tribu des Nelsonieae avec deux genres, Nelsonia
(N. campestris) et Tubiflora (T. acaulis
et T. madagascariensis).
3 - Enfin, la sous famille des Acanthoideae est
composée de cinq (5) tribus, à savoir les Ruellieae, les
Crabeeae, les Barlerieae, les Acantheae et les Justicieae.
23
Matériels et méthodes
? La tribu des Ruellieae est représentée par 14
genres :
V' Ruellia est le plus riche avec trente et une
espèces ;
V' Dyschoriste et Strobilanthes suivent avec
huit espèces ;
V' Mimulopsis et Hygrophila ont chacun 7
espèces ;
V' Eusiphon, Forsythiopsis et Micranthus
possèdent chacun deux espèces ; et enfin
V' Les restes sont monospécifiques, à
savoir Brillantaisia madagascariensis,
Ionacanthus calcaratus, Lankesteria granduosa,
Paulowilhelmia decayi,
Pseudoruellia perrieri et Zygoruellia richardii.
? La tribu des Crabeeae possède cinq
genres :
V' Podoriungia est le plus riche avec cinq
espèces ;
V' Suivi par Lasiocladus avec quatre
espèces ;
V' Pseudodicliptera en a trois; et enfin
V' Deux genres sont monospécifiques, à
savoir Periblema cuspidatum et Pericalypta biflora.
? La tribu des Barlerieae est composée
de trois genres : V' Barleria est le plus riche avec 27 espèces
;
V' Lepidagathis est représenté par six
espèces ; et enfin V' Neuracanthus possède quatre
espèces.
? La tribu des Acantheae est
représentée par quatre genres :
V' Crossandra est le plus riche avec seize
espèces ;
V' Blepharis et Stenandriopsis ont chacun sept
espèces ; et enfin V' Achyrocalyx a trois espèces.
II.1.2. Morphologie des Acanthaceae
La morphologie est l'étude de la forme de la plante.
Les caractères des feuilles, des fleurs et des fruits sont
relevés pour cette étude. Les Acanthaceae sont des plantes
herbacées, arbustives et rarement à port lianescent ou des
arbrés (Schatz, 2001) et caractérisées par la
présence des cytolithes (Patil et Patil, 2011). Les espèces
herbacées se développent en formant un tapis sur le sol et des
fois des touffes. D'une manière générale, les Acanthaceae
sont des plantes à fleurs bisexuelles et zygomorphes, inermes ou
épineuses. La racine de ces espèces est superficielle et souvent
en fascicule. Mendoncia delphina et Barleria kitchingii,
espèces lianescentes, développent des racines au niveau des
noeuds quand ces derniers touchent le sol.
24
Matériels et méthodes
Les tiges des représentants de la famille des
Acanthaceae présentent parfois des épines. Elles sont souvent
pubescentes au stade jeune et lisses au stade adulte. La section des tiges est
arrondie, hexagonale ou sub-arrondie. Des fois, quelques espèces ont une
tige présentant des fentes ou des rainures longitudinales.
Ce sont des plantes à feuilles simples, opposées
et décussées, sans stipule, entières ou
plurilobées. Quelques espèces présentent des feuilles
terminées par une pointe, d'autres ont des feuilles à
épines sur les marges. Les feuilles sont lisses et des fois couvertes de
poils fins sur la face inférieure. Des espèces accusent la
présence d'écailles à la base des pétales et
d'autres non. Les feuilles ont une dimension très variable, petites,
allant de 11mm à 12mm de longueur chez Thunbergia et de 2mm
à 6mm de largeur chez Lasiocladus et grandes, allant de 4cm
à 15cm de longueur chez Brillantaisia et de 3cm à 8cm de
largeur chez Mendoncia.
Le calice peut être présent ou absent. S'il est
présent, il est composé de quatre à cinq sépales
lisses ou couverts de poils fins ou même pluridentés pour
certaines espèces. Les pièces peuvent être libres ou
soudées, formant un tube à préfloraison valvaire. Pour les
espèces épineuses, le calice et la corolle sont présents
à la fois sinon, l'une de ces pièces manque.
Les Acanthaceae ont des fleurs solitaires ou diversement
groupées en inflorescences axillaires ou terminales, hermaphrodites et
zygomorphes. La fleur est souvent petite, sauf chez Ruellia geayi et
Brillantaisia madagascariensis. La corolle est gamopétale,
pentamère et zygomorphe, à préfloraison soit tordue, soit
quinconciale, soit diversement imbriquée. Quand les pétales sont
soudés, ils forment un tube terminé par quatre à cinq
lobes. Les pétales sont parfois vivement colorés.
L'androcée est composé de deux ou de quatre étamines
fertiles et parfois avec zéro à trois staminodes,
insérés sur le tube de la corolle. Les filets peuvent être
libres ou soudés. Les anthères sont introrses avec un ou deux
loges s'ouvrant par des fentes longitudinales ou rarement par des pores
apicaux. Le gynécée est pourvu d'un ovaire supère. Ovaire
biloculaire, rarement uniloculaire par suite de l'avortement d'une des loges
avec deux à dix ovules axiles par loge. Style allongé et souvent
filiforme; stigmate bifide ou simple, parfois infundibuliforme ou
bilabié. Le fruit est ordinairement une capsule à 2 loges,
à déhiscence loculicide, contenant dans chaque loge un à
deux ou un plus grand nombre de graines. Généralement, chaque
graine est supportée par une expansion lignifiée du funicule
appelée retinacle et seule Mendoncia possède des baies.
Graine ordinairement sans albumen, discoïde, globuleuse à
tégument lisse, granuleux, poilu ou écailleux. Les Acanthaceae
présentent deux types de fruits, un fruit sec déhiscent ou une
baie. Les graines sont arrondies ou ailées, albuminées ou non
(Clarke 1900 ; Benoist, 1967; Daniel, 1997 ; Scotland et Vollesen 2000).
25
Matériels et méthodes
II.2. MÉTHODES D'ETUDE
Pour atteindre les objectifs, les méthodes
adoptées au cours de ce travail de recherche se divisent en deux, sur le
terrain et en laboratoire. Le travail sur site a consisté à
étudier les différents types d'habitats de la zone d`étude
suivi de la collecte les spécimens et enfin à enquêter la
population locale sur l'utilisation des plantes. Le travail en laboratoire est
axé sur l'étude morphologique, suivant la méthode de
Capuron (1957), des échantillons collectés qui sont
comparés ensuite avec les données des herbaria détenteurs
de spécimens d'Acanthaceae malagasy qui sont les herbaria de Tsimbazaza
(TAN) et de California Academy of Sciences (CAS) de San Francisco. Les
spécimens absents au CAS ont été empruntés au
Muséum National d'Histoire Naturelle de Paris.
II.2.1. Travaux sur le terrain
Les travaux de terrain se divisent en deux étapes
à savoir, les enquêtes et l'étude des habitats et les
collectes de spécimens d'Acanthaceae rencontrés. Toutes ces
investigations ont été faites dans les zones de collectes
mentionnées sur la carte 4 ci-dessous où sont aussi situés
les sites de relevé lors des inventaires.
a. Enquêtes ethnobotaniques
Des enquêtes ont été faites auprès
de la population riveraine de la zone d'étude pour avoir les
informations sur les plantes, en particulier par rapport à leurs
utilisations. Deux types d'enquêtes ont été menés,
l'entrevue individuelle et l'entretien par groupe avec, pour les deux, une
fiche à questionnaire ouvert élaborée au préalable
(Annexe 1). La première, individuelle, est une enquête-discussion
où aucune prise de notes n'est faite car la transcription des
données obtenues est réalisée plus tard en aparté.
La deuxième méthode est menée au niveau de certains
villages situés sur les axes d'inventaire auprès de groupe de
personnes qui ont l'habitude d'aller dans la forêt et qui connaissent
bien les plantes, en particulier les Acanthaceae. Après le regroupement
par groupe, certaines personnes clés sont approchées
séparément par `visite à domicile'.
Au cours de l'étude, au moins une personne, par village
a été interviewée. En tout, 30 personnes ont
été enquêtées. Elles se répartissent en 10
personnes au niveau de la partie humide, 10 dans la zone de transition et 10
dans la partie sèche.
Matériels et méthodes

27
Matériels et méthodes
b. Prospection écologique des habitats de
la zone d'étude
A chaque site de relevé, une observation minutieuse de
l'habitat a été faite pour avoir les informations sur la
préférence écologique des espèces. Les
caractéristiques écologiques, à savoir la physionomie et
les espèces dominantes, de tous les écosystèmes de la zone
d'étude, de la forêt jusqu'à la savane ont
été relevées. Quant ce fut possible, des groupements
végétaux ont été identifiés.
L'étude de l'habitat des espèces d'Acanthaceae a
concerné les différents facteurs écologiques suivants,
à savoir la subdivision climatique, l'altitude, l'ouverture du milieu,
ainsi que le type de sol où se développe la plante.
Au cours de la présente étude, la
délimitation des régions climatiques de Cornet (1974)
était prise en considération. La zone d'étude se divise en
trois parties qui sont les parties humides à l'Est, de transition au
centre et sèche à l'Ouest. Le choix même de cette zone
d'étude a été conditionné par trois conditions
climatiques différentes mais dans un espace restreint. En effet,
l'existence de la faille pluviothérmique mentionnée par cet
auteur offre bien des habitats différents où pourrait être
vérifiée la problématique mentionnée en
introduction.
L'analyse des altitudes des sites de relevé est ensuite
entreprise par la prise des coordonnées
géoréférencées des endroits de collecte à
l'aide d'un GPS. Trois altitudes, couramment considérées comme
limites de végétation en botanique, ont été
choisies ici, à savoir, zéro, 400m et 800m. Ainsi, l'analyse de
la repartition des espèces d'Acanthaceae a tenu compte de trois gammes,
basse altitude de 0 à 400m, moyenne de 400 à 800 et haute pour
une altitude au-delà (ANGAP, 2001).
L'étude du facteur `ouverture du milieu' a
concerné l'évaluation de la proportion de la surface du sol qui
serait recouvert si on y projetait verticalement les parties aériennes
des plantes. Ce facteur est obtenu à l'aide de la «Charte pour
l'estimation visuelle des rapports de surface» d'Emberger et al.,
(1983) qui fût adaptée aux différents types de milieu car
les Acanthaceae se trouvent en majorité en strate basse :
- Recouvrement > 75% : milieu fermé ;
- 50% < Recouvrement < 75% : milieu semi ouvert ;
- Recouvrement <50% : milieu ouvert.
L'étude du sol était faite à travers sa
couleur et sa texture pour avoir une idée sur le type du milieu
où se trouve chaque plante. Sont choisis, au cours de cette
étude, les trois types de sol les plus fréquents dans la zone qui
sont les sols noirs de forêt (sol ferralitique peu evolué et
humifère), sols alluvionnaires (sableux de couleur roux) et un sol
calcaire (sol ferrugineux sur calcaire) (CIRAD et al., 2002, De la
Roche 1958, Majmundar, 1961).
28
Matériels et méthodes
c. Inventaire biologique
Des missions de collecte ont été
organisées dans la zone d'étude. Suivant les informations
figurées au niveau des étiquètes d'herbier, telle que la
localisation de la collecte, on sait que les Acanthaceae se trouvent dans des
milieux ouverts, hypothèse à vérifier au cours de la
présente étude. Un itinéraire, au niveau des voies
d'accès, est alors défini suivant ces indications pour faire les
reconnaissances et les collectes.
La collecte a été faite en suivant les
réglementations systématiques en vigueur pour pouvoir
entreprendre une étude morphologique de chaque plante afin de les
identifier et pour pouvoir proposer la clé de détermination
partielle des Acanthaceae de la zone d'étude, objectif de cette
étude. Les parties de la plante collectées concernent, selon le
cas, des fleurs, des fruits, des feuilles, des tiges et de la racine. Pour les
plantes de petite taille, l'individu entier est collecté. Chaque fois
que c'est possible pour une plante observée, la collecte est faite au
minimum en quatre exemplaires conformément aux règlements en
vigueur à Madagascar. Une part est déposée au TAN et les
restes sont envoyés aux herbaria partenaires.
Chaque échantillon a fait l'objet d'une première
description, sur place avec une loupe portative avant de le mettre sous presse
herbier. Au cours de cette observation, les caractères qui ne sont plus
visibles sur les herbiers sont munitieusement notés ; ce sont le port,
les aspects de l'appareil végétatif et la couleur.
La méthode de confection des herbiers suit celle de
Bridson et Forman (1992). Les spécimens ont été
arrimés dans des papiers journaux puis pressés et
séchés directement sur terrain ou imbibés d'alcool avant
un séchage ultérieur. Par la suite, les spécimens sont
montés pour être déposés à l'herbarium de
Tsimbazaza. Des échantillons de fleurs, de feuilles et de fruits ont
été mis dans une boîte contenant du gel de Silice quand les
spécimens présents dans la nature le permettaient pour conserver
le matériel génétique pour une étude
ultérieure d'ADN. Des collections photographiques de toutes les
espèces recensées et de leurs habitats respectifs ont
été faites.
II.2.2. Travaux en laboratoire
Les travaux en laboratoire ont été surtout faits
au niveau des hebaria de référence, où des loupes
binoculaires beaucoup plus performantes ont été utilisées.
Des spécimens du genre Dicliptera ont été
empruntés au Muséum National d'Histoire Naturelle (MNHN) pour
compléter les échantillons. Ces travaux ont concerné
surtout l'analyse morphologique des plantes collectées et des
spécimens d'herbiers.
29
Matériels et méthodes
L'élaboration de la clé de détermination
des espèces de la zone d'étude et l'étude statistique des
données relatives aux préférences écologiques de
chaque plante ont été ensuite faites. Cette dernière
concerne surtout la similarité entre les sites de relevé,
habitats des Acanthaceae.
Des études à part ont été faites
sur les échantillons indéterminés de TAN contenant des
spécimens d'Acanthaceae ou d'autres familles, même si ces derniers
étaient localisés hors de la zone d'étude. Cette approche
a été entreprise pour assurer une étude complète
des echantillons disponibles à Madagascar.
Les travaux effectués concernent :
a- La manipulation et la description des spécimens
d'herbier existants et l'identification des espèces rencontrées
au cours des missions d'inventaire en comparaison avec les isotypes,
les syntypes et les lecotypes déposés au TAN.
Les types biologiques proposés par Raunkiaer (1904) selon le
positionnement des bourgeons en période défavorable ont
été utilisés. Les caractères biologiques
étudiés concernent les feuilles, les tiges, les fruits et les
graines et leurs éléments connexes (poils, épines, ...)
;
b- La révision de la nomenclature et des
classifications des espèces collectées confirmées par
`The International Plant Names Index'. A cet effet, quelques flores
des Acanthaceae ont été utilisées comme celle de la Chine
et celle de la Guyane Française ;
c- Le dessin des planches des spécimens pour les
illustrations ;
d- Cartographie la localisation des spécimens
collectés avec le logiciel Arc View ;
e- Les analyses statistiques des données relatives aux
paramètres écologiques du milieu sont faites avec les logiciels
STATISTICA 6 et XLSTAT sous windows ;
f- Enfin, les résultats de toutes les descriptions des
échantillons ont été traités avec le logiciel
Botanical Research and Herbarium Management System (BRAHMS) pour
pouvoir proposer une clé partielle de détermination des
plantes.
II.2.3. Traitement des données
Les données collectées relatives aux plantes
étudiées ont été analysées pour
déterminer leurs préférences écologiques en
fonction des variations des paramètres écologiques
considérés qui sont les subdivisions climatiques, l'altitude,
l'ouverture du milieu et les types de sol. Ces variations seront
présentées en resultats sous forme de tableaux pour analyser
cette préférence écologique de chaque espèce.
Une étude de similarité entre les sites de
relevé est ensuite faite selon la méthode de Jaccard (1901) avec
le logiciel XLSTAT 6, pour connaître le nombre des espèces
communes entre deux
30
Matériels et méthodes
sites et confirmer si les Acanthaceae sont réellement des
indicateurs écologiques. L'indice de similitude de Jaccard est
donné par la formule :
SJac=c/(a+ b+c)
où
- a et b résentent le
nombre d'espèces inventoriées respectivement dans les
relevés A et B ;
- c représente le nombre des
espèces communes entre les deux sites de relevés.
Si l'indice de similitude de Jaccard (SJac) augmente, un
nombre important d'espèces communes sont rencontrées pour les
deux sites comparés.
Une analyse factorielle des correspondances (AFC) est enfin
entreprise avec le logiciel STATISTICA pour voir la relation entre les
espèces et leur site de collecte et plus précisément, pour
définir les groupes de plantes qui se forment en fonction des
paramètres écologiques étudiés.
RÉSULTATS ET INTERPRETATIONS
|
31
Résultats et interprétations
Les résultats de cette étude ont concerné
les habitats de la zone d'étude, qui seraient présentés en
premier lieu, suivis de l'analyse morphologique des spécimens
collectés et de la clé de détermination proprement dites.
Ces informations ont ensuite permis de décrire les Acanthaceae du site
de relevé et d'analyser leurs préférences
écologiques.
III.1. RESULTATS DES ENQUETES ETHNOBOTANIQUES
Conformément à la méthodologie
d'enquête appliquée relative aux utilisations des Acanthaceae, les
résultats (Tableau 5) montrent que la plupart de ces espèces ne
sont pas connues. Seule Crossandra longipes a été
utilisée par la population en ornementation. En plus de cette
dernière, Barleria humbertii, Ecbolium syringifolium et
Lasiocladus anthospermifolius ont chacune un nom vernaculaire.
Tableau 5 : Les utilisations des plantes par la population
|
Noms scientifiques
|
Nom
vernaculaire
|
Famille
|
Parties
utilisées
|
Utilisations
|
|
Abrahamia grandidieri
|
Sohihy
|
Anacardiaceae
|
Bois
|
Construction, arôme de rhum local
|
|
Adansonia za
|
Za
|
Malvaceae
|
Ecorces, Fruits
|
Construction, Alimentation
|
|
Aloe suzanae
|
Vahondrandra
|
Liliaceae
|
Bois, feuilles
|
Construction, fourrage
|
|
Alluaudia procera
|
Fantsiolitse
|
Didiereaceae
|
Bois, Branches
|
Bois de construction, bois de chauffe
|
|
Breonadia salicina
|
Soaravy
|
Rubiaceae
|
Bois, branches
|
Construction, bois de chauffe, arôme de rhum local
|
|
Capuronia mahafalensis
|
Mantsake
|
Meliaceae
|
Bois
|
Construction
|
|
Cedrelopsis grevei
|
Katrafay
|
Rutaceae
|
Bois, Branches, Écorces
|
Construction, bois de chauffe, médicinale
|
|
Crossandra longipes
|
Sonsonsoy
|
Acanthaceae
|
Plante entière
|
Ornementation
|
|
Dipsys decaryi
|
Lafa
|
Palmae
|
Feulles, graines
|
Construction, alimentation
|
|
Dioscorea sosa
|
Sosa
|
Dioscoreaceae
|
Tubercules
|
Alimentation
|
|
Dioscorea acuminata Gyrocarpus americanus
|
Ovy ala Sirosiro
|
Dioscoreaceae Hernandiaceae
|
Tubercules Bois
|
Alimentation Construction
|
|
Hymenodiction sp.
|
Beholitsy
|
Rubiaceae
|
Bois
|
Construction
|
|
Hyppocratea urceolus
|
Vahipindy
|
Hippocrateaceae
|
Tiges
|
Corde d'assemblage
|
|
Mendoravia sp.
|
Mendoravy
|
Fabaceae
|
Bois
|
Construction
|
|
Neobeguea mahafalensis
|
Handy
|
Meliaceae
|
Feuilles
|
Médicinale
|
|
Ravenala
madagascariensis
|
Ravinala
|
Strelitziaceae
|
Feuilles
|
Construction
|
32
Résultats et interprétations
Tableau 5 : Les utilisations des plantes par la population
(suite)
|
Noms scientifiques
|
Nom
vernaculaire
|
Famille
|
Parties
utilisées
|
Utilisations
|
|
Strychnos
madagascariensis
|
Dagoa
|
Loganiaceae
|
Fruits
|
Alimentation
|
|
Tetrapterocarpon geayi
|
Vaovy
|
Fabaceae
|
Bois
|
Construction
|
|
Uncarina stellulifera
|
Farehitse
|
Pedalliaceae
|
Fruits, feuilles
|
Pièges à rats et shampooing
|
|
Vanilla humblotii
|
Vahinamalo
|
Orchidaceae
|
Tiges
|
Médicinale
|
|
Dracaena reflexa
|
Hasina
|
Dracaenaceae
|
Plante entière, tige
|
Ornementale, médicinale
|
III.2. DESCRIPTION DES HABITATS DE LA ZONE D'ETUDE
Les observations directes et les références
bibliographiques ont permis de déterminer trois types d'habitat des
Acanthaceae. D'Est en Ouest, on a la forêt dense humide de l'Est, la
formation végétale de transition à Dypsis decaryi
et enfin la forêt sèche associée au fourré
typique du Sud. Chaque habitat a ses taxons d'Acanthaceae spécifiques
correspondant à des conditions édaphoclimatiques du milieu.
Quelques exceptions existent car des plantes peuvent se rencontrer à la
fois dans les trois types de milieu. En effet, globalement, ces habitats
correspondent aux différentes parties de la zone d'étude
mentionnées plus haut, à savoir la partie humide, la zone de
transition et enfin la partie sèche dans un milieu nettement plus
sec.
III.2.1. Habitats de la partie humide
La formation végétale y est du type forêt
humide de moyenne ou de basse altitude. La canopée peut atteindre 25m en
moyenne altitude et seulement 10 à 12m en haute altitude. La forêt
est composée d'arbres à cime contiguë, à branches
basses portant de nombreuses épiphytes et présente un sous-bois
assez clair. Les espèces dominantes sont Ravenala madagascariensis
(Strelitziaceae), Tambourisssa (Monimiaceae), Sloanea
rhodantha (Eleocarpaceae), Weinmannia rutenbergii (Cunoniaceae)
Canarium madagascariensis (Burseraceae), Chrysophyllum boivianum
(Sapotaceae), Uapaca sp. (Euphorbiaceae), Pandanus sp.,
Marojejya insignis (Arecaceae), Ocotea leavis (Lauraceae),
Cyathea andohahelensis, Asplenium normale (Aspleniaceae) etc... Une
brousse éricoïde à Phillipia sp. (Ericaceae) et
Imperata sp. (Poaceae) se rencontre sur le sommet et la partie
sommitale. Le sol y est noir en général, argileux et peu profond
parsemé par des affleurements granitiques qui se rencontrent un peu
partout, même dans la forêt.
33
Résultats et interprétations
III.2.2. Habitats de la zone de transition
La partie Centre de la zone d'étude se présente
sous forme d'une bande d'orientation Nord Sud depuis Esomony-Maromby au Nord,
Ankariera Bevilany et le col de Ranopiso au Sud. La localité d'Esomony
est pourvue de formation végétale de transition entre la
forêt dense humide orientale et la formation sèche du Sud. Celle
de Ranopiso présente une formation végétale de type haut
fourré arboré à Dypsis decaryi (Arecaceae) avec
une association d'Alluaudia procera et d'Alluaudia ascendens
(Didiereaceae). Cette formation végétale est
entrecoupée par des savanes herbeuses à Heteropogon contortus
(Poaceae). Cette partie présente aussi une formation
végétale de type forêt sèche à canopée
ouverte tout à fait au Nord.
III.2.3. Habitats de la partie sèche
La partie Sud de la zone d'étude présente une
série de formations végétales climaciques de type
forêts denses sèches associée au fourré à
Didiereaceae et à Euphorbia. Ces deux formations
végétales sont entrecoupées par des forêts galeries
caractérisées par de grands arbres sur sols fertiles longeant les
cours d'eau. En plus de ces taxons caractéristiques, cet habitat
comporte des espèces telles que Cedrelopsis grevei, Diospyros
humbertiana, Commiphora simplicifolia, Grewia cyclea, Aloe suzannae, Croton
mongue, Strychnos decussata, Ravenea xerophila, Aloe helenae, Barleria decaryi
et B. humbertii. Un sol de type ferrugineux tropicaux et des carapaces de
sols ferralitiques de couleur roux, sur un soubassement cristallin, s'y
rencontrent.
Plusieurs groupements végétaux y ont
été identifiés en fonction des conditions
édaphiques :
- des hauts fourrés arborés à
Alluaudia procera sur sol sableux, alluvionnaire ;
- des hauts fourrés arborés à
Alluaudia procera, Megistostegium perrieri sur sol calcaire dans le
moyen bassin de la Mandrare ;
- et de la forêt sèche caducifoliée
à Dalbergia (Fabaceae), Commiphora (Burseraceae) et
Didiereaceae au niveau d'Ihazofotsy.
Cet habitat comporte aussi des formations secondaires
dûes surtout à la pratique de la culture itinérante sur
abattis brûlis. Le sisal, dont les graines sont disséminées
par les oiseaux à partir des plantations d'Amboasary-Atsimo, envahit ces
milieux naturels dégradés.
Résultats et interprétations
III.3. MORPHOLOGIE ET BIOLOGIE DES ESPECES ETUDIEES
Pour pouvoir élaborer une clé de
détermination, une étude détaillée de la
morphologie de tous les échantillons de plantes récoltées
était nécessaire conformément à la
méthodologie développée auparavant.
III.3.1. Caractéristiques morphologiques des
Acanthaceae
L'analyse morphologique des spécimens
étudiés a permis de mettre en évidence les points
importants de leurs caractères. Il s'agit de l'analyse des feuilles, des
fleurs, des fruits et des graines. Différents ports et types biologiques
ont été rencontrés. Les conditions
édapho-climatiques du milieu influent beaucoup sur ces caractères
morphologiques.
III.3.1.1. Type biologique
Les Acanthaceae étudiées présentent
différents types biologiques, à savoir des
nanophanérophytes, des chaméphytes lianescents ou non, des
hémicryptophytes ou même des lianes. Le tableau 6 et la figure 3
ci-dessous les montrent par ordre d'importance.
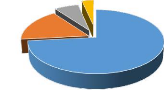
17%
76%
7% 3% Hémicryptophytes
Chaméphytes
Lianes
Nanophanérophytes
34
Figure 3 : Distribution des espèces par type
biologique.
35
Résultats et interprétations
Tableau 6 : Distribution des espèces par `type
biologique'
|
N°
|
Genres et espèces
|
Types biologiques
|
|
Hémi-
cryptophytes
|
Chamé-
phytes
|
Lianes
|
Nano-
phanérophyte
|
|
1
|
Asystasia gangetica
|
X
|
|
|
|
|
2
|
Blepharis calcitrapa
|
X
|
|
|
|
|
3
|
Blepharis maculata
|
X
|
|
|
|
|
4
|
Brillantaisia madagascariensis
|
X
|
|
|
|
|
5
|
Camarotea sp.
|
X
|
|
|
|
|
6
|
Crossandra longipes
|
X
|
|
|
|
|
7
|
Dicliptera decaryi
|
X
|
|
|
|
|
8
|
Ecbolium madagascariensis
|
X
|
|
|
|
|
9
|
Ecbolium syringifolium
|
X
|
|
|
|
|
10
|
Hypoestes decaryana
|
X
|
|
|
|
|
11
|
Hypoestes longituba
|
X
|
|
|
|
|
12
|
Hypoestes poissonii
|
X
|
|
|
|
|
13
|
Hypoestes sp.1
|
X
|
|
|
|
|
14
|
Hypoestes sp.2
|
X
|
|
|
|
|
15
|
Hypoestes sp.3
|
X
|
|
|
|
|
16
|
Hypoestes strigata
|
X
|
|
|
|
|
17
|
Justicia sp.1
|
X
|
|
|
|
|
18
|
Lasiocladus anthospermifolius
|
X
|
|
|
|
|
19
|
Neuracanthus richardianus
|
X
|
|
|
|
|
20
|
Forsythiopsis vincoides
|
X
|
|
|
|
|
21
|
Phaulopsis imbricata
|
X
|
|
|
|
|
22
|
Stenandriopsis carduacea
|
X
|
|
|
|
|
23
|
Barleria alluaudii
|
|
X
|
|
|
|
24
|
Barleria decaryi
|
|
X
|
|
|
|
25
|
Barleria humbertii
|
|
X
|
|
|
|
26
|
Barleria her
|
|
X
|
|
|
|
*27
|
Barleria kitchingii
|
|
X
|
X
|
|
|
28
|
Mendoncia delphina
|
|
|
X
|
|
|
29
|
Ruellia geayi
|
|
|
|
X
|
|
TOTAL :
|
22
|
5
|
2
|
1
|
Sur les vingt-neuf (29) espèces collectées, un
seul représentant des Acanthaceae de la zone d'étude est de type
nanophanérophyte, c'est Ruellia geayi. Cinq (5) espèces
sont des chaméphytes et enfin vingt-deux (22) sont des
hémicryptophytes, soit 76% des espèces collectées. On
constate que Barleria kitchingii évolue biologiquement en
arbuste au stade jeune, en liane en présence de support au stade adulte,
tandis que Mendoncia delphina est toujours une liane. On peut donc
affirmer que la majorité des Acanthaceae du Sud de Madagascar sont des
hémicryptophytes.
36
Résultats et interprétations
III.3.1.2. Appareil végétatif a.
Tiges
Le tableau 5 suivant donne le détail de la
répartition des espèces, par ordre d'importance, en fonction de
la forme des tiges en section.
Les spécimens collectés montrent que les tiges de
ces plantes se présentent sous forme quadrangulaire, subquadrangulaire,
hexagonale ou plus ou moins arrondie ou subcylindrique (Figure 4). Certaines
espèces de cette famille ont des tiges présentant des stries
longitudinales.
Tableau 5 : Répartition des espèces par rapport aux
sections des tiges
|
N°
|
Genre et espèce
|
Section des tiges en coupe transversale
|
|
Quadrangulaire
|
Sub-
quadrangulaire
|
Hexagonale
|
Sub-
cylindrique
|
|
1
|
Asystasia gangetica
|
X
|
|
|
|
|
2
|
Barleria decaryi
|
X
|
|
|
|
|
3
|
Barleria kitchingii
|
X
|
|
|
|
|
4
|
Brillantaisia madagascariensis
|
X
|
|
|
|
|
5
|
Hypoestes decaryana
|
X
|
|
|
|
|
6
|
Hypoestes longituba
|
X
|
|
|
|
|
7
|
Hypoestes sp. 3
|
X
|
|
|
|
|
8
|
Hypoestes strigata
|
X
|
|
|
|
|
9
|
Lasiocladus anthospermifolius
|
X
|
|
|
|
|
10
|
Forsythiopsis vincoides
|
X
|
|
|
|
|
11
|
Ruellia geayi
|
X
|
|
|
|
|
12
|
Stenandriopsis carduacea
|
X
|
|
|
|
|
13
|
Barleria alluaudii
|
|
X
|
|
|
|
14
|
Barleria humbertii
|
|
X
|
|
|
|
15
|
Blepharis calcitrapa
|
|
X
|
|
|
|
16
|
Blepharis maculata
|
|
X
|
|
|
|
17
|
Camarotea sp.
|
|
X
|
|
|
|
18
|
Crossandra longipes
|
|
X
|
|
|
|
19
|
Hypoestes sp.2
|
|
X
|
|
|
|
20
|
Justicia sp. 1
|
|
X
|
|
|
|
21
|
Hypoestes poissonii
|
|
X
|
|
|
|
22
|
Mendoncia delphina
|
|
X
|
|
|
|
23
|
Neuracanthus richardianus
|
|
X
|
|
|
|
24
|
Hypoestes sp.1
|
|
|
X
|
|
|
25
|
Phaulopsis imbricata
|
|
|
X
|
|
|
26
|
Dicliptera decaryi
|
|
|
X
|
|
|
27
|
Ecbolium madagascariense
|
|
|
X
|
|
|
28
|
Ecbolium syringifolium
|
|
|
X
|
|
|
29
|
Barleria her
|
|
|
|
X
|
|
TOTAL :
|
12
|
11
|
5
|
1
|
Résultats et interprétations
3%
17% Quadrangulaire
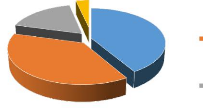
41%
38%
Sub-quadrangulaire
Hexagonale
Sub-cylindrique
Figure 4 : Répartition des espèces par type de tige
vue en section
En section, les tiges peuvent être quadrangulaires pour
douze espèces sur vingt-neuf qui représente 41% des
espèces collectées, subquadrangulaires pour onze, hexagonales
pour cinq et enfin subcylindriques pour une espèce qui est Barleria
her. La section des tiges ne semble pas intervenir dans le système
de classification des Acanthaceae proposé au cours de la présente
étude.
Les représentants de la famille des Acanthaceae ont
plusieurs types de pilosité (Figure 5) et le tableau 7 en donne les
détails.
21% Pubescentes
34%
24%
Villeux
37
Figure 5 : Répartition des espèces par la
présence ou non de poils
38
Résultats et interprétations
Tableau 7 : Répartition des espèces par rapport
à la présence de poils
|
N°
|
Genre et espèce
|
Type de poils
|
|
Pubescents
|
Glabres
|
Hirsutes
|
Villeux
|
|
1
|
Barleria alluaudii
|
X
|
|
|
|
|
2
|
Barleria decaryi
|
X
|
|
|
|
|
3
|
Barleria her
|
X
|
|
|
|
|
4
|
Barleria kitchingii
|
X
|
|
|
|
|
5
|
Blepharis calcitrapa
|
X
|
|
|
|
|
6
|
Blepharis maculata
|
X
|
|
|
|
|
7
|
Camarotea sp.
|
X
|
|
|
|
|
8
|
Justicia sp. 1
|
X
|
|
|
|
|
9
|
Phaulopsis imbricata
|
X
|
|
|
|
|
10
|
Stenandriopsis carduacea
|
X
|
|
|
|
|
11
|
Crossandra longipes
|
|
X
|
|
|
|
12
|
Dicliptera decaryi
|
|
X
|
|
|
|
13
|
Ecbolium syringifolium
|
|
X
|
|
|
|
14
|
Hypoestes longituba
|
|
X
|
|
|
|
15
|
Hypoestes sp.1
|
|
X
|
|
|
|
16
|
Hypoestes sp.2
|
|
X
|
|
|
|
17
|
Lasiocladus anthospermifolius
|
|
X
|
|
|
|
18
|
Ecbolium madagascariense
|
|
|
X
|
|
|
19
|
Asystasia gangetica
|
|
|
X
|
|
|
20
|
Mendoncia delphina
|
|
|
X
|
|
|
21
|
Ruellia geayi
|
|
|
X
|
|
|
22
|
Hypoestes poissonii
|
|
|
X
|
|
|
23
|
Hypoestes sp. 3
|
|
|
X
|
|
|
24
|
Barleria humbertii
|
|
|
|
X
|
|
25
|
Brillantaisia madagascariensis
|
|
|
|
X
|
|
26
|
Forsythiopsis vincoides
|
|
|
|
X
|
|
27
|
Hypoestes decaryana
|
|
|
|
X
|
|
28
|
Hypoestes strigata
|
|
|
|
X
|
|
29
|
Neuracanthus richardianus
|
|
|
|
X
|
|
TOTAL :
|
10
|
7
|
6
|
6
|
Les tiges jeunes sont parfois couvertes de poils et deviennent
glabres au stade adulte. Toutes les parties de la plante peuvent être
couvertes de poils. Dix espèces sur les vingt-neuf collectées
sont pubescentes, sept glabres, six hirsutes et six sont villeuses. Sur les
tiges, les feuilles et les bractées, ces poils peuvent être fins,
denses, longs, courts, grands ou petits. En comparant les genres et les
espèces, la présence de poils n'entre pas dans les
critères de classification de la famille des Acanthaceae utilisés
dans la clé proposée.
Résultats et interprétations
b. Feuilles
En général, les Acanthaceae ont des feuilles
simples, entières et opposées, grandes (Brillantaisia
madagascariensis et Ruellia geayi) surtout dans les zones humide
et de transition ou petites dans la zone sèche pour Lasiocladus
et enfin très petites pour Barleria humbertii. Les
feuilles sont en faux verticilles dans le cas du genre Crossandra longipes
et en verticilles chez Stenandriopsis carduacea. Le limbe est
lancéolé sauf pour Mendoncia qui est multilobé.
Certaines espèces ont des épines au niveau des marges des
feuilles comme le cas de Barleria decaryi, B. humbertii, B. kitchinghii,
Blepharis calcitrapa et Stenandriopsis carduacea ; alors que les
autres espèces sont toutes à bordures lisses. L'adaptation
à la sècheresse des plantes est parfois montrée par
l'apparition du phénomène de spinescence ; elle est
confirmée pour les Acanthaceae sauf pour Neuracanthus richardianus
qui se rencontre à la fois en zones humide et sèche.
c. Epines
Certaines Acanthaceae présentent des épines au
niveau des tiges, des feuilles et même des fruits. Vingt et une (21)
espèces sur les 29 étudiées sont inermes, soit 72% (Figure
6, Tableau 8). Le caractère `épine' n'entre pas dans le
critère de classification proposée dans la présente
étude car les genres Blepharis, Stenandriopsis et
Barleria ont tous des épines mais ne peuvent pas être
groupés dans un seul genre à cause de leurs autres
caractères très différents.
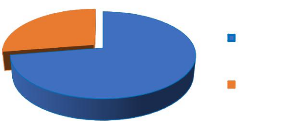
28%
Inermes
Epineuses
72%
39
Figure 6 : Répartition des espèces par la
présence ou non d'épines
40
Résultats et interprétations
Tableau 8 : Répartition des espèces par rapport
à la présence des épines
|
N
|
Genre et espèce
|
Plante
|
|
Inerme
|
Epineuse
|
|
1
|
Asystasia gangetica
|
X
|
|
|
2
|
Barleria kitchingii
|
X
|
|
|
3
|
Brillantaisia madagascariensis
|
X
|
|
|
4
|
Camarotea sp.
|
X
|
|
|
5
|
Crossandra longipes
|
X
|
|
|
6
|
Dicliptera decaryi
|
X
|
|
|
7
|
Ecbolium madagascariense
|
X
|
|
|
8
|
Ecbolium syringifolium
|
X
|
|
|
9
|
Hypoestes decaryana
|
X
|
|
|
10
|
Hypoestes longituba
|
X
|
|
|
11
|
Hypoestes poissonii
|
X
|
|
|
12
|
Hypoestes sp. 3
|
X
|
|
|
13
|
Hypoestes sp.1
|
X
|
|
|
14
|
Hypoestes sp.2
|
X
|
|
|
15
|
Hypoestes strigata
|
X
|
|
|
16
|
Justicia sp. 1
|
X
|
|
|
17
|
Lasiocladus anthospermifolius
|
X
|
|
|
18
|
Mendoncia delphina
|
X
|
|
|
19
|
Forsythiopsis vincoides
|
X
|
|
|
20
|
Phaulopsis imbricata
|
X
|
|
|
21
|
Ruellia geayi
|
X
|
|
|
22
|
Blepharis calcitrapa
|
|
X
|
|
23
|
Blepharis maculata
|
|
X
|
|
24
|
Barleria alluaudii
|
|
X
|
|
25
|
Barleria decaryi
|
|
X
|
|
26
|
Barleria her
|
|
X
|
|
27
|
Barleria humbertii
|
|
X
|
|
28
|
Neuracanthus richardianus
|
|
X
|
|
29
|
Stenandriopsis carduacea
|
|
X
|
|
TOTAL :
|
21
|
8
|
III.3.1.3. Appareil reproducteur
Les Acanthaceae sont toutes des plantes à fleurs mais
des fois les graines sont stériles. En plus de leur capacité de
se multiplier par voie sexuée, les herbes ont la possibilité de
développer des racines au niveau des noeuds facilitant ainsi la
multiplication végétative. Les fleurs sont
généralement petites et vivement colorées.
Il faut mentionner un cas très particulier et unique
pour Hypoestes strigata qui ne se multiplie que par voie
végétative car ses fleurs sont stériles.
41
Résultats et interprétations
a. Inflorescences
La répartition des espèces par type
d'inflorescence est donnée dans le tableau 9 et la figure 7. Les types
d'inflorescence rencontrés sont des cymes, des capitules, des panicules
ou des épis. Quelques espèces ont des fleurs solitaires. Les
inflorescences sont des fois axillaires, subterminales ou terminales. La
majorité des espèces étudiées ont des
inflorescences du type épi.
Tableau 9 : Répartition des espèces par rapport aux
types d'inflorescence
|
Genres et espèces
|
Type d'inflorescence
|
|
Epi
|
Panicule
|
Cyme
bipare
|
Solitaire
|
Capitule
|
Grappe
|
|
Asystasia gangetica
|
X
|
|
|
|
|
|
|
Barleria alluaudii
|
X
|
|
|
|
|
|
|
Barleria decaryi
|
X
|
|
|
|
|
|
|
Barleria her
|
X
|
|
|
|
|
|
|
Barleria kitchingii
|
X
|
|
|
|
|
|
|
Blepharis calcitrapa
|
X
|
|
|
|
|
|
|
Blepharis maculata
|
X
|
|
|
|
|
|
|
Camarotea sp.
|
X
|
|
|
|
|
|
|
Crossandra longipes
|
X
|
|
|
|
|
|
|
Ecbolium madagascariense
|
X
|
|
|
|
|
|
|
Ecbolium syringifolium
|
X
|
|
|
|
|
|
|
Hypoestes sp. 1
|
X
|
|
|
|
|
|
|
Hypoestes sp. 2
|
X
|
|
|
|
|
|
|
Hypoestes sp. 3
|
X
|
|
|
|
|
|
|
Justicia sp. 1
|
X
|
|
|
|
|
|
|
Neuracanthus richardianus
|
X
|
|
|
|
|
|
|
Phaulopsis imbricata
|
X
|
|
|
|
|
|
|
Stenandriopsis carduacea
|
X
|
|
|
|
|
|
|
Brillantaisia madagascariensis
|
|
X
|
|
|
|
|
|
Hypoestes decaryana
|
|
X
|
|
|
|
|
|
Hypoestes strigata
|
|
X
|
|
|
|
|
|
Ruellia geayi
|
|
X
|
|
|
|
|
|
Dicliptera decaryi
|
|
|
X
|
|
|
|
|
Forsthyopis vincoides
|
|
|
X
|
|
|
|
|
Hypoestes poissonii
|
|
|
X
|
|
|
|
|
Barleria humbertii
|
|
|
|
X
|
|
|
|
Mendoncia delphina
|
|
|
|
X
|
|
|
|
Hypoestes longituba
|
|
|
|
|
X
|
|
|
Lasiocladus anthospermifolius
|
|
|
|
|
|
X
|
|
18
|
4
|
3
|
2
|
1
|
1
|
Résultats et interprétations
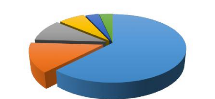
14%
62%
7% 3% 3%
10%
Epi
Panicule Cyme bipare Solitaire Capitule Grappe
Figure 7 : Répartition des espèces par type
d'inflorescence
Dix (18) espèces sur les vingt-neuf (29)
collectées, soit 62% ont le type d'inflorescence en épi.
b. Fruits et graines
Les fruits sont de type silique sauf pour Mendoncia
delphina qui a une baie. Les graines, en général, ont un
diamètre compris entre 2 et 4mm, arrondies, aplaties, ovales, ou
oblongues et la plupart du temps ornementées. Mendoncia delphina
constitue une exception avec une grosse graine de 15mm de diamètre
(Figure 8) et le tableau 10 en donne les détails.
Nombre d'espèces
|
12
10
|
|
3-4mm
2-3mm
1-2mm
1mm
4-5mm
15mm
Diamètres des graines
|
|
8 6 4 2 0
|
42
Figure 8 : Répartition des espèces par classe de
diamètre des graines
43
Résultats et interprétations
Tableau 10 : Répartition des espèces par classe de
diamètre de graines
|
N°
|
Genre et espèce
|
Diamètre des graines
|
|
1
|
Mendoncia delphina
|
15mm
|
|
2
|
Lasiocladus anthospermifolius
|
5mm
|
|
3
|
Ecbolium syringifolium
|
5mm
|
|
4
|
Ruellia geayi
|
5mm
|
|
5
|
Forsythiopsis vincoides
|
4mm
|
|
6
|
Brillantaisia madagascariensis
|
4mm
|
|
7
|
Hypoestes sp. 3
|
3-4mm
|
|
8
|
Justicia sp. 1
|
3-4mm
|
|
9
|
Camarotea sp.
|
3-4mm
|
|
10
|
Asystasia gangetica
|
3-4mm
|
|
11
|
Barleria alluaudii
|
3-4mm
|
|
12
|
Barleria decaryi
|
3-4mm
|
|
13
|
Barleria her
|
3-4mm
|
|
14
|
Hypoestes decaryana
|
3mm
|
|
15
|
Barleria kitchingii
|
3mm
|
|
16
|
Blepharis calcitrapa
|
3mm
|
|
17
|
Blepharis maculata
|
3mm
|
|
18
|
Phaulopsis imbricata
|
3mm
|
|
19
|
Stenandriopsis carduacea
|
3mm
|
|
20
|
Hypoestes longituba
|
2,5mm
|
|
21
|
Hypoestes sp. 2
|
2mm
|
|
22
|
Hypoestes strigata
|
2mm
|
|
23
|
Crossandra longipes
|
2mm
|
|
24
|
Barleria humbertii
|
2mm
|
|
25
|
Neuracanthus richardianus
|
1,5 - 1,8mm
|
|
26
|
Hypoestes poissonii
|
1,5mm
|
|
27
|
Ecbolium madagascariense
|
1-1,5mm
|
|
28
|
Hypoestes sp. 1
|
1mm
|
|
29
|
Dicliptera decaryi
|
0,5mm
|
Les graines des espèces étudiées sont
lisses, parfois rugueuses. Comme elles sont petites, elles sont donc facilement
emportées par le vent et par l'eau de ruissellement en savane. Sinon, en
présence de barrière écologique en forêt ou en
fourré, leur dispersion se fait à courte distance. Ainsi, on
constate que les plantules ne se développent qu'autour de la plante
mère. Une des possibilités de dispersion des graines est aussi
l'exozoochorie. Les capsules, ayant des poils, s'accrochent sur les duvets des
oiseaux et tombent sur leurs parcours de vol. C'est le cas du genre
Barleria. Un spécimen du genre Barleria alluaudii a
été trouvé à 20km à l'Ouest d'Ambovombe,
dans la région de Manja très loin de la zone d'étude.
L'endozoochorie est rencontrée chez le genre Mendoncia
delphina. Les oiseaux frugivores mangent les fruits et assurent la
dispersion des graines qui peuvent alors se retrouver très loin de
l'ensemble du lot.
44
Résultats et interprétations
III.3.2. Clé de détermination des genres
Les premiers critères de classification utilisés
dans la clé proposée ici sont les types biologiques à
savoir ici le port `liane' (1a) et `non liane' (1b),
c'est-à-dire les caractères morphologiques évidents
perçus sur le terrain. Cette clé traite directement les
données relatives aux 29 espèces collectées sans
considération des tribus des Acanthaceae.
1a. Liane, feuilles multilobées, calice en cupule
à bord entier, sinué ou pluridenté ; ovaire biloculaire ou
uniloculaire, chaque loge contenant 2 ovules collatéraux, sessiles,
insérés, par presque toute leur longueur ; graine sans
retinacles, ovaire à une seule loge fertile, calice velu,
bractéole couverte de poils roussâtres, le fruit
est une drupe. .. Mendoncia
1b. Herbe, arbuste ou rarement un arbre.
2a.Arbuste ou rarement un arbre.
3a. Préfloraison tordue.
4a. Tube de la corolle étroite et longuement cylindrique,
sépale concrescent dans leur moitié
inférieure au moins .... Eusiphon =
Ruellia
2b. Herbe
5a. Préfloraison tordue
6a. Les deux étamines de chaque côté
concrescentes par la base des filets.
7a. Corolle profondément bilabiée, la
lèvre supérieure en casque ; les 2 étamines
antérieures stériles, les 2 latérales fertiles, la
postérieure réduite à un minime staminode
Brillantaisia
7b. Corolle bilabiée ou non, dans ce dernier cas la
lèvre supérieure plane, étamines antérieures
toujours fertiles. Corolle non fendue, bilabiée ou 5 lobes
subégaux.
8a. Quatre étamines fertiles, rarement seulement deux et
pas de staminode.
9a. Etamine mutique ou apiculée à la base. A la
déhiscence de la capsule, chaque demi-cloison restant adhérant
à la valve. Corolle à 5 lobes subégaux, non distinctement
bilabiée. Loge des anthères mutiques.
10a. Ovaire contenant dans chaque loge 3 ovules et plus rarement
un seulement
Ruellia
10b. Ovaire contenant 2 - 3 ovules dans chaque loge.
11a. Bractéole petite, plus courte que le calice, non
concrescente.
11b. A la déhiscence de la capsule, chaque demi-cloison
se séparant de la valve sur une partie de sa longueur, sépale
postérieur plus grand que les autres.
8b. 2 étamines et 2 staminodes, corolle bilabiée,
lèvre plus longue que le
tube ......... ..................... ...... ...
Forsythiopsis = Oplonia
45
Résultats et interprétations
5b. Préfloraison quinconciale ou imbriquée. 12a.
Préfloraison quinconciale.
13a. Calice actinomorphe, des cystolithes.
14a. Pièce de l'involucre libre ou brièvement
concrescente à la base de la corolle longue de 10-25mm. Fleur en
glomérule, sessile, axillaire, chacune entourée de 6-8
bractées
libres. Bractée entourant chaque fleur linéaire
ou lancéolée, aiguë Lasiocladus
13b. Calice zygomorphe ; des cystolithes.
15a. Corolle à 5 lobes subégaux ou
l'antérieur plus grand, calice divisé en 4 segments,
l'antérieur et le postérieur plus grands.
Barleria
15b. Corolle bilabiée.
16a. Anthères des étamines postérieures
disjointes ..... Neuracanthus
5c. Préfloraison imbriquée.
17a. Sépale imbriqué ; 4 étamines fertiles
à anthère uniloculaire ; pas de cystolithes.
18a. Corolle unilabiée à 5 lobes peu profonds, les
latéraux parfois indistincts.
19a. Tube de la corolle très court, filet des
étamines antérieures appendiculé
Blepharis
19b. Tube de la corolle allongé, cylindrique, filet des
étamines non appendiculé
... Crossandra
18b. Corolle soit à 5 lobes bien individualisés
soit bilabiée.
20a. Corolle bilabiée.
Stenandriospsis
21a. Feuille entière fortement dentée
épineuse sur les bords.
17b. Sépale non imbriqué ; 2 à 4
étamines fertiles ; des cystolithes.
22a. 2 étamines fertiles.
23a. Plante arbustive ou herbacée. Tige en zigzag ou
rectiligne, quadrangulaire, subquadrangulaire, glabre ou finement pileuse,
pourvue des lignes longitudinales ou transversales.. Limbe ovale,
lancéolée, glabre, parsemée des poils fins. Inflorescence
en épi, en cyme, en panicule, ou en grappe.
24a. Inflorescence en épis terminal et sub-terminal, 2
bractées lancéolées et ovales. Calice en 5 pièces
soudées à la base, linéaire, violette, tube tordu,
lèvres supérieures soudées en une seule pièce de
même que la lèvre inférieure. Carpelle uniloculaire, 4
graines aplaties, 10-11mm de long, fixées au niveau de la partie
antérieure du tube de
la corolle Justicieae sp.1
25a. Feuille de base
triangulaire à subcordée ou cordée avec des sinus,
bractée caduque, graine 4 à 5mm parfois 6mm de longueur. Tube de
la corolle toujours vert (rarement blanche ou violette), tube toujours plus
court que les lobes floraux, lèvre
46
Résultats et interprétations
supérieure bifide, bractée caduque,
étamine érectile et parallèle avec les lèvres
supérieures, filament toujours court, anthère courbée, et
parallèle avec les filaments.
Ecbolium
25b. Tige hexagonale au niveau des sections, inflorescence
axillaire et terminale, cyme pédonculée avec 3 à 5
pédoncules des cymes, Fleurs 2 à 6 par cymule, deux à
trois fleurs développées, bractéole des cymes et
bractéole des capitules de forme ovale à deltoïde. Corolle
toujours tordue (c.-à-d., le tube tordu à 180°)
Dicliptera
25c. Bractée linéaire en 5 pièces, 8mm
de long et de 1mm de large, parsemée des courts poils. Calice
soudé à la base, 5 pièces, linéaire, corolle
tubulaire, lèvre inférieure bilobée, et lèvre
supérieure trilobée, carpelle uniloculaire contenant 4
graines aplaties Camarotea
24b. Inflorescence
axillaire et terminale, en épi, en grappe, en panicule, calice
formé par 4 à 5 sépales linéaires, triangulaires ou
ovales. Corolle 5 lobes, soudés ou libres,
tubulaire, tordue. Hypoestes
22b. 4 étamines fertiles.
26a. Inflorescence terminale, et axillaire, en racème,
en épi, en grappe, calice formé par cinq sépales, corolle
par cinq pièces en forme de tube, deux étamines, carpelle
à deux ou quatre ovules, uniloculaire. Herbe dressée ou
prostrée.
27a. Herbe pérenne dressée. Inflorescence cyme
unipare scorpioïde, terminal spiciforme, lâche. Corolle tubulaire
blanche, longue de 15-17mm, pubescente, 5 lobes irréguliers. Etamines
soudées sur le tube de la corolle. Graines 2 loge ou 4 par capsule,
rugueuse, marge crénelée, face finement
ornementée. Asystasia
27b. Herbe pérenne prostrée. Limbe bilabié,
ovale ou elliptique.
28a. Limbe ovale. Inflorescence terminale ou axillaire,
dorsi-ventrale, bractée foliacée, imbriquée, à
leure aisselle une cyme de 1 à 5 fleurs. Première,
deuxième bractée deltoïde. Calice à 5 pièces
libres, inégal, tube cylindrique, pubescent à l'intérieur.
Corolle glabre, lèvre supérieure bilobée, lèvre
inférieure trilobée. Etamines didynames, incluses,
insérées vers le milieu du tube de la corolle, filet de 2
étamines voisines soudées à la base
Phaulopsis
28b. Limbe elliptique. Bractée de l'épi
opposée et linéaire. Inflorescence axillaire et en épi.
Involucre de bractée obdeltoïde à obelliptique. Calice de 5
sépales, lisses, linéaires, terminé par un mucron. Corolle
tubulaire, de couleur blanche puis devenue rose claire. Deux étamines.
Ovaire uniloculaire, lisse, contenant deux ovules, fruit
47
Résultats et interprétations
de 2mm de long et 1mm de large, avec une fente de
déhiscence longitudinale, deux
graines arrondies par loge, plates environ 0,5mm de
diamètre Justicia sp.2
III.4. DESCRIPTIONS DES TAXONS
A partir de toutes les informations obtenues dans la
présente thèse, une proposition de clé partielle de
détermination est donnée avant de décrire les taxons
étudiés. Les nouvelles données recueillies ont permis de
placer chaque échantillon dans un niveau taxonomique précis.
Vingt-neuf (29) espèces d'Acanthaceae regroupées dans 17 genres
ont été récoltées (Tableau 11) dans leurs lieux de
collecte respectifs et précisant les 12 espèces figurant dans le
tome I de Benoist (1967), mentionnées avec un astérisque. Cinq
nouvelles espèces ont été identifiées et
décrites ici. Il s'agit d'une espèce du genre Camarotea,
trois de Hypoestes et une de Justicia. On a constaté
aussi que quelques espèces n'ont pas pu être rencontrées
dans le site de relevé suite à la transformation des lieux de
collecte en terrain de culture ou en zone d'habitation. Il s'agit de
Crossandra vestita, Ruellia monathos et Ruellia
nummularia.
III.4.1. Asystasia Jussieu
Herbe pérenne dressée, peu ramifiée,
glabre ou hispide sur les nervures et les pétioles. Tiges finement
pubescentes, quadrangulaires, deux côtés opposés arrondis.
Feuilles opposées, décussées, pétiolées,
lancéolées à ovales, simples et entières,
présentant une large gouttière sur la face supérieure.
Limbe ovale, à la base en coin large à arrondie, au sommet aigu
ou brusquement acuminé, légèrement pubescent sur les deux
faces, marge entière, nervures pennées. Inflorescence en
racème terminal spiciforme, lâche. Bractées et
bractéoles petites, triangulaires, acuminées et ciliées.
Pédicelles finement hirsutes avec des poils. Calice formé de cinq
sépales linéaires, soudés à la base,
parsemés de poils. Corolle imbriquée, courtement pubescente,
blanche, composée de 5 pièces soudées en tube à la
base, extrémité du tube s'ouvrant en cinq lobes
irréguliers arrondis, étalés. Lèvre
inférieure striée et tachetée de violet. 2 étamines
fertiles et zéro staminode, soudées sur moitié
supérieure du tube de la corolle, 8-10mm de long. Pistil long de 2-3cm
et ovaire pubescent glanduleux. Deux carpelles soudées, uniloculaire.
Placentation pariétale. Fruit capsulaire en forme de silique à
deux valves, long de 2cm. Etroit à la base, 1 à 2 importants
renflements dans la partie supérieure, parsemée de petits poils
glandulaires. Graines arrondies, 2 ou 4 par capsule, rugueuses, grises, marge
crénelée, faces ornementées.
Résultats et interprétations
Tableau 11 : Liste des taxa collectés
|
N
|
Spécimens
|
Genre et espèce
|
Auteurs
|
Lieux de collecte
|
|
1
|
BVM 057
|
Asystasia gangetica
|
(L.) Anders ex Thwaites
|
Antananolobe, Fenoevo Efita
|
|
2
|
BVM 054
|
Barleria alluaudii*
|
Benoist
|
Bereketa, Behara
|
|
3
|
BVM
113/BVM 132
|
Barleria decaryi*
|
Benoist
|
Bassin de Mandrare près d'Amboasary.
|
|
4
|
BVM 182
|
Barleria her *
|
Benoist
|
Tolagnaro
|
|
5
|
BVM 129
|
Barleria humbertii*
|
Benoist
|
Entre Behara et Tranomaro
|
|
6
|
BVM 117
|
Barleria kitchingii*
|
Baker
|
Ankirikiriky,
Tsimelahy, Esomony
|
|
7
|
BVM
053/BVM 122
|
Blepharis calcitrapa*
|
Benoist
|
Mangatsiaka
|
|
8
|
BVM 101
|
Blepharis maculata*
|
Benoist
|
Tsimelahy
|
|
9
|
BVM 143
|
Brillantaisia madagascariensis*
|
Anderson ex
Lindau
|
Col de Manangotry
|
|
10
|
BVM 120
|
Camarotea sp.
|
|
Nord d'Ankodida
|
|
11
|
BVM
051/BVM 127
|
Crossandra longipes
|
Moore
|
Tranomaro.
|
|
12
|
BVM 159
|
Dicliptera decaryi*
|
Benoist
|
Manangotry
|
|
13
|
BVM 114
|
Ecbolium madagascariense
|
(Vahl) Vollesen
|
Fanota, Isaka Ivondro
|
|
14
|
BVM 070
|
Ecbolium syringifolium
|
(Vahl) Vollesen
|
Antsapana, Tranomaro
|
|
15
|
LRK 756
|
Forsythiopsis vincoides*
|
Benoist
|
Behara
|
|
16
|
BVM 130/
BVM 185
|
Hypoestes decaryana
|
Benoist
|
Bevilany
|
|
17
|
BVM 187
|
Hypoestes longituba
|
Benoist
|
Entre Mahamavo et Ampahiso
|
|
18
|
BVM 118
|
Hypoestes poissonii
|
Phillipson
|
Manangotry
|
|
19
|
BVM 172
|
Hypoestes sp.1
|
|
Manangotry
|
|
20
|
BVM 131/
BVM 143
|
Hypoestes sp.2
|
|
Mangatsiaka
|
|
21
|
BVM
061/BVM 158
|
Hypoestes strigata
|
Benoist
|
Manangotry
|
|
22
|
BVM 131
|
Hypoestes sp. 3
|
|
Entre Tsimelahy et Ebosiky
|
|
23
|
BVM 166
|
Justicia sp. 1
|
|
Manangotry
|
|
24
|
BVM 052
|
Lasiocladus anthospermifolius*
|
Bojer et Ness
|
Bassin de réception de Mananara
|
|
25
|
BVM 163
|
Mendoncia delphina
|
Magnaghi
|
Manangotry
|
|
26
|
LRK 139
|
Neuracanthus richardianus
|
(Nees) Boivini ex Benoist
|
Tsimelahy
|
|
27
|
BVM 123/
BVM 168
|
Phaulopsis imbricata
|
Manktelow
|
Manangotry
|
|
28
|
Eboroke 811
|
Ruellia geayi
|
Benoist
|
Mahamavo, Ambatoabo
|
|
29
|
BVM 111
|
Stenandriopsis carduacea*
|
Benoist
|
Haut bassin de Mandrare
|
49
Résultats et interprétations
III.4.1.1. Asystasia gangetica (L.) Anders ex
Thwaites
Herbe dressée, peu ramifiée, haute de 30-50cm,
glabre ou finement pileuse sur les nervures et les pétioles. Tiges
finement pubescentes, épaisses et violacées au niveau des noeuds,
ramifiées dès la base, quadrangulaires, 2 côtés
opposés arrondis. Feuilles à limbe lancéolé
à ovale, simples et entières, pétiole long de 1-5cm
présentant une large gouttière sur la face supérieure.
Limbe à la base en coin large à arrondie, à sommet aigu ou
brusquement acuminé, 9cm de long et 5cm de large,
légèrement pubescent sur les deux faces, marge entière,
à poils plus denses sur les 4 à 5 paires de nervures
pennées. Inflorescence en racème terminal spiciforme,
lâche, long de 10-15cm. Bractées et bractéoles
triangulaires, acuminées et ciliées. Pédicelles 4mm de
long et finement hirsutes avec des poils. Calice formé de 5
sépales linéaires, soudés à la base, longs de 7mm
et parsemés de poils. Corolle blanche, longue de 15-17mm, pubescente,
extrémité du tube, large de 15mm, s'ouvre en 5 lobes
irréguliers arrondis, étalés. Lèvre
inférieure striée et tachetée de violet. Etamines
soudées sur la moitié supérieure du tube de la corolle,
8-10mm de long. Pistil long de 2-3cm et ovaire pubescent glanduleux.
Bicarpellé, uniloculaire, ovaire supère, placentation
pariétale. Fruit capsulaire en forme de silique à 2 valves, long
de 2cm. Etroit à la base, 1 à 2 importants renflements dans la
partie supérieure, parsemée de petits poils glandulaires. Graines
2 ou 4 par capsule, rugueuses, grises, 3-4mm de diamètre, marge
crénelée, faces finement ornementées.
Floraison : décembre - avril, fructification :
mai-septembre. Nom vernaculaire : Anamalemy.
BVM 057 collecté le 29 octobre 2005 par MBOLA, B.
et PAUL, D., Antananolobe, Manangotry 442.122m/ 155.582m, alt. 520m,
Forêt dense humide, milieu ouvert, sol noir de forêt. Daniel TF et
al. 11108, mars 2007, PN Isalo, 22°29,11'S ; 45°22'77'E, Gautier et
Chatelaine LG 2843, RS Manongarivo. 14°04'S ; 48°17'E ; Daniel TF et
al. 11107, mars 2007, RN1, pk.55, 18°59'08"S ; 47°05'45"E.
50
Résultats et interprétations

Photo 10 : Fragment de tige de Asystasia gangetica
Anders et Thwaites

Photo 11 : Fleur de Asystasia gangetica Anders et
Thwaites
51
Résultats et interprétations

Figure 9 : Asystasia gangetica Anders et Thwaites
Clé de l'espèce
1a. 4 étamines fertiles.
2a. 2 ovules par loge, 4 graines par capsule, pollen avec des
marges et sans épines.
3a. Corolle tubulaire blanche, longue de 15-17mm, pubescente
Asystasia gangetica
52
Résultats et interprétations
III.4.2. Barleria Linné
Herbes ou arbustes à feuilles opposées. A
l'aisselle des feuilles se développent tantôt seulement un
bourgeon feuillé, tantôt deux bourgeons superposés,
l'inferieur produisant un rameau court, constitué par 4 épines
décussées. Inflorescence en racème, terminale ; fleurs
tantôt solitaires à l'aisselle des feuilles supérieures,
tantôt en épis terminaux. Fleur zygomorphe. Calice divisé
en 4 segments, les deux externes plus grands, l'un postérieur à
sommet entier, l'autre antérieur à sommet souvent
émarginé ou brièvement bilobé et deux internes plus
étroits. Corolle en tube cylindrique dans sa partie basilaire ou
évasé portant 5 lobes inégaux, l'antérieur se
détachant du tube soit bien au-dessous des autres, soit presque au
même niveau, les deux postérieurs souvent concrescents un peu plus
haut que les autres. Androcée comprenant 2 étamines
antérieures fertiles, 2 staminodes latéraux beaucoup plus courts,
à anthères rudimentaires et ordinairement un staminode
postérieur réduit à un court filet ; étamines et
staminodes soudés au tube de la corolle. Ovaire biloculaire, chaque loge
contenant un ou deux ovules ; style linéaire terminé par deux
stigmates inégaux. Capsule comprimée, atténuée au
sommet en bec stérile plus ou moins prononcé, contenant 2
à 4 graines discoïdes couvertes de poils hygroscopiques.
III.4.2.1. Barleria alluaudii Benoist
Arbuste pérenne pouvant atteindre une hauteur de 1,5m ;
rameaux subquadrangulaires, pubescents, sillonnés et tomenteux sur deux
faces opposées. Feuilles simples, opposées, brièvement
pétiolées, à pétiole long de 2 à 3mm. Limbes
à bord entier, ovales ou oblongues, atténués à la
base, aigus et spinescents au sommet, glabres au-dessus, portant des poils
épars sur la nervure principale, longues de 20 à 45mm, larges de
9 à 14mm ; un rameau court, épineux à l'aisselle des
feuilles. Inflorescences terminales en épis denses, longs de 2 à
3cm ; bractées disposées sur 4 rangs, les inférieures
semblables aux feuilles, les supérieures progressivement plus
réduites, sessiles, ovales, spinescentes à
l'extrémité, glabres mais ornées sur les bords d'une
frange de cils, longues de 8mm, larges de 4mm, chacune portant à son
aisselle une fleur sessile pourvue de deux bractéoles
lancéolées, spinescentes à l'extrémité,
glabres, ciliées sur les bords dans leur moitié
supérieure, longues de 5mm. Calice composé de 4 pièces.
Sépales externes ovales brièvement acuminés, spinescents
à l'extrémité, ciliés sur les bords dans leur
moitié supérieure, mesurant 8mm de longs sur 5mm de larges,
parcourus par 11 à 13 nervures longitudinales assez saillantes.
Sépales internes lancéolés, brièvement
acuminés ciliés sur les bords, longs de 5mm, larges de 2,5mm.
Corolle composée de 5 pièces, jaune, longue de 40mm,
bilabiée, son tube long de 14mm ; lèvre antérieure
oblongue, entière, longue de 15mm, lèvre postérieure
longue de 25mm environ, semi-tubuleuse à la base, terminée par 4
lobes ovales longs de 15mm. Androcée comprenant 2 étamines
antérieures fertiles à filets pubescents et à
anthères
53
Résultats et interprétations
biloculaires et 2 staminodes petits et stériles
à anthères très réduites. Disque cupuliforme.
Ovaire bicarpellé, biloculaire, conique, glabre, chaque loge contenant
un ovule ; style filiforme, stigmate linéaire. Placentation
pariétale. Capsule longue de 8 à 9mm glabre, ovoïde
comprimée, terminée par un bec court plein contenant dans chaque
loge une graine aplatie, ovale.
Floraison et fructification : toute l'année.
MBOLA, B., BVM 054, 10 juin 2005, Apres le village de
Manja sur la RN 10, 329.982m/ 96.882m, alt. 130m, Haut fourré
arboré à Didiereaceae et Euphorbia, milieu ouvert, sol
alluvionnaire du type argileux de couleur grise. MBOLA, B., BVM 141, 3 mai
2008, Ampilira (croisement route d'Esomony et Tsivory) 403.874m/ 143.182m, alt.
305m, forêt sèche, milieu ouvert, sol alluvionnaire du type
argilo-sableux de couleur roux. PHILLIPSON, P. 2673, parcelle 3, Parc National
Andohahela, 25°01'S 40°38', alt. 100m. CROAT, T. 31844, Ambondro
Ambovombe. Tim WILLING, 13, 4 novembre 1075, entre Tsihombe et Ambovombe.
MABBERLEY, 893, Lakandrano nord d'Ihosy, 22°23'3S 46°08'E, alt.
800-1000m. RAUH, 1188, Beloha, 16 novembre 1959. RAKOTONALA, J.P.P. et al. 186,
Ambatoabo, mars 1994, 24°49'817"S 46°32'244"E, alt. 30-50m.
RAKOTONIANA, 3453, 10 mars 1951, Ranopiso sur terrain sec.

Photo 12 : fragment de tige de Barleria alluaudii
Benoist

Photo 13 : Fleur de Barleria alluaudii Benoist
54
Résultats et interprétations

Figure 10 : Barleria alluaudii Benoist
III.4.2.2. Barleria decaryi Benoist
Herbe pérenne, 35cm de hauteur, rameaux jeunes
quadrangulaires pubescents. Feuilles très brièvement
pétiolées à pétiole long de 1-2mm ; ovales à
oblongues, atténuées à la base, aiguës et
spinescentes au sommet, en dessous pubescent, à nervures secondaires
indistinctes, glabres au-dessus, longues de 1-2cm, larges de 4-6mm ; court
rameau, présence d'épines au niveau de la
55
Résultats et interprétations
tige à l'aisselle des feuilles. Inflorescences
terminales en épis denses, longs de 4-5cm ; bractées
decussées disposées sur 4 rangs, ovales, spinescentes à
l'extrémité, blanchâtres, à 5 nervures, pubescentes,
les unes simples, les autres glanduleuses, capitées, longues de 14-16mm,
larges de 7-10mm ; chacun portant à son aisselle une fleur sessile avec
2 bractéoles linéaires, spinescentes à
l'extrémité, longue de 6mm. Calice composé de 5
sépales, poilus, les segments externes étroitement
lancéolés, acuminés, terminés par un mucron
spinescent, longs de 12mm, large de 3mm, les deux internes semblables mais un
peu plus étroits. Corolle jaune, composée de cinq pétales
soudés à la base formant un tube long de 45mm, large de 35mm,
étroitement cylindrique dans ses deux tiers inférieurs,
légèrement élargi dans son tiers supérieur,
terminé par 5 lobes subégaux, ovales. Androcée comprenant
2 étamines fertiles antérieures exertes et 2 staminodes, petits,
stériles, inclus dans le tube de la corolle, insérées
comme les étamines vers le quart supérieur du tube. Disque en
cupule entourant la base de l'ovaire. Ovaire supère, biloculaire,
bicarpellé, glabre, contenant un ovule dans chaque loge ; style
filiforme, stigmate linéaire. Placentation pariétale. Capsule
longue de 9mm, glabre, ovoïde, comprimée parallèlement
à la cloison, terminée par un bec très court plein,
contenant dans chaque loge une graine aplatie, oblongue.
Floraison et fructification : Toute l'année.
MBOLA, B. et MANERA, F., BVM 113, 30 avril 2007,
Mangatsiaka, 441.642m/ 128.109m, alt. 428m, haut fourré arboré
à Didiereaceae et Euphorbia, milieu ouvert, dépôt argileux
de couleur grise de basse vallée. DECARY 9272, 19 octobre 1931, Ouest
d'Ambovombe. MALCOMBER 1109, 1991, parelle 2 Parc National Andohahela, Nord Est
d'Amboasary, 24°50'S 46°32'E, alt. 100m. DESCOING 233, février
1955, Antanimora. DESCOING 417, Février 155, Tolagnaro.
56
Résultats et interprétations

Figure 11 : Barleria decaryi Benoist
57
Résultats et interprétations

Photo 14 : Fragment de tige de Barleria decaryi
Benoist

Photo 15 : Fleur de Barleria decaryi Benoist
III.4.2.3. Barleria humbertii Benoist
Arbuste pérenne pouvant atteindre une hauteur de 3m,
à tiges parfois sarmenteuses ; rameaux jeunes subquadrangulaires villeux
sur deux faces opposées et marquées par de nombreux cystolithes.
Feuilles opposées, décussées, très
brièvement pétiolées, pétiole long de 1mm, limbes
ovales ou oblongues, obtuses ou arrondies à la base, aiguës au
sommet et terminées par un mucron aigu, spinescentes, à bord
rarement entier, pourvu de chaque côté de 1 à 3 dents
terminées par une spinule dure et aiguë ; face supérieure
portant des poils épars, l'inférieure à poils plus
nombreux. Limbe long de 10 à 15mm, large de 4 à 10mm. Fleurs
axillaires solitaires, sessiles pourvue de 2 bractéoles linéaires
ou lancéolées-linéaires, aplanies spiniformes, vertes,
pourvues de chaque côté de 2-3 dents épineuses, à
pilosité blanche, longues de 15 à 20mm. Calice composé de
cinq pièces. Les pièces externes du calice sont ovales à
oblongues ou oblongues-arrondies à la base, aiguës et spinescentes
au sommet, à bord denté-épineux, nervures formant une
réticulation saillante, couvertes d'une pilosité blanche peu
dense en dehors, longues de 20 à 25mm, large de 8 à 12mm. Les
pièces internes sont lancéolées-linéaires,
aiguës, poilues en dehors. Corolle bleu violacée, longue de 4
à 5cm, composées de cinq pièces soudées à la
base, son tube long de 25 à 28mm un peu élargi à la gorge
; lobes ovales, subégaux. Deux étamines
58
Résultats et interprétations
antérieures fertiles, deux staminodes latéraux
réduits à un court filet, tous insérés vers le
milieu du tube de la corolle. Anthères basifixes et introrses. Ovaire
supère, biloculaire, glabre, chaque loge biovulée. Placentation
pariétale. Capsule glabre, longue de 15mm contenant 4 graines arrondies
et aplaties des deux côtés.
Floraison et fructification : toute l'année.
Nom local : Sitry.
MBOLA, B., et MANAROMBAKA, M., BVM 129, 29 octobre 2007,
Behara, 318.081m/150.916m, alt. 115m, Haut fourré arboré à
Didiereaceae et Euphorbia, sable roux avec affleurement rocheux calcaire.
ALMEDA, RANARIVELO, RANDRIAMBOLOLONA et MBOLA., ALMEDA, 8742, 18 mars 2004,
Station forestière de Sakaraha. DESCOING 2457, 14 février 1957,
route entre Ambovombe et Antanimora, sable roux. BOSSER J., 112, Efoetse
Beheloka, sable beige. T.F. DANIEL, MAGNAGHI, et RANARIVELO, T.F. DANIEL,
11095, 15 mars 2007, RN7, 15km de Toliara, 23°26'89"S 43°45'78"E,
spiny forest. BOSSER, J., 129, 12 juin 1951, Ejeda sur plateau calcaire. TF
DANIEL, 10420, 9 octobre 2003, 28 km au nord de Toliara.

Photo 16 : Fragment de tige de Barleria humbertii
Benoist

Photo 17 : Fleur de Barleria humbertii Benoist
59
Résultats et interprétations

Figure 12 : Barleria humbertii Benoist
60
Résultats et interprétations
III.4.2.4. Barleria her Benoist
Herbe pérenne pouvant atteindre 2m de hauteur, rameaux
jeunes subcylindriques et âgés arrondis, glabres sauf sur deux
sillons longitudinaux opposés pubescents. Feuilles opposées
décussées, subsessiles ; limbes lancéolées ou
lancéolées-linéaires, atténuées à la
base, terminées par un mucron spiniforme, bord entier, glabres, longues
de 1,5 à 7cm, larges de 3 à 7mm, rameau épineux, court
à l'aisselle des feuilles. Inflorescences terminales en épis
assez denses ; bractées longues de 15mm, ces dernières portent
une fleur sessile et porte à son aisselle deux bractéoles
lancéolées - linéaires, spinescentes à
l'extrémité, glabres, longues de 16 à 20mm. Calice de 4
sépales, les deux externes lancéolés - linéaires,
atténués jusqu'à l'extrémité et spinescents
au sommet, longs de 15mm, larges à la base de 2,5 à 3mm, les deux
internes un peu plus effilés. Corolle jaune, rouge ou orangée,
composée de cinq pétales soudés à la base, longue
de 35-40 mm, bilabiée. Tube long de 10mm terminé par cinq lobes
formés par la lèvre antérieure, oblongue, entière,
longue de 18mm environ et les lèvres postérieures longues de 28
mm environ, semi tubuleuses à la base. Deux étamines
antérieures, versatiles, biloculaires, fertiles à filets glabres
et deux staminodes postérieurs à anthères réduites
et stériles ; Ovaire biloculaire, bicarpellée, glabre à
loges uniovulées. Placentation pariétale. Capsule glabre,
ovoïde, comprimée, contenant deux graines arrondies et aplaties des
deux côtés.
Floraison et fructification : Toute l'année.
MBOLA, B., FARAMALALA, M. H., LETSARA, R., RAMAVOVOLOLONA,
et RAZANATSOA, J., BVM 182, 2 août 2009, nord d'Ankodida sur la RN 13,
404.186m/ 124.787m, alt. 99m, Haut fourré arboré à
Didiereaceae et Euphorbia, milieu ouvert, sols argileux brun. MALCOMBER, et
PIGEON, 14955, à 7km à l'Est Près d'Amboasary, route de
Lac Anony, RN 10, 29 Janvier 1990. JAMES et RANDRIANASOLO, 6107, 24 mars 1991,
RN7 entre 18 et 30 km de Toliara.
61
Résultats et interprétations
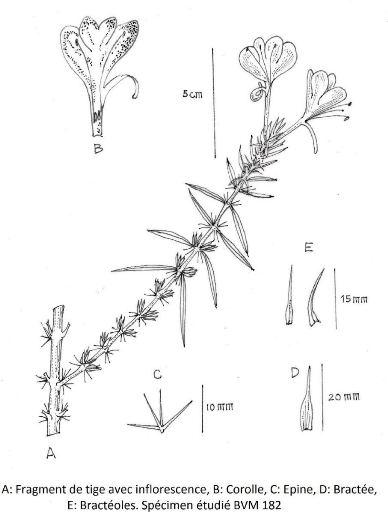
Figure 13 : Barleria her Benoist
62
Résultats et interprétations

Photo 18 : Fragment de tige et inflorescence de Barleria her
Benoist

Photo 19 : Fleur de Barleria her Benoist
III.4.2.5. Barleria kitchingii Baker
Plante pérenne de sous-bois pouvant atteindre une
hauteur de 1,50m, rameuse. Tige quadrangulaire, au stade jeune pubescente et au
stade âgé tomentellé, un peu renflée au niveau des
noeuds. Feuilles simples et entières, pétiolées, à
pétiole 5-6mm de long, lancéolées, aiguës à la
base et décurrentes sur le pétiole, acuminées au sommet,
à bord entier, pubescentes sur les 2 faces, 5-7cm de longues, 1,5-3,5cm
de large. Inflorescences en épi, terminales et axillaires, assez denses
; bractées lancéolées avec les bordures dentées
longues de 10mm et large de 4mm, pubescentes. Bractéoles de dimensions
inégales, la plus grande de forme deltoïde, aiguë à
l'extrémité, 18-22mm de long, 14-20mm de large, scarieuses, ayant
8 à 10 nervures longitudinales partant de la base, anastomosées
en réseau, la plus petite de forme deltoïde, bilobée au
sommet, 16-20mm de long, 8 à 10mm de large et présente des
nervures en réseau ; pourvues sur les bords de 12-15 dents
inégales terminées par un poil ; segments latéraux
lancéolés, courts, poilus sur les bords. Corolle bleue
violacé, 7-8cm de long, tube de la corolle long de 5cm, 5 lobes ovales,
les deux postérieurs un peu plus hautement concrescents que les autres.
Androcée comprenant 2 étamines antérieures fertiles et 2
staminodes à anthères réduites et stériles, tous
insérés vers le tiers inférieur du tube de la corolle.
Disque en cupule entourant la base de l'ovaire.
63
Résultats et interprétations
Ovaire glabre contenant deux ovules par loge. Placentation
pariétale. Capsule glabre, longue de 15-16mm à 4 graines
arrondies et aplatie sur les deux côtés.
Floraison et fructification : toute l'année.
MBOLA, B. et MANAROMBAKA, M., BVM 117, 18 mai 2007, entre
Ankirikiriky et Andragiragy (Behara), 400.309m/140.640m, alt. 98m, haut
fourré arboré à Didiereaceae et Euphorbia et forêt
galerie à Tamarindus indica et Cryptostegia madagascariensis, sol
alluvionnaire. MBOLA, B., MANAROMBAKA, M. et LAMBO, BVM 132, 4 mars 2008,
près bureau du parc à Tsimelahy, 418.785m/130.985m, alt. 178m,
haut fourré arboré à Didiereaceae et Dypsis decaryi, sol
alluvionnaire. BOSSER, J., 15857, juin 1962, entre Amboasary et Tolagnaro,
pk.434. DESCOING B., 2409, 8 juin 1957, St Augustin Toliara. DESCOING, B., 664,
février 1955, Androy.

Photo 20 : Fragment de tige Barleria kitcthingii
Baker

Photo 21 : Fleur de Barleria kitcthingii Baker
Résultats et interprétations
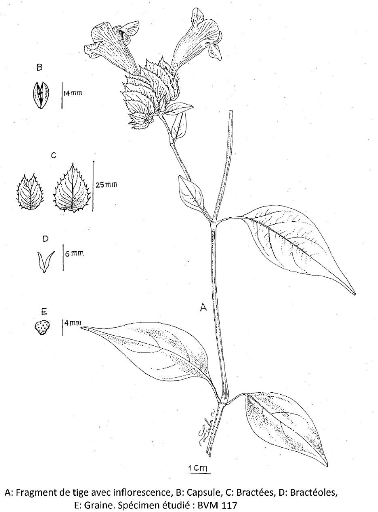
64
Figure 14 : Barleria kitcthingii Baker
65
Résultats et interprétations
Clé des espèces
1a. Un seul ovule dans chaque loge de l'ovaire. Des faisceaux
d'épines à l'aisselle des feuilles de la tige.
2a. Fleur en épis, solitaire à l'aisselle des
bractées de forme différente de celle des feuilles.
Bractées lancéolées ou ovales.
3a. Bractées ovales ; 2-3 nervures secondaire partant de
la nervure principale. Barleria decaryi 4a. Fleur soit isolée,
soit en cyme unipare scorpioide courte à l'aisselle des feuilles
supérieures ; celle-ci, vers l'extrémité
de l'inflorescence souvent réduite en bractéiforme. Corolle
longue, plus de 30mm. Lobe antérieur de la corolle se détachant
du tube beaucoup plus bas que les autres.
5a. Lobes du calice portant sur les bords une frange de courts
poils serrés, les externes
ovales, mucronés. .. Barleria alluaudii
5b. Lobes du calice non frangés sur les bords.
6a. Bractées, au moins les supérieurs glandulaires
; sépales externes étroits, rétrécis
progressivement vers le sommet Barleria her
1b. Ovaire contenant deux ovules dans chaque loge. Segment
externe du calice à bord denté-épineux.
7a. Bractées rigides, terminées par une pointe
spiniforme dure et souvent à bords dentés-
épineux à la base Barleria humbertii
7b. Bractées molles, foliacées, non
terminées par une pointe. Dents des bords du calice
nombreuses ; corolle longue de 5cm. Barleria
kitchingi
III.4.3. Blepharis Jussieu
Herbes ou arbustes à feuilles
généralement épineuses, opposées, mais le plus
souvent rapprochées par 4 en pseudoverticilles, la paire inferieure
souvent plus petites que les autres. Feuilles, sessiles à subsessiles,
limbe à marge entière des fois dentée. Fleur solitaire,
terminale ou axillaire. Fleurs disposées en épis strobiliformes,
ou bien isolées et munies à leur base de 6-8 bractées
stériles ou réunies en cymes lâches ou condensées.
Bractées très souvent épineuses, bractéoles
présentes ou absentes, linéaires au lancéolées,
uninerves, aiguës. Calice 5 sépales inégaux, les deux
latéraux plus étroits, les postérieurs plus larges, les
deux antérieurs concrescents jusque près du sommet. Corolle
gamopétale, unilabiée, fendue du côté
postérieur, à 5 lobes plus ou moins distincts. Androcée de
4 étamines didynames mais peu différentes, soudées
près de l'apex du tube de la corolle ; anthères uniloculaires,
leur bord frangé de poils blancs Ovaire glabre, supère,
biloculaire, chaque loge contient 1 ou 2 ovules. Capsule ovoïde
66
Résultats et interprétations
contenant 2-4 graines. Style linéaire, stigmate bifide.
Capsule oblongue à ellipsoïde. Graines discoïdes, couvertes
par de longs poils hygroscopiques.
III.4.3.1. Blepharis calcitrapa var. decaryi
Benoist.
Herbe pérenne pouvant atteindre 40cm de hauteur
à rameaux jeunes subquadrangulaires, pubescents. Feuilles
disposées par 4 en pseudoverticilles à chaque noeud, subsessiles
ou pétiolées ; limbe décurrent, lancéolé,
atténué à la base, acuminé et aigu à
l'extrémité, à bord entier, pourvus de chaque
côté 1 à 2 épines, long de 4 à 10cm, large de
14 à 35mm, glabre sauf sur les nervures qui portent des poils
épars. Inflorescences en épis terminaux, denses, sessiles,
simples, ayant jusqu'à 12cm de longueur sur 12mm d'épaisseur,
toutes fertiles ; bractées opposées, disposées sur 6 rangs
longitudinaux. Ovales, acuminées et spinescentes au sommet,
atténuées en coin à la base, portant de longs poils blancs
épars, pourvues de 5 à 7 nervures, longues de 10mm, larges de 7
à 8mm ; fleurs sessiles munies de deux bractéoles
lancéolées, acuminées. Calice composé de quatre
pièces, la première terminée par 3 épines, long de
7mm et large de 2mm, la seconde terminée par une épine long de
8mm et large de 3mm, les deux autres terminées par des épines.
Corolle composée de cinq pétales, blanche tachetée de
violet en dedans, longue de 10mm, son tube long de 6mm terminé par cinq
lobes, la lèvre supérieure bilobée, l'inférieure
trilobée. 4 étamines didynames insérées vers le
tiers supérieur du tube de la corolle ; filets courts ; les
antérieures à anthères basifixes, à loge
rudimentaire, versatiles, biloculaire ; les deux postérieures avec une
loge bien développée. Ovaire bicarpellée, biloculaire,
glabre, contenant 2 ovules inégaux dans chaque loge ; style glabre.
Placentation pariétale Capsule longue de 6mm, comprimée,
contenant une seule graine dans chaque loge, terminée par un court bec.
Graine très petite de l'ordre de demi-millimètre de
diamètre, arrondie et aplatie des deux côtés.
Floraison et fructification : toute l'année.
MBOLA, B. et LAMBO, BVM 053, 10 mai 2005, Site Touristique
de Mangatsiaka, 398.058 m/ 150.249m, alt. 422m, Haut fourré
arboré à Didiereaceae et Euphorbia, sableux de couleurs rouge
avec affleurement de roches calcaires. DECARY, 9898, 8 juin 1952, Entre Behara
et Tranomaro, (Isotype). PHILLIPSON, et RABESIHANAKA, 3167, 11 juin 1989, RNI
Tsimanampesotse, 24°04'S 43°46'E, T.F. DANIEL, et al. 10403, 8
octobre 2003, La table, Toliara, 23°24'36"S 43°41'12"E. LETSARA, et
al. LRK 758, entre Behara et Tranomaro à 30km d'Amboasary,
24°45'15"S 46°25'68"E, alt. 173m. T.F. DANIEL, et al. 11 novembre
2007, à 16 km à l'Est de La table, Toliara, 23°24'65"S
43°47'12"E, alt. 50-60m.
Résultats et interprétations

67
Figure 15 : Blepharis calcitrapa Benoist var.
decaryi Benoist.
68
Résultats et interprétations

Photo 22 : Fragment de tige Blepharis calcitrapa
Benoist

Photo 23 : Fleur de Blepharis calcitrapa Benoist
III.4.3.2. Blepharis maculata Benoist
Petite herbe pérenne haut de 50cm à tiges
grêles, pubescente, couchées, radicantes aux noeuds,
subquadrangulaires avec 2 lignes longitudinales. Feuilles opposées et
decussées, simples et entières, groupées par 4 à
chaque noeud, subsessiles avec un pétiole de 1 à 3mm. Limbe
lancéolé ou ovale, atténué ou obtus à la
base, obtus ou arrondi à l'extrémité, pubescente sur les
deux faces, longues de 6 à 38mm, larges de 4 à 13mm. Fleurs
sessiles ou axillaires, les autres sessiles à l'extrémité
d'un rameau et entourées d'un verticille de 4 feuilles ; 8
bractées, pubescente en dehors, les inférieures linéaires,
uninerves, pourvue des taches de couleur brun violace au sommet,
terminées par 1 à 3 épines couvertes de fins poils, les
supérieures lancéolées, échancrées à
l'extrémité avec un appendice étroit au milieu de
l'échancrure. Calice formé par 4 segments inégaux, le
postérieur lancéolé, incisé à
l'extrémité et terminé par un appendice oblong, parcouru
par 3 à 5 nervures, pubescent sur la face externe, long de 10mm, large
de 3,5mm, les latéraux étroitement lancéolés,
uninerves, longs de 10mm, l'antérieur lancéolé,
binervé, bidenté, long de 5mm. Corolle composée de cinq
pièces soudées à la base formant un tube, terminé
par cinq lobes regroupé en deux lèvres inférieures
réduites et trois supérieures, de couleur jaune poussin à
jaune verdâtre, longue de 10mm, son tube long de 3mm, orné en
dedans d'un anneau de poils en dessous
69
Résultats et interprétations
de l'insertion des étamines. Androcée de 4
étamines insérées à la gorge de la corolle, les
postérieures à filet épais, les antérieures
à filet comprimé portant un appendice étroit et aigu ;
anthères ornées d'une frange de poils d'un côté.
Ovaire biloculaire et bicarpellée, style glabre. Placentation
pariétale. Graines arrondies et aplatie des deux côtés.
Floraison et fructification : toute l'année.
MBOLA, B., BVM 101, 14 septembre 2006 (TAN), Tsimelahy,
418.756m/ 130.959m. alt. 217m, Forêt de transition entre la forêt
sèche et le haut fourré arboré, sol gris de forêt.
BOSSER, J., 19075, mars 1964, vallée de Mandrare, Analafaly. MORAT,
4446, Avril 1974, Port Bergé.
70
Résultats et interprétations

Figure 16 : Blepharis maculata Benoist.
71
Résultats et interprétations
Clé des espèces
1a. Plante herbacée. Segment postérieur du
calice ovale ou lancéolé, acuminé ou brusquement
rétréci avant l'extrémité et prolongé par un
appendice étroit. Corolle terminée par trois lobes larges et
obtus, épines des bractées assez longues. Segments
latéraux du calice bien plus court que les autres. . Blepharis
maculata
1b. Arbuste ramifié, segment postérieur du
calice ovale, terminée par un acumen court et aigu,
épine des bractées courtes, aigues. ..
Blepharis calcitrapa
III.4.4. Brillantaisia Beauv.
Herbes à tiges subquadrangulaires; feuilles
opposées, décussées, sans stipules. Fleurs en panicules
terminales amples ou spiciformes, formées de cyme unipare
scorpioïde, lâches ou contractées, situées à
l'aisselle de feuilles ou de bractées. Calice de 5 sépales libres
jusqu'à la base, inégaux, le postérieur souvent plus
grand. Corolle profondément bilabiée, son tube court, brusquement
dilate sous la gorge ; lèvre supérieure en forme de casque
allongé, l'inferieure trilobée ; préfloraison tordue ; 2
étamines fertiles (les latérales) et 2 staminodes
antérieurs, tous insères sur le tube de la corolle;
étamine postérieure représentée par un staminode
réduit à un filet très court; les 2 étamines
fertiles a anthères dorsifixes, les loges disjointes au-dessous du point
d'insertion sur le filet. Ovaire a deux loges contenant chacune plus de 2
ovules ; style filiforme. Capsule subquadrangulaire, loculicide, fertile
dès la base, chaque valve creusée de 3 sillons longitudinaux
retinacles arqués; graines plates, ovales ou orbiculaires.
III.4.4.1. Brillantaisia madagascariensis Anders. ex
Lindau
Herbe pérenne de 50cm de hauteur, à tiges
rameuses souvent couchées et renflées aux noeuds à la
base, puis redressées, au début quadrangulaires et villeux puis
devenant subcylindriques et glabres, renflés au-dessus des noeuds.
Feuilles opposées décussées, simples et entières,
pétiolées, à pétiole long de 1-9cm, ovales,
longuement décurrentes sur le pétiole. Limbe
lancéolé, aigu au sommet, à bord
irrégulièrement festonné portant des poils blancs
au-dessus, pour les stades jeunes pubescentes en dessous sur les nervures,
à nombreux cystolithes visibles surtout au-dessus, 6-30cm de long et
4-15cm de large. Fleurs disposées en panicules spiciformes terminales
longues de 3-22cm, l'axe d'inflorescence portant des bractées
opposées, ovales ou oblongues, un peu concaves, arrondies à
l'extrémité, un peu rétrécies à la base,
velues en dehors, 1-3cm de long ; à l'aisselle de chaque bractée
se trouve un petit cyme unipare scorpioïde de 3 fleurs souvent
réduites à une seule fleur. Bractéoles
lancéolées-spatulées, foliacées, velues, long de
2,5cm et large de 1cm. Calice composé de 5 sépales, libres, un
peu inégaux, linéaires, faiblement élargis
72
Résultats et interprétations
et obtus à l'extrémité, villeux,
glanduleux, 12-25mm de long, large de 1mm. Corolle bleue ou violet claire,
longue de 25mm, tube long de 10mm, lèvre supérieure un peu
élargie dans son tiers inférieur, légèrement
échancrée à l'extrémité, l'inférieur
brusquement renflée et bosselée à la gorge,
trilobée. Etamines antérieures stériles, les
latérales fertiles, filets munis de longs poils à la base,
anthères biloculaires, basifixes, les loges insérées
à la même hauteur sur le filet, divergentes en bas. Disque petit,
régulier. Ovaire uniloculaire et pluriovulée (jusqu'à 40),
poilu. Placentation pariétale. Capsule 20-30mm de long, poilue au
sommet, graines discoïdes, arrondies ou ovales, glabres et lisses,
mesurant 4mm de diamètre.
Floraison et fructification toute l'année.
MBOLA, B., PAUL, D. et ZEROME (TAN), BVM 143, 30 mai 2008,
Col de Manangotry, le long de la RIP 118. Coordonnées GPS (Laborde) :
42.405/153.223, alt : 458m. Forêt dense humide, bord de route. Sol noir
de forêt. P.B. PHILIPSON, et al. 3458, RN13, Parcelle 1 Parc National
Andohahela, 24°45'10"S ; 46°51'15"E, alt. 480m. CREMES G. 2141, mai
1972, route de Ranomafana Ifanadiana, PK 28. PHILLIPSON, P.B., 3585, mars 1992,
RNI Andohahela, parcelle 1, RIP 118, pk 33. 24°45'10"S ; 46°51'15"E,
alt. 480m. De BLOCK, RAKOTONASOLO, F., RANDRIAMBOAVONJY, T., 678. Manangotry
Andohahela, 24°45'39"S; 46°51'49"E, alt. 319m. SANJY., 6312, avril
1954, Marovato Ambanja.

Photo 24 : Inflorescence de Brillantaisia madagascariensis
Anders. ex Lindau

Photo 25 : Fleur de Brillantaisia madagascariensis
Anders. ex Lindau
Résultats et interprétations
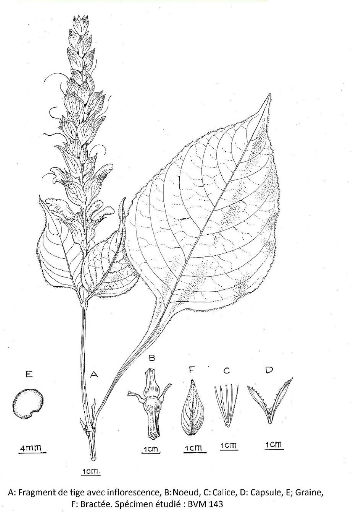
73
Figure 17 : Brillantaisia madagascariensis Anders. ex
Lindau
74
Résultats et interprétations
Clé de l'espèce
1a. Herbe, arbuste ou rarement un arbre.
2a. Préfloraison tordue.
3a. Les deux étamines de chaque côté
concrescentes par la base des filets.
4a. Corolle profondément bilabiée, la lèvre
supérieure en casque ; les 2 étamines antérieures
stériles, les 2 latérales fertiles, la
postérieures réduite à un minime staminode
Brillantaisia madagascariensis
III.4.5. Camarotea Scott-Elliot
Herbe pérenne dressée pouvant atteindre 120cm de
hauteur. Feuilles ovales, arrondies au sommet et atténuées
à la base ; face supérieure glabre, face inférieure
pubescente, nervures secondaires marquées. Inflorescences en épis
terminaux et sub-terminaux. 5 bractées linéaires, pubescentes.
Calice formé de 5 pièces, linéaire soudé à
la base. Corolle rose clair ou blanche, tubulaire, lèvre
inférieure à 2 lobes, et lèvre supérieure à
3 lobes, la partie externe pubescente, la partie intérieure glabre, tube
tordu, lèvre supérieure soudée de même que les
inférieures. Androcée composé de 2 étamines
fertiles, filets pubescents, soudés à la partie antérieure
du tube de la corolle. Anthère basifixe, biloculaire avec une fente de
déhiscence longitudinale. Ovaire uniloculaire contenant 2 à 4
graines, style couvert par des petits poils blancs, stigmate bifide. Graines
ovales, aplaties et rugueuses.
III.4.5.1. Camarotea sp.
Herbe pérenne de sous-bois à tiges
dressées, 120cm de hauteur. Feuilles ovales, arrondies au sommet et
atténuées à la base, 3-12cm de long, et 1-4cm de large ;
face supérieure glabre, couverte de cire, de couleur vert foncé
en général, face inférieure pubescente lui donnant une
couleur mauve, nervures secondaires distinctes. Rameau parfois court.
Inflorescences en épis terminaux et sub-terminaux. Bractées
linéaires vert pâle, formées par 5 pièces, 8mm de
long et 1mm de large, pubescentes. Calice formé par 5 pièces
linéaires, soudé à la base, long de 6mm et large de 0,5mm.
Corolle tubulaire longue de 2,5cm, de couleur rose clair, tordue ; lèvre
inférieure 2-lobées et lèvre supérieure
3-lobées ; partie externe pubescente, donnant sa couleur très
claire, partie intérieure lisse, glabre et de couleur violette.
Lèvre supérieure soudée en une seule pièce de
même que la lèvre inférieure. Androcée
composé de 2 étamines fertiles ; filet long de 10-11mm, couvert
par de petits poils blancs, fixé au niveau de la partie
antérieure du tube de la corolle. Anthère basifixe, biloculaire
avec une fente de déhiscence longitudinale. Ovaire composé de
deux carpelles uniloculaires contenant 4 graines ; style long de 20-25mm,
couvert
75
par de petits poils blancs, stigmate bifide. Fruit sec
à déhiscence longitudinale avec une seule loge contenant 2
à 4 graines. Graines ovales, aplaties, rugueuses, 3-4mm de
diamètre.
Floraison toute l'année, fructification : toute
l'année.
MBOLA, B., FARAMALALA, M. H., LETSARA, R., RAMAVOVOLOLONA
et RAZANATSOA, J., BVM 120, 22 août 2009, RN10, nord Ankodida, entre
Amboasary et Ankariera Bevilany, 403.978m/ 124.755m, alt. 115m, haut
fourré arboré à Didiereaceae et Euphorbia, milieu ouvert,
sol alluvionnaire pourvu d'affleurement de calcaire, de couleur blanche.
PHILLIPSON, 2874 décembre 1989, RN 10, PK 45, Andrevo - Toliara,
23°01'S ; 43°36'E, alt. 25m. DECARY, M.R., 1089, Octobre 1961,
Ampasipolaka - Ambovombe.

Photo 26 : Fragment de tige Camarotea sp.

Photo 27 : Fleur de Camarotea sp.
Résultats et interprétations

76
Figure 18 : Camarotea sp.
77
Résultats et interprétations
III.4.6. Crossandra Salisb.
Arbustes ou plantes herbacées a feuilles parfois
groupées en pseudoverticilles de 4, sans stipules. Inflorescences
terminant la tige et les rameaux, sessiles ou portées par un
pédoncule dénudé. Fleurs sessiles disposées en
épis parfois strobiliformes, chacune située à l'aisselle
d'une bractée et pourvue de deux bractéoles. Calice de 5
sépales inégaux, le postérieur parfois pourvu à la
base d'une callosité saillante, entier, bidenté ou
quadridenté. Corolle longuement et étroitement tubuleuse à
la base ; limbe fendu du côté postérieur jusqu'à la
gorge, à 5 lobes inégaux, les deux postérieurs parfois peu
distincts. Androcée de 4 étamines fertiles, incluses ; filets
courts insérés sur la partie supérieure du tube de la
corolle ; anthères uniloculaires. Ovaire biloculaire contenant 2 ovules
dans chaque loge. Capsule à 4 graines.
III.4.6.1. Crossandra longipes Benoist.
Plante pérenne de sous-bois, 50cm de hauteur. Tige
arrondie, rameau jeune très finement pubescente puis glabre,
entre-noeuds alternativement longs et très courts de sorte que les
feuilles sont presque groupées en verticille de 4, brièvement
pétiolées, atténuées à la base sur le
pétiole ; limbes obtus à l'extrémité pubescente,
3-6cm de long, 10-22mm de large. Inflorescences axillaires portées par
une hampe florale très finement pubescent, 5-25cm de long ; épis
strobiliformes 2-4cm de long. Fleurs zygomorphes, disposées sur 4
rangées longitudinales, situées chacune à l'aisselle d'une
bractée et pourvues de 2 bractéoles. Bractées
inférieures stériles, ovales, les supérieures fertiles,
oblongues, toutes obtuses avec présence de mucrons au sommet,
pubérulentes en dehors et pourvues d'une frange de petits poils blancs
sur les bords, les inférieures longues de 5-8mm, larges de 4-5mm, les
supérieures longues de 10-12mm, larges de 5-6mm. Bractéoles
linéaires, longues de 5mm, larges de 2mm. Calice formé de 5
sépales libres, inégaux, glabres, le postérieur ovale,
long de 7 mm, large de 4 mm, les deux antérieurs
lancéolés, longs de 5mm, larges de 2mm, les deux latéraux
longs de 4mm, larges de 1,5mm. Corolle de couleur saumon ou jaune, longue de
4cm, tube long de 25mm, pubescent en dehors, subcylindrique,
légèrement élargi sous la gorge, lèvre à 5
lobes ovales. Androcée composé de 4 étamines dorsifixes
à anthères uniloculaires incluses dans la partie
supérieure du tube de la corolle ; anthères barbues d'un
côté. Ovaire uniloculaire contenant 2 à 4 graines, glabres.
Placentation pariétale. Capsule longue de 1cm, graine arrondie, aplatie
2mm de diamètre environ, surfaces ornementées.
Floraison et fructification : toute l'année.
78
Résultats et interprétations
MBOLA, B., BVM 051, 10 avril 2005, 408.815m/ 143.401m,
alt. 138m, Zone d'Ihazofotsy sur un milieu ouvert, Haut fourré
arboré à Didiereaceae et Euphorbia, milieu ouvert, de la
vallée de la Mananara entouré d'une formation
végétale type haut fourré arboré arborée
à Alluaudia procera et Cedrelopsis sp. Piste entre le village
d'Ihazofotsy et la limite du parc (parcelle 2), sableux de couleur roux.
PHILLIPSON, P.B., 3458, 7 février 1990, Ampanihy, route d'Androka.
GUILLAUMET, J.L., 2481, avril 1969, au Sud de Betioky-Sud. CROAT, T., 32087, 22
février 1975, RN 13 entre Isoanala et Betroka.
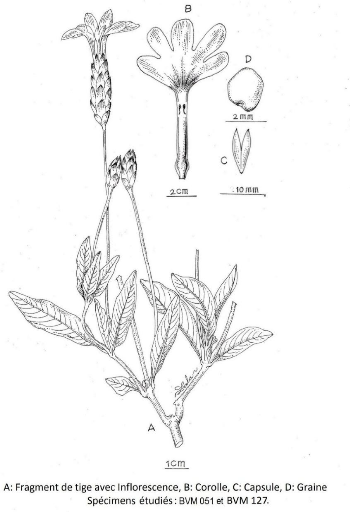
Figure 19 : Crossandra longipes Benoist
79
Résultats et interprétations

Photo 28 : Fragment de tige et inflorescence de Crossandra
longipes Benoists

Photo 29 : Fleur de Crossandra longipes Benoists
Clé de l'espèce
1a. Corolle unilabiée à 5 lobes peu profonds, les
latéraux parfois indistincts. 2a.Tube de la corolle allongé,
cylindrique, filets des étamines non appendiculés.
3a. Feuilles disposées en pseudoverticilles de 4 ;
inflorescence avec hampe florale
Crossandra longipes
III.4.7. Dicliptera Jussieu
Herbe pérenne, grêle peu ramifiée, glabre
ou finement poilue sur les nervures et les pétioles. Les jeunes tiges
ont 6 angles dans leur section. Feuille simple et entière, de couleur
vert clair à foncé. Feuilles opposées
décussées, pétiolées, lancéolées
à elliptiques, arrondies à la base, lisses. Inflorescence en cyme
bipare, axillaire et terminale, 3 à 5 pédoncules de cyme de
couleur vert à marron, long de 3 à 70mm, pédoncules de
cymules longs de 2 à 16,5mm. Bractée accompagnant les divisions
de l'axe d'inflorescence linéaire. Bractéoles des capitules de
formes ovales à deltoïdes arrondies à la base et un peu
arrondies et terminées au sommet par une pointe, parsemées par
des poils épars surtout sur les bordures. Fleurs 2 à 6 par
cymule, deux à trois fleurs
80
Résultats et interprétations
développées. Calice de couleur verte, long de 3
à 4mm. Corolle en cinq pièces, tubulaire. Corolle toujours tordue
à 180°, glabre à la surface externe, lèvre
supérieure entière, lèvre inférieure
3-lobées. Les étamines sont insérées près du
sommet de tube de la corolle, les anthères sont soudées presque
superposées l'une à côté de l'autre, de même
dimension. Ovaire ellipsoïde à obovoide, et le style sort de tube
de la corolle, stigmate à 2 lobes égaux. Graines plus ou moins
arrondies, la surface est couverte de tubercules qui portent des poils
dressés vers le bas.
II.4.7.1. Dicliptera decaryi Benoist
Herbe pérenne, grêle peu ramifiée, haute
de 70cm, glabre à pubescente sur les nervures et les pétioles.
Les jeunes tiges ont 6 angles dans leur section. Feuilles opposées
décussées, simples et entières, portée par un
pétiole long de 2mm à 50mm, feuille lancéolée
à elliptique arrondie à la base et avec 15mm à 90mm de
large et de 45mm à 90mm de long, lisse, à nervures secondaires au
nombre de 4 à 7 sur chaque côté de la nervure principale.
Inflorescence en cyme bipare, axillaire et terminale, 3 à 5
pédoncules de cyme de couleur vert à marron, longs de 3 à
70mm, pédoncules de cymules longs de 2 à 16,5mm. Bractée
accompagnant les divisions de l'axe d'inflorescence linéaire, longue de
2 à 3mm et large de 0,5mm. Bractéoles des capitules de forme
ovales à deltoïdes arrondies à la base et un peu arrondies
et terminées au sommet par une pointe, longues de 2 à 6mm, une
paire de dimension différente longues de 3mm, larges de 2mm, pour la
plus petite et longues de 20mm, larges de 14mm pour la plus grande,
parsemées de poils épars surtout sur les bordures. Corolle longue
de 12mm, son tube long de 7mm. Fleurs 2 à 6 par cymule, deux à
trois fleurs développées. Calice de couleur verte, long de 3
à 4mm. Corolle toujours tordue à 180°, de couleur rose,
glabre de la surface externe, tube long de 4,5 à 30mm, lèvre en
position supérieure entière, longue de 4 à 8mm et large de
2 à 8mm, lèvre en position inférieure 3-lobées, 3
à 9mm de long et 1 à 2mm de large. Les étamines sont
insérées près du sommet du tube de la corolle, les
anthères sont soudées presque superposées l'une à
côté de l'autre, de même dimension, 0,75mm de long et 0,5mm
de large. Ovaire bicarpellée, biloculaire, ellipsoïde à
obovoide, 2mm de long et 1 mm de large, et le style dépasse le tube de
la corolle, 8-9mm de long, stigmate à 2 lobes égaux. Placentation
pariétale, capsule longue de 5mm. Graines plus ou moins arrondies, 1
à 1,5mm de long et 0,5mm de large, la surface est couverte de
protubérance qui portent des poils dressés vers le bas.
Floraison et fructification : toute l'année.
MBOLA, B., FARAMALALA, M. H., LETSARA, R., RAMAVOVOLOLONA,
et RAZANATSOA, J. , BVM 159, 30 mai 2009, Manangotry, 442.298m/ 152.928m, alt.
420m, Forêt dense humide de moyenne altitude type de l'est, sol noir
forestière. MBOLA, B., FARAMALALA, M. H., LETSARA, R., RAMAVOVOLOLONA et
RAZANATSOA, J., BVM 165, 30 mai 2009, Site de Manangotry, sur le RIP 118, PK
48, sous forêt dense humide de moyenne altitude,
Résultats et interprétations
442.300m/ 152.919m, at. 517m, Forêt dense humide de
moyenne altitude type de l'est, sol noir forestière. BOSSER, J., 3625,
Octobre 1952, Tsimbazaza. LETSARA, R., RAKOTONASOLO, F., ANDRIAMIHAJARIVO,
RAZAKAMALALA, LRK 907, Ambinda, Beanka. 18°03'06"S ; 44°31'58"E, alt.
310m. MORAT, P., 61, Octobre 1963, Tsiandro, RS N°9. PHILLIPSON, P., 2249,
Aout 1987, RN Bemaraha, 18°45'S ; 44°48'E, alt. 400m.

Figure 20 : Dicliptera decaryi Benoist
81
82
Résultats et interprétations

Photo 30 : Inflorescence de Dicliptera decaryi
Benoist

Photo 31 : Fleur de Dicliptera decaryi Benoist
Clé de l'espèce
1a. Bractéoles des capitules ovales à
deltoïdes, longues de 2 à 6mm. Corolle long de 12mm, tube long de
7mm. Capsule longue de 5mm. Fleurs 2 à 6 par cymule, deux à trois
fleurs développées.
Calice long de 3 à 4mm. Dicliptera decaryi
III.4.8. Ecbolium Kurz
Herbe ou arbuste ; jeunes tiges pubescentes, tiges
âgées glabres. Feuilles entières et
pétiolées, denses ou espacées, pubescentes avec des larges
nervures secondaires très marquées. Limbes ovales, rarement
elliptiques ou oblongs, obcordées à la base et elliptiques
à oblongs au sommet. Épi, caduc au sommet et présent au
plus bas, couvert par des poils longs et denses, sans glandes. Bractée
souvent semblable à des cônes, elliptique à largement
ovale, bractée couverte par des cils, terminée dans sa partie
supérieure par des glandes. Bractéoles présentes. Calice
linéaire souvent pourvu de cils. Corolle imbriquée, tubulaire ;
lobe central présente la lèvre inférieure, les lobes
latéraux sont plus étroits ; lèvre supérieure
lancéolée linéaire. Ovaire biloculaire, bicarpellé.
Capsule 2-4 côtés, presque lisse à rugueuse, les deux
côtés semblables. 4 graines arrondies par capsule.
83
Résultats et interprétations
III.4.8.1. Ecbolium madagascariense Vollesen
Herbe pérenne, haut de 1,50cm ; jeunes tiges à
six angles, hispite, tiges âgées, arrondies, glabre. Feuilles
entières et pétiolées, denses ou espacées,
pubescentes avec de larges pétioles longs de 223 mm, des nervures
secondaires très marquées. Limbes ovales, rarement elliptiques ou
oblongs, obcordés à la base et elliptiques à oblongs au
sommet, 1-5cm de long et 0,5-3cm de large. Épi long de 1-18cm, caduc au
sommet et présent en bas, pubescent à hispite, sans glandes.
Bractée longue de 2cm, souvent semblable à des cônes,
elliptique à largement ovale, bractée couverte par des cils,
terminée dans sa partie supérieure par des glandes.
Bractéoles longues de 2-6mm. Calice, linéaire, 1-1,5mm de long,
pubescent. Corolle, blanche à jaune citron ou avec une teinte
verdâtre à violette ; tube long de 2,5-4cm ; lobe central
présente la lèvre inférieure jusqu'à 15x10mm de
dimension, les lobes latéraux sont plus étroits ; lèvre
supérieure lancéolée linéaire, jusqu'à
13x15mm. 2 étamines introrses, fertiles long de 5mm. Anthères
basifixes à ouverture en fente longitudinale. Ovaire allongée,
bicarpellée, biloculaire. Placentation pariétale. Capsule 2-4
côtés, 5,5-6,5 de long à 4-5mm de large, presque lisse
à rugueuse, les deux côtés semblables. Graines arrondies et
aplaties des deux côtés avec 5mm de diamètre.
Floraison et fructification : toute l'année.
MBOLA, B. et MANAROMBAKA, M., BVM 114, 30 avril 2007,
Mangatsiaka, limite Ouest de la parcelle 2 du parc, 411.487m/ 128.687m, alt.
428m, forêt sèche, sol argilo-sablonneux de couleur noir.
ANDRIAMIHAJARIVO, et RAKOTOARIVONY, 1193, 20 janvier 2007, entre Beraketa et
Bevia, CR Behara, 24°55'56"S ; 46°25'11"E, 50m. DANIEL, T.F., McDADE,
RANARIVELO, 10419, Ifaty, pk 28 RN9, 23°05'54"S ; 43°36'36"E, alt.
5m. McPHERSON, G., PIGEON, M., 14935, janvier 1990, près village
d'Ihazofotsy, Behara. BOSSER, J., 17271, février 1963, Ankazoabo.
PHILLIPSON, P.B., 2611, novembre 1987, RS Beza Mahafaly, 23°39'S ;
44°38'E, alt. 140m. DESCOINGS, B., 234, février 1935, Antanimora,
Androy. DANIEL, T.F., McDADE, RANARIVELO, 10412, octobre 2003, Piste
d'Ambohimahavelona, Toliara, pk 13, 23°26'36"S ; 43°54'12"E.

Photo 32 : Ecbolium madagascariense Vollesen
84
Résultats et interprétations
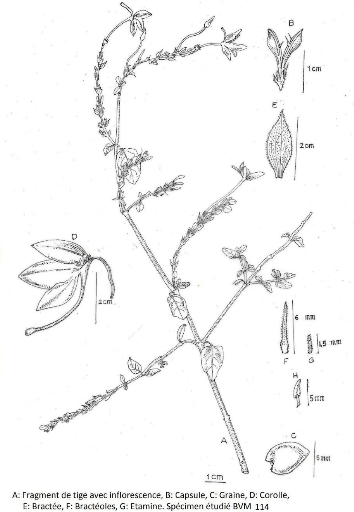
Figure 21 : Ecbolium madagascariense Vollesen
85
Résultats et interprétations
III.4.8.2. Ecbolium syringifolium (Vahl) Vollesen
Herbe ou arbuste pérenne haut de 2m, jeunes tiges
lisses et rarement pubescentes. Feuilles entières et
pétiolées, lisses ou faiblement pubescentes avec des
pétioles longs de 5 à 60 millimètres, des nervures
secondaires très marquées. Limbe ovale, des fois elliptique ou
oblong, subcordé à la base et elliptique à oblong au
sommet, de dimensions 40 à 85mm de long et 5 à 15mm de large. Epi
long de 3 à 19cm, pédoncule 5 à 15mm de long, glabre,
rarement pubescent, sans glandes. Bractées lancéolées,
elliptiques à obovales, longues de 15 à 25mm et 5 à 12mm
de large y compris une pointe qui est long de 2 à 5mm, couvertes de
poils long de 1mm, bractéoles
1 à 3mm de longueur. Calice à 5 lobes,
linéaires, 4 à 7mm de long. Corolle de couleur jaune poussin
à jaune citron, composée de cinq pièces soudées
à la base et au sommet ouverte par cinq lobes inégaux.
Lèvre inférieure, longue jusqu'à 10 à 17mm et large
de 12mm, lèvre supérieure linéaire, longue jusqu'à
13 à 15mm sur 1mm. Le tube de la corolle est de 2,5 à 3,5cm de
longueur.
2 étamines introrses, fertiles, long de 3,5mm.
Anthères basifixes à ouverture en fente longitudinale. Ovaire
bicarpellé et biloculaire. Placentation pariétale. Capsule 2
à 4 graines par loge, de dimension 10 à 14mm de long et 4
à 5,5mm de large, presque lisse à rugueuse, les deux
côtés semblables. Graines arrondies et aplaties des deux
côtés avec 5mm de diamètre.
Floraison et fructification : toute l'année.
Nom local : Sandrisandry
MBOLA, B., BVM 070, 23 Novembre 2005, Analamantsaky,
420.559m/ 123.869m, alt. 220m, haut fourré arboré à
Didiereaceae et Euphorbia, milieu ouvert, sol alluvionnaire de bas fond, lit de
cours d'eau. MALCOMBER, 1113, 1991, Parcelle 2, Parc National Andohahela,
Befandily, 24°50'S 46°32'E, alt. 100m, near Ihazofotsy, LETSARA, et
al. LRK 721, 7 décembre 2008, Andohahela, Befandily 24°56'49"S
46°88'44"E, alt. 196m, LETSARA, et al. LRK 737, 7 décembre 2008,
Andohahela, Befandily 24°56'49"S ; 46°88'44"E. McPHERSON, G., et al.,
14762, janvier 1990, Petriky, Tolagnaro, 25°05'S ; 46°52'E, alt. 15m.
De BLOCK, RAKOTONASOLO, RANDRIAMBOAVONJY, LEYMAN, RESSEIN, 714, janvier 1999,
Andohahela entre parcelle 1 et parcelle 2, 25°12'24"S ; 46°38'23"E,
alt. 149m. McPHERSON, G., et al., 14872, janvier 1990, Mandena, Tolagnaro,
24°57'S ; 47°00'E, alt. 2-15m. R.N., 5021, janvier 1953, RNI
Andohahela, Behara-Amboasary Sud.
Résultats et interprétations
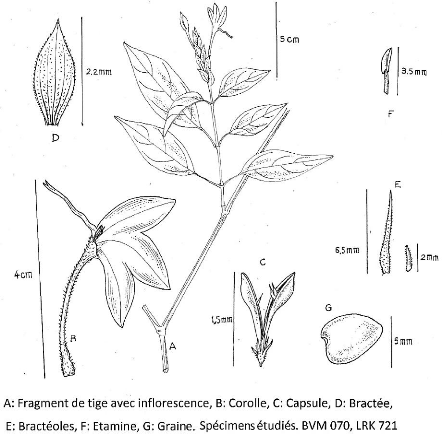
86
Figure 22 : Ecbolium syringifolium (Vahl) Vollesen
87
Résultats et interprétations

Photo 33 : Fragment de tige et inflorescence de Ecbolium
syringifolium (Vahl) Vollesen

Photo 34 : Fleur de Ecbolium syringifolium (Vahl)
Vollesen
Clé des espèces
1a. Tube de la corolle long de 2 à 4cm, tiges
arrondies, présence des glandes au niveau des nodules. Bractée en
forme de cône, 4 graines par capsule, graines à 4mm de
diamètre et 7mm de longueur. Fleurs 3 à 5 dans la partie basse de
l'inflorescence, avec un pédicelle de 1mm de longueur.
2a. Jeunes tiges glabres ou couvertes par des fins poils,
feuilles large de 2,5 ou 4 à 15cm de longueur, bractée en forme
de cône, la petite 15-25mm de longueur ... Ecbolium
syringifolium
2b. Jeunes tiges couvertes de poils épars ou denses,
la plus large feuille, de dimension 1-4(5)cm de longueur, bractée
souvent en forme de cône, la plus petite 5-12(15)mm de longueur
Ecbolium madagascariense
III.3.9. Forsythiospis Baker
Herbes ou arbustes ramifiés, à feuilles
opposées, entières, sans stipules, insérées sur des
coussinets saillants. Fleurs axillaires, solitaires ou en ombelles ;
pédicelles munis vers leur base de 2 bractéoles opposées
très petites, linéaires ou lancéolées. Calice de 5
sépales subégaux, libres jusque près de la base. Corolle
assez distinctement bilabiée à tube cylindrique ; lèvre
supérieure
88
Résultats et interprétations
bilobée, l'inferieure trilobée ;
préfloraison tordue. Androcée de 2 étamines fertiles (les
antérieures) et 2 staminodes, tous insérés un peu
au-dessous de la gorge de la corolle, chacune des étamines ayant la base
de son filet concrescente avec le staminode situé du même
côté. Ovaire biloculaire contenant deux ovules dans chaque loge ;
style filiforme très brièvement bilobé à
l'extrémité. Capsule pleine et stérile dans sa
moitié inferieure contenant 4 graines aplaties.
III.3.9.1. Forsythiopsis vincoides Benoist
Arbuste pérenne atteignant 3 m de hauteur,
ramifié, à feuilles opposées, entières, sans
stipules, pétiole long de 2-6 mm, lancéolées ou ovales,
atténuées à la base, obtuses au sommet, à bord
entier, jeunes pubescentes devenant bientôt glabres, longues de 3-8 cm,
larges de 22-34 mm, insérées sur des coussinets saillants. Fleurs
axillaires, solitaires ou en cymes tri-florées, portées par des
pédicelles longs de 1-3 cm., munis à leur base de deux petites
bractées lancéolées, pubescentes puis glabres, longues de
3mm. Calice composé de 5 sépales subégaux, aigus, au
début pubescents puis à peu près glabres, libres jusque
près de la base, long de 4-5mm. Corolle assez distinctement
bilabiée à tube cylindrique, blanche, bleue ou violette, longue
de 15 mm, tube long de 6 mm, glabre ou pubescent en dedans; lèvre
supérieure bilobée, l'inférieure trilobée;
préfloraison tordue. Androcée constituée de 2
étamines fertiles (les antérieures) et 2 staminodes, tous
insérés un peu au-dessous de la gorge de la corolle, chacune des
étamines ayant la base de son filet concrescente avec le staminode
situé du même côté ; filets glabres ou pubescents ;
anthères basifixes, biloculaires, environ 4 fois aussi longues que
larges. Ovaire bicarpellée, biloculaire, contenant deux ovules dans
chaque loge glabre ; style éparsement poilu ; style filiforme
très brièvement bilobé à l'extrémité.
Placentation pariétale. Capsule, longue de 20 mm, glabre, 4 graines
lisses, arrondies et aplaties des deux côtés.
Floraison juillet - décembre, fructification juillet -
janvier.
LETSARA, R., FARAMALALA, M. H., RAKOTONASOLO, F., MBOLA,
B., PAUL, LRK 756, 8 décembre 2009, 24°51'15"S/ 46°25'68"E,
à 30km de Behara, route de Tsivory, sur un milieu ouvert, Haut
fourré arboré à Didiereaceae et Euphorbia, milieu ouvert,
de la vallée de la Mandrare entouré d'une formation
végétale type haut fourré arboré arborée
à Alluaudia procera et Cedrelopsis sp., sol sableux de couleur roux.
RABEVOHITRA 2403, à l'Ouest d'Ambovombe. MESSMER, N., RAKOTOMALAZA,
P.J., ANDRIATSIFERANA, M.N., 185, Parcelle 2, Ihazofotsy, 24°49,0'S
46°36,6'E, alt. 120m. HUMBERT, H. et PERRIER DE LA BATHIE, H., 2506,
septembre 1924,
Environ de Toliara. DECARY, 4273, janvier 1927 Behara,
Amboasary Sud.
Résultats et interprétations

89
Figure 23 : Forsythiopsis vincoides Benoist
90
Résultats et interprétations

Photo 35 : Fragment de tige et inflorescence de Forsythiopsis
vincoides Benoist

Photo 36 : Fleur de Forsythiopsis vincoides Benoist
Clé de l'espèce
1a. Feuille ovale ou lancéolée,
pétiolée .. Forsythiopsis vincoides
III.3.10. Hypoestes Solander ex Brown
Arbuste ou herbe pérenne dressée, très
ramifiée, glabre ou finement pileuse sur les nervures et les
pétioles et se développe surtout dans les endroits
dégagés. Tiges finement pubescentes, épaisses au niveau
des noeuds, ramifiées dès la base, quadrangulaires. Feuilles
pétiolées simples et entières. Limbe ovale avec une base
et un sommet arrondi, glabre sur les deux faces, marge entière.
Inflorescence en épi. Bractées très petites triangulaires,
pubescentes, acuminées et ciliées. Calice formé de 4
sépales linéaires, soudés à la base et
parsemés de poils dans sa partie interne et lisse dans sa partie
externe. Corolle tubulaire, imbriquée, blanche, ornementée de
rose au milieu, tube 1mm de long. Lèvre supérieure
trilobée et lèvre inférieure soudée en un seul
lobe. 2 étamines fertiles insérées sur le tube de la
corolle, style blanc et sac pollinique rose. Pistil blanc et stigmate rose.
Ovaire ovoïde, biloculaire, chaque loge contenant environ 2 ovules. Fruit
sec à déhiscence longitudinale, en forme de capsule conique, long
de 10mm, avec à l'intérieur 2 graines par loge. Graine arrondie,
3mm de diamètre et aplatie, surfaces ornementées.
91
Résultats et interprétations
III.3.10.1. Hypoestes decaryana Benoist
Herbe pérenne dressée, très
ramifiée, 90cm de hauteur, glabre ou tomentellée sur les nervures
et les pétioles, se développe surtout dans les endroits
dégagés. Tiges pubescentes, épaisses au niveau des noeuds,
ramifiées dès la base, quadrangulaires. Feuilles simples et
entières, portées par un pétiole long de 1-2cm. Limbe
ovale avec une base et un sommet arrondi, 2cm de long et 1,5cm de large, glabre
sur les deux faces, marge entière. Inflorescence disposée en
panicule. Bractées lancéolées, très petites
triangulaires, pubescentes, acuminées et ciliées. Calice
formé de 4 sépales linéaires, soudés à la
base, 3-4mm de longs et parsemés de poils dans sa partie interne et
lisse dans sa partie externe. Corolle composée de cinq pétales,
blanche ornementée de rose au milieu, tube 1mm de long. Lèvre
supérieure trilobée et lèvre inférieure
soudée en un seul lobe. 2 étamines fertiles
insérées sur le tube de la corolle, filament blanc et sac
pollinique rose. Anthère basifixe, biloculaire. Ovaire ovoïde,
biloculaire, chaque loge contenant environ 2 ovules. Pistil blanc et stigmate
rose. Placentation pariétale. Fruit sec à déhiscence
longitudinale, en forme de capsule conique, long de 10mm, avec à
l'intérieur 2 graines par loge. Graine arrondie, 3mm de diamètre
et aplatie avec une épaisseur de 0,5mm, surfaces ornementées.
Floraison : toute l'année, fructification : toute
l'année.
Nom local : Heja
MBOLA B. et F. MANERA., BVM 086, 23 novembre 2005,
Bevilany, 416.18m! 124.259m, alt. 78m, Milieu ouvert dans un
écosystème de haut fourré arboré à
Didiereaceae et Euphorbia, sol alluvionnaire. MBOLA B. et F. MANERA., BVM 130,
29 octobre 2007, Tsimelahy, 418.756m! 130.959m, alt. 217m, haut fourré
arborée à Didiereaceae et Euphorbia sp., habitat ouvert, sol
alluvionnaire. MBOLA, B., FARAMALALA, M. H., RAMAVOVOLOLONA, et LETSARA, R.,
BVM 185, 22 août 2009, Bevilany, 416.18m! 124.259m, alt. 77m, haut
fourré arboré à Didiereaceae et Euphorbia, milieu ouvert,
sol alluvionnaire. HUMBERT, H., 11243, octobre 1933, Forêt de Besomanty
entre Fiherenana et Isahaina (Mangoky). BOSSER, J., 4075, novembre 1952,
Ifotaka, Amboasary-Sud. MANJAKAHERY, et PHILLIPSON, 119, Ankililoaka, Toliara
II, 22°45'50"S ; 45°34'07"E, alt. 86m. DECARY, M., 10666, janvier
1939, Andrahomana, Tolagnaro. PHILLIPSON et al., 3771, octobre 1990,
forêt de Zombitse, Sakaraha, 22°53'S ; 44°40'E, alt.
700m.
92
Résultats et interprétations

Figure 24 : Hypoestes decaryana Benoist
93
Résultats et interprétations

Photo 37 : Fragment de tige et inflorescence de Hypoestes
decaryana Benoist

Photo 38 : Fleur de Hypoestes decaryana Benoist
III.3.10.2. Hypoestes longituba Benoist
Herbe pérenne de sous-bois, de 50cm à 60cm de
hauteur, à tiges dressées et radicantes aux noeuds à la
base, au début quadrangulaires et pubescentes au stade jeune, puis
devenant subcylindriques et glabres au stade âgé, renflées
au-dessus des noeuds. Feuilles opposées, décussées,
entières, pétiolées, à pétiole long de 1,5-3
cm, obliques, lancéolées et parsemées par de petits poils
blanchâtres, de dimension 8 à 14,5cm de long et de 2,5 à
5,5cm de large. Nervures très développées. Fleurs
terminales et axillaires au moins sur les deux derniers entre-noeuds.
Inflorescence en capitule avec chancue portant 3 à 5 épis,
pédoncules courts. Bractée couverte de fins poils
blanchâtres, triangulaire au niveau de chaque épi, long de 1mm.
Calice à 5 lobes soudés à la base long de 6 mm, large de
1mm. Corolle, blanche, tubulaire, terminée par 5 lobes, deux lobes
inférieurs et trois supérieurs, avec un tube long de 1,5cm. 2
staminodes. 2 étamines fertiles soudées au tube de la corolle,
à anthère biloculaire, long de 1 cm. Pistil avec un stigmate
à deux lobes, long de 3 cm. Ovaire bicarpellée, biloculaire.
Placentation pariétale. Chaque loge
Résultats et interprétations
contient 2 à 4 graines aplaties, discoïdes, arrondies
ou ovales, glabres et lisses, mesurant 2,5mm de diamètre.
Floraison : juin - juillet ; fructification : Août
Nom local : Velatra
BVM 178 collecté le 23 août 2009 par MBOLA,
B., FARAMALALA, M. H., RAMAVOVOLOLONA, LETSARA, R., ZEROME et RAZANATSOA, J.,
Tanatana, Manangotry. 441.520m/
152.527m. al. 510m.
Forêt humide, milieu fermé. Sol noir de bas fond, type rendzine.
BOSSER, J., 6987, septembre 1953, Route d'Anosibe Moramanga, pk 15. DECARY,
MR., 7922, bassin d'Antsalova, juin 1930. RALIMANANA, H., 970, Analabe,
Ambatofinandrahana, 20°37'15"S ; 46°50'19"E. DESCOING, B., 3787,
juillet 1958, Vallée d'Ihosy, pk. 18. MORAT, P., 4615, juillet 1974,
Bongolava, Ouest de Tsiroanomandidy, alt 1300m.

Photo 39 : Fragment de tige et inflorescence de Hypoestes
longituba Benoist

94
Photo 40 : Boutons floraux de Hypoestes longituba
Benoist
95
Résultats et interprétations

Figure 25 : Hypoestes longituba Benoist
96
Résultats et interprétations
III.3.10.3. Hypoestes poissonii Phillipson
Herbe pérenne près des zones humides ou flaques
d'eau permanentes, en touffe, haut de 50cm. Tige subquadrangulaire, avec quatre
stries longitudinales, lisse, présence de quelques poils au niveau des
stries le long de la tige au stade adulte et hirsute au stade jeune.
Entre-noeuds très longs jusqu'à 8cm, noeud très
renflé et lisse. Feuilles opposées décussées,
entières, pétiolées, pétiole long de 4mm. Limbes
à nervures principales très marquées surtout sur la face
inférieure, lancéolés, glabres, long de 4,5cm, large de
1,8cm. Inflorescence en épi lache terminale et axillaire.
Bractées opposées, linéaires longs de 7mm,
bractéoles lisses, 5 lobes, terminées par une pointe. Calice, 4
lobes, lisses, longs de 7mm. Corolle rose ornée de blanc,
composée de 5 pièces soudées à la base, tubulaire,
tordue à 360°, long de 10mm, lèvres inférieures, deux
lobes soudés, en position ventrale, et lèvre supérieure 3
lobées soudées. Etamine composée de 2 anthères
fertiles soudées au tube de la corolle. Anthère basifixe à
ouverture en pore. Ovaire bicarpellé et biloculaire. Placentation
pariétale. Capsule longue de 8mm, glabre, avec chaque loge contient 2
à 4 graines. Pistil lisse, bifide, long de 18mm. Graines arrondies, 2mm
de diamètre, aplaties, 0,5mm d'épaisseur.
Floraison et fructification : toute l'année.
MBOLA, B. et MANAROMBAKA, M., BVM 112, 28 septembre 2006,
Tsimelahy, près piscine naturelle, 418.756m / 123. 689m, alt. 170m, Haut
fourré arboré à Didiereaceae et Euphorbia, milieu ouvert,
sol gris avec affleurement des roches calcaires. HUMBERT, H., 2651, Belavenoka,
vallée de l'Onilahy Toliara (Isotype).

Photo 41 : Hypoestes poissonii Phillipson
Résultats et interprétations

97
Figure 26 : Hypoestes poissonii Phillipson
98
Résultats et interprétations
III.3.10.4. Hypoestes strigata
Herbe pérenne de 50cm de hauteur, à tiges
ailées, renflées au-dessus des noeuds, quadrangulaires,
pubescente au stade jeune et glabre au stade âgé. Feuilles
opposées, décussées, simples et entières,
pétiolées, à pétiole long de 1-9cm, ovales,
longuement décurrentes sur le pétiole. Limbe
lancéolé, aigue au sommet, à bord normal lisse sur les
deux faces, long jusqu' à 8cm et large jusqu'à 2,8cm.
Inflorescence en grappe d'épi axillaire, l'axe d'inflorescence portant
des bractées de déférentes dimensions, ovales ou
oblongues, arrondies à l'extrémité, un peu
rétrécies à la base, longues de 1-3cm ; à
l'aisselle de chaque bractée se trouve un petit cyme de 3 fleurs souvent
réduit à une seule fleur. Calice composé de 5
sépales soudés à la base, linéaire, long de 3mm.
Corolle violet ou rose claire, longue de 8mm, le tube de la corolle long de
2mm, lèvre supérieure à 3 lobées et la lèvre
inférieure 2 lobées. Androcée composé de 2
étamines fertiles, basifixes, filet soudé au niveau du tube de la
corolle. Loge pollinique à fente de déhiscence longitudinale.
Aucune capsule n'est rencontrée au cours des misions
sur le terrain.
MBOLA, B., BVM 061, 09 novembre 2005. Manangotry, sur le
RIP 118, PK 31. 442.617m/ 150.841m, al. 417m. Forêt humide, milieu
ouvert. Sol noir de forêt. MBOLA, B. et PAUL, D., BVM 158, 30 mai 2009,
Manangotry, endroit dégagé, le long d'un ruisseau longeant la
route RIP 118, près du village de Fanota. 442.758m/150.192m, alt. 410m,
Forêt humide, milieu ouvert. Sol noir de forêt. MBOLA, B.,
FARAMALALA, M. H., RAMAVOVOLOLONA, et LETSARA, R., BVM 175, 23 août 2009,
Manangotry, circuit touristique. 442.621m/ 150.852m, alt. 417m, Forêt
humide, milieu fermé. Sol noir de forêt.

Photo 42 : Tige de Hypoestes strigata

Photo 43 : Fleur de Hypoestes strigata
99
Résultats et interprétations

Figure 27 : Hypoestes strigata
100
Résultats et interprétations
III.3.10.5. Hypoestes sp. 1
Herbe pérenne, 40 à 50cm de hauteur, rameuse.
Tige subquadrangulaire, en zigzag, lisse et renflée aux entre-noeuds.
Feuilles opposées, décussées, simples et entières.
Limbes lancéolés, atténués et plus ou moins
asymétriques à la base, l'un très grand et l'autre petit,
anisophylles, à position alternée, pétiolée,
pétiole 5 à 6mm de long, nervation très marquée,
à bord entier, lisse, face supérieure couverte de cire, vert
foncé, luisant, limbe 5 à 7cm de long, 1,5 à 3,5cm de
large. Inflorescences en ombelle terminales et axillaires, assez denses,
pédonculées formées de deux à trois cymes;
bractées linéaires avec les bordures dentées long de 5mm,
pourvues de poils épars. Bractéoles lisses, 5 lobes,
terminés par une pointe. Calice à 4 lobes plus ou moins
linéaires soudés à la base, long de 7 mm, pourvu de poils
épars. Corolle violette, composée de 5 pièces
soudées à la base en tube, long de 10mm, tordue à
360°, lèvres inférieures, deux lobes et lèvre
supérieure 3 lobées. 4 étamines composées de 2
staminodes et 2 anthères fertiles soudées au tube de la corolle.
Anthère basifixe. Loge pollinique à fente de déhiscence
longitudinale. Pistil lisse, bifide, long de 18mm. Ovaire à deux
carpelles, glabre contenant 2 à 4 ovules par loge. Placentation
pariétale. Capsule glabre, longue de 6mm et 2 à 4 graines
terminées par un mucron. Graine arrondie de 1,5mm de diamètre et
aplatie.
Floraison et fructification : toute l'année.
MBOLA, B. et PAUL, D., BVM 064, 9 novembre 2005,
Manangotry, 442.841m/ 151.324m, alt. 512m, forêt dense humide, milieu
fermé, sol noir de forêt. MBOLA, B., FARAMALALA, M. H.,
RAMAVOVOLOLONA, LETSARA, R., RAZANATSOA, J., et JEROME., BVM 172, 22 Août
2009, Manangotry, 442.850m/ 151.389m, alt. 505m, forêt dense humide,
milieu fermé, sol noir de forêt.

101
Figure 28 : Hypoestes sp. 1
Résultats et interprétations

Photo 44 : Fragment de tige de Hypoestes sp. 1

Photo 45 : Fleur et fruit de Hypoestes sp. 1
III.3.10.6. Hypoestes sp. 2
Herbe pérenne dressée, touffue haute
d'environ 100cm, se développe bien dans un endroit dégagé.
Tige de couleur rose au stade jeune et à six angles, tomentellée
à glabre et arrondie au stade adulte. Feuilles opposées
décussées, entières et pétiolées,
lancéolées. Pétioles parsemés de poils et longs de
5 à 15mm. Limbe lisse sur sa face supérieure et rugueuse
sur sa face inférieure, lancéolé, souvent
atténué à l'apex. Limbes parsemés de poils fins des
deux côtés surtout au niveau des nervures principales et
secondaires. Il mesure jusqu'à 6 à 85mm de long et 12 à
32mm de large. Inflorescence en épi dense. Bractée double,
triangulaire, lisse, et de couleur verte. 5 pétales, libres et
linéaires forment le calice. Corolle 6mm de longueur, tordue à
360°, composée de 5 pétales soudés à la base
et forme un tube long de 1mm, de couleur blanche et devenus jaunes au stade
âgé, deux lèvres inférieures, trois
supérieures, soudées. Etamines au nombre de 2 et les filets
soudés au niveau du tube de la corolle. Ovaire bicarpellé,
uniloculaire avec 1 à 2 graines. Placentation pariétale. Fruit
sec à déhiscence longitudinale et long de 5 à 6mm et large
de 2mm. Graine aplatie de 1mm diamètre, rugueuse.
102
Floraison et fructification : toute l'année.
Résultats et interprétations
MBOLA, B., FARAMALALA, M. H., RAMAVOVOLOLONA, et LETSARA,
R., BVM 171, 22 août 2009, Manangotry, sur la piste reliant la RIP 118 et
Tanatana, 441.614m/ 151.183m, alt. 415m, forêt dense humide, milieu
ouvert, sol noir de bas-fonds. MBOLA, B., FARAMALALA, M. H., RAMAVOVOLOLONA, et
LETSARA, R., BVM 176, 23 août 2009, Manangotry, près
d'Androangaloaka, PK 42, 442.841m/ 151.324m, alt. 315m, forêt dense
humide, milieu ouvert, sol noir de forêt.

Photo 46 : Fragment de tige et inflorescence de Hypoestes
sp. 2

103
Photo 47 : Fleur de Hypoestes sp. 2
Résultats et interprétations
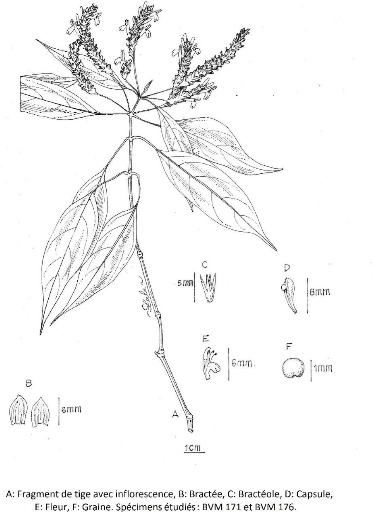
104
Figure 29 : Hypoestes sp. 2
105
Résultats et interprétations
III.3.10.7. Hypoestes sp. 3
Plante herbacée, pérenne, de sous-bois à
tiges subquadrangulaires, dressées pouvant atteindre 120cm de hauteur.
Feuilles pétiolées et entières, pétiole 2-3mm de
long. Limbe ovale et arrondi au sommet, atténué à la base,
la face supérieure glabre, couverte de cire, de couleur vert
foncé en général, la face inférieure pubescente
luire donnant une couleur mauve, nervures secondaires distinctes, 3-12 cm de
long, et 1-4 cm de larges. Rameau parfois court. Inflorescences en épis
terminaux et sub-terminaux, 2 bractées lancéolées et
ovales, un peu rétrécies à la base, vertes, 2mm de long et
0,5mm de large. Calice soudé à la base, 5 pièces,
linéaire, long de 6mm et large de 0,5mm. Corolle, la partie externe
couverte par de courts poils blancs donnant sa couleur blanche, la partie
intérieure lisse sans poils de couleur violette, tube tordu à
360°, long de 8-9mm, lèvres supérieures terminées par
trois lobes et lèvre inférieure par deux lobes.
Gynécée composé de carpelle uniloculaire contenant 4
graines. Style couvert par des petits poils blancs, long de 22-24mm, pistil
bifide. Androcée composé de 2 étamines fertiles. Filet,
couvert par des petits poils blancs, 10-11mm de long, soudé au niveau de
la partie antérieure du tube de la corolle. Anthère basifixe avec
une fente de déhiscence longitudinale. Ovaire biloculaire et
bicarpellée. Placentation pariétale. Fruit sec à
déhiscence longitudinale avec une seule loge contenant 2 à 4
graines. Graines rugueuses, aplaties, ovales, 3-4mm de diamètre.
Floraison toute l'année, fructification : toute
l'année.
MBOLA, B., BVM 131, 4 mars 2008, Ebosiky, 422.257m/
130.837m, alt. 175m, Entre Tsimelahy et Ebosiky dans une forêt galerie
à Breonadia salicina, sol alluvionnaire dans une formation de
forêt galerie. MBOLA, B. et PHILIBERT, BVM 143, 4 avril 2008, Ebosiky,
422.261m/ 130.839m, alt. 178m, Entre Tsimelahy et Ebosiky dans une forêt
galerie à Breonadia salicina, sol alluvionnaire.
Résultats et interprétations
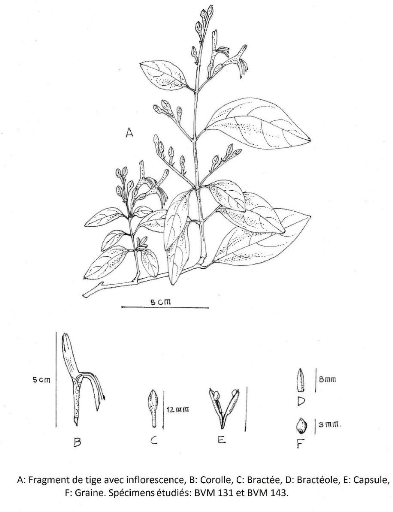
106
Figure 30 : Hypoestes sp. 3
107
Résultats et interprétations

Photo 48 : Fragment de tige et inflorescence de Hypoestes
sp. 3

Photo 49 : Inflorescence de Hypoestes sp. 3
Clé des espèces
1a. Calice à 4 lobes.
2a. Inflorescence en panicule ou en épi, bractée
triangulaire.
3a. Bractées triangulaires, inflorescence en épi
... Hypoestes decaryana
3b. Bractées triangulaires, inflorescence en capitule
Hypoestes longituba
2b. Inflorescence en épi ou en panicule.
4a. Bractées linéaires, inflorescence en
épi. Hypoestes poissonii
4b. Bractées ovales, inflorescence en panicule.
Hypoestes strigata
4c. Inflorescence en ombelle Hypoestes sp.1
4d. Inflorescence en épi dense Hypoestes
sp.2
4e. Bractée de l'épi opposée et
linéaire, inflorescence en épi axillaire. Involucre de
bractée obdeltoïde à obelliptique. Calice de 5
sépales, lisses, linéaires, terminé par un mucron. Corolle
tubulaire, de couleur blanche puis devenue rose claire. Deux étamines
introrses. Ovaire uniloculaire, lisse, contenant deux ovules, fruit de 2mm de
long et 1mm de large,
deux graines arrondies par loge, plates d'environ 0,5mm de
diamètre Hypoestes sp.3
108
Résultats et interprétations
III.3.11. Justicia Linnaeus
Herbe pérenne à port prostré, formant un
tapis sur le sol. Tige grêle, quadrangulaire, pubescente, parsemé
de petits poils épars, chaque entre-noeud touchant le sol
développe des racines grêles. Feuille pétiolée,
glabre, simple, entière. Limbe elliptique, arrondi à la base et
un peu arrondi au sommet. Bractée de l'épi opposée et
linéaire. Inflorescence axillaire et en épi dense. Fleurs
disposées en quatre rangées. Involucre de bractée
obdeltoïde à obelliptique, atténuée à la base
et arrondie au sommet. Calice formé de 5 sépales, lisses,
linéaires terminés par un mucron. Corolle tubulaire, de couleur
blanche puis devenue rose claire. Androcée composé de 2
étamines fertiles, filet couvert par de petits poils blancs, 1-2mm de
long, fixé au niveau de la partie antérieure du tube de la
corolle. Loge pollinique basifixe avec une fente de déhiscence
longitudinale. Ovaire uniloculaire, lisse, contenant deux ovules ; fruit
allongé avec une fente de déhiscence longitudinale, graines
arrondies et plates.
III.3.11.1. Justicia sp. 1
Herbe pérenne à port prostré, formant un
tapis sur le sol. Tige grêle, quadrangulaire, pubescente, chaque
entre-noeud touchant le sol développe des racines grêles. Feuilles
opposées, décussées, glabres, simples, entières,
pétiolées, pétiole long de 6mm. Limbe elliptique, arrondi
à la base et un peu arrondi au sommet, jusqu'à 15mm de long et
9mm de large. Bractée de l'épi opposée et linéaire,
longue de 2mm et large de 1mm. Inflorescence axillaire et en épi dense,
long de 2cm et épaisse de 5mm. Fleurs disposées en quatre
rangées. Involucre de bractée obdeltoïde à
obelliptique, atténuée à la base et arrondie au sommet,
5mm de long et 3mm de large. Calice formé de 5 sépales, libres,
lisses, linéaires longs de 2mm, terminés par un mucron. Corolle
composée de 5 pièces soudées à la base, tubulaire
s'ouvrant en cinq lobes, de couleur blanche puis devenue rose claire, deux
lèvres inférieures et trois supérieures. Androcée
composé de 2 staminodes et de 2 étamines fertiles, filet
pubescent 1-2mm de long, fixé au niveau de la partie antérieure
du tube de la corolle. Loge pollinique basifixe avec une fente de
déhiscence longitudinale. Ovaire uniloculaire, lisse, contenant deux
ovules. Placentation pariétale. Fruit sec de 2mm de longs et 1mm de
large, avec une fente de déhiscence longitudinale, graines arrondies et
plates d'environ 0,5mm de diamètre.
Floraison et fructification : toute l'année.
MBOLA, B., FARAMALALA, M. H., LETSARA, R., RAMAVOVOLOLONA
et RAZANATSOA, J., BVM 166, 22 Août 2009 (TAN), Manangotry, sur la RIP
118. 442.849m/ 151.326m, alt. 510m, Forêt dense humide de moyenne
altitude, milieu ouvert, sol gris de forêt. LETSARA, R., FARAMALALA,
RAKOTONASOLO, MBOLA, PAUL, LRK 722, décembre 2008, Befandily-Tsimelahy,
CR Ankariera Bevilany, 24°56'49"S ; 46°88'47"E, alt. 169m.
Résultats et interprétations
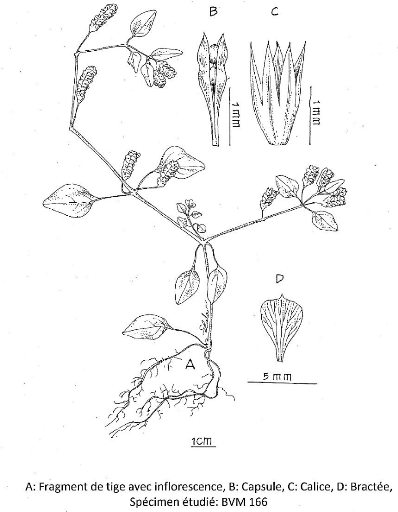
109
Figure 31 : Justicia sp. 1
110
Résultats et interprétations

Photo 50 : Fragment de tige et inflorescence de Justicia
sp. 1

Photo 51 : Inflorescence de Justicia sp. 1
Clé de l'espèce
1a. Sépales non imbriqués ; 2 à 4
étamines fertiles ; des cystolithes.
2a. 2 étamines fertiles.
3a. Plante arbustive ou herbacée, tiges en zigzag ou
rectiligne, quadrangulaires, subquadrangulaires, glabre ou finement pileuse,
limbe ovale, lancéolé, glabre, parsemé de poils fins.
Inflorescence en épi ou en grappe.
4a. Plante arbustive ou herbacée, tige pourvue des
lignes longitudinales ou transversales. 5a. Inflorescences en épis
terminaux et sub-terminaux, 2 bractées lancéolées et
ovales. Calice en 5 pièces soudées à la base,
linéaires, violettes, tube tordu, lèvres supérieures
soudées en une seule pièce de même que la lèvre
inférieure. Carpelle uniloculaire, 4 graines aplaties, 10-11mm de long,
fixée au niveau de la partie antérieure du tube de la corolle
Justicia sp.1
III.3.12. Lasiocladus Bojer ex Ness
Herbes pérennes à feuilles opposées,
pétiolées ou sessiles, dépourvues de stipules.
Inflorescences axillaires en cymes pauciflores sessiles très
contractées. Fleurs hermaphrodites, zygomorphes, chacune pourvue d'un
involucre forme de 6-8 bractées fibres. Calice à 5 sépales
inégaux ou subégaux. Corolle composée de cinq
pétales, préfloraison quinconciale, bilabiée, la
lèvre inférieure trilobée, la supérieure plus
courte, bilobée. Androcée composé de 4 étamines
111
Résultats et interprétations
didynames insérées isolement sur la corolle, un
petit staminode postérieur filiforme. Disque court, annulaire. Ovaire
bicarpellé, biloculaire, chaque loge contenant deux ovules
superposés. Capsule atténuée et aigue au sommet, contenant
4 graines discoïdes.
III.3.12.1. Lasiocladus anthospermifolius (Bojer et
Ness) Benoist.
Herbe pérenne dressée, peu ramifiée,
haute de 30-50cm, glabre ou pubescente sur les nervures et les pétioles.
Tige ramifiée dès la base, quadrangulaire, à stries sur
les deux faces opposées, les 2 autres côtés sont arrondis,
épaissies et de couleur violacée au niveau des noeuds. Feuilles
simples et opposées, portées par un pétiole de 1-5cm de
long, présentant une large gouttière sur la face
supérieure. Limbe ovale à arrondi à la base, aigue au
sommet ou brusquement acuminé, 9cm de long et 5cm de large,
légèrement pubescent sur les deux faces, marge entière.
Poils plus denses sur les 4 à 5 paires de nervures pennées.
Inflorescence disposée en racème terminal spiciforme,
lâche, long de 10-15cm. Bractées et bractéoles petites,
triangulaires, acuminées et ciliées. Pédicelle 4mm de
long, finement hirsute avec des poils non glandulaires et glandulaires. Calice
formé de 5 sépales linéaires égaux, soudés
à la base, longs de 7mm et hirsute. Corolle blanche, composée de
cinq pièces soudées à la base, longue de 15-17 mm,
à base tubulaire un peu bosselée, pubescente.
L'extrémité du tube, large de 15mm, s'ouvre en 5 lobes
irréguliers arrondis, étalés. Lèvre
inférieure striée et tachetée de violet. 4 étamines
insérés isolément sur environ 3,5mm du fonds du tube de la
corolle, 8 à 10mm de long. Anthère basifixe, deux loges. Ovaire
bicarpellé et uniloculaire, pubescente, glanduleuse. Placentation
pariétale. Pistil long de 2-3cm. Fruit capsulaire en forme de silique
à 2 valves, long de 2cm. Capsule étroite à la base,
à 1 à 2 importants renflements dans la partie supérieure
et uaiguë au sommet, parsemée de petits poils glandulaires et non
glandulaires. Graines au nombre de 2 ou 4 par capsule, rugueuses, grises, 3-4mm
de diamètre ; marge crénelée, faces finement
ornementées.
Floraison et fructification : toute l'année.
MBOLA, B., RYVAH, R., BVM 052, 10 mai 2005, Mangatsiaka,
près site de camping du parc, 411.724m/ 128.151m. alt. 120m, Haut
fourré arboré à Didiereaceae et Euphorbia, milieu ouvert,
sol beige de bas-fonds (Alluvionnaire). HUMBERT 12591, décembre 1933,
Vallée d'Anadabolava, Mandrare moyen (Isotype). Du PUY et Du PUY, LABAT,
PHILLIPSON, MB 604, 3 février 1990, La table Toliara sur la RN7,
23°23'S 44°50'E. LETSARA et al. LRK 719, 8 décembre 2008,
Tsimelahy.
Résultats et interprétations
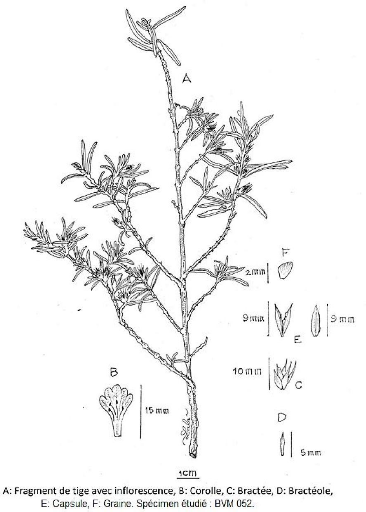
112
Figure 32 : Lasiocladus anthospermifolius Benoist.
113
Résultats et interprétations

Photo 52 : Fragment de tige et inflorescence de Lasiocladus
anthospermifolius Benoist.

Photo 53 : Fleur de Lasiocladus anthospermifolius
Benoist.
Clé de l'espèce
1a. Plantes à poils blancs ou blanchâtres ;
feuilles à bords roulés ou incurvés en dessous. Face
inférieure des feuilles à pilosité blanche assez longue.
Corolle longues de 12-18mm
.. Lasiocladus anthospermifolius
III.3.13. Mendoncia Velloso in Vandelli
Lianes à feuilles opposées, sans stipules.
Fleurs axillaires, pédicellées, munies chacune de 2 grandes
bractéoles opposées enveloppant la fleur. Calice, tordu, en
cupule à bord entier ou sinué. Corolle à tube
infundibuliforme et à 5 lobes plus ou moins inégaux, parfois
groupés en 2 lèvres assez distinctes. Androcée
composée de 4 étamine didyname; anthères biloculaires,
aiguës au sommet, à loges arrondies et velues à la base,
s'ouvrant par de très courtes fentes subapicales. Disque annulaire.
Ovaire uniloculaire contenant 2 ovules collatéraux. Drupe renfermant 1
à 2 graines.
114
Résultats et interprétations
III.3.13.1. Mendoncia delphina Magnaghi
Haute liane pérenne à tiges jeunes grêles,
tomenteux dans leur jeunesse, devenant ensuite glabres. Feuilles
opposées décussées à pétiole long de 5
à 14mm, hérissées de poils roussâtres; limbe ovale,
4 à 8cm de long, 2 à 4,5cm de large, arrondi à la base,
plus ou moins longuement acuminé au sommet, à bord entier et
nervation pennée, très rugueuses en dessus et portant des poils
roussâtres sur les nervures principales, les dessous de toutes les
nervures sont velues. Fleurs solitaires ou disposées par 2 à
l'aisselle des feuilles ; pédicelles longs jusqu'à 4cm,
hérissés de poils roussâtres ; bractéoles ovales
à elliptiques, longues de 22mm, couvertes en dehors de poils
roussâtres, glabres en dedans, cohérentes par leurs bords avant la
floraison et formant un sac rempli d'eau mucilagineuse. Calice en 5
pièces soudées, cupuliforme, couvert de poils roussâtres,
à bord entier. Corolle en 5 pièces soudées à la
base, rouge, violette ou violette foncée, longue de 4cm, glabre, son
tube long de 3cm, étroit, courbe dans son tiers supérieur,
s'élargissant un peu sous la gorge, lobes un peu inégaux. 4
étamines composées de deux staminodes et de deux étamines
fertiles, insérées vers les deux tiers du tube de la corolle ;
anthères biloculaires, dorsifixes, pubescentes à la base,
s'ouvrant par de très courtes fentes subapicales. Disque cupuliforme,
glabre. Ovaire uniloculaire, composé de deux carpelles, densément
velu ; style glabre ; stigmate brièvement bilabié. Placentation
centrale. Fruit de type drupe, violette ou noir rougeâtre, ovale à
elliptique, glabre, de dimension 13,5mm sur 8,5mm.
Floraison : Juin -juillet, Fructification : Août
-septembre.
MBOLA, B., FARAMALALA, M. H., RAMAVOVOLOLONA, LETSARA, R.
et RAZANATSOA, J. (TAN), BVM 163 collecté le 22 août 2009.
Manangotry, 442.298m/ 152.928m, alt. 460m. Forêt humide, milieu
fermé, bord de route, Sol noir de forêt. MALCOMBER et al. 1665,
oct. 1992, RNI n°11, 24°45'S, 46°51'E, alt. 500m. Forêt
humide. RAZAKAMALALA et al. 2302, Brickaville, Andaka, 18°25'16",
48°48'23"E, alt. 698m, Oct. 2005. N. DUMETZ 1395, Nov. 1990, Forêt
de Manantantely Fort-Dauphin. SCHATS et al. 3189, RS Manombo, RN 12 sur
latérite, 9-11 mars 1977. MALCOMBER 2119, 6-13 février 1993, RNI
Andohahela, parcelle 1, Nord Est d'Eminiminy, 24°38'S 46°46'E, alt.
500-1000m. BERNARD et RASOAMIADANA 945, Amparihy, CR Maitsoandakana, Mahajanga,
14°55'50"S 49°25'16"E.
Résultats et interprétations

Photo 54 : Fragment de tige de Mendoncia delphina
Magnaghi.

115
Photo 55 : Fruit de Mendoncia delphina Magnaghi.
Résultats et interprétations
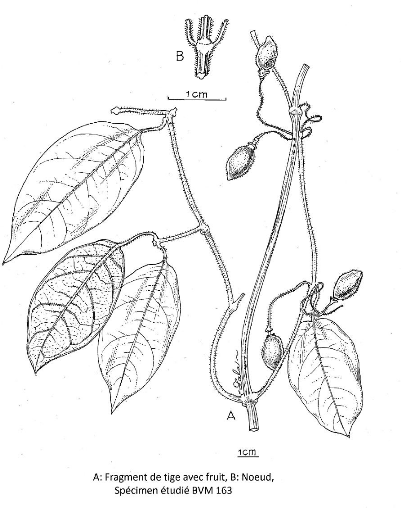
116
Figure 33 : Mendoncia delphina Magnaghi.
117
Résultats et interprétations
III.3.14. Neuracanthus Ness.
Arbuste ou herbe pérenne à tiges jeunes
subquadrangulaires, poilues et à tiges âgées arrondies ;
feuilles opposées décussées, subsessiles ou
pétiolées, à limbe décurrent sur le pétiole,
lancéolées, longuement atténuées à la base,
acuminées et aiguës à l'extrémité, à
bord entier, glabres sauf sur les nervures qui sont pubescentes. Inflorescences
en épis axillaires, subterminales, denses, sessiles, simples ;
bractées de l'épi opposées triangulaires, pubescentes.
Bractéoles de chaque fleur, acuminées, spinescentes au sommet,
atténué en coin à la base, pubescentes, pourvues de 5
à 7 nervures. Fleurs ovales, disposées sur plusieurs rangs
longitudinaux, toutes fertiles. Calice bilabié, pubescente ;
lèvre inférieure bifide jusqu'au milieu, la supérieure
trifide jusqu'au milieu. 4 étamines didynames ; filets très
courts, insérées séparément au tube de la corolle
vers son tiers supérieur ; les antérieures à
anthères biloculaires, les postérieures à une loge bien
développée et l'autre rudimentaire. Ovaire bicarpellé,
biloculaire, glabre contenant 2 ovules inégaux dans chaque loges ; style
glabre. Capsule comprimée, contenant une seule graine dans chaque loge,
terminée par un court bec plein.
III.3.14.1. Neuracanthus richardianus (Nees) Boivini
ex. Benoist
Herbe pérenne pouvant atteindre 40 cm de hauteur
à tiges jeunes subquadrangulaires, pubescentes et à tiges
âgées arrondies ; feuilles subsessiles ou pétiolées,
à limbe décurrent sur le pétiole, longues de 4 à
10cm, larges de 14 à 35mm, lancéolées, longuement
atténuées à la base, acuminées et aiguës
à l'extrémité, à bord entier, glabres sauf sur les
nervures qui sont pubescentes. Inflorescences axillaires, subterminales, en
épi, ayant jusqu'à 12cm de longueur sur 12mm, denses ;
bractées de l'épi opposées triangulaires, couvertes de
poils fins, longues de 5mm et large de 1mm. Bractéoles acuminées,
spinescentes aux sommets, atténués à la base, portant de
longs poils blancs épars, pourvues de 5 à 7 nervures, 12 mm de
long et 8 mm de large. Fleurs blanches, ovales, disposées sur plusieurs
rangs longitudinaux, toutes fertiles. Calice bilabié, long de 8mm,
portant de longs poils blancs épars. Corolle blanche, bleue ou
violacée, longue de 6-7mm, bilabiée, son tube long de 5mm.
Lèvres peu distinctes, la supérieure bilobée,
l'inférieure trilobée. 4 étamines didynames
insérées vers le tiers supérieur du tube de la corolle ;
filets courts, long de 2mm ; les antérieures à anthères
biloculaires, les postérieures à une loge bien
développée et l'autre rudimentaire. Ovaire bicarpellé,
biloculaire, glabre contenant 2 ovules inégaux dans chaque loge ; style
glabre. Placentation pariétale. Capsule longue de 6mm et large de 33mm,
comprimée, contenant une seule graine dans chaque loge, terminée
par un court bec.
MBOLA, B., BVM 139, 3 mai 2008, route d'Esomony
après Marohotro, 416 496m/ 182 715m, alt. 410m, Haute vallée de
la Mandrare, forêt sèche à Dalbergia sp et Commiphora sp,
sur bas-fonds avec des affleurements rocheux,
118
Résultats et interprétations
sol gris, alluvionnaire argilo sableux. MBOLA, B.,
FARAMALALA, M. H., LETSARA, R., RAMAVOVOLOLONA et RAZANATSOA, J., BVM 169, 22
août 2009, Manangotry, 442.842m/ 151.318m, alt. 310m, vallée
d'Efaho, forêt dense humide de moyenne altitude à Uacapa sp, sol
gris, alluvionnaire argilo sableux. DESCOING 2721, 27 février 1957,
vallée moyen de la Mandrare, Evasia, CR Ambatoabo. BOSSER J., 417, 16
juillet 1951, Betioky Sud. PHILLIPSON, P.B., RABESIHANAKA, 3124, Janvier 1989,
Itombony, Route d'Anakao, Betioky Sud, 23°53'S ; 44°12'E, Alt. 25m.
PHILLIPSON, P.B., 1806, mai 1987, RS Beza Mahafaly, Parcelle I, 23°39'S ;
44°30'E, alt. 130m.

119
Résultats et interprétations
Figure 34 : Neuracanthus richardianus Benoist

Photo 56 : Fragment de tige et inflorescence de Neuracanthus
richardianus Benoist

Photo 57 : Inflorescence de Neuracanthus richardianus
Benoist
Clé de l'espèce
1a. Bractées disposées en 4 rangées
longitudinales.
2a. Lèvres du calice distinctes presque jusqu'à la
base, les dents étroites aigues.
3a. Etamines à anthères bien
développées Neuracanthus richardianus
III.3.15. Phaulopsis Willdenow
Herbe pérenne plus ou moins rampante, souvent
prostrée, plus ou moins dressée pour un endroit couvert d'arbre
et rampante pour endroit dégagé. Tige jeune, parsemée des
petits poils épars, à 6 angles et plus ou moins arrondies et
glabre au stade adulte. Feuille pétiolée, simple et
entière, faiblement crénelée, souvent inégale dans
une paire. Feuilles parsemées par des petits poils surtout au niveau des
bordures. Limbe lancéolé un peu arrondie à la base et
acuminé au sommet. Inflorescences terminales ou axillaires,
imbriquées, ayant à leur aisselle une petite cyme de 1 à 5
fleurs ; bractéoles absentes. Corolle petite, assez distinctement
bilabiée, tube un peu élargi au sommet, lèvre
supérieure bilobée, lèvre inférieure
trilobée. Calice composé de 5 pièces libres,
120
Résultats et interprétations
inégales, l'un axial, ovale, foliacée,
ressemblant à une bractée, penninerve, et les quatre autres
linéaires uninerves et plus courtes. Corolle glabre à 5 lobes, 2
lobes supérieurs et 3 inférieurs, de couleur blanche, tube
cylindrique, pubescent à l'intérieur. Limbe bilabié,
ovale. Etamines 4, didynames, gamopétales, filet glabre et 2
étamines voisines soudées à la base, anthères
à loge pointue à la base. Ovaire ovoïde, un peu
pubérulent au sommet, stigmate bilobé. Capsule aiguë au
sommet, parfois latérale, fragile. Graines lenticulaires,
marginées, portant des poils hygroscopiques sur les marges.
III.3.15.1. Phaulopsis imbricata ssp. madagascariensis
M. Manktelow
Herbe pérenne plus ou moins rampante, souvent
prostrée, haut de 30-50cm, plus ou moins dressée à un
endroit couvert d'arbre et rampante dans un endroit dégagé. Tige
jeune, pubescente, à 6 angles et plus ou moins arrondies, glabre au
stade adulte. Feuilles opposées décussées, simples et
entières, faiblement crénelées, souvent inégales
dans une paire. Les deux feuilles d'un même noeud ont différentes
dimensions, l'une plus grande que l'autre, portées par un pétiole
long de 20-42mm. Limbe lancéolé à nervure principale et
secondaire très marquée, 26-80mm de long et 12-31mm de large, un
peu arrondi à la base et acuminé au sommet. Feuilles pubescentes
surtout au niveau des bordures. Inflorescences en épi terminales ou
axillaires, souvent dorsi-ventrales, à bractées foliacées,
imbriquées, ayant à leurs aisselles un épi de 1 à 5
fleurs ; bractéoles absentes. La première bractée est
longue de 10mm et 6mm de large, parsemée de poils, la deuxième de
forme deltoïde, de dimension 4mm de long et 1mm de large. Calice
composé de 5 pièces libres, inégales, l'une axiale, ovale,
foliacée, ressemblant à une bractée, penninerve 8-12 sur
5-6mm, et les quatre autres linéaires uninerves et plus courtes et
longues de 2mm. Corolle à cinq pièces soudées à la
base et s'ouvrant en 5 petits lobes, assez distinctement bilabiée, tube
un peu élargi au sommet, lèvre supérieure bilobée,
lèvre inférieure trilobée, de couleur blanche, 6-10mm de
long, tube cylindrique, pubescent à l'intérieur. Etamines 4,
didynames, incluses, insérées vers le milieu du tube de la
corolle ; filets glabres. 2 étamines voisines soudées à la
base, anthères longues d'à peu près 1mm, à loge
pointue à la base. Ovaire ovoïde, un peu pubérulent au
sommet, style long de 3mm, stigmate bilobé. Placentation
pariétale. Capsule longue de 5-8mm, aiguë au sommet, fragiles.
Graines lenticulaire, marginées, 1,5-1,8mm de diamètre, portant
des poils hygroscopiques sur les marges.
Floraison et fructification toute l'année.
MBOLA, B., BVM 123, 28 juillet 2007, Manangotry, sur la
RIP 118, 442.839m/ 151.321m, alt. 510m, Forêt dense humide de moyenne
altitude type forêt de l'est, milieu ouvert, sol noir de forêt.
MBOLA, B., FARAMALALA, M. H.,
121
Résultats et interprétations
LETSARA, R., RAMAVOVOLOLONA et RAZANATSOA, J., BVM 168, 22
août 2009, Manangotry, 442.843m/ 151.325m, alt. 512m, Milieu ouvert d'une
forêt dense humide de moyenne altitude type de l'est, noir de
forêt. MBOLA, B., FARAMALALA, M. H., LETSARA, R., RAMAVOVOLOLONA et
RAZANATSOA, J., BVM 174, 22 août 2009, Manangotry, 442.841m/ 151.324m,
alt. 511m, Milieu ouvert d'une forêt dense humide de moyenne altitude
type de l'est, noir de forêt. KOTOZAFY, RANDRIAMANATENA, RANDRIANASOLO,
198, Aout 1993, PN Ranomafana, parcelle 2, 21°14'S ; 47°22'E, alt.
1200m. HUMBERT, H., 1095, novembre 1924, chaines de Vohibory (Ouest d'Ivohibe).
HUMBERT, H., 6012, octobre 1928, Bassin de Manampanihy, col de Fita. BOSSER,
J., 16243, juin 1962, Antsahadinta.

Photo 58 : Fragment de tige et inflorescence de Phaulopsis
imbricata ssp. madagascariensis Manktelow

Photo 59 : Fleur de Phaulopsis imbricata ssp.
madagascariensis Manktelow
Clé de l'espèce
1a. Limbe ovale. Inflorescence en épi terminale ou
axillaire, dorsi-ventrale, bractées foliacées, imbriquées,
à leurs aisselles une cyme de 1 à 5 fleurs. Bractées
deltoïdes. Calice à 5 pièces libres, inégales.
Corolle en tube cylindrique, pubescent à l'intérieur. Etamines
didynames, incluses, insérées vers le milieu du tube de la
corolle, les filets de 2 étamines voisines sont soudés à
la
base Phaulopsis imbricata ssp.
madagascariensis
122
Résultats et interprétations
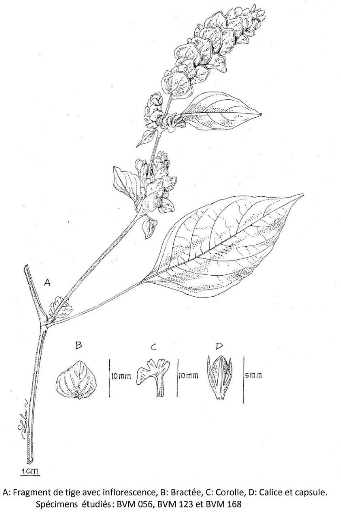
Figure 35 : Phaulopsis imbricata ssp.
madagascariensis M. Manktelow
123
Résultats et interprétations
III.3.16. Ruellia Benoist
Arbustes ou herbe à feuilles opposées.
Inflorescences terminales, en panicules à rameaux opposés. Calice
de 5 sépales subégaux, valvaires, concrescents dans leur moitie
inferieure au moins. Corolle à 5 pétales soudés à
la base formant un tube longuement cylindrique, terminée par 5 lobes
oblongs, les deux supérieurs un peu plus petits. Androcée de 4
étamines didynames exsertes, insérées un peu au-dessous de
la gorge de la corolle, les deux de chaque côté assez longuement
concrescentes par la base de leurs filets; anthères biloculaires. Ovaire
biloculaire contenant 2 ovules dans chaque loge.
III.3.16.1. Ruellia geayi var. tomentosus Benoist
Arbuste pérenne haut de 1,5 à 2m, au
début à tige quadrangulaire et villeux, puis devenu
subcylindrique, pubescent et renflé au-dessus des noeuds. Feuilles
opposées, simples et entières, pétiolées, à
pétioles pubescents. Limbe lancéolé, obtus à la
base, acuminé au sommet, 6 à 8cm de long et 2 à 3cm de
large, pubescent sur les deux faces, de couleur blanche. Face supérieure
des feuilles de couleur verte et face inférieure de couleur beaucoup
plus claire par la pubescence. Inflorescences en cymes, terminales et
subterminales ; les inflorescences inférieures sont situées
à l'aisselle des feuilles supérieures ; bractées et
bractéoles foliacées, linéaires, pubescentes. Calice long
de 4mm, couvert de poils fins, formé de 5 sépales
subégaux, lancéolés linéaires, aigus, concrescents
jusqu'au milieu. Corolle de couleur blanc à blanc cassé, longue
de 35mm, tube allongé, long de 28mm, cylindrique, 5 lobes presque
égaux avec une préfloraison imbriquée. Etamines 4
didynames, soudées au niveau du tube de la corolle, les deux de chaque
côté brièvement concrescents par la base des filets. Disque
petit, régulier. Ovaire uniloculaire et pluriovulée, glabre et
style pourvu de poils. Placentation pariétale. Capsule 20mm de long,
lisse, graines discoïdes, arrondies ou ovales, glabres et lisses, mesurant
4mm de diamètre.
Floraison et fructification : toute l'année.
EBOROKE, S., 811, 10 mai 1994, Piste entre Mahamavo et la
forêt humide, 428.405m/ 151.490m, alt. 1215m, Forêt dense humide,
sol gris de forêt. HUMBERT 13778, Piste entre Ampahiso et Mahamavo,
46°7000' S, 24°7800' E, alt. 1321m. Forêt dense humide de
moyenne altitude, sol gris de forêt. HUMBERT, H., 12971, Environ
d'Esomony, vallée de Manambolo, haut bassin de Mandrare, 24°5200'
S, 46°6100' E, Forêt dense humide de moyenne altitude, sol gris de
forêt. DESCOING 253, Février 1964, Tolagnaro., MORAT, 678,
Forêt entre Betioky-Sud et Toliara. CROAT, T., 31083, 13 février
1975, Andranovory entre pk. 880 et 885.
124
Résultats et interprétations

Figure 36 : Ruellia geayi var. tomentosus
Benoist
125
Résultats et interprétations

Photo 60 : Inflorescence de Ruellia geayi var.
tomentosus Benoist
Clé de l'espèce
1a. Corolle longue de 6-7cm Ruellia geayi
III.3.17. Stenandriopsis Moore
Arbuste ou herbe à rameaux jeunes pubescents, devenant
ensuite glabres. Feuilles opposées très rapprochées
formant un faux verticille. Feuilles pétiolées, dentées,
lancéolées ou subovales, aiguës à la base,
terminées par un mucron au sommet. Inflorescence en épi,
terminale, parfois axillaire. Bractées disposées en quatre
rangées longitudinales, lancéolées, pourvues de dents vers
le sommet, à nervures longitudinales saillantes ; bractéoles
lancéolées linéaires à extrémité
terminée par un mucron, les latérales sont toutes
lancéolées et linéaires à sommet épineux et
à surface glabre. Calice à 5 sépales linéaires,
libres, le postérieur un peu plus grand que les autres. Corolle poilue
en dehors, de couleur violet clair, subcylindrique. Lobes peu inégaux,
ovales, arrondis à l'extrémité. 4 étamines fertiles
insérées vers le quart supérieur du tube de la corolle,
filet court, anthères versatiles, uniloculaires. Ovaire
bicarpellée, biloculaire. Capsule glabre. Graine plus ou moins
triangulaire et rugueuse.
III.3.17.1. Stenandriopsis carduacea Benoist
Herbe pérenne à rameaux jeunes pubescents,
devenant ensuite glabres. Feuilles opposées très
rapprochées formant un faux verticille. Feuilles
pétiolées, lancéolées ou subovales, aigues à
la base, terminées par un mucron au sommet, chaque côté du
bord du limbe est pourvu de 3 à 5 épines, glabre sur la face
supérieure, pubescent sur la face inférieure. Limbe 3-5cm de long
et 2-20mm de large. Inflorescence en épi, terminale, parfois axillaire,
15-25mm de long ; bractées disposées en quatre rangées
longitudinales, lancéolées, pourvues de 3 dents vers le sommet,
dents aigues et la médiane est plus longue, couvertes d'une fine
pubescence éparse, à nervures
Résultats et interprétations
longitudinales saillantes, bractées
lancéolées de dimension 9-10mm de long et 3mm de large.
Bractéoles latérales longues de 6mm et larges de 2mm, toutes
lancéolées et linéaires à sommet épineux et
à surface glabres, extrémité terminée par un
mucron. Calice à 5 sépales linéaires, libres, le
postérieur un peu plus grand que les autres, long de 7mm et large de
1,5mm, Corolle composée de 5 pièces soudées à la
base, tube 10-12mm de long, poilue en dehors, de couleur violet clair, longue
de 16mm, subcylindrique. Lobes peu inégaux, ovales et arrondis à
l'extrémité. 4 étamines fertiles, insérées
vers le quart supérieur du tube de la corolle, filet court,
anthères versatiles, uniloculaires, filaments poilus. Ovaire
bicarpellée, biloculaire. Placentation pariétale. Capsule glabre
longue de 10mm. Graine plus ou moins triangulaire et rugueuse.
Floraison et fructification : toute l'année.
MBOLA, B. et MANAROMBAKA, M., BVM 111, 28 septembre 2006,
Tsimelahy, près de la piscine naturelle, site touristique du parc,
418.259m/ 130.613m, alt. 199m, haut fourré arboré à
Didiereaceae et Euphorbia, milieu plus ou moins ouvert, sol gris avec
affleurement des roches calcaires. LETSARA, R., FARAMALALA, RAKOTONASOLO,
MBOLA, PAUL, LRK 753, décembre 2008, Takamy, Mangatsiaka, Amboasary Sud,
24°57'61"S ; 46°33'52"E, alt. 137m. DECARY, M.R., 10695, janvier
1933, Samborano Tolagnaro (ISOTYPE). LETSARA, R., FARAMALALA, RAKOTONASOLO,
MBOLA, PAUL, LRK 749, décembre 2008, Bafandily, rivière Tarantsy,
24°57'19"S ; 46°37'34"E, alt. 132m.

126
Photo 61 : Fragment de tige et inflorescence de
Stenandriopsis carduacea R Benoist
127
Résultats et interprétations
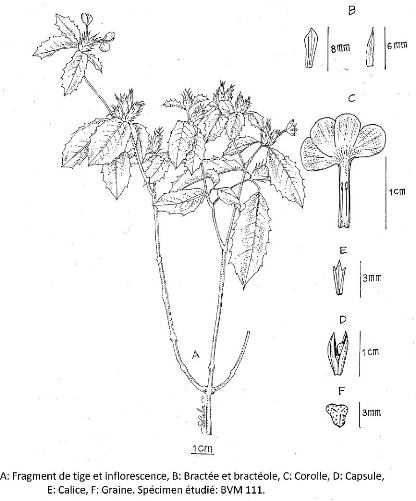
Figure 37 : Stenandriopsis carduacea R Benoist
Clé des espèces
1a. Préfloraison imbriquée.
2a. Sépale imbriqué ; 4 étamines fertiles
à anthère uniloculaire ; pas de cystolithe 3a. Corolle soit
à 5 lobes bien individualisés soit bilabiée.
4a. Corolle bilabiée. . Stenandriospsis
5a.
Feuille entière fortement dentée épineuse sur les
bords.
6a. Sépale libre ; corolle longue de 20mm ; feuille longue
de 3 à 5cm
Stenandriopsis carduacea
128
Résultats et interprétations
III.3.18. Les Acanthaceae de l'herbarium de Tsimbazaza
Conformement à la méthodologie définie
plus haut, les espèces d'Acanthaceae inderminées de l'herbarium
de Tsimbazaza ont été identifiées au cours de la
présente étude. Trente-sept (37) espèces réparties
dans 20 genres ont été ainsi nommées car certaines
étaient mal classées, à l'exemple de 331 spécimens
d'Hypoestes et de 173 de Justicia. Toutefois, quatre
espèces d'Anisotes, de Dicliptera, d'Hypoestes
et de Pseuderanthemum restent indéterminées.
Tableau 12 : Liste des espèces identifiées
|
N°
|
Genres
|
Espèces
|
Nombre d'échantillons
|
|
1
|
Anisotes
|
divaricatus
|
1
|
|
2
|
indet.
|
3
|
|
3
|
subcoreaceus
|
5
|
|
4
|
Barleria
|
humbertii
|
3
|
|
5
|
kitchingii
|
1
|
|
6
|
lupulina
|
3
|
|
7
|
parsvispina
|
3
|
|
8
|
paucidenta
|
6
|
|
9
|
puberula
|
1
|
|
10
|
Blepharis
|
citrina
|
1
|
|
11
|
maculata
|
1
|
|
12
|
Brachystephanus
|
africanus
|
9
|
|
13
|
lyallii
|
4
|
|
14
|
Brillantaisia
|
madagascariensis
|
6
|
|
15
|
Crossandra
|
rupestris
|
1
|
|
16
|
strobilifera
|
1
|
|
17
|
Dicliptera
|
insularis
|
6
|
|
18
|
madagascariensis
|
1
|
|
19
|
ocimoides
|
1
|
|
20
|
indet.
|
14
|
|
21
|
Ecbolium
|
frimbriatum
|
5
|
|
22
|
madagascariense
|
8
|
|
23
|
palmatum
|
7
|
|
24
|
syringifolium
|
3
|
|
25
|
Forsythiopsis
|
vincoides
|
2
|
|
26
|
Hypoestes
|
longituba
|
11
|
|
27
|
strigata
|
2
|
|
28
|
indet.
|
1
|
|
29
|
Lankesteria
|
grandulosa
|
3
|
|
30
|
Lasiocladus
|
anthospermifolius
|
13
|
|
31
|
villosus
|
1
|
|
32
|
Mendoncia
|
cowanii
|
1
|
|
33
|
flagellaris
|
2
|
|
34
|
Neuracanthus
|
richardianus
|
1
|
|
35
|
Forsythiopsis
|
minor
|
1
|
|
36
|
Phaulopsis
|
imbricata
|
9
|
|
37
|
Pseuderanthemum
|
indet.
|
5
|
129
Résultats et interprétations
III.4. RELATIONS ENTRE LES ACANTHACEAE ET LES FACTEURS
ECOLOGIQUES
Les données collectées relatives aux plantes
étudiées ont été analysées pour
déterminer leurs préférences écologiques en
fonction des variations des paramètres écologiques
considérés qui sont les subdivisions climatiques, l'altitude,
l'ouverture du milieu et les types de sol. La similarité entre les sites
de relevés est étudiée avec la méthode de Jaccard
qui permet d'évaluer les affinités entre les sites de
relevé prises deux à deux. L'analyse factorielle des
correspondances a été choisie pour analyser l'association entre
les espèces et entre les paramètres écologique du site de
relevé.
III.4.1. Répartition des espèces selon le
paramètre `subdivision climatique'
Les espèces rencontrées dans les sites de
relevé se répartissent dans les trois subdivisions climatiques
(humide, transition et sèche). Le tableau 12, la carte 5 et la figure 38
présentent les espèces selon leur distribution climatique et les
indices de Jaccard sont montrés par le tableau13.
Tableau 13 : Répartition des espèces selon la
`subdivision climatique' des sites de relevé
|
Genre et espèce
|
Subdivisions climatiques des sites de
relevé
|
|
Humide
|
Transition
|
Sèche
|
|
Asystasia gangetica
|
X
|
|
|
|
Brillantaisia madagascariensis
|
X
|
|
|
|
Dicliptera decaryi
|
X
|
|
|
|
Ecbolium madagascariense
|
X
|
|
|
|
Ecbolium syringifolium
|
X
|
|
X
|
|
Hypoestes longituba
|
X
|
|
|
|
Hypoestes poissonii
|
X
|
|
|
|
Hypoestes sp.1
|
X
|
|
|
|
Hypoestes sp.2
|
X
|
|
|
|
Hypoestes strigata
|
X
|
|
|
|
Justicia sp. 1
|
X
|
|
|
|
Mendoncia delphina
|
X
|
|
|
|
Phaulopsis imbricata
|
X
|
|
|
|
Barleria alluaudii
|
|
X
|
|
|
Barleria kitchingii
|
|
X
|
X
|
|
Blepharis calcitrapa
|
|
X
|
|
|
Blepharis maculata
|
|
X
|
|
|
Barleria her
|
|
X
|
|
|
Forsythiopsis vincoides
|
|
X
|
|
|
Hypoestes sp.3
|
|
X
|
|
|
Lasiocladus anthospermifolius
|
|
X
|
|
|
Neuracanthus richardianus
|
|
X
|
|
|
Ruellia geayi
|
|
X
|
|
|
Stenandriopsis carduacea
|
|
X
|
|
|
Barleria decaryi
|
|
|
X
|
|
Barleria humbertii
|
|
|
X
|
|
Camarotea sp.
|
|
|
X
|
|
Crossandra longipes
|
|
|
X
|
|
Hypoestes decaryana
|
|
|
X
|
|
TOTAL
|
13
|
11
|
7
|
Résultats et interprétations

130
Carte 5 : Répartition des Acanthaceae dans les districts
de Tolagnaro et d'Amboasary-Atsimo
131
Résultats et interprétations

24% Humide
45%
Transition
Sèche
38%
Figure 38 : Répartition des espèces selon le
paramètre `subdivision climatique' des sites de relevé
Tableau 14 : Indice de similitude de Jaccard des sites de
relevé pour le paramètre `subdivision climatique'
|
Site de relevé
|
Humide
|
Transition
|
Sec
|
|
Humide
|
1
|
0
|
0,05
|
|
Transition
|
0
|
1
|
0,05
|
|
Sec
|
0,05
|
0,05
|
1
|
Les Acanthaceae collectées se divisent en trois
groupes. Treize (13) se rencontrent dans les sites de la partie humide, onze
(11) dans la zone de transition contre sept (7) dans la partie sèche.
Deux (2) espèces tolèrent deux types d'habitat, à savoir
Barleria kitchingii en zones de transition et sèche, et
Ecbolium syringifolium en zones humide et sèche. Pour
Barleria kitchingii la localité de collecte a un climat sec.
Par contre pour Ecbolium syringifolium, la zone de collecte est proche
des plans d'eau en zone de transition. Le site de Manangotry, sous couvert
forestier, est le plus riche en Acanthaceae avec 13 espèces ; les icones
superposés dans la carte de répartition des Acanthaceae le
prouvent.
L'indice de similitude de Jaccard est nul entre les sites de
relevé en zones humide et transition ; celui entre les sites de
relevé en zones de transition et sèche sont identiques avec une
valeur de 0,05. Ce sont toutes des très faibles valeurs qui sont
voisines de zéro. Aucune similarité en termes d'espèces
n'est constatée pour ces sites de relevé. Ces derniers ont leurs
propres espèces.
III.4.2. Altitude
L'altitude et le relief ont des influences sur le
développement des espèces d'Acanthaceae. Une altitude est prise
ici comme basse quand elle est entre 0 et 400m, moyenne entre 400m et 800m et
enfin haute au-delà de 800m. Les représentants de la famille des
Acanthaceae ont une préférence et se divisent en trois groupes
par rapport à l'altitude (Tableau 14 et Figure 39).
Résultats et interprétations
Tableau 15 : Répartition des espèces en fonction de
l'altitude des sites de relevé
|
Genre et espèce
|
Altitudes des sites de relevé
|
|
Basse
|
Moyenne
|
Haute
|
|
Asystasia gangetica
|
X
|
X
|
X
|
|
Barleria alluaudii
|
X
|
X
|
X
|
|
Barleria kitchingii
|
X
|
X
|
X
|
|
Phaulopsis imbricata
|
X
|
X
|
X
|
|
Barleria her
|
X
|
X
|
-
|
|
Crossandra longipes
|
X
|
X
|
-
|
|
Ecbolium madagascariense
|
X
|
X
|
-
|
|
Ecbolium syringifolium
|
X
|
X
|
-
|
|
Hypoestes poissonii
|
X
|
X
|
-
|
|
Hypoestes sp.2
|
X
|
X
|
-
|
|
Justicia sp.1
|
X
|
X
|
-
|
|
Neuracanthus richardianus
|
X
|
X
|
-
|
|
Brillantaisia madagascariensis
|
-
|
X
|
X
|
|
Dicliptera decaryi
|
-
|
X
|
X
|
|
Hypoestes longituba
|
-
|
X
|
X
|
|
Mendoncia delphina
|
-
|
X
|
X
|
|
Ruellia geayi
|
-
|
X
|
X
|
|
Barleria humbertii
|
X
|
-
|
-
|
|
Blepharis calcitrapa
|
X
|
-
|
-
|
|
Blepharis maculata
|
X
|
-
|
-
|
|
Camarotea sp.
|
X
|
-
|
-
|
|
Hypoestes sp.3
|
X
|
-
|
-
|
|
Forsythiopsis vincoides
|
X
|
-
|
-
|
|
Lasiocladus anthospermifolius
|
X
|
-
|
-
|
|
Stenandriopsis carduacea
|
X
|
-
|
-
|
|
Hypoestes decaryana
|
-
|
X
|
-
|
|
Barleria decaryi
|
-
|
X
|
-
|
|
Hypoestes sp.1
|
-
|
X
|
-
|
|
Hypoestes strigata
|
-
|
X
|
-
|
|
20
|
21
|
9
|

69%
Basse Moyenne Haute
72%
31%
132
Figure 39 : Représentation graphique de la
répartition des espèces en fonction de l'altitude
Résultats et interprétations
Vingt une (21) espèces préfèrent les
sites en altitudes moyennes, vingt (20) sont visibles en basse altitude et
enfin neuf (9) site en haute altitude. Il est constaté que des
espèces se développent exlusivement dans une classe d'altitude,
huit (8) en basse altitude, quatre (4) en moyenne. D'autres espèces par
contre semblent être indifférentes à l'altitude car quinze
(15) se rencontrent sur deux classes (basse et moyenne) et enfin quatre (4)
espèces sur les trois classes considérées ; ce sont
Asystasia gangetica, Barleria alluaudii, Barleria
kitchingii et Phaulopsis imbricata.
Le tableau 15 présente les indices de Jaccard de chaque
paramètre `altitude'. Tableau 16 : Représentation des indices de
Jaccard pour le paramètre `altitude'
|
Sites de relevé
|
Basse altitude
|
Moyenne altitude
|
Haute altitude
|
|
Basse altitude
|
1
|
0,41
|
0,16
|
|
Moyenne altitude
|
0,41
|
1
|
0,25
|
|
Haute altitude
|
0,16
|
0,25
|
1
|
Les sites de relevé en altitudes basses et moyennes ont
un degré de similarité de 0,41. Ils ont plus d'espèces
communes par rapport aux deux autres sites (haute et basse altitudes).
III.4.3. Ouverture du milieu
La répartition des espèces dans les des sites de
relevé est en relation étroite avec le degré d'ouverture
du milieu (Tableau 16, Figure 40). Treize (13) espèces sur les vint neuf
collectés, soit 31% se développent sur un site semi-fermé.
Les Acanthaceae sont en général des espèces heliophiles.
Le site dont on parle ici est soit une forêt dense humide, soit une
forêt sèche ou un fourré.
45%
31%
28%
Ouvert Semi-fermé Fermé
133
Figure 40 : Répartition des espèces en fonction de
la variable `ouverture du milieu'
134
Résultats et interprétations
Tableau 17 : Répartition des espèces en fonction du
paramètre `ouverture du milieu' des sites de relevé
|
Genre et espèce
|
Degré d'ouverture des sites de
relevé
|
|
Ouvert
|
Semi-fermé
|
Fermé
|
|
Asystasia gangetica
|
X
|
|
|
|
Barleria alluaudii
|
X
|
|
|
|
Barleria decaryi
|
X
|
|
|
|
Barleria her
|
X
|
|
|
|
Barleria humbertii
|
X
|
|
|
|
Crossandra longipes
|
X
|
|
|
|
Ecbolium syringifolium
|
X
|
|
|
|
Phaulopsis imbricata
|
X
|
|
|
|
Barleria kitchingii
|
|
X
|
|
|
Blepharis calcitrapa var decaryi
|
|
X
|
|
|
Blepharis maculata
|
|
X
|
|
|
Brillantaisia madagascariensis
|
|
X
|
|
|
Camarotea sp.
|
|
X
|
|
|
Forsythiopsis vincoides
|
|
X
|
|
|
Hypoestes decaryana
|
|
X
|
|
|
Hypoestes poissonii
|
|
X
|
|
|
Hypoestes sp.1
|
|
X
|
|
|
Hypoestes strigata
|
|
X
|
|
|
Lasiocladus anthospermifolius
|
|
X
|
|
|
Stenandriopsis carduacea
|
|
X
|
|
|
Dicliptera decaryi
|
|
|
X
|
|
Ecbolium madagascariense
|
|
|
X
|
|
Hypoestes longituba
|
|
|
X
|
|
Hypoestes sp.2
|
|
|
X
|
|
Hypoestes sp.3
|
|
|
X
|
|
Justicia sp.1
|
|
|
X
|
|
Mendoncia cowanii
|
|
|
X
|
|
Ruellia geayi
|
|
|
X
|
|
Neuracanthus richardianus
|
X
|
X
|
|
|
9
|
13
|
8
|
Le tableau 17 présente les détails sur les indices
de similarité de Jaccard des sites de relevé pour le
paramètre `ouverture du milieu'.
Tableau 18 : Indices de Jaccard des sites de relevé pour
le paramètre `ouverture du milieu'
|
Sites de relevé
|
Ouvert
|
Semi-fermé
|
Fermé
|
|
Ouvert
|
1,00
|
0,043
|
0,00
|
|
Semi-fermé
|
0,043
|
1,00
|
0,00
|
|
Fermé
|
0,00
|
0,00
|
1,00
|
135
Résultats et interprétations
Le total des espèces évaluées (30
espèces) dépasse le nombre des collectes car Neuracanthus
richardianus se trouve à la fois dans un milieu ouvert ou
semi-fermé ; c'est la seule espèce tolérante de deux types
d'ouverture. L'indice de similarité de Jaccard 0,043 des sites de
relevé pour les paramètres `milieu ouvert' et `milieu
semi-fermé' explique qu'une très faible similitude existe entre
eux par rapport à l'ouverture de milieu. On constate en effet que chaque
site de relevé a ses propres espèces par rapport au
paramètre ouverture du milieu.
III.4.4. Sols
Les Acanthaceae se développent sur différents
types de sol. La figure 41 et le tableau 18 montrent que treize (13)
espèces se rencontrent sur un sol ferrugineux tropicaux souvent
érodés en sol alluvionnaire, sept (7) sur un sol du type carapace
calcaire et enfin quinze (15) sur sol ferralitique peu évolué de
couleur noir sous forêt.
Tableau 19 : Répartition des espèces par rapport au
paramètre `type de sol' des sites de relevé
|
Genres et Espèces
|
Types de sol des sites de relevé
|
|
Sol noir de forêt
|
Sol alluvionnaire
|
Sol calcaire
|
|
Ecbolium syringifolium
|
X
|
X
|
X
|
|
Barleria her
|
|
X
|
X
|
|
Barleria kitchingii
|
|
X
|
X
|
|
Blepharis calcitrapa var decaryi
|
|
X
|
X
|
|
Blepharis maculata
|
|
X
|
X
|
|
Barleria alluaudii
|
|
X
|
|
|
Barleria decaryi
|
|
X
|
|
|
Barleria humbertii
|
|
X
|
|
|
Crossandra longipes
|
|
X
|
|
|
Hypoestes decaryana
|
|
X
|
|
|
Hypoestes sp.2
|
|
X
|
|
|
Lasiocladus anthospermifolius
|
|
X
|
|
|
Stenandriopsis carduacea
|
|
X
|
|
|
Asystasia gangetica
|
X
|
|
|
|
Brillantaisia madagascariensis
|
X
|
|
|
|
Dicliptera decaryi
|
X
|
|
|
|
Ecbolium madagascariense
|
X
|
|
|
|
Hypoestes longituba
|
X
|
|
|
|
Hypoestes poissonii
|
X
|
|
|
|
Hypoestes sp.1
|
X
|
|
|
|
Hypoestes sp.3
|
X
|
|
|
|
Hypoestes strigata
|
X
|
|
|
|
Justicia sp.1
|
X
|
|
|
|
Mendoncia delphina
|
X
|
|
|
|
Phaulospis imbricata
|
X
|
|
|
|
Ruellia geayi
|
X
|
|
|
|
Forsythiopsis vincoides
|
X
|
|
|
|
Camarotea sp.
|
|
|
X
|
|
Neuracanthus richardianus
|
|
|
X
|
|
TOTAL
|
15
|
13
|
7
|
Résultats et interprétations
52%
45%
24%
Sol noir de forêt
Sol alluvionnaire
Sol calcaire
136
Figure 41 : Répartition des espèces en fonction du
type de sol
Le tableau 19 donne les détails des indices de Jaccard des
sites de relevé pour le paramètre `type de sol'.
Tableau 20 : Indice de Jaccard des sites de relevé pour le
paramètre `type de sol'
|
Sites de relevé
|
Sol alluvionnaire
|
Sol noir de forêt
|
Sol calcaire
|
|
Sol alluvionnaire
|
1,00
|
0,04
|
0,25
|
|
Sol noir de forêt
|
0,04
|
1,00
|
0,06
|
|
Sol calcaire
|
0,25
|
0,06
|
1,00
|
La répartition des espèces d'Acanthaceae est
conditionnée par le type de sol. Seule Ecbolium syringifolium
se rencontre sur trois types de sol. Quatre (4) espèces,
Barleria kitchingi, B. her, Blepharis maculata et B. calcitrapa
poussent sur deux types de sol (alluvionnaire et calcaire). Huit (8) sont
propres au sol alluvionnaire, quatorze (14) sur sol noir de forêt et deux
(2) sur du sol calcaire. Donc, la majorité des espèces de la zone
d'étude se développent sur un sol noir de forêt avec 52%
des échantillons étudiés.
Les sites de relevé pour les paramètres `sol
alluvionnaire' et `sol calcaire' ont un indice de similarité de Jaccard
de 0,25. Les indices présentés sur le tableau 17 sont très
faibles ; donc il y a peu de similitude entre ces deux sites de relevé
pour le paramètre types de sol en termes d'espèces d'Acanthaceae.
Chaque site de relevé, ayant des types de sol différents, a ses
propres espèces.
III.4.5. Relations entre les espèces et les
paramètres écologiques des sites de relevé
Les espèces se répartissent dans la nature selon
leurs préférences écologiques. Les interprétations
des données obtenues au cours des travaux de terrain et de laboratoire
ont permis de dire que les Acanthaceae se rencontrent dans les milieux ouverts
mais aussi dans les endroits secs ou
137
Résultats et interprétations
humides, altitude et latitude confondues et même en
milieu aquatique. La figure 42 suivante donne la représentation
géométrique des analyses des correspondances des espèces
en fonction des paramètres écologiques des sites de
relévé.
Trois groupes d'espèces sont formés suivant
leurs affinités. Le premier est représenté par
Brillantaisia madagascariensis, Ruellia geayi, Forsythiopsis vincoides,
Dicliptera decaryi, Asystasia gangetica, Justicia sp.1 et Hypoestes
sp.2 qui sont proches ou manifestent une attraction par rapport aux
paramètres écologiques `climat humide', `moyenne altitude',
`haute altitude', `milieu fermé' et `sol noir'. Cela correspond aux
sites qui se trouvent dans la partie humide de la zone d'étude.
Le deuxième regroupe Blepharis maculata, Blepharis
calcitrapa, Camarotea sp., Ecbolium madagascariensis, Hypoestes
strigata, Hypoestes decaryi, Phaulopsis imbricata, Mendoncia delphina et
Stenandriopsis carduacea, qui sont quant à eux proches par
rapport aux paramètres écologiques `partie sèche', `milieu
ouvert' et `sol alluvionnaire'.
Le troisième groupe est composé par
Crossandra longipes, Barleria decaryi, Barleria alluaudii et
Barleria her qui sont proches par rapport aux paramètres
écologiques `partie transition', `milieu semi-fermé' et `sol
calcaire'.
Quelques espèces comme Ecbolium syringifolium,
Hypoestes sp.1, Hypoestes sp.2, sont beaucoup plus libres ou
indépendantes.
Aucune attraction n'est constatée entre les
paramètres `milieu ouvert', `milieu semi-fermé' et `milieu
fermé'. Les sites sur sol alluvionnaire et sur calcaire ont des
attractions ; ils sont rapprochés. Par contre le site sur sol noir est
répulsé par ces deux derniers.
Résultats et interprétations

138
Figure 42 : Représentations géométriques des
espèces et des paramètres écologiques des sites de
relevé.
DISCUSSIONS ET RECOMMANDATIONS
|
139
Discussions et recommandations
Deux points seront discutés dans cette partie. Le
premier se rapporte à la méthodologie de classification
utilisée et le second sur la rareté des recherches sur les
Acanthaceae à Madagascar.
IV.1. DISCUSSIONS SUR LA METHODOLOGIE
Selon Benoist (1967), la famille des Acanthaceae comporte
trois sous familles, les Thunbergioidae incluant Mendoncia, les
Tubiflora et les Acanthoideae. En 2008, Mabberley (2008) a
amélioré la classification en considérant quatre (4) sous
familles qui sont les Nelsonoideae incluant Tubiflora, les
Thunbergioideae, les Acanthoideae et les Mendoncioideae. La présente
étude a simplifié l'approche en considérant le type
biologique `liane' comme premier critère de distinction morphologique,
critère qui permet tout de suite de séparer les
échantillons étudiés en deux. L'espèce Mendocia
delphina constitue un groupe à part car c'est la seule liane du
site de relevé. Ensuite, les autres types biologiques ont
été utilisés pour séparer Ruellia qui est
un nanophanérophyte, du reste qui sont des hémicryptophytes. La
suite de la détermination a été faite surtout à
partir des fleurs.
Cette démarche simplifiée devrait être
appliquée à tous les taxons de Madagascar dans la finalisation
des autres tomes de la clé de détermination des Acanthaceae de
Madagascar incluant des sous familles et/ou des tribus. Aussi, d'autres
investigations sont nécessaires pour couvrir l'ensemble de Madagascar.
En effet, les analyses des étiquettes d'herbiers existantes montrent que
les lieux de collectes visités jusqu'à ce jour ne concernent que
les aires protégées et les voies d'accès. Si l'on veut
terminer les deux tomes de la clé de détermination
préconisée par Benoist en 1967, il faudrait étendre les
collectes aux autres régions de Madagascar. En plus, pour diverses
causes d'accessibilité, plusieurs localités comportant
sûrement des représentants de cette famille n'ont pas pu
être visitées.
La méthodologie adoptée au cours de ce travail
de recherche ne concerne que l'étude morphologique des espèces
récoltées. Ceci a permis de proposer la clé de
détermination donnée en résultat qui considère
surtout des caractères biologiques visibles directement sur le terrain
et renforcés en laboratoire avec du matériel plus performant. On
peut recommander ici que des études plus poussées
phylogénétiques et polliniques soient à faire pour avoir
le maximum de données de description taxonomique mais l'étude
morphologique devrait toujours constituer une base fondamentale de la
systématique. En effet, actuellement, des études plus
poussées relatives à la phylogénie moléculaire et
aux études polliniques permettraient une meilleure distinction
systématique entre les différentes espèces d'Acanthaceae.
Avec la première méthode, à partir des
140
Discussions et recommandations
spécimens des herbaria de Genève, de Paris, de
Kew et de Missouri, Callmander et al. (2014) ont pu par exemple
améliorer la classification du genre Eusiphon qui est en fait
un Ruellia.
IV.2. RARETE DE LA RECHERCHE SUR LES ACANTHACEAE A
MADAGASCAR
Plusieurs constats sont faits à travers l'étude
des spécimens déposés dans l'herbarium de Tsimbazaza, site
principal de recherche en laboratoire au cours de cette thèse. Plusieurs
dépôts y existent avec environ 1000 spécimens
d'Acanthaceae. Ce sont le fruit des collectes de plusieurs auteurs depuis
plusieurs années, sans que des recherches spécifiques sur les
Acanthaceae aient été menées.
Plusieurs chercheurs s'intéressent aux Acanthaceae mais
peu de résultats concernent directement Madagascar. En effet, des
études relatives aux Acanthaceae ont été faites avec
quelques spécimens Malagasy sans qu'aucune espèce du site de
relevé ne fût considérée :
- Furness, C.A., et Grant, M.C., 1996. Pollen morphology of
some Ruellia species (Acanthaceae) from Africa and Madagascar,
Grana, 35 (4): 231-239) ;
- Daniel, T.F. 2006. Chromosome numbers of miscellaneous
Malagasy Acanthaceae. Brittonia, The New York Botanical Garden Press,
Bronx, 58 (4) : 291-300 ;
- Daniel, T. F., Mbola, V. A. B., Almeda, F. et Phillipson,
P., 2007. Anisotes in Madagascar. Proceedings of the California
Academy of Sciences, fourth series, Vol. 58: 121-131 ;
- Daniel, T. F., Letsara, R., Santiago, M-B., 2013. Four new
species of Anisotes (Acanthaceae) from Madagascar. A journal for
Botanical nomenclature, Missouri Botanical Garden. 22 (4) : 396-408.
Initiées par CAS, ces deux dernières
études ont été faites dans le cadre de la révision
de la famille des Acanthaceae Malagasy pour faire avancer l'élaboration
finale de sa clé de determination.
IV.3. APPLICATIONS
La richesse de Madagascar en Acanthaceae est inestimable,
seulement beaucoup d'entre elles sont méconnues par la population et une
mauvaise gestion de ces ressources est constatée.
Beaucoup d'espèces d'Acanthaceae sont horticoles mais
très peu sont visibles dans les jardins publics ou botaniques de
Madagascar que ce soit pour les autochtones ou les introduites. Seules
Crossandra longipes et Barleria lupiluna, espèces
endémiques Malagasy existent dans quelques jardins publics à
Toliara et à Antananarivo. Seules Acanthus albus A. mollis,
Aphelandra sp. Dyschoriste sp., Elytraria
sp. et Hemigraphis sp. ont
été introduites ; elles sont surtout visibles à
Antananarivo. Pourtant, les Acanthaceae Malagasy sont reconnues ornementales
à cause de leurs
141
Discussions et recommandations
belles fleurs, à l'exemple de Barleria lupiluna,
qui fut exportée en Guyane (Crener et Hoff, 1995) et dans diverses
régions tropicales.
On constate aussi qu'aucune collection vivante d'Acanthaceae
n'existe à Madagascar. L'Université de Toliara qui possède
déjà un Laboratoire de phytobiologie devrait étoffer son
herbarium avec les Acanthaceae de la région et mettre en place une
pépinière et un jardin botanique pour alimenter les jardins
publics de la ville. De même, l'herbarium du Département de
Biologie et Ecologie Végétales de l'Université
d'Antananarivo devrait être mis à jour pour pouvoir donner des
outils de formation adéquats en Acanthaceae.
Si toutes ces propositions se réalisent, des programmes
de sensibilisation pourraient être développés auprès
des horticulteurs professionnels afin de pouvoir conserver ces espèces
et de les gérer durablement.
142
Conclusion
CONCLUSION
La famille des Acanthaceae, principal objet de la
présente thèse, possède environ 4000 espèces
reparties sur 221 genres dans le monde tropical. Cette thèse a pour
objectif de compléter les informations sur cette famille afin de pouvoir
plus tard finaliser sa clé de détermination pour Madagascar.
La zone d'étude est le Parc National d'Andohahela et
ses environs, dans les districts de Tolagnaro et d'Amboasary-Atsimo. Cette zone
a été choisie parce qu'elle est subdivisée en trois
parties selon le paramètre `subdivision climatique', parties qui sont
supposées comporter différentes espèces d'Acanthaceae. Ces
domaines phytogéographiques correspondent en effet à trois
conditions climatiques délimitées à partir de l'existence
d'une faille pluviométrique au niveau de Ranopiso-Bevilany qui
définit par conséquent trois habitats distincts de ces
espèces à savoir la forêt humide de l'Est de la Parcelle 1
du Parc National d'Andohahela, une zone de transition à Dypsis
decaryi au niveau de Ranopiso-Bevilany et les formations sèches de
l'Ouest (forêt, fourré et savane).
Les méthodes utilisées dans cette thèse
sont surtout les inventaires de terrain des espèces d'Acanthaceae et
leur étude morphologique en laboratoire qui sont les herbaria du Parc
Botanique et Zoologique de Tsimbazaza, Antananarivo et de California Academy of
Sciences de San Francisco. Les inventaires ont consisté à la
collecte des spécimens et à l'étude de leur habitat. Pour
cela, les paramètres écologiques relevés sont le facteur
lumière traduit par l'ouverture du milieu, l'altitude et le type de
sol.
Vingt-neuf (29) espèces d'Acanthaceae,
regroupées dans 17 genres, ont été collectés au
cours de la présente étude. Leurs études morphologiques
ont montré que ces Acanthaceae sont surtout des
`hémicryptophytes' sauf Ruellia geay qui est un
`nanophanérophyte', sont pour la plupart inermes (72%) et leur
inflorescence est surtout de type épi.
Une clé de détermination partielle a
été faite en considérant ces espèces. Cinq (5)
nouvelles espèces regroupées dans trois (3) genres ont
été identifiées, à savoir Camarotea,
Justicia et Hypoestes. Pour ce faire, le type biologique a
été utilisé car c'est le premier caractère visible
sur terrain et les conditions du paysage permettaient de le voir facilement.
L'analyse des spécimens de l'herbarium du TAN, principal
bénéficiaire des résultats produits ici permettra
d'améliorer l'état des données sur les Acanthaceae afin
d'atteindre l'objectif principal défini dans le cadre de cette
thèse, à savoir la finalisation des différents tomes de
cette clé.
143
Conclusion
Treize (13) espèces sont visibles dans la partie
humide, onze (11) au niveau de la zone de transition et sept (7) dans la partie
sèche. Quelques espèces se retrouvent dans deux types d'habitat ;
une seule espèce (1) est vue dans la zone humide et dans la partie
sèche (Barleria kicthingi) et une seule espèce se
rencontre dans les zones de transition et sèche (Ecbolium
syringifolium). La première hypothèse énoncée
dans le cadre de cette étude qui stipule que `les Acanthaceae sont des
bons indicateurs climatiques' est vérifiée car chaque subdivision
bioclimatique a ses propres espèces sauf pour les cas des deux
espèces mentionnées ci-dessus. Vingt une (21) espèces se
rencontrent en moyenne altitude, neuf (9) en haute, et vingt (20) en basse
altitude. Quatre (4) espèces se développent exclusivement en
moyenne altitude. Ce sont Hypoestes decaryana, H.
strigata, H. sp.1 et Barleria decaryi. Trois
espèces se trouvent dans quatre (4) types d'altitude différents,
à savoir Asystasia gangetica, Barleria alluaudii,
B. kitchingii et Phaulopsis imbricata.
Contrairement aux données obtenues à partir des
analyses des étiquettes des herbiers du TAN, les Acanthaceae ne se
retrouvent pas seulement en milieu ouvert. En effet, seules huit (8) sur les
vingt-neuf (29) espèces étudiées, soit 27% y poussent. Le
reste se développe respectivement en milieu fermé avec neuf (9)
espèces et treize (13) en milieu semi-fermé. La deuxième
hypothèse qui stipule que `les espèces de la famille des
Acanthaceae à Madagascar se développent en milieu ouvert',
énoncée dans le cadre de cette étude n'est pas
vérifiée car chaque type d'ouverture de milieu a ses propres
espèces.
Les différents types de sol du site de relevé
ont leurs propres Acanthaceae sauf pour quatre espèces ; Ecbolium
syringifolium est indifférente et pousse sur les trois types de
sols ; Barleria her et B. kitchingii, Blepharis calcitrapa et
B. maculata sont visibles sur deux types de sol, alluvionnaire et
calcaire. La repartition globale montre bien une exclusivité car douze
(12) espèces se trouvent sur sol alluvionnaire, deux (2) sur calcaire et
quatorze (14) sur sol noir de forêt. Donc, les Acanthaceae à
quelques exceptions près sont exigeants par rapport aux conditions
édaphiques.
Ces constats sont seulement valables dans le cas de la
présente étude et les situations dans les autres régions
de Madagascar doivent être étudiées à partir de plus
vastes prospections. Les détails y relatives sont données plus
haut en récommandations.
Pouvoir finaliser la clé entière des Acanthaceae
Malagasy, on peut suggérer en persperctive, les thèmes de
recherches suivants :
a. Réviser les Acanthaceae malagasy pour finaliser
leurs classifications et arriver au niveau `espèce'. A titre d'exemple,
les Justicieae possèdent plusieurs espèces qui se
répartissent dans tout Madagascar, en particulier les cinq (5) nouvelles
espèces (Justisia sp, Camarotea
144
Conclusion
sp, Hypoestes sp.1, Hypoestes sp.2,
Hypoestes sp.3) découvertes au cours de la présente
étude ;
b. Revoir la liste des herbiers de la famille des Acanthaceae
dans l'herbarium de Tsimbazaza et ceux à travers le monde ;
c. Mener une prospection complète de toutes les
régions de Madagascar pour actualiser les données existantes. En
effet, dans le cadre de cette étude, plusieurs espèces n'ont pas
été rencontrées suite à la transformation du site
en terrain de culture ou même en zone d'habitation. C'est le cas de
Crossandra vestita, Ruellia monathos et R.
nummularia.
d. Mener des études sur les pollens de toutes les
espèces déjà déposées à l'herbarium
de Tsimbazaza afin de les comparer pour compléter les informations pour
la classification ;
e. Mener une étude pour déterminer la
biogéographie des Acanthaceae Malagasy car des espèces
d'Acanthaceae sont endémiques locales et d'autres sont à large
distribution ;
f. Continuer la determination des 500 spécimens
d'Acanthaceae de l'herbarium de Tsimbazaza car seules 163 ont été
étudiées et déterminées dans le cadre de cette
étude ;
g. Mener des études phylogéniques sur les
espèces car peu de données sur les ADN sont disponibles pour les
Acanthaceae Malagasy.
REFERENCES BIBLIOGRAPHIQUES
|
145
Références bibliographiques
REFERENCES BIBLIOGRAPHIQUES
1. ANDRIARIMALALA, T., 2014. Analyses floristiques,
structurales, phénologiques et distribution spatiale des habitats de
Microcebus spp. dans le Parc National d'Andohahela. Thèse de
Doctorat. Faculté des Sciences, Département de Biologie et
Ecologie Végétales, Université d'Antananarivo. 113p.
2. ANGAP, 2001. Plan de gestion du réseau des aires
protégées. 121p.
3. ANGAP, 2004. Plan de gestion de conservation du Parc
National d'Andohahela. Document de travail. 75p.
4. ANISA, S. AL-HAKIMI, HAJA, MAIDEEN, et A. LATIFF., 2015.
Pollen and Seed Morphology of Rhinacanthus Nees and Hypoestes
Sol. ex R. Br. (Acanthaceae) of Yemen. Sains Malaysiana 44(1): 7
- 15.
5. APG III. 2009. An update of the Angiosperm Phylogeny Group
classification for the orders and families of flowering plants: APG III.
Botanical Journal of the Linnean Society 161: 105 - 121.
6. BADEN, C., 1981. New taxa in Anisotes
(Acanthaceae). Nord. J. Bot. n°1 : 143-153.
7. BAZOT, G., 1976. Contribution à l'étude des
formations métamorphiques précambriennes du Sud-Est de
Madagascar. Thèse de Doctorat, Mention `Sciences'. Université de
Clermont Ferrand. 33p.
8. BENNET, J. R., and SCOTLAND R.W., 2003. A revision of
Strobilanthes (Acanthaceae) in Java. Kew Bulletin 58 : 1 - 82 .
9. BENOIST, R., 1920. Nouvelle espèce d'Hypoestes
de Madagascar. Bulletin du Muséum National d'Histoire
Naturelle. Paris : 186 - 190.
10. BENOIST, R., 1929. Une nouvelle espèce du genre
Hypoestes. Bulletin du Muséum National d'Histoire
Naturelle. Paris. Série II : 222 - 223.
11. BENOIST, R., 1947. Nouvelles Acanthacées de
Madagascar. Notul. Syst. Paris. Vol.13:1 - 13.
12. BENOIST, R., 1949. Les Dicliptera Malgaches.
Mémoire de l'Institut Scientifique de Madagascar. Série
B. Tome II. Fascicule 2 : 213 - 217.
13. BENOIST, R., 1967. Flore de Madagascar et des Comores
(Plantes vasculaires), Acanthaceae. 82e famille, Tome I.
Muséum National d'Histoire Naturelle. Laboratoire de
Phanérogamie. 230p.
14. BIDGOOD, S. and BRUMMITT, R. K., 1998. A revision of the
genus Neuracanthus (Acanthaceae). Kew Bulletin, 53 : 1 -
76.
146
Références bibliographiques
15. BOSSER, J., 1969. Graminées des pâturages et
des cultures à Madagascar. Mémoire ORSTOM, n°35.
Paris. 440p.
16. BREMEKAMP, C. E. B., 1938. Notes on the Acanthaceae of
Surinam. Rec. Trav. Bot. Neerl 35: 130 - 174.
17. BREMEKAMP, C. E. B., 1957. New Scrophulariaceae and
Acanthaceae from New Guinea. Nova Guinea Ser. 2 8: 129 - 155.
18. BRIDSON, D., and FORMAN, L., 1992. The herbarian handbok.
Revised edition. Whistable Litho Printers, Kew. 310p.
19. CALLMANDER, M. W., TRIPP, E.A. and PHILLIPSON, P.B.,
2014. A new name in Ruellia L. (Acanthaceae) for Madagascar. Candollea
69 : 81 - 83.
20. CAPURON, R., 1957. Introduction à l'étude
de la flore forestière de Madagascar. 120p. + annexes.
21. CARINE, M. A., and SCOTLAND, R. W., 1998. Pollen
morphology of Strobilanthes Blume (Acanthaceae) from Southern India and Sri
Lanka. Review of Palaeobotany and Palynology 103: 143 - 165.
22. CHAPERON, P., DANLOUX, J., and FERRY, L., 1993. Fleuves
et rivières de Madagascar. IRD, DMH et CNRE. 874p.
23. CLARKE, C. B., 1900. Acanthaceae. Tiselton-Dyer, W.
T., (Ed.), Flora of tropical Africa. London Vol. 5 : 1 - 262.
24. CORNET, A., 1974. Essai de cartographie bioclimatique
à Madagascar, Notice explicative, ORSTOM, Paris, n°55. 27p.
25. CRENER, R. et HOFF, M., 1995. Inventaire taxonomique des
plantes de la Guyane Française. 1ère Partie :
Acanthaceae à Bixaceae. 248p.
26. DANIEL, T.F., 1997. The Acanthaceae of California and the
peninsula of baja California. Proceedings of the California Academy of
Sciences. Sciences. Vol. 49 : 309-372.
27. DANIEL, T.F., 2006. Chromosome numbers of miscellaneous
Malagasy Acanthaceae. Brittonia. The New York Botanical Garden Press,
Bronx, Vol. 58, n°4 : 291-300.
28. DANIEL, T. F., MBOLA, V.A.B., ALMEDA, F., and PHILLIPSON,
P.B., 2007. Anisotes (Acanthaceae) in Madagascar. Proc. Calif.
Acad. Sci. 58 : 121 - 131.
29. DANIEL, T. F., McDADE, L. A., MANKTELOW, M., and KIEL, C.
A., 2008a. The «Tetramerium lineage» (Acanthaceae; Acanthoideae,
Justicieae): Delimitation and intra-lineage relationships based on cp and nrITS
sequence data. Systematic Botany 33 : 416 - 436.
147
Références bibliographiques
30.DANIEL, T.F., McDADE, L.A., MANKTELOW, M., and KIEL, C.A.,
2008b. Toward a comprehensive understanding of phylogenetic relationships among
lineages of
Acanthaceae s.l. (Lamiales). American Journal of
Botany. Vol. 95 (9) : 1136 - 1152.
31. DANIEL, T.F., and FIGUEIREDO, E., 2009. The California
Academy of Sciences Gulf of Guinea Expeditions (2001, 2006, 2008) VII.
Acanthaceae of Saõ Tomé and Principe. Proc. Calif. Acad. Sci.
60 : 623- 674.
32. DANIEL, T. F., LETSARA, R., and SANTIAGO, M-B., 2013.
Four new species of Anisotes (Acanthaceae) from Madagascar. A
journal for Botanical nomenclature, Missouri Botanical Garden. Vol. 22,
n°4 : 396 - 408.
33. De la ROCHE, H., 1958. Etude géologique de
l'extrême Sud-Est de Madagascar. Bull. Soc. Géol. France,
6e série, n°4 : 259 - 302.
34. EBOROKE, S., 1994. Approche phytosociologique de la
parcelle III de la Réserve Naturelle Intégrale d'Andohahela,
Département des Sciences Naturelle, Université de Tuléar.
(Rapport de stage). 81p. + annexe.
35. EMBERGER, L., GODRON, M., DAGET, P., LONG, G., LE FLOC'H,
E., POISSONET, J., SAUVAGE C., et WACQUANT, J.P., 1983. Code pour le
relevé méthodique de la végétation et du
milieu, Edition du Centre National de la Recherche Scientifique.
Paris. 292 p.
36. EZCURRA, C., 1993. Systematics of Ruellia
(Acanthaceae) in Southern South America. Ann. Missouri Bot. Gard. 80 : 787
- 845.
37. FURNESS, C.A. and GRANT, M.C., 1996. Pollen morphology of
some Ruellia species (Acanthaceae) from Africa and Madagascar,
Grana, Vol. 35, n°4 : 231 - 239.
38. GAUSSEN, H., 1955. Détermination des climats par
la méthode des courbes ombrothermiques. C.R./Acad. Sc., Vol. 2.
647p.
39. GOODMAN, S.M., 2008. Paysages naturels et
biodiversité de Madagascar. Muséum National d'Histoire
Naturelle. Paris. 694p.
40. GOODMAN, S.M., BENSTEAD, J.P., 2005. Uptadet estimate of
biotic diversity and endemism for Madagascar. Oryx. Vol. 39, n°1
: 73 - 77.
41. GUILLANI, 2007. La géologie et la
minéralogie à Madagascar avant la colonisation. In
Parcours de recherche à Madagascar. IRD. Paris : 297 -
319.
42. GUILLAUMET, J.L., 1983. Forêts et fourrés de
montagne à Madagascar. Candollea, n°38 Hennig : 481 -
502.
43. HUMBERT, H., 1955. Les territoires
phytogéographiques de Madagascar. Leur cartographie. Colloque sur les
régions écologiques du globe. Ann. Biol. Paris, Fasc.
5-6 : 439 - 448.
148
Références bibliographiques
44. HUMBERT, H. et COURS-DARNE, G., 1965. Notice de la carte
de Madagascar, Extrait des travaux de la section scientifique et technique de
l'Institut Français de Pondichéry. Hors-série n°6.
162p.
45. JACCARD, P., 1901. Bulletin de la Société
Vaudoise des Sciences Naturelles, 37 : 241 - 272.
46. KOECHLIN, J., GUILLAUMET,
J-L. et MORAT, P., 1974. Flore et
végétation de Madagascar. J. Cramer. (Vaduz). 626p.
47. KOEKEMOER, M., STEYN, H.M. and BESTER, S.P., 2014. Guide
to plant families of southern Africa. Strelitzia 31. South African National
Biodiversity Institute, Pretoria.
48. KULUS, E., 2001. L'impact des feux dans la zone d'Esomony
(Ouest du Parc National d'Andohahela). Note de fin de stage, Université
Louis Pasteur, Strasbourg France.42p.
49. LEONARD, E.C., 1951. The Acanthaceae of Colombia I.
Contributions of the United States National Herbarium 31 : 1 - 117.
50. MABBERLEY, D.J., 2008. Mabberley's plant-book. A portable
dictionary of plants, their classification and uses. 3rd. ed. Cambridge :
Cambridge University Press. 1021p.
51. MAEP, 2003. Monographie de la Région Anosy.
187p.
52. MAGNAGHI, B.E., and DANIEL, T.F., 2014. Mendoncia
(Acanthaceae) from Madagascar, in Proceeding of the California Academy
of Sciences, California Academy of Sciences. Allen Press Inc, Lawrence
Kansas, USA. Vol. 23, n°2 : 187 - 196.
53. MAJMUNDAR, H., 1961. Contribution à l'étude
minéralogique et géochimique des pyroxènes et de micas
dans les pyroxénites à phlogopite et dans les charnockites du
Sud-Est de Madagascar. Thèse de Nancy. Service Géologique de
Tananarive. 117p.
54. McDADE, L, DANIEL, T.F., MASTA, S.E. and RILEY, K.M.,
2000. Phylogenetic relationships within the tribe Justicieae (Acanthaceae) :
evidence from molecular sequences, morphology, and cytology. Annals of the
Missouri Botanical Garden. n°87 : 435 - 458.
55. McDADE, L.A., MASTA, S.E., MOODY, M. L., and WATERS, E.,
2000. Phylogenetic relationships among Acanthaceae: evidence from two genomes.
Syst. Bot. 25: 106 - 121.
56.McDADE, L.A., DANIEL, T.F., KIEL, C.A., and VOLLESEN, K.,
2005. Phylogenetic Relationships among Acantheae (Acanthaceae) : Major Lineages
Present Contrasting Patterns of Molecular Evolution and Morphological
Differentiation, The American Society of Plant Taxonomists, Systematic
Botany, 30 (4) : 834 - 862.
57. McDADE, A.L., DANIEL, T.F. and CARRIE, A. K., 2008. Toward
a comprehensive understanding of phylogenetic relationship among lineages of
Acanthaceae S.L. (Lamiales). American Journal of Botany. Vol. 95,
n°9 : 1136 - 1152.
149
Références bibliographiques
58. MOAT, J., et SMITH, P., 2007. Atlas de la
végétation de Madagascar. Royal Botanical Garden, Kew.
UK. 124p.
59. NOIZET, G., 1959. Les formations de métamorphisme
d'intensité élevée de l'Androy Mandrareenne (Sud-Est de
Madagascar). Thèse de Doctorat, Mention `Sciences'. Université de
Nancy. France. 266p.
60. NURUL AINI, C. A. C., T. NORAINI, A. LATIFF., A. J.
AMIRUL-AIMAN, and NOOR-SYAHEERA, M.R., 2017. Pollen morphology of selected
species of Ruellioideae (Acanthaceae) in Peninsular Malaysia. Malayan
Nature Journal, 69(2) : 39 - 51.
61. PATIL, A.M., and PATIL, D.A., 2011. Occurrence and
significance of cystoliths in Acanthaceae. Curr. Bot. 2(4) : 1-5.
62. PAULIAN, R., BLANC, Ch., GUILLAUMET, J-L., BETSCH, J-M.,
GRIVAUD, P., et PEYRIERAS, A., 1973. Etude des écosystèmes
montagnards dans la région Malgache. Les chaînes Anosyennes.
Géomorphologie, climatologie et groupement végétaux.
Bul. Mus. Nat. Hist. Nat. Série 3, n°118. 40p.
63. PBZT, 2017. Liste des spécimens
déposés à l'herbarium du Parc Botanique et Zoologique de
Tsimbazaza. Département Flore (Document inédit). 47p.
64. PERVEEN, A., & QAISER, M., 2010. Pollen flora of
Pakistan - LXVII : Acanthaceae. Pakistan Journal Bototany Special
Issues 42 : 175 - 191.
65. PETIT, M., 1998. Présentation physique de la
Grande île Madagascar. FTM. 192p + planches.
66. PHILLIPSON, P.B., SCHATZ, G.E., LOWRY, II P.P., and
LABAT, J-N., 2006. A catalogue of the vascular plant of Madagascar. In
Goodman S.M., Beentje H.J. (Edis). Taxonomy and ecology of African plant.
Their conservation and sustainable use. Kew Royal Botanic Garden : 613
- 627.
67. RAJERIARISON, C., et FARAMALALA, M. H., 1999.
Nomenclature des formations végétales de Madagascar. Edition
ANGAP, 68p.
68. RAKOTONDRASOA, E. F., 2011. Etude comparative de la
diversité des reptiles suivants les différents types d'habitats
(forêt sèche, forêt galerie, forêt de transition) dans
le Parc National d'Andohahela (Madagascar). Mémoire de DESS, en Sciences
de l'Environnement. Faculté des Sciences. Université
d'Antananarivo. 91p.
69. RATSIVALAKA-RANDRIAMANGA, S., 1987. Recherche sur le
climat de Fort-Dauphin (ex-Tolagnaro) (Extrême Sud Est de
Madagascar). Mad. Rev. Géo. n°46 : 47 - 67.
70. RAUNKIAER, C., 1904. Types biologiques pour la
géographie botanique. Bull. Acad. Roy. Sc. Etablissement des
Lettres. Danemark, Copenhague : 347 - 437.
150
Références bibliographiques
71. SCHATZ, G.E., 2001. Flore générique des
arbres de Madagascar. Missouri Botanical Garden, Saint Louis; Royal
Botanical Garden, Kew. 503p.
72. SCOTLAND, R.W., and VOLLESEN, K., 2000. Classification of
Acanthaceae. Kew bulletin, Vol. 55, n°3 : 513 - 589.
73. SHENDAGE, S. M., and YADAV, R. V., 2009. Pollen
morphology of Barleria L. (Acanthaceae) from India.
Phytomorphology, 59(3/4) : 121 - 126
74. TRIPP, E. A., and McDADE, L.A., 2012. New synonymies for
Ruellia (Acanthaceae) of Costa Rica and notes on other neotropical
species. Brittonia, 64 : 305 - 317.
75. VOLLESEN, K. 1989. A revision of Megalochlamys
and Ecbolium (Acanthaceae: Justicieae). Kew Bulletin, 44
: 601 - 680.
76. WASSHAUSEN, D.C., and WOOD, J.R.I., 2004. Acanthaceae of
Bolivia. Contributions from the United States National Herbarium,
Department of Botany National Museum of Natural History. Washington,
DC, vol. 49 : 152p, including 42 figures.
77. WHITE, F., 1983. The vegetation of Africa, a descriptive
memoir to accompany the UNESCO/ AETFAT/UNSO vegetation map of Africa.
UNESCO, Natural Ressources Rechearch, n° 20 : 356p.
78. WOOD, J.R., 1997. A Hand Book of the Yemen Flora. Kew.
Royal Botanic Garden. 480p.
79. ZHEHNGYI, W., RAVEN, P., et DEYUAN, H., 2011. Flora of
China, Science Press, Beijing, China. vol. 19: 884p.
151
Références bibliographiques
WEBOGRAPHIE
1.
http://wikipedia.org : La famille des
Acanthaceae Juss. (consulté, mars 2009).
2. Anonyme, 2005. Botanical Research And Herbarium Management
System (BRAHMS). University of Oxford (consulté, 2014)
http://herbaria.plants.ox.ac.uk.
3. Anonyme. The families of flowering plant, the Acanthaceae
(consulté, mars 2010)
http://www.jstor.org,
4. De Candolle, Augustin-Pyramus, 1824 à
1873. Théorie élémentaire de la botanique, ou
exposition des principes de la classification naturelle et de l'art de
décrire et d'étudier les végétaux.
(consulté, août 2018) Prodromus Systematis Naturalis Regni
Vegetabilis in
http://wikipedia.org
5. Direction Général de la
Météorologie. Surveillance du Climat, (consulé
décembre 2018) www.maproom dgm.
6. Hennig, W., 1950. Grundzüge einer theorie der
phylogenetischen Systematik, Deutscher Zentralverlag, Berlin. Traduit par
Dwight, D. Davis, 1966. Phylogenetics systematics, University of Illinos Press.
263p. (consulté, août 2011) http://wikipedia.org/ Willi Hennig
7. Hu Jiaqi, Deng Yunfei, John R. I. Wood, Thomas F., Daniel
: Acanthaceae de Chine : 369477 (consulté, mars 2017).
http://www.efloras.org/florataxon.aspx?flora_id=2
&taxon
8. Grundzüge E., 1950. Chronologie de la classification
botanique. Theorie der phylogenetischen Systematik, Deutscher Zentralverlag,
Berlin (consulté, mars 2017).
http://wikipedia.org.
9. ITIS. Catalogue of Life : Indexing the word's known
species. The naturalist Biodiversity Center (consulté, mai 2017)
http://www. catalogueoflife.org/ col/browse /tree
10. Lecointre. G., Le Guyader. H. 2001. Classification
phylogénétique du vivant, Belin, p. 17 (consulté, janvier
2017).
http://wikipedia.org
11. Parc National d'Andohahela (consulté, mai
2008).
https://www.parcs-madagascar.com/aire-protégée/parc-national-andohahela
12. Magnaghi, E.B. and Daniel, T.F. 2004 : Three new
species of Mendoncia (Acanthaceae) from Madagascar. Novon a journal
for botanical nomenclature. Vol. 23, n°2 : 187-196. (consulté le 18 juin
2014). Novon : a journal for botanical nomenclature.
13. Missouri Botanical Garden. 2014. Catalogue of the
Vascular Plants of Madagascar . St. Louis & Antananarivo (consulté,
mars 2017).
http://www.efloras.org/madagascar
14. Missori Botanical Garden. Vascular plant of Madagascar
(consulté, mars 2010).
http://WWW.TROPICOS.org
15. The International Plant Names Index (consulté,
novembre 2010).
http://IPNI.org
152
Références bibliographiques
16. Reich, K. Esticos criticos sobre la Flore de Chile.
81e Familia. (consulté, fevrier 2017)
http://images.mobot.org/efloras/FloraData/060/PDF/V05/
vol5-81 Acanthaceae .pdf
I
Article
PROCEDINGS OF THE CALIFORNIA ACADEMY OF SCIENCES
Fourth Series
Volume 58, No 8, pp, 121-131, 4 figs May 11, 2007
Anisotes (Acanthaceae) in Madagascar
DANIEL1, T. F., MBOLA3, V. A. B.,
ALMEDA 1, F. et PHILLIPSON2, P.,
1Department of Botany, California
Academy of Sciences, 875 Howard Street, San Francisco, California 941-3 ;
2Missouri Botanical Garden, P.O. Box 299, St. Louis, MO, 63166-0299,
U.S.A. and Muséum National d'Histoire Naturelle, Département
Systématique et Evolution. Boîte postale 39, rue Cuvier 57, 75231
Paris, Cedex 05, France ; 3PNM-ANGAP, Parc National
d'Andohahela, BP.73 Tolagnaro 624,
Madagascar.
balzacmbola2018@gmail.com
Two species of Anisotes (Acanthaceae,
Justicieae) are recognized from Madagascar.
Anisotes madagascariensis is endemic to
sandy, coastal regions of southern Madagascar. New morphological,
distributional, and ecological data are presented for this species. A newly
described species, A. divaricatus, is known only from calcareous,
rocky substrates near Lake Tsimanampetsotsa in southwestern Madagascar. Major
morphological distinctions between A. divaricatus and A.
madagascariensis include : habit (divaricate branches vs. ascendant
branches), length (5--11 vs. 14--45 mm long) and shape (obcordate to obdeltate
vs. broadly ovate to elliptic) of leaf blades, corolla form (corolla tube :
corolla 0.46--0.56 vs. 0.23--0.38; lower lip not spirally coiled vs. spirally
coiled), and capsule pubescence (lacking trichomes vs. trichomes present).
Images of flowers and pollen, morphological descriptions, and mapped
distributions are provided for both species.
Nineteen species occurring from tropical and southern Africa
to Madagascar and tropical Arabia were recognized in the most recent revision
of Anisotes (Baden 1981). The genus has traditionally been included in
subfamily Acanthoideae, tribe Justicieae based on its 2-lipped,
«imbricate» (presumably ascending cochlear aestivation in
Anisotes and its relatives) corollas with bithecous stamens. Baden
(1981) and others have noted that generic delimitation in Justicieae is
problematic. Indeed, Anisotes does not appear well circumscribed in
the tribe based on either macromorphological or palynological characters. No
morphological synapomorphies can be identified for it, and it is reasonable to
suppose that it is not necessarily monophyletic. Baden (1981) did not identify
a probable closest relative for Anisotes, but he noted some
morphological similarities (especially corollas) between Anisotes and
Metarungia Baden (as Macrorungia C.B. Clarke). He
distinguished them by the presence of elastically separating placenta (with
attached retinacula) in capsules of the latter genus; placentas/retinacula
remain attached to the inner surface of capsules in Anisotes.
A single species of Anisotes has been reported from
Madagascar, A.madagascariensis; however, Lindau (1898) described
Symplectochilus madagascariensis Lindau from Madagascar, and
Symplectochilus is now treated as congeneric with Anisotes.
Baden (1981) indicated that the taxonomic position of S. madagascariensis
«remains doubtful due to inadequate type material.» Indeed, the
type (Antsiranana : Bay of Diego Suarez, forest, 1833, Goudot s.n. at
G, photo!) of this name was collected in a different habitat far from the known
distribution of A. madagascariensis, does not represent the same taxon
as the type of A. madagascariensis, and does not necessarily pertain
to the genus Anisotes. However, plants resembling Anisotes
have been reported (e.g., Gautier 2002) in other regions of Madagascar
;
II
Article
122 PROCEDING OF THE CALIFORNIA ACADEMY OF
SCIENCES
Fourtth series, Volume 58, n°8
Benoist annotated specimens at P with names of several new
species and new combinations for plants from Madagascar and the Comoros, but he
did not publish them ; and several herbarium specimens of unknown taxa (e.g.,
Phillipson 1904 at CAS from Mahajanga, Bosser 5917 at TAN
from Montagne d'Ambre) show affinities to Anisotes. This summary of
Anisotes in Madagascar incorporates information not known to or
reported by Baden (1981) for A. madagascariensis, such as :
descriptions of capsules and seeds, fruiting phenology, detailed distributional
data, chromosome number, and phylogenetic position based on molecular sequence
data. In addition, a second distinctive species from southwestern Madagascar is
newly described and compared to A. madagascariensis. Recent molecular
phylogenetic studies that included Old World «justicioids» (McDade et
al. 2000 ; Kiel et al. 2006) reveals that Anisotes is closely related
to several species of Justicia from the Old World (e.g., J.
adhatoda L., J. anagalloides (Nees) T. Anderson, and
J. campylostemon (Nees) T. Anderson) and Duvernoia
aconitiflora A. Meeuse. Based on limited sampling of Anisotes,
Kiel et al. (2006) showed that the African species A. rogersii S.
Moore is sister to J. adhatoda, and that this pair is sister to A.
madagascariensis in a strongly supported clade. Additional molecular
markers as well as more samples of Anisotes and Old World
«justicioids» will be necessary to test monophyly for the 19 species
treated by Baden (1981) and to resolve possible phylogenetic relationships
among them, but there appears to be little doubt that A. madagascariensis
is closely related to African species of Anisotes. Metarungia
appears to be monophyletic (based on a sampling of two species ; Kiel et
al. 2006) and basal to a larger clade that includes Anisotes. Daniel
(2007) reported a chromosome count of n = 17 for Anisotes
madagascariensis, the first number reported for the genus. Chromosome
numbers of n =17 have been reported elsewhere among Old World
«justicioids» in two species of Justicia (Daniel and Chuang
1998 ; Daniel 2000 ; Daniel et al., 2000), in Duvernoia aconitiflora
(Daniel et al., 2000), and in Rungia repens Nees (Subramanian and
Govindarajan 1980 ; however, most counts for this species are n = 10 :
Narayanan 1951 ; Ellis 1962 ; Ranganath 1981 ; Krishnappa and Ranganath 1982 ;
Saggoo 1983). Counts of n = 17 are well established in both J.
betonica L. of section Betonica and J. adhatoda of
section Vasica. The common chromosome number shared by A.
madagascariensis, J. adhatoda, and D. aconitiflora
undoubtedly reflects their close phylogenetic relationships as revealed by
molecular markers (Kiel et al. 2006). In spite of counts of n = 17 in
J. betonica and Rungia repens, neither the former species nor
the latter genus appear especially closely related to the clade consisting of
Anisotes and J. adhatoda based on molecular phylogenies
(McDade et al. 2000 ; Kiel et al. 2006). The sole chromosome count for
Metarungia, posited as a potentially close relative of Anisotes
by Baden (1981), is n = 14 (Daniel et al. 2000). Muller et al.
(1989) described and figured pollen of A. madagascariensis (from
Humbert & Swingle 5295) as 3-colporate and 6-pseudocolpate. Baden
(1981; based on Perrier de la Bâthie 9494) described it as
3-porate with a trema area having reticulate bands (i.e., 3-colporate,
6-pseudocolpate). Variation among other species of the genus (Baden 1981)
includes pollen 2--4-porate (or 2-colporate) with the trema area having two or
four rows of insulae rather than pseudocolpi. Baden (1981) used palynological
differences to help delimit six sections of Anisotes. Pollen of the
newly described species from Madagascar, A. divaricatus, resembles
that of A. madagascariensis in size, aperture type and number, and
exine microsculpturing (i.e., bireticulate).
III
Article
Daniel et al., ANISOTES (ACANTHACEAE) IN
MADAGASCAR 123
Anisotes Nees
Type : A. trisulcus (Forssk.) Nees
Himantochilus T. Anders. ex Benth. Type: H.
sessiliflorus T. Anders. Symplectochilus Lindau Type: S.
formosissimus (Klotzsch) Lindau
Suffrutescent perennials, shrubs, or small trees. Leaves
deciduous or evergreen, opposite, sessile to petiolate, margin entire to
crenate. Inflorescences of axillary or terminal spikes, racemes, or thyrses ;
bracts opposite; bracteoles present or absent; dichasia 1-3- flowered, sessile
to pedunculate. Flowers sessile to pedicellate ; calyx 5-lobed; corolla
reddish, purple, orange, or yellowish, tube #177; cylindric to ampliate
distally, limb 2-labiate, upper lip convavo-convex, usually #177; arched,
entire to 2-fid at apex, internally rugulate, lower lip deeply 3-lobed,
straightforwardly splayed, recoiled, or spirally twisted, aestivation
presumably ascending cochlear (lower lip outermost in bud, but lobes of lower
lip either not evident or not overlapping in buds observed). Stamens 2,
inserted near apex of corolla tube, exserted from corolla tube but not
extending beyond upper lip (under which they are #177; enclosed), anthers
2-thecous, thecae parallel to #177; oblique, subequally to unequally inserted,
#177; equal to unequal in size, basal appendage present or absent; staminodes
absents ; pollen 2-, 3-, or 4-aperturate, colporate/pseudocolpate or porate (to
colporate) in trema regions studded with circular insulae. Stigma 2-lobed or
lobes obscure. Capsule stipitate, retinacula remaining attached to inner wall
of mature capsule. Seeds 4 (or fewer by abortion), lenticular.
Key to Species of Anisotes in
Madagascar
1. Leaf blades broadly ovate to broadly elliptic to elliptic
to ovate-elliptic, 14--45 mm long, 10-- 38 mm wide, 1.1--2.7 times longer than
wide, rounded (to emarginate) at apex ; calyx 2--3 mm long, margin of lobes
#177; densely ciliate ; corolla with the internal surface conspicuously lighter
colored than the external surface, corolla tube : corolla = 0.23--0.38, corolla
tube 9--15 mm long, upper lip 20--35 mm long and distally whitish to pinkish
along the margin, lower lip spirally coiled, 18--28 mm long, lobes 12--14 mm
long; stamens 26--33 mm long; capsule pubescent with flexuose to antrorse
eglandular trichomes 0.05--0.1 mm long; growing on sandy
flats and dunes. A. madagascariensis
1. Leaf
blades broadly obovate to subcircular to obcordate to obdeltate to oblate,
5--11mm long, 4.5--11.5 mm wide, 0.7--1.3 times longer than wide, emarginate to
truncate at apex ; calyx 1.3--2.7 mm long, margin of lobes eciliate to sparsely
ciliate; corolla with the internal surface not conspicuously lighter in color
than the external surface, corolla tube : corolla = 0.46--0.56, corolla tube
14--19 mm long, upper lip (11-) 14--17 mm long, lacking a pale margin distally,
lower lip recurved to reflexed (not spirally coiled), 11--16 mm long, lobes
linear, 8--12 mm long ; stamens 15--18.5 mm long; capsule #177; scurfy but
lacking noticeable eglandular trichomes;
growing on limestone A. divaricatus
Anisotes madagascariensis R. Ben. Bull. Soc.
Bot. France 76:1036. 1929.
TYPE : MADAGASCAR. Toliara:
dunes, rives du Lac Mananpetsa [Tsimanampetsotsa], coté Mahafaly, June
1910, H. Perrier de la Bâthie 9494 (lectotype, designated by
Baden in 1981: P!; isolectotype: P, digital image!). Figure 1.
IV
Article
124 PROCEDING OF THE CALIFORNIA ACADEMY OF
SCIENCES
Fourtth series, Volume 58, n°8
Shrubs to 2 m tall; older branches (#177; divaricate to)
ascending usually at angles of 40-70°. Young stems quadrate-ridged,
densely and evenly pubescent with (flexuose to) antrorse to antrorsely
appressed eglandular trichomes 0.2-0.4 mm long, pubescence resulting in a
grayish green coloration. Leaves coriaceous to subsucculent, petiolate,
petioles to 12 mm long, blades

Figure 1. Flowers of Anisotes divaricatus (a--b) and
A. madagascariensis (c). a. Almeda 9237, scale = 4 mm, photo
by F. Almeda. b. Almeda 9237, scale = 5 mm, photo by H. Mally. c.
Almeda 7705, scale = 8 mm, photo by F. Almeda.
V
Article
Daniel et al., ANISOTES (ACANTHACEAE) IN
MADAGASCAR 125
broadly ovate to broadly elliptic to elliptic to
ovateelliptic, 14-45 mm long, 10-38 mm wide, 1.12.7 times longer than wide,
acute at base, rounded (to emarginate) at apex, venation obscure, surfaces
densely and evenly pubescent with cauline type trichomes, pubescence resulting
in a grayish green coloration. Inflorescence of #177; congested, axillary
(usually only in axils of distalmost 1-3 pairs of leaves) and/or terminal,
pedunculate spikes, peduncles to 24 mm long, rachis densely pubescent like
young stem, internodes usually very short or inconspicuous. Bracts opposite,
all fertile, subtending a single flower, broadly triangular, 1-2 mm long,
1.5-2.2 m wide, abaxial surface densely pubescent with antrorse eglandular
trichomes 0.1-0.2 mm long. Bracteoles absent. Flowers sessile (to subsessile),
usually more than 2 per spike. Calyx 5-lobed, 2-3.5 mm long, tube ca. equal to
lobes (i.e., slightly longer to slightly shorter than lobes), lobes ovate to
triangular, 0.6-1.8 mm long, 0.9-1.1 mm wide, 0.30-0.69 times as long as calyx,
abaxial surface nearly glabrous to sparsely pubescent (especially near apex)
with antrorse to antrorsely appressed eglandular trichomes 0.05-0.1 mm long,
margin #177; densely ciliate with flexuose to antrorse eglandular trichomes.
Corolla maroon to dull red to pink-brown externally, pinkish to whitish
internally, externally glabrous, (30-) 33-47 mm long, tube proximally
cylindric, #177; gradually expanded distally, 9-15 mm long, 3-4 mm in diameter
near midpoint, corolla tube : corolla = 0.23-0.38, upper lip 20-35 mm long,
internally rugulate, entire at apex, distal portion with whitish to pinkish
margin, lower lip spirally coiled, 18-28 mm long, lobes 12-14 mm long, 1.6-3.2
mm wide. Stamens 26-33 mm long, inserted near apex of corolla tube, exserted
from mouth of corolla but not surpassing upper lip, filaments whitish,
glabrous, thecae maroon, unequal in size (distal theca longer, 3.2-4 mm long,
proximal theca shorter, 2.4-3.5 mm long), unequally inserted (overlapping by
1-1.6 mm), glabrous, lacking basal appendages (or proximal theca with an
inconspicuous appendage to 0.1 mm long); pollen 62.5-74 ìm polar
diameter (P), 32.5-42.5 ìm equatorial diameter (E), P:E = 1.52-1.90.
Style 29-36 mm long, glabrous, stigma lobes obscure or 0.2 mm long. Nectar disk
cupuliform and often visible between calyx lobes. Capsule 18-24 mm long,
externally pubescent with flexuose to antrorse eglandular trichomes 0.05-0.1 mm
long, stipe 9-11 mm long, head ellipsoid to obovoid. Seeds 4, discoid, (3.2-)
4.5-5 mm long, (3.6-) 4.1-5 mm wide, surfaces minutely papillose or rugose and
shiny, margin #177; swollen and smooth. (n = 17, Daniel 2007).
Floral Phenology. Flowering : throughout the
year ; fruiting : May, August-October.
Distribution and habitats. Endemic to
southern Madagascar; plants occur on dunes and in sandy flats in regions of
spiny forest (with Diderea madagascariensis and Euphorbia
stenoclada) generally along the coast of the Mozambique Channel at
elevations from sea level to 20 m (Fig. 2). Two apparently wild collections
were made in coastal regions of the Indian Ocean near Ambovombe Androy, well to
the east of Cap Sainte Maire and about 225 km disjunct from the nearest locale
of the species at Itampolo on the west coast (Allorge 428 and
Decary 2729--not seen, but cited among the syntypes of A.
madagascariensis in the protologue). The coastal region between Itampolo
and Ambovombe remains poorly collected, and it is possible that the range of
the species is somewhat more extensive than presently known in coastal regions
of southern Madagascar.
Local names. «Berava,»
«beravo» (Koechlin 25) ; «hazontsohy»
(Ravelonanahary 3286-RN) ; «hazontsoy» (Ravelonanahary
2787-RN, 3926-RN, 4287-RN) ; «hazontsohy lahy»
(Ravelonanahary 4159-RN) ; «voamany» (Koechlin 1)
; «voamamy» (Koechlin 2). Baden (1981) treated
species of Anisotes in six sections based primarily on characters of
the inflorescence and pollen. Anisotes madagascariensis was placed
into section Spiciflori based on its inflorescence
VI
Article
126 PROCEDING OF THE CALIFORNIA ACADEMY OF
SCIENCES
Fourtth series, Volume 58, n°8
spikes, non-reticulate nerved bracts, «large»
corolla: tube ratio, and 3-aperturate pollen with apertures flanked by
pseudocolpi (Fig. 3). The only other species placed into this section by Baden
(1981), A. formosissimus (Klotzsch) Milne-Redhead, occurs in
Mozambique, Zimbabwe, and Malawi in southeastern Africa. It differs from A.
madagascariensis by its longer spikes, bracts, and calyx; glabrate leaves;
and bracteoles, which are present. Although Baden (1981) cited only 5
collections of this species, plants are rather frequent along the southwestern
coast of Madagascar. Because plants often either persist or become established
in disturbed habitats, this attractive plant is well suited for horticultural
use in local landscaping. Cours 4643 differs from other collections by
having spikes 2-flowered and flowers subsessile (i.e., borne on pedicels to 1
mm long); in other respects, it resembles most other collections. Although
pollinators have not been documented for A. madagascariensis,
Long-billed Green Sunbirds (Nectarinia notata) were observed visiting
flowers of Daniel & Butterwick 6736 in November.
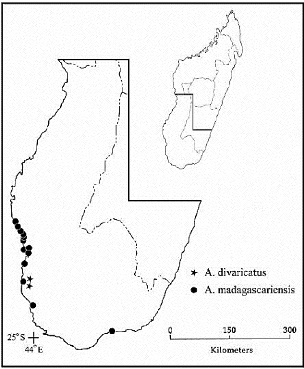
Figure 2. Map of southwestern Madagascar showing distributions of
A. divaricatus and A. madagascariensis.
Additional specimens examined. MADAGASCAR.
Toliara : Ambovombe, chemin 100 m avant l'océan, 11 September 1992
(flr), L. Allorge 428 (P) ; just N of Itampolo on Mozambique Channel,
1 April 1996 (flr), F. Almeda 7705 (CAS) ; Itampolo, 13 May 1951 (flr,
frt), J. Bosser 78 (P, TAN) ; Efoetsy, 12 May 1951 (flr), J.
Bosser 116 (P), 126 (TAN); environs de Tulear, November 1956
(flr), J. Bosser 10598 (P, TAN) ; au nord du Fiherenana,
27 June 1958 (flr), R. Capuron I8623-SF (P) ; without
locality, M. Cours 4643 (P) ; ca. 5 km S of Hotel Lakana Vezo in Ifaty
toward Toliara (ca. 20 km N of Toliara along coast), ca. 0m, 5 November 1993
(flr), T. Daniel & M. Butterwick 6736 (CAS) ; ca.
28 km N of Toliara, just N of Ifaty along road to
Ankaraobato, 23°6'S,
43°36.5'E, 5 m, 9 October
2003 (flr, frt), T. Daniel et al. 10422 (CAS, K, MO, P, PH, TAN) ;
District Betioky, Anakao, 21 October 1940 (flr), R. Decary 16077 (P);
environs terrain d'aviation Tuléar, J. DeQuaire 27548 (P) ;
Itampolo, 1945 (flr), A.-M.
VII
Article
Daniel et al., ANISOTES (ACANTHACEAE) IN
MADAGASCAR 127
Homolle 1650 (P) ; environs de Tuléar, delta
du Fiherenana, 6 August 1928 (flr, frt), H. Humbert & C. Swingle 5164
(P, TAN), 14--26 September 1924 (flr), H. Humbert et al. 2425 (P)
; du Lac Manampetsotsa au delta de La Linta (Cote Sud-Ouest), 1--10 m, 17--24
August 1928 (flr), H. Humbert & C. Swingle 5295 (P, UC); Anakao, 3
September 1967 (flr), B. Koechlin 1 (P) ; Manombo, Anjevo, 4 November
1967 (flr), B. Koechlin 2 (P) ; Songeritelo, 23 September 1967
(sterile), B. Koechlin 25 (P) ; environs de Tuléar, route de
Manombo, 3 November 1960 (flr), J. Leandri & R. Jean de Dieu 3626
(P), 3634 (P) ; 21 km N of Ifaty on track from Manambo to
Tsifota, near Fiserenamasay, 22°48'S, 43°18'E, 0 m, 4 August 1993
(flr), B. Lewis et al. 515 (TAN); Tsivonoa N., 11 September 1961
(flr), J. Peltier & M. Peltier 3384 (P, TAN) ; environs de Tulear,
August 1919 (flr, frt), H. Perrier de la Bâthie 19811 (P) ; 20km
N of Tulear, near resort of Mora Mora, Ifaty [«Italy»], 23°11'S,
43°07'E, 10 m, 7 May 1987 (flr), P. Phillipson 1776 (CAS, P, TAN)
; N of Itampolo, 24°41'S, 43°57'E, 10 October 1990 (flr), P.
Phillipson et al. 3739 (CAS, P, TAN) ; Befanany, 2 August 1921 (flr),
H. Poisson 277 (P) ; outskirts of village of Itampolo, 24°42'S,
43°57'E, 0 m, 9 February 1990 (flr), B. du Puy et al. MB686 (P,
TAN) ; NW of Toliara, Forest of Mikea, ca. 13 km N of mouth of Manombo River,
22°53'S, 43°24'E, 10 m, 4 December 1993 (flr), D. du Puy et al.
M691 (P, TAN) ; Tulear, Raich 1097 (TAN); District Tulear,
Réserves Naturelles, 22 May 1951 (flr), L. de G. Ravelonanahary
2787-RN (P, TAN) ; District Betioky, Réserves Naturelles, 31
December 1951 (flr), L. de G. Ravelonanahary 3286-RN (P) ; District
Betioky, Réserves Naturelles, L. de G. Ravelonanahary 3926-RN
(P, TAN) ; District Betioky, Réserves Naturelles, July 1952 (flr),
4159-RN (P, TAN) ; District Tulear, Réserves Naturelles, 24
October 1952 (flr), L. de G. Ravelonanahary 4287-RN (P, TAN) ; 35 km N
of Toliara along coast road, 23°04'S, 43°35'E, 20 m, 12 December 1988
(flr), G. Schatz & J. Miller 2471 (CAS) ; ca. 19 km NNW of Tulear
on RN 9, ca. 2 km N of Songoritela toward Morombe, 0 m, 9 June 1991 (flr),
23°12'S, 43°37'E, J. Zarucchi et al. 7575 (TAN).
Anisotes divaricatus TF Daniel, Mbola, Almeda,
and Phillipson, sp. Nov.
TYPE: MADAGASCAR. Toliara: Réserve du
Lac Tsimanampetsotsa (No. 10), NW corner of reserve, 24°04'S,
43°46'E, escarpment of calcareous plateau, 50 m, 8 October 1990, P.
Phillipson et al. 3721 (holotype: TAN!; isotypes: CAS!, P!). Figures 1,
4.
Frutex usque ad 1 m, ramis vetustis #177; divaricatis. Folia
petiolata, laminae subsucculentae, late ovatae vel subcirculares vel obcordatae
vel obdeltatae vel oblatae, 5-11 X 4.5-11 mm, 0.7-1.3-plo longiores quam
latiores, emarginatae vel truncatae ad apicem, costa solum manifesta (in pagina
abaxiali). Spicae axillares, pedunculares. Bracteae triangulares vel late
triangulares, 1-1.3 mm longae. Calyx 1.3-2.7 mm longus. Corolla rubra-purpurea
intus, 27-36 mm longa. Capsula 13-20 mm longa, trichomatibus eglandulosis
nullis. Semina 3.8-4.8 mm longa. Scrambling to clambering or arching shrub to 1
m; older branches #177; divaricate. Young stems quadrate-ridged [similar to
subhexagonal but only small valleys at corners], densely and evenly pubescent
with (flexuose to) antrorse to antrorsely appressed eglandular trichomes
0.2-0.4 mm long, resulting in a grayish cast. Leaves petiolate, petioles to 5
mm long, blades subsucculent, broadly obovate to subcircular to obcordate to
obdeltate to oblate, 5-14 mm long, 4.5-14.3 mm wide, 0.7-1.3 times longer than
wide, acute at base, emarginate to truncate at apex, venation obscure, midvein
only evident on abaxial surface, surfaces and margin #177; densely and evenly
pubescent with cauline type trichomes, resulting in a grayish cast.
Inflorescence of #177; congested, axillary (in axils of distalmost 1-3 pairs of
leaves), pedunculate spikes; peduncles 2-11 mm long, rachis densely to sparsely
pubescent like young stems, internodes to 1 mm long. Bracts opposite, all
fertile, each subtending a single flower, triangular to broadly triangular,
1-1.3 mm long, 0.8-1.2 mm wide, abaxial surface densely pubescent with
(flexuose to) antrorse to antrorsely appressed eglandular
VIII
Article
128 PROCEDING OF THE CALIFORNIA ACADEMY OF
SCIENCES
Fourtth series, Volume 58, n°8
trichomes 0.1-0.2 mm long. Bracteoles absent. Flowers sessile
to subsessile (i.e., pedicels to 0.3 mm long). Calyx 5-lobed, 1.3-2.7 mm long,
tube shorter than lobes, lobes triangular to lanceolate
to ovate to obovate, 1-1.7 mm long, 0.7-1 mm wide, 0.63-0.87
times as long as calyx, abaxial
surface sparsely pubescent (especially distally) with antrorse
to antrorsely appressed eglandular trichomes 0.05-0.1 mm long, margin eciliate
to sparsely ciliate with flexuose to antrorse
eglandular trichomes. Corolla #177; concolorous, dull
purple-red (maroon), or #177; discolorous with the tube pink-brown (bronze
colored) and the lobes of the lower lip maroon, externally mostly glabrous
(occasionally with a few flexuose to retrorse eglandular trichomes to 0.2 mm
long
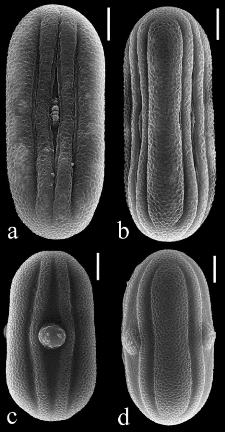
Figure 3. Pollen of Anisotes divaricatus (a, b;
Almeda 9237) and A. madagascariensis (c,d; Daniel et al.
10422). a. apertural view. b. interapertural view. c. apertural view. d.
interapertural view. Scale lines = 10 ìm.
proximally),27-40 mm long, tube 14-20 mm long, narrow proximal
portion subcylindric, 4-7 mm long, #177; abruptly expanded into a funnelform
throat, throat 9-14 mm long, 2.8-4 mm I diameter near midpoint, corolla
tube:corolla = 0.46-0.56, upper lip (11-) 14-18 mm long, internally rugulate,
entire to emarginate at apex, lacking a pale colored margin distally, lower lip
recurved to reflexed, 11-19 mm long, lobes linear, 8-12 mm long, 1-4 mm wide,
central lobe larger than lateral lobes. Stamens 15-18.5 mm long, inserted near
apex of corolla tube, exserted from mouth of corolla but usually not surpassing
upper lip (or surpassing it by up to 1 mm), filaments cream, glabrous, thecae
yellow and brown, unequal in size (distal larger, 2.4-3.3 mm long; proximal
shorter, 2-2.8 mm long), unequally inserted (overlapping by 0.8-1.2 mm),
parallel to slightly oblique, glabrous, lacking basal appendages or proximal
theca with an inconspicuous basal appendage 0.05 mm long; pollen 72 ìm
polar diameter (P), 28-30.5 ìm equatorial diameter (E), P:E= 2.4-2.6.
Style 24-32 mm long, glabrous, stigma 0.2-0.3 mm long, lobes obscure. Nectar
disk asymmetric, often with a lobe-like appendage to 0.6 mm long protruding
between outermost (i.e., farthest from rachis) pair of calyx lobes. Capsule
13-20 mm long, surface #177; scurfy but lacking noticeable eglandular
trichomes, stipe 5-8 mm long, head ovoid-ellipsoid to obovoid-ellipsoid with a
slight medial constriction. Seeds 4, discoid, 3.8-4.8 mm long, 3.7-5.1 mm wide,
surfaces minutely
papillose or rugose, shiny, margin sometimes #177; swollen and
smooth.
IX
Article
Daniel et al., ANISOTES (ACANTHACEAE) IN
MADAGASCAR 129
Phenology. Flowering : August-May; fruiting :
August-October, April.
Distribution and habitat. Endemic to
southwestern Madagascar; plants occur on limestone rock (Mahafaly Plateau
limestone) with spiny forest (dominated by Alluaudia comosa,
Commiphora madagascariensis, Delonix floribunda, D.
adansonioides, Adansonia rubrostipa, Euphorbia
oncoclada, E. laroo, and Uncarina stellulifora) in the
vicinity of Lake Tsimanampetsotsa at elevations from 22--50 m (Fig. 2).
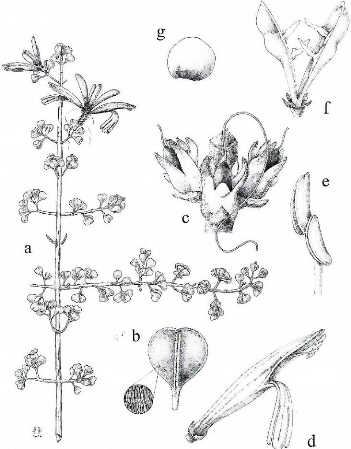
Figure 4. Anisotes divaricatus. a. Habit (Leandri
& Saboureau 4441), ×0.5. b. Leaf (abaxial surface), ×2.1,
with close-up of pubescence, ×16 (Leandri & Saboureau 4441).
c. Inflorescence nodes with corollas removed (Bosser 1762), ×8.
d. Flower (Bosser 15609 & Perrier de la Bâthie
19156), ×1.7. e. Distal portion of stamen (Perrier de la
Bâthie 19156), ×7.8. f. Capsule (Perrier de la Bâthie
19156), ×3. g. Seed (Perrier de la Bâthie 19156),
×3.5. Drawn by Erin Hunter.
X
Article
130 PROCEDING OF THE CALIFORNIA ACADEMY OF
SCIENCES
Fourtth series, Volume 58, n°8
Anisotes divaricatus is both morphologically and
ecologically distinct from A. madagascariensis. However, based on its
spicate inflorescence, bracts lacking reticulate venation, and 3-aperturate
pollen (Fig. 3), A. divaricatus would also appear to belong to section
Spiciflori. Like A. madagascariensis and unlike A.
formosissimus, the only other species of section Spiciflori,
A. divaricatus lacks bracteoles. Indeed, the two Malagasy species are
the sole representatives of the genus in which these structures are not
present. Both Malagasy species occur in the vicinity of Lake Tsimanampetsotsa
and the general collecting locales are sometimes identical for each (e.g.,
«Efoetsy» or «near Lake Tsimanampetsotsa»). In most such
instances, however, plants of A. madagascariensis were noted to have
been collected from maritime sands or dunes, and at lower elevations than those
of A. divaricatus.
A collection from a rocky forest on «Plateau
Mahafaly» (Perrier de la Bâthie 9521 at P) contains a small
branch that likely pertains to A. divaricatus. Like that species, it
has round to oblate leaves on petioles up to 5 mm long and corollas with the
tube:corolla=0.47. However, it differs from that species by its slightly longer
vegetative trichomes (up to 0.5 mm long), larger leaves (up to 15 × 17
mm), smaller corollas (to 17 mm long) with the tube to 8 mm long and not or but
barely expanded distally, and smaller anther thecae (1.3--1.5 mm long). Until
additional material is available that resembles this collection or that links
it to specimens noted herein, Perrier de la Bâthie 9521 is not
included within the circumscription of A. divaricatus.
Paratypes. MADAGASCAR. Toliara: NE sector of Tsimanampetsotsa
National Park, 24°02.895'S, 43°45.138'E, ca. 22 m, 25 August 2006
(flr, frt), F. Almeda 9237 (CAS, TAN); Efoetsy, 12 May 1951 (flr),
J. Bosser 187 (P) ; Lac Tsimanampetsotsa, R.N. 10, February 1962
(flr), J. Bosser 15,609 (P, TAN) ; Réserve 10, Lac
Tsimanampetsotsa, February 1962 (flr), M. Keraudren 1403 (P); environs
du Lac Tsimanampetsa, environs de l'aven de Mitoho, 23 November 1960 (flr),
J. Leandri & P. Saboureau 4441 (P); Manampetsa, April 1933 (flr,
frt), H. Perrier de la Bâthie 19156 (P); Réserve de
Tsimanampetsotsa, SW corner of reserve, 24°11'S, 43°48'E, 50 m, 13
January 1989 (flr), P. Phillipson & S. Rabesihanaka 3169 (CAS, P,
TAN).
ACKNOWLEDGMENTS
Field studies in 2003 were supported by a grant from the
National Science Foundation (DEB 0108589) to L.A. McDade and T.F. Daniel. We
thank De Ada and Henry Mally for photographing A. divaricatus at Lake
Tsimanampetsotsa; Heritiana Ranarivelo, Lucinda McDade, and Randriambololona
Tantely Nirina for assisting with field work and logistics in Madagascar; and
Erin Hunter, the 2006 Fellows Intern in Biological Illustration at the
California Academy of Sciences, for her exceptional drawing, which helped to
relocate the new species at Lake Tsimanampetsotsa in 2006. We are grateful for
courtesies extended by the following herbaria from which specimens were
examined: CAS, P, TAN, and UC. We also thank MO for sending duplicate specimens
of Anisotes and other Malagasy Acanthaceae to CAS. The staff at G
kindly provided a digital image of the type of Symplectochilus
madagascariensis.
XI
Article
Daniel et al., ANISOTES (ACANTHACEAE) IN
MADAGASCAR 131
LITERATURE CITED
BADEN, C. 1981. The genus Anisotes (Acanthaceae), a
taxonomic revision. Nordic Journal of Botany 1:623--664.
BENOIST, R. 1929. Descriptions d'espèces nouvelles
d'acanthacées de Madagascar. Bulletin de la Société
Botanique de France 76:1031--1038.
DANIEL, T.F. 2000. Additional chromosome numbers of American
Acanthaceae. Systematic Botany 25:15-25.
DANIEL, T.F. 2007. Chromosome numbers of miscellaneous
Malagasy Acanthaceae. Brittonia 58: in press.
DANIEL, T.F. AND T.I. CHUANG. 1998. Chromosome numbers of
cultivated Acanthaceae and systematic implications. Pp. 309--330. In :
P. Mathew and M. Sivadasan (eds.), Diversity and Taxonomy of Tropical
Flowering Plants. Mentor Books, Calicut.
DANIEL, T.F., K. BAKLWILL, AND M.-J. BALKWILL. 2000.
Chromosome numbers of South
African Acanthaceae. Proceedings of the California Academy
of Sciences 52:143--158. ELLIS, J.L. 1962. Chromosome numbers in some
members of Acanthaceae. Science & Culture
28:191--192.
GAUTIER, L. 2002. Liste commentée des
phanérogames de la Réserve Spéciale de Manongarivo,
Madagascar. Boissiera 59 : 105--239.
KIEL, C.A., L.A. MCDADE, AND T.F. DANIEL. 2006. The
species-rich and biologically diverse «justicioids» (Acanthaceae :
Justicieae) : phylogeny, biogeography, and morphological evolution. Botany
2006: Abstracts, Scientific Meeting, Abstract 622:284.
KRISHNAPPA, D.G. AND R.M. RANGANATH. 1982. Acanthaceae. Pp.
364--365. In: A. Löve (ed.) IOPB chromosome number reports LXXV.
Taxon 31:342--368.
LINDAU, G. 1898. Einige neue acanthaceen. Annuaire du
Conservatoire et du Jardin Botaniques de Genève 2:1--140.
Mc.DADE, L. A., T.F. DANIEL, S.E. MASTA, AND K.M. RILEY. 2000.
Phylogenetic relationships within the tribe Justicieae (Acanthaceae): evidence
from molecular sequences, morphology, and cytology. Annals of the Missouri
Botanical Garden 87:435-- 458.
MULLER, J., M. SCHULLER, H. STRAKA, AND B. FRIEDRICH. 1989.
Palynologia Madagassica et Mascarenica. Fam. 182 : Acanthaceae. Tropische
und Subtropische Pflanzenwelt 67:138--187.
NARAYANAN, C.R. 1951. Somatic chromosomes in the Acanthaceae.
Journal of the Madras University 21:220--231.
RANGANATH, R.M. 1981. Morphological and cytological studies in
Acanthaceae. Ph.D. Thesis, Bangalore. [not seen, cited in :
Monographs in Systematic Botany from the Missouri Botanical Garden
8:33. 1984]
SAGGOO, M.I.S. 1983. Cytomorphological studies on plants of
economic importance of Bicarpellatae from India. Ph.D. Thesis, Punjabi
University, Patiala. [not seen, cited in:
Monographs in Systematic Botany from the Missouri Botanical
Garden 13:18. 1985]
SUBRAMANIAN, D. AND T. GOVINDARAJAN. 1980. Cytotaxonomy of
some species of Acanthaceae. Journal of Cytology and Genetics
15:90--92.
Copyriths (c) 2007 by the California Academy of Sciences
San
Francisco, Californie, U.S.A.
XII
Poster
POSTER : Mbola, V. A. B.,
Daniel, T.F., Letsara, R., Ramavovolona, Faramalala, M.H.,
Distribution des représentants de la famille des
Acanthaceae dans le Sud de Madagascar. AETFAT, 2010 Antananarivo.

XIII
Annexe
Annexe 1 : Questionnaire d'enquête
Date : N°
Commune : Localité :
Zone: sèche transition humide
Chef de famille :
homme femme
Activités principales :
agriculteur éleveurs autres
1. Est-ce qu'il y a, au niveau de ce village, une matrone ou un
tradipraticien ?
Oui Non
2. Quelles types de plante prélevez-vous dans la
forêt ?
arbres lianes herbes
3. Quelle espèce la plus prisée?
3. Quelle partie de la plante prélevez-vous dans la
forêt ?
tige feuille racine fruits
4. Quelle est l'utilisation de chaque espèce ?
construction médicinale alimentaire autres
5. Par rapport à cinq années passées,
à quelle distance du village vous récoltez ces plantes ?
proche loin très loin
6. En cas de récolte, avez-vous des autorisations de la
part de l'organisme chargé de la gestion de la forêt (ANGAP,
Service forestier ou autres) ?
OUI Non
7. Est-ce que vous trouvez encore les plantes que vous cherchez
? En quelle quantité ?
OUI Non
peu suffisante
8. Est-ce que vous n'avez pas l'idée de planter ces
espèces près de votre village pour pouvoir en collecter
facilement plus tard ?
OUI Non
9. Au niveau de votre village, connaissez-vous d'autre(s)
personne(s) qui collecte(nt) les plantes au même endroit que vous ?
OUI Non
10. En cas d'existence d'autres collecteurs, à quoi ces
derniers utilisent -ils les plantes ?
OUI Non
XIV
Annexe
Annexe 2 : Répartition des espèces
rencontrées en fonction des paramètres du milieu.
|
N°
|
Code
|
Genre et espèce
|
Altitude
|
Ouverture du milieu
|
Type de sol
|
Subdivion climatique
|
|
Basse
Altitude
|
Moyenne Altitude
|
Haute
altitude
|
Ouvert
|
Semi-
fermé
|
Fermé
|
Alluvionnaire
|
Sol noir
de forêt
|
Sol
calcaire
|
Humide
|
Transition
|
Sèche
|
|
1
|
BVM 057
|
Asystasia gangetica
|
1
|
1
|
1
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
|
2
|
BVM 054
|
Barleria alluaudii
|
1
|
1
|
1
|
1
|
0
|
0
|
1
|
0
|
0
|
0
|
1
|
0
|
|
3
|
BVM 113/ BVM 132
|
Barleria decaryi
|
0
|
1
|
0
|
1
|
0
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
4
|
BVM 182
|
Barleria her
|
1
|
1
|
0
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
|
5
|
BVM 129
|
Barleria humbertii
|
1
|
0
|
0
|
1
|
0
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
6
|
BVM 117
|
Barleria kitchingii
|
1
|
1
|
1
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
1
|
1
|
|
7
|
BVM 053/ BVM 122
|
Blepharis calcitrapa
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
|
8
|
BVM 101
|
Blepharis maculata
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
|
9
|
BVM 143
|
Brillantaisia madagascariensis
|
0
|
1
|
1
|
0
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
|
10
|
BVM 120
|
Camarotea sp.
|
1
|
0
|
0
|
0
|
1
|
0
|
0
|
0
|
1
|
0
|
0
|
1
|
|
11
|
BVM 051/ BVM 127
|
Crossandra longipes
|
1
|
1
|
0
|
1
|
0
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
12
|
BVM 159
|
Dicliptera decaryi
|
0
|
1
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
0
|
|
13
|
BVM 114
|
Ecbolium
madagascariense
|
1
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
1
|
0
|
0
|
|
14
|
BVM 070
|
Ecbolium syringifolium
|
1
|
1
|
0
|
0
|
0
|
1
|
1
|
1
|
1
|
1
|
0
|
1
|
|
15
|
LRK 756
|
Forsythiopsis vincoides
|
1
|
0
|
0
|
0
|
1
|
0
|
0
|
1
|
0
|
0
|
0
|
1
|
|
16
|
BVM 130/ BVM 185
|
Hypoestes decaryana
|
0
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
17
|
BVM 187
|
Hypoestes longituba
|
0
|
1
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
0
|
|
18
|
BVM 118
|
Hypoestes poissonii
|
1
|
1
|
0
|
0
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
|
19
|
BVM 172
|
Hypoestes sp.1
|
0
|
1
|
0
|
0
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
XV
Annexe
Annexe 2 : Répartition des espèces
rencontrées en fonction des paramètres du milieu (suite).
|
N°
|
Code
|
Genre et espèce
|
Altitude
|
Ouverture du milieu
|
Type de sol
|
Subdivion climatique
|
|
Basse
Altitude
|
Moyenne Altitude
|
Haute
altitude
|
Ouvert
|
Semi-
fermé
|
Fermé
|
Alluvionnaire
|
Sol noir
de forêt
|
Sol
calcaire
|
Humide
|
Transition
|
Sèche
|
|
20
|
BVM 131/ BVM 143
|
Hypoestes sp.2
|
1
|
1
|
0
|
0
|
0
|
1
|
1
|
0
|
0
|
1
|
0
|
0
|
|
21
|
BVM 131
|
Hypoestes sp.3
|
1
|
0
|
0
|
0
|
1
|
0
|
0
|
1
|
0
|
0
|
1
|
0
|
|
22
|
BVM 061/ BVM 158
|
Hypoestes strigata
|
0
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
0
|
|
23
|
BVM 166
|
Justicia sp.1
|
1
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
0
|
|
24
|
BVM 052
|
Lasiocladus anthospermifolius
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
0
|
1
|
0
|
|
25
|
BVM 163
|
Mendoncia delphina
|
0
|
1
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
0
|
|
26
|
LRK 139
|
Neuracanthus richardianus
|
1
|
1
|
0
|
1
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
|
27
|
BVM 123/ BVM 168
|
Phaulopsis imbricata
|
1
|
1
|
1
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
|
28
|
Eboroke 811
|
Ruellia geayi
|
0
|
1
|
1
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
1
|
0
|
|
29
|
BVM 111
|
Stenandriopsis carduacea
|
1
|
0
|
0
|
0
|
1
|
0
|
1
|
0
|
0
|
0
|
1
|
0
|
|
TOTAL :
|
20
|
21
|
9
|
8
|
14
|
8
|
14
|
14
|
7
|
13
|
10
|
8
|
First name : Andrianandrasana Balzac
Surname : MBOLA VERSENE
Thesis title : Systematic inventory of genera and species
representing the Acanthaceae family in Tolagnaro and Amboasary-Atsimo district,
Madagascar.
ABSTRACT
The district of Tolagnaro and Amboasary-Atsimo, from East to
West presents humid forest with Dombeya seyrigii, Psiadia dracaenifolia,
Helichrysum fulvescens, Helichrysum sp., Kalanchoe sp. and Xerophyta
dasylirioides, followed by a transitional forest with Dypsis
decaryi, and dry forest with Adansonia za, Commiphora aprevalii, C.
lamii, Alluaudia procera, A. ascendens and Gyrocarpus americanus.
The present thesis was undertaken with the aim at having complementary data to
advance the key of determination of malagasy Acanthaceae family. Different
bioclimatic and ecological factors were analyzed for different types of habitat
and specimens collected which are describe morphologogicaly. Twenty-nine
species were collected during this study. Five new species grouped in three
genera, Camarotea, Hypoestes and Justicia were found.
Three kinds of habitas are defined, from Est to West, as humid with
thirteen species, transitional area with ten species and eight species in the
dry area. Eacht kind of habitat has its species except Ecbolium
syringifolium and Barleria kitchingii which are seen in two. Only
one specie represents tree type, Ruellia geayi and one species it's a
liana Mendoncia delphina, five species are shrubs and finally
twenty-two are herbaceous. The majority of Acanthaceae have spike
inflorescence. Twenty one are seen in middle altitude, twenty at low altitude
and nine at high altitude. The Acanthaceae plants grow in some kind of habitat
and just eight species are developed in open habitats. Thirteen species occur
on black forest soil. Acanthaceae are good climate indicate except Ecbolium
syringifolium which is seen in humid and transitional zones and
Barleria kitchingii is found in transitional and dry zones. After a
description, a partially species-specific identification key is given relative
to twenty nine species in this study.
Key words : Acanthaceae Systematic, Biogeography, Tolagnaro
and Amboasary-Atsimo districts, Madagascar.
Thesis Advisor : Pr. Miadana Harisoa FARAMALALA
Nom : MBOLA VERSENE
Prénoms : Andrianandrasana Balzac
Titre de la thèse : Inventaire systématique des
genres et espèces représentants de la famille des Acanthaceae
dans les districts de Tolagnaro et d'Amboasary-Atsimo, Madagascar.
RESUME
Les districts de Tolagnaro et d'Amboasary-Atsimo, d'Est en
Ouest, présentent une forêt dense humide à Dombeya
seyrigii, Psiadia dracaenifolia, Helichrysum fulvescens,
Helichrysum sp., Kalanchoe sp., Xerophyta dasylirioides,
suivie d'une forêt de transition à Dypsis decaryi, et
d'une forêt dense sèche associée à un fourré
à Adansonia za, Commiphora aprevalii, C. lamii, Alluaudia procera,
A. ascendens et Gyrocarpus americanus. La présente
thèse a été menée dans cette zone dans le but
d'avoir des données complémentaires pour faire avancer la
clé de determination de la famille des Acanthaceae à Madagascar.
Différents facteurs bioclimatiques et écologiques ont
été analysés pour définir les
préférences écologiques des Acanthaceae collectés
et qui ont été décrits morphologiquement. Vingt-neuf
espèces ont été recoltées au cours du
présent travail d'inventaire. Cinq nouvelles espèces
regroupées dans trois genres à savoir Camarotea,
Hypoestes et Justicia ont été trouvées.
Trois types d'habitat ont été définis d'Est en Ouest,
à savoir une partie humide comportant treize espèces, une zone de
transition avec dix espèces et une partie sèche avec huit
espèces. Si toutes les espèces sont propres à un type
d'habitat, Ecbolium syringifolium et Barleria kitchingii se
rencontrent dans deux. Une seule espèce est de type
nanophanérophyte, Ruellia geayi ; une seule espèce est
de type liane, Mendoncia delphina ; cinq espèces sont des
chaméphytes et enfin, vingt-deux sont des hémicryptophytes.
Presque la moitié des Acanthaceae étudiés ont des
inflorescences en épi. Vingt-une espèces se trouvent en moyenne
altitude, vingt en basse et neuf en haute altitude. Les Acanthaceae colonisent
d'autres types de milieu et seules huit espèces se développent en
milieu ouvert. La majorité des Acanthaceae du site étudié
poussent sur sol noir de forêt (16/29 espèces). Les Acanthaceae de
la zone d'étude sont des bons indicateurs climatiques sauf pour
Ecbolium syringifolium qui se développe en zones humide et
sèche et Barleria kitchingii en zones de transition et
sèche. Apres description, une clé de détermination
partielle relative aux espèces étudiées est donnée,
mais elle doit être complétée.
Mots clé : Acanthaceae, Systématique,
Biogéographie, Districts de Tolagnaro et d'Amboasary-Atsimo,
Madagascar.
Directeur de thèse : Pr. FARAMALALA Miadana Harisoa
|
|


