|

UNIVERSITÉ D'ÉTAT
D'HAÏTI
(UEH)
FACULTÉ D'AGRONOMIE ET DE MÉDECINE
VÉTÉRINAIRE
(FAMV)
DÉPARTEMENT DES RESSOURCES
NATURELLES ET ENVIRONNEMENT
(DRNE)
Évaluation du stock de carbone d'un peuplement
d'Acacia mangium âgé de 6 ans, à Bois Nago,
section Bois-Poux dans la commune de Mont-Organisé,
Haïti
Mémoire de fin d'études
Préparé par : Anne-Doty
HÉRISSÉ
Pour l'obtention du diplôme d'Ingénieur
Agronome
Option : Ressources Naturelles et Environnement
(RNE)
Juin 2021
Ce mémoire intitulé :
Évaluation du stock de carbone d'un peuplement
d'Acacia mangium âgé de 6 ans, à
Bois Nago,
section Bois-Poux dans la commune de Mont-Organisé, Haïti
A été approuvé par le jury composé de
:
Signature Date
Neudy JEAN-BAPTISTE ..../..../....
Président du jury
Steevensen ALCIUS ..../..../....
Membre du jury
Eric junior VILMONT ..../..../....
Membre du jury
Odré VALBRUN ..../..../....
Membre, Conseiller Scientifique
i
AVANT-PROPOS
Ce mémoire rentre dans le cadre de l'obtention du
diplôme d'Ingénieur-Agronome de fin d'études
réalisées à la Faculté d'Agronomie et de
Médecine Vétérinaire (FAMV). La FAO (2010) a
souligné l'importance des boisements à croissance rapide dans le
stockage du carbone. Or, les études visant à quantifier le stock
de carbone sont assez récentes et peu nombreuses dans le pays. Ainsi,
cette étude se veut être une contribution à la
communauté scientifique haïtienne en étant un modèle
pour tous ceux qui auront à approfondir cette thématique. Ce
travail décrit de manière concise la méthodologie
appliquée pour la bonne compréhension de tous ceux qui auront
à consulter ce mémoire. Une liste de référence
regroupant l'essentiel des faits scientifiques sur le sujet accompagne
également le document. Les difficultés rencontrées dans la
réalisation de ce mémoire concernent particulièrement la
non disponibilité des équations allométriques
spécifiques à la réalité écologique de la
zone d'étude d'où l'utilisation d'un modèle
généralisé.
ii
REMERCIEMENTS
Je tiens à remercier l'architecte de l'Univers, le
grand Dieu tout puissant qui ne cesse chaque jour de me faire don de sa
grâce.
Je voudrais exprimer toute ma gratitude au professeur
Odré Valbrun, mon Conseiller-Scientifique pour avoir accepté de
m'encadrer dans cette étude. Je le remercie pour sa
disponibilité, sa patience et la qualité de ses conseils.
Je souhaite également adresser mes sincères
remerciements au corps professoral et administratif de la Faculté
d'Agronomie et de Médecine Vétérinaire (FAMV), pour la
richesse et la qualité de leur enseignement.
Je désire aussi remercier tous les membres du Staff de
Ouanaminthe particulièrement Aide Apollon, Gilbert Goimbert, William
Metellus, Marline Raphaël, Dapheney Lumas, pour leur support et leur
aide.
Je tiens à témoigner toute ma reconnaissance
envers mes amis et collègues qui m'ont apporté leur soutien moral
intellectuel tout au long du travail.
Je remercie également toute la famille Apollon de
m'avoir chaleureusement accueilli chez eux à Ouanaminthe, mais aussi de
leurs encouragements, et de tout l'amour manifesté à mon
égard.
Un infini merci à mon oncle Gabriel
Frédéric, pour sa confiance et son soutien inestimable. Je le
remercie également pour sa patience, sa disponibilité, ses
encouragements et ses précieux conseils.
Et enfin je remercie mon infatigable mère Juliette Jean
Philippe ainsi que mon frère Jean Guilloux Hérissé qui ne
se lassent jamais de m'encourager.
iii
RÉSUMÉ
L'Acacia mangium est l'essence forestière
favorisée lors des activités de plantation entreprises par la
Welthungerhilfe (WHH); une évaluation des performances de ces
plantations en termes de stockage de carbone est importante si Haïti
compte respecter ses différents engagements internationaux sur le
changement climatique. D'où le principal objectif de cette étude,
est d'estimer le stock de carbone de la biomasse d'un peuplement d'Acacia
mangium âgé de six ans, situé à Bois Nago et
s'étendant sur une parcelle de 7.15 hectares de superficie. Un
inventaire a d'abord été réalisé par
échantillonnage systématique en implémentant 35 placettes
provisoires de 8 m de rayon, donnant pour chacune une superficie de 199
m2, après avoir appliqué un facteur de correction de
la pente. Avec un diamètre de précomptage de 5 cm, les mesures
prises dans chaque placette sont celles du diamètre à hauteur de
poitrine (DHP) et de la hauteur totale (H). Au total 1189 arbres ont
été inventoriés et les résultats obtenus ont
montré une densité moyenne de 1707 tiges/ha. Par la suite, la
biomasse aérienne a été quantifiée en utilisant
deux modèles d'équation allométriques, celui de
Traoré et al. (2018) considérant les paramètres (DHP et H)
et celui de Chave et al. (2005) considérant en plus la densité du
bois sèche au four (ñ). Une comparaison a été faite
à partir des valeurs obtenues pour ces différents modèles
et le choix a été porté sur le modèle
développé par Chave et al. (2005). Et l'équation de Cairns
et al. (1997) a ensuite été utilisée pour
déterminer la biomasse souterraine. Les résultats obtenus sont
respectivement 50.54#177;6.39 t/ha, 11.45#177;1.35 t/ha et 61.98#177;7.73 t/ha
pour la biomasse aérienne, souterraine et totale. Pour finir, le stock
de carbone total a été évalué en appliquant une
fraction carbone recommandé par GIEC, (2006) valant 47 % de la biomasse
totale. Le résultat obtenu est 29.13#177;3.63 t/ha dont a
été déduite la quantité de CO2 estimée
à 106.82#177;16.32 t/ha. L'hypothèse de l'étude affirmant
que le peuplement de Bois Nago stocke moins de carbone que le peuplement
d'Acacia mangium de 7 ans étudié par Traoré et
al. (2018) est vérifiée. Toutefois, les résultats
quantitatifs observés sur ce jeune peuplement, n'ayant
bénéficié d'aucun traitement sylvicole, permettent
d'espérer une bonne performance de cette espèce à Bois
Nago.
iv
TABLE DES MATIÈRES
LISTE DES TABLEAUX vii
LISTE DES FIGURES viii
LISTE DES SIGLES ET ACRONYMES ix
LISTE DES ANNEXES x
CHAPITRE I : INTRODUCTION 1
1.1 Problématique 1
1.2 Objectifs 3
1.2.1 Objectif général 3
1.2.2 Objectifs spécifiques 3
1.3 Hypothèse 3
1.4 Limites de l'étude 3
CHAPITRE II : REVUE DE LA LITTERATURE 4
2.1 Généralités sur l'Acacia mangium
4
2.1.1 Caractéristiques de l'espèce 4
2.1.2 Ecologie et climat 4
2.1.3 Conditions édaphiques 4
2.2 Estimation de la biomasse des arbres 5
2.2.1 Méthode d'estimation de la biomasse des arbres
vivants 6
2.2.2 Méthodes d'estimation de la biomasse des bois
morts 7
2.2.3 Estimation à partir des équations
allométriques 7
2.3 Méthode d'évaluation du stock de carbone
9
CHAPITRE III : MÉTHODOLOGIE 11
3.1 Présentation de la zone d'étude 11
3.1.1 Localisation 11
3.1.2 Pluviométrie 12
3.1.3 Température 14
3.1.4 Topographie et sol 14
v
5.1 Conclusion 35
3.1.5 Végétation et faune 14
3.1.6 Mode de gestion actuelle 14
3.2 Matériels et méthodes 15
3.2.1 Matériels utilisés 15
3.2.2 Méthode de collecte des données 15
3.2.2.1 Détermination de la superficie du site 15
3.2.2.2 Inventaire des arbres 15
3.2.2.2.1 Pré inventaire 15
3.2.2.2.2 Plan d'échantillonnage 16
3.2.2.3 Réalisation de l'inventaire 17
3.2.2.4 Calcul de la biomasse aérienne et souterraine
des arbres 19
3.2.2.4.1 Calcul de la biomasse aérienne (AGB) 19
3.2.2.4.2 Calcul de la biomasse souterraine (BGB) 21
3.2.2.4.3 Calcul de la biomasse totale (TB) 21
3.2.2.5 Évaluation du stock de carbone et de la
quantité de CO2 équivalente 21
CHAPITRE IV : RÉSULTATS ET DISCUSSION 22
4.1 Inventaire du peuplement 22
4.1.1 Densité 22
4.1.2 Diamètre à hauteur de poitrine 23
4.1.3 Surface terrière 25
4.1.4 Hauteur totale 26
4.2 Biomasse 27
4.2.1 Biomasse aérienne avec l'équation de Chave
et al. (2005) 27
4.2.2 Biomasse aérienne avec l'équation de
Traoré et al. (2018) 28
4.2.3 Comparaison des résultats de la biomasse
aérienne 29
4.2.4 Biomasse souterraine 30
4.2.5 Biomasse totale 31
4.2.6 Relation entre la biomasse totale et la densité
31
4.3 Stock de carbone et quantité de CO2
équivalente 32
CHAPITRE V : CONCLUSION ET RECOMMANDATIONS 35
vi
5.2 Recommandations 36
RÉFÉRENCES BIBLIOGRAPHIQUES 37
ANNEXES 44
vii
LISTE DES TABLEAUX
Tableau 1: Croissance de certains peuplements
d'Acacia mangium en fonction de
l'âge et de la densité 5
Tableau 2:
Valeurs de l'estimation de la biomasse épigée de
certains peuplements
d'Acacia mangium situés en Indonésie par rapport
à l'âge 6
Tableau 3: Équations
allométriques pour l'estimation de la biomasse des arbres
d'Acacia mangium. 8
Tableau 4:
Équations allométriques pour l'estimation de la biomasse
aérienne des
arbres en forêts tropicales humides 9
Tableau 5:
Stock de carbone évalué pour certains peuplements
d'Acacia mangium
en fonction de l'âge et de la densité
10
Tableau 6: Densité du peuplement d'Acacia mangium
de Bois Nago à l'hectare 23
Tableau 7: DHP du peuplement d'Acacia mangium de
Bois Nago 24
Tableau 8: Surface terrière du peuplement
26
Tableau 9: Hauteur totale du peuplement 27
Tableau 10: Biomasse aérienne du
peuplement calculée à partir de l'équation de
Chave et al. (2005) 28
Tableau 11: Biomasse
aérienne du peuplement calculée à partir de
l'équation de
Traoré et al (2018) 29
Tableau 12: Biomasse souterraine du peuplement
30
Tableau 13: Biomasse totale du peuplement 31
Tableau 15: Stock de carbone du peuplement 32
Tableau 16: Quantité de CO2
équivalente du peuplement 34
viii
LISTE DES FIGURES
Figure 1: Vue aérienne de la parcelle
d'étude 11
Figure 2: Carte de localisation de la zone
d'étude 12
Figure 3: Pluviométrie moyenne mensuelle
de Ouanaminthe (1988-2019) 13
Figure 4: Variations annuelles de la
pluviométrie totale de Ouanaminthe (1988-
2019) 13
Figure 5: Distribution des placettes
d'échantillonnage 17
Figure 6: Carte présentant la
densité du peuplement de Bois Nago 22
Figure 7: Histogramme montrant le nombre de
tiges par classe de diamètre 24
Figure 8: Histogramme présentant la
surface terrière par classe de diamètre 25
Figure 9: Boite à moustache
présentant la distribution de la hauteur totale du
peuplement 26
Figure 10 : Diagramme de
corrélation entre la biomasse et la densité du
peuplement 31
ix
LISTE DES SIGLES ET ACRONYMES
AGB : Above Ground Biomass (Biomasse aérienne)
ANEVERT : Action Noble pour un Environnement Vert
BEF : Facteurs d'expansion de la biomasse
BGB : Below Ground Biomass (Biomasse souterraine)
CCNUCC : Convention Cadre des Nations Unies sur le Changement
Climatique
CGAAER : Conseil Général de l'Alimentation, de
l'Agriculture et des
Espaces Ruraux
CIRAD : Centre Internationale de Recherche Agronomique pour
le
Développement
CO2 : Dioxyde de carbone
CPDN : Contribution prévue déterminée au
niveau national
DHP : Diamètre à Hauteur de Poitrine
FAO : Food and Agriculture Organization
GES : Gaz à Effet de Serre
GIEC : Groupe d'experts Intergouvernemental sur l'Evolution du
Climat
IGN : Institut de l'information geographique et foresterie
MDE : Ministère De l'Environnement
PNUD : Programme des Nations Unies pour le
Développement
REDD+ : Réduction des émissions dues à la
déforestation et à la
dégradation des forêts
SIG : Système d'Information Géographique
USAID : United States Agency for International Development
WHH : Welthungerhilfe (Agro Action Allemande)
x
LISTE DES ANNEXES
Annexe 1: Coordonnées des centres des
placettes
Annexe 2: Précipitation moyenne
mensuelle de Ouanaminthe pour la période
(1988-2019)
Annexe 3: Précipitation totale
annuelle de Ouanaminthe pour la période (1988-
2019)
Annexe 4: Classification des zones
bioclimatiques de Holdridge
Annexe 5: Paramètres
dendrométriques comptés, mesurés et calculés par
placette
Annexe 6: Biomasse ( aérienne,
souterraine et totale) des arbres échantillonnés par
placette
Annexe 7: Fiche de collecte des
données
Annexe 8: Quelques prises de vue du site
1
CHAPITRE I : INTRODUCTION
1.1 Problématique
L'existence du changement climatique ne fait malheureusement
aucun doute; c'est un phénomène d'actualité mondiale. Sa
gravité est largement démontrée dans le 5ième
rapport du Groupe d'experts intergouvernemental sur l'évolution du
climat (GIEC) publié en 2014. Elle se traduit par une
élévation de la température moyenne du globe à
cause de la hausse des concentrations de gaz à effet de serre dans
l'atmosphère, particulièrement le dioxyde de carbone (GIEC,
2007). Cependant, ce dernier peut être absorbé grâce au
mécanisme de la photosynthèse des végétaux
composant les forêts, qu'elles soient d'origine naturelle ou anthropique.
Elles se comportent alors comme des puits capables de stocker du carbone
(Madignier et al., 2015). D'ailleurs, l'ensemble des forêts mondiales
représenterait selon la FAO (1997), 80 % des stocks de carbone au-dessus
du sol, et 40 % du carbone incorporé dans le sol. De même, la
déforestation peut-être une source d'émission de CO2 dans
l'atmosphère. Les scientifiques du GIEC (2007) estiment que les
émissions annuelles mondiales de CO2 dues à la
déforestation tropicale sont comprises entre 5 et 25 %.
Ce phénomène, à savoir la
déforestation s'accroit de plus en plus en Haïti et devient de nos
jours très préoccupant pour le pays. D'après Singh et
Cohen (2014), Haïti est l'un des pays les plus déboisés au
monde et selon une classification, faite par Churches et al (2014) seulement
32,3% de la superficie totale du pays était couverte d'arbres en
2010-2011. Le Ministère de l'Environnement dans sa deuxième
communication sur les changements climatiques publiée en 2013, avait
clairement affirmé que les forêts étaient en voie de
disparition. Mais le souci est que le pays est exposé à plusieurs
menaces naturelles liées directement ou indirectement à la
déforestation dont les plus courants sont les cyclones et les
inondations (MDE, 2006). Et même, Haïti subit de plus en plus les
conséquences de la déforestation tel que la dégradation du
sol, l'érosion et la diminution des ressources en eau (Singh et Cohen,
2014). Ces conséquences ne sont nulles autres que des effets du
changement climatique et Haïti à travers divers accords
multilatéraux sur l'environnement, reconnaît l'importance du
secteur forestier
2
comme moyen de les atténuer. Le pays a signé et
ratifié la Convention Cadre des Nations Unies sur le Changement
Climatique (CCNUCC) respectivement en juin 1992 et en septembre 1996, mais
aussi l'Accord de Paris (COP 21), qui fut signé en avril 2016 et
ratifié en juillet 2017 (PNUD, 2016). Dans le but d'apporter sa
contribution dans cette lutte mondiale de réchauffement
planétaire, Haïti s'est engagé lors de sa participation
à l'Accord de Paris en 2015, à réduire de 31 % ses
émissions de gaz à effet de serre à l'horizon 2030. A cet
effet, plusieurs mesures d'atténuation consistant entre autres à
augmenter son stock de carbone ont été prévues, parmi
lesquelles la plantation de 137.500 ha de forêt d'ici à 2030 en
privilégiant les espèces locales dont 100.000 ha de
manière conditionnelle entre 2020 et 2030 (MDE, 2015). Afin de respecter
ses engagements, il est important pour Haïti de réaliser ses
inventaires forestiers pour quantifier les stocks de carbone de ses boisements
ou ses reboisements. Cependant dans le pays, très peu de travaux
d'inventaire forestier ont été réalisés
jusqu'à date. De plus, plusieurs organisations nationales ou
internationales incluent dans leurs projets des activités de reboisement
sans effectuer pour autant des inventaires et procéder à
l'évaluation de leur stock de carbone.
Dans le cadre de son projet sur la sécurité
alimentaire, la Welthungerhilfe (WHH) avait entrepris des activités de
reboisement particulièrement dans le département du Nord-Est du
pays. La présente étude consiste alors à évaluer le
stock de carbone d'un peuplement d'Acacia mangium de l'une des
parcelles boisées située à Bois Nago. Ce travail
consistant à déterminer à la fois la biomasse
aérienne et la biomasse souterraine pour quantifier le stock de carbone,
permettra d'évaluer l'efficacité des peuplements d'Acacia
mangium quant à l'atténuation du changement climatique.
Quelle est la quantité de biomasse stockée dans le peuplement
d'Acacia mangium de Bois Nago ? Quel est le stock de carbone qui peut
en être déduite ? Ce sont là, les questions auxquelles ce
présent travail tente d'apporter des éléments de
réponse qui contribueront à l'acquisition de nouvelles
connaissances relatives au secteur forestier.
3
1.2 Objectifs
1.2.1 Objectif général
Cette étude envisage d'évaluer la
capacité de stockage de carbone d'un peuplement d'Acacia mangium
en vue de comprendre l'importance de ces plantations quant à
l'atténuation des effets du changement climatique.
1.2.2 Objectifs spécifiques
y' Faire l'inventaire par échantillonnage d'un petit
peuplement monospécifique d'Acacia mangium ;
y' Quantifier la biomasse aérienne et souterraine des
arbres de toute la parcelle ;
y' Évaluer le stock de carbone des arbres et la
quantité de CO2 équivalente.
1.3 Hypothèse
Le peuplement d'Acacia mangium de Bois Nago
âgé de six (6) ans et deux (2) mois, dont la densité est de
1707 tiges/ha stocke moins de carbone que celui de Côte d'Ivoire
étudié par Traoré et al. (2018) âgé de 7 ans,
et dont la densité est de 715 tiges/ha.
1.4 Limites de l'étude
Les limites de cette étude consistent tout d'abord au
fait que seulement le stock de carbone de la biomasse des arbres d'Acacia
mangium de la parcelle boisée à Bois Nago ont
été évalués. Ensuite, des modèles
d'équations allométriques généralisées et
spécifiques à l'espèce d'Acacia mangium et non au
site de l'étude ont été utilisés.
4
CHAPITRE II : REVUE DE LA LITTERATURE
2.1 Généralités sur l'Acacia
mangium
2.1.1 Caractéristiques de
l'espèce
L'Acacia mangium est un grand arbre originaire de la
Papouasie Nouvelle-Guinée, de l'Indonésie et de l'Australie qui
peut atteindre jusqu'à 30 m de haut. Cependant dans des conditions peu
favorables c'est un petit arbre, parfois avec des tiges multiples et
buissonnantes. Son fût est long et droit certaines fois
légèrement cannelé à la base. L'écorce est
claire, gris-brunâtre ou brune, rugueuse avec des fissures
longitudinales. Sur le tronc, les branches persistent longuement,
l'élagage naturel est donc mauvais. Le système racinaire est
vigoureux et traçant (CIRAD, 2003). Caractérisé par un
bois léger de densité égale à 0.6 g/cm3
(FAO, 1979) ; l'Acacia mangium est exploitable entre 6 et 9 ans. Sa
une durée de vie est relativement courte car elle ne dépasse pas
une trentaine d'années dans son aire d'origine (Pinyopusarerk et al.,
1993).
2.1.2 Ecologie et climat
L'Acacia mangium est une espèce des
forêts claires et des savanes boisées, partiellement liée
au passage des feux. En général, on la trouve dans les
régions côtières, surtout en dessous de 300 m d'altitude,
au maximum à 800 m. Exigeante en lumière, c'est une espèce
à croissance rapide qui particulièrement, ne présente pas
d'arrêt de croissance quand les conditions sont adéquates. Formant
des associations symbiotiques avec des Rhizobiums, des
Bradyrhizobium, des ecto et endomycorhizes, elle la capacité de
croître sur des sols particulièrement pauvres (CIRAD, 2003).
L'Acacia mangium peut supporter des conditions climatiques très
diverses : des précipitations d'été ou uniformément
réparties, de 1000 à 4000 mm avec une saison sèche de 0
à 6 mois. La température moyenne du mois le plus chaud qu'elle
tolère est d'environ 30-34oC et la moyenne du mois le plus
frais 15-22oC (Mergen et al., 1983).
2.1.3 Conditions édaphiques
L'Acacia mangium est capable de se produire
derrière les mangroves, dans les marécages saisonniers, le long
des ruisseaux et sur les plateaux bien drainés, les
5
crêtes basses et les collines de montagne (Pinyopusarerk
et al., 1993). A la différence des autres légumineuses qui
nécessitent un sol de pH supérieur à 5.5, l'Acacia
mangium est apte à se développer sur des sols acides de pH
4.5-6.5 pauvres, légers et bien drainés en surface (CIRAD,
2003).
2.1.4 Croissance
L'Acacia mangium a une forte croissance initiale ;
dès l'âge de 3 ans elle peut déjà atteindre 5
à 6 m de hauteur et une quinzaine de mètres à 15 ans en
conditions moyennes. A partir de 3 ans, l'accroissement en circonférence
est influencé par la densité pour les plantations les plus
serrées. Après 8 ans, sa croissance diminue aussi rapidement
(Dupuy et N'Guessan, 1990). Le tableau suivant présente
différentes valeurs de croissance en hauteur et en diamètre des
arbres d'Acacia mangium des peuplements de différents
âges à partir des études réalisées en
Indonésie et en Côte d'Ivoire en fonction de l'âge et de la
densité
Tableau 1: Croissance de certains peuplements d'Acacia
mangium en fonction de l'âge et de la densité
|
Références
|
Age (ans)
|
Densité (arbres/ha)
|
DHP moyen Hauteur
(cm) (m)
|
Surface terrière (m2/ha)
|
|
Hériansyah et al., 2007
|
3
|
1838
|
7.25
|
7.33
|
8.78
|
|
5
|
758
|
17.89
|
17.41
|
20.12
|
|
Traoré et al., 2018
|
3
|
845
|
12.5
|
12.5
|
10.42
|
|
7
|
715
|
23.4
|
19.0
|
30.75
|
|
Ilyas, 2013
|
3
|
1100
|
12.73
|
5
|
14.40
|
|
5
|
1100
|
15.28
|
6
|
21.54
|
|
7
|
1100
|
24.19
|
8
|
48.54
|
2.2 Estimation de la biomasse des arbres
Selon Brown (2002), deux composantes sont à
considérer afin d'estimer la biomasse des arbres. Tout d'abord, la
composante aérienne constituée de feuilles, de branches et de
tiges d'où la biomasse aérienne ; ensuite le sous-sol comprenant
les racines d'où la biomasse souterraine. La biomasse de certains
peuplements
6
d'Acacia mangium a déjà
été évaluée lors de précédents
travaux réalisés sur différents sites d'étude en
Indonésie. Le tableau suivant présente les valeurs
trouvées en fonction de l'âge.
Tableau 2: Valeurs de l'estimation de la biomasse
épigée de certains peuplements d'Acacia mangium
situés en Indonésie par rapport à l'âge
Age (ans) Biomasse (t/ha)
Références
1 17.2 Hardiyanto et al., 2004
2 46.1 Hardiyanto et al., 2004
3 94.25 Ilyas, 2013
5 137.85 Ilyas, 2013
7 292.02 Ilyas, 2013
7 177.08 Ilyas, 2013
7 63.46 Hériansyah et al.,
2007
2.2.1 Méthode d'estimation de la biomasse des
arbres vivants
L'estimation de la biomasse des arbres peut se faire par des
méthodes directes et indirectes (Brown, 2002). Les méthodes
directes consistent à récolter l'arbre et à prendre des
mesures de la masse réelle des différents compartiments (troncs,
branches, feuilles, racines). Ces méthodes sont destructives (Kangas et
Maltamo, 2006). Les méthodes indirectes utilisent des modèles de
biomasse et des facteurs d'expansion de la biomasse (BEF) pour exprimer la
biomasse des arbres, elles sont efficaces dans le temps (Peltier et al.,
2007).
Des deux composantes (aérienne et souterraine) de la
biomasse des arbres, l'estimation de la biomasse aérienne (AGB) suscite
beaucoup d'intérêt ces dernières années et est
devenu un sujet de recherche de grande importance (Zolkos et al., 2013). Deux
méthodes indirectes d'évaluation de la biomasse sont
préconisées. Tout d'abord, les méthodes se basant sur des
techniques de télédétection et adaptées pour la
mesure et le suivi des forêts sur de grandes études spatiales
(Picard et al., 2012). Ensuite, les méthodes fondées sur
l'utilisation des équations allométriques en appliquant des
données d'inventaire. Etant donné que
7
cette étude se réalise sur une petite parcelle
et que cette dernière méthode est moins coûteuse, peu
exigeante et de bonne précision, c'est elle qui est appliquée.
Quant à la deuxième composante, à savoir la biomasse
souterraine (BGB) ; elle est considérée comme étant le
réservoir de carbone le plus difficile à évaluer.
Cependant, elle peut être estimée à partir de la biomasse
aérienne plus facilement mesurable en utilisant le ratio racine/tige
(R/S) : AGB = AGB X R/S (Nasi et al., 2008). De même, plusieurs
équations allométriques applicables à différents
types de forêt testées et validées dans des publications
scientifiques, ont été développées pour estimer la
biomasse souterraine à partir de la biomasse aérienne.
2.2.2 Méthodes d'estimation de la biomasse des
bois morts
Le bois mort peut représenter jusqu'à 40 % de la
biomasse aérienne totale des systèmes forestiers tropicaux
(CIRAD, 2011). Pearson et Brown (2005), définit quatre catégories
de bois morts : la première catégorie est celle des arbres avec
les branches et les rameaux ressemblant à un arbre vivant (sauf pour le
feuillage), la deuxième comprend les arbres sans rameaux mais avec
encore des branches grandes et petites, la troisième catégorie
concerne les arbres avec seulement de grandes branches et la quatrième
et dernière comprend le tronc seulement, sans les branches. Pour cette
dernière catégorie, la hauteur de l'arbre et son diamètre
au niveau du sol sont à mesurer et il faut estimer le diamètre de
la cime. Les bois morts du peuplement de Bois Nago appartiennent à la
première catégorie. Alors, les équations
allométriques générales peuvent être
utilisées pour calculer la biomasse des arbres morts qui ont encore des
branches et des rameaux. La quantité de biomasse dans un arbre mort
encore debout peut être similaire à celle d'un arbre vivant.
Cependant, un facteur de désintégration du bois mort doit
être utilisé, par exemple 0,1 (ou 10 %) parce que le bois mort qui
se décompose au sol est moins dense que le bois de l'arbre encore debout
(Mcghee et al., 2016).
2.2.3 Estimation à partir des équations
allométriques
Les équations allométriques sont des relations
qui relient la biomasse à une ou deux variables indépendantes
telles que le diamètre à hauteur de poitrine et la hauteur des
arbres (Lotfi, 2008). Beaucoup d'auteurs ont proposé des
modèles
8
allométriques sur la base d'une, deux ou trois
variables indépendantes: 1) le DHP uniquement; 2) le DHP et la
densité du bois, et 3) le DHP, la densité du bois et la hauteur
totale (Dorvil, 2010). Certains modèles cités dans
différentes publications scientifiques ont été
développées pour estimer la biomasse aérienne des arbres
d'Acacia mangium spécifiquement (tableau 3). D'autres plus
généralisées permettent également de calculer la
biomasse aérienne mais ont été développés en
fonction de la zone agroécologique (tableau 4). Les modèles
présentés dans le tableau ci-dessous ont été
développés afin d'estimer la biomasse aérienne des arbres
d'Acacia mangium spécifiquement. D'après les
modèles du tableau ci-dessous la biomasse aérienne peut
être calculée par la sommation de la biomasse des tiges, des
branches et des feuilles.
Tableau 3: Équations allométriques pour
l'estimation de la biomasse des arbres d'Acacia mangium.
Équations Références
3.0294
Btiges = 0.0116D Hardiyanto et Wicaksono,
2008
2.21179
Bbranches = 0.0712D Hardiyanto et Wicaksono,
2008
0.515
Bfeuilles = 2.1195D Hardiyanto et Wicaksono,
2008
Btiges=exp(-3.228+1.681×ln(??)+1.056×ln(??))
Traoré et al., 2018
Bbranches=exp(-0.865+0.498×ln(??2×??))
Traoré et al., 2018
Bfeuilles=exp(-0.882+1.339×ln(??))
Traoré et al., 2018
Btiges=0.0193 (D2H) 0.9934 Hériansyah et al., 2007
Bbranches=0.0193(D2H)0.9934
Hériansyah et al., 2007
Bfeuilles=0.0245(D2H)0.6173
Hériansyah et al., 2007
Btiges = biomasse tiges, Bbranches = biomasse branches,
Bfeuilles = Biomasse feuilles, D = diamètre à hauteur de
poitrine, H = hauteur totale.
Ces équations ont été
développées dans des peuplements situés en zone de climat
tropical, notamment en Côte d'Ivoire et en Indonésie. Elles
permettent d'estimer la biomasse aérienne des arbres d'Acacia
mangium. Les équations allométriques de Traoré et al.
(2018) ont permis d'estimer la biomasse des différentes composantes
9
aériennes (troncs, branches, feuilles)
séparément. Elles ont été développées
en Côte d'Ivoire, pour des peuplements d'Acacia mangium;
situés dans une zone de forêt tropicale, plantés sur des
sols ferrugineux à basse altitude (40 à 100 m) et recevant une
précipitation moyenne de 1766 mm Les peuplements d'Acacia mangium
étudiés par Hardiyanto et Wicaksono (2008) et
Hériansyah et al. (2007) reçoivent annuellement plus de 2000 mm
et sont situés à moins de 200 m d'altitude. Le tableau suivant
présente plusieurs modèles d'équation allométrique
généralisées développées en fonction de la
zone agroécologique et qui permettent de calculer la biomasse
aérienne des peuplements forestiers.
Tableau 4: Équations allométriques pour
l'estimation de la biomasse aérienne des arbres en forêts
tropicales humides
Equations Références
AGB=exp(-2.557 +0.940ln(pD2 H??0.0776x(pD2
H)0.940 Chave et al., 2005
AGB =1.276 + 0.034× (D2 H ) Brown et yverson,
1992
AGB = Exp (-2.134× 2,53 ln (D)) Brown, 1997
AGB = 38,4908 - 11,7883 (D) + 1,1926 (D2) Brown et al.,
1989
AGB = 42,69-12,8D + 1,242D2 FAO, 1997
Source : Dorvil, 2010
Selon Ketterings et al. (2001) cité par Dorvil (2010),
la plupart des modèles ont été développés en
utilisant des données provenant de zones équatoriales. Ces
équations allométriques reposent sur le DHP et la hauteur totale
mais aussi sur la densité du bois (Chave et al., 2005). Cairns et al.
(1997) ont développé une équation allométrique
permettant d'estimer la biomasse souterraine en forêt tropicale à
partir de la biomasse aérienne: BGB = 0.347 ×
(AGB)0.884.
2.3 Méthode d'évaluation du stock de
carbone
Le bois sec contient environ 50 % de carbone; cela signifie
que près de 25% d'un arbre vivant est composé de carbone, comme
la fraction couramment appliquée par
10
défaut est de 0,47 entre la biomasse sèche et la
biomasse vivante (GIEC, 2006). Ainsi, le carbone stocké dans les plantes
ligneuses peut être estimé par la relation:
Cw=k×Bw, avec Cw, le stock de carbone, Bw la
biomasse exprimée en matière sèche par unité de
superficie en (t/ha) et k0,47 (Nasi et al., 2008).
Quand le bois est brûlé, il réagit avec
l'oxygène de l'air et produit du dioxyde de carbone, de l'eau, de la
chaleur et de la lumière. A partir du tableau périodique des
éléments la masse atomique du carbone (C) est de 12 et celle de
l'oxygène (O) est de 16. La molécule du dioxyde de carbone (CO2)
a un atome de carbone (C) et deux atomes d'oxygène (O). La masse
atomique du dioxyde de carbone (CO2) est la somme des masses atomiques de ces
atomes (44). Le rapport de la masse atomique du dioxyde de carbone à
celle du carbone est 44/12, qui est sensiblement égal à 3.667.
Pour convertir la quantité de carbone en dioxyde de carbone. Il faut
multiplier la quantité de carbone par 3.667 ou 44/12.
Les études concernant l'évaluation des stocks de
carbone des peuplements forestiers sont très nombreuses de nos jours. Le
tableau suivant présente les valeurs des stocks de carbone de quelques
peuplements d'Acacia mangium en fonction de l'âge et de la
densité.
Tableau 5: Stock de carbone évalué pour certains
peuplements d'Acacia mangium en fonction de l'âge et de la
densité
|
Age
|
Densité
|
Stock de carbone
|
Références
|
|
Ans
|
arbres/ha
|
t/ha
|
|
|
1
|
1050
|
0.22
|
Herdiyanti et Sulistyawati, 2009
|
|
3
|
889
|
12.45
|
Herdiyanti et Sulistyawati, 2009
|
|
3
|
845
|
27.79
|
Traoré et al., 2018
|
|
5
|
839
|
28.59
|
Herdiyanti et Sulistyawati, 2009
|
|
7
|
728
|
31.48
|
Herdiyanti et Sulistyawati, 2009
|
|
7
|
715
|
86.33
|
Traoré et al., 2018
|
|
11
|
553
|
105.00
|
Traoré et al., 2018
|
11
CHAPITRE III : MÉTHODOLOGIE
3.1 Présentation de la zone
d'étude
Malgré le fait que l'altitude optimale de
développement de l'Acacia mangium est en-dessous de 300 m
(CIRAD, 2003), le peuplement étudié a été
installé depuis les mois de janvier et février de l'année
2014 en montagne à une altitude variant de 687 m à 889 m.
D'après la classification de Holdridge axée sur la zone de vie,
le site de l'étude correspond à une zone d'agriculture de
montagne humide.

Source : Google earth
Figure 1: Vue aérienne de la parcelle d'étude
3.1.1 Localisation
Le peuplement étudié est situé à
Mont-Organisé dans une localité appelée Bois Nago. Les
coordonnées géographiques du site sont comprises entre
19°24' de latitude Nord et 71°48' de longitude Ouest. La figure 2,
réalisée en fonction de la classification de Holdridge, permet
d'observer la localisation de la parcelle sur la carte d'Haïti.
12

Figure 2: Carte de localisation de la zone d'étude
3.1.2 Pluviométrie
Faute de station agro-climatologique fonctionnelle dans la
zone, les données pluviométriques recueillies sont celles
collectées par la station la plus proche du Centre Chrétien de
Ouanaminthe, située à une distance de 19 km de la zone
d'étude. Les figures suivantes présentent la pluviométrie
moyenne par mois (figure 3) et la pluviométrie totale par année
(figure 4) enregistrées à Ouanaminthe durant la période
1988-2019.
13
140
120
Pluviométrie (mm]
100
80
60
40
20
0

Pluviométrie (mm]
3500
3000
2500
2000
1500
1000
500
0
Figure 4: Variations annuelles de la pluviométrie
totale de Ouanaminthe (19882019)
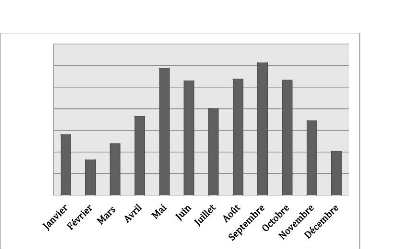
Figure 3: Pluviométrie moyenne mensuelle de Ouanaminthe
(1988-2019)
Les mois de septembre et de mai sont les plus pluvieux avec le
maximum de précipitations moyennes (122.4 mm) enregistré au mois
septembre. Le mois de février est le moins pluvieux (32.7 mm de pluie).
A Ouanaminthe, la saison estivale est beaucoup plus arrosée
comparativement à la saison hivernale.
14
L'évolution de la pluviométrie s'est traduite
par des écarts élevés entre les années ; le maximum
(2881.2 mm) de pluie a été enregistré en 2007 et le
minimum (815.3 mm) en 1991. La courbe tendance traduit une augmentation des
précipitations avec le temps.
3.1.3 Température
Pour la même raison évoquée dans la
section précédente, les données climatiques de
température, pour la commune de Mont-Organisé sont presque
inexistantes. Il a été estimé qu'elle varie de 15 à
30°C sur toute l'année et peut descendre jusqu'à 12°C
en décembre (Valbrun, 2005).
3.1.4 Topographie et sol
Le site de l'étude est une montagne avec un relief
très élevé et dont les pentes varient de 42 à 80 %
avec une moyenne de 60 %. Les sols de Mont-Organisé sont majoritairement
rouges à dominance argileuse, constitués de l'évolution de
roches éruptives principalement basaltiques (Valbrun, 2005). Ce sont des
sols fersialitiques assez perméables dont la profondeur corrèle
avec les différentes classes de pentes. Ainsi pour les terrains à
forte déclivité comme c'est le cas pour le site de
l'étude, la profondeur est faible. Ce sont des sols très
sensibles à l'érosion.
3.1.5 Végétation et faune
Au niveau de la parcelle la végétation naturelle
est constituée de deux strates : la strate arborée
constituée d'arbre forestier énergétique : l'Acacia
mangium et la strate herbacée composée de plantes comme la
citronnelle (Cymbopogon citratus), le zèb guinen (Panicum
maximum). Quant à la faune, une observation directe a permis de
constater la présence de pintade marron (Numida meleagris), de
guêpe (Vespula vulgaris), de reptiles tel l'anolis et le
mabouillard (Ameiva chrysoloema).
3.1.6 Mode de gestion actuelle
La plantation étudiée est gérée
par une jeune association nommée ANEVERT (Action Noble pour un
Environnement Vert) chargée de prendre toutes les mesures relatives
à la protection des arbres du site. Dès le début de la
plantation, des
15
opérations de désherbage ont été
effectuées ainsi qu'un débroussaillement en amont de la parcelle.
Depuis, aucun traitement sylvicole n'a été appliqué
à la parcelle jusqu'à date. Aucun traitement n'a
été effectué sur les arbres (éclaircissage,
élagage,..) ou sur le site (amendement du sol,..), aucun
procédé de régénération (coupe progressive
régulière, coupe totale,...). Le site n'est pas
clôturé, cependant les animaux ne représentent pas une
menace au développement de la plantation.
3.2 Matériels et méthodes
3.2.1 Matériels utilisés
Les matériels utilisés pour réaliser ce
travail de mémoire sont les suivants :
V' GPS : pour la localisation des points sur le terrain
V' Appareil photographique : pour des prises de vue sur le
terrain
V' Compas forestier : pour la mesure du diamètre à
1,30 m des tiges
V' Ruban métrique : pour la mesure des distances et des
diamètres
V' Dendromètre SUUNTO : pour la mesure des angles
permettant de calculer
la hauteur des arbres et la pente du terrain
V' Fiche d'inventaire : pour la saisie des données sur le
terrain
3.2.2 Méthode de collecte des
données
Dans le cadre de cette étude, il a fallu tout d'abord
déterminer la superficie de la
parcelle. Ensuite les données qui ont été
collectées sont celles relatives à
l'inventaire ; à savoir le nombre de tiges, leur DHP et
les angles qui ont permis de
calculer la hauteur totale.
3.2.2.1 Détermination de la superficie du
site
La superficie du site a été calculée
à l'aide d'un GPS calibré de façon à enregistrer
les coordonnées du périmètre toutes les 30 secondes. Elle
a été estimée à 7,15 ha soit 71500 m2
3.2.2.2 Inventaire des arbres
3.2.2.2.1 Pré inventaire
Le pré-inventaire a été
réalisé dans la parcelle deux ans après la plantation des
arbres d'Acacia mangium. Un total de 23 arbres répartis dans 3
placettes, de 25 m2
16
chacune a été compté et mesuré. Le
coefficient de variation pour le nombre d'arbres inventoriés est
estimé à 7.44 %.
3.2.2.2.2 Plan d'échantillonnage
Pour cet inventaire on a priorisé un type
d'échantillonnage qui a permis de couvrir toute la parcelle
forestière. Et d'après une visite de reconnaissance du site,
aucune hétérogénéité n'a été
constatée. Ainsi, le choix a été porté pour un
échantillonnage systématique aléatoire car elle permet
d'avoir une meilleure estimation des variables et elle offre également
une certaine sécurité des estimations à cause de
l'uniformité de la parcelle.
V' Forme et dimension des placettes
:
Nous avons opté pour des placettes de forme circulaire
parce que selon Lecomte et Rondeux (2002), cette forme est facile à
installer et elle ne présente aucune direction
préférentielle. Et aussi, elle possède le plus petit
périmètre à surface égale ce qui limite les risques
d'erreur. La dimension des placettes quant à elle, dépend de
l'exactitude, la précision, le temps et le coût de la mesure
(Pearson et Brown, 2005). Chacune des placettes d'échantillonnage avait
un rayon (r =8 m) et (r1= 7.92 m) en tenant compte du facteur correcteur de
pente. La superficie de chaque placette calculée en appliquant la
formule s = it X r X r1 est de 198.97
m2.
V' Taux d'échantillonnage :
Pour cette étude, l'effort d'échantillonnage de
l'inventaire a été fixé à 10 %. Tenant compte du
facteur correcteur de la pente, l'échantillon a couvert une superficie
de 6963.98 m2.
V' Nombres de placettes à
échantillonner :
Le nombre de placettes d'échantillonnage est de 35. Il
a été estimé à partir du taux de sondage (f), de la
superficie totale de la parcelle (S) et de la superficie à
échantillonner (s) en appliquant la formule : n= (f×S) / s.
V' Erreur d'échantillonnage :
Le calcul de l'erreur d'échantillonnage permet de
vérifier la validité de l'échantillon. Pour un niveau de
confiance de 95 %, elle a été estimée à 2.46 %
en
17
?? × ????
appliquant la formule : e = v?? , « t »
représente le t de student, « CV » le
coefficient de variation et « n » le nombre de
placettes d'échantillonnage.
V' Répartition des placettes
d'échantillonnage :
La figure suivante montre la répartition des placettes
suivant un échantillonnage aléatoire systématique. La
première placette a été choisie au hasard et les autres
sont placées de façon systématique à une distance
moyenne de 45 m l'une de l'autre. Les coordonnées des centres des
placettes sont mentionnées en annexe.
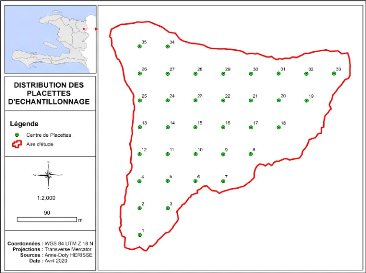
Figure 5: Distribution des placettes d'échantillonnage
3.2.2.3 Réalisation de l'inventaire
V' Équipe d'inventaire
L'équipe d'inventaire était constituée de
deux personnes, un compteur qui a implémenté les placettes et
mesuré les diamètres à hauteur de poitrines et un pointeur
qui a noté les mentions dictées par le compteur après les
avoir répétées à voix haute et qui a aussi
mesuré et noté les angles pour le calcul de la hauteur totale
ainsi que les distances entre l'arbre à mesurer et lui.
18
V' Diamètre de
précomptage
Les mesures se sont portées sur tous les arbres
d'Acacia mangium du site (âgés de 6 ans et 2 mois) dont
le DHP est supérieur ou égal à 5 cm. Les arbres vivants
comme les morts ont été mesurés.
V' Cheminement et localisation des
placettes
L'échantillonnage avait été
préalablement réalisé sur le logiciel ArcGis (version
10.2.1). Les coordonnées des centres des placettes
générées par le logiciel ont été
préalablement enregistrées dans la mémoire du
récepteur GPS. Pour localiser la première placette, il a suffi de
sélectionner le point dans le GPS, lorsque la distance était
nulle le centre était atteint. On a cheminé 45 m, toujours dans
la direction Sud-Nord afin de localiser la placette 2. Dans cette même
direction, on a parcouru toute la première rangée située
à l'ouest jusqu'à la placette 35. La placette 34 a ensuite
été localisé en cheminant la même distance dans la
direction Ouest-Est. Une fois à la 2ème rangée,
on procède de la même manière jusqu'à la placette 3,
sauf que c'est dans la direction Nord-Sud. La 6ème placette a
ensuite été localisée en parcourant 63 m dans la direction
Sud-Ouest. On a recommencé les mêmes procédures et
cheminé ainsi de suite en suite jusqu'à localiser toutes les
placettes.
V' Implantation des placettes
Une fois au centre des placettes, le compteur tend le ruban
métrique au pointeur, ce dernier mesure la distance 8 m correspondant au
rayon de la placette et marque la limite avec un piquet enfoncé dans le
sol. Cette opération avait été effectuée quatre
fois à partir du centre de chaque placette dans des directions
différentes. Ensuite le compteur fait le tour de la placette en
maintenant la ficelle tendue à un des jalons qui délimitent la
placette ou à un arbre dans certains cas.
V' Mesure des arbres
Les différentes mesures des arbres prises dans le cadre
de cet inventaire sont celles du diamètre à hauteur de poitrine
et celles permettant de calculer la hauteur totale. Pour chaque arbre des
placettes le diamètre a été mesuré à 1.30 m
en utilisant le compas forestier gradué jusqu'à 50 cm, qui a
permis d'obtenir une mesure directe du diamètre des arbres.
19
Afin de déterminer la hauteur totale le
dendromètre SUUNTO est l'instrument qui a été
utilisé, cela a impliqué de mesurer des angles. Trois cas ont
été rencontrés sur le terrain et d'après Mille et
Louppe (2015), il faut faire dans ces cas une ou deux lectures en visant soit
le sommet ou à la fois le sommet et la base de l'arbre. AB
représente la hauteur des arbres à mesurer, h la hauteur de
l'oeil de l'observateur, d la distance de l'observateur à l'arbre et
á et f3 sont les angles obtenus en visant soit le sommet ou la base de
l'arbre. Suivant les trois cas rencontrés sur le terrain, les formules
trigonométriques suivantes doivent être appliquées selon
Mille et Louppe (2015) pour déterminer la hauteur totale.
1er cas : Le terrain est horizontal et l'oeil de l'observateur
se trouve entre le sommet et la base de l'arbre, la formule appliquée a
été: AB = h + d.tgá
2e cas : Le terrain est incliné et l'oeil de
l'observateur se trouve entre le sommet et la base de l'arbre, la formule
utilisée : AB = d (tgf3 + tgá)
3e cas : Le terrain est incliné et l'oeil de
l'observateur se trouve en dessous de la base de l'arbre, alors on applique la
formule : AB = d (tgá - tgf3)
? Traitement des cas litigieux
Les cas litigieux que nous avons traités concernent ;
tout d'abord les arbres se trouvant sur la limite de la placette, alors
l'inclusion dans la placette a été faite quand plus de 3/4 de son
tronc s'est trouvé à l'intérieur de celle-ci. Ensuite, le
cas des arbres fourchus au-dessous de 1,30 m ; alors les diamètres des
différentes fourches à 1,30 m ont été
totalisés. Pour les arbres avec contreforts à 1.30 m de hauteur,
on a considéré la moyenne des circonférences
mesurées à 10 cm directement au-dessus et en-dessous de
l'anomalie. Et enfin, le cas litigieux rencontré le plus souvent lors de
la collecte des données est celui des arbres situés sur une
pente. Dans ce cas, le diamètre à hauteur de poitrine (1.30 m)
des arbres a été compté en amont du pied de l'arbre.
3.2.2.4 Calcul de la biomasse aérienne et
souterraine des arbres
3.2.2.4.1 Calcul de la biomasse aérienne
(AGB)
? Arbres vivants
Afin de déterminer la biomasse aérienne des arbres
vivants, les modèles
allométriques de Traoré et al. (2018) et de Chave
et al. (2005) ont été utilisés. Les
20
modèles allométriques développés
par Traoré et al. (2018) sont les suivants et elles ont permis
l'estimation respective de la biomasse du tronc, des branches et des
feuilles:
B1= exp(-3.228+1.681×ln(D)+1.056×ln(H))
B2 = exp(-0.865+0.498×ln(D2×H))
B3 = exp(-0.882+1.339×ln(D))
La biomasse aérienne totale a été
déterminée en additionnant la biomasse des différentes
composantes : AGB = B1+ B2+B3.
L'équation allométrique de Chave et al. (2005) a
permis une estimation directe de la biomasse. Elle a tenu compte explicitement
de la densité spécifique du bois. Pour l'espèce Acacia
mangium, la densité du bois (0.6g/cm3) a
été obtenue dans une publication faite par la FAO en 1979,
donnant des informations sur les ressources forestières. Selon Dorvil
(2010), la densité du bois à 12 % d'humidité (D12) peut
être convertie en densité spécifique du bois
séché au four à partir d'une relation
élaborée par Chave et al. (2006) à travers des
données obtenues dans des forêts tropicales : ñ = 0.872 x
D12.
L'équation allométrique de Chave et al. (2005)
applicable en forêt tropicale utilisée dans le cadre de cette
étude est la suivante:
AGB = exp(-2.557 + 0.940ln(ñD2H)) ?
0.0776×(ñD2H)0.940.
Pour cette étude, les deux résultats obtenus
pour la biomasse aérienne ont été comparés afin
d'adopter le meilleur pouvant être utilisé pour le peuplement de
Bois Nago.
? Bois morts
Se basant sur la classification faite dans le guide de mesure
et de suivi du carbone élaboré par l'USAID en 2005, les bois
morts du site appartiennent à la première catégorie :
arbre avec branches et rameaux ressemblant à un arbre vivant (sauf pour
le feuillage). La même méthode utilisée pour
déterminer la biomasse des arbres vivants a donc été
appliquée pour les arbres morts (Pearson et Brown, 2005).
NB : La biomasse des feuilles n'a pas été
calculée pour les bois morts lorsqu'il a fallu appliquer le
modèle Traoré et al. (2018).
21
3.2.2.4.2 Calcul de la biomasse souterraine
(BGB)
Selon Cairns et al. (1997), la biomasse souterraine peut
être calculée en appliquant un modèle allométrique
après avoir estimé la biomasse aérienne des arbres
mesurés. La formule suivante de Cairns et al. (1997) applicable en
forêt tropicale a été utilisée afin de
déterminer la biomasse souterraine.
BGB = 0.347 × (AGB)0.884
3.2.2.4.3 Calcul de la biomasse totale (TB)
La biomasse totale a été évaluée
en additionnant la biomasse aérienne à la biomasse souterraine
:
TB = AGB + BGB
3.2.2.5 Évaluation du stock de carbone et de la
quantité de CO2 équivalente D'après les
recommandations faites dans les directives du GIEC (2006), la fraction de
carbone couramment appliquée par défaut est de 0.47. Pour
évaluer le stock de carbone, la fraction (0.47) a été
multipliée par la biomasse.
Selon le guide de quantification des émissions de gaz
à effet de serre élaboré en 2019 par le Ministère
de l'Environnement et de la lutte contre les Changements Climatiques (MELCC) du
Québec, la quantité de dioxyde carbone (CO2) équivalente
peut être évaluée en multipliant le stock de carbone (C)
par 44/12 qui est le résultat du rapport de la masse atomique du dioxyde
de carbone à celle du carbone. C'est la méthode qui a
été appliquée dans le cadre de cette étude afin de
de déduire le stock de CO2 de celui du carbone.
Tout en respectant les règles des inférences
statistiques tous les résultats obtenus à partir de
l'échantillon ont été généralisés
à toute la population d'où l'échantillon est
tiré.
22
CHAPITRE IV : RÉSULTATS ET DISCUSSION 4.1
Inventaire du peuplement
4.1.1 Densité
Les différentes placettes inventoriées à
Bois Nago ont un total variant de 15 à 85 tiges ce qui a donné
une moyenne de 34 tiges/placette. Majoritairement (77%), les placettes
d'échantillonnage comptent de 21 à 40 tiges. La distribution du
nombre de tiges des placettes est donc très
hétérogène. Afin de mieux apprécier la
densité du peuplement, la carte suivante représente par des
boules le nombre de tiges des différentes placettes. Ainsi les endroits
plus ou moins denses du peuplement sont localisés.
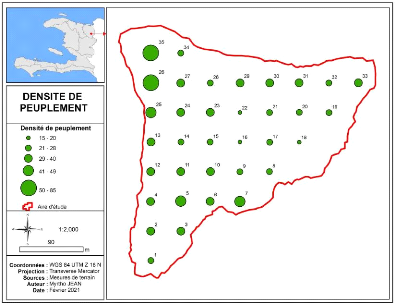
Figure 6: Carte présentant la densité du peuplement
de Bois Nago
Le peuplement est plus dense au Nord-Ouest à
l'extrémité de la parcelle d'où la pente est plus raide
tandis qu'à l'Est la densité est plus faible car le nombre de
tiges des placettes varie de 15 à 20 (figure 6). Le nombre de tiges de
chaque placette a été ramené à l'hectare (tableau
6), et le coefficient de variation ainsi que
23
les valeurs minimum et maximum de densité montrent que
la densité moyenne du peuplement n'est pas représentative de
celle de toutes les placettes. Le coefficient de variation des arbres morts est
plus élevé, cette distribution est donc très
hétérogène comparativement aux arbres vivants. Les arbres
morts ne représentent qu'1 % de la population, les résultats de
cette distribution n'influencent pas beaucoup la distribution totale comprenant
les deux catégories d'arbres. Le tableau suivant présente la
densité du peuplement à l'hectare :
Tableau 6: Densité du peuplement d'Acacia mangium
de Bois Nago à l'hectare
|
Densité
|
Effectif
|
Moyenne/ ha
|
Coefficient
de variation
|
Erreur-type
|
Min
|
Max
|
|
Arbres vivants
|
1180
|
1694
|
0.40
|
226
|
704
|
4272
|
|
Arbres morts
|
9
|
13
|
3.98
|
17
|
0
|
302
|
|
Tous
|
1189
|
1707
|
0.39
|
222
|
754
|
4272
|
Le peuplement est beaucoup plus dense que ceux qui ont
été mentionnés dans la littérature et dans
l'étude de l'hypothèse en se référant aux travaux
réalisés par Herdiyanti et Sulistyawati (2009), Hériansyah
et al. (2007), Ilyas (2013) et Traoré et al (2018) dans des peuplements
d'Acacia mangium âgé respectivement de 5, 5.5 et 7 ans.
Ces écarts s'expliquent par le fait que les objectifs de plantation
diffèrent de celui de Bois Nago qui a été principalement
de couvrir le sol préalablement dénudé. De plus, le
peuplement étudié n'a bénéficié d'aucun
traitement sylvicole à la différence de ceux de la
littérature.
4.1.2 Diamètre à hauteur de
poitrine
? Structure diamétrique du
peuplement
Les diamètres des tiges (arbres morts y compris)
varient de 5 cm à 29 cm et ont été regroupés en 6
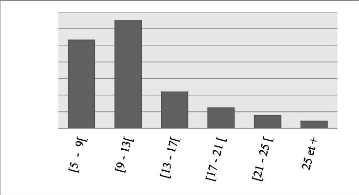
classes de 4 cm d'amplitude. Les tiges de classe [5 cm, 9 cm) et [9 cm, 13 cm)
sont en plus grande proportion à Bois Nago et représentent
à eux seuls 91.34 % de la distribution avec la fréquence de tiges
la plus élevée (701) enregistrée dans la classe [5 cm, 9
cm). Cette distribution est illustrée par le diagramme en barres
ci-dessous :
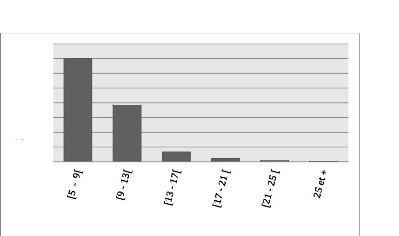
24
Classes de diamètre (cm)
800
700
Nombre de tiges
600
500
400
300
200
100
0
Le DHP moyen du peuplement étudié est
très faible par rapport à celui (23.4 cm) de l'étude de
l'hypothèse réalisé par Traoré et al. (2018).
Quoiqu'âgé de 7 ans, la
Figure 7: Histogramme montrant le nombre de tiges par classe de
diamètre
La distribution diamétrique du peuplement montre
qu'à Bois Nago les tiges de petits diamètres dominent, la forte
présence de ces individus pourrait être justifiée par la
densité trop élevée des tiges qui implique un espacement
trop petit entre les arbres. En effet, l'augmentation de la densité
réduit la croissance en diamètre des arbres (Fonton et al., 2002;
Tonini et al., 2018; Prégent, 1998). Les DHP mesurés
présentent une asymétrie positive et une distribution
générale en forme de « J » inversé, traduisant
un peuplement jeune et non exploitable. D'après le tableau 7, la
dispersion de la distribution du DHP des arbres morts et des arbres vivants est
presque similaire puisque le coefficient de variation présente seulement
une différence de 2 %, cependant la distribution du DHP des arbres
vivants est plus proche du DHP moyen.
Tableau 7: DHP du peuplement d'Acacia mangium de Bois
Nago
|
DHP (cm)
|
Effectif
|
Moyenne
|
Coefficient de variation
|
Erreur-type
|
Min
|
Max
|
|
Arbres vivants
|
1180
|
8.82
|
0.36
|
0.18
|
5
|
29
|
|
Arbres morts
|
9
|
9.04
|
0.34
|
1.99
|
5.40
|
14.90
|
|
Tous
|
1189
|
8.83
|
0.36
|
0.18
|
5
|
29
|
25
différence est énorme par rapport au DHP moyen
du peuplement étudié, deux principales raisons expliquent ces
écarts ; tout d'abord, à Bois Nago les tiges évoluent en
exerçant une forte concurrence entre elles surtout par rapport aux
ressources limitées, ce qui a conduit à l'obtention des tiges de
faibles diamètres. Ensuite, l'absence d'application de technique
sylvicole, notamment l'éclaircissage qui selon Prégent (1998) est
très utile pour obtenir du bois de grosses dimensions.
4.1.3 Surface terrière
La surface terrière des arbres varie de 0.002
m2 à 0.066 m2. Répartie entre les
différentes classes de diamètre, elle varie de 3.25 m2
à 0.22 m2. La distribution de la surface terrière
occupée par les arbres de chaque classe de diamètre est
présentée par l'histogramme ci-dessous.
Surface terrière (m2)
2.5
0.5
3.5
1.5
2
0
3
1
Classe de diamètre (cm)
Figure 8: Histogramme présentant la surface
terrière par classe de diamètre
Les arbres de la classe [9 cm, 13 cm) occupent la plus grande
surface terrière quoiqu'elle ne compte pas le plus grand nombre de
tiges. Cependant le DHP des arbres qui représentent cette classe est
plus élevé par rapport à la première classe. Les
tiges de plus gros diamètres de ce peuplement obtiennent les faibles
valeurs de surface terrière car elles sont en plus petites proportions.
Nos résultats confirment les affirmations de Prégent (1998)
stipulant que la surface terrière est un paramètre tenant
à la fois compte de la grosseur et du nombre de tiges. La surface
terrière obtenue par les différentes catégories d'arbres
de chaque placette a été ramenée à
26
l'hectare (tableau 8). Les résultats montrent que les
arbres vivants occupent la plus grande surface terrière à Bois
Nago comparativement à celle des arbres morts. Les valeurs totales et
moyennes de la surface terrière du peuplement sont
présentées dans le tableau ci-dessous :
Tableau 8: Surface terrière du peuplement
|
Surface
terrière (m2/ha)
|
Effectif Total Moyenne Coefficient Erreur-
de variation type
|
Min Max
|
|
Arbres vivants
|
1180
|
11.67
|
0.33
|
0.31
|
0.03
|
0.15
|
0.55
|
|
Arbres morts
|
9
|
0.09
|
0.003
|
4.71
|
0.004
|
0
|
0.074
|
|
Tous
|
1189
|
11.77
|
0.34
|
0.30
|
0.03
|
0.15
|
0.55
|
Nos résultats corroborent avec celui (11.1
m2/ha) trouvé par Dupuy et N'Guessan (1990) pour un
peuplement d'Acacia mangium âgés de 5 ans. Cependant, par
rapport à l'étude de l'hypothèse réalisée
par Traoré et al. (2018) la surface terrière du peuplement de
Bois Nago corrobore à celui de 3 ans estimée à (10.42
m2/ha) mais elle est beaucoup plus faible que celle
(30.75m2/ha) du peuplement de 7 ans.
4.1.4 Hauteur totale
Les résultats de cette étude montrent que la
hauteur totale des arbres au niveau du site est non uniformément
répartie, elle oscille entre 4.87 m et 27.62 m. La variabilité de
la distribution est illustrée par la boîte à moustache
ci-dessous.
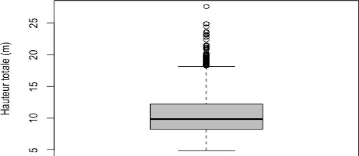
Figure 9: Boite à moustache présentant la
distribution de la hauteur totale du peuplement
27
La boîte à moustache (figure 9) montre une
distribution asymétrique car la valeur de la médiane (9.84 m) est
inférieure à la hauteur moyenne (10.70 m). En plus de cela, 75 %
des arbres ont une hauteur totale supérieure à 8.24 m. La
boîte à moustache permet également de constater que la
hauteur totale du peuplement de Bois Nago présente un bon nombre de
valeurs extrêmes et une valeur aberrante. Et d'après le tableau 9,
les arbres vivants affichent un coefficient de variation plus
élevé, qui traduit une plus forte dispersion de la distribution
de la hauteur totale des arbres vivants comparativement aux arbres morts. La
hauteur totale dominante c'est-à-dire la hauteur moyenne des 100 plus
hauts arbres à l'hectare du peuplement ainsi que la hauteur totale
moyenne est présentée dans le tableau 9:
Tableau 9: Hauteur totale du peuplement
|
Hauteur totale (m)
|
Effectif
|
Hauteur dominante
|
Moyenne
|
Coefficient de variation
|
Erreur-type
|
Min
|
Max
|
|
Arbres vivants
|
1180
|
20.01
|
10.70
|
0.33
|
0.20
|
4.87
|
27.62
|
|
Arbres morts
|
9
|
-
|
9.08
|
0.21
|
1.24
|
6.47
|
12.73
|
|
Tous
|
1189
|
-
|
10.69
|
0.33
|
0.20
|
4.87
|
27.62
|
La valeur moyenne de la hauteur totale du peuplement
étudié est inférieure à celle (18.95 m) de
l'étude de l'hypothèse pour le peuplement de 7 ans. Il en est de
même pour le peuplement de 3 ans dont la hauteur moyenne est de 12.58 m.
Nos résultats se rapprochent plutôt de celle trouvée dans
une étude réalisée par Dupuy et N'Guessan (1990) pour un
peuplement d'Acacia mangium âgé de 5 ans. Le rapport
entre la hauteur moyenne et le DHP moyen du peuplement de Bois Nago donne un
facteur d'élancement égal à 121, cela traduit selon
Massenet (2010) que le peuplement est très fragile et à ce stade,
un éclaircissage pourrait augmenter le risque de chablis.
4.2 Biomasse
4.2.1 Biomasse aérienne avec l'équation de
Chave et al. (2005)
En appliquant l'équation de Chave et al. (2005), les
résultats obtenus pour la biomasse aérienne totale de
l'échantillon inventorié est de 34.96 tonnes. Ramené
à
28
l'hectare (tableau 10), chaque placette a une quantité
de biomasse variant de 18.22 t/ha à 87.53 t/ha. La biomasse des
placettes n'ayant aucun arbre mort est estimée à 0 t/ha. Les
résultats trouvés en moyenne par catégorie sont
présentés dans le tableau suivant :
Tableau 10: Biomasse aérienne du peuplement
calculée à partir de l'équation de Chave et al. (2005)
|
Biomasse aérienne (t/ha)
|
Effectif
|
Moyenne
|
Coefficient de variation
|
Erreur-type
|
Min
|
Max
|
|
Arbres vivants
|
1180
|
50.21
|
0.39
|
6.43
|
18.22
|
87.53
|
|
Arbres morts
|
9
|
0.33
|
4.94
|
0.54
|
0
|
9.71
|
|
Tous
|
1189
|
50.54
|
0.38
|
6.39
|
18.22
|
87.53
|
D'après les valeurs de coefficient de variation des
deux catégories d'arbres, la distribution de la biomasse aérienne
des arbres vivants est beaucoup plus homogène. Cependant, ces derniers
ne représentent que 0.66 % de la biomasse aérienne du site, cela
explique le fait que ces résultats n'influencent pas trop la
distribution totale pour laquelle le coefficient de variation est sensiblement
égal à celui des arbres vivants. Comparativement aux
résultats (63.47 t/ha) de l'étude réalisée par
Hériansyah et al. (2007) dans un peuplement d'Acacia
mangium de 7 ans, la biomasse du peuplement de Bois Nago est moins
élevée. Le résultat est également inférieur
à la valeur (90 t/ha) présentée par Aalde et al. (2006)
pour les plantations de forêt du système montagneux tropical dans
les lignes directrices 2006 du GIEC pour les inventaires nationaux de gaz
à effet de serre.
4.2.2 Biomasse aérienne avec l'équation
de Traoré et al. (2018)
La biomasse aérienne totale des arbres
échantillonnés en appliquant l'équation de Traoré
et al. (2018) est de 50.34 tonnes, dont les arbres vivants constituent 99.45 %
de ce stock. Les résultats trouvés pour chaque placette ont
été ramenés à l'hectare et la biomasse
aérienne a été estimée à une valeur moyenne
par placette de 72.68#177;8.15 t/ha. La biomasse aérienne du peuplement
par catégorie ainsi que
29
les valeurs minimum et maximum des placettes
échantillonnées sont présentées dans le tableau
ci-dessous :
Tableau 11: Biomasse aérienne du peuplement
calculée à partir de l'équation de Traoré et al
(2018)
|
Biomasse aérienne (t/ha)
|
Effectif
|
Moyenne
|
|
|
CV
|
Erreur-type
|
Min
|
Max
|
|
T
|
B
|
F
|
BA
|
|
|
Arbres vivants
|
1180
|
37.74
|
20.93
|
13.62
|
72.29
|
0.34
|
8.25
|
29.36
|
122.96
|
|
Arbres morts
|
9
|
0.24
|
0.15
|
0
|
0.39
|
4.78
|
0.62
|
0
|
11.16
|
|
Tous
|
1189
|
37.98
|
21.08
|
13.62
|
72.68
|
0.34
|
8.15
|
29.36
|
122.96
|
T : Tronc, B : branches, F : feuilles, BA : biomasse
aérienne, CV : coefficient de
variation
Le coefficient de variation des arbres morts étant plus
élevé, cela traduit que la distribution de la biomasse
aérienne des arbres morts est beaucoup plus dispersée. Le
résultat trouvé en appliquant l'équation de Traoré
et al. (2018) est supérieur aux résultats de l'étude
réalisée par Hériansyah et al. (2007) dans le peuplement
d'Acacia mangium de 7 ans ; mais inférieur à la valeur
présentée par Aalde et al. (2006) pour les plantations du
système montagneux tropical.
4.2.3 Comparaison des résultats de la biomasse
aérienne
Le premier modèle d'équation allométrique
appliqué c'est-à-dire celui de Chave et al. (2005) a permis
d'obtenir des résultats plus faibles comparativement au second
modèle, celui de Traoré et al. (2018). Le coefficient de
variation obtenue de la distribution de la biomasse aérienne à
partir de l'équation de Chave et al. (2005) est de 2 % plus
élevé que celui de Traoré et al. (2018), cela traduit que
la distribution des résultats de l'application du premier modèle
est plus dispersée autour de la moyenne. Les raisons expliquant cette
différence dans les résultats de l'application des deux
modèles sont le fait que : d'une part, l'équation de Chave et al.
(2005) fait une estimation directe de la biomasse aérienne et est
applicable aux zones de forêts tropicales humides, la
réalité écologique du site étudié y a donc
été
30
prise en compte. Ce modèle considère
également la densité spécifique du bois, en plus du DHP et
de la hauteur totale, ce qui permet d'avoir une meilleure précision des
estimations. D'autre part, l'équation de Traoré et al. (2018) a
permis une estimation séparée de la biomasse de chaque
compartiment aérien (tronc, branches, feuilles) des arbres afin de
calculer la biomasse aérienne, d'où une surestimation des valeurs
par rapport aux précédents modèles. De plus, c'est un
modèle développé seulement pour un site en Côte
d'Ivoire dont les réalités climatiques et d'altitudes
décrites dans la section 2.2.3, sont très différentes de
Bois Nago. Ainsi, le modèle qui convient le mieux dans le cadre de cette
étude est l'équation développée par Chave et al.
(2005) et c'est celui dont les résultats ont été
utilisés afin de poursuivre le travail.
4.2.4 Biomasse souterraine
La biomasse souterraine de l'échantillon a
été estimée à 7.92 tonnes au total. Convertie
à l'hectare elle varie entre les placettes de 4.35 t/ha à 19.06
t/ha. La biomasse souterraine du peuplement à l'hectare est
présentée dans le tableau suivant :
Tableau 12: Biomasse souterraine du peuplement
|
Biomasse
souterraine (t/ha)
|
Effectif
|
Moyenne
|
Coefficient de variation
|
Erreur-type
|
Min
|
Max
|
|
Arbres vivants
|
1180
|
11.37
|
0.36
|
1.37
|
4.35
|
19.06
|
|
Arbres morts
|
9
|
0.08
|
4.85
|
0.12
|
0
|
2.22
|
|
Tous
|
1189
|
11.45
|
0.36
|
1.35
|
4.35
|
19.06
|
Étant donné que c'est un seul et même
échantillon et que la biomasse souterraine est étroitement
liée à la biomasse aérienne, les constats sont pareils. En
effet, la distribution de la biomasse des arbres vivants est beaucoup plus
homogène. Comparativement aux études de la littérature
(tableau 2) dont les résultats de la biomasse aérienne sont plus
élevés ou plus faibles à celle de Bois Nago, la biomasse
souterraine du peuplement étudié sera plus faible et
inversement.

Biomasse totale (t/ha)
120
100
40
80
60
20
0
0 1000 2000 3000 4000 5000
Densité
y = 0.0144x + 37.439 R2 = 0.1709
Figure 10: Diagramme de corrélation entre la biomasse et
la densité du peuplement
31
4.2.5 Biomasse totale
La biomasse totale de l'échantillon est de 42.88 tonnes.
Les résultats ramenés à l'hectare sont
présentés dans le tableau 13 et ils montrent à partir de
l'erreur-type, une différence de 7.73 t/ha si tous les arbres du
peuplement avaient été mesurés.
Tableau 13: Biomasse totale du peuplement
|
Biomasse totale (t/ha)
|
Effectif
|
Moyenne
|
Coefficient de variation
|
Erreur-type
|
Min
|
Max
|
|
Arbres vivants
|
1180
|
61.58
|
0.38
|
7.79
|
22.57
|
105.64
|
|
Arbres morts
|
9
|
0.40
|
4.92
|
0.66
|
0
|
11.93
|
|
Tous
|
1189
|
61.98
|
0.38
|
7.73
|
22.57
|
105.64
|
La biomasse aérienne représentant 81.53 % de la
biomasse totale influence davantage les résultats que la biomasse
souterraine d'où la similitude du coefficient variation de la biomasse
aérienne avec celui obtenu pour la biomasse totale.
4.2.6 Relation entre la biomasse totale et la
densité
Comme vu dans la section 4.1.1, la densité du
peuplement d'Acacia mangium de Bois Nago est élevée et
de plus très hétérogène. La relation existant entre
la biomasse de ce peuplement et la densité de plantation est
illustrée par le diagramme ci -dessous :
32
Chaque nuage de point de la figure 10 représente la
biomasse totale et la densité de chaque placette. On peut constater que
la densité la plus élevée ne correspond pas au plus grand
stock de biomasse. La figure 10 montre également que la relation entre
la biomasse et la densité n'est pas linéaire car les nuages de
point n'ont pas la forme d'une ligne. Etant donné que certains points de
la figure sont proches et que d'autres sont éloignés, on peut
dire que la relation entre la biomasse et la densité est
modérée et le signe du coefficient de corrélation indique
une relation positive. Donc on peut conclure que les résultats de cette
étude montrent qu'il existe effectivement une relation, quoique non
linéaire entre la biomasse et la densité du peuplement, mais ce
dernier paramètre n'influence pas à lui seul le stock de biomasse
trouvé pour le peuplement d'Acacia mangium de Bois Nago.
4.3 Stock de carbone et quantité de CO2
équivalente
Le stock de carbone total de l'échantillon est
évalué à 20.29 tonnes. La distribution des données
de stock de carbone présente presque les mêmes
caractéristiques (sauf pour l'erreur-type) que celle de la biomasse
aérienne, souterraine et totale à partir desquelles a
été évalué le stock de carbone. Les valeurs
moyennes à l'hectare par placette sont présentées dans le
tableau suivant :
Tableau 14: Stock de carbone du peuplement
|
Stock de carbone (t/ha)
|
Moyenne Coefficient
de variation
|
Erreur-type
|
Min Max
|
|
Partie aérienne
|
23.75
|
0.38
|
3.00
|
8.56
|
41.14
|
|
Partie souterraine
|
5.38
|
0.34
|
0.64
|
2.05
|
8.96
|
|
Totale
|
29.13
|
0.38
|
3.63
|
10.61
|
49.65
|
Le stock moyen de carbone du peuplement de Bois Nago
(29.13#177;3.63 t/ha) est inférieur à la quantité (86.33
t/ha) stockée par le peuplement d'Acacia mangium de 7 ans de
l'étude réalisée par Traoré et al. (2018)
mentionnée dans notre hypothèse, il est également
inférieur au stock de carbone (53.72 t/ha) préconisé par
Aalde et al. (2006). Lorsqu'on compare le résultat de l'étude de
Traoré et al. (2018) à celle de Bois Nago, les deux raisons
suivantes pourraient expliquer les écarts observés.
Comparativement à la valeur (139.8 t/ha) de l'étude
réalisée par Jean Baptiste
33
? La forte abondance des tiges de plus petits diamètres
au niveau du peuplement ; 50 % des arbres ont un diamètre
inférieur à 9 cm et une hauteur inférieure à 10 m.
Le stock de carbone est étroitement lié à la biomasse, or
les individus de petits diamètres représentent selon Mille et
Louppe (2015) une faible fraction de la biomasse aérienne ; d'où
le faible stock de carbone.
? La différence des conditions écologiques et
d'âge du peuplement de Traoré et al. (2018) à celui de Bois
Nago. En effet, le peuplement étudié par Traoré et al.
(2018) âgé 7 ans, est situé à une altitude variant
de 40 à 100 m, plus favorable au développement de
l'espèce, car l'altitude optimale de développement de
l'espèce est en-dessous de 300 m (CIRAD, 2003). La croissance des arbres
diminuent dans les conditions peu favorables dont l'altitude, ce qui entraine
une faible portion de la biomasse et par la même occasion un faible stock
de carbone. Des auteurs dont, Fayolle et al (2013), Ifo et al. (2018), Ifo et
Binsangou (2019), Mugnier et al. (2009) ont effectivement observées des
variations du stock de carbone selon l'altitude, le microclimat et l'âge
de la forêt.
Par rapport à la valeur présentée par
Aalde et al. (2006), les lignes directrices du GIEC sont très peu
explicites quant à la méthode utilisée pour
déterminer la biomasse dont dépend le stock de carbone, mais
confirment que ce ne sont que des valeurs approximatives. De plus ces
études ont été réalisées à
très grande échelle et l'absence de données pour les
régions d'un pays donne parfois lieu à des omissions. A
côté de cela, la nature des sources de données peut
être à l'origine des incertitudes, car les méthodes du GIEC
sont généralement basées sur des données existantes
mises à jour périodiquement (GIEC, 2003). Or à nos jours
une base de données sur les inventaires forestiers en Haïti est
pratiquement inexistante, ce qui écarte encore davantage la
possibilité que la valeur approximative d'Aalde et al. (2006)
corresponde à la réalité de Bois Nago. Dans cette
étude, les données ont été collectées
directement sur le terrain par échantillonnage systématique, ce
qui permet d'avoir une bonne précision des estimations du stock de
carbone.
34
(2018) dans un peuplement multispécifique, le stock de
carbone du peuplement d'Acacia mangium est beaucoup moins
élevé. En effet, la méthodologie appliquée dans les
deux études est similaire car il a été question
d'appliquer des modèles d'équations allométriques
généralisées. Cependant, le peuplement
étudié par Jean Baptiste (2018) a un DHP très
appréciable, qui va au-delà de 100 cm pour certains arbres et de
plus, 40 % du peuplement a une hauteur qui dépasse 20 m. Contrairement
au jeune peuplement d'Acacia mangium qui est monospécifique et
dont moins de 2 % des arbres ont une hauteur qui dépasse 20 m et le DHP
du peuplement est compris seulement entre 5 cm et 29 cm. D'où la raison
expliquant le faible stock de carbone du peuplement d'Acacia mangium
de Bois Nago par rapport au peuplement de l'étude de Jean Baptiste
(2018).
La quantité de CO2 du site a été
déduite directement du stock de carbone total. Les résultats
trouvés pour le peuplement de Bois Nago sont présentés
dans le tableau suivant :
Tableau 15: Quantité de CO2 équivalente du
peuplement
Stock de carbone Quantité moyenne de CO2 Min Max
total
29.13#177;3.63 106.82#177;16.32 38.90 182.06
Nos résultats montrent que le peuplement de Bois Nago
est capable de stocker 106.82#177;16.32 t/ha de CO2. Toujours est-il que ce
stock est faible par rapport à celui de l'étude de Traoré
et al. (2018) car il a été déduit du stock de carbone.
Selon la banque mondiale (2016), chaque habitant du pays émettait par an
0.3 tonne de CO2, ce qui fait que cette plantation est capable de stocker
l'équivalent de CO2 émise annuellement par 2545 habitants.
35
CHAPITRE V : CONCLUSION ET RECOMMANDATIONS
5.1 Conclusion
A partir d'un inventaire statistique systématique
réalisé sur le site, un total de 1189 tiges d'Acacia mangium
parmi lesquels 9 bois morts a été
échantillonné. Les résultats ont montré que le
peuplement est très dense car il compte en moyenne 1707 tiges
d'Acacia mangium par hectare, les bois morts y compris. Étant
donné qu'il n'existe pas encore de modèle allométrique
spécifique à la zone d'étude, pour le calcul de la
biomasse aérienne le choix a été porté sur le
modèle généralisé de Chave et al (2005)
considérant trois variables indépendantes ; le DHP (en cm), la
hauteur totale (en m) et la densité spécifique du bois
sèche au four (en g/cm3). Ensuite l'équation de Cairns
et al. (1997) a été utilisée pour estimer la biomasse
souterraine. Les résultats obtenus sont : 50.54#177;6.39 t/ha,
11.45#177;1.35 t/ha et 61.98#177;7.73 t/ha respectivement pour la biomasse
aérienne, souterraine et totale. Le stock de carbone a été
déduit de la biomasse totale en appliquant une fraction carbone (0.47)
recommandée par le GIEC (2006), et le résultat obtenu pour le
peuplement de Bois Nago est de 29.13 #177; 3.63 t/ha ; de ce stock, la
quantité de CO2 équivaut à 106.82#177;16.32 t/ha.
Le stock de carbone du peuplement d'Acacia mangium de
Bois Nago est plus faible que celui du peuplement étudié par
Traoré et al. (2018) en Côte d'Ivoire, l'hypothèse de
l'étude est confirmée. Cependant, il ressort des résultats
obtenus que l'Acacia mangium peut servir dans les projets de
reboisement à objectif de stockage de carbone. Les plantations
assistées de cette espèce sont à encourager dans les zones
de terre dégradées ou déboisées. Par
conséquent, ces plantations forestières à grande
échelle pourraient non seulement jouer un rôle important en tant
que puits de carbone, mais également procurer des avantages
économiques à la communauté rurale comme bois
énergétique. Toutefois, cette étude doit être
complétée et devra être étendue à d'autres
compartiments importants du cycle biogéochimique du carbone tels que le
sol.
36
5.2 Recommandations
Sur la base des observations faites et des différentes
analyses effectuées dans le cadre de cette étude, les
recommandations s'adressent tout d'abord à la WHH qui avait pris en
charge les activités de reboisement, ensuite à l'ANEVERT
responsable de la gestion de la parcelle et enfin au propriétaire de la
parcelle.
A la WHH, nous faisons les recommandations suivantes : y'
Effectuer des études sur d'autres peuplements d'Acacia mangium
du projet afin de comparer les performances de cette espèce dans
divers sites écologiques du département.
y' Développer des équations allométriques
de la biomasse, spécifique à Acacia mangium et à
la réalité écologique de la zone de l'étude.
y' Faire des études complémentaires qui
tiendraient compte du carbone du sol représentant selon Widhanarto et
al. (2016), le principal contributeur aux stocks totaux de carbone dans les
peuplements d'Acacia mangium jusqu'à 7 ans.
A l'ANEVERT nous recommandons de :
y' Mettre en place un comité d'entretien qui serait
chargé de l'entretien régulier du peuplement, entre autres le
contrôle de la densité de plantation.
y' Effectuer des travaux sylviculturaux dont, des coupes rases
afin d'essayer de rattraper le retard de croissance des arbres et de favoriser
leur croissance en diamètre.
Au propriétaire de la parcelle, il serait judicieux
d'envisager de marquer les limites de la plantation afin qu'elles soient
visibles ; cela facilitera l'identification de la parcelle et évitera
d'empiéter sur les plantations avoisinantes lorsqu'il faudra
entreprendre les différentes opérations sylvicoles.
37
RÉFÉRENCES BIBLIOGRAPHIQUES
Aalde H., Gonzalez P., Gytarsky M., Krug T., Kurz W.
A., Ogle S., Raison J., Schoene D. et Ravindranath N. H., 2006. Terre
forestière. Lignes directrices 2006 du GIEC pour les inventaires
nationaux de gaz à effet de serre. GIEC, 65 p.
Baker, T.R., Phillips, O.L., Malhi, Y., Almeida, S.,
Arroyo, L., Di Fiore, A., Erwin, T., Killeen, T.J., Laurance, S.G., Laurance,
W.F., Lewis, S.L., Lloyd, J., Monteagudo, A., Neill, D.S., Patiño, S.,
Pitman, N.C.A., Silva, J.N.M. and Martínez, R.V., 2004.
Variation in wood density determines spatial patterns in Amazonian forest
biomass. Global Change Biology, 10: 201-220.
Banque mondiale. 2021. Statistiques :
émissions de dioxyde de carbone (CO2), tonnes métriques par
habitant. [Consulté le 20.04.21]. Disponible sur internet: URL :
https://donnees.banquemondiale.org
Brown S., 2002. Measuring carbon in forests:
current status and future challenges, Environmental pollution, 116 (3):
363-372.
Cairns, M.A., Brown, S., Helmer, E.H. et Baumgardner,
G.A., 1997. Root biomass allocation in the world's upland forests.
Oecologia, 111 (1): 1-11.
Chave J., Andalo C., Brown S., Cairns M. A., J.
Chambers Q., Eamus D., Fölster, H., F. Fromard F., Higuchi N., Kira T.,
Lescure J.-P., Nelson B. W., Ogawa H., Puig H., Riera B., Yamakura T., 2005.
Tree Allometry and Improved Estimation of Carbon Stocks and Balance in
Tropical Forests. Oecologia, 145 (1): 87-99.
Chave J., Helene C. M.-L., Timothy R. B., Tomás
A. E., Hans T. S., et Campbell O. W., 2006. Regional and Phylogenetic
Variation of Wood Density. Neotropical Tree Species. Ecological Applications.
16 (6): 2356-2367.
Churches C. E., Wampler P. J., Sun W., Smith A. J.,
2014. Evaluation of forest cover estimates for Haiti using supervised
classification of Landsat data. In: International Journal of Applied Earth
Observation and Geoinformation. 30: 203216
38
CIRAD (Centre de coopération Internationale en
Recherche Agronomique pour le Développement). 2003. Acacia
mangium : [Fiche technique]. Montpellier : CIRAD-Forêt, 2 p.
CIRAD (Centre de coopération Internationale en
Recherche Agronomique pour le Développement). 2011.
Dégradation de la biomasse ligneuse dans une forêt de
Guyane, 8 p.
Dorvil W., 2010. Evaluation de la biomasse et
des stocks de carbone sur des placettes forestières en forêts
tropicales humides de la Guadeloupe. Thèse d'université,
Écosystèmes naturels et exploités. Université des
Antilles et de la Guyane, la Guadeloupe, 44 p.
Dupuy B. et N'Guessan K., 1990. Sylviculture
de l'Acacia mangium en basse Côte d'Ivoire. Bois et Forêts des
Tropiques, 225: 24-32.
Fayolle, A., Doucet, J. L., Gillet, J. F., Bourland,
N., Lejeune, P. 2013. Tree allometry in Central Africa: Testing the
validity of pantropical multi-species allometric equations for estimating
biomass and carbon stocks. Forest Ecology and Management, 305: 29-37.
Fonton N. H., Kakai R. G., Rondeux J., 2002.
Étude dendrométrique d'Acacia auriculiformis, A.
Cunn. ex Benth. en mélange sur vertisol au Bénin.
Biotechnologie, Agronomie, Société et Environnement, 6 (1):
29-37.
Food and Agriculture Organization (FAO), 1979.
Essais d'Acacia mangium Willd. comme essence de plantation
à Sabah. In : Informations sur les ressources génétiques
forestières. [Consultée le 01.08.21]. Disponible sur internet :
URL:
http://www.fao.org/3/n2511f/N2511F08.htm
GIEC, 2006. Autres terres. Dans : Lignes
directrices 2006 du GIEC pour les inventaires nationaux de gaz à effet
de serre, préparé par le Programme pour les inventaires nationaux
de gaz à effet de serre. [Sous la direction de l'équipe de
rédaction principale, Jennifer C. J., Hector D. G., Stephen O.] : IGES,
Japon, 4 (9): 4-10.
GIEC, 2007. Bilan 2007 des changements
climatiques. Contribution des Groupes de travail I, II et III au
quatrième Rapport d'évaluation du Groupe d'experts
39
intergouvernemental sur l'évolution du climat
[Équipe de rédaction principale, Pachauri, R.K. et Reisinger,
A.]. GIEC, Genève, Suisse, 103 p.
GIEC, 2014. Changements climatiques 2014:
Rapport de synthèse. Contribution des Groupes de travail I, II et III au
cinquième Rapport d'évaluation du Groupe d'experts
intergouvernemental sur l'évolution du climat [Sous la direction de
l'équipe de rédaction principale, Rajendra K. Pachauri et Leo A.
Meyer]. GIEC, Genève, Suisse, 161 p.
GIEC. 2003. Recommandations du GIEC en
matière de bonnes pratiques pour le secteur de l'utilisation des terres,
changements d'affectation des terres et foresterie. Programme du GIEC sur les
inventaires nationaux de gaz à effet de serre. [Sous la direction de
l'équipe de rédaction principale, Jim Penman, Michael Gytarsky,
Taka Hiraishi, Thelma Krug, et Dina Kruger]. IGES, Kanagawa, Japon, 594 p.
Hardiyanto E.B., Anshori, S., et Sulistyono, D.,
2004. Early results of site management in Acacia mangium plantations
at PT Musi Hutan Persada, South Sumatra, Indonesia. In: Site management and
productivity in tropical plantation forests, p.93-108.
Hardiyanto E.B. et Wicaksono, A., 2008.
Inter-rotation site management, stand growth and soil properties in
Acacia mangium plantations in South Sumatra, Indonesia. In: Site management and
productivity in tropical plantation forests, p.107-122.
Hériansyah I., Miyakuni K., Kato T., Kiyono Y.,
Kanazawa Y. 2007. Growth characteristics and biomass accumulations of
Acacia mangium under different management practices in Indonesia. Journal of
Tropical Forest Science 19 (4): 226-235.
Ifo S. A., et Binsangou S. 2019.
Variabilité spatiale du stock de carbone de la biomasse
aérienne de la zone urbaine vers la forêt tropicale dense dans
cinq localités de la république du Congo. Conférence
OSFACO: Des images satellites pour la gestion durable des territoires en
Afrique, Cotonou, Bénin. hal-02189395, 15 p.
40
Ifo S.A., Binsangou S., Ibocko Ngala L., Madingou M.,
Sanchez A.C., 2018. Seasonally flooded, and terra firme in northem
Congo: Insights on their structure, diversity and biomass. In : African journal
of ecology, 57: 92-103.
Ilyas S., 2013. Allometric equation and
carbon sequestration of Acacia mangium willd. in coal mining
reclamation a firme in northern Congo: Insights on their structure, diversity
and biomass. African Journal of Ecology, 12 p.
Jean Baptiste K., 2018. Evaluation du stock
de carbone forestier dans les peuplements multispécifiques et
inéquiennes: Cas de l'arboretum de Damien, Haïti. 43 p.
Kangas A., Maltamo M., 2006. Forest
inventory: methodology and applications, Springer, 11 p.
Krisnawati, H., Kallio, M. and Kanninen, M. 2011.
Acacia mangium Willd : ecology, silviculture and
productivity. CIFOR, Bogor, Indonesia, 26 p.
Lecomte H., Rondeux J. 2002. Les inventaires
forestiers nationaux en Europe : Tentative de Synthèse. Les Cahiers
Forestiers de Gembloux, 29 p.
Lotfi A. 2008. Durabilité
écologique des paysages agricoles et production de bois, bocage et
néobocage. Ecologie, Environnement. Université Rennes 1, France,
190 p.
Madignier M.L., Benoit G., Roy C., 2015. Les
contributions possibles de l'agriculture et de la forêt à la lutte
contre le changement climatique. Synthèse du rapport CGAAER, Paris, 83
p.
Massenet J. Y., 2010. Caractérisation et
mesure des peuplements réguliers, 25 p. McGhee W., Saigle W.,
Padonou E.A., Lykke A.M. 2016. Méthodes de calcul de la
biomasse et du carbone des arbres en Afrique de l'Ouest. Annales des Sciences
Agronomiques, 20: 79-98.
MDE (Ministère De l'Environnement), 2013.
Deuxième communication nationale sur les changements
climatiques, Port-au-Prince, Haïti. Ministère De l'Environnement,
14 p.
41
MDE (Ministère De l'Environnement), 2015.
Contribution Prévue Déterminée au niveau National
(CPDN), Port-au-Prince, Haïti. Ministère de l'Environnement, 11
p.
MDE (Ministère de l'Environnement), 2006.
Plan d'Action National d'Adaptation (PANA). Programme Changements
Climatiques République d'Haïti. 61 p.
MELCC (Ministère de l'Environnement et de la
lutte contre les Changements Climatiques), 2019. Guide de
quantification des émissions de gaz à effet de serre,
Québec, 107 p.
Mergen F, Hodges C, Nicholson DI, Popenoe HL, Wiersum
KF. 1983. Mangium and other acacias of the humid tropics. National
Academy Press, Washington, DC, USA, 62 p.
Mille G., Louppe D. 2015. Mémento du
Forestier Tropical. Editions Quae. Versailles, France; 1200 p.
Mugnier A., Cassagne B., Bayos N., Lafon C., 2009.
Estimation des stocks de carbone des forêts du bassin du Congo
pour la REDD: Etude comparative conduite sur 22 types forestiers, 4 pays et un
dispositif d'aménagement 4,8 millions d'ha. Buenos Aires, Argentina, 12
p.
Nasi R., Mayaux P., Devers D., Bayol N., Eba'a Atyi
R., Mugnier A., Cassagne B., de Marc ken P., Billand A., Sonwa D., 2008.
Un aperçu des stocks de carbone et leurs variations dans les
Forêts du Bassin du Congo. Luxembourg, Belgique, Office des publications
de l'Union européenne, p. 199-216.
Pearson T. H. et Brown S., 2005. Guide de
Mesure et de Suivi du Carbone dans les Forêts et Prairies Herbeuses,
Arlington, USA. Winrock International, 39 p.
Peltier R., Forkong N.C., Ntoupka N., Manlay R., Henry
M., Morillon V., 2007. Evaluation du stock de carbone et de la
productivité en bois d'un parc à karités du Nord Cameroun.
Bois et Forêts des Tropiques. 294 (4): 39-50.
Picard N., Saint-André L., Henry M. 2012.
Manuel de construction d'équations allométriques pour
l'estimation du volume et la biomasse des arbres : de la mesure de terrain
à la prédiction, Rome, Italie et Montpellier, France.
Organisation des
42
Nations Unies pour l'alimentation et l'agriculture, et Centre de
Coopération Internationale en Recherche Agronomique pour le
Développement, 220 p.
Pinyopusarerk K., Liang S.B., Gunn B.V. 1993.
Taxonomy, distribution, biology and use as an exotic. In: Acacia
mangium - Growing and Utilization. Winrock International and FAO, Bangkok,
Thailand, p 56-62.
Prégent, G., 1998. L'éclaircie
dans les plantations. Gouvernement du Québec, ministère des
ressources naturelles, Direction de la recherche forestière.
Mémoire de recherche forestière, 38 p.
Programme des Nations Unies pour le
Développement (PNUD). 2016. Programme de pays pour Haïti,
Port-au-Prince, Haïti. PNUD Haïti, 19 p.
Singh B. et Cohen M.J., 2014. Adaptation aux changements
climatiques. Le cas d'Haïti. Rapport de recherche Oxfam. 39 p.
Tonini H., Schwengber D.R., Morales M. M., Magalhaes
C. A., Franco J. M., 2018. Growth, biomass, and energy quality of
Acacia mangium timber grown at different spacings. Pesq. agropec.
bras., Brasiìlia, 53 (7): 791-799.
Traoré S., Djomo A.N., N'Guessan A.K.,
Coulibaly B., Ahoba A., Gnahoua G.M., N'Guessan E. K., Adou Yao C.Y., N'Dja J.
K., Guéde N. Z., 2018. Stand structure, allometric equations,
biomass and carbon Sequestration Capacity of Acacia mangium Willd.
(Mimosaceae) in Côte d'Ivoire. In: Open Journal of Forestry, 8: 42-60.
Tsoumou B.R., Lumandé K.J., Kampé J.P.,
Nzila J.D. 2016. Estimation de la quantité de carbone
séquestré par la Forêt Modèle de Dimonika (Sud-ouest
de la République du Congo). Revue Scientifique et Technique Forêt
et Environnement du Bassin du Congo, 6: 39-45.
Valbrun O., 2005. Monographie agricole de la
commune de Mont-Organisé, Programme de Relance Economique (PROREC)/
MARNDR, Haïti, 57 p.
Wiliam M. Ciesla, 1997. Les arbres et les
forêts, sources et puits de GES et de carbone. In : Le Changement
Climatique, les Forêts et l'Aménagement Forestier : Aspects
Généraux [En ligne]. [Organisation des Nations Unies pour
l'alimentation
43
et l'agriculture] 1997 [Consultée le 14.03.21]. Disponible
sur internet : URL :
http://www.fao.org/docrep/v5240f/v5240f09.ht
Zolkos S.G., Goetz S.J. et Dubayah R., 2013. A
meta-analysis of terrestrial aboveground biomass estimation using lidar remote
sensing. Remote Sens. Environ. 128: 289-298.
ANNEXES
Annexe 1: Coordonnées des centres des
placettes
|
# placettes
|
Longitude
|
Latitude
|
|
1
|
-71.81500156
|
19.40441313
|
|
2
|
-71.81499646
|
19.40478903
|
|
3
|
-71.81458986
|
19.40478189
|
|
4
|
-71.81499107
|
19.40516914
|
|
5
|
-71.81458439
|
19.40516582
|
|
6
|
-71.81418789
|
19.40515695
|
|
7
|
-71.81378736
|
19.40514802
|
|
8
|
-71.81338136
|
19.40552301
|
|
9
|
-71.81377995
|
19.40552809
|
|
10
|
-71.81417645
|
19.40553696
|
|
11
|
-71.81457702
|
19.40554399
|
|
12
|
-71.81498161
|
19.40555109
|
|
13
|
-71.81497022
|
19.40592918
|
|
14
|
-71.81456764
|
19.40592212
|
|
15
|
-71.81417311
|
19.40591519
|
|
16
|
-71.81377053
|
19.40590813
|
|
17
|
-71.81337000
|
19.40589919
|
|
18
|
-71.81297145
|
19.40589219
|
|
19
|
-71.81256351
|
19.40626333
|
|
20
|
-71.81296400
|
19.40627418
|
|
21
|
-71.81336460
|
19.40627930
|
|
22
|
-71.81376316
|
19.40628629
|
|
23
|
-71.81416369
|
19.40629523
|
|
24
|
-71.81456224
|
19.40630223
|
|
25
|
-71.81496486
|
19.40630738
|
|
26
|
-71.81495745
|
19.40668746
|
|
27
|
-71.81455495
|
19.40667657
|
|
28
|
-71.81415635
|
19.40667149
|
|
29
|
-71.81375377
|
19.40666442
|
|
30
|
-71.81335522
|
19.40665743
|
|
31
|
-71.81295864
|
19.40665238
|
|
32
|
-71.81255811
|
19.40664343
|
|
33
|
-71.81215952
|
19.40663835
|
|
34
|
-71.81454750
|
19.40705856
|
|
35
|
-71.81495206
|
19.40706757
|
Annexe 2: Précipitation moyenne mensuelle
de Ouanaminthe pour la période (19882019)
|
Janv
|
Fév
|
Mar
|
Avr
|
Mai
|
Juin
|
Juil
|
Août
|
Sept
|
Oct
|
Nov
|
Déc
|
|
56.0
|
32.7
|
47.7
|
73.0
|
117.4
|
105.9
|
80.3
|
107.5
|
122.6
|
106.6
|
68.9
|
40.7
|
Annexe 3: Précipitation totale annuelle
de Ouanaminthe pour la période (1988-2019)
|
Années
|
Précipitations (mm)
|
Années
|
Précipitations (mm)
|
Années
|
Précipitations (mm)
|
|
1988
|
1715.5
|
1999
|
1487.1
|
2010
|
2488.5
|
|
1989
|
1146.9
|
2000
|
1241.4
|
2011
|
1514.8
|
|
1990
|
1161.8
|
2001
|
1347.5
|
2012
|
1894.1
|
|
1991
|
815.3
|
2002
|
1044
|
2013
|
1604.9
|
|
1992
|
1084
|
2003
|
1487.3
|
2014
|
1977.5
|
|
1993
|
1328.2
|
2004
|
1302.5
|
2015
|
1500.3
|
|
1994
|
1145.6
|
2005
|
1705.2
|
2016
|
2301.6
|
|
1995
|
1043.9
|
2006
|
1793
|
2017
|
1907
|
|
1996
|
1294.2
|
2007
|
2881.2
|
2018
|
1292.3
|
|
1997
|
867
|
2008
|
1623.7
|
2019
|
1805.2
|
|
1998
|
1207.2
|
2009
|
2215.1
|
|
|
Annexe 4: Classification des zones bioclimatiques de
Holdridge
Annexe 5: Paramètres
dendrométriques comptés, mesurés et calculés par
placette
|
#
placette
|
Nombre de tiges
|
N
arbres vivants
|
N bois morts
|
Surface
terrière
(m2/ha)
|
Hauteur moyenne
|
Diamètre moyen
|
|
1
|
25
|
25
|
0
|
0.31
|
12.12
|
9.9
|
|
2
|
38
|
38
|
0
|
0.41
|
11.33
|
9.45
|
|
3
|
37
|
37
|
0
|
0.25
|
9.26
|
7.51
|
|
4
|
38
|
38
|
0
|
0.50
|
10.97
|
9.83
|
|
5
|
46
|
46
|
0
|
0.36
|
8.50
|
7.82
|
|
6
|
32
|
31
|
1
|
0.41
|
11.85
|
9.98
|
|
7
|
46
|
45
|
1
|
0.45
|
10.01
|
8.74
|
|
8
|
24
|
24
|
0
|
0.31
|
12.95
|
10.25
|
|
9
|
27
|
27
|
0
|
0.28
|
9.98
|
9.1
|
|
10
|
34
|
33
|
1
|
0.27
|
12.52
|
8.24
|
|
11
|
31
|
31
|
0
|
0.27
|
12.40
|
10.91
|
|
12
|
40
|
40
|
0
|
0.31
|
9.79
|
7.98
|
|
13
|
36
|
36
|
0
|
0.42
|
11.48
|
9.66
|
|
14
|
23
|
23
|
0
|
0.22
|
11.17
|
8.95
|
|
15
|
26
|
26
|
0
|
0.24
|
11.49
|
8.78
|
|
16
|
18
|
18
|
0
|
0.15
|
8.71
|
8.26
|
|
17
|
26
|
26
|
0
|
0.30
|
12.25
|
9.57
|
|
18
|
20
|
14
|
6
|
0.29
|
11.14
|
10.18
|
|
19
|
28
|
28
|
0
|
0.33
|
9.47
|
9.25
|
|
20
|
25
|
25
|
0
|
0.37
|
8.51
|
10.36
|
|
21
|
24
|
24
|
0
|
0.33
|
9.26
|
10.07
|
|
22
|
15
|
15
|
0
|
0.20
|
10.06
|
10.19
|
|
23
|
39
|
39
|
0
|
0.53
|
12.02
|
10.03
|
|
24
|
35
|
35
|
0
|
0.44
|
15.36
|
10.26
|
|
25
|
49
|
49
|
0
|
0.54
|
10.91
|
9.23
|
|
26
|
69
|
69
|
0
|
0.55
|
10.09
|
8.08
|
|
27
|
37
|
37
|
0
|
0.37
|
12.34
|
9.05
|
|
28
|
27
|
27
|
0
|
0.19
|
8.93
|
7.69
|
|
29
|
39
|
39
|
0
|
0.24
|
9.72
|
7.22
|
|
30
|
35
|
35
|
0
|
0.29
|
11.79
|
8.44
|
|
31
|
31
|
31
|
0
|
0.32
|
8.48
|
9.25
|
|
32
|
28
|
28
|
0
|
0.29
|
9
|
9
|
|
33
|
34
|
34
|
0
|
0.21
|
10.40
|
7.19
|
|
34
|
22
|
22
|
0
|
0.24
|
12.79
|
9.83
|
|
35
|
85
|
85
|
0
|
0.40
|
9.83
|
6.41
|
AGB1 : Biomasse aérienne obtenue à partir des
équations de Traoré et al (2018)
AGB2 : Biomasse aérienne obtenue à partir
l'équation de Chave et al (2005)
Annexe 6: Biomasse (aérienne,
souterraine et totale) des arbres échantillonnés par placette
|
#
Placette
|
Biomasse troncs (t)
|
Biomasse branches (t)
|
Biomasse feuilles
(t)
|
AGB1 (t)
|
AGB2 (t)
|
BGB (t)
|
Biomasse totale (t)
|
|
1
|
0.69
|
0.35
|
0.23
|
1.27
|
0.92
|
0.20
|
1.12
|
|
2
|
0.91
|
0.50
|
0.32
|
1.73
|
1.20
|
0.27
|
1.47
|
|
3
|
0.48
|
0.35
|
0.23
|
1.07
|
0.63
|
0.15
|
0.78
|
|
4
|
1.03
|
0.52
|
0.35
|
1.89
|
1.42
|
0.31
|
1.73
|
|
5
|
0.64
|
0.44
|
0.31
|
1.39
|
0.86
|
0.21
|
1.07
|
|
6
|
0.92
|
0.46
|
0.29
|
1.67
|
1.25
|
0.27
|
1.52
|
|
7
|
0.89
|
0.53
|
0.35
|
1.78
|
1.19
|
0.28
|
1.47
|
|
8
|
0.82
|
0.37
|
0.23
|
1.42
|
1.08
|
0.24
|
1.32
|
|
9
|
0.51
|
0.31
|
0.22
|
1.04
|
0.69
|
0.16
|
0.85
|
|
10
|
0.72
|
0.41
|
0.24
|
1.36
|
0.90
|
0.21
|
1.12
|
|
11
|
1.27
|
0.51
|
0.33
|
2.11
|
1.74
|
0.36
|
2.10
|
|
12
|
0.63
|
0.42
|
0.27
|
1.32
|
0.81
|
0.20
|
1.01
|
|
13
|
0.99
|
0.50
|
0.32
|
1.80
|
1.31
|
0.29
|
1.61
|
|
14
|
0.45
|
0.27
|
0.17
|
0.89
|
0.58
|
0.14
|
0.72
|
|
15
|
0.58
|
0.32
|
0.20
|
1.10
|
0.74
|
0.17
|
0.91
|
|
16
|
0.27
|
0.18
|
0.13
|
0.58
|
0.36
|
0.09
|
0.45
|
|
17
|
0.73
|
0.36
|
0.23
|
1.32
|
0.96
|
0.21
|
1.18
|
|
18
|
0.63
|
0.29
|
0.14
|
1.05
|
0.87
|
0.18
|
1.05
|
|
19
|
0.61
|
0.34
|
0.24
|
1.18
|
0.85
|
0.19
|
1.04
|
|
20
|
1.22
|
0.66
|
0.49
|
2.36
|
1.72
|
0.38
|
2.10
|
|
21
|
0.63
|
0.32
|
0.23
|
1.18
|
0.90
|
0.19
|
1.09
|
|
22
|
0.38
|
0.20
|
0.14
|
0.73
|
0.52
|
0.12
|
0.64
|
|
23
|
1.20
|
0.57
|
0.37
|
2.13
|
1.64
|
0.35
|
1.99
|
|
24
|
1.31
|
0.59
|
0.33
|
2.23
|
1.70
|
0.37
|
2.07
|
|
25
|
1.19
|
0.62
|
0.41
|
2.22
|
1.60
|
0.36
|
1.96
|
|
26
|
1.22
|
0.75
|
0.48
|
2.45
|
1.59
|
0.37
|
1.97
|
|
27
|
0.90
|
0.48
|
0.30
|
1.69
|
1.17
|
0.27
|
1.44
|
|
28
|
0.36
|
0.26
|
0.17
|
0.79
|
0.47
|
0.11
|
0.58
|
|
29
|
0.52
|
0.37
|
0.23
|
1.12
|
0.66
|
0.16
|
0.82
|
|
30
|
0.74
|
0.42
|
0.25
|
1.42
|
0.94
|
0.22
|
1.16
|
|
31
|
0.51
|
0.34
|
0.26
|
1.11
|
0.70
|
0.17
|
0.86
|
|
32
|
0.52
|
0.32
|
0.23
|
1.07
|
0.70
|
0.16
|
0.87
|
|
33
|
0.49
|
0.33
|
0.20
|
1.02
|
0.61
|
0.15
|
0.76
|
|
34
|
0.60
|
0.32
|
0.19
|
1.11
|
0.78
|
0.18
|
0.96
|
|
35
|
0.90
|
0.71
|
0.43
|
2.03
|
1.10
|
0.28
|
1.38
|
Annexe 7: Fiche de collecte des
données
Date.../.../... No placette : Coordonnées
des placettes :
|
# arbre Catégorie
Arbre vivant
|
|
DHP (cm) Circ (cm) Do (m)
|
Cas
|
á (°) f (°)
|
|
Mort sur pied
|
1e 2e 3e 4e
|
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
2
|
|
|
|
|
|
|
|
|
|
|
|
|
3
|
|
|
|
|
|
|
|
|
|
|
|
|
4
|
|
|
|
|
|
|
|
|
|
|
|
|
5
|
|
|
|
|
|
|
|
|
|
|
|
|
6
|
|
|
|
|
|
|
|
|
|
|
|
|
7
|
|
|
|
|
|
|
|
|
|
|
|
|
8
|
|
|
|
|
|
|
|
|
|
|
|
|
9
|
|
|
|
|
|
|
|
|
|
|
|
|
10
|
|
|
|
|
|
|
|
|
|
|
|
|
11
|
|
|
|
|
|
|
|
|
|
|
|
|
12
|
|
|
|
|
|
|
|
|
|
|
|
|
13
|
|
|
|
|
|
|
|
|
|
|
|
|
14
|
|
|
|
|
|
|
|
|
|
|
|
|
15
|
|
|
|
|
|
|
|
|
|
|
|
|
16
|
|
|
|
|
|
|
|
|
|
|
|
|
17
|
|
|
|
|
|
|
|
|
|
|
|
|
18
|
|
|
|
|
|
|
|
|
|
|
|
|
19
|
|
|
|
|
|
|
|
|
|
|
|
|
20
|
|
|
|
|
|
|
|
|
|
|
|
|
21
|
|
|
|
|
|
|
|
|
|
|
|
|
22
|
|
|
|
|
|
|
|
|
|
|
|
|
23
|
|
|
|
|
|
|
|
|
|
|
|
|
24
|
|
|
|
|
|
|
|
|
|
|
|
Annexe 8: Quelques prises de vue du site
a- Vue aérienne du site b- Feuilles d'Acacia
mangium

c- Mesure du DHP d- Mesure du DHP

e- Partie dense du site

| 

