|
Mémoire
Présenté
à
L'Université de Tunis El Manar
Faculté des
Sciences de Tunis
Pour obtenir
Le diplôme de Mastère
Dynamique Tectono-sédimentaire et Caractérisation
des Réservoirs
Par
MOHAMED SOUA
|
Biostratigraphie de haute résolution des
Foraminifères
planctoniques
du passage Cénomanien - Turonien
et impact de
l'événement anoxique EAO-2 sur
ce groupe dans la marge sud de
la Téthys,
exemple : régions de Jerissa et Bargou
|
Soutenu le 20/07/2005
Jury
Président : Pr. Mohamed Moncef TURKI Faculté des
Sciences de Tunis
Directeur de recherche : Pr. Dalila ZAGHBIB-TURKI Faculté
des Sciences de Tunis
Examinateur : Pr. Habib BELAYOUNI Faculté des Sciences de
Tunis
Je suis reconnaissant aux personnels de l'Office National des
Mines et en particulier à Mr. Abdelbaki Mansouri, ingénieur en
chef et directeur de la recherche minière à
Avant propos
Je voudrais saisir l'occasion de l'élaboration de ce
Mémoire pour exprimer ma profonde gratitude à tous ceux et celles
qui, directement ou indirectement, m'ont encouragé à suivre et/ou
facilité mon engagement dans la voie de la Micropaléontologie et
la Paléoécologie. Je m'adresse tout particulièrement
à Madame Dalila ZAGHBIB-TURKI professeur à la Faculté des
Sciences de Tunis et directeur de mes travaux de recherche de Mastère ;
elle a activement contribué à sa réalisation. Je lui suis
reconnaissant de m'avoir donné l'opportunité de travailler sur ce
sujet, un sujet qui demande plus d'exploration et de recherche. Grâce
à ce travail et à l'encadrement de Pr. Dalila ZAGHBIB-TURKI, j'ai
mis un pied dans l'étonnant royaume des Foraminifères. Ses
remarques, ses conseils, ses critiques constructifs et ses idées
pertinentes m'ont permis d'évoluer sur le plan de la rigueur et la
précision scientifique. Qu'elle soit assurée de ma profonde
gratitude.
J'exprime également ma profonde gratitude à
Monsieur Mohamed Moncef TURKI professeur à la Faculté des
Sciences de Tunis et directeur du Département de Géologie, pour
m'avoir accompagné sur le terrain dans la région de Bargou ainsi
que de m'avoir fait profiter de sa valeureuse expertise. Je le remercie aussi
d'avoir accepté de juger mon modeste travail.
De même, Je remercie profondément Monsieur Habib
BELAYOUNI, professeur à la Faculté des Sciences de Tunis pour
avoir accepté de faire partie du jury
Ma gratitude va de droit à toute ma famille : mes
Parents, Mahmoud et Najet, dont l'apport moral et matériel n'ont
d'égal que leur compréhension et leur gentillesse durant toute la
période de réalisation de ce travail, mes deux frères
Lassâad et Chokri pour m'avoir donné de leurs temps et m'avoir
accompagné maintes fois sur le terrain et en me fournissant le soutien
moral et l'encouragement nécessaire pour la finalisation de ce modeste
travail. Je leur dédie ce Mémoire.
l'ONM pour ses chaleureux encouragements, sa
disponibilité et son aide précieuse pour la confection des lames
minces. De même je remercie vivement Mr. Ali Ouertani ingénieur
principal à l'ONM pour m'avoir accompagné sur le terrain et
m'avoir indiqué le lieu de la coupe de Oued Es Smara dans la
région de Jerissa.
Je ne saurais remercier Dr. Radouen AZIZI KADMIRI, assistant
à la Faculté des Sciences de Tunis de m'avoir accompagné
sur le terrain et d'avoir partagé ses connaissances en matière de
sédimentologie.
Je suis reconnaissant à Mr. Bahri REZIG pour `avoir
permis de faire des séances de M.E.B à l'ENIT (École
Nationale des Ingénieurs de Tunis). De même je remercie Mlle
Raoudha BRINI pour m'avoir donné de son temps pour mener à bien
ces séances.
Je voudrais témoigner de toute ma reconnaissance
à Monsieur Fredj Chaâbani Maître de Conférence
à la Faculté des Sciences de Tunis, pour m'avoir permis et
facilité la tâche d'effectuer les analyses calcimétriques
de mes échantillons dans son laboratoire.
Je tiens à remercier mes collègues de
laboratoire, Ramadan BENASSI, Lamia ZILI, Noujoud GALLALA, Amal CHAKROUN, Semia
ABIDI et tous mes camarades de classe de Mastère « Dynamique
Tectono-Sédimentaire et Caractérisation des Réservoirs
» notamment Riadh CHEBBI qui m'a accompagné sur le terrain et qui
n'a pas cessé de me soutenir.
L'amitié et la collaboration sans faille des amies et
collègues Lamia Kouzena et Abir Guidara de la Faculté des
sciences de Tunis m'ont permis d'avancer dans mes travaux aussi bien pour les
analyses calcimétriques que pour l'élaboration de cartes
numérisées.
Je ne saurais oublier mes amis Hichem, Adel, Sadok, Sinda,
Asma, Nessrine, Azzouz, Atef, Zied, Semi, Kamel, Mohamed, Sabri, Ramzi, Kais,
Jalloul, Sofien, Nabil, Ramla, Imène, Tarek, Adnène, Hajer, Anis,
Wahid et Walid pour leur soutien moral et leur encouragement qu'ils m'ont
témoigné tout au long de ce travail.
Je tiens aussi à remercier tout le personnel du
Département de Géologie notamment Adel et Meriam.
J'adresse mes plus sincères remerciements à ma
fiancée, Amira Khedhri, non seulement pour m'avoir patiemment soutenu et
encouragé mais aussi pour m'avoir fourni une aide précieuse.
Qu'elle trouve dans ces lignes l'expression de ma profonde gratitude.
Enfin, mes hommages vont aux nombreux chercheurs auteurs de
publications et articles qui à travers la richesse et la qualité
de leurs travaux j'ai tant appris. Je pense entre autres à Alfred
Loeblich et Helen Tappan desquels je me suis inspiré maintes fois pour
la compréhension des structures internes des Foraminifères
planctoniques (du passage Cénomanien - Turonien) et leurs biologies. Je
pense aussi à Madlène Neumann, Mark Leckie, Michèle Caron,
Gerta Keller, Francis Robaszynski et bien d'autres.
1
TABLE DES MATIÈRES
PREMIERIÈRE PARTIE
INTRODUCTION GÉNÉRALE PAGES
I - GENERALITES 07
1 - LES CARACTERISTIQUES GENERALES DE L'ÉVENEMENT ANOXIQUE
OCEANIQUE
DURANT LA LIMITE CENOMANIEN - TURONIEN (EAO-2) 12
2 - LA COUPE TYPE ET LES COUPES LES MIEUX ETUDIEES AU MONDE
CONCERNANT LA LIMITE CENOMANIEN - TURONIEN 18
a - Le Niveau Bonarelli : 18
b - La coupe de Pueblo : 20
II - LA LIMITE CENOMANIEN - TURONIEN EN TUNISIE 21
III - CARACTERISTIQUES ET EXTENSION DES DEPOTS DE LA LIMITE C/T
DANS LE DOMAINE
TETHYSIEN 30
A - L'ALGERIE 30
B - LE MAROC 31
C - L'ESPAGNE 32
D - LA FRANCE 33
E - L'ITALIE 33
IV - OBJECTIF DU TRAVAIL 34
V - MATERIEL ET METHODE 35
DEUXIÈME PARTIE
DESCRIPTION DES FACIÈS ET ENVIRONNIMENTS DE
DÉPÔTS
I - INTRODUCTION 39
A - ÉTUDE DE LA COUPE D'OUED ES SMARA : CES 40
1 - Historique des travaux antérieurs 40
2 - Cadre géographique et géologique 42
a - Cadre géographique 42
b - Cadre géologique 43
3 - Coupe d'Oued Es Semara (CES) 45
a - L'Unité basale U1 [CES-1 - CES-2] : 45
b - Unité U2 [CES-3 - CES-6] 47
c - Unité U3 [CES-7 - CES-18] 47
d - Unité U4 [CES-19 - CES-37] 49
e - Unité U5 [CES-38 - CES-61] 51
f - Unité U6 [CES-47 - CES-67] 53
B - ÉTUDE DE LA COUPE DE OUED EL KHARROUB : COK 53
1 - Travaux antérieurs 53
2 - Cadre géographique et géologique 54
a - Cadre géographique 54
b - Cadre géologique 55
3 - Étude stratigraphique: 57
a) L'Unité basale U1 [COK-1 - COK-15] 59
b) L'Unité U2 [COK-16 - COK-30] 59
c)
2
L'Unité U3 [COK-30 - COK-37] 62
d) L'Unité U4 [COK-38 - COK-61] 62
e) L'Unité U5 [COK-62 - COK-77] 63
f) L'Unité U6 [COK-78 - COK-89] 63
g) L'Unité U7 [COK-90 - COK-100] 64
II - INTERPRETATION DES ENVIRONNEMENTS DE DEPOTS 64
TROISIÈME PARTIE
BIOSTRATIGRAPHIE
I - BIOSTRATIGRAPHIE 67
A - BIOZONATION DES FORAMINIFERES PLANCTONIQUE DU PASSAGE
CENOMANIEN -
TURONIEN. 67
1 - Historique 67
2 - Biozonation des Foraminifères planctoniques 71
a) Zone à Rotalipora cushmani. 72
- La sous-zone à Dicarinella hagni 72
b) Zone à Whiteinella archaeocretacea 73
- La sous-zone à Globigerinelloides bentonensis
74
- La sous-zone à Dicarinella hagni 74
- La sous-zone à Heterohelix moremani 74
c) Zone à Helvetoglobotruncana helvetica 75
3 - Conclusion : 75
B - BIOSTRATIGRAPHIE DES RADIOLAIRES DU PASSAGE CENOMANIEN -
TURONIEN. 77
Historique 77
QUATRIÈME PARTIE
PALÉOÉCOLOGIE DES FORAMINIFÈRES
PLANCTONIQUES ET DES RADIOLAIRES DU PASSAGE
CÉNOMANIEN -
TURONIEN
A - PALEOECOLOGIE DES FORAMINIFERES PLANCTONIQUES DU PASSAGE
CENOMANIEN - TURONIEN 80
1) INTRODUCTION 80
2) PALEOECOLOGIE DES FORAMINIFERES PLANCTONIQUES DE L'INTERVALLE
DU PASSAGE
CENOMANIEN - TURONIEN. 82
a) Faunes de mer épicontinentale (<100m) 82
b) Faunes de subsurface habitant les mers
épicontinentales 82
c) La faune de mer profonde 83
a) Les habitants de subsurface : les formes
carénées 83
b) Les habitants de la zone photique inférieur : les
espèces faiblement
carénées 84
c) Les habitants de la zone à oxygène minimum :
les Hétérohélicidés 84
d) Les habitants de surface de faible salinité :
Hedbergella planispira 86
e) Les habitants de surface à salinité normale
86
f) Les opportunistes, habitants des surfaces eutrophiques :
Guembelitria 86
3) LES IDICATEURS PALEOECOLOGIQUES DU PASSAGE CENOMANIEN -
TURONIEN DE
LA COUPE D'OUED ES SMARA (CES) 86
1) Assemblage de
Foraminifères planctoniques diversifiés dans une masse d'eau
stratifiée 86
2)
3
L'installation de la ZOM et prolifération des
Heterohelicacea 87
3) Expansion de la ZOM, hypoxie -anoxie et multiplication des
opportunistes 89
4) Retour aux conditions normales : renouvellement faunique
91
B - PALEOECOLOGIE DES RADIOLAIRES DU PASSAGE CENOMANIEN -
TURONIEN. 94
CINQUIÈME PARTIE
SYSTÉMATIQUE DES FORAMINIFÈRES
PLANCTONIQUES DU PASSAGE CÉNOMANIEN -
TURONIEN
ORDRE : FORAMINIFERIDA EICHWALD, 1830 101
SOUS-ORDRE : GLOBIGERININA DELAGE & HEROUARD, 1896 101
SUPERFAMILLE : HETEROHELICACEA CUSHMAN, 1927 101
FAMILLE : HETEROHELICIDAE CUSHMAN, 1927 101
SOUS-FAMILLE : HETEROHELICINAE CUSHMAN, 1927 101
GENRE : HETEROHELIX EHRENBERG, 1843 101
ESPECE : HETEROHELIX MOREMANI (CUSHMAN, 1938) 101
ESPECE : HETEROHELIX NAVARROENSIS LOEBLICH, 1951 102
ESPECE : HETEROHELIX PULCHRA (BROTZEN, 1936) 103
ESPECE : HETEROHELIX REUSSI (CUHMAN, 1938) 103
SUPERFAMILLE : HETEROHELICACEA CUSHMAN, 1927 104
FAMILLE : GUEMBELITRIIDAE MONTANARO GALLITELLI, 1957 104
SOUS-FAMILLE : GUEMBELITRIINAE MONTANARO GALLITELLI, 1957 104
GENRE : GUEMBELITRIA CUSHMAN, 1933 104
ESPECE : GUEMBELITRIA CENOMANA (KELLER, 1935) 104
ESPECE : GUEMBELITRIA ALBERTENSIS STELCK ET WALL, 1954 105
SUPERFAMILLE : PLANOMALINACEA BOLLI, LOEBLICH & TAPPAN, 1957
106
FAMILLE : GLOBIGERINELLOIDIDAE LONGORIA, 1974 106
SOUS-FAMILLE : GLOBIGERINELLOIDINAE LONGORIA, 1974 106
GENRE : GLOBIGERINELLOIDES CUSHMAN & TEN DAM, 1948
106
ESPÈCE : GLOBIGERINELLOIDES BENTONENSIS (MORROW,
1934) 107
ESPÈCE : GLOBIGERINELLOIDES ULTRAMICRA (SUBBOTINA, 1949)
108
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 108
FAMILLE: HEDBERGELLIDAE LOEBLICH & TAPPAN, 1961 108
SOUS-FAMILLE: HEDBERGELLINAE LOEBLICH & TAPPAN, 1961 108
GENRE: CLAVIHEDBERGELLA BANNER & BLOW, 1959 108
ESPÈCE-TYPE : HASTIGERINELLA
(=CLAVIHEDBERGELLA) SUBCRETACEA TAPPAN, 1943 109
ESPÈCE: CLAVIHEDBERGELLA SIMPLEX (MORROW, 1934)
109
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 110
FAMILLE: HEDBERGELLIDAE LOEBLICH & TAPPAN, 1961 110
SOUS-FAMILLE: HEDBERGELLINAE LOEBLICH & TAPPAN, 1961 110
GENRE: HEDBERGELLA BRÖNNIMANN & BROWN, 1958
110
ESPÈCE : HEDBERGELLA DELRIOENSIS (CARSEY, 1926)
110
ESPÈCE : HEDBERGELLA PLANISPIRA (TAPPAN, 1940) 111
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 112
FAMILLE: HEDBERGELLIDAE LOEBLICH & TAPPAN, 1961 112
SOUS-FAMILLE: HEDBERGELLINAE LOEBLICH & TAPPAN, 1961 112
GENRE : WHITEINELLA PESSAGNO, 1967 112
ESPECE: WHITEINELLA APRICA (LOEBLICH & TAPPAN, 1961)
112
ESPÈCE : WHITEINELLA ARCHAEOCRETACEA PESSAGNO,
1967 113
ESPÈCE : WHITEINELLA BALTICA DOUGLAS &
RANKIN, 1969 114
ESPÈCE: WHITEINELLA BRITTONENSIS (LOEBLICH &
TAPPAN, 1961) 115
4
ESPECE: WHITEINELLA PARADUBIA (SIGAL, 1952) 115
ESPÈCE: WHITEINELLA PRAEHELVETICA (TRUJILLO,
1960) 116
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 117
FAMILLE: HEDBERGELLIDAE LOEBLICH & TAPPAN, 1961 117
SOUS-FAMILLE : ROTUNDININAE BELLIER & SALAJ, 1977 117
GENRE : PRAEGLOBOTRUNCANA BERMUDEZ, 1952 117
ESPECE : PRAEGLOBOTRUNCANA DELRIOENSIS (PLUMMER, 1931)
117
ESPECE : PRAEGLOBOTRUNCANA GIBBA KLAUS, 1960 118
ESPECE : PRAEGLOBOTRUNCANA STEPHANI (GANDOLFI, 1942)
119
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 120
FAMILLE: HEDBERGELLIDAE LOEBLICH & TAPPAN, 1961 120
SOUS-FAMILLE : HELVETOGLOBOTRUNCANINAE LAMOLDA, 1976 120
GENRE : DICARINELLA PORTHAULT, 1970 120
ESPECE : DICARINELLA ALGERIANA (CARON, 1966) 121
ESPECE : DICARINELLA HAGNI (SCHEIBNEROVA, 1962) 122
ESPECE : DICARINELLA IMBRICATA (MORNOD, 1949) 122
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 123
FAMILLE: HEDBERGELLIDAE LOEBLICH & TAPPAN, 1961 123
SOUS-FAMILLE: HELVETOGLOBOTRUNCANINAE LAMOLDA, 1976 123
GENRE: HELVETOGLOBOTRUNCANA REISS, 1957 123
ESPECE : HELVETOGLOBOTRUNCANA HELVETICA (BOLLI, 1945)
124
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 125
FAMILLE: ROTALIPORIDAE SIGAL, 1958 125
SOUS-FAMILLE: ROTALIPORINAE SIGAL, 1958 125
GENRE: ANATICINELLA EICHER, 1972 125
ESPECE : ANATICINELLA MULTILOCULATA (MORROW, 1934)
125
SUPERFAMILLE: ROTALIPORACEA SIGAL, 1958 126
FAMILLE: ROTALIPORIDAE SIGAL, 1958 126
SOUS-FAMILLE: ROTALIPORINAE SIGAL, 1958 126
GENRE: ROTALIPORA BROTZEN, 1942 126
ESPÈCE : ROTALIPORA CUSHMANI (MORROW, 1934)
PL.1, FIG.16-17/PL.5, FIG.1 127
ESPECE : ROTALIPORA GREENHORNENSIS (MORROW, 1934)
128
ESPÈCE : ROTALIPORA MONTSALVENSIS (MORNOD,
1949) 129
SUPERFAMILLE: GLOBOTRUNCANACEA BROTZEN, 1942 129
FAMILLE: GLOBOTRUNCANIDAE BROTZEN, 1942 129
SOUS-FAMILLE : GLOBOTRUNCANINAE BROTZEN, 1942 129
GENRE : MARGINOTRUNCANA HOFKER, 1956 129
ESPECE - TYPE : MARGINOTRUNCANA RENZI (GANDOLFI, 1942)
130
CONCLUSION GÉNÉRALE
131
REFERENCES BIBLIOGRAPHIQUES
134
5
Liste des Figures et des Tableaux
Figure 01 - Carte paléogéographique au passage
Cénomanien - Turonien (-93 ,5 Ma)
Figure 03 - Biozonation comparée des Foraminifères
planctoniques et des Ammonites de l'intervalle
Cénomanien - Turonien en domaine téthysien
(d'après les données de Salaj, 1980 ; Robasz.
et al., 1990, Amedro et Robaszynski, 1993 ; Thomel, 1993 et Gale
et al., 1993)
Figure 04 - Les variations relatives temporelles du taux de
production de croûtes océaniques au
Figure 05 - Situation du EAO-2. Comparaison entre la courbe
eustatique du niveau marin et la courbe de
surface de terres emmergées (d'après Jones et
Jenkyns, 2001)
Figure 06 - Modèle général
caractérisant le passage Cénomanien - Turonien
Figure 07 - modèle conceptuel montrant que les
activités hydrothermales liées à l'expansion des
croûtes
océaniques peuvent induire des EAOs (d'après Jones
et Jenkyns, 2001)
Figure 08 - Réchauffement global des eaux
océaniques au passage Cénomanien - Turonien
Figure 09 - Log lithologique et position du niveau Bonarelli
(d'après Premoli Silva et al., 1999)
Figure 10 - Variation du TOC dans la coupe de Pueblo
(d'après Ferry et al., 2003)
Figure 11 - Carte des isopaques de la Formation Bahloul
(Cénomanien - Turonien) et de ses équivalents
latéraux. (D'après les données de :
Burollet, 1956 ; Benzarti-Said, 1981 ; Zargouni, 1985 ; Ben
Ferjani et al., 1991 ; Razgallah et al., 1994 ; Layeb et
Belayouni, 1999 ; Abdallah, 2000 ;
Lüning et al., 2004)
Figure 12 - Extension des dépôts de la limite C/T
dans le domaine téthysien (Lüning et al., 2002, modifiée
concernant la Tunisie)
Figure 13 - carte paléogéographique du passage
Cénomanien - Turonien montrant le domaine téthysien.
(Philip et al., 2000)
Figure 14 - Carte de distribution des différents domaines
d'âge Cénomanien - Turonien dans la marge
provençale du bassin subalpin (Ferry et al., 2003).
Figure 15 - carte de localisation de la région de
Tajerouine
Figure 16 - cadre structural général des deux
coupes (d'après Layeb, 1990 simplifiée)
Figure 17 - Vue panoramique de la coupe CES
Figure 18 - description lithologique de la coupe CES
Figure 19 - répartition des Foraminifères
planctonisues dans la coupe de Oued Es Smara
Figure 20 - Répartition et courbe de richesse
spécifique dans la coupe CES.
Figure 21 - Répartition des fossiles,
éléments minéraux et courbe de variation du taux de
CaCO3 dans la
coupe CES
Figure 22 - Carte de situation
Figure 23 - Extrait de la carte géologique N°47 de
Bargou au 1/50000
Figure 24 - Description lithologique de la Formation Bahloul au
niveau de la coupe COK
Figure 25 - répartition des Foraminifères
planctoniques dans la coupe de Oued El Kharroub (région de
Bargou)
Figure 26 - variation d'épaisseur entre les deux coupes
(COK et CES)
Figure 27 - Abondances relatives des espèces de
Foraminifères planctoniques dans la fraction de taille
>63um de la Formation Bahloul de la coupe CES. A noter que
l'association est dominée par les Heterohelicidés tolérant
l'oxygène minimum et les Guembelitriidés marqueurs des eaux de
surface à caractère eutrophique et à salinité
relativement normale, ceci est due à une ZOM bien
développée
Figure 28 - le renouvellement faunique mis en évidence
à Oued Es Smara
Figure 29 - Abondances relatives des Genres de
Foraminifères planctoniques dans la fraction de taille
>63um de la Formation Bahloul de la coupe CES. A noter le
comportement antagoniste des
Heterohelicidés et des Guembelitriidés
Figure 30 - Les quatre phases
Figure 31 - comparaison entre la fréquence relative du
genre Heterohelix
Figure 32 - comparaison entre la fréquence relative de
Guembelitria cenomana
Tableau 1 - Variations d'épaisseur de la Formation Bahloul
dans la région de Tajerouine
Tableau 2 - Variation d'épaisseur de la Formation Bahloul
dans la région de Bargou
Tableau 3 - Les indicateurs paléoécologiques
Tableau 4 - tableau de répartition des
Foraminifères planctoniques dans les différentes niches
écologiques Tableau 5 - Fréquences relatives en pourcentage des
Foraminifères planctoniques dans la coupe CES (première
partie)
Tableau 6 - Fréquences relatives en pourcentage des
Foraminifères planctoniques dans la coupe CES (deuxième
partie)
6
PREMIÈRE PARTIE :
INTRODUCTION GÉNÉRALE
7
I - Généralités
Les dépôts enregistrant l'événement
géologique global, qui a eu lieu durant le passage
Cénomanien-Turonien (EAO-C/T : Événement Anoxique
Océanique de la limite Cénomanien-Turonien, EAO-2, ELCT :
Événement de la limite Cénomanien - Turonien ou
CTBE : Cenomanian Turonian Boundary Event des auteurs anglo-saxons),
ont été abordés sous plusieurs thèmes. Divers
travaux multidisciplinaires, qui ont fait l'objet de publications ou de
thèses, ont permis la caractérisation de cet
événements sur plusieurs plans : biostratigraphique,
paléoécologique, géochimique,
paléogéographique et océanographique (Leckie, 1984 ;
Arthure et al., 1990 ; Robaszynski et al, 1993 ; Hancock,
1993 ; Gale et al., 1993 ; Premoli Silva et Sliter, 1994) . Les
spécialistes se sont rendus compte qu'à la fois les
sédiments de bassin comme de plateforme, de l'intervalle de ce passage,
sont marqués par un enrichissement en matière organique
conservée et ont enregistré des changements faunistiques majeurs.
Sur le plan géochimique, cette matière organique est
considérée en liaison avec l'installation de conditions
hypoxiques voire anoxique. Ces changements ont eu lieu simultanément
dans différents cadres paléogéographiques (Fig. 1) et ont
été par conséquent considérés comme le
résultat de processus ou d'événements océaniques
globaux. Ces processus pourraient aussi être influencés par des
facteurs locaux (Caus et al., 1993).
Durant cet intervalle et précisément entre 93,5
et 93 Ma (Gale et al., 1993 ; Kuypers et al., 2002),
l'événement anoxique a pris de l'ampleur en s'exprimant à
l'échelle mondiale par des dépôts analogues. (Schlanger
et al., 1987 ; Haq et al., 1987).
Scholle et al., (1980) mentionnent que les
sédiments riches en matière organique d'âge
Cénomanien - Turonien ont été déposés
à travers les régions téthysienne et boréale sous
des conditions anoxiques. Un tel modèle de bassins anoxiques
évoque la présence d'un taux élevé de
productivité primaire résultant probablement d'un upwelling
(Pederson et Clavert, 1990 ; Layeb, 1990 ; Layeb et Belayouni, 1991 ; 1999),
vue la présence de sédiments riches en silice biogène ou
de radiolaires interprétés comme étant des organismes
indicateurs de tels phénomènes (De Wever et al., 1985 ;
Pederson et al., 1990). L'accumulation de tels organismes dans les
séries sédimentaires aboutit à la
8
formation de niveaux carrément siliceux comme le niveau
Bonarelli en Italie (Arthur et Premoli Silva, 1982 ; Premoli Silva
et al., 1999) ; Oued El - Kharroub, Tunisie (Turki, 1975 ; 1985 ;
Rami, 1998).
-60° -30° 0° 30° 60°

-30°
60°
30°
0°
Amérique du
Amérique du Afrique
Sud
Nord
Atlantique
Nord
P
Europe
T
Australie
Asie
-30°
-60°
0°
30°
-90° -60° -30° 0° 30°
|
Terre
mer peu profonde mer profonde
|
P: T:
|
ODP DSDP Passerelle
Tunisie
rivage actuel
|
Fig. 1 : Carte paléogéographique au passage
Cénomanien - Turonien (-93 ,5 Ma)
(Obtenue à partir de GEOMAR
map generator)
La faune de Foraminifère planctonique du passage
Cénomanien - Turonien des faciès pélagiques
déposés en milieu marin profond est dominée par des formes
opportunistes (Caus, 1993 ; Robaszynski et Caron, 1995) de la stratigie r
(r-selected) tels que les Whiteinelles, les Hedbergelles , les
Hétérohélicidés et les Guembelitriidés (Fig.
2). Les
9
espèces de Rotalipores se sont éteintes à
la fin du Cénomanien pour être remplacées par les
Helvetoglobotruncana au Turonien inférieur à moyen ainsi
que par les Dicarinelles et les Marginotruncanidés. (Caron, 1985 ;
Robaszynski et al., 1990 ; Caus et al, 1993 ; Balla et
al, 1993 ; Premoli Silva and Sliter, 1994 ; Maâmouri et
al., 1994 ; Robaszynski et Caron, 1995 ; Rami et al., 1997 ;
Rami, 1998 ; Georgescu, 2000 ; Kuypers et al., 2002).
|
ÉTAGE
|
CÉNOMANIEN
|
TURONIEN
|
|
Inférieur Moyen Supérieur
|
Inférieur Moyen Supérieur
|
|
AGE (Ma)
|
93,2
|
|
Zone à
Foraminifères
|
Rotalipora
brotzeni
|
Rotalipora R.
montsalve nsis
|
cushmani
R.
cushmani
|
W.
archaeo-
cretacea
|
Helvetoglobotruncana
helvetica
|
M.
Schneeg.
|
|
Whiteinella
W. aprica
W. archaeocretacea
W. baltica
W. brittonensis
W. paradubia
W. praehelvetica
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Dicarinella
D. algeriana D. hagni
D. imbricata
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rotalipora
R. cushmani
R. montsalvensis
|
|
|
|
|
|
|
|
Marginotruncana
M. renzi
|
|
|
|
|
|
|
|
|
Helvetoglobotruncana H. helvetica
|
|
|
|
|
|
|
Hedbergella
Cl. simplex H. delrioensis H. planispira
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Heterohelix
H. reussi
H. moremani H. navarroensis
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Guembelitria
G. cenomana G. albertensis G. sp
|
|
|
|
|
|
|
|
|
|
|
|
Fig. 2 - Aperçu sur la répartition des
Foraminifères planctoniques de l'intervalle Cénomanien -
Turonien
(d'après E.W.G.P.F., 1979.1 légèrement
modifiée)
De tel renouvellement faunique a été
enregistré dans les dépôts de plateforme. Dans ces
dépôts, les foraminifères planctoniques de stratégie
K (K-selected) (Caus et al., 1993 ;
10
Robaszynski et Caron, 1995), ont disparu à la limite
Cénomanien - Turonien pour être remplacés au Turonien
inférieur par d'autres formes de même stratégie
écologique. Dans le domaine boréal, les Rotalipores et surtout
Rotalipora cushmani et Rotalipora greenhornensis
disparaissent avant la fin du Cénomanien durant la zone à
Ammonites, Metoicoceras geslinianum, (Fig. 3) alors qu'en domaine
téthysien leur extinction est lié à l'installation d'un
milieu anoxique qui a eu lieu au début du Cénomanien
supérieur (Robaszynski et Caron, 1995). L'espèce
Helvetoglobotruncana helvetica se manifeste dès le début
du Turonien marqué par la zone d'Ammonite à Pseudaspidoceras
flexuosum (Fig. 3) alors que l'espèce Whiteinella praehelvetica
apparaît déjà à la fin du Cénomanien.
Robaszynski et al., (1990) mentionnent que les
Helvetoglobotruncana helvetica disparaissent avant la fin du Turonien
moyen.
Dans le domaine téthysien, la limite Cénomanien
- Turonien est enregistrée généralement dans une
séquence de dépôt de troisième ordre qui
débute dans la partie terminale du Cénomanien et se termine dans
le Turonien inférieur (Segura et al., 1993 ; Hardenbol et
al., 1993). Cette séquence intègre le cortège
transgressif lié à la transgression rapide de l'océan
Atlantique (Vilas et al., 1986 in Segura et al.,
1993). Vu que la monté du niveau marin était très
importante, des sédiments hémipélagiques ont
été déposés même dans les régions
proximales situées en marge de cet océan (Garcia et al.,
1987 ; Segura et al., 1993).
Sur la marge occidentale du bassin intérieur
nord-américain, durant cet intervalle, la géométrie du
prisme côtier montre que les calcaires de Bridge Creek sont largement
transgressifs sur les faciès distaux (les Shales de Hartland) du dernier
prisme sableux côtier régressif de Belle Fourche.
La transgression correspond à la reprise du
régime de flexuration du bassin d'avant-chaîne (Ferry et
al., 2003). Elle est dans ce cas d'origine tectonique et marquée
corrélativement par une recrudescence des bentonites (couches de cendres
volcaniques) et qui souligne la reprise d'activité concomitante des
volcans de la cordillère de Sevier.
11
Age
|
(Ma)
|
Étage
|
Zone à Ammonites
|
Zone à Foraminifères
|
|
92,1
92,6
93,2
94,7
96,2
|
TURONIEN
|
Sup.
|
Subprionocyclus neptuni
|
Marginotruncana schneegansi
|
|
Moy.
|
Collignoceras woollgari
|
Helvetoglobotruncana helvetica
|
|
Inf.
|
Mammites nodosoides
|
|
Pseudaspidoceras flexuosum
|
|
Watinoceas coloradoense
|
Whiteinella archaeocretacea
|
|
CÉNOMANIEN
|
Supérieur
|
Neocardioceras judii
|
|
Metoicoceras geslinianum
|
|
Rotalipora cushmani
|
|
Calycoceras naviculare
|
|
Moyen
|
Acanthoceras jukesbrownei
|
|
Acanthoceras rhotomagense
|
|
Rotalipora montsalvensis
|
|
Inférieur
|
Mantelliceras gr. Dixoni
|
Rotalipora brotzeni
|
|
Mantelliceras cantianum
|
Fig. 3 - Biozonation comparée des Foraminifères
planctoniques et des Ammonites de l'intervalle Cénomanien - Turonien en
domaine téthysien (d'après les données de Salaj, 1980 ;
Robaszynski et al., 1990, Amedro et Robaszynski, 1993 ; Thomel, 1993 et Gale et
al., 1993)
En effet, la partie médiane du Crétacé
(-124 à -90 Ma) était une période de grandes
manifestations magmatiques, avec une production énorme de larges
provinces volcaniques et de dorsales médio-océaniques (Fig.4)
(Anderson, 1994 ; Coffin et al., 1994 ; Zhao et al., 2004). A
partir de ces données plusieurs auteurs admettent que les
émissions du dioxyde de carbone CO2 volcanique
libérés par les larges provinces volcaniques durant cette
période du crétacé, ont pu causer le réchauffement
exceptionnel du Crétacé supérieur (Tarduno et
al., 1998, Zhao et al., 2004) et par la suite provoquer
deux à sept Événements Anoxiques
Océaniques (EAOs) durant la partie médiane du
Crétacé supérieur (Leckie et al., 2002 ; Zhao
et al., 2004). En outre, le paléochamps magnétique
durant cette période était d'une manière non
caractéristique stable de telle manière qu'il n'a pas
changé durant près de 40 Ma (de l'Aptien au Santonien : de -121
à -83 Ma) (Zhao et al., 2004).

Crétacé
Tertiaire
(%) changement du taux de production de croûte
Jur
-20
60
C-T
40
20
0
OAE-2
12
140 120 100 93.5 80 60 40 20
Age (Ma)
Fig. 4 - Les variations relatives temporelles du taux de
production de croûtes océaniques au niveau des rides
médio-océaniques à cause des activités volcaniques
intenses (d'après Jones et
Jenkyns, 2001)
Le Programme de Forage Océanique (ODP : Ocean
Drilling Program) et son prédécesseur le Project de Forage
dans les Mers Pélagiques (DSDP : Deep See Drilling Project) ont
beaucoup contribué à la compréhension des
événements crétacés et surtout, ils ont fourni de
nombreuses données permettant la reconstitution des conditions et des
scénarios déroulés.
1 - Les caractéristiques générales de
l'Événement Anoxique Océanique durant la limite
Cénomanien - Turonien (EAO-2)
Le concept « Événement Anoxique
Océanique » (EAO) en général a été
crée par Schlanger et Jenkyns (1976) en définissant deux
épisodes majeurs au Crétacé
13
d'appauvrissement des eaux de fond en oxygène. De
telles conditions ont été à l'origine de formation des
couches riches en matière organique. Ces événements
globalement décelables par les dépôts de Black Shales ont
été appelés les « Événements
Anoxiques Océaniques » (EAOs) parce qu'ils sont
considérés comme des périodes de crise d'oxygène
dans des océans relativement chauds (Forester et al., 2002). En
effet, Le Cénomanien Turonien représente une période dans
l'histoire de la Terre qui est en contraste avec le climat global
récent. En général, les paléotempératures et
la surface d'inondation maximale durant cette période de temps
coïncidaient avec un volcanisme intense et une création de
croûte océanique. (Forester et al., 2002).
Schlanger et Jenkyns (1976) ont constaté que le second
événement océanique (EAO-2) coïncidait avec la limite
Cénomanien - Turonien (Fig. 5). La caractéristique essentielle de
cet événement qui le distingue fondamentalement des autres EAOs,
est son extension globale à l'échelle planétaire (en
Europe, en Afrique, en Amérique, en Australie et en Asie) et sa courte
durée (intervalle de temps très court : de 93,5 à 93 Ma,
Kuypers et al., 2002).

300
Tertiaire
Crétacé
O A E -2
Land area
Sea level
150
140 130 120 110 100 90 80 70 60 50 40 30 20 10 (My)
Crétacé inférieur
Crétacé supérieur
Paléogène
Néogène
200
100
0
Niveau marin relatif (m I
110
120
130
140
Terre emergée 1 0 k m 2
6
Fig. 5 - Situation du EAO-2. Comparaison entre la courbe
eustatique du niveau marin et la courbe de la
surface de terres
emmergées (d'après Jones et Jenkyns, 2001)
Toutefois, la précision chronostratigraphique de cet
événement EAO-2 est fort controversée. En effet, des
micropaléontologistes et des géochimistes ainsi que des
sédimentologues ont fourni de multiples efforts pour mieux cerner cet
événement. Muller et al., (1984), dans un rapport de
synthèse sur les dépôts pris par le programme international
de forage océanique pour la recherche scientifique ODP, ont
mentionné que cet événement coïncide avec la zone de
Foraminifère planctonique à Whiteinella
14
archaeocretacea. Cette biozone du passage
Cénomanien Turonien est suivie par celle à
Helvetoglobotruncana helvetica du Turonien inférieur à
moyen (Fig. 6). Robaszynski et al., (1995) avaient aussi situé
cet événement dans la zone à Whiteinella
archaeocretacea en
|
Age
|
Foraminifères Planctoniques
|
Ammonites
|
Radiolaires
|
Niveau marin+
250 aao ,w
|
b Séquences
3 2nd1
|
8t(%0)&
2 3 4-4
|
O (%0)
-3 -2 -1
|
TOC (%)
0 4.5 9
|
|
CénomanienTuronien
93,5
|
D. concavata
|
S. neptuni
|
|
UZA - 3
|
8944
Tu4
90.74 Tu3
Tu2
92.36
Tu1
93.09
Ce5
|
R
T
R
T
|
13C Ç U
|
EAO-2
|
|
|
|
M. schneegansi
|
|
H. helvetica
|
|
C. woollgari
sompedia
Alievium
superbum
A
um m
eh
W. coloradoense
|
M. nodosoides
|
|
|
|
|
|
|
|
|
91,5
|
|
|
|
|
|
Dorypyle silviae
|
Guttacapsa biacuta
|
|
|
Ce4
96.15
Ce3
Ce2
|
|
Patellula
|
|
A. rhotomagense Patellula
|
nakasekoi
|
|
R. reicheli
|
|
R.
globotruncanoides
|
spica
|
|
|
Ce1
97.95
|
|
|
Dorypyle anisa
|
Dorypyle anisa
|
Fig. 6 - Modèle général
caractérisant le passage Cénomanien - Turonien
indiquant cette fois que cette zone intervallaire
caractérise le passage Cénomanien - Turonien. D'autres
auteurs en se basant sur la répartition des nannofossiles calcaires, le
situaient dans la zone à Lithraphidites acutum du
Cénomanien supérieur (Muller et al., 1984 ; Balla et
al., 1993 ; Bengston, 1996). Arthur et Premoli Silva (1982), indiquent que
l'événement anoxique du passage Cénomanien - Turonien dans
le niveau Bonarelli en Italie ne dépasse pas le 0,7 Ma. Cette
interprétation sera plus tard controversée par Arthur et
al., (1990) en signalant que cet événement est de 0,5 Ma (de
-91 à -90,5 Ma). Plus tard, Gale et al., (1993) ont
débattu cet âge pour dire que l'Événement Anoxique
Océanique du Passage Cénomanien - Turonien débute à
-94,3 Ma et se termine à -93,5 Ma. Pour ces auteurs la durée de
cet événement est de 0,8 Ma et la limite Cénomanien -
Turonien se situe alors à -93,5 Ma.
Certains auteurs mettent l'accent sur la
paléotopographie et le rôle de la paléogéographie
pour expliquer le modèle de dépôt du faciès riche en
matière organique du passage Cénomanien Turonien ou Black Shales
(Layeb, 1990 ; Layeb et Belayouni 1999) mais pour d'autres ce modèle
leur parait insuffisant pour expliquer l'anoxie dans des mers
épicontinentales ouvertes (Nzoussi-Mbassani, 2003).
Cet Événement EAO-2 est
caractérisé aussi par une grande transgression rapide (Fig. 6).
Nzoussi-Mbassani (2003) a mentionné que à part l'influence
suggérée par plusieurs auteurs de la productivité
organique, cette transgression semble induire à son apogée
à la limite C/T une limitation du taux sédimentaire et que
l'expansion du domaine océanique (Fig.7) aurait induit un blocage des
apports terrigènes loin des bassins sédimentaires
15
océaniques. Ce qui explique la non dilution de la
matière organique par les apports considérables de constituants
minéraux.
En ce qui concerne le passage C/T, deux modèles ont
été décrits afin d'expliquer les caractéristiques
de l'EAO-2 et le dépôt des Black Shales riches en Carbone
Organique. Le premier modèle est basé sur le concept de la
préservation de cette M.O qui caractérise un milieu relativement
euxinique (Rayan et Cita, 1977 ; Bralower et al., 1987, Kuhnt
et al., 1990). Ce modèle euxinique évoque la
présence de deux couches d'eau à densités
différentes, l'une supérieure, donc peu dense et oxique, alors
que la deuxième est plus inférieure et anoxique très riche
en H2S. En général, ce modèle nous renseigne
sur l'extension de l'anoxie vers les eaux les plus profondes voire même
abyssales (Smith et al., 2001 ; Kuypers et al., 2002 ;
Nzoussi Mbassani, 2003), vu les teneurs élevées du Carbone
Organique Total (COT) dans les forages de l'Atlantique : DSDP, sites 105 et
603B (Kuhnt et al., 1990 ; Kuypers et al., 2000 ; 2002). Le
deuxième modèle est basé sur le concept d'une
productivité primaire élevée à fin d'expliquer les
teneurs élevées de COT dans les Black Shales (Layeb, 1990 ; Layeb
et Belayouni, 1989 ; 1999), conformément aux idées de Schlanger
et Jenkyns (1976) ; Jenkyns (1980)
La cause de EAO-2 est considérée comme
liée aux changements du climat et du niveau marin (Jenkyns, 1980 ;
Arthur et al., 1990 ; Huber et al., 1999). Les traits du
changement du niveau de la mer autour de la limite Cénomanien - Turonien
ont été examinés par plusieurs auteurs (Schlanger et
Jenkyns 1976 ; Herbin et al., 1986 ; Haq et al., 1987 ; Hart
et al, 1993 ; Lüning, 2001), mais la relation exacte entre le
changement du niveau marin et les autres processus
paléogéographiques pendant cette limite est encore
controversée. Cela vient de la difficulté de la reconstitution
des changements du niveau marin à une échelle de temps de l'ordre
de 100 ka (Yasukochi et al., 2001). En effet, ces auteurs ont
déduit que le niveau Oyubari (à Hokkaido, au Japon, d'âge
Cénomanien supérieur à Turonien inférieur)
représente une séquence à intercalations de petits niveaux
turbiditiques d'épaisseur centimétrique et que la
fréquence de ces turbidites sont de périodicité de l'ordre
de 1000 ka, 100 ka, 40 ka et de 20 ka coïncidant avec les cycles de
Milankovic. L'intervalle de la limite Cénomanien - Turonien a
été aussi étudié dans le Site 1050 à
Blake Nose (Océan Atlantique) par Huber et al., (1999)
et par Bellier et al., (2000).
16
Fig. 7 - modèle conceptuel montrant que les
activités hydrothermales liées à l'expansion des
croûtes
océaniques peuvent induire des EAOs (d'après
Jones et Jenkyns, 2001)

Grands Flux
des éléments
nutritifs
Cycle
hydrologique
accéléré
Grande expansion de croûte océanique et peut
être flux intense de CO2 volcanique
Réchauffement global
Upwellings intenses
Haute productivité biologique dans les océans
Production
croissante des
eaux chaudes et
salées dans les mers peu profondes
Fumées issues des activités
hydrothermales
induisent
la fertilisation du Fer
Activité volcanique intense induite par une production
de croûte océanique
Dans ce travail, les auteurs se sont appuyés sur les
résultats de l'analyse des isotopes stables de foraminifères
planctoniques et benthiques non altérés (Huber et al.,
1999). Ils ont constaté une baisse importante des valeurs de
?18O des Foraminifères benthiques indiquant une
élévation importante des paléotempératures des eaux
bathyales (Fig.8). En
17
effet, ils ont estimé que la
paléotempérature de ces eaux bathyales était
déjà élevée (15°C) avant la limite C/T.
Néanmoins elle atteignit approximativement 19°C au niveau de
l'événement EAO-2. Ce réchauffement des eaux profondes ne
paraît pas être influencé par les températures de
l'eau de surface alors qu'un effet de serre s'était propagé
à travers les eaux de fond. Ainsi, ces auteurs suggèrent que le
réchauffement a pu être responsable de l'extinction des genres de
Foraminifères planctoniques de subsurface tel que Rotalipora
(Huber et al., 1999 ; Bellier et al., 2000) et que la
sédimentation du carbone organique, pendant l'Événement
EAO-2, a séquestré le CO2 atmosphérique et a
initié un effet de serre inverse (Clarke, 2003).

Fig. 8 - Réchauffement global des eaux
océaniques au passage Cénomanien - Turonien
En outre, la limite Cénomanien - Turonien a
été abordée aussi sur le plan chimiostratigraphique
(Accarie et al., 1999) et isotopique concernant les isotopes
légers 13C et 18O (Schlanger et al., 1987 ; Hart
et al., 1993).
Fisher et al., (2003) ont réalisé une
analyse géochimique et isotopique des Foraminifères des
sédiments de l'intervalle Cénomanien - Turonien provenant du Site
ODP 762. Ils ont mis en évidence l'enregistrement de
l'événement EAO-2 dans cette région et ont confirmé
sa liaison d'une part avec un réchauffement global (selon les
données de ä18O) et d'autre part avec une
productivité primaire élevée (selon les données de
ä13C). Keller et al., (2001) en se basant sur la
lithologie, les isotopes stables et les foraminifères planctoniques de
la coupe de Eastbourne en Gun Gardens (Angleterre Sud-Est), ont indiqué
que l'intervalle du passage C/T est marqué par une
élévation du niveau marin . Les auteurs ont constaté aussi
que l'augmentation des valeurs de ?13C déterminant une
productivité primaire, caractérise un intervalle de temps assez
long débutant de la zone à Rotalipora
18
cushmani et couvrant toute la zone à
Whiteinella archaeocretacea. Cette augmentation de la
productivité a fort influencé le comportement des
foraminifères planctoniques. Ainsi, ces auteurs ont distingué
deux phases de renouvellement faunistique (I et II) qui ont
coïncidé avec les pulsations positives du ?13C. La
première pulsation de ?13C (augmentation de 1,8 %o)
coïncide avec l'extinction des Rotalipores qui vivent loin de la ligne de
rivage vers le large et l'abondance des espèces d'Heterohelix
tolérant un faible taux d'Oxygène. Selon ces auteurs ceci
serait en liaison avec une baisse du niveau marin simultanée à
une augmentation de la productivité et une expansion de la zone à
oxygène minimum. La deuxième phase coïncide avec la
disparition de presque 50% des assemblages de Foraminifères
planctoniques, au profit des habitants de surface (Guembelitria et
Whiteinella), qui serait en liaison avec l'augmentation de la
productivité primaire et l'expansion de la zone à oxygène
minimum associées à un réchauffement climatique.
D'après ces mêmes auteurs, cette seconde phase de
renouvellement faunistique est stratigraphiquement corrélée avec
l'Événement Anoxique Océanique global (EAO-2) en Italie
(le niveau de Bonarelli) et en Tunisie (la Formation
Bahloul), alors que la première phase se corrèle avec
les premiers niveaux riches en matière organique dans la partie
supérieure de la zone supérieure de Rotalipora cushmani
de ces régions.
Pour Arthur (2003), l'augmentation isotopique de
ä13C n'est pas clairement correlable ni avec le dégazage
du CO2 volcanique ni avec la libération du méthane
De même, pour Bralower (2003), l'événement
EAO-2 enregistré dans les sédiments du Site 174AX (du forage ODP)
ne serait pas lié à la dissociation de l'hydrate du
méthane, qui a pu épuiser l'Oxygène dans le fond des
océans.
2 - La Coupe Type et les Coupes les mieux
étudiées au monde concernant la limite Cénomanien -
Turonien
a - Le Niveau Bonarelli :
En Italie, le niveau Bonarelli ou Livello Bonarelli
défini un enregistrement lithologique spectaculaire exceptionnel de
l'EAO-2. De ce fait, il représente la localité type pour la
limite Cénomanien - Turonien. Il a été défini dans
le bassin de Umbria - Marche. Il mesure approximativement 1,5 m et il
correspond à des alternances de marnes noires
19
(black shales) et des couches sileuses riches en radiolaires
(Fig.9). Il est caractérisé aussi par une accumulation et une
bonne préservation de carbone organique. Les couches siliceuses
s'avèrent exclusivement dépourvues de carbonates. La coupe de
référence de ce niveau est représentée dans la
Valle del Bottaccione. On parle généralement de
l'événement de Bonarelli (Bonarelli Event) qui se
confond avec l'événement anoxique : OAE-2).

Fig. 9 - Log lithologique et position du niveau Bonarelli
(d'après Premoli Silva et al., 1999)
Concernant les Foraminifères planctoniques, les
espèces de Rotalipores et Globigerinelloides bentonensis
disparaissent vers la base du niveau Bonarelli. Les Foraminifères
planctoniques de petites tailles telles que Heterohelix moremani, H.
reussi, Hedbergella planispira deviennent abondant dans le niveau
Bonarelli. Il est attribué à la zone de
Foraminifère planctonique à Whiteinella archaeocretacea.
(Arthur et Premoli Silva, 1982 ; Premoli Silva et Sliter, 1994 ; Premoli Silva
et al., 1999 ; Adatte et al, 2003 ; Coccione, 2003 ; Coccione
et Luciani, 2003, Musavu-Moussavou et al, 2003)
20
b - La coupe de Pueblo :

Fig. 10 - Variation du TOC dans la coupe de Pueblo
(d'après Ferry et al., 2003)
Dans la Vallée de Colorado, les formations
déposées dans le Bassin du Western Interior au cours du
Crétacé supérieur renferment de nombreux horizons de
cendres volcaniques (bentonites) facilement reconnaissables et
corrélables à l'échelle du bassin. Une dizaine de niveaux
de bentonites ont été reconnus dans l'intervalle du passage
Cénomanien - Turonien (Fig. 10), sur plus de 1000 km depuis la bordure
occidentale du bassin (Arizona) jusqu'à sa marge orientale stable
(Kansas). Ces évennements semblent être de courtes durées
et s'étendent sur une période de 2 millions d'années,
comprise entre les zones
21
d'ammonites à Sciponoceras gracile et à
Mammites nodosoides. (Ferry et al., 2003, Desmares et
al., 2003)
II - La limite Cénomanien - Turonien en
Tunisie
Cet intervalle est représenté en Tunisie
septentrionale et centro - septentrionale par une entité lithologique
essentiellement formée d'alternances marno-calcaires. Les calcaires y
sont laminés, cartonnés, de texture micritique, noirâtres
ou blanchâtres et se débitant en plaquettes. Les marnes y sont
indurées ; elles sont aussi feuilletées et noirâtres. Ces
alternances marno-calcaires passent latéralement vers le sud (Tunisie
méridionale et
centro-méridionale) à des dépôts
carbonatés (Fig. 11) (calcaire ou dolomie)
caractéristiques
d'un environnement de type plateforme carbonatée (calcaire et
dolomitique) (Abdallah, 1995 ; Rasgallah et al., 1994 ; Layeb et
Belayouni, 1999).
Ces alternances marno-calcaires ont été
repèrées et nommées « marnes schisteuses » par
Pervinquière (1903) ou « Première masse calcaire » par
Berthe (1949) ou « calcaire en plaquette » par Schijfsma (1955).
Elles ont été attribuées à la « Formation
Bahloul » ou à « l'horizon Bahloul » par Burollet et
al., (1954) puis Burollet (1956). La localité-type de cette
formation, Oued Bahloul est située au Sud Est du Makthar. En
décrivant cette Formation, ces auteurs précisent qu'elle
démarre dans la base du Turonien et qu'elle correspond à un
horizon stratigraphique caractérisé par l'expansion du genre
Guembelina EGGER. (Dalbiez, 1956). Selon son auteur (Burollet, 1956),
la Formation Bahloul est formée par des calcaires finement lités
d'aspect feuilleté ou cartonné, souvent marneux et alternant
parfois avec des marnes (alternances marno-calcaires) brunes, gris foncé
ou noires et dégageant une odeur bitumineuse lors de la cassure. Les
surfaces de stratification de cette formation présentent des empreintes
aplaties d'Ammonites et parfois des restes de poisson. L'auteur mentionne que
cette entité n'est pas résistante à l'érosion
toutefois, elle forme un « ressaut dans la topographie entre les deux
larges ensembles
8° 9° 10° 11°

37°
36°
34°
35°
35°
30 Les isopaques de Fm. Bahloul
40
Algerie
10 Fahdène/Zebbag supérieur
avec Guettar
Mer Méditerranée
* Localité type "Oued Bahloul"
Touzeur
?
Bahloul Black shale facies
C-T lithofacies zones
Puit pétrolier Coupes C-T
Bahloul absent
La coupe CES
Tebessa
Légende
Kalâat Senan
?
80
Kasserine
Gafsa
Tajerouine
Tabarka
Kef
Jendouba
Évaporites & dépôts
détritiques
20
?
Kebili
Carbonates
*
Bizerte
Skhira
Kairouan
0 Échelle 100 km
22
Gabès
Bahloul
pauvre en M.O
Tunis
Golfe de Gabès
Bioaccumulation
West Gabes well
Morjene-1 well
Golfe de Hammamet
Medenine
30
Kerkenah West Permit
Sfax
Djerba
Mahdia
Kerkenah
N
37°
35°
34°
35°
36°
8° 9° 10° 11°
22
Fig. 11 - Carte des isopaques de la Formation Bahloul
(Cénomanien - Turonien) et de ses équivalents
latéraux.
(D'après les données de : Burollet, 1956 ; Benzarti-Said, 1981 ;
Zargouni, 1985 ; Ben Ferjani et al.,
1991 ; Razgallah et al., 1994 ; Layeb
et Belayouni, 1999 ; Abdallah, 2000 ; Lüning et al., 2004)
23
argilo-marneux qui l'entourent » (idem,
p.81) tout en reposant directement sur la Formation Fahdène (Burollet,
1956) et en étant surmontée par la Formation Kef (Fournié,
1978) qui sont de nature essentiellement argileuse. Quant à
l'attribution chronostratigraphiqe de cette formation est controversée.
Dans le même document, Burollet (idem), mentionne que la
Formation Bahloul, comprise entre les Formations Fahdène et Aleg, est du
passage Cénomanien - Turonien (fig. 19 - 20, p. 86), alors que dans le
tableau de corrélation des unités stratigraphiques en Tunisie
Centrale, la Formation Bahloul est attribuée au Turonien basal
conformément à la définition qu'il a proposé en
page 81.
Jauzein (1967) a décrit lui aussi la formation Bahloul
dans de nombreuses coupes qu'il a levées en Tunisie
centro-septentrionale. Cet auteur considère que la base de cette
entité est du Turonien basal ou du Turonien inférieur mais sans
donner d'arguments paléontologiques pour fonder une telle attribution
chronostratigraphique.
Salaj (1976 ; 1980), a signalé le caractère
typique des calcaires en plaquette dits calcaires du Bahloul et leur abondance
en Tunisie septentrionale. Toutefois, en ce qui concerne certaines
régions (Zaghouan, Pont du Fahs, Jebel Fkerine et Enfida), l'auteur a
noté la réduction voire l'absence de ces calcaires. En outre, en
se basant sur la distribution des Foraminifères planctoniques, il
précise que selon les régions la Formation Bahloul peut
être du passage Cénomanien - Turonien ou du Turonien basal.
Tlatli et al., (1977) ont eu le mérite de
découvrir le « Complexe récifal » de Borj Cédria
(Potinville). Il s'agit d'un dépôt à faune mixte (des
Foraminifères planctoniques et des Rudistes). Ces auteurs ont conclu que
ce complxe est d'âge Cénomanien - Turonien.
Benzarti-Saïd (1981) a reconnu la Formation Bahloul dans
le forage de Djebiniana 1 (DJB.1) réalisé dans le Sahel Tunisien
(Tunisie Orientale). Elle note que les calcaires de ladite Formation sont
constitués d'une biomicrite brune riche en matière organique et
contenant des Hedbergelles, des Pithonelles et des filaments. Elle attribue la
Formation Bahloul à la base du Turonien en se basant sur les
associations d'Ostracodes.
24
Bismuth et al., (1981) attribuent la Formation
Bahloul affleurant dans la région de Jebel Bou Zer (Tunisie Centrale) au
Turonien basal en se basant sur les associations de Foraminifères et
d'Ostracodes. Dans la même année Boltenhagen arrive à la
même conclusion en publiant une carte des faciès du Turonien
montrant la répartition des calcaires Bahloul et leurs
équivalents latéraux.
Gargouri (1981 ; 1983) a essayé d'effectuer des
analyses stratigraphique et paléoécologique sur le
Cénomanien de la Tunisie Centrale. De ce fait, elle a surtout
établi une étude d'ordre phylogénétique,
taxonomique et paléoécologique sur l'espèce de
Foraminifère benthique Thomassinnella punica (SCHLUMBERGER)
qu'elle considère marqueur de l'étage Cénomanien. Ainsi,
vu que cette espèce figure aussi dans les calcaires du Bahloul de
Tunisie Centrale, elle conclut que ces calcaires sont du Cénomanien
supérieur.
Phillip (1981 ; 1982) a étudié spécialement
les Rudistes du « Complexe récifal ».
Bellier (1983) dans son travail sur les Foraminifères
du Crétacé superieur, a abordé du point de vue
biostratigraphique la Formation Bahloul du Jebel el Oust (région de Bir
M'cherga). Il a proposé une biozonation basée sur les
Foramininifères planctoniques. Concernant le passage Cénomanien -
Turonien, il précise qu'il est caractérisé par la zone
à Whiteinella aprica.
Marie et al., (1984) en s'intéressant à
la paléogéographie de la Tunisie, ont publié une carte
paléogéographique du Turonien en se basant sur l'extension des
dépôts de la Formation Bahloul et de ses équivalents
latéraux. Cette carte a été ultérieurement
améliorée et modifiée par de nombreux autres travaux
(Layeb, 1990 ; Ben Ferjani et al., 1991).
Orgeval et al., (1986) ont mis en évidence
l'intérêt minier de la Formation Bahloul et sa richesse en
sulfures de Pb, Zn, S, notamment dans le gisement de Bougrine (Tunisie Nord
Occidentale).
25
Dali-Rossot (1987), en se basant sur les
Calcisphaerulidés, a noté que la Formation Bahloul n'est pas
synchrone. Elle peut être du Cénomanien terminal - Turonien basal
comme au Jebel Bou El Ahnèche ou du Turonien basal comme au Jebel Sif El
Kohol (Tunisie Centrale). Le même auteur a indiqué que cet
intervalle durant lequel le climat était chaud, le dépôt de
la Formation Bahloul correspond à une transgression majeure.
Ben Haj Ali (1987) a effectué une étude
biostratigraphique du Crétacé (Aptien - Cénomanien) dans
la Région de Téboursouk (Tunisie Septentrionale) en se basant sur
les Foraminifères planctoniques. Dans ce travail, elle a reconnu le
faciès typique de la Formation Bahloul qu'elle a attribué au
Turonien basal.
Dans la même année, Batik et al.,
(1987), dans leur étude sur les dépôts
crétacés du Jebel Trozza (Tunisie Centrale), ont reconnu la
Formation Bahloul composée de calcaires en plaquettes feuilletés,
de couleur gris foncé, et riches en matières organique. Ces
calcaires ont livré d'après ces auteurs une association de
Foraminifères composée de Dicarinella imbricata, D. hagni,
Whiteinella gegantea, W. paradubia, W. aprica, Heterohelix globulosa.
Cette association est attribuée au Turonien inférieur.
Abdallah (1987 ; 1995 ; 2000) a signalé la
présence du faciès Bahloul dans la chaîne Nord des
Chotts.
Par la suite, Layeb et Belayouni (1989 ; 1991 ; 1999) et Layeb
(1990) ont réalisé une étude géochimique
approfondie sur les faciès riches en matière organique de la
Formation Bahloul en Tunisie Centro-Septentrionale. En établissant une
carte des isopaques de ladite Formation, ils ont dégagé un
modèle de dépôt conduisant à une bonne roche
mère de pétrole. De ce fait, Ils ont attribué la Formation
Bahloul au passage Cénomanien - Turonien.
Robaszynski et al., (1990 ; 1993) ont entrepris une
étude sur la Formation Bahloul de plusieurs coupes levées dans la
région de Kalâat Es Senan (Tunisie Centrale). L'âge de
ladite Formation a été discuté en se basant d'une part sur
les Ammonites qu'ils ont récoltées dans cette formation et
d'autre part sur les associations de Foraminifères
26
planctoniques et de nannofossiles calcaires. Ces auteurs
concluent que ladite formation est d'âge Cénomanien
supérieur.
En (1991) Burollet et Robaszynski décrivent de «
gros bancs de calcaires » à la base de la Formation Bahloul qu'ils
intègrent dans ladite Formation des secteurs de Kalâat Es Senan et
d'Oued Bahloul. Ces « gros bancs » sont de natures gréseuses
et riches en bioclastes.
Au cours de la même année Matmati et al.,
(1991) ont publié sur le membre Gattar, qui est un
équivalent latéral d'une partie de la Formation Bahloul,
affleurant dans la chaîne Faïd - Chaabet El Attaris (Axe Nord Sud :
Tunisie Centro - orientale). Dans ce travail, les auteurs attribuent ce membre
au passage Cénomanien - Turonien.
Par la suite, Hardenbol et al., (1993) ont repris les
analyses séquentielle et sédimentologique sur la Formation
Bahloul dans la région de Kalâat Es Senan. Ils ont
précisé que cette formation s'était déposée
au cours d'un cycle transgressif.
Une année plus tard, Chaâbani et al.,
(1994) ont découvert un horizon riche en matière organique
à cachet Bahloul au sein d'une dalle carbonatée de 80 m
d'épaisseur mise en équivalence par les auteurs avec le membre
Gattar (équivalent latérale d'une partie de la Formation Bahloul
en Tunisie Centro - méridionale) au Jebel Berda (Tunisie
centro-méridionale).
Maâmouri et al., (1994) ont
étudié plusieurs coupes levées dans différentes
localités où affleure la Formation Bahloul : Koudiat Nahala (Garn
Halfaya : Feuille Tajerouine), Jebel Bou Grine (les Salines), J. Es Seif
(Ouergha), Koudiat Guenaoua, Hammam Mellègue (El Kef), Kalâat Es
Senan, Melalis (Nébeur) et même dans sa localité type (Oued
Bahloul). En se basant sur la distribution des Foraminifères
planctoniques, les auteurs attribuent un âge Cénomanien terminal -
Turonien basal à cette formation en éliminant les gros bancs de
base de ladite Formation. Les auteurs précisent que les Ammonites
récoltées dans la coupe de Oued Bahloul, Calycoceras
naviculare, Pseudaspidoceras cf. angulanum recueillies
dans le niveau OB 8 sont considérées appartenant à la Zone
à Metoicoceras geslinianum du Cénomanien
supérieur, alors que Eucalycoceras gothicum, Mantelliceras
villei ; Fagesia sp, Pseudaspidoceras flexuosum, Mammites
sp, Kamerunoceras sp.,
27
déterminent le Turonien inférieur. Quant
à la Zone à Neocardioceras judii du Cénomanien
sommital n'a pas été reconnue.
Sur le plan séquentiel, les mêmes auteurs
admettent que les conditions d'anoxie coïncidant avec
l'événement global du passage Cénomanien - Turonien
(EAO-2) ont été réalisées postérieurement
à l'augmentation globale du niveau marin de la période
transgressive déterminant le cycle eustatique global UZA-2.5 de Haq
et al., (1987), sans dépasser toutefois la période
d'inondation maximale induite il y a 91 Ma. D'ailleurs ces auteurs
précisent que la base de la séquence de troisième ordre
est bien soulignée par un niveau conglomératique.
Au cours de la même année, Ben Haj Ali et
al., (1994) ont confirmé que la Formation Bahloul dans sa
localité type est d'âge Cénomanien - Turonien en se basant
d'une part sur les Ammonites étudiées par l'un des signataires
(J. W. Kennedy) et d'autre part sur les Foraminifères planctoniques. Ces
auteurs ont identifié la zone à Calycoceras guerangeri
indiquée par Calycoceras sp et l'association des
Foraminifères planctoniques parmi lesquels Rotalipora cushmani, R.
greenhornensis. Puis la Zone à Metoicoceras geslinianum et
la Zone à Neocardioceras judii qui caractérisent le
Cénomanien sommital où la microfaune est pauvre et
représentée essentiellement par Whiteinella spp., des
Dicarinelles et Hedbergella spp.. Ces auteurs indiquent que la base du
Turonien est soulignée par l'apparition du genre Fagesia alors
que Praeglobotruncana helvetica n'apparaît que plus haut au sein
des marnes de la Formation Kef.
En outre, Razgallah et al., (1994) ont accomplis un
travail sur le passage Cénomanien - Turonien de la Tunisie
Centro-méridionale. Ils ont apporté des précisions sur le
membre Gattar et les faciès intermédiaires entre les deux
unités lithologiques Formation Bahloul et Membre Gattar en admettant que
le faciès de la Formation Bahloul n'affleure que dans le massif de
Chemsi, le reste du secteur étudié est caractérisé
par le développement du membre Gattar. En se basant sur les Ammonites et
les Foraminifères, ces auteurs concluent que le membre Gattar est
d'âge Turonien basal.
Bagga et al., (1995) ont signalé l'existence
de récifs d'âge Cénomanien - Turonien à Borj
Cédria. Ces auteurs indiquent que ces récifs correspondent en
effet, à l'établissement de
28
deux cycles de sédimentation récifale locaux et
sont formés essentiellement par des débris de Rudistes.
Kassaâ et al., (1995) en effectuant une
étude minéralogique dans la région de Garn Halfeya
(Tunisie Nord occidentale), rajoutent que le passage Cénomanien -
Turonien représenté par des laminites à faciès de
Black Shales caractéristique de la Formation Bahloul, contiennent des
fissures miniralisées riches en calcite et en galène
témoignent d'une tectonique synsédimentaire.
Quelques années plus tard, Rami (1998) dans son travail
de Thèse intitulé « Stratigraphie,
micropaléontologie et environnements de dépôt du
Crétacé supérieur de la Tunisie Centro-Septentrionale
» a abordé entre autre les dépôts de la Formation
Bahloul en indiquant son milieu de dépôt en se basant sur la
répartition des Foraminifères planctoniques en Tunisie centro -
septentrionale, nord orientale et septentrionale. Il a précisé
que cette Formation s'étend sur l'intervalle des Zones à
Rotalipora cushmani, Whiteinella archaeocretacea et
Helvetoglobotruncana helvetica.
Ensuite, Troudi et al., (1999) dans leur analyse
séquentielle de la Formation Bahloul en Tunisie centrale, confirment que
cette formation riche en matière organique serait déposée
au cours d'un cycle transgressif.
Accarie et al., (1999) et Caron et al.,
(1999) ont effectué des analyses chimiostratigraphiques,
cyclostratigraphique et biostratigraphique sur les dépôts riches
en matière organique de la Formation Bahloul dans la région de
Kalâat Es Senan et dans sa localité Type : Oued Bahloul. Ils ont
conclu que ces faciès typiques à Black Shales représentent
l'enregistrement de l'Événement Anoxique Océanique global
du passage Cénomanien - Turonien en Tunisie. Caron et al.,
(1999) ont ajouté que la Formation Bahloul dans sa localité
type montre une répétition cyclique d'une séquence
enregistrant deux paléoenvironnements. Un milieu eutrophique mais
anoxique sur le fond (les calcaires noirs finement laminés) et un milieu
oligotrophe et faiblement oxygéné sur le fond (les marnes
bioturbées). Ces mêmes auteurs admettent que la Formation Bahloul
serait déposée lors d'un grand cycle d'excentricité du
globe de durée égale à 400 ka.
29
Pour Nederbragt et Fiorentino (1999), la coupe d'Oued
Mellègue (région d'El Kef, Tunisie Nord Occidentale) levée
à intervalles serrés, a fourni des renseignements
biostratigraphiques de haute résolution sur l'Événement
Anoxique Océanique du Cénomanien sommital (EAO-2) en milieu
bathyal supérieur et pélagique à
hémiplégique. L'analyse isotopique de ä13C et
ä18O effectuée sur les dépôts de la
Formation Bahloul du passage Cénomanien - Turonien ainsi que l'analyse
des associations d'Hétérohélicidés sont en faveur
du développement des conditions anoxiques maximales, bien
exprimées dans cette coupe. Ils rajoutent que la diversité des
associations est très faible dans les sédiments de la limite
Cénomanien - Turonien. Cette faible diversité est
attribuée par ces auteurs à l'installation de conditions hostiles
mais également à la dissolution des tests des
Foraminifères au cours de la sédimentation de la Formation
Bahloul. Une baisse dans l'abondance des indicateurs de haute
productivité parmi les nannofossiles pourrait aussi être le
résultat de cette dissolution. Les mêmes auteurs ont
confirmé que la Formation Bahloul s'était déposée
au cours d'une période de haut niveau marin majeur. A la même
période des sédiments clastiques ont été
piégés en domaine de plateforme. Ultérieurement au cours
du Turonien durant la période de dépôt du membre Annaba
(Burrolet, 1956), les conditions océaniques hostiles s'estompent et la
diversification des associations de foraminifères planctoniques et des
nannofossiles augmente.
Srarfi (1999) a attribué un âge Cénomanien
- Turonien à la Formation Bahloul affleurant dans les régions des
Jebels Sekarna et Bargou.
Ferry et al., (2003) dans leur contribution sur la
crise de la limite Cénomanien - Turonien en Tunisie méridionale
(bassin de Tataouine), indiquent que la barre dolomitique Gattar, localement
à rudistes et madréporaires, est transgressive. Plus au Nord dans
la Chaîne des Chotts, l'équivalent latéral d'une partie du
black shale Bahloul de Tunisie centrale repose au Jebel Asker sur des calcaires
à ammonites du Cénomanien supérieur. Au Jebel Zitouna et
Jebel Naimia, le faciès Bahloul passe à des calcaires noduleux
moins profonds. Ces auteurs ont conclu que la surface d'émersion connue
plus au Sud se raccorde à la base du Bahloul qui peut être
considéré, à la manière de la fosse vocontienne
occidentale, comme un faciès confiné de bas niveau marin,
déposé dans la partie la plus profonde des bassins
résiduels en période d'émersion de la marge gondwanienne
et que
30
la barre Gattar transgressive correspond à la remise en
eau de cette marge méridionale gondwanienne.
Ouertani (2004) a entrepris des études
minéralogique et structurale dans la région de Tajerouine. Dans
ce travail, les séries crétacées et
paléogènes ont été précisées.
Particulièrement, la Formation Bahloul a été
attribuée au passage Cénomanien - Turonien.
III - Caractéristiques et extension des
dépôts de la limite C/T dans le domaine
téthysien

facies
"Black shale"
Terre émergée
Milieu de Platforme
Environnement marin profond

369
Atlantique
11%
Essaouira
18%
Tarfaya Maroc
397
Maroc
3,5%
Dakar Senegal
W.S
Maroc
28%
Rif
Maroc
5%
g 11 e N Atlas
Algérie
3%
Tebessa
Algerie
Tunisie 7%
Lybie Lybie Egypt
Est
18.7%
Gafsa
>
<---
NW
v ?
13% Tunisie
9%
West
Kerkennah
a
Tunisie
Sicilie
Mediterranean sea
<---
26 %
Calabianca
Guidaloca
2%
Egypt centrale
<---
Fig. 11 - Extension des dépôts de la limite C/T
dans le domaine téthysien (Lüning et al., 2002,
modifiée
concernant la Tunisie)
a - L'Algérie
Naïli et al., (1995) ont signalé
l'existence des calcaires en plaquettes riches en matière organique dans
la Région de Tebessa et de Morsott (Algérie orientale, environ 45
km de Kalâat Es Senan). Ils attribuent ces calcaires de la Formation
Bahloul (Burollet, 1956) au Cénomanien terminal en s'appuyant sur la
présence de l'Ammonite Pseudaspidoceras pseudonosoides en
association avec des Foraminifères planctoniques parmi lesquels
Heterohelix sp, Whiteinella sp, Whiteinella archaeocretacea,
Dicarinella
31
spp, Praeglobotruncana sp, Praeglobotruncana
stephani. Ils ajoutent que le Turonien inférieur ne débute
qu'à la base du membre Annaba (Burollet, 1956) avec la présence
de Fagesia
sp. et Watinoceras sp.
en association avec Hedbergella sp, Whiteinella archaeocretacea,
Guembelitria cenomana, Dicarinella sp et Heterohelix spp.
Des couches siliceuses riches matière organique
d'âge Cénomanien - Turonien ont été signalées
dans plusieurs localités en Algérie notamment au nord de
l'accident Sud Atlasique (Busson et Cornée, 1996 ; Busson et al.,
1999). Ce faciès a formé une bande continue le long de la
plateforme nord africaine au cours du Cénomanien - Turonien,
s'étendant de la Tunisie à l'Est jusqu'au Pré-Rif marocain
à l'Ouest (Kuhnt et al., 1990).
b - Le Maroc
Dans la région d'Éssaouira (Hauts Atlas),
Içame (1994) et Jalal et al., (2002) ont mentionné
l'existence d'un faciès dolomitique qui rappelle celui du membre Gattar
en Tunisie. Alors que Jati et al., (2001), ont signalé que
certaines séquences étudiées dans la région
d'Agadir (Sud ouest du Maroc) sont émersives, notamment au passage
Cénomanien - Turonien, qui est marqué par la présence de
galets roulés et des figures de karstification. Les auteurs ont
rajouté que l'analyse séquentielle montre que les calcaires
laminés anoxiques de la base du Turonien sont réellement
transgressifs. Le maximum d'inondation est marqué par la Zone
d'Ammonites à Neocardioceras vibrayeanus qui se situe entre la
Zone à Rotalipora cushmani et la Zone à Whiteinella
archaeocretacea (Charrière et al., 1998). L'âge de
la Zone à N. vibrayeanus au Marroc correspond à
l'âge de la surface transgressive du Cénomanien sommital en
Tunisie (post Ce5 TS) (Hardenbol et al., 1998). En effet, les
données géochimiques ont montré que l'anomalie de
?13C est enregistrée dans la dernière séquence
coïncidant avec des oscillations mineures du niveau marin relatif
représentées par des lumachelles. Les associations des
foraminifères planctoniques représentées par
Rotalipora cushmani associée à Whiteinella
archaeocretacea, et autres indiquent un milieu assez favorable dans la
série située sous la première discontinuité, avec
toutefois des périodes de stress marquées par des associations
à petits individus peu différenciés tels que
Guembelitria cenomana et G. cretacea (Ettachfini et al.,
2004). Dans les niveaux laminés sus-jacents, les associations sont
principalement représentées par les mêmes formes à
loges globuleuses alors que les formes carénées ne
réapparaissent que temporairement. Ces derniers auteurs ont
32
interprété ces particularités par une
persistance du régime de confinement local malgré l'augmentation
du niveau marin. Par ailleurs Thurow et Kuhnt (1986) en travaillant dans la
région de l'Arch de Gibraltar et dans le Rif (Maroc septentrionale),
montrent que les dépôts du passage Cénomanien Turonien sont
caractérisés par un épanouissement important des
Radiolaires. Ceci coïncide avec plusieurs événements
importants dans l'évolution des Foraminifères planctoniques
(première apparition de Whiteinella archaeocretacea, extinction
du genre Rotalipora, apparition de nouvelles espèces chez le
genre Praeglobotruncana et l'épanouissement des genres à
double carène tels que Marginotruncana et Dicarinella.

Atlantique Central
Tethys
Terre
Fig. 12 - carte paléogéographique du passage
Cénomanien - Turonien montrant le domaine téthysien.
(Philip et al., 2000)
c - L'Espagne
Lamolda et al., (1997) ont essayé d'appliquer
une approche biostratigraphique intégrée pour la limite
Cénomanien - Turonien de la série affleurant à Ganuza en
Espagne. Et
33
c'est en se basant sur la répartition des
Foraminifères planctoniques, des nannofossiles calcaires et ldes
Ammonites. Ces auteurs ont détaillé trois Biozones de
Foraminifères planctoniques : la Zone à Rotalipora
cushmani, la Zone à Whiteinella archaeocretacea et la Zone
à Helvetoglobotruncana helvetica ; Ils les ont correlées
avec les Zones à Eiffellithus turriseiffelii et à
Quadrum gartneri des nannofossiles calcaires. Bien que la macrofaune
soit rare, ces auteurs ont mis en évidence les Zones d'Ammonites
à Metoicoceras geslinianum et à Mammites
nodosoides. Ces mêmes auteurs ont indiqué que la limite C/T
pourrait être établie entre la première occurrence de
Kamerunoceras
sp. et Mytiloides submytiloides et
la première occurrence de Mytiloides kossmati kossmati. Les
premières occurrences successives de Whiteinella praehelvetica
et Q. gartneri sont indiquées par ces auteurs comme
indicateurs probables de la limite C/T.
d - La France
Ferry et al., (2003) ont affirmé que la limite
Cénomanien - Turonien dans la marge provençale du bassin subalpin
est caractérisée par le faciès de black shale Thomel. Ce
faciès n'existe que dans la partie occidentale de cette marge de mer
profonde. Ces auteurs mentionnent que ce faciès de black shales
disparaît dans le domaine de talus (Fig.13) (ou bassin suspendu) qui
serait situé à l'Est de la faille de Rouaine. Le black shale
Thomel de la partie occidentale du bassin vocontien pourrait être
interprété comme un faciès anoxique de milieu restreint
localisé dans un bassin résiduel (black shale de bas niveau).
Tandis que la transgression du passage Cénomanien - Turonien a
déposé sur toute la vallée du Rhône des grés
gloconieux de couleur rougeâtre. L'inondation maximale caractérise
la Zone d'Ammonites à nodosoides.
e - L'Italie
La crise globale EAO-2 est enregistrée en Italie dans
les dépôts du Livello Bonarelli (voir pp 19-20 de ce
document). Ce niveau a l'aspect d'une bande noire caractérisant le
sommet de la Formation Scaglia Bianca. Cet horizon est dépourvu de
carbonates. Il est formé de chert à la base et au sommet
délimitant ainsi des sables et des silts à radiolaires.
Généralement, le Livello Bonarelli mesure de 0.8
à 1.5 m (Arthure et Premoli Silva, 1982 ; Baudin et al.,
1998).

34
Fig. 13 - Carte de distribution des différents domaines
d'âge Cénomanien - Turonien dans la marge
provençale du
bassin subalpin (Ferry et cl., 2003).
IV - Objectif du travail
Dans le cadre du présent travail, nous contribuons par
l'étude micropaléontologique et biostratigraphique de haute
résolution des Foraminifères de l'intervalle du passage
Cénomanien - Turonien de deux coupes levées en Tunisie afin de
:
) multiplier l'information sur les différentes
espèces en association du passage Cénomanien - Turonien.
) participer à une meilleure caractérisation
biostratigraphique de l'intervalle du passage Cénomanien - Turonien, en
se basant sur un échantillonage très serré et en adoptant
les normes de la biostratigraphie de haute résolution.
) mettre en évidence les événements
biologiques enregistrés dans les dépôts de l'intervalle du
passage Cénomanien - Turonien (C/T) par le suivi du comportement
différentiel des Foraminifères planctoniques et benthiques et
préciser leurs changements s'il y a eu lieu le long de ce passage en
s'appuyant sur l'analyse statistique des composants de cette faune.
) participer à une meilleure reconstitution des
conditions paléoenvironnementales au niveau des deux coupes
étudiées et vérifier s'il y a bien une relation entre
ces
35
conditions locales avec celles reignant à l'échelle
globale durant l'intervalle du passage C/T.
V - Matériel et méthode
Une première campagne de terrain nous a permis
d'arrêter notre choix sur deux coupes dans la série de la
Formation Bahloul. La première coupe levée dans la vallée
d'Oued Es-Smara (CES) dans la région de Jerissa au Centre - Ouest de la
Tunisie. La deuxième coupe est celle d'Oued El-Khrroub (COK) dans la
région de Bargou prés de la ville de Siliana à peu
prés à la même latitude que la première mais
beaucoup plus à l'Est. Notre étude est d'ordre
micropaléontologique et paléoécologique. Elle est
basée sur la détermination des Foraminifères
prélevés dans les échantillons de ces deux coupes, leur
état de conservation et leur diversité. Elle est également
d'ordre biostratigraphique de haute résolution basée sur la
répartition verticale détaillée des espèces
marqueurs de Foraminifères planctoniques. Ces derniers vont nous servir
pour la subdivision et la biozonation de l'intervalle du passage
Cénomanien - Turonien s'intégrant dans la Formation Bahloul et
pour discuter les autres biozonations proposées par nos
prédécesseurs, en définissant les limites,
inférieure et supérieur, de ladite Formation.
Le travail sur le terrain consiste en un
échantillonnage plus ou moins serré tout en adoptant des
méthodes de biostratigraphie qualifiée de haute
résolution, surtout dans la zone où se situe la limite des deux
étages géologiques. Ainsi dans les deux coupes
étudiées, les échantillons sont prélevés
à intervalle de 20 cm et 40 cm (voire même 10 cm) pour la
première coupe (CES) et de 30 cm à 50 cm (voire même 20 cm)
pour la deuxième coupe (COK), surtout dans les niveaux sombres de la
Formation Bahloul. De part et d'autre de cette Formation l'intervalle
d'échantillonnage est moins serré. L'espacement entre les
échantillons peut être de 80 cm ou 1 m si le faciès est
inchangé. Les échantillons sont stockés dans des sacs en
plastique portant des étiquettes où sont écrit, la cote de
l'échantillon, son numéro et sa provenance.
La deuxième étape du travail consiste en la
préparation de ces échantillons au laboratoire. Chaque
échantillon est subdivisé en deux parties égales, une qui
servira comme témoin (et pouvant faire objet d'autres types d'analyse),
et l'autre est trompée dans de l'eau de robinet
36
pendant un certain nombre de jours (selon la dureté de
l'échantillon, sans toutefois dépasser une semaine).
Le lavage de ces échantillons consiste en un tamisage
sous un écoulement d'eau afin d'en extraire des éléments
de taille donnée à travers une colonne de quelques tamis
métalliques calibrés. Dans le cadre de ce présent travail,
quatre tamis de type AFNOR (63 um, 100 um, 250 um, 500 um) ont servi pour le
tamisage des échantillons des deux coupes étudiées. Ces
tamis sont placés suivant une colonne à maille
décroissante, c'est-à-dire celui ayant les mailles les plus fines
étant en bas. Cette opération est utilisée à la
fois pour extraire des fossiles ou microfossiles et pour étudier la
répartition granulométrique des éléments
sédimentaires.
En versant le contenu de l'échantillon trompé
sur le tamis supérieur, l'eau du robinet va évacuer les
particules sablo-argileuses inférieures à 500um vers les tamis de
mailles plus fines tout en barbotant le sédiment. Ainsi, quand le
résidu retenu dans chacun des tamis devient propre et réfutant
une eau limpide, il sera récupéré. Les résidus
seront recueillis dans des boîtes à pétri, puis
séchés à l'étuve à moins de 50° C
pendant 4 à 5 jours. Ces résidus séchés sont
ensuite gardés dans des pellules codés et hermétiquement
fermées. L'examen à la loupe binoculaire des résidus est
suivi de tri des microfossiles et leur récupération dans des
cellules à usage micropaléontologique simples ou multiples.
Notons que le passage d'un échantillon à un
autre est toujours précédé par un lavage des tamis avec
une solution du Bleu de Bromothymol, afin de colorer tous les grains proveneant
des échantillons antérieurement tamisés, ensuite les tamis
sont dûment rincés à l'eau de robinet. Ceci évitera
toute contamination lors de la détermination de la microfaune.
Les échantillons calcaires indurés ont fait
l'objet de confection de lames minces afin d'analyser le microfaciès et
le contenu micropaléontologique en section.
L'étude systématique au cours de ce travail n'a
concerné que les Foraminifères planctoniques en particulier. Les
Foraminifères que contiennent les fractions des échantillons
observés sous la loupe binoculaire sont identifiés
spécifiquement en se basant sur les travaux de E.W.G.P.F en
coopération avec F. Robaszynski et M. Caron, sur les
Foraminifères planctoniques du Crétacé Supérieur
(1979. 1 et 2), les traités de micropaléontologie de Loeblich et
Tappan (1961), (1964) et (1988) et d'autres travaux illustrant et discutant les
espèces de Foraminifères planctoniques de l'intervalle de passage
Cénomanien - Turonien (Stelck et Wall, 1954 ; Pessagno, 1967 ; Eicher et
Worstell, 1970 ; Chitta, 1979 ; Salaj,
37
1980 ; Bellier, 1983 ; Robaszynski et al., 1990 ;
Premoli Silva et Sliter, 1994 ; Rami, 1998 ; Georgescu, 2000 ; Keller et
al., 2001 et d'autres...). Pour l'analyse statistique, on a eu
recours à la subdivision des trois fractions de chaque
échantillon (les fractions : 63 um, 100 um et 250 um) à l'aide
d'un microsplitter de type « Otto ». Chaque fraction est
subdivisée en deux parties égales : la première est remise
dans la boite initiale et l'autre est mélangée avec les autres
moitiés de fractions inférieures du même
échantillon. Ce fractionnement va nous aider à obtenir une
association de Foraminifères (aussi bien planctoniques que benthiques)
comportant au minimum 300 individus dans un résidu global >63 um par
échantillon. Ces 300 individus serviront pour une analyse statistique
adéquate. Le matériel étudié peut être riche
ou pauvre en Foraminifères. Pour obtenir seulement 300 individus de
Foraminifères planctoniques. Dans le cas où le résidu
global >63mm est très riche en foraminifère, nous avons
été amenés à le fractionner aussi à l'aide
du microsplitter afin d'obtenir une association de foraminifère
planctonique ne dépassant pas 400 individus. Quand l'échantillon
est pauvre en Foraminifères, nous les avons triés dans l'ensemble
du résidu global.
Concernant les analyses calcimétriques on a eu recours
à un dosage des carbonates à l'aide du calcimètre Bernard
en utilisant une balance de précision, HCl (10%) placé dans une
pissette, une pissette d'eau distillée, un calcimètre Bernard, un
étalon de calcite et une spatule. Les échantillons
prélevés sur le terrain vont servir à la
détermination des teneurs en CaCO3. Cette analyse
calcimétrique consiste tout d'abord à l'étalonnage de
l'appareil en introduisant dans une erlen de 250ml 0,30g de CaCO3 pur
préalablement séché à 50°C. Ensuite on place
dans l'erlen un tube a essai rempli de HCl (10%) en le maintenant dans la
position verticale. Puis l'erlen est fermée afin de prendre une
première lecture L1 sur la burette en ramenant la surface du
liquide que l'erlen contient au même niveau que celui de la burette.
Ensuite le contenu de l'erlen est agité en gardant les surfaces du
liquide dans l'ampoule et dans la burette au même niveau. Enfin on fait
une deuxième lecture L2 sur la beurette. Après cet
étalonage, la determination du taux de CaCO3 dans chaque
échantillon est effectué comme suit : la méthode consiste
à peser 300mg de poudre de l'échantillon préalablement
broyé. Ensuite la même méthode d'attaque que celle
adoptée pour l'étalon est effectuée sur
l'échantillon. L'expression des résultats est :
38
DEUXIÈME PARTIE :
DESCRIPTION DES FACIÈS ET ENVIRONNIMENTS
DE DÉPÔTS
39
I - Introduction
Les deux coupes que nous étudions, intéressant
l'intervalle du passage Cénomanien - Turonien, ont été
levées dans la Formation Bahloul. L'une concerne l'affleurement de la
région de Jerissa au niveau de Oued Es Smara (CES) (Fig.14) en Tunisie
Nord occidentale et l'autre coupe (COK) est levée plus au nord est
à Oued El Kharroub.
|
Legend
|
|
Road Car track
|
|
Railroad Hydrographic network
|

Tajerouine
Towards
Kalâat
Senan
0 Scale 2 km
O. El Morr
644
El Hameima
Aïn el
Zoumiat
879
Aïn bit Hafi
790
649
Jerissa
822
Fig. 14 - carte de localisation de la région de
Tajerouine
Sur le plan structural, le massif de Jerissa s'intègre
dans l'atlas central. Cette unité structurale est
caractérisée par des plis orientés NE-SW
entrecoupée par de nombreux fossés d'effondrement (Fig.15) parmi
lesquels celui de Kalâat Khesba situé en bordure occidentale de
l'anticlinale de Jerissa à coeur de l'Aptien.
Le massif de Bargou fait partie de la dorsale tunisienne et
forme aussi un anticlinarium à coeur de l'Aptien orienté NE-SW.
Dans cette région, la série du passage C/T est bien
développée au tour du massif

Jerissa
Légende
Garn Halfaya
El Kef Seliana
CES
Nebeur
Diapire
Crétacé supérieur
J. Kebbouch
Bougrine
Zone des fossés
Aïn
Zakkar
Anticlinal
Synclinal
Graben Faille
0 Scale 10 km
Bargou
N
40
Fig. 15 - cadre structural général des deux
coupes (d'après Layeb, 1990 simplifiée)
A - ÉTUDE DE LA COUPE D'OUED ES SMARA : CES
1 - Historique des travaux antérieurs
Cette région de Tajerouine, bien qu'elle ait faite
l'objet de nombreux travaux depuis le début du 20ème
siècle, elle demeure encore un secteur insuffisamment
étudié sur certains plans. Parmi ces travaux, ceux de
Pervinquière (1903), Burollet (1956), Ben Ayed (1986), Negra (1994),
Kassaa et al., (1995) ; Zaier (1999), Chikhaoui (2002) et Ouertani
(2004) ont concerné l'aspet structural de la région et/ou
sédimentologique et lithostratigraphique des différentes
séries sédimentaires afin de reconstituer les environnements de
dépôt ou
42
encore de caractériser les substances utiles, mais
aucune étude biostratigraphique détaillée basée sur
les Foraminifères planctoniques et surtout en ce qui concerne
l'intervalle du passage Cénomanien - Turonien n'a été
élaborée jusqu'ici. Les travaux de Robaszynski et al.,
(1990 ; 1993) sur les Foraminifères planctoniques de la
région de Kalâat Es Senan (Oued Smara, Oued Zitoun) située
plus Sud Ouest, sont de faible résolution. Le travail de Nederbragt et
Fiorentino (1999) qui est de haute résolution a intéressé
la région d'El Kef située plus au Nord. Ce dernier travail a
concerné uniquement les Heterohelicidae.
La Formation Bahloul dans la région de Jerissa et les
régions limitrophes est d'épaisseur variable (Tableau - 1)
|
La région de Tajerouine - Jerissa
|
|
Localité
|
épaisseur (m)
|
|
Bougrine
|
26
|
|
Garn Halfeya
|
17
|
|
Jerissa
|
24
|
|
Oued Hammam
|
18
|
|
Oued Amor
|
18
|
|
Oued Semara (KS)
|
23
|
Tableau 1 - Variations d'épaisseur de la Formation Bahloul
dans la région de Tajerouine
(KS) : Kalâat Es Senan (d'après les
données de Layeb, 1990 et Ouertani, 2004)
2 - Cadre géographique et géologique a
- Cadre géographique
La coupe que nous avons levée dans l'intervalle de
passage C/T passe par Oued Es Smara. Ce cours d'eau est localisé
géographiquement au Nord-Ouest de la Tunisie (Fig.14) assez près
de la frontière tuniso-algérienne, à
l'extrémité Sud Est de la feuille
43
topographique et géologique de Tajerouine (n° 51)
au 1/50.000. Il est situé à l'Ouest de Oued Amor, Aïn Rahala
et les collines de Fej Tameur ; à l'Est du massif de Djebel Oum Khelil
et Oued El Abiod.
Il est limité au Nord par Aïn Halfi et les
prolongements Sud Ouest des reliefs de Kef Es Slougui et au Sud par la ville de
Jerissa. L'accès à cette coupe se fait par la Route Nationale
GP. 18 menant à la ville de Jerissa et son complexe minier. Plus au
nord se dresse le flanc légèrement redressé de Djebel El
Hout. L'altitude moyenne des reliefs entourant le secteur est de 800 m. La
région est dominée par un réseau hydrographique
représenté par de grands ou petits lits d'oueds (Oued Amor, Oued
El Abiod, Oued Hammam) qui sont tous des affluents d'Oued Er Retem qui est
situé plus au Sud.
b - Cadre géologique
La région de Tajerouine est considérée
comme étant une zone de transition entre l'Atlas septentrional et
l'Atlas central. Les travaux cartographiques de la région ont
été élaborés par Burollet et al., (1953 ;
1954) et Burollet et Sainfeld, (1956). Elle est dominée par des
fossés d'effondrement (fossés de Kalâat Khesba,
fossé de Rouhia) et des structures diapiriques à coeur de Trias.
A l'Est, le secteur est limité par le rebord oriental du fossé de
Kalâat Khesba (Zaier, 1999 ; Ouertani, 2004) et au Sud Est par le
prolongement du fossé de Rouhia. Le secteur est dominé aussi par
un réseau de failles et de décrochements de directions multiples
(Fig. 17) : N-S, NW-SE, E-W et NE-SW. Le secteur est limité dans sa
partie sud par le flanc nord peu redressé du monoclinal de Jebel
Jerissa.
Au niveau de la coupe CES, les niveaux de marnes les bancs de
calcaires argileux de la Formation Bahloul sont tous de direction N140. Ils
pendent de 35° vers le Nord Est (N140-35°NE). Plus loin les couches
varient de direction. Au voisinage de la coupe, ces couches sont
affectées par un décrochement senestre de direction Nord Sud et
elles comportent des figures de glissement synsédimentaires (Ouertani,
2004).

44
Lower Santonian Lower Campanian Middle Campanian Lower
Maastrichtian
Middle Maastrichtian Maastrichtian - Paleocene
Lutitian
Quaternary
Lower Aptian Upper Aptian Lower Albian Middle Albian
Upper Albian Cenomanian
Cenomanian-Turonian Lower Coniacian
N
<
O.
O
.
a
ra
m
45 T
S
---
-
s
e
.
O
S
E
50T
C
55T
Jerissa
--
-
<
<---- Towards Tajerouine
Towards Kalâat Senan
l
O.
el
Abiod
Aïn bit Halfi
Jebel Jerissa
Am
o r
Aïn Rahala
Ham
m
F aej Tameur
m Hills
Towards Dahmani
Piste Route
Faille
Réseau hydrographique
0 0 Échelle 20 km
CES Coupe de Oued Smara (CES)
Fig. 17 : Extrait de la carte géologique N° 51de
Tejerouine 1/50
00
45
3 - Coupe d'Oued Es Semara (CES)
La coupe CES est levée au niveau du point aux
coordonnées :
35°E50'E56,3036'' de la feuille topographique de Tejerouine
n°51 au
1/50.000 ; Prés de la piste qui représente le
prolongement vers le Nord Est de la route Nationale GP. 18, au niveau du point
de rencontre des deux affluents de Oued Es Semara.
Cette coupe a concerné seulement la Formation Bahloul.
L'attribution chronostratigraphique de ladite formation a été
discutée par plusieurs auteurs ayant travaillé dans les
régions voisines. Elle a été attribuée soit au
Cénomanien supérieur
(Gargouri, 1983 ; Robaszynski et al., 1990) soit au
Turonien inférieur (Burollet,
1956 ; Bismuth et al., 1981 ;
Benzarti-Saïd, 1981 ; Robazsynski et al., 1993) soit encore au
passage Cénomanien - Turonien (Dali - Rossot, 1987 ; Ben Haj Ali et
al., 1994 ; Maâmouri et al., 1994 ; Robaszynski et al.,
1995 ; Rami, 1998, Accarie et al., 1999 ; Nederbragt et
Fiorentino, 1999 ; Caron et al., 1999). La série que nous
détaillons est épaisse de 23,2m, elle couvre la totalité
de la Formation Bahloul et déborde vers le bas sur la partie terminale
de la Formation Fahdène (Burollet, 1956) et vers le haut sur la partie
basale de la Formation Kef (Fournié, 1978). Dans cette série,
nous avons distingué, au point de vue lithostratigraphie, six
unités différentes :
a - L'Unité basale U1 [CES-1 - CES-2] :
Elle s'étend sur 3,5m. Elle est formée
essentiellement d'argiles à intercalations de calcaires argileux.
L'ensemble est de couleur grisâtre devenant de plus en plus sombre. A la
cassure fraîche, cet ensemble se débite le plus souvent en
nodules. Toutefois dans sa partie médiane, il englobe un niveau calcaire
argileux se débitant plutôt en plaquettes de couleur noir en
patine comme en cassure fraîche.
Ce terme a livré une association de
Foraminifères planctoniques composée de Rotalipora
montsalvensis (MORNOD), Rotalipora cushmani (MORROW),
Rotalipora greenhornensis (MORROW), Praeglobotruncana stephani
(GANDOLFI), Praeglobotruncana delrioensis (PLUMMER),
Praeglobotruncana gibba KLAUS, Dicarinella algeriana
(CARON),
|
Stage
|
Formation
|
Zones
|
Thickness
|
Sample
|
Lithology
|
Unit
|
Lithology description
|
|
Mid. Cenomanian Upper Cenomanian Lower Turonian
|
Kef
|
Rotalipora cushmani Whiteinella archaeocretacea H.
helvetica
|
22 20 18 16 14 12 10 8 6 4 2
|
|
|
|
Nodulous and fracturated limestone Dark marls
Grey to greenish marls
Calcarous marls layered with sheets
Black bioturbated and organic-rich limestone (Bioturbation
level)
Black laminated limestone with Ammonites mould (Ammonites
level)
Black clayey limestone interbedded with grey limestone
Marls level layered in sheets Grey limestone layered in sheets
Marls and bituminous grey to black clayey limestone
alternation
Nodulous grey marls (1.5m) interbedded with grey to dark-grey
clayey limestone beds (30cm) layered
in sheets
|
|
65
|
|
|
|
Bahloul
|
|
|
U6
|
|
|
|
|
60
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
55
|
|
|
|
|
|
|
50
|
|
|
|
|
|
|
|
|
|
|
|
|
|
U5
|
|
45
|
|
|
|
|
|
|
40
|
|
|
|
|
|
|
35
|
|
|
|
|
|
|
|
|
|
30
|
|
|
|
|
U4
|
|
25
|
|
|
|
|
|
|
20
|
|
|
|
|
|
|
|
|
|
|
|
|
15
|
|
|
|
10
|
|
U3
|
|
|
|
|
|
|
|
5
4
3
2
1
|
|
U2
|
|
Fahden
|
|
U1
|
|
|
1m
|
46
Fig. 18 - description lithologique de la coupe CES
Globigerinelloides ultramicra (SUBBOTINA),
Globigerinelloides bentonensis
(MORROW), Clavihedbergella simplex (MORROW),
Hedbergella planispira (TAPPAN), Whiteinella aprica (LOEBLICH
& TAPPAN), Whiteinella brittonensis (LOEBLICH & TAPPAN),,
Whiteinella baltica DOUGLAS & RANKIN, Heterohelix moremani
(CUSHMAN), Heterohelix navarroensis LOEBLICH, Heterohelix
aff. Pulchra (BROTZEN), Heterohelix sp., Guembelitria cenomana
(KELLER) et des Foraminifères benthiques représentées
essentiellement par (Cassidella, Frondicularia, Gaudryina, Gavelinella,
Buliminella, Textularia)
La présence de Rotalipora montsalvensis en
association avec Rotalipora cushmani et Rotalipora greenhornensis
témoigne d'un âge cénomanien moyen.
b - Unité U2 [CES-3 - CES-6]
Les 3,5m suivant sont représentés par des marnes
schisteuses alternant avec des calcaires argileux bitumineux. Ce terme a
livré Rotalipora cushmani (MORROW) Rotalipora
greenhornensis (MORROW), Praeglobotruncana
stephani (GANDOLFI),
Praeglobotruncana delrioensis (PLUMMER),
Praeglobotruncana gibba (KLAUS), Dicarinella algeriana
(CARON), Globigerinelloides ultramicra (SUBBOTINA),
Globigerinelloides bentonensis (MORROW), Hedbergella simplex
(MORROW) , Hedbergella planispira (TAPPAN), Hedbergella
delrioensis (CARSEY), Whiteinella aprica (LOEBLICH & TAPPAN),
Whiteinella brittonensis (LOEBLICH & TAPPAN), Whiteinella
baltica DOUGLAS & RANKIN, Whiteinella paradubia (SIGAL),
Whiteinella archaeocretacea PESSAGNO, Heterohelix moremani
(CUSHMAN), Heterohelix navarroensis LOEBLICH, Heterohelix
aff. Pulchra (BROTZEN), Heterohelix reussi (CUSHMAN),
Guembelitria cenomana (KELLER), Guembelitria albertensis
STELCK & WALL et des Foraminifères benthiques
représentés par (Cassidella, Frondicularia, Gavelinella,
Buliminella, Textularia) qui commencent à être rare. On note
que Whiteinella archaeocretacea n'apparaît qu'à partir de
l'échantillon CES-4 (7,75m), ce qui indique qu'elle coexiste bien avec
les derniers Rotalipores.
La présence de Rotalipora cushmani et
Rotalipora greenhornensis en association avec Whiteinella
archaeocretacea et sans Rotalipora montsalvensis indique un
âge Cénomanien supérieur.
47
c - Unité U3 [CES-7 - CES-18]
|
Stage
|
Formation
|
Zones
|
Subzones
|
Thickness
|
Sample
|
Lithology
|
Unit
|
R. montsalvensis P. delrioensis
R. cushmani R. greenhornensis P. stephani W.
paradubia
Gl. bentonensis D. hagni-imbricata A. multiloculata P.
gibba
D. algeriana
Gl. ultramicra
G. cenomana Hd. delrioensis
Hd. planispira Cl. simplex
H. reussi
H. moremani H. navarroensis H. pulchra
W. aprica W. baltica
W. inornata
W. aumalensis
W. brittonensis
W. archaeocretacea
D. imbricata
Guembelitria sp.
W. praehelvetica
D. hagni
Hl. helvetica
M. renzi
M. sigali
M. sp
|
Species richness
|
|
Mid. Cenomanian Upper Cenomanian Lower
Turonian
|
Kef
|
H. helvetica
|
22 20 18 16 14 12 10 8 6 4 2
|
|
|
|
|
|
|
65
|
|
U6
|
|
|
|
Fahden Bahloul
|
|
|
|
H. helvetica FA
|
|
|
|
|
|
|
|
Whiteinella archaeocretacea
|
G. bentonensis Dicarinella hagni Heterohelix moremani
|
|
60
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
55
|
|
|
|
|
|
|
|
|
|
|
50
|
|
|
|
|
|
|
|
|
r-strategy
|
|
|
|
Heterohelix
|
|
|
|
|
|
|
|
|
U5
|
|
|
|
45
|
|
|
shift
|
|
|
|
|
|
|
|
40
|
|
|
|
|
|
|
|
|
|
|
35
|
|
|
|
|
|
|
|
Very low
|
|
|
|
30
|
diversity
|
|
|
|
|
|
Benthic
|
|
|
U4
|
|
Foraminifera
|
|
25
|
|
|
|
absence
|
|
|
|
|
|
|
20
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
15
|
|
|
|
|
|
10
|
|
U3
|
|
|
|
|
|
R. cushmani
|
LA
|
|
|
|
|
|
|
5
4
3
2
1
|
|
U2
|
|
K-strategy
|
|
Rotalipora cushmani
|
R. montsalvensis D. algeriana
|
|
U1
|
|
|
|
|
1m
|
|
0 15 30
|
48
Fig. 19 - répartition des Foraminifères
planctonisues dans la coupe de Oued Es Smara (région de Jerissa)
49
Cette unité s'étend sur 2,30m, représente
un petit banc calcaire gris clair en patine comme en cassure fraîche se
débitant en feuillets et surmonté par un niveau marneux finement
lité qui se débite en plaquettes alternant avec un banc calcaire
en plaquettes plus foncé. Ce terme a livré la même
association précédante en plus de Dicarinella hagni
(SCHEIBNEROVA) et Dicarinella imricata (MORNOD).
Cette unité, comme la précédante, est
caractérisée par l'abondance des Calcisphères
(Pithonella sp, Bonetocardiella sp), des Radiolaire, des filaments,
débris d'Echinodermes, des Ostracodes et des cristaux de Pyrite
associés parfois à des cristaux de calcite.
Notons que au-delà de l'échantillon CES-6
(8,20m) les dernières Rotalipores (Rotalipora greenhornensis)
disparaissent et qu'à partir de CES-9 (9,35m) Whiteinella
praehelvetica apparaît.
En résumé, dans ces trois unités à
dominante marneuse et à intercalation sporadique de petits bancs
calcaires finement lités de couleur gris en patine qui se
débitent en petits feuillets, on assiste d'une part à
l'extinction des Rotalipores et d'autre part à l'apparition de
Whiteinella archaeocretacea ainsi que de Whiteinella
praehelvetica. En ce qui concerne les
Hétérohélicidés et les Guembelitriidés qui
sont de petite taille (<100um) et très abondants tels H. moremani
(CUSHMAN), H. reussi (CUSHMAN), G. cenomana (KELLER) et
G. albertensis STELCK & WALL, on remarque deux pics très
nets succédant la disparition des Rotalipores (Fig.25). Le premier est
celui de H. moremani accompagné par un pique de
Guembelitria cenomana. Le deuxième est uniquement de H.
moremani, H. reussi.
Dans ces niveaux où la microfaune est trop petite
(<250 um) on note entre autre une dissolution dans les échantillons
CES-17 et CES-18.
d - Unité U4 [CES-19 - CES-37]
Cet ensemble est constitué essentiellement d'une masse
calcaire laminé bien développée. Ce calcaire est beige en
patine et noir à la cassure fraîche se débitant en
feuillets. Il s'éclaircit vers CES-24 (13,70m) où l'association
des Foraminifères planctoniques est représentée par
Heterohelix moremani (CUSHMAN), H. reussi (CUSHMAN)
H.
51
navarroensis LOEBLICH, H. aff. Pulchra
(BROTZEN), Guembelitria cenomana (KELLER), G. albertensis
STELCK & WALL, Whiteinella praehelvetica (TRUJILLO), W.
aprica (LOEBLICH & TAPPAN), Clavihedbergella simplex
(MORROW), Hedbergella delrioensis (CARSEY), puis il redevient
encore noir foncé vers CES-30 (16,10m). Notons que l'échantillon
CES-26 (14,30m) comprend une microfaune très dissoute et pauvre en terme
de diversité (quelques Hétérohélicidés,
Guembelitria cenomana, Hedbergella delrioensis et presque
dépourvu de Foraminifères benthiques). On note aussi la
disparition de Globigerinelloides bentonensis au-delà de CES-23
(13,4m) et l'absence des Foraminifères benthiques au-delà de
CES-27 (14,9m). Les éléments associés sont
représentés par de rares Ostracodes, une abondance de grains de
quartz (Fig.21) et présence sporadique de Calcisphères et de
Radiolaires.
e - Unité U5 [CES-38 - CES-61]
Cette unité mesure 5,3m. Elle est composée par
des calcaires à aspect massif mais qui se débitent en plaquettes.
Ils sont gris à gris clair en patine et noirâtre à la
cassure. La partie basale de cette unité contient des Ammonites (Niveau
à Ammonites). Ce dernier niveau est surmonté par un autre niveau
calcaire noir peut être riche en matière organique et très
bioturbé (niveau à bioturbation). Il est de couleur gris
foncé en patine et noir à noir foncé en cassure. Ces deux
niveaux sont séparés par une passée marneuse.
L'association des Foraminifères planctoniques livrée reste
presque inchangée dominée par les
Hétérohélicidés, Guembelitriidés,
Hedbergelles et quelques Whiteinelles avec la présence encore de
Dicarinella algeriana (CARON), Dicarinella hagni
(SCHEIBNEROVA), parfois Globigerinelloides ultramicra (SUBBOTINA)
et Praeglobotruncana gibba KLAUS.
Au niveau de cette unité on note que
Praeglobotruncana gibba disparaît au-delà de CES-50
(22,25m). La diversité spécifique des Whiteinelles devient de
plus en plus pauvre dominée par Whiteinella praehelvetica avec
présence sporadique de W. archaeocretacea et W.
aprica. L'association est représentée par de rares
Dicarinella algeriana, Dicarinella imbricata (rares
individus), parfois Globigerinelloides ultramicra, quelques
Clavihedbergella simplex, Hedbergella delrioensis, Hedbergella
planispira. Cette association est dominée par les
Hétérohélicidés et les Guembelitriidés
(surtout
52
Stage
|
Formation
|
Zones
|
Subzones
|
Thickness
|
Sample
|
Lithology
|
Unit
|
Pyrite
|
Calcite
|
Benthic
Foraminifera
|
Ammonites
|
Glauconites
|
Ostracods
|
Radiolarians
|
Filaments
|
Calcispheres
|
Quartz
|
CaCO
3
(%)
|
|
Mid. Cenomanian Upper Cenomanian Lower Turonian
|
Kef
|
H. helvetica
|
22 20 18 16 14 12 10 8 6 4
2
|
|
|
+
+
+
+
+
+
+ + + + + +
+
+
+
+
+
+ + + + + + + + +
+
+
+
|
+
+
|
+
+ + + + + + ++
+ + + +
+
+
+
+
++
+
+ + + +
+
++
+
+
+
+
+
+++
+
+
+
+
+
+ + + + + +
+ + + + + + + + + +
+
+
+
+
|
+
+
|
+
|
+
+
+
+
+
|
+
+
+
+
+
+
+
+ + + + + + + +
+
+
+
+
+
|
+
+ + + + +
+
+
+
+
+
+
+
+
+
+
+
+
+
+
|
+
+
+
+
+
+
+
+ + + +
+ + + + + + + +
+
+
+
+
+
|
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
|
0 40 80
|
|
65
|
|
|
|
Bahloul
|
|
|
U6
|
|
60
|
|
|
|
Whiteinella archaeocretacea
|
G. bentonensis Dicarinella hagni Heterohelix
moremani
|
|
|
|
|
|
|
|
|
|
|
55
|
|
|
|
|
|
|
|
|
|
50
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
45
|
|
U5
|
|
|
|
|
|
|
|
40
|
|
|
|
|
|
|
35
|
|
|
|
|
|
|
|
|
|
30
|
|
|
|
|
U4
|
|
25
|
|
|
|
|
|
|
20
|
|
|
|
|
|
|
|
|
|
|
|
|
15
|
|
|
|
10
|
|
U3
|
|
|
|
|
|
|
|
Rotalipora cushmani
|
Rot. montsalvensis D. algeriana
|
|
5
4
3
2
1
|
|
U2
|
|
Fahden
|
|
U1
|
|
|
1m
|
|
Fig. 21 - Répartition des fossiles,
éléments minéraux et courbe de variation du taux de CaCO3
dans la
coupe CES
Guembelitria cenomana). Dans ce niveau on note le
retour des Foraminifères benthiques (fort pourcentage) surtout
Buliminella et Gavelinella
f - Unité U6 [CES-47 - CES-67]
Sur cette dernière barre calcaire se développe
une série peu épaisse à dominante marneuse de 2,25m qui
disparaît sous la piste. Ces marnes sont au début claires et
lithées puis deviennent noirâtres.
Les éléments présents sont
représentés par des filaments, une richesse en grains de calcite
et de quartz, débris d'Echinodermes, présence très rare de
Calcisphères (Pithonella et Bonitocardiella) et des
radiolaires, de rares Ostracodes, parfois des cristaux de pyrites
Dès la base de cette unité CES-62 (26,30m) on
assiste à l'apparition de Helvetoglobotruncana helvetica
(BOLLI) et Marginotruncana renzi. Ces marnes ont livré une
association de Foraminifères planctoniques composée
essentiellement de Heterohelix moremani, H. reussi, H. aff. Pulchra, H.
navarroensis, Guembelitria cenomana, G. albertensis (signalons que les
niveaux CES-62 et CES-63 respectivement 26,20m et 26,80m sont dépourvus
de Guembelitria), abondance de Whiteinella praehelvetica,
Helvetoglobotruncana helvetica, présence sporadique de W.
aprica et W. archaeocretacea, de rares Hedbergella planispira, H.
delrioensis, H. simplex, Marginotruncana renzi, Marginotruncana sigali,
Marginotruncana sp., Dicarinella algeriana, parfois de petits
Globigerinelloides ultramicra
La microfaune est représentée par des individus
de très petite taille même <100um. La présence de
Helvetoglobotruncana helvetica en association avec les premier
Marginotruncana témoigne d'un âge Turonien
inférieur à moyen.
B - ÉTUDE DE LA COUPE DE OUED EL KHARROUB : COK
53
1 - Travaux antérieurs
54
La région de Bargou a fait l'objet de plusieurs
travaux. Les plus anciens écrits sont d'Aubert (1892). En
présentant la carte géologique provisoire de la Tunisie au
1/800.000 l'auteur a décrit les séries affleurant dans la
région et les principales structures géologiques qui la
caractérisent. Il a particulièrement évoqué
l'anticlinal des massifs Bargou et de Serj. Ce travail a été
succédé par ceux de Pervinquière (1903), Volpière
(1933), De Spengler (1948), Castany (1951), qui sont d'ordre stratigraphique et
structural, Zebidi (1963), d'ordre hydrogéologique, Bismuth (1964),
Jauzein (1967), Guirand (1968), Turki (1975 ; 1985), Sedjil (1981), El Ouerdi
(1992) qui est d'ordre structural. Layeb, (1990) d'ordre géochimique.
Ceux qui sont d'ordre paléontologique, citons ceux Khayati - Ammar
(1996) concernant les ostracodes de l'Aptien, de Rami (1998) (les
Foraminifères du Crétacé supérieur) et de Srarfi
(1999). Ainsi, peu de travaux ont concerné la Formation Bahloul du
passage Cénomanien - Turonien et son contenu en Foraminifères
planctoniques suivant le mode de haute résolution.
2 - Cadre géographique et géologique a
-Cadre géographique
Le massif de Bargou culminant à 1270m d'altitude qui
s'étend sur presque 20 km de long et 10 km de large et se situe
géographiquement dans l'Atlas tunisien central (Fig. 15). Il occupe la
partie centrale de la feuille n° 47 de Jebel Bargou au 1/50.000. En
général, cette région est de direction majeure NE - SW qui
représente la direction de ce massif considéré comme un
complexe montagneux. Au flanc nord occidental du Jebel Bargou, la coupe de Oued
El Kharroub (COK) a été levée au niveau de Oued El
Kharroub. Elle est limitée à l'Est par le massif de Bargou,
à l'Ouest par Oued Touirellil, Hinchir El Bouria et Bled El Hofra. Au
nord, elle est limitée par Merdja Aouab et Oued Fredj, alors qu'au sud,
elle est limitée par Aïn zakkar et Oued Zakkar (Fig.22).
L'épaisseur de la Formation Bahloul varie tout au tour du
paléo-haut fond du Bargou (Layeb, 1990) (Tableau-2.)
Les principaux reliefs qui représentent ce complexe
sont jebel Bargou et Seba Koudiat. Ces montagnes sont entaillées par de
nombreuses vallées dont la plus importante et la plus proche est celle
de Oued Drija. L'altitude moyenne entourant cette région est de 1000
m.
55
Le climat dominant cette province montagneuse est
généralement continental subhumide à contraste thermique
fort influant sur le couvert végétal caractérisé
par quelques arbustes qui poussent prés des petits ravins.
|
La région de Bargou
|
|
Localité
|
épaisseur (m)
|
|
Aïn Zakkar
|
39
|
|
Dir Ouled Yahia
|
37
|
|
Oued Gsab
|
13
|
|
Oued Drija
|
30
|
|
Jebel Boukéhil
|
23
|
|
Sidi Naoui
|
3
|
Tableau-2. Variation d'épaisseur de la Formation Bahloul
dans la région de Bargou
(D'après les données de Layeb, 1990 et Rami,
1998)
b - Cadre géologique
Ce secteur fait partie de la zone des plis atlasiques. Il se
situe en position à faciès mixte (El Ouerdi, 1996) où une
sédimentation différentielle se manifeste. Cette
caractéristique s'exprime par la présence de sédimentation
marine profonde de sillon et une deuxième de type néritique de
plate-forme. Le massif de Bargou est considéré comme un
anticlinorium de direction générale NE - SW. Il est formé
par un complexe de plis anticlinaux séparés par des synclinaux
étroits (Turki, 1975 ; 1985). Cette vaste structure anticlinale
présente une ossature d'âge Aptien supérieur à
Albien inférieur (Castany, 1951 ; Jauzein, 1967 ; Turki, 1975 ; 1985 ;
El Ouerdi, 1996). L'évolution structurale de la région est
marquée par le scellement des structures d'âge
Crétacé inférieur où un régime de distension
est

Siliana
Merdja Aouab
G.P.4
759
O.
Kharroub
Dir Od Yahia
Bargou
1268
Aïn Zakkar
0 2 4 Km
56
Fig. 22 - Carte de situation
57
décelé. Cette activité persiste pour
former des structures en horsts et grabens (Martinez et al., 1984 ;
Layeb, 1990) qui ont peut être facilité le dépôt des
sédiments riches en matière organique dans les zones les plus
subsidentes à la limite Cénomanien - Turonien.

1 - Crétacé inférieur
2 - Cénomanien - Turonien
3 - Crétacé supérieur
4 - Maastrichtien-Danien
5 - Paléocène -
Éocène
6 - Quaternaire
1207
665
795
966
689
40
Dap Cheikh Ammar
Dir Ouled Yahia
Ain Zakkar
St Kanassi
Dret Haj Hssine
1078
904
S. Med ben Khlifa
Fig.23 - Extrait de la carte géologique N°47 de
Bargou au 1/50000
3 - Étude stratigraphique:
La coupe COK est levée au niveau du point aux
coordonnées :
36 °~ 2 '~ 7,9480 ''~des feuilles topographique et
géologique de Bargou n°47
au 1/50.000 ; prés de la piste
menant à Aïn Bou Saâdia.

Cénomanien supérieur Turonien inférieur
Cénomanien moyen
Fahdène
Kef
Bahloul
Whiteinella archaeocretacea
Foraminifères
Rotalipora cushmani
H. helvetica
Radiolaires
Alievium superbum
Rhopalosyringium hessi
Pessagno 1976
Rhopalosyringium majuroensis Crucella cashensis
Thurow 1988
O'Dogherty
Superbum
Biacuta
/ Jud
1994
Unité 1 Unité 2 Unité 3 Unité 4
Unité 5 Unité 6 Unité 7
58
Fig. 24. Description lithologique de la Formation Bahloul au
niveau de la coupe COK
59
Au niveau de cette coupe qui n'a concerné aussi que la
Formation Bahloul, nous avons distingué, au point de vue
lithostratigraphie, sept unités différentes :
a) L'Unité basale U1 [COK-1 - COK-15]
Cette Unité est essentiellement marneuse de couleur
beige clair à quelques intercalations marno-calcaires à
calcaires. Cette unité de 10m d'épaisseur a livré
l'association de Foraminifères planctoniques suivante : Rotalipora
appenninica (RENZ), R. montsalvensis MORNOD, R.
greenhornensis (MORROW), R. cushmani (MORROW),
Globigerinelloides bentonensis (MORROW), Praeglobotruncana gibba
KLAUS, P. delrioensis (PLUMMER), P. stephani
(GANDOLFI), Whiteinella aprica (LOEBLICH & TAPPAN), W.
aumalensis, W. paradubia (SIGAL), Globigerinelloides ultramicra
(SUBBOTINA), Heterohelix moreman (CUSHMAN)i, H. reussi
(CUSHMAN), H. pulchra (BROTZEN), Hedbergella delrioensis
(CARSEY), H. planispira (TAPPAN), Clavihedbergella. Simplex
(MORROW), Guembelitria cenomana (KELLER). On note l'apparition de
Dicarinella algeriana dés le niveau COK-8 et la disparition de
Rotalipora appenninica à partir du niveau COK-10. La
présence des Foraminifères benthiques est aussi importante, elles
sont représentées par l'association composée de :
Gaudryina, Tappanina, Gavellinella, Lunatriella, Textularia, Trochamina,
Saccammina, Cassidella, Frondicularia, Ammobaculoides. Cette
dernière association est dominée par les Textularia.
La présence de la faune à R. appenninica et R.
montsalvensis indique un âge Cénomanien moyen à
supérieur
b) L'Unité U2 [COK-16 - COK-30]
Cette unité est caractérisée par des
alternances de niveaux calcaires de couleur beige clair, se débitant en
petits parallélipipèdes et des marnes grisâtres se
débitant en plaquettes. Elle comporte trois niveaux à Ammonites.
Le toit de cette unité est coiffé par une base ravinante. Cette
partie a livré presque la même association de Foraminifères
planctoniques avec la présence sporadique de Whiteinella
brittonnensis. On note aussi la présence de Heterohelix
navarroensis. Cette unité est caractérisée par
l'abondance de Rotalipora greenhornensis, Dicarinella algeriana,
Whiteinella aprica et des Foraminifères

Cénomanien supérieur Turonien inférieur
Cénomanien moyen
Age
Fahdène
Kef
Bahloul
Formation
Foraminifères
Whiteinella archaeocretacea H. helvetica
Rotalipora cushmani
Radiolaires
Alievium superbum
Rhopalosyringium hessi
Pessagno 1976
Rhopalosyringium majuroensis Crucella cashensis
Thurow 1988
O'Dogherty / Jud 1994
Superbum
Biacuta
Épaisseur
Échantillon
Lithologie
C. puga
E. communis
X. aff. picenus
Guttacapsa sp
Extinction Résistance Évolution
S. elatica
Novixitus sp
S. stocki
M. gracilis
C. messinae
S. fossilis
P. pseudomacrocephala
R. radiosum
U. pacifica
R. hispidum
D. montisserei
P. lipmanae
S. communis
A. aff. vitalis
T.veneta
P. glascockensis
C. euganea A.? mellifera
Phalangites (?) sp
P. spinosa
C. californiaensis
Age
|
Formations
|
Biozones
|
Sous-Zones
|
Épaisseur
|
Échantillons
Lithologie
|
R. appenninica R. montsalvensis
P. delrioensis R. cushmani
R. greenhornensis
Gl. bentonensis
P.gibba
P. stephani W.aprica
W. aumalensis
W. inornata
W. baltica
W. paradubia
Gl. ultramicra
W. brittonensis
Het. moremani
Het. navarroensis
Het. reussi
Het. pulchra
Het. sp
G. cenomana
G. albertensis
H. delrioensis
H. simplex H. planispira
D. algeriana
W. archaeocretacea A. multiloculata
D. hagni
W. praehelvetica
D.imbricata
H. helvetica
M. renzi
M. pseudolineana
|
|
Cénomanien moyen Cénomanien supérieur
Turonien inférieur
|
Fahdène Bahloul Kef
|
H.
helvetica
|
57 54 51 48 45 42 39
36 33 30 27 24 21
18
15 12 9 6 3
|
99
97 96 95 94 93 92 91 90 89 88 87 86 85 84 83
81
80
78 77 76 75 74 73
72
70 69 68 67 66 65 64 63 62 61 60 59 58 57 56
55
54
51 50 49 48 47 46 45 44 43 42
38
37
36
34
33
32 31 30 29 28
27
26
25 24 23 22 21 20
17
16
15
13
12
11
10
9
8
|
100
98
|
|
|
U7
|
|
|
|
Rotalipora cushmani Whiteinella archaeocretacea
|
R. Gl.
Dicarinella algeriana Heterohelix moremani
greenhornensis benton ensis D. hagni
|
|
|
|
U6
|
|
79
|
|
|
|
71
U5
|
|
|
|
|
53
52
|
|
|
U4
|
|
|
|
4401 39
|
|
|
|
35
|
|
U3
|
|
|
|
U2
|
|
|
|
19
18
|
|
|
|
|
|
|
14
|
|
|
|
|
U1
|
|
7
6
5
4
3
2
1
3 m
|
|
60
Fig.25a répartition des foraminifères
planctoniques et des Radiolaires dans la coupe de Oued El Kharroub
(région de Bargou)
61
Age
|
Formations
|
Biozones
|
Sous-Zones
|
Radiolaires
|
Épaisseur
|
Échantillons
|
Lithologie
|
COT
0 5 10
|
513C
-25 -24 -23 -22
|
Richesse spécifique
des
Foraminifères
et Radiolaires
0 5 10 15 20 25
|
|
Cénomanien moyen Cénomanien supérieur
Turonien inférieur
|
Fahdène Bahloul Kef
|
H.
helvetica
|
Guttacapsa biacuta Alievium superbum
|
57
54
51
48
45
42
39
36 33 30 27
24
21
18
15
12
9
6
3
|
|
99 98 97 96 95 94 93 92 91 90 89 88 87
86
84
81 80 79 78
77
75
72
69 68 67 66 6654 63 62 61 60 59 58 57 56
55
54
52 51 50 49 48
43 42 41 40
35 34 33 32 31 30 29 28 27 26 25 24
23
21
18
16
14
13
12
11
9
7
4
|
100
|
|
47 46
La limite C/T
|
|
|
|
|
|
7
|
|
Unité
|
|
|
|
Whiteinella archaeocretacea
|
Gl. Dicarinella hagni Heterohelix moremani
benton
|
|
|
|
|
|
|
|
85
83
82
|
|
|
|
U
|
|
|
|
76
74
73
|
|
|
71
70
|
|
|
|
|
|
|
|
U
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
53
|
|
|
|
Un
|
III
|
|
|
|
|
45
44
|
|
|
EAO-2
ECM: Événement
|
|
|
|
|
39
38
|
|
37
|
|
Unité
|
|
II
|
|
|
36
|
|
|
Rotalipora cushmani
|
R.
greenhornensis Dicarinella algeriana
|
|
|
|
|
|
|
|
|
I
du Cénomanien
|
moyen
|
|
|
|
22
|
|
20
19
|
|
17
|
|
|
15
|
|
|
10
8
6 5
3
|
|
|
2
1
|
Fig.25b - Carbone organique total, d13C (Barrett, 1998); et
richesse spécifique des Foraminifères planctoniques et des
Radiolaires dans la coupe Oued El Kharroub (région de Bargou)
62
benthiques représentées par des Gavelinella,
Cassidella et Dentalina. Notons que R. montsalvensis disparaît dans
l'échantillo COK-25 récolté au tiers supérieur de
cette unité. Cette association indique un âge Cénomanien
moyen supérieur.
c) L'Unité U3 [COK-30 - COK-37]
Le faciés change progressivement. L'unité
U3 débute par un niveau calcaire argileux de couleur beige
épais de 2m supportant des calcaires laminés durs à patine
gris foncé et se débitant en plaquettes suivis par une alternance
de calcaires beiges et des calcaires argileux de couleur noirâtre. Cette
unité qui mesure 4,5m, a livré : Rotalipora cushmani, R.
greenhornensis, Praeglobotruncana stephani, P. gibba, Whiteinella aprica, W.
paradubia, W. aumalensis, W. inornata, Heterohelix moremani, H. navarroensis,
H. pulchra, H. reussi, Hedbergella planispira, H. simplex, H. delrioensis,
Globigerinelloides bentonensis, G. ultramicra. Au sein de cette
unité on note la disparition de Praeglobotruncana delrioensis
dés l'échantillon COK-35 et l'apparition de deux
espèces : Whiteinella archaeocretacea COK-30 et
Anaticinella multiloculata COK-37.
A partir de l'échantillon COK-36 et surtout COK-37,
l'association de Radiolaires est marquée par un épanouissement
important. Cette association est représentée essentiellement par
la famille des Nassellaria. On note la présence de Novixitus
mclaughlini, Novixitus sp, Pseudodictyomitra tiara, Xitus picenus, X.
spicularius, Pseudodictyomitra carpatica, P. pentacolensis, P.
pseudomacrocephala, Stichomitra communis. L'association de
Foraminifères planctoniques est caractéristique du
Cénomanien supérieure à Cénomanien sommital.
d) L'Unité U4 [COK-38 - COK-61]
Cette unité est composée essentiellement par des
alternances de niveaux marneux et/ou des calcaires argileux de couleur
noirâtre se débitant en plaquettes et des niveaux calcaires
siliceux de couleur beige clair se débitant en petits prismes. C'est
dans cette unité que se fait la disparition des derniers Rotalipores
(Rotalipora greenhornensis et R. cushmani) et celle de
Globigerinelloides bentonensis et Praeglobotruncana gibba,
coincidant avec l'apparitions de Dicarinella hagni, D. imbricata et
Whiteinella praehelvetica. L'association de Foraminifères
planctoniques est dominée par les
Hétérohélicidés (H. moremani, H. reussi, H.
pulchra, H. navarroensis et H. sp) ainsi que
63
Hedbergella planispira, Guembelitria cenomana et
G. albertensis. Dans les premiers niveaux de cette unité
l'association est dominée par les Radiolaires. Leur fréquence est
d'environ 60% de l'association si l'on tient compte de tous les autres
microorganismes, Foraminifères planctoniques, Foraminifères
benthiques, Ostracodes). Dans l'échantillon COK-56, ces Radiolaires sont
représentés essentiellement par la fammille des Sphaerellaria de
très petite taille et on note bien un renouvellement faunique concernant
ce groupe planctonique. De tel renouvellement noté au sein du groupe a
été signalé par plusieurs auteurs en Italie (O'Dorghety,
1994, Salvini et Marcucci Passerini, 1998 ; Premoli Silva et al,
1999).
Cette unité est caractérisée aussi par
l'abbondance de grains de quartz, grains de phosphate. La présence de W.
archaeocretacea sans les Rotalipores indiquerait le passage Cénomanien
sommital - Turonien basal.
e) L'Unité U5 [COK-62 - COK-77]
Elle est composée de deux barres calcaires, à
patine beige et cassure fraîche noire, séparées par un
niveau marneux à silex. La deuxième barre est soulignée
par un niveau comportant des nodules d'oxyde de Fer.
Elle mesure 8m. L'association de Foraminifères
planctoniques est représentée par : Whiteinella aprica, W.
archaeocretacea, quelques occurrences de P. stephani, W.
aumalensis. Toutefois, cette association est dominée par
Heterohelix moremani, H. reussi, H. pulchra, H. planispira, H.
delrioensis. On note aussi quelques occurrences timides de
Anaticinella multiloculata. Cette association idique le passage
Cénomanien sommital-Turonien basal.
f) L'Unité U6 [COK-78 - COK-89]
Cette unité est marquée par la présence
des figures de slumps accompagnés par des blocs
décimétriques à métriques enracinés dans les
dépôts de la Formation Bahloul, ce sont les Olistolites. Ces blocs
qui témoignent de la mobilité du plancher sédimentaire
durant le dépôt de ladite formation, ont été
signalés déjà par plusieurs auteurs (Turki, 1975 ; Layeb,
1990 ; Rami, 1998 ; Layeb et Belayouni, 1999). Ce terme a livré :
Whieteinella archaeocretacea (présence sporadique), W.
aprica, W. baltica, W.
64
aumalensis, Dicarinella hagni, D. imbricata et la
dominance des forme bisériées et trisériées
(Heterohelix moremani, H. reussi, Guembelitria cenomana, G.
albertensis). On note la disparition de Anaticinella multiloculata
à partir de l'échantillon COK-79 et l'abondance de W.
praehelvetica. Cette association de Foraminifères planctoniques
indiquerait un âge Turonien basal.
g) L'Unité U7 [COK-90 - COK-100]
Ce terme est essentiellement formé par des calcaires
argileux ou siliceux. Ce terme a livré l'association de
Foraminifères planctoniques suivante : Whiteinella praehelvetica,
Dicarinella imbricata, Heterohelix reussi, H. moremani, Guembelitria cenomana,
G. albertensis, Helvetoglobotruncana helvetica, Marginotruncana renzi,
Marginitruncana sp. On note l'apparition de H. helvetica dés
l'échantillon COK-92 et les Marginotruncana dés
l'échantillon COK-93. Cette association est caractéristique du
Turonien inférieure à moyen.
II - Interprétation des environnements de
dépôts
La présence de débris d'échinodermes et
l'abondance des calcisphères nous laisse supposer qu'ils ont
été déplacés par des courrants forts. La
présence de Foraminifères planctoniques bien diversifiées
dans les premiers niveaux associés à des filaments (=
lamellibranches) et de rares Foraminifères benthiques nous renseigne sur
des conditions marines normales de mer ouverte de type bathyal
supérieur. Les dernières Rotalipores disparaissent dans les
niveaux à black shales riches en matière organique (à
odeur de bitume) enregistrant l'installation d'un environnement hypoxyque voire
anoxique, là où la productivité primaire est
élevé selon les données isotopiques de
ä13C (Keller et al., 2001 ; Keller et Pardo, 2004 ; Keller al.,
2004). Cet événement biologique a été
déjà signalé dans plusieurs régions à savoir
le bassin Vocontien (Crumière, 1989), Chalk sea (Jarvis et al.,
1988), l'Océan Atlantique (Kuhnt et al., 1986), en Italie
(Premoli Silva et Sliter, 1994 ; Premoli Silva et al., 1999), en
Espagne (Lamolda et al., 1997), en Amérique du Nord (Schlanger
et Jenkyns, 1977 ; Keller et Pardo, 2004 ; Keller et al., 2004) et
même en Turkie (Sükrü Yurtsever et al., 2003).
Après la disparition des Rotalipores les Foraminifères sont
représentées exclusivement par des formes à loges
globuleuses qui peuvent être des
65
formes bisériées, trisériées ou
trochospiralées. Leur diversité diminue considérablement
(5 espèces). L'alternance entre des niveaux clairs et des niveaux
sombres indique que le milieu devient par périodes à
oxygène minimum et même épuisé en oxygène. De
temps en temps, les eaux de surface seraient peut être enrichies en
oxygène et en nutriments probablement par l'intervention des eaux
hyposalées venant du continent. Les bancs riches en Ammonites et
à surface bioturbée sont généralement de couleur
noirâtre, mais les bioturbations nous laisse supposer un milieu peu
oxygéné au moins à l'interface eau/sédiment.
La disparition des débris d'échinodermes et des
calcisphères est interprétée comme étant le
début d'une phase transgressive (Robaszynski et al., 1990 ;
1993).
La rareté voire l'absence des Foraminifères
benthiques ni le fait que le fond marin était oxygéné
durant l'intervalle CES-25 - CES-30 ; COK-39 - COK42.
La dissoulution observées au niveau des
échantillons CES-17 et CES-18 nous renseigne sur une période de
forte
L'abondance des radiolaires dans les niveaux COK-37
jusqu'à COK-46 nous laisse supposer que le milieu à
été favorable pour le développement de ces planctoniques
au profit des foraminifères. Leur distribution verticale montre que le
changement dans leur assaemblage était brusque. Les conditions
eutrophiques ont favouri le développement des radiolaires.
Concernant la coupe COK, la présence de figures de
slumps et d'olistolithes indiquent la mobilité du plancher
sédimentaire lors du dépôt de la Formation Bahloul au moins
dans la région de Bargou.
66
TROISIÈME PARTIE
BIOSTRATIGRAPHIE
67
I - Biostratigraphie
A - Biozonation des Foraminifères planctonique du
passage Cénomanien - Turonien.
1 - Historique
De nombreux travaux biostratigraphiques et
micropaléontologiques ont discuté la répartition
biostratigraphique de la microfaune du passage Cénomanien-Turonien et
ont abordé les limites de cet intervalle :
Sigal (1955), en se basant sur la microfaune, a définie
originellement la Zone à « Grandes Globigerines seules et
dominantes ». Considérées comme des «
Globigérines de grande taille et massives », elles ont
été observées par Schijfsma (1955) puis Lehmann (1962).
Bolli (1966) a définit la Zone à Praeglobotruncana gigantea
en se référant à Lehmann et à Van Hinte
(1965). Salaj (1976), quant à lui, définie en cet intervalle les
Zones à Rotundina cretacea (synonyme de W.
archaeocretacea) et Praeglobotruncana imbricata.
Et pour mieux cerner cette limite entre le Cénomanien
et le Turonien de multiples travaux concernant plusieurs groupes de fossiles
(Foraminifères planctoniques , Radiolaires, nannofossiles calcaires) de
l'intervalle Cénomanien supérieur - Turonien inférieur ont
été élaboré. Concernant les Foraminifères
planctoniques, les principaux sont :
Salaj (1980) a mentionné la zone à
Rotalipora turonica, caractéristique du passage
Cénomanien - Turonien. L'auteur précise que cette zone à
R. turonica détermine la partie supérieure de la zone
à Rotalipora cushmani s.l. Elle est succédée par
la zone à Rotundina cretacea - Praeglobotruncana imbricata
(qui correspond à la zone à grandes globigérines
seules de Sigal (1955) et détermine la partie supérieure du
Turonien inférieur.
Concernant Rotalipora turonica telle qu'elle a
été figurée par Salaj (idem, pl.8 fig.11) est très
proche de R. montsalvensisnt aussi à la zone à Rotalipora
cushmani cushmani sens large.
68
La zone à Rotundina cretacea -
Praeglobotruncana imbricata. Elle correspond à la partie
supérieure du Turonien inférieur.
La zone à Helvetoglobotruncana helvetica
succédant la zone R cretacea - P imbricata indique le
Turonien moyen, il l'a subdivisé en trois sous-zones à savoir
:
· la sous-zone à Praeglobotruncana oraviensis
trigona.
· La sous-zone à Praeglobotruncana
biconvexa
· La sous-zone à Globotruncana turona.
La Formation Bahloul s'étend sur les Zones à
Rotalipora turonica et à R. creatacea-P. imbricata. Ainsi, pour Salaj,
elles est d'âge Cénomanien supérieur pp - Turonien
inférieur.
Le même auteur propose un tableau de corrélation
des ces biozones de Foraminifères planctoniques avec celles des
Ammonites (Tableau p.72). Il considère donc que la Zone à R.
turonica s'étend sur les Zones à Ammonite à C. naviculare
(partie supérieur) à C. crassum indiquant pour lui le
Cénomanien supérieur, à M. geslinianum et à F
supertes du Turonien basal à Turonien inférieur.
Bellier (1983), quant à lui, concernant Rotalipora
montsalvensis, il signale que cette espèce est rare. Il rajoute que
la sous-zone à Rotalipora montsalvensis, dont l'indicateur
apparaît avant Rotalipora cushmani, caractérise le
Cénomanien moyen. Ainsi, Bellier propose de subdiviser la Zone
à Rotalipora cushmani en remarquant que Dicarinella
algeriana s'individualise au sein de celle-ci. C'est pour cette
raison qu'il a conçu une zone accessoire, comprise entre l'apparition de
Dicarinella algeriana et la disparition de Rotalipora
cushmani.
Le même auteur propose une zonation différente
de celle de Salaj (1980), il attribue le passage Cénomanien - Turonien
à la zone d'intervalle à Whiteinela aprica.
La limite inférieure de cette est indiquée par la
disparition de Rotalipora cushmani. Sa limite supérieure est
définie par la première occurrence de Helvetoglobotruncana
helvetica du Turonien inférieur à moyen. Il a
constaté que la constance de l'apparition simultanée
de Whiteinela aprica, Whiteinella paradubia et
Dicarinela imbricata, coïncide assez bien avec
la disparition de Rotalipora cushrnani et de Rotalipora turonica.
Ainsi l'auteur n'a pas dégagé, en Tunisie, de
décalage entre l'apparition des « Grandes Globigérines
» (qui se confondent avec les Whiteinelles) et la disparition des
Rotalipores, conformément à ce qui a été
signalé par Sigal (1955).
69
Sigal (1985) distingue dans le Cénomanien moyen et
supérieur la zone à Rotalipora cushmani qu'il l'a
subdiviséeen trois sous-zones à savoir :
· sous-zone à Rotalipora appenninica qui
caractérise la partie inférieure de la zone à
Rotalipora cushmani.
· sous-zone à Globotruncana algeriana
déterminant la partie moyenne de la zone à Rotalipora
cushmani.
· sous-zone à Helvetoglobotruncana
paradubia.
Sigal (1987) reprend l'intervalle le Turonien qu'il subdivise
en Zone à Whiteinella paradubia - Whiteinella aprica
(caractérisant le passage Cénomanien-Turonien).Il
dégage la zone à Globotruncana helvetica du Turonien
inférieur et la zone à Globotruncana sigali - Globotruncana
schneegansi du Turonien supérieur.
Robaszynski et al. (1990) ont attribué
l'ensemble de la Formation Bahloul affleurant dans la région de
Kalâat Senan au Cénomanien supérieur en s'appuyant sur
l'existence dans la partie supérieure de ladite Formation d'une
espèce d'ammonite Euomphaloceras cf. septemseriatum (CARGIN)
qu'il considère du Cénomanien terminal.
Maâmouri et al., (1994), en étudiant la
Formation Bahloul en Tunisie centro - septentrionale, admettent que le
Cénomanien supérieur est caractérisé par
l'extension de l'espèce Rotalipora cushmani qui est
associée à Whiteinella archaeocretacea ou à
Whiteinella brittonensis. Les mêmes auteurs indiquent que la
Formation Bahloul, dans les différents affleurements
étudiés, débute dans la partie supérieure de la
biozone à Rotalipora cushmani du Cénomanien
supérieur et persiste dans la biozone à Whiteinella
archaeocretacea du passage Cénomanien - Turonien. La zone à
H. Helvetica étant décrite dans la Formation sus-jacente, la
Formation Kef (Fournié, 1978)
Ils ont admis que la biozone à W. archaeocretacea
correspond à celle originellement nommée à «
grandes Globigérines seules et dominantes » par Sigal (1955), ou
à « Praeglobotruncana gigantea » par Lehmann (1962),
Van Hinte (1965) et de Bolli (1966), ou encore zone intervallaire à
Whiteinella aprica définie par Bellier (1983).
70
Ils ont considéré aussi que l'espèce
Praeglobotruncana gigantea est synonyme de Whiteinella
archaeocretacea.
Ils ont attribué ainsi un âge Cénomanien
supérieur - Turonien inférieur à la Formation Bahloul dans
sa localité type qu'ailleurs.
Concernant les Ammonites,
Dans la même année Ben Haj Ali et al.,
(1994) arrivent à la même conclusion concernant l'âge
de la Formation Bahloul. Cette formation englobe la partie terminale de la Zone
terminal à Rotalipora cushmani d'âge Cénomanien
supérieur et la Zone à Whiteinella archaeocretacea
déterminant la passage C/T. Le toit de ladite Formation coincide
avec la zone à Praeglobotruncana helvetica.
Les mêmes auteurs reconnaissent dans la partie
inférieure de la Formation Bahloul. Des espèces d'Ammonites
indicatrices des zones à Metoicoceras geslinianum et à
Neocardioceras judii du Cénomanien supérieur. Les
Ammonites du genre Fagesia déterminant l'entrée au
Turonien, ont été récoltées au toit de la Formation
Bahloul
Dans leurs travaux sur la limite Cénomanien - Turonien
en Tunisie centrale et méridionale, Razgallah et al., (1994)
ont attribué le membre Gattar au Turonien basal en s'appuyant sur
l'existence de Pseudaspidoceras flexuosum qui indique le début
du Turonien.
Au cours de ses études sur la Tunisie du Centre et du
Sud, Abdallah (1995) a constaté que les dépôts
situés à la limite Cénomanien - Turonien au J. Asker et au
J. Zitouna (chaîne nord des Chotts), comprennent :
É des alternances argilo-carbonatées
d'âge Cénomanien supérieur à huîtres, rudistes
(Praeradiolites biskraensis), polypiers, foraminifères
(Thomasinella punica).
· des calcaires blancs micritiques à
huîtres naines et ammonites (Neolobites vibrayeaus, Calycoceras
sp.) qui indiquent le Cénomanien sommital.
· des alternances de la Formation Bahloul à
matière organique telles que à Khanguet Besbessa, ou bien leurs
équivalents calcaires blancs alternativement feuilletés et
massifs à organismes nectoniques et planctoniques d'âge
Cénomanien supérieur terminal - Turonien inférieur basal
(Abdallah, 1987 ; 1995)
·
71
des dolomies du membre Gattar bioclastiques et à
stratifications obliques de grandes tailles sur la partie inférieure, au
J. Asker, contenant des rudistes au J. Zitouna.
Le même auteur ajoute que la transgression du
Cénomanien supérieur - Turonien inférieur est
considérée comme étant la plus importante parmi celles du
Crétacé, qui selon lui, toutefois, elle serait surestimée,
étant donnée qu'elle est secondée par une anoxie. Cette
anoxie a, en effet, maintenu l'apparition des groupes néritiques sur les
plates-formes carbonatées et a favorisé, au contraire,
1'etalement d'un faciès a caché bathyal sur des plates-formes
superficielles et ceci contrairement un reste des transgressions du
Crétacé qui avaient favorisé une importante vie
néritique sur les mêmes aires paléogéographiques
(Abdallah, 1995).
Rami et al., (1997) et Rami (1998) définissent
la zone à Whiteinella archaeocretacea comme étant une
zone intervallaire à Whiteinella archaeocretacea correspondant
à l'intervalle compris entre l'extinction de tous les Rotalipores et
l'apparition de Helvetoglobotruncana helvetica. Elle marque le passage
Cénomanien - Turonien comme elle a été adoptée par
Robaszynski et al. (1995). Ainsi ils la considèrent synonyme de
la zone à Whiteinella aprica de Bellier (1983). Dans la zone
à Whiteinella archaeocretacea,Les mêmes auteurs ont
constaté que de nombreuses autres espèces du même genre
s'épanouissent.
Dans les travaux inédits effectués par Bismuth
(1997, in Layeb et Belayouni 1999) dans le cadre d'un projet
pour l'Office Nationale des Mines dans la région de Oued
Bahloul, l'auteur a mentionné l'extinction des Rotalipora cushmani
au mur de ladite Formation et il a souligné l'absence des
Helvetoglobotruncana helvetica au toit de la Formation. Seule
Whiteinella praehelvetica est présente. Ceci est non-conforme
aux travaux de Maâmori et al., (1994) et de Bel Haj Ali et
al., (1994) et semble être non-conforme à la
réalité.
2 - Biozonation des Foraminifères
planctoniques
Un échantillonage serré dans l'intervalle des
dépôts du Cénomanien supérieur à Turonien
inférieur nous a permis d'affiner la biostratigraphie selon le
modèle appliqué pour la biostratigraphie à haute
résolution. Pour les Zones, nous avons adopté la biozonation
72
proposée par Caron (1985). Concernant la subdivision en
sous-zones, nous avons adopté le modèle proposé par Keller
et al., (2001) pour la zone à Rotalipora cushmani du
Cénomanien supérieure et Keller et Pardo (2004) pour la zone
à Whiteinella archaeocretacea du passage Cénomanien
Turonien.
N'ayant travaillé dans le détail que la partie
supérieure de la zone à R. cushmani nous avons reconnu
la sous-zone à Dicarinella algeriana.
a) Zone à Rotalipora cushmani.
Elle est définie par l'extension totale de
l'espèce index Rotalipora cushmani (TRZ : Total Range Zone).
Comme l'ont adopté d'autres auteurs (Sigal, 1977, Salaj, 1980 ; Bellier,
1983, Sigal, 1987, Maâmouri et al., 194 ; Premoli Silva et
Sliter, 1995), la Zone à R. cushmani caractérise le
Cénomanien moyen à supérieur. Cette Zone est
définie à l'origine par Bolli (1966). Elle correspond à la
Zone à Rotalipora de Dalbiez (1956) (zones 7a et 7b), la Zone
UC2 et UC3 de Van Hinte (1976). Nous avons reconnu la
Zone à Rotalipora cushmani dans la région de Tajerouine
(CES) et dans la région de Bargou (COK). Dans les deux coupes
l'espèce Rotalipora montsalvensis est présente. Cette
dernière espèce s'éteint bien avant R. cushmani.
La Zone à Rotalipora cushmani a été reconnue dans
le domaine téthysien (Barr, 1972 ; Sigal, 1977 ; EWGPF, 1979.1 ;
Wonders, 1980 ; Bellier, 1983 ; Robaszynski et Caron, 1995 ; Premoli Silva et
Sliter, 1995) et dans le domaine boréale (Sigal, 1987, Georgescu, 1995 ;
2000).
Dans la coupe d'Oued El Kharroub (COK), la Zone à R.
cushmani englobe partiellement la sous-zone à Rotalipora
greenhornensis et la totalité de la sous-zone à
Dicarinella algeriana A propos de la coupe d'Oued Es Smara (CES)
l'échantillonnage n'a concerné que la partie supérieure de
la sous-zone à D. algeriana.
- La sous-zone à Dicarinella hagni
Cette sous-zone est définie dans sa limite
inférieure par l'apparition de l'espèce Dicarinella algeriana
et dans sa limite supérieure par la disparition de Rotalipora
cushmani et toutes les Rotalipores. Dans la coupe COK cette sous-zone est
observée dans sa totalité. L'échantillon
73
COK-8 correspond à l'horizon dans lequel se fait la
première occurrence de l'espèce indicatrice. Cette sous-zone est
épaisse de 23m dans la coupe COK. N'ayant pas observé
l'apparition de Dicarinella algeriana dans la coupe CES, l'ensemble de
cette sous-zone est supposé incomplet. Dans cette sous-zone, s'effectue
la disparition de R. montsalvensis suivie par celle de
Praeglobotruncana delrioensis dans la partie supérieure de
cette sous-zone. Ces deux disparitions sont suivies par l'apparition de
Whiteinella archaeocretacea bien avant la diparition de R.
cushmani. Entre autre, au niveau de cette sous-zone on a noté un
pic dans la courbe d'abondance relative de Guembelitria cenomana (30%)
suivi d'un pic pour Heterohelix moremani (50%). On remarque aussi
l'abondance des Foraminifères benthiques (25%)
représentées par Cassidella, Frondicularia, Gavelinella,
Buliminella, Cibicidoides et Textularia rioensis dans la partie moyenne de
cette sous-zone. Cet intervalle mésure 5m
b) Zone à Whiteinella archaeocretacea
La Zone intervavallaire à Whiteinella
archaeocretacea correspond à l'intervalle compris entre
l'extinction de toutes les Rotalipores et la première occurrence de
Helvetoglobotruncana helvetica. Elle marque le passage
Cénomanien - Turonien comme elle a été adoptée par
plusieurs auteurs (EWGPF, 1979.1 ; Maamouri et al., 1994 ; Robaszynski et
Caron, 1995 ; Premoli Silva et Sliter, 1995 ; Georgescu, 2000). Nous la
considérons équivalente à la partie sommitale de la zone
à Rotalipora turonica et la partie basale de Rotalipora
cretacea - Praeglobotruncana imbricata de Salaj (1980) à
la Zone à Whiteinella aprica de Bellier (1983), à la Zone
à Whiteinella paradubia - W. aprica de Sigal (1987). Nous l'avons
reconnue dans les deux coupes (CES et COK) dans la région de Tajerouine
et dans la région de Bargou. Cette Zone intervallaire a
été reconnue en Tunisie septentrionale (Maamouri et al.,
1994 ; Rami, 1992 ; 1998) en Tunisie centro-septentrionale et en Tunisie
centrale (Robaszynski et al., 1990 ; 1993 ; Ben Haj Ali et al.,
1994 ; Maamouri et al., 1994) et en Tunisie nord-orientale (Rami,
1998). Plusieurs auteurs (Schijfsma, 1955 ; Sigal, 1955 ; Lehman, 1962) ont
remarqué l'épanouissement de nombreuses autres espèces du
même genre définie par Pessagno (1967).
L'espèce Whiteinella archaeocretacea a
été retenue par plusieurs auteurs comme marqueur du passage
Cénomanien - Turonien aussi bien en domaine téthysien (Caron,
1985 ; Maamouri et al., 1994 ; Ben Haj Ali et al., 1994 ;
Robaszynski et
74
Caron, 1995, Premoli Silva et Sliter, 1995) qu'en domaine
boréal (EWGPF, 1979.1 ; Georgescu, 2000).
Nous subdivisons cette Zone en trois sous-zones : la sous-zone
à Globigerinelloides bentonensis, la sous-zone à
Dicarinella hagni et la sous-zone acmé à Heterohelix
moremani.
- La sous-zone à Globigerinelloides
bentonensis
La sous-zone à Gl. bentonensis est
définie par l'intervalle compris entre la disparition de R. cushmani
et la disparition de Gl. bentonensis. Dans la région de
Jerissa comme celle de Bargou cette sous-zone marque le tiers inférieur
de la Zone à W. archaeocretacea. Cette sous-zone s'etend sur 5m
dans la coupe d'Oued Es Smara (région de Jerissa) et seulement sur 3m
à Oued El Kharroub (région de Bargou). La sous-zone à
Globigerinelloides bentonensis est marquée par l'apparition de
Whiteinella praehelvetica suivie par celle de Dicarinella hagni
et par la disparition de Anaticinella multiloculata (uniquement
à CES) chose qu'on n'observe pas au niveau de la coupe COK.
Concernant W. praehelvetica, Keller et al.,
(2001) et Keller et Pardo (2004) indiquent qu'elle disparaît bien
avant la disparition de R. cushmani alors que dans notre
matériel W. praehelvetica n'apparaît que dans la partie
inférieure de la zone à W. archaeocretacea. Cette
sous-zone est dominée par une association diversifiée de
Heterohelix moremani, Guembelitria cenomana, Hedbergella delrioensis,
Dicarinella hagni, Whiteinella aprica et Globigerinelloides ultramicra.
- La sous-zone à Dicarinella hagni
Cette sous-zone est définie par l'intervalle compris
entre la disparition de Gl. bentonensis et le niveau à
abondance des Heterohelix. Au niveau de la coupe CES, Dicarinella
hagni apparaît dans l'échantillon CES-16 avant la disparition
de Gl. Bentonensis qui se fait dans l'échantillon CES-23. Cette
sous-zone s'étens sur 6m dans la coupe CES et sur 9m dans la coupe
COK.
- La sous-zone à Heterohelix moremani
75
La limite inférieure de cette sous-zone est
déterminée par l'abondance ou l'acmé des
Heterohelicidés (Heterohelix shift). Sa limite
supérieure est déterminée par l'apparition de
Helvetoglobotruncana helvetica. L'acmé ou le pic de
fréquence des Hétérohélicidés a
été reconnu dans plusieurs localités du monde : à
Rock Canyon (Western Interior), (Leckie et al., 1998) ; à Oued
Mellègue en Tunisie (Nederbraght et Fiorentino, 1999) ; à
Eastbourne en Angleterre (Keller et al., 2001) ; à Pueblo
(Keller et Pardo, 2004). Cette sous-zone s'étend sur 6,5m dans la coupe
CES et sur 16m dans la coupe COK.
c) Zone à Helvetoglobotruncana helvetica
La Zone à Helvetoglobotruncana helvetica est
définie par l'extension totale du fossile index. Cette zone a
été définie à l'origine sous le nom de
Globotruncana helvetica, par (Sigal, 1955 ; Dalbiez, 1955). Par la
suite, Bolli (1966) a fixé ses limites d'une manière nette. Telle
qu'elle a été considérée ici, la Zone à
Helvetoglobotruncana helvetica indique le Turonien inférieur
à moyen comme l'a adoptée Sigal (1977 ; 1987). Pour Bellier
(1983) et Robaszynski et Caron (1995), cette Zone se termine peu avant la fin
du Turonien moyen. Cependant, auparavant, Schijfsma (1955) a retenu la Zone
à Globotruncana helvetica pour marquer le début du
Coniacien et Postuma (1971) l'a fait correspondre à tout le Turonien.
Nous avons reconnu cette Zone à Helvetoglobotruncana helvetica
dans les deux coupe (CES et COK) respectivement dans la région de
Tajerouine et dans la région de Bargou. Dans cette zone, nous remarquons
que plusieurs autres espèces s'épanouissent notamment ceux du
genre Marginotruncana et particulièrement Marginotruncana
renzi.
Notons que la Zone à Helvetoglobotruncana helvetica est
reconnue mondialement. Dans le domaine téthysien (Schijfsma, 1955 ;
Postuma, 1971 ; Barr, 1972 ; Van Hinte, 1976 ; Sigal, 1977 ; Wonders, 1980 ;
Salaj, 1980 ; Bellier, 1983 ; Sigal, 1987 ; Robaszynski et Caron, 1995 ;
Premoli Silva et Sliter, 1995), en domaine boréal (Van Hinte, 1976 ;
EWGPF, 1979.1 ; Georgescu, 1995 ; 2000), en Amérique (Douglas, 1969 ;
Keller et Pardo, 2004).
Conclusion : les épaisseurs de ces différentes
zones et sous-zones ce sont revélées inégales en passant
d'une région à une autre (Fig.26)
|
Age
|
Formations
|
Biozones
|
Sous-Zones
|
Radiolaires
|
Épaisseur
|
Échantillons
|
Lithologie
|
|
Cénomanien moyen Cénomanien
supérieur Turonien inférieur
|
Fahdène Bahloul Kef
|
H.
helvetica
|
Guttacapsa biacuta Alievium superbum
|
57
54
51
48
45
42
39
36
33
30
27
24
21
18
15
12
9
6
3
|
99 98 97 96 95 94 93 92 91 90 89 88 87 86 85 84 83
81 80 79 78 77 76 75 74 73 72
69 68 67 66 65 64 63 62 61 60 59 58 57 56
55
54
52 51 50 49 48 47 46 45 44 43 42
40
39
38
31 30 29 28 27 26
25
22
21
19
18
16
14
12
11
9
4
|
100
|
|
|
|
|
7
|
|
Unité
|
|
|
|
|
Whiteinella archaeocretacea
|
Gl. Dicarinella hagni Heterohelix moremani
benton
|
|
|
|
82
nité 6
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
71
70
|
|
|
|
U
|
|
|
|
|
|
|
|
|
|
53
|
|
|
|
|
|
|
U
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Unité 2 Unité 3
|
|
37
36
35
34
33
32
|
|
|
Rotalipora cushmani
|
R.
greenhornensis Dicarinella algeriana
|
|
|
24
23
|
|
|
20
|
|
17
|
|
15
|
|
13
10
8
7
6
5
3
|
|
|
2
1
|
zone à
Sous zone à
Dicarinella
algeriana
Sous zone à
Heterohelix moremani
Sous zone à
Globigerinelloides bentonensis
Sous zone à Dicarinella hagni
|
Age
|
Formation
|
Zone
|
Sous-zone
Épaisseur
Échantillon Lithologie
|
Unité
|
|
Cen. moy Cénomanien supérieur Tur. inf
|
Fahdène Bahloul Kef
|
Rotalipora cushmani Whiteinella archaeocretacea H.
helvetica
R. montsalvensis D. algeriana G. bentonensis D. hagni H.
moremani
|
22 65
|
U6
|
|
|
|
|
|
|
|
20 60
|
|
|
|
|
|
|
|
|
|
|
|
|
55
|
|
|
|
|
|
|
18
|
|
|
|
50
|
|
|
|
|
|
|
|
|
|
|
|
|
16 45
|
|
U5
|
|
|
|
|
40
|
|
|
|
|
|
|
14 35
|
|
|
|
|
|
|
|
|
|
30
|
|
|
|
12
|
|
U4
|
|
25
|
|
|
|
|
|
|
0 20
|
|
|
|
|
|
|
|
|
|
15
8
10
|
U3
|
|
|
|
5
6
4
3
4 2
1
2
|
U2
|
|
U1
1m
|

76
Fig. 26 - variation d'épaisseur entre les deux coupes
(COK et CES)
77
B - Biostratigraphie des Radiolaires du passage Cénomanien
- Turonien. Historique
Plusieurs arrangements biostratigraphiques ont
été proposés pour les Radiolaires du Crétacé
supérieur. Dans ce travail, nous allons suivre les biozones
proposées par Pessagno (1976, 1977), Thurrow (1988), O'Dogherty (1994),
Salvini et Marcucci Passerini (1998), Premoli Silva et al., (1999) et Sukru
Yurtsever et al., (2003) concernant l'intervalle Cénomanien -
Turonien. Nous avons reconnu des Radiolaires aussi bien dans la coupe de Bargou
(COK) en abondance que dans la coupe de Tajerouine (CES) qui peuvent être
confondus avec les Calcisphaerulidae. Dans la Coupe d'Oued El Kharroub
(région de Bargou), l'épanouissement des Radiolaires survient
dans la partie supérieure de la Zone à Rotalipora cushmani
et se poursuit sur quelques mètres de la zone à
Whiteinella archaeocretacea. Ensuite, dans la partie inférieure
de la Zone à Whiteinella archaeocretacea. Deux associations
différentes ont été distinguées : celle du
Cénomanien supérieur est dominée par les Nassellaria et en
particulier Pseudodictyomitra et celle du Turonien est
caractérisée par l'abondance des Sphaericularia.
Quelques espèces demeurent communes à ces deux associations
cénomano-turoniennes. Le fait que la majorité des espèces
qui y sont différentes cela nous permet de mettre en évidence un
renouvellement faunique marquant aussi les Radiolaires du passage C/T.
De tel renouvellement a été signalé
également par Marcucci et al., (1991), O'Dorghety (1994),
Erbacher et Thurrow (1997), Salvini et Marcucci Passerini (1998) et Premoli
Silva et al., (1999). En effet, ces auteurs mentionnent qu'au cours de
l'intervalle Cénomanien - Turonien un grand renouvellement faunique
concernant les Radiolaires s'est produit. Ces auteurs ont dégagé
alors deux associations de Radiolaires importantes. L'association
« inférieure » est caractérisée
particulièrement par Dactyliosphaera silviae
et
Rhopalosyringium majuroensi marqueurs de zone (O'Dorghety,
1994) du Cénomanien supérieur. L'association «
supérieure » est caractérisée particulièrement
par l'apparition de plusieurs espèces parmi lesquelles Alievium
superbum et Crucella cachensis indicateurs de zone du passage C/T
qui peut être correlée avec celle à W. archaeocretacea
des Foraminifères planctoniques. Plusieurs espèces de cette
association appartiennent au
78
Turonien inférieur. Salvini et Marcucci Passerini (1998)
admettent que cette association appartient à la Zone à
Crucella cachensis.
79
QUATRIÈME PARTIE :
PALÉOÉCOLOGIE DES
FORAMINIFÈRES
PLANCTONIQUES ET DES RADIOLAIRES DU
PASSAGE CÉNOMANIEN - TURONIEN
80
A - Paléoécologie des
Foraminifères planctoniques du passage Cénomanien - Turonien
1) Introduction
Les foraminifères planctoniques ont prouvé leur
rôle capital pour la biostratigraphie à haute résolution en
particulier pour l'intervalle Albien - Turonien. Leur répartition a
été utilisée pour fonder les reconstitutions
paléoécologiques et paléobiogéographiques.
Toutefois leur usage dans les études de paléoécologie
détaillée et la détermination précise des
conditions environnementales reste beaucoup trés limitée. Faute
de données disponibles sur l'écologie des Foraminifères
vivants en termes des limites de leurs tolérances aux différents
paramètres écologiques (température, salinité,
bathymétrie, taux d'oxygène dissous et l'impact des Variations de
ces paramètres sur les populations et les communautés de ce
groupe d'organismes). Toutefois les interprétations
paléoécologiques des espèces disparues et des assemblages
associés doivent être abordées avec soin.
Les Foraminifères planctoniques actuels sont
généralement limités aux environnements marins ouverts, de
salinité océanique normale et d'eaux claires. Ils sont d'autant
plus diversifiés si ces eaux sont stratifiées. Ils sont abondants
dans la Zone Euphotique, bien que les exigences physiologiques et
écologiques s'expriment en une certaine distribution verticale, par les
espèces hermatypiques (c'est-à-dire vivant en symbiose avec les
algues planctoniques ou spinose) limitées aux rayons solaires
et aux eaux de surface, tandis que les espèces vivant
indépandamment de ces algues (nonspinose) tendent à
occuper les eaux de plus en plus profondes (Bé, 1982 ; Hemleben et
Spindler, 1983 ; Leckie, 1987). Cette distribution peut dépendre
partiellement de la densité des eaux marines et spécialement dans
les basses latitudes où la variation de la température et de la
salinité de la tranche d'eau supérieure peut favoriser une niche
hautement diversifiée. La diversité des associations des
Foraminifères planctoniques dans les environnements océaniques
ouverts de basses latitudes est probablement très influencée par
la disponibilité des nutriments et la fertilité de la surface de
l'eau (Lipps, 1979 ; Leckie, 1987). Les Foraminifères planctoniques sont
rares dans les environnements littoraux des plateaux continentaux et ils
augmentent en terme d'abondance avec l'augmentation de la profondeur (Murray,
1976 ; Rottman, 1978 ; Leckie, 1987). Ainsi, même dans les
dépôts anciens caractérisant les mers marginales, leur
augmentation a été vérifiée et souvent les auteurs
relient l'augmentation du rapport des Foraminifères
planctoniques/Foraminifères benthiques
81
(P/B) des sédiments et l'augmentation de la
diversité des Foraminifères planctoniques à
l'approfondissement des environnements marins (Murray, 1976). Ces augmentations
peuvent être en liaiason avec l'augmentation du niveau marin, de l'apport
nutritif ou autre.
Les modèles de distribution
paléogéographique et paléobiogéographique des
Foraminifères planctoniques sont utiles pour les reconstitutions
paléogéographiques et peuvent refléter de larges gradients
de températures des océans crétacés et des mers
épicontinentales. Les distributions paléobiogéographiques
sont aussi utiles pour la reconstitution des fluctuations du niveau marin
global. Entre autre, d'autres applications paléocéanographiques
comportent des interprétations des systèmes d'upwelling anciens
en se basant sur la composition des Foraminifères, la faune et la flore
associées et l'étude de faciès (Butt, 1982 ; Leckie,
1987).
La répartition en profondeur dans la colonne d'eau des
Foraminifères planctoniques du Crétacé est
considérée analogue à celles des Foraminifères
planctoniques actuelles (Leckie, 1987 ; Keller et al., 1995 ;
Zaghbib-Turki et al., 2000 ; Karoui Yaakoub et al., 2001 ;
Keller et Pardo, 2004). Les morphotypes à loges globuleuses auraient
habiter les eaux de surface, et les formes carénées
trochospiralées biconvexes et planoconvexes sont cantonnées aux
habitats de subsurface (Hart et Bailey, 1979). De telles interprétations
sont appuyées par les analyses statistiques et isotopiques de
l'oxygène O18 des Foraminifères des séries
crétacées (Eicher et Worstell, 1970 ; Leckie, 1987 ; Keller
et al., 2001 ; Keller et Pardo, 2004).
Eicher et Worstell (1970) ont constaté que les
morphotypes à loges globuleues telles que Hedbergella et
Heterohelix sont les premières à apparaître durant
les transgressions et les dernières à disparaître durant
les régressions marines du Crétacé. Ces données
paléoécologiques ont été adoptées par Leckie
(1987).
Ce modèle de répartition des
Foraminifères planctoniques et le type des niches écologiques
occupées par les différentes espèces peuvent être
déduits à travers la diversité et les fréquences
spécifiques, les morphologies des espèces et leur distribution
paléogéographique (Eicher et Worstell, 1970). Récemment,
pour mieux argumenter les
82
interprétations paléoécologiques, les
auteurs ajoutent des analyses isotopiques de l'Oxygène O18 et
du Carbone C13 (Shackleton et al., 1994).
2) Paléoécologie des Foraminifères
planctoniques de l'intervalle du passage Cénomanien - Turonien.
Selon Leckie (1987), la faune du Crétacé
(Gargasien - Cénomanien supérieur) peut être
subdivisée en trois catégories :
a) Faunes de mer épicontinentale (<100m)
Cette faune est caractérisée par les
espèces appartenant aux genres Heterohelix, à test
bisérié et les Guembelitria, à test
trisérié. Ces genres sont particulièrement les
diagnostiques des milieux à faible tranche d'eau des mers
épicontinentales du Crétacé. Elles sont peu
fréquentes dans les milieux marins ouverts. Il est admis que ces genres
habitaient les eaux de surface ou prés de la surface et pourraient avoir
eu de larges tolérances trophiques. Ils étaient capables de vivre
dans les milieux à condition écologiques très variables.
Ils sont qualifiés d'Eurytopiques. Ces genres englobent les
Foraminifères planctoniques opportunistes durant l'intervalle
Gargasien-Cénomanien supérieur. Par ailleurs ces
Hétérohélicacés sont considérés comme
des habitants de surface.
b) Faunes de subsurface habitant les mers
épicontinentales
Dans ces mêmes mers relativement peu profonde (<100m)
Hedbergella spp et Globigerinelloides spp seraient les
habitants de subsurface (deep-water taxa). Ces espèces sont
dominantes dans les mers ouvertes riches en faune pélagique.
Particulièrement Clavihedbergella notammnet
C. simplex à loges étirées habitaient les
tranches méso et bathypélagiques de ces mers ouvertes. Ces
Clavihedbergella vivaient dans des tranches d'eau plus épaisse
que Hedbergella delrioensis et Hedbergella planispira
à loges globuleuses
83
c) La faune de mer profonde
Les espèces carénées Rotalipora
spp. et Praeglobotruncana spp sont
supposées les habitants de subsurface dans les eaux photiques. Cette
interprétation est basée sur les données de
18O, la distribution actuelle de leurs analogues qui sont les
Globorotaliidae. Ces espèces sont aussi supposées comme
étant les plus sensibles aux changements envirenmentaux (stenotopique)
de tous les Foraminifères planctoniques durant le Cénomanien
(Leckie, 1987).
Ces différentes indications
paléoécologiques ne sont pas toutes conformes avec ce qu'ont
proposé d'autres auteurs. Ainsi, selon Lipps (1979) ; Leckie et
al., (1998) ; Keller et al., (2001) ; Price et Hart, (2002);
Keller (2002) ; Keller et Pardo, (2004), l'évolution, la diversification
et l'extinction des Foraminifères planctoniques sont
généralement liées à la stratification de la
colonne d'eau, les variations des apports trophiques, les gradients de
température et de densité. Ces auteurs indiquent que la
stratification de la masse d'eau et le type des niches écologiques
occupées par les différentes espèces peuvent être
déduits des fréquences spécifiques, des morphologies des
taxa et des données isotopiques. Notamment, Keller et Pardo (2004) ont
distingué les attributs suivants :
a) Les habitants de subsurface : les formes
carénées
Ces auteurs ont constaté que les Rotalipores occupaient
les eaux de subsurface les plus profondes (dans le sens bathypélagique
et oligotrophique) généralement peu au dessous de la thermocline.
Leur extinction est par conséquent due aux changements des conditions
écologiques. Ces Rotalipores sont très sensibles aux variations
des paramètres écologiques notamment à la stratification
des eaux.
84
|
CATÉGORIE
|
ESPÈCES
|
MORPHOTYPES
|
|
Habitants des eaux de subsurface zone photique
supérieur
|
Rotalipores
|
Formes crènées
|
|
Habitants des eaux
de subsurface
zone photique
inférieur
|
Praeglobotuncana
Dicarinella
Anatecinella
|
formes faiblement
carènées
|
|
Habitants de la
Zone à Oxygène Minimum
|
Heterohelix
|
Biseriés
|
|
Habitants des eaux
Salinité faible
|
Hedbergella planispira alternating with
Heterohelix, when Hetero - helicids increase the number of
Hedbergella decrease
|
Trochospiralés
avec une trochospire basse
et
biseriés
|
|
Habitants des eaux
de surface
Salinité normale
|
Clavihedbergella simplex
Hd. delrioensis, Hd.
planispira
Globigerinelloides ultramicra
Gl. bentonensis, Whiteinella
spp.
|
Trochospiralés
avec une trochospire basse
et des loges étirées
ou des formes
planispiralées
|
|
Habitants des eaux
de surface
eutrophiques
|
Guembelitria cenomana
Guembelitria albertensis
|
Trisériés trés petits
|
Tableau 3 - Les indicateurs
paléoécologiques
b) Les habitants de la zone photique inférieur : les
espèces faiblement carénées
Ce groupe est caractérisé par le
développement des espèces à grandes loges et à
carènes faiblement développées : Anaticinella
multiloculata, Praeglobotruncana spp, Dicarinella algeriana, D. hagni. Ces
espèces ont été interprétées comme
appartenant à la zone photique inférieure.
c) Les habitants de la zone à oxygène minimum :
les Hétérohélicidés Les
Hétérohélicidés, de l'intervalle Cénomanien
- Turonien, s'épanouissaient dans des environnements marins pauvres en
Oxygène.

Paleoecological parameters
Upper water column depth
Oxygene rate
Salinity
Trophic input
Subsurface
Hypoxic
environment
Subsaline surface
Normal saline surface
Upper photic zone
Lower photic zone
Eutrophic surface
Surface
O optimum
2
O2 minimum
Rotalipora montsalvensis
Rotalipora cushmani
Rotalipora greenhornensis
Heterohelix reussi
Heterohelix pulchra
Heterohelix moremani
Helvetoglobotrunacana helvetica
Marginotruncana sigali Marginotruncana renzi
Praeglobotruncana gibba
Praeglobotruncana stephani
Dicarinella algeriana
Dicarinella hagni
Anaticinella multiloculata
Globigerinelloides bentonensis
Gl. ultramicra
Hedbergella planispira Hedbergella delrioensis Hedbergella
simplex Guembelitria cenomana Guembelitria albertensis
Whiteinella archaeocretacea Whiteinella aprica
Whiteinella brittonensis Whiteinella paradubia
Whiteinella baltica
Whiteinella aumalensis
Whiteinella inornata
Whiteinella praehelvetica
85
Tableau 4 - tableau de répartition des
Foraminifères planctoniques dans les différentes niches
écologiques
d) 86
Les habitants de surface de faible salinité :
Hedbergella planispira
Les analyses isotopiques indiquent que toutes les
espèces du genre Hedbergella seraient des habitants de surface
ou prés de la surface. Parmi ces espèces, Hedbergella
planispira se développait en alternance avec les
Hétérohéliicidés quand la salinité des eaux
de surface diminue. Ceci est appuyé par l'augmentation des
éléments détritiques dans les sédiments.
e) Les habitants de surface à salinité normale
Il groupe les autres espèces de Hedbergella
(H. delrioensis, H. simplex), les espèces de
Globigerinelloides (G. bentonensis, G. ultramicra) et les
whiteinelles (W. aprica, W. brittonensis, W. archaeocretacea, W.
paradubia).
f) Les opportunistes, habitants des surfaces eutrophiques :
Guembelitria Les espèces de Guembelitria
s'épanouissaient au niveau des eaux de surface eutrophiques dans
des environnements marins marginaux à faible tranche d'eau. La
salinité pourrait être variable. Durant des périodes de
stress écologiques sévères où peu d'espèces
survivaient. Guembelitria cenomana et G. albertensis
deviennent abondantes. Ainsi elles sont considérées comme
des espèces opportunistes habitant les eaux de surface eutrophiques.
3) Les indicateurs paléoécologiques du
passage Cénomanien - Turonien de la coupe d'Oued Es Smara (CES)
Dans ce qui suit nous nous sommes basés sur les
résultats de l'analyse statistiques des différentes
espèces (Fig.25) et des morphotypes en adoptant les attributs
considérés par Leckie (1987) et Keller et Pardo (2004).
Nous cernons le comportement des Foraminifères
planctoniques en plusieurs étapes :
1) Assemblage de Foraminifères planctoniques
diversifiés dans une masse d'eau
stratifiée
Dans les dépôts d'Oued Es Smara (CES), les formes
carénées telles que les Rotalipores sont
représentées par Rotalipora cushmani, Rotalipora
montsalvensis, Rotalipora greenhornensis. Elles sont présentes
à la base de la série étudiée. Leur
fréquence est faible (elle est globalement inférieure à
10%). Même rares, les Rotalipores indiquent une mer ouverte, là
où la stratification de la masse d'eau est normale. De tel habitat a
été signalé ailleurs au Maroc (Leckie, 1984), en
Angleterre (Keller et al., 2001), en Amérique du nord (Keller
et Pardo ; Keller et al., 2004). Toutefois, la fréquence des
Hétérohélicidés, des Guembelitriidés et des
Whiteinelles reste peu élevée dans ces niveaux de la base de la
séries étudiée (respectivement 25%, inférieur
à 3% et 13%).
2) L'installation de la ZOM et prolifération des
Heterohelicacea
Peu avant leur extinction, les Rotalipores deviennent rares et
les fréquences des trois autres groupes ont augmenté. Celles des
Hétérohélicidés de toute petite taille atteint 75%,
celle des Guembelitria atteint 25% et celle des Whiteinelles atteint 20%. Nous
pensons que la rareté des Rotalipores et l'augmentation des
fréquences de ces trois groupes sont évocatrices de changement
des conditions paléoécologiques. D'ailleurs, ces
évènements biologiques coincident avec le changement des
faciès, tout au moins en apparence ; c'est-à-dire avec
l'apparition dans la série des niveaux de marnes et calcaires argileux
de couleur noirâtre, se débitant en feuillets et dégageant
une odeur de bitume à la cassure fraîche qui devraient être
riches en matière organique.
Ainsi, l'augmentation du taux à (70%) des petits
Heterohelix (H. moremani, H. reussi, H. pulchra, H. navarroensis)
qui sont considérés comme étant les habitants des
eaux marines à faible taux d'Oxygène dissous dénote de
l'installation de la zone à Oxygène minimum (ZOM).
En outre, nous constatons que dans ces niveaux, quand la
fréquence des Hétérohélicacés
bisériés du genre Heterohelix augmente celle des
Hétérohélicacés trisériés du genre
Guembelitria diminue et vis-versa.
87
Étant donné que ces derniers morphotypes sont
indicateurs d'habitats très faiblement
88
Age
|
Formation
|
Zone
|
Sous-zone
|
Épaisseur
|
Echantillon
|
Lithologie
|
|
R. m ntsalvensis P.delrioensis
R. cushmani
R. greenhornensis P. stephani
W.paradubia
Gl.bentonensis
D. hagni-imbricata P. gibba
D.algeriana
Gl.ultramicra G.cenomana
Hd.delrioensis Hd.planispira
Clav.simpl ex H.reussi
H.moremani
H.navarroensis H.aff.pulchra
W.aprica
W.baltica
W.archaeocretacea
D.imbricata G.albertensis
W.praehelvetica
D.hagni
Helv.helvetica
M.renzi M.sigali
Benthonic
Foraminifera
|
|
C. moy Cénomanien supérieur Tur. inf.
|
Kef
|
H. helvetica
|
22
20
18
14 12 10 8 6 4 2
|
|
65
|
|
|
|
|
|
|
|
|
|
|
0 (%) 50 (%) 100 (%)
|
|
|
|
|
|
|
|
|
Fahdène Bahloul
|
|
|
U6
|
|
60
|
|
|
Whiteinella archaeocretacea
|
G. bentonensis D. hagni H. moremani
|
|
|
|
|
|
|
|
|
|
|
55
|
|
|
|
|
|
|
|
|
|
50
|
|
|
|
|
|
|
|
|
|
|
U5
|
|
45
|
|
|
|
|
|
|
40
|
|
|
|
|
|
|
35
|
|
|
|
|
|
|
30
|
|
|
|
|
U4
|
|
|
|
|
25
|
|
|
|
|
|
|
20
|
|
|
|
|
|
|
|
|
|
15
10
|
U3
|
|
|
|
5
4
3
2
1
|
U2
|
|
R. cushmani
|
R. montsalvensis D. algeriana
|
|
U1
1m
|
Fig. 27 - Abondances relatives des espèces de
Foraminifères planctoniques dans la fraction de taille >63um de la
Formation Bahloul de la coupe CES. A noter que
l'association est
dominée par les Heterohelicidés tolérant l'oxygène
minimum et les Guembelitriidés marqueurs des eaux de surface à
caractère eutrophique et à salinité
relativement
normale, ceci est due à une ZOM bien développ
89
oxygénés voire hypoxiques, cette alternance des
morphotypes bisériés et trisériés serait en
relation avec la diminution et l'augmentation alternées du taux d'O2
dissous dans l'eau.
3) Expansion de la ZOM, hypoxie - anoxie et
multiplication des opportunistes
Après l'extinction des Rotalipores, la fréquence
des Whiteinelles, des Hedbergelles, des Guembelitria et des
Heterohelix ont considérablement augmenté
(respectivement 22%, 12%, 25%, 75%). Cette disparition des Rotalipores coincide
entre autre avec la diversification des Whiteinelles (W. archaeocretacea,
W. aprica, W. praehelvetica). De ce fait, nous pensons que le changement
total du faciés, c'est-à-dire l'apparition des niveaux à
black shales qui alternent avec des marno-calcaires de couleur noirâtre
supposés riches en matière organique, et
l'élévation des fréquences des groupes cités sont
évocatrices de l'expansion de la zone à Oxygène minimum
(ZOM) préalablement installée. Cette expansion a favorisé
l'épanouissement plutôt des Heterohelicidae de toute petite taille
et à test bisérié en alternance avec les Guembelitriidae
à test trisérié. L'alternance de ces deux morphotypes
renseigne encore sur la diminution et l'augmentation alternées du taux
d'oxygène dissous dans l'eau et aussi sur l'apport trophique. Nous
savons que les Guembelitria prolifèrent dans les environnements
eutrophiques. Il a été verifié par ailleurs que leur pic
d'abondance coincide avec les valeurs maximales de äC13
indiquant un taux de productivité primaire élevé.
La disparition de Globigerinelloides bentonensis
coïncide bien avec une chutte brusque des fréquences de
Whiteinelles, d'Hedbergelles et de Dicarinelles (ces dernières sont
respectivement de 4%, 2% et inférieur à 1%) et l'augmentation des
fréquences des Guembelitria et de Heterohelix
(respectivement 40% et 70%). Cette diminution dans la fréquence des
espèces supposées comme habitants de surface (Whiteinlles et
Hedbergelles à loges globuleuses) et de subsurface de la zone photique
inférieure (Dicarinelles à test faiblement caréné)
qui coincide avec l'élévation des fréquences des
Hétérohélicacés et la diminution voire l'absence
des Foraminifères benthiques, nous renseigne sur l'expansion de la ZOM
dans la couche superfivielle des eaux océaniques ainsi que le fond marin
au moins à l'interface eau-sédiment. Le retour des
Foraminifères
90
CA
|
113
|
Formation
|
Zones
|
in
m
N N
|
[Thickness
·
|
ID
É
|
a
01
ô
J
|
c
|
Bioevent
|
CES section faunal turnover CaCO3rate
|
|
Mid. Cenomanian Upper Cenomanian ILowerTuronian
|
Fanden Bahloul kef
|
V v
4 v
tl
6 ÿ
c
· ~ _ --
r7. ne7 Verfco
|
ÿ40
G. bentonensis I fJirarinelia hagni Jhlererahelix
moremanl
|
" 65
|
111
·1
11
~ ü6
.
|
M. renzi
N. helvetira
s
P. gibba
Ichnofossils level
Ammonites level
et Berrthonic Foraminifera
return
'tl Benthonic Foraminifer.;
` i temperetydisappea Lai i.c
GI. benronensis
D. hagnf
Z A. multiloculara
W.praehelvetira
$
R. R,
cus~omermoninsis
* green
J W. archaearretareo
1.
P. ,,~ delrioerrsis
R. montsalvensis
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1
55
50
--.
--
|
|
|
|
|
|
F,..
|
|
pt °$
|
-.-
|
|
CI"~35EMI
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
30
|
-
|
|
|
|
|
|
Pa
|
--L
|
4
|
|
|
|
|
|
|
|
25
|
|
|
|
|
|
|
IDI
|
--
|
|
|
a an
|
|
|
|
|
|
|
:
|
|
|
jts
|
|
|
L73
|
|
to
|
|
|
|
Ratalipora [ushmani
|
R. montsalvensis R
algeriana
|
|
6
|
--
|
|
s
a
3
·
|
^rt
2
ui
1
·11
|
Fig.28 -- le renouvellement faunique mis en
évidence à Oued Es Smara
91
benthiques coincide entre autre avec l'élévation
des fréquences des groupes cités anterieurement (respectivement
13%, 9%,5%). La présence des Ammonites dans les niveaux sus-jascents
indiquerait un milieu oxygèné et favorable à la vie
planctonique (augmentation du taux d'oxygène dans la masse d'eau). En
plus les niveaux à Ichnofossiles (terriers) indiquent aussi une
élévation du taux d'oxygène dans l'interface
eau-sédiment. Dans ces niveaux à Ammonites et à
bioturbation correspendant à des calcaires feuilletés ou à
aspect massif de couleur moins foncé, bien que l'association des
Foraminifères planctoniques reste dominée par les
Guembelitria et Heterohelix, la présence des
espèces Globigerinelloides ultramicra, Praeglobotruncana
gibba, Dicarinella imbricata, D. hagni, des Whiteinelles, des Hedbergelles
qui est sporadique nous laissent supposer que l'impact de la 7OM s'est
atténué.
4) Retour aux conditions normales : renouvellement
faunique
Dés l'apparition de Helvetoglobotruncana
helvetica, on assiste à un changement de faciès traduit par
l'apparition de dépôts marneux de couleur beige. Ce changement de
faciès coincide avec une chutte dans la fréquence des
Guembelitria (2%) et de Heterohelix (40%) et
l'épanouissement de nouveux genres tels que Marginotruncana et
en particulier M. renzi (3%)
En récapitulation, on dénote l'existence de quatre
phases :
Phase 1 - les masses d'eau devraient être normalement
stratifiées au moment où les Rotalipores étaient
présentes et diversifiées (sous la thermocline).
Phase 2 - Avant l'extinction des Rotalipores, on assiste
à l'installation d'une 7OM décelée par l'augmentation de
la fréquence des petits Hétérohélicidés.
Phase 3 - Après l'extinction des Rotalipores, la 7OM
devient alors plus répandue, attesté par l'abondance
Heterohelix et Guembelitria.
Phase 4 -L'apparition de H. helvetica et des
Marginotruncana, formes carénées supposées
habiter en subsurface (sous la thermocline) quand la masse d'eau est de nouveau
stratifiée , laisse supposer l' atténuation de l'impact de la
7OM.
92
Age
|
Formation
|
Zone
|
Sous-zone
|
paisseur
|
chantillon Lithologie
|
Unité
|
Rotalipora
Globogerinelloides Praeglobotruncana
Whiteinella & Hedbergella
Whiteinella Hedbergella
Dicarinella
Guembelitria
Hetrohelix
GH
u e
cao
lix
ria &
C
O
T
C
a
C
O
3
|
|
Tur. inf
|
Fahdène Bahloul Kef
|
Rotalipora cushmani Whiteinella archaeocretacea H.
helvetica
|
22 20 18 16 14 12 0 8 6 4 2
|
65
U6
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
60
|
|
|
|
Cénomanien supérieur
|
|
|
55
|
|
|
|
50
|
|
|
|
U5
|
|
45
|
|
|
40
|
|
|
|
|
|
|
|
|
|
|
35
|
|
|
|
30
U4
|
|
|
25
|
|
|
20
|
|
|
|
15
U3
10
|
|
|
|
|
|
|
|
|
|
5
4
3 U2
2
1
|
|
Cen. moy
|
|
0 1 20 40 80
(%) (%)
0 50 100
(%)
|
|
U1
1m
|
Fig. 29 - Abondances relatives des Genres de Foraminifères
planctoniques dans la fraction de taille >63um de la Formation Bahloul de la
coupe CES. A noter le comportement antagoniste des Heterohelicidés et
des Guembelitriid

continent
Surface Subsurface
mer
Zone photique inférieure
93
Fig.30 a- première phase : masse d'eau
stratifiée

continent
Surface Subsurface
mer
Zone photique inférieure
Fig.30 b- deuxième phase : installation de la ZOM

continent
Surface Subsurface
mer
Zone photique inférieure
Fig.30 c- troisième phase : expansion de la ZOM

continent
Surface Subsurface
mer
Zone photique inférieure
94
Fig.30 d- quatrième phase : renouvellement
faunique
Fig.30 - Les quatre phases
B - Paléoécologie des Radiolaires du passage
Cénomanien - Turonien.
Les changements, évoquées dans l'intervalle
Cénomanien - Turonien, concernant les associations des
Foraminifères planctoniques et benthiques ont été mis en
évidence par plusieurs auteurs (Arthur et Premoli Silva, 1982 ;
Robaszynski et Caron, 1995 ; Premoli Silva et Sliter, 1995 ; ect...), dans
plusieurs localités et en particulier dans le domaine téthysien.
Ces auteurs admettent que ces changements sont survenus suite à des
conditions eutrophiques qui ont atteint probablement leur apogée
à la limite Cénomanien - Turonien (Zone à Whiteinella
archaeocretacea) et ont continué lors des dépôts des
sédiments du Turonien inférieur jusqu'à peu avant ou peu
après l'apparition de Helvetoglobotruncana helvetica. Les
associations des Radiolaires ont aussi subi un renouvellement faunique majeur,
qui serait peut être l'expression de l'impact de
l'évènement EAO-2 sur ce groupe, comme cela a été
appuyé par plusieurs auteurs (Kuhnt et al., 1986 ; Thurrow et Kuhnt,
1986 ; Erbacher, 1994 ; Lambert et De Wever, 1996). selon O'Dorghety (1994) ce
changement ne serait manifesté qu'à la fin de l'EAO-2. Salvini et
Marcucci Passerini (1998) ont constaté que les changements dans les
associations des Radiolaires ont eu lieu à l'apogée de
l'épisode de Haute productivité. La répartition verticale
des assemblages montre que le changement était soudain et que les
conditions eutrophiques ont favorisé le développement des
Radiolaires, différemment des Foraminifères planctoniques. Selon
Coccioni et al., (1992), les Radiolaires sont plus tolérants
aux conditions eutrophiques.
95
|
Age
|
C moyen
|
|
|
|
|
|
Cénomanien supérieur
|
|
|
|
|
|
|
|
|
|
Zone
|
Rotalipra cushmani
|
|
|
|
|
|
|
Whiteinella archaeocretacea
|
|
|
|
|
|
|
|
|
|
Sous-Zone
|
Dicarinella algeriana
|
|
|
|
|
|
|
Globigerinelloides bentonensis
|
|
|
|
Dicarinella hagni
|
|
|
|
|
|
Numéro d'échantillon
|
1 2 3 4 5
|
6 7
|
8
|
9
|
10
|
11
|
12 13 14 15 16 17 18 19 20 21 22
|
23
|
24
|
25
|
26 27 28 29
|
30
|
31
|
32
|
33
|
|
Profondeur (cm)
|
0 105 220 475 500
|
520 540
|
550
|
575
|
605
|
635
|
665 695 725 755 785 805 835 855 950 980 1010
|
1040
|
1070
|
1100
|
1130 1190 1250 1280
|
##
|
1340
|
1370
|
1395
|
|
Dicarinella algeriana
|
5
|
6
|
2
|
6
|
8
|
3
|
1
|
3
|
3
|
1
|
4
|
1
|
2
|
1
|
1
|
0
|
1
|
3
|
1
|
0
|
1
|
0
|
0
|
0
|
0
|
x
|
x
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Dicarinella hagni
|
3
|
2
|
2
|
1
|
2
|
5
|
3
|
|
1
|
3
|
1
|
1
|
0
|
1
|
0
|
|
|
1
|
1
|
3
|
3
|
3
|
x
|
x
|
x
|
x
|
x
|
x
|
0
|
0
|
0
|
0
|
1
|
|
Dicarinella imbricata
|
|
|
|
|
|
0
|
1
|
0
|
0
|
0
|
1
|
0
|
x
|
x
|
x
|
x
|
0
|
x
|
x
|
x
|
x
|
x
|
x
|
x
|
x
|
x
|
x
|
x
|
x
|
x
|
0
|
0
|
0
|
|
Globigerinelloides bentonensis
|
12
|
11
|
12
|
6
|
3
|
0
|
0
|
1
|
|
|
2
|
|
|
x
|
1
|
|
|
|
|
|
|
|
1
|
|
|
x
|
|
|
|
|
|
|
|
|
Globigerinelloides ultramicra
|
2
|
3
|
2
|
3
|
|
0
|
0
|
0
|
|
|
1
|
0
|
0
|
x
|
0
|
|
|
0
|
|
0
|
|
|
|
|
|
x
|
|
|
|
|
|
|
|
|
Guembelitria cenomana
|
1
|
1
|
6
|
30
|
23
|
12
|
4
|
2
|
13
|
16
|
9
|
10
|
17
|
5
|
15
|
3
|
10
|
25
|
18
|
21
|
17
|
16
|
6
|
7
|
8
|
34
|
7
|
17
|
15
|
22
|
14
|
15
|
17
|
|
Guembelitria albertensis
|
x
|
x
|
x
|
x
|
x
|
2
|
4
|
1
|
3
|
7
|
5
|
2
|
9
|
2
|
3
|
1
|
3
|
4
|
5
|
3
|
1
|
3
|
1
|
2
|
3
|
6
|
5
|
8
|
2
|
6
|
1
|
4
|
5
|
|
Hedbergella delrioensis
|
0
|
0
|
1
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0
|
2
|
1
|
4
|
0
|
5
|
3
|
3
|
3
|
0
|
3
|
5
|
1
|
|
2
|
|
|
0
|
1
|
|
|
2
|
|
Hedbergella planispira
|
1
|
5
|
5
|
14
|
1
|
4
|
2
|
1
|
2
|
2
|
3
|
2
|
3
|
2
|
1
|
1
|
5
|
3
|
1
|
2
|
3
|
3
|
2
|
2
|
1
|
x
|
x
|
x
|
0
|
1
|
1
|
0
|
1
|
|
Clavihedbergella simplex
|
10
|
2
|
3
|
10
|
5
|
|
3
|
7
|
3
|
10
|
1
|
9
|
4
|
2
|
9
|
|
2
|
1
|
3
|
2
|
2
|
1
|
0
|
0
|
x
|
x
|
x
|
x
|
x
|
|
|
|
2
|
|
Helvetoglobotruncana helvetica Heterohelix globulosa
|
3
|
5
|
2
|
2
|
15
|
10
|
4
|
12
|
8
|
8
|
18
|
22
|
17
|
30
|
15
|
21
|
18
|
16
|
16
|
15
|
13
|
18
|
25
|
27
|
22
|
23
|
31
|
19
|
27
|
22
|
33
|
30
|
22
|
|
Heterohelix moremani
|
11
|
12
|
26
|
14
|
27
|
20
|
36
|
51
|
54
|
22
|
28
|
21
|
19
|
39
|
18
|
46
|
24
|
30
|
30
|
31
|
40
|
34
|
28
|
37
|
44
|
34
|
27
|
33
|
37
|
37
|
36
|
40
|
31
|
|
Heterohelix navarroensis
|
4
|
2
|
2
|
4
|
4
|
6
|
6
|
4
|
6
|
8
|
4
|
8
|
3
|
5
|
3
|
3
|
4
|
4
|
4
|
1
|
5
|
6
|
8
|
2
|
3
|
x
|
|
3
|
4
|
|
3
|
1
|
3
|
|
Heterohelix aff. pulchra
|
0
|
0
|
|
|
3
|
0
|
|
0
|
|
0
|
|
|
|
|
|
0
|
|
|
|
3
|
8
|
1
|
19
|
17
|
13
|
x
|
24
|
19
|
13
|
6
|
9
|
10
|
4
|
|
Marginotruncana renzi
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Margininotruncana sp
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Praeglobotruncana delrioensis
|
1
|
|
2
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Praeglobotruncana gibba
|
3
|
6
|
2
|
1
|
1
|
5
|
1
|
0
|
0
|
0
|
3
|
1
|
2
|
1
|
1
|
0
|
0
|
1
|
0
|
0
|
0
|
0
|
x
|
x
|
x
|
x
|
|
|
|
|
|
|
|
|
Praeglobotruncana stephani
|
1
|
0
|
0
|
1
|
1
|
0
|
0
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rotalipora cushmani
|
3
|
4
|
3
|
1
|
2
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rotalipora greenhornensis
|
4
|
2
|
2
|
0
|
2
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rotalipora montsalvensis
|
1
|
0
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Whiteinella aprica
|
3
|
9
|
13
|
11
|
3
|
5
|
7
|
6
|
1
|
5
|
3
|
3
|
2
|
2
|
3
|
1
|
2
|
2
|
3
|
2
|
|
1
|
0
|
0
|
0
|
x
|
|
|
0
|
1
|
1
|
|
1
|
|
Whiteinella archaeocretacea
|
|
|
|
1
|
2
|
2
|
3
|
4
|
2
|
6
|
6
|
4
|
8
|
3
|
7
|
7
|
5
|
2
|
5
|
3
|
3
|
2
|
0
|
|
x
|
x
|
|
|
|
|
|
|
2
|
|
Whiteinella aff. archaeocretacea
|
0
|
0
|
1
|
1
|
4
|
0
|
0
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Whiteinella baltica
|
2
|
4
|
5
|
5
|
5
|
3
|
2
|
2
|
3
|
5
|
3
|
4
|
1
|
2
|
2
|
6
|
5
|
2
|
1
|
x
|
|
2
|
|
|
|
x
|
|
|
|
|
|
|
0
|
|
Whiteinella brittonensis
|
0
|
0
|
|
|
|
|
0
|
1
|
0
|
3
|
1
|
1
|
0
|
0
|
|
|
0
|
|
|
x
|
|
|
|
|
|
x
|
|
|
|
|
|
|
|
|
Whiteinella paradubia
|
0
|
|
|
|
|
|
|
|
|
|
1
|
0
|
1
|
x
|
|
|
|
|
|
0
|
|
|
|
|
|
x
|
|
|
|
|
|
|
|
|
Whiteinella praehelvetica
|
|
|
|
|
|
|
|
|
|
|
3
|
6
|
9
|
4
|
13
|
1
|
12
|
|
7
|
8
|
3
|
3
|
|
1
|
|
x
|
1
|
|
1
|
4
|
2
|
|
3
|
|
Benthiques
|
28
|
26
|
17
|
23
|
14
|
6
|
4
|
8
|
3
|
5
|
6
|
6
|
3
|
3
|
4
|
10
|
5
|
6
|
3
|
2
|
1
|
3
|
5
|
4
|
6
|
1
|
4
|
x
|
x
|
x
|
x
|
x
|
6
|
|
Nombre total
|
452
|
431
|
681
|
374
|
111
|
424
|
1319
|
414
|
477
|
615
|
305
|
300
|
491
|
1661
|
478
|
398
|
394
|
386
|
496
|
445
|
284
|
371
|
739
|
568
|
668
|
68
|
165
|
487
|
279
|
209
|
535
|
312
|
281
|
Tableau-5. Fréquences relatives en pourcentage des
Foraminifères planctoniques dans la coupe CES (première
partie)
96
|
Age
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Turonien
inférieur
|
|
|
|
Zone
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Helvetoglobotruncana helvetica
|
|
|
Sous-Zone
|
Dicarinella hagni
|
|
|
|
|
|
|
|
|
|
|
Heterohelix moremani
|
|
|
|
|
|
|
|
|
|
Numéro d'échantillon
|
34 35 36 37
|
38
|
39
|
40
|
41
|
42
|
43
|
44
|
45
|
46
|
47
|
48 49 50 51 52
|
53
|
54
|
55
|
56
|
57
|
58
|
59
|
60 61
|
62 63 64 65
|
66
|
|
Profondeur (cm)
|
1425 1455 1485 1545
|
1575
|
1605
|
1635
|
1665
|
1695
|
1725
|
1755
|
1785
|
1805
|
1835
|
1865 1895 1925 1955 1985
|
2015
|
2045
|
2075
|
2105
|
2135
|
2165
|
2195
|
2225 2270
|
2320 2380 2425 2455
|
2495
|
|
Dicarinella algeriana
|
1
|
4
|
2
|
1
|
0
|
1
|
1
|
1
|
|
0
|
0
|
2
|
1
|
|
|
0
|
0
|
0
|
1
|
0
|
|
2
|
|
1
|
1
|
|
0
|
1
|
0
|
|
|
0
|
0
|
|
Dicarinella hagni
|
1
|
1
|
1
|
1
|
0
|
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Dicarinella imbricata
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0
|
0
|
|
1
|
|
|
|
|
|
|
|
|
0
|
0
|
|
Globigerinelloides bentonensis
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Globigerinelloides ultramicra
|
|
|
|
|
|
|
|
0
|
|
|
0
|
|
|
|
|
|
0
|
|
|
|
|
0
|
|
|
0
|
|
|
|
|
|
1
|
|
0
|
|
Guembelitria cenomana
|
26
|
31
|
28
|
29
|
38
|
42
|
44
|
39
|
32
|
30
|
26
|
34
|
17
|
35
|
42
|
46
|
32
|
40
|
31
|
38
|
30
|
25
|
23
|
25
|
32
|
32
|
43
|
16
|
|
|
10
|
29
|
24
|
|
Guembelitria albertensis
|
6
|
9
|
13
|
6
|
6
|
8
|
4
|
7
|
5
|
4
|
18
|
11
|
5
|
6
|
7
|
12
|
11
|
13
|
16
|
9
|
11
|
8
|
12
|
3
|
6
|
2
|
6
|
3
|
|
|
1
|
2
|
2
|
|
Hedbergella delrioensis
|
1
|
2
|
1
|
1
|
0
|
1
|
|
0
|
|
0
|
1
|
|
1
|
|
|
0
|
|
0
|
0
|
|
0
|
|
1
|
|
|
0
|
|
1
|
0
|
|
1
|
|
0
|
|
Hedbergella planispira
|
0
|
1
|
0
|
1
|
1
|
|
|
|
|
1
|
|
1
|
|
|
1
|
3
|
2
|
|
|
1
|
3
|
1
|
1
|
2
|
1
|
2
|
1
|
1
|
2
|
1
|
1
|
1
|
|
|
Clavihedbergella simplex
|
|
1
|
0
|
0
|
1
|
|
0
|
0
|
1
|
|
|
0
|
|
0
|
|
|
0
|
|
1
|
|
0
|
|
1
|
|
|
|
|
|
0
|
0
|
1
|
|
|
|
Helvetoglobotruncana helvetica
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2
|
3
|
2
|
1
|
|
|
|
Heterohelix globulosa
|
14
|
12
|
15
|
12
|
11
|
12
|
11
|
9
|
6
|
14
|
9
|
8
|
12
|
18
|
6
|
6
|
9
|
8
|
9
|
6
|
8
|
5
|
7
|
10
|
15
|
17
|
11
|
15
|
20
|
27
|
18
|
11
|
9
|
|
Heterohelix moremani
|
31
|
22
|
22
|
33
|
29
|
23
|
21
|
34
|
49
|
36
|
33
|
38
|
50
|
31
|
36
|
20
|
26
|
25
|
22
|
19
|
21
|
14
|
23
|
25
|
26
|
23
|
11
|
19
|
22
|
33
|
34
|
16
|
24
|
|
Heterohelix navarroensis
|
2
|
5
|
5
|
4
|
9
|
6
|
5
|
0
|
2
|
5
|
2
|
0
|
3
|
3
|
2
|
4
|
9
|
2
|
0
|
2
|
2
|
0
|
1
|
3
|
3
|
3
|
3
|
2
|
4
|
2
|
7
|
2
|
2
|
|
Heterohelix aff. pulchra
|
10
|
5
|
3
|
4
|
1
|
4
|
5
|
3
|
1
|
5
|
0
|
0
|
3
|
|
|
0
|
1
|
|
2
|
4
|
2
|
4
|
1
|
0
|
|
0
|
6
|
14
|
12
|
17
|
|
3
|
4
|
|
Marginotruncana renzi
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2
|
1
|
1
|
2
|
|
|
Margininotruncana sp
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1
|
1
|
2
|
1
|
|
|
Praeglobotruncana delrioensis
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Praeglobotruncana gibba
|
1
|
|
1
|
0
|
|
|
0
|
1
|
0
|
0
|
1
|
|
|
|
|
|
0
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Praeglobotruncana stephani
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rotalipora cushmani
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rotalipora greenhornensis
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Rotalipora montsalvensis
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Whiteinella aprica
|
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
1
|
0
|
1
|
|
|
|
|
|
|
|
|
|
|
|
0
|
|
|
0
|
|
|
1
|
|
1
|
|
0
|
|
Whiteinella archaeocretacea
|
1
|
2
|
2
|
2
|
0
|
1
|
3
|
3
|
2
|
1
|
3
|
2
|
4
|
1
|
3
|
2
|
1
|
1
|
0
|
|
0
|
|
1
|
0
|
0
|
1
|
|
|
1
|
|
3
|
|
0
|
|
Whiteinella aff. archaeocretacea
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Whiteinella baltica
|
|
|
0
|
|
|
|
|
|
0
|
|
0
|
|
|
|
|
|
|
0
|
|
|
|
|
0
|
|
|
|
|
|
0
|
|
2
|
0
|
0
|
|
Whiteinella brittonensis
|
|
|
|
|
|
|
|
|
0
|
|
|
|
|
|
|
|
|
0
|
|
|
|
|
0
|
|
|
0
|
|
|
|
|
|
|
|
|
Whiteinella paradubia
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Whiteinella praehelvetica
|
2
|
4
|
4
|
5
|
2
|
2
|
5
|
2
|
1
|
4
|
6
|
3
|
2
|
4
|
2
|
5
|
6
|
2
|
1
|
2
|
1
|
1
|
3
|
2
|
1
|
1
|
|
1
|
4
|
|
6
|
3
|
1
|
|
Benthiques
|
5
|
5
|
4
|
2
|
1
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
2
|
1
|
2
|
1
|
3
|
8
|
16
|
18
|
22
|
40
|
25
|
29
|
15
|
19
|
18
|
27
|
28
|
16
|
10
|
30
|
32
|
|
Nombre total
|
191
|
226
|
328
|
325
|
362
|
429
|
452
|
445
|
377
|
332
|
434
|
409
|
374
|
416
|
394
|
411
|
228
|
393
|
509
|
520
|
570
|
248
|
546
|
384
|
620
|
480
|
404
|
527
|
629
|
621
|
407
|
656
|
600
|
Tableau-5. Fréquences relatives en pourcentage des
Foraminifères planctoniques dans la coupe CES (deuxième
partie)

97
Fig. 31 - comparaison entre la fréquence relative du
genre Heterohelix

98
Fig. 32 - comparaison entre la fréquence relative de
Guembelitria cenomana
99
Premoli Silva et al., (1999) ont indiqué que
l'association des Radiolaire est dominée par les Nassellaires (Ordre des
Nassellaria) et précisément les genres Archaeodictyomitra,
Dictyomitra, Cryptamphorella, Holocryptocanium, Pseudodictyomitra, Stichomitra
et Thanarla. A partir de l'état de conservation de ces
Nassellaires, ces auteurs suggèrent que ces derniers seraient peut
être les analogues des habitants de subsurface actuels au cours du
Crétacé. Ces habitants de subsurface s'épanouissaient soit
au cours des périodes où le flux de matière organique est
supposé élevé soit au cours des périodes de haute
productivité, ce qui a laissé penser que ce groupe sont des
indicateurs des conditions eutrophiques et des taux élevés de
matière organique (Salvini et Marcucci Passerini, 1998 ; Primoli Silva
et al., 1999).
Dans notre matériel d'étude nous constatons
l'abondance de Dictyomitra spp et Pseudodictyomitra spp qui
sont des Nassellaires dans les niveaux du Cénomanien supérieur de
la zone à Rotalipora cushmani et des Sphaecularia dans les
niveaux de la zone à W. archaeocretacea riches en
matière organique.
Ce renouvellement faunique que nous mettons en évidence
serait étudié dans le détail ultérieurement.
100
CINQUIÈME PARTIE
SYSTÉMATIQUE DES FORAMINIFÈRES
PLANCTONIQUES DU PASSAGE CÉNOMANIEN -
TURONIEN
101
Systématique des Foraminifères
Dans le présent chapitre nous décrivons 33
espèces réparties en 12 genres et 6 familles. La classification
de ces Foraminifères adoptée est celle de LOEBLICH et TAPPAN
(1988).
ORDRE : FORAMINIFERIDA
EICHWALD, 1830
SOUS-ORDRE : GLOBIGERININA
DELAGE & HEROUARD, 1896
SUPERFAMILLE : HETEROHELICACEA
CUSHMAN, 1927
FAMILLE : HETEROHELICIDAE
CUSHMAN, 1927 SOUS-FAMILLE : HETEROHELICINAE
CUSHMAN, 1927
GENRE : Heterohelix
EHRENBERG, 1843
Espèce - Type : Textularia (=Heterohelix)
americana EHRENBERG, 1843
Synonymie du genre :
1843 - Heterohelix EHRENBERG, p. 429
1844 - Spiroplecta EHRENBERG, p. 75
1899 - Guembelina EGGER, p. 31
1985 - Heterohelix EHRENBERG, Caron, p. 23
1988 - Pseudoplanoglobulina LOEBLICH et TAPPAN, p.
455
1990 - Heterohelix EHRENBERG, Nederbragt, p. 89
2000 - Heterohelix EHRENBERG, Georgescu, p. 162
Diagnose du genre :
Le genre Heterohelix est caractérisé par
un test bisérié, présentant (ou pas) un enroulement
planispiral initial. Il est finement à grossièrement
perforé et/ou orné par des côtes.
Espèce : Heterohelix moremani (CUSHMAN,
1938)
Pl.3, fig.5-6
Synonymie:
1938 - Guembelina moremani CUSHMAN, pl. 2, fig. 1-3, p.
10
1940 - Guembelina washitensis TAPPAN, pl.19, fig.1, p.
115
1984 - Heterohelix moremani (CUSHMAN), Leckie, pl. 10,
fig. 1-2, p. 593
1985 - Heterohelix moremani (CUSHMAN), Caron, fig. 24,
6-7
1990 - Heterohelix moremani (CUSHMAN), Nederbragt, pl.2,
fig. 6a-7; pl.3, fig.1a-2b, p88, 93
1991 - Heterohelix moremani (CUSHMAN), Matmati et al,
pl.8, fig.2, p. 62
1995 - Heterohelix moremani (CUSHMAN), Georgescu, fig.
3-5, 13, p. 403
1998 - Heterohelix moremani (CUSHMAN), Rami, pl. 4, fig.
9, p. 144
2000 - Heterohelix moremani (CUSHMAN), Georgescu, p.
162
2001 - Heterohelix moremani (CUSHMAN), Keller et al.,
fig.8, 5, p. 404
2001 - Heterohelix moremani (CUSHMAN), El Amri, pl. VI,
fig. 10
2004 - Heterohelix moremani (CUSHMAN), Keller et Pardo.
Pl.6, fig.15-17
Caractères généraux:
C'est une espèce qui présente un arrangement
bisérié (de 6 à 9 paires de loges). Les loges augmentent
de taille progressivement donnant un aspect mince au test. Les loges sont
globuleuses et finement perforées. L'ouverture est en forme d'arc
bordée par une lèvre.
Répartition stratigraphique :
H. moremani est connue de l'Albien supérieur
jusqu'au Santonien.
Dans notre matériel :
H. moremani est l'espèce la plus
fréquente de l'association. Son pic d'abondance (acmé) nous a
permis d'établir une sous-zone. Sa fréquence maximale (55%)
coincïde avec l'échantillon CES-42.
Espèce : Heterohelix navarroensis
LOEBLICH, 1951
Pl.3, fig.3-4
Synonymie:
1844 - Spiroplecta americana EHRENBERG, p. 75
1951 - Heterohelix navarroensis LOEBLICH, pl. 12, fig.
3
1985 - Heterohelix navarroensis LOEBLICH, Caron, fig.
24, 8-9 1990 - Heterohelix navarroensis LOEBLICH, Nederbragt, pl.3,
fig. 5a-b 1991 - Heterohelix navarroensis LOEBLICH, Matmati et al,
pl.8, fig.1, p. 62 1998 - Heterohelix navarroensis LOEBLICH, Rami, pl.
5, fig. 1-2, p. 144 2001 - Heterohelix navarroensis LOEBLICH, El Amri,
pl. VI, fig. 5
Caractères généraux:
C'est une espèce bisériée. Cette
espèce montre généralement un stade initial planispiral
mais à croissance continue. Le test est de petite taille, avec un apex
non pointu, il présente une surface finement perforée et finement
costulée. L'ouverture est bordée par un bourrelet. Cette
espèce se rapproche de H. globulosa, qui peut aussi être
planispirale à sa phase initiale, toutefois elle en diffère par
les dernières paires de loge qui sont nettement moins
dilatées.
102
Répartition stratigraphique :
103
H. navarroensis est connue depuis le Cénomanien
supérieur jusqu'au Maastrichtien.
Espèce : Heterohelix pulchra (BROTZEN, 1936)
Pl.3,
fig.7-8
Synonymie :
1936 - Guembelina pulchra BROTZEN, p.121, pl.9, fig.
3a-b
1983 - Heterohelix pulchra (BROTZEN), Weiss, p.46-47,
pl.2, fig.9-13
1990 - Laeviheterohelix pulchra (BROTZEN), Nederbragt,
p.101,pl.5, fig. 7a-b, pl.6 fig.1
1995 - Heterohelix pulchra (BROTZEN), Keller et al.,
pl.3 fig.3
1998 - Heterohelix pulchra (BROTZEN), Rami, p.145, pl.5,
fig.3-4
2001 - Heterohelix pulchra (BROTZEN), El Amri, p.66,
pl.6, fig.9
Caractères généraux :
Le test est H. pulchra est lisse à profile
aplati. Cette espèce est caractérisée par des loges en
croissant qui se recouvrent de telle sorte que la stuture médiane
déprimée acquière une forme en dent de scie. L'ouverture
chez cette espèce est assez large.
Dans notre matériel :
H. pulchra est fréquente aussi dans la coupe CES
comme dans la coupe COK. Sa fréquence maximale (25%) coincïde avec
l'échantillon CES-42.
Espèce : Heterohelix reussi (CUHMAN, 1938) Pl.3,
fig.1-2
Synonymie:
1938 - Guembelina reussi CUSHMAN, pl.2, fig.6-9, p.11
1970 - Heterohelix globulosa (EHRENBERG), Eicher et
Worstell,
1985 - Heterohelix reussi (CUSHMAN), Caron, fig. 24,
10-11
1990 - Hetrohelix globulosa (CUSHMAN), Nederbragt, pl.
2, fig. 1a-2b, p88
2001 - Heterohelix reussi (CUSHMAN), Keller et al,
Fig.8, 1-3
2004 - Heterohelix reussi (CUSHMAN), Keller et Pardo,
Pl. 6 fig. 11-14
Caractères généraux :
C'est une espèce qui présente un agencement
bisérié avec des loges globuleuses qui augmentent de taille
graduellement, à l'exception des dernières loges, qui croissent
plus rapidement. Toutes les loges présentent une ornementation en fines
côtes et sont finement perforées. Cette espèce se distingue
de H. globulosa par sa forme plus allongée.
104
Répartition stratigraphique :
H. reussi été signalée dans les
dépôts du Crétacé supérieur.
C'est-à-dire du Cénomanien supérieur au Maastrichtien.
Dans notre matériel :
H. reussi est présente dans les niveaux de
base des séries étudiées jusqu'au dernier
échantillon. Sa fréquence est relativement élevée
par rapport aux autres Hétérohélicidés. Cette
espèce atteint 35% de l'association dans la partie inférieure de
la zone à W. archaeocretacea.
SUPERFAMILLE : HETEROHELICACEA
CUSHMAN, 1927
FAMILLE : GUEMBELITRIIDAE
MONTANARO GALLITELLI, 1957
SOUS-FAMILLE : GUEMBELITRIINAE
MONTANARO GALLITELLI, 1957 GENRE :
Guembelitria CUSHMAN, 1933
Espèce - Type : Guembelitria cretacea
CUSHMAN
Pl.2, fig.9
Synonymie du genre :
1933 - Guembelitria CUSHMAN, p. 37
1954 - Guembelitria CUSHMAN, Stelck et Wall, p.
1956 - Guembelina EGGER, Dalbiez, p. 35-37
1964 - Guembelitria CUSHMAN, Loeblich et Tappan, p.
c652
1970 - Neobulimina CUSHMAN et WICKENDEN, Eicher et
Worstell
1985 - Guembelitria CUSHMAN, Caron, p. 23
1988 - Guembelitria CUSHMAN, Loeblich et Tappan, p.
473
Diagnose du genre :
C'est une espèce de très petite taille que nous
rencontrons dans les résidus fins (<100um). Ce genre comporte des
espèces à un test subconique trisérié et à
des loges globuleuses. Ce test est finement perforé, chaque pore est
entouré par un cône le plus souvent émoussé (par
fois il y a deux pores par cône). De telle microstructure est dite
à « Pore-mounds » par Olsson et al., (1999)
Espèce : Guembelitria cenomana (KELLER,
1935)
Pl.2, fig.1-3
105
Synonymie :
1935 - Guembelina cenomana KELLER, pl. 3, fig. 13-14,
p. 547
1940 - Guembelitria harrisi TAPPAN, pl. 19, fig. 2,
p. 115
1964 - Guembelitria cenomana (KELLER), Loeblich et
Tappan, p. 652
1970 - Guembelitria harrisi TAPPAN, Eicher et
Worstell, Pl.8, fig.1-2.
1985 - Guembelitria cenomana (KELLER), Caron, fig.
24.3-4, p. 55
1988 - Guembelitria cenomana (KELLER), Loeblich et
Tappan, pl. 485, fig. 8-12
2001 - Guembelitria cenomana (KELLER), Keller et
al., fig.8. 6-8, p. 404
2004 - Guembelitria cenomana (KELLER), Keller et
Pardo fig.8. 6-8, p. 404
Caractères généraux :
Guembelitria cenomana, possède un test
trisérié non allongé, à surface perforée.
Elle diffère de G. cretacea du Maastrichtien-Danien seulement
par son ouverture subcirculaire relativement plus petite.
Répartition stratigraphique :
Guembelitria cenomana est connue comme un indicateur
paléoécologique habitant les surfaces marines eutrophiques
à la période du passage Cénomanien Turonien, son
épanouissement (bloom) coincide avec l'événement anoxique
EAO-2. Elle a été signalée dans l'Albien supérieur
(zone à Rotalipora appenninica) et s'étend à la
partie supérieure de la zone à Whiteinella archaeocretacea
c'est-à-dire prés de la limite Cénomanien -
Turonien.
Dans notre matériel :
C'est la première fois que cette espèce est
reconnue en Tunisie. Cette espèce est mondialement reconnue comme un
marqueur de période de crise ou stress écologique. Elle est
présente depuis les premiers niveaux jusqu'au dernier échantillon
de chaque coupe.
Espèce : Guembelitria albertensis STELCK et
WALL, 1954
Pl.2, fig.4-6
Synonymie :
1954 - Guembelitria cretacea CUSHMAN Var.
albertensis STELCK et WALL, Pl. 2, Fig. 19, p. 23 1955 - Guembelitria
cretacea CUSHMAN Var. spiritensis STELCK et WALL, Pl. 2, Fig. 11,
p. 44 1962 - Neobulimina albertensis (STELCK et WALL), Tappan, Pl. 48,
Fig. 3-6, p. 184
1970 - Neobulimina albertensis (STELCK et WALL), Eicher
et Worstell, Pl. 4, Fig. 2-4, p. 290 2001 - Guembelitria albertensis
STELCK et WALL, Keller et al., fig.8. 9-10, p. 404
106
2004 - Guembelitria cretacea CUSHMAN, Ettachfini et
al., fig.7. 12, p.66
Caractères généraux :
Cette espèce se caractérise par un test
trisérié allongé. La partie où s'effectue la
trisériation chez cette espèce est remarquablement plus
allongée que chez Guembelitria cenomana. L'ouverture en arc
chez Guembelitria albertensis est nettement plus courte. Le test
trisérié allongé est orné par des petits pores.
Cette espèce se distingue aussi de G. cenomana par sa forme
plus mince et peu allongée.
Répartition stratigraphique :
Guembelitria albertensis est connue l'Albien
supérieur dans la zone à Rotalipora apenninica au
Turonien (partie inférieure de la zone à Helvetoglobotruncana
helvetica).
Dans notre matériel :
C'est la première fois que cette espèce est
reconnue en Tunisie. Concernant la coupe CES G. albertensis coexiste
avec les Rotalipores, alors que pour la coupe COK elle est déjà
présente dans les marnes de Cénomanien attribuées à
la Formation Fahdènes.
SUPERFAMILLE : PLANOMALINACEA
BOLLI, LOEBLICH & TAPPAN, 1957
FAMILLE : GLOBIGERINELLOIDIDAE
LONGORIA, 1974 SOUS-FAMILLE :
GLOBIGERINELLOIDINAE LONGORIA, 1974 GENRE :
Globigerinelloides CUSHMAN & TEN DAM, 1948
Espèce - Type : Globigerinelloides algeriana
CUSHMAN & TEN DAM, 1948
Synonymie du genre :
1948 - Globigerinelloides CUSHMAN & TEN DAM
1964 - Globigerinelloides CUSHMAN & TEN DAM,
Loeblich et Tappan, p. c656 1985 - Globigerinelloides CUSHMAN &
TEN DAM, Caron, p. 22, 47
1988 - Globigerinelloides CUSHMAN & TEN DAM,
Loeblich et Tappan, p. 22, 47
Diagnose du genre :
Ce genre groupe les espèces à test
planispiralé doublement ombiliquées (biombiliqué) qui peut
être involute soit évolute à loges globuleuses dont la
dernière peut se deviser surtout
l'épaisseur du test. Les représentants du genre
Globigerinelloides possèdent des sutures
déprimées sur les deux faces pouvant être radiales, droites
ou sigmoïdes. La périphérie du test est arrondie et
dépourvue de carènes. Comportant des ouvertures successives
considérées comme étant des ouvertures résiduelles
sur les deux cotés, elles sont typiques pour les tests
planispiralés. Généralement, Globigerinelloides
diffère de Biticinella par l'absence des ouvertures
supplémentaires et de Planomalina par l'absence de
carène périphérique.
Espèce : Globigerinelloides bentonensis
(MORROW, 1934)
Pl.1, fig.11-12
Synonymie
1934 - Anomalina bentonensis MORROW, Pl. 30, Fig. 4, p.
201
1961 - Globigerinelloides bentonensis (MORROW), Loeblich
et Tappan, Pl. 2, Fig. 8-10, p. 267
1970 - Globigerinelloides bentonensis (MORROW), Eicher
et Worstell, pl. 8, Fig. 17-19, Pl. 9,
Fig. 3, P. 297
1979 - Globigerinelloides bentonensis (MORROW), Chitta, p.77
2001 - Globigerinelloides bentonensis (MORROW), Keller
et al., texte seulement.
2004 - Globigerinelloides bentonensis (MORROW), Keller
et Pardo, Pl.5, fig. 15-18
Caractères généraux :
G. bentonensis possède un test
biombiliqué, planispiral evolute à partiellement involute. Le
test contient de 6 à 8 loges (généralement 6). La taille
augmente graduellement. Les premières loges sont étroitement
comprimées alors que les dérnières sont moins
comprimées et tendent à devenir évolute. La
périphérie est lobée, les sutures bsont radiales qui
peuvent être légérement courbées. Le test est
finement perforé. Cette espèce diffère de G. ultramicra
par la dernière loge qui est relativement plus grande et qui englobe le
test.
Répartition stratigraphique :
Cette espèce est signalée dans l'Albien et elle
disparaît dans la partie inférieure de la zone à
Whiteinella archaeocretacea. G. bentonensis forme une
sous-zone dans cette zone.
107
Dans notre matériel :
108
C'est la première fois que cette espèce est
reconnue en Tunisie. Concernant les deux coupex CES et COK, G. bentonensis
coexiste avec les Rotalipores. Dans la partie supérieure à
R. cushmani cette espèce est fréquente.
Espèce : Globigerinelloides ultramicra
(SUBBOTINA, 1949)
Pl.1, fig.13/Pl.5, fig.6
Synonymie
1949 - Globigerinella ultramicra SUBBOTINA, pl. 2, fig.
17-18, p. 33
1964 - Globigerinelloides ultramicra (SUBBOTINA),
Loeblich et Tappan, c656 1985 - Globigerinelloides ultramicra
(SUBBOTINA), Caron, fig. 29.18-19, p. 66 1988 - Globigerinelloides
ultramicra (SUBBOTINA), Loeblich et Tappan, 2001 - Globigerinelloides
ultramicra (SUBBOTINA), Keller et al., fig.8.11-13, p. 404 2004 -
Globigerinelloides ultramicra (SUBBOTINA), Keller et Pardo, Pl.5,
10-14
Caractères généraux :
Globigerinelloides ultramicra possède un test
planispiralé biombiliqué à enroulement évolute. Les
loges sont globuleuses est peuvent souvent être en nombre de huit. Chez
cette é*espèce les sutures sont typiquement
déprimées sur les deux côtés, radiales et droites.
La periphérie est à surface arrondie et dépourvue de
carènes. A la dernière loge l'ouverture est marginao-ombilicale
sur les loges précédante on observe des relicats d'ouverture au
niveau des deux ombilics. Généralement, Globigerinelloides
ultramicra diffère des autres Globigerinelloides
essentiellement par sa petite taille et son grand nombre de loges. Cette
espèce de G. bentonensis par son contour moins lobé et
son enroulement moins involute.
Répartition stratigraphique :
Selon Caron (1985) Globigerinelloides ultramicra est
connue de l'Albien supérieur au Maastrichtien inférieur, (la zone
à Rotalipora appenninica à la zone à
Globotruncana havanensis).
Dans notre matériel :
Cette espèce est présente dans notre
matériel d'une manière sporadique.
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958
FAMILLE: HEDBERGELLIDAE
LOEBLICH & TAPPAN, 1961 SOUS-FAMILLE:
HEDBERGELLINAE LOEBLICH & TAPPAN, 1961 GENRE:
Clavihedbergella BANNER & BLOW, 1959
109
Espèce-type : Hastigerinella
(=Clavihedbergella) subcretacea TAPPAN, 1943
Synonime du genre
1959 - Praeglobotruncana (Hedbergella) BANNER &
BLOW, p.18 1959 - Praeglobotruncana (Clavihedbergella) BANNER &
BLOW, p.18 1961 - Planogyra ZAKHAROVA-ATAKBEKJAN, p.50
1979 - Hedbergella BRÖNNIMANN & BROWN, E.W.G.P
.F.1, p. 115 1985 - Hedbergella BRÖNNIMANN & BROWN, Caron, p.
23
2000 - Hedbergella BRÖNNIMANN & BROWN,
Georgescu, p. 163
Diagnose du genre
Test trochospiralé à spire basse, biconvex,
à ombilic large, périphérie est subcirculaire et
lobée. Ce test est dépourvu de carènes. Les
premières loges sont globuleuse à ovales, alors que les
dernières sont étirées à allongées. Les
sutures sont fortement retrécies, radiales, droites ou courbées.
Les loges sont à surface finement perforée, granuleuse et lisse.
L'ouverture est estraombilical-obilicale en forme d'arc. Elle est bordée
par une lèvre.
Espèce: Clavihedbergella simplex
(MORROW, 1934)
Pl.4, fig.4-6
Synonymie:
1934 - Hastigerinella simplex MORROW, pl. 30, fig. 6, p.
198
1954 - Hastigerinella simplicissima MAGNÉ &
SIGAL, pl. 14, fig. 11a-b-c
1961 - Hedbergella amabilis LOEBLICH & TAPPAN, pl.
13, fig. 1a-b-c, p.273
1961 - Clavihedbergella simplex (MORROW), Loeblich et Tappan,
pl.3, fig.11-14
1970 - Clavihedbergella simplex (MORROW), Eicher et Worstell, pl.
10, fig. 4, 6-7
1971 - Hedbergella simplississima (MAGNÉ &
SIGAL), Magniez-Jannin, pl.19, fig.12-16, p361
1977 - Clavihedbergella simplex (MORROW), Master
1979 - Hedbergella simplex (MORROW), E.W.G.P .F.1,pl.
29,fig. 1-3, pl. 30, fig.1-2, p.147,149
1979 - Clavihedbergella simplex (MORROW), Chitta, p.98
1985 - Hedbergella simplex (MORROW), Caron, fig. 25,
15-16, p.58
1983 - Clavihedbergella simplex (MORROW), Gargouri-Razgallah,
p.140
1987 - Hedbergella amabilis (MORROW), Ben Haj Ali, pl.
4, fig. 9-11, p. 97
1993 - Hedbergella simplex (MORROW), Balla et al, fig.
5, 4, p. 436
2000 - Hedbergella amabilis (MORROW), Georgescu, pl.1,
fig. 3-4, p. 165
2001 - Hedbergella simplex (MORROW), Keller et al.,
fig.8, 14, p. 404
Caractères généraux:
Test trochospiral à spire basse. Le dernier tour est
formé généralement par quatre à six loges
légérement étirées radialement à surface
granuleuse (ou épineuse). Elles sont à
110
croissance rapide avec un contour très lobé
à digité. Les sutures sont typiquement radiales et
déprimées. La face ombilicale est faiblement convexe.
Répartition stratigraphique :
C. simplex est connue de l'Albien inférieur
à la limite Turonien - Coniacien. (Caron, 1985).
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958
FAMILLE: HEDBERGELLIDAE
LOEBLICH & TAPPAN, 1961
SOUS-FAMILLE: HEDBERGELLINAE
LOEBLICH & TAPPAN, 1961 GENRE:
Hedbergella BRÖNNIMANN & BROWN, 1958
Espèce - Type : Anomalina (=Hedbergella)
lorneiana D'ORBIGNY var. trochoidea GANDOLFI, 1942
Synonymie du genre :
1959 - Praeglobotruncana (Hedbergella) BANNER &
BLOW, p.18 1959 - Praeglobotruncana (Clavihedbergella) BANNER &
BLOW, p.18 1961 - Planogyra ZAKHAROVA-ATAKBEKJAN, p.50
1979 - Hedbergella BRÖNNIMANN & BROWN,
E.W.G.P .F.1, p. 115 1985 - Hedbergella BRÖNNIMANN & BROWN,
Caron, p. 23
2000 - Hedbergella BRÖNNIMANN & BROWN,
Georgescu, p. 163
Diagnose du genre:
Ce genre est caractérisé par un test
trochospiralé finement perforé et calcaire, biconvexe
représenté par un contour lobé. Les loge sont globuleuses
à ovales, lobés à profile arrondi ne contenant ni
carène ni bandeau périphérique imperforé.
L'ouverture primaire chez Hedbergella est extraombilicale-ombilicale,
s'étendant jusqu'à la périphérie. Les sutures sont
déprimées, radiales, courbées ou droites. La surface des
loges peut être lisse comme elle peut être couverte soit par des
rugosités ou par des épines.
Espèce : Hedbergella delrioensis
(CARSEY, 1926)
Pl.4, fig.1-3
Synonymie:
1926 - Globigerina cretacea D'ORBIGNY var.
delrioensis CARSEY, p.43
1961 - Hedbergella delrioensis (CARSEY), Loeblich et
Tappan, pl.2, fig. 11-13, p. 275 1967 - Hedbergella delrioensis
(CARSEY), Pessagno, pl. 48, fig. 1-5, p. 282
1971 - Hedbergella delrioensis (CARSEY), Magniez-Jannin,
pl. 13, fig. 13-34, p.376
111
1979 - Hedbergella delrioensis (CARSEY), E.W.G.P.F.1,
pl. 22, fig.1-2, pl. 23,fig.1-3, p.125,127
1979 - Hedbergella delrioensis (CARSEY), Chitta,
p.90, pl.2, fig.1-2
1983 - Hedbergella delrioensis (CARSEY),
Gargouri-Razgallah, p. 140, pl.24, fig. 1-3
1985 - Hedbergella delrioensis (CARSEY), Caron, fig.
25, 6-7, p.58
1993 - Hedbergella delrioensis (CARSEY), Caus et
al., fig.7, 1a-c, p. 541
1994 - Hedbergella delrioensis (CARSEY), Premoli
Silva & Sliter, pl. 1, fig. 7
2000 - Hedbergella delrioensis (CARSEY), Georgescu,
pl. 1, fig. 5, p. 165
2001 - Hedbergella delrioensis (CARSEY), Keller
et al., fig.8, 15-16, p. 404
2004 - Hedbergella delrioensis (CARSEY), Keller et
Pardo, Pl.5 5-9
Caractères généraux :
Cette espèce montre un agencement trochospiralé
du test au contour lobé, contenant au dernier tour 5 à 7 loges.
Les deux dernières loges sont décalées du
côté ventral. Les loges sont à croissance rapide. La
surface des premières loges montre un leger aspect rugueux tant disque
la dernière loge est lisse. Les sutures sont radiales et
déprimées. L'ombilic est assez étroit.
Répartition stratigraphique :
H. delrioensis est connue de l'Aptien moyen au
Coniacien. (Caron, 1985). Dans notre matériel, cette espèce est
présente depuis la partie supérieure de la zone à
Rotalipora cushmani.
Espèce : Hedbergella planispira (TAPPAN, 1940)
Pl.6, fig.3
Synonymie :
1940 - Globigerina planispira TAPPAN, pl. 19, fig. 12,
p. 122
1961 - Hedbergella planispira (TAPPAN), Loeblich et
Tappan, pl. 5, fig. 4-10, p. 276
1979 - Hedbergella planispira (TAPPAN), E.W.G.P .F.1,pl.
27,fig.1-3, pl. 28, fig. 1-4, p.140,142
1979 - Hedbergella planispira (TAPPAN), Chitta, p.92
1983 - Hedbergella planispira (TAPPAN),
Gargouri-Razgallah, p.139, pl.2-4, fig.8
1984 - Hedbergella planispira (TAPPAN), Leckie, pl. 9,
fig. 6-7, p. 599
1985 - Hedbergella planispira (TAPPAN), Caron, fig. 25,
23-24, p. 58
1994 - Hedbergella planispira (TAPPAN), Premoli Silva
& Sliter, pl. 1, fig. 1, 10
2000 - Hedbergella planispira (TAPPAN), Georgescu, pl.
1, fig. 8, p. 165
2001 - Hedbergella planispira (TAPPAN), Keller et
al., fig. 8, 17-18, p. 404
Caractères généraux :
Cette espèce possède un test de petite de taille
trochospiralé à trochospire basse et à contour
lobé. Les loges peuvent être de six à huit au dernier tour,
globuleuses et à surface
112
lisse. Les loges sont à croissance lente. Les sutures
sont radiales et déprimées. Chez cette espèce le profil
est très faiblement dissymétrique
Répartition stratigraphique :
H. planispira a été signalée
dans l'Aptien inférieur au Turonien. (Caron, 1985). Dans notre
matériel, cette espèce est présente depuis la partie
supérieure de la zone à Rotalipora cushmani et continue
jusqu'au Turonien inférieur à moyen (Zone à H.
helvetica).
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958
FAMILLE: HEDBERGELLIDAE
LOEBLICH & TAPPAN, 1961
SOUS-FAMILLE: HEDBERGELLINAE
LOEBLICH & TAPPAN, 1961 GENRE :
Whiteinella PESSAGNO, 1967
Espèce - Type : Whiteinella
archaeocretacea, PESSAGNO, 1967
Synonymie du genre :
1967 - Whiteinella PESSAGNO, p. 298
1979 - Whiteinella PESSAGNO, E.W.G.P.F.1, p.155
1983 - Hedbergellita MASLAKOVA, p. 29
1985 - Whiteinella PESSAGNO, Caron, p. 79
1988 - Whiteinella PESSAGNO, Loeblich et Tappan, p.
462
1998 - Whiteinella PESSAGNO, Rami, p. 177
2000 - Whiteinella PESSAGNO, Georgescu, p. 163
Diagnose du genre :
Le test est caractérisé par trochospire basse,
biconvexe et à contour lobé. En profil il est parfois
hémisphérique. Il test ne contient pas de véritable
carènes parfois au niveau de la marge, il peut être marqué
par une rangée de pustules comme chez Whiteinella
archaeocretacea. L'ouverture primaire est principalement
extraombilicale-ombilicale généralement restreinte. L'ombilic est
relativement large et partiellement recouvert par des portici qui se recouvrent
sans se souder. Les sutures spirales et ombilicales sont
déprimées et radiales.
Espèce: Whiteinella aprica (LOEBLICH
& TAPPAN, 1961)
Pl.5, fig.5
113
Synonymie:
1961 - Ticinella aprica LOEBLICH & TAPPAN, pl. 4,
fig. 14-16, p. 292
1979 - Whiteinella aprica (LOEBLICH & TAPPAN),
E.W.G.P.F.1, pl.32, fig.1-2, p.157
1983 - Whiteinella aprica (LOEBLICH & TAPPAN),
Bellier, pl.12, fig. 1-2, p.50-52
1990 - Whiteinella aprica (LOEBLICH & TAPPAN),
Robaszynski et al., pl. 27, fig.4, 6, 7, p.328
1994 - Whiteinella aprica (LOEBLICH & TAPPAN),
Premoli silva & Sliter, pl.10, fig. 3, 8
1998 - Whiteinella aprica (LOEBLICH & TAPPAN),
Rami, pl. 10, fig. 6-8
2000 - Whiteinella aprica (LOEBLICH & TAPPAN),
Georgescu, pl.1, fig. 11-13, p. 165
2001 - Whiteinella aprica (LOEBLICH & TAPPAN),
Keller et al., fig. 9, 4
Caractères généraux :
Cette espèce montre un profil à trochospire
basse très faiblement dissymétrique avec une face spirale plane
à légèrement convexe. Les loges de W. aprica sont
globuleuses à surface uniformément pustuleuse. Elles montrent une
croissance lente à profil circulaire. L'ouverture primaire est
extraombilicale-ombilicale, s'étendant vers la périphérie,
bordée par un porticus. Les sutures sont radiales et
déprimées, le contour est nettement lobé. L'ombilic est
peu large et comporte des portici.
Répartition stratigraphique :
Cette espèce est commune du Cénomanien
supérieur au Turonien moyen (E.W.G.P.F., 1979). Dans notre
matériel de la coupe CES comme de la coupe COK, W. aprica est
est associée à R. cushmani.
Espèce : Whiteinella archaeocretacea
PESSAGNO, 1967
Pl.1, fig.21
Synonymie
1967 - Whiteinella archaeocretacea PESSAGNO, pl.51,
fig.2-4, p.298
1979 - Whiteinella archaeocretacea PESSAGNO,
E.W.G.P.F.1,pl.33-34,fig1-3 ;fig1-2 ;p.163, p.165
1985 - Whiteinella archaeocretacea PESSAGNO, Caron,
fig.37;4-5, p.79
1990 - Whiteinella archaeocretacea PESSAGNO, Robaszynski
et al., pl.29, fig.1-2, p.332 1998 - Whiteinella archaeocretacea
PESSAGNO, Rami, pl.10, fig9; pl.11, fig.1
2003 - Whiteinella archaeocretacea PESSAGNO,
Aguilera-Franco, pl.1, fig.11 p.208 2004 - Whiteinella archaeocretacea
PESSAGNO, Ettachfini et al., fig.7 ; 14a-c, p.66
Caractères généraux :
Chez W. archaeocretacea, le test présente une
trochospire basse, il est formé généralement de cinq loges
initialement globuleuses. Les loges suivantes sont allongées suivant le
sens de l'enroulement. En profil, les loges apparaissent comprimées
114
latéralement et montrent un bord
périphérique faiblement perforé et parfois subanguleux.
L'ouverture primaire, extraombilicale-ombilicale, est recouverte partiellement
par des portici. Les sutures sont déprimées et radiales.
Répartition stratigraphique :
W. archaeocretacea est connue du Cénomanien
supérieur jusqu'au Santonien (Caron, 1985). Dans notre matériel,
cette espèce est présente déjà dans la partie
supérieure de la zone de R. cushmani.sous-zone à D.
algeriana et dans tout l'intervalle étudié de la coupe CES.
Dans la coupe COK, elle démarre également dans la sous-zone
à D. algeriana du Cénomanien supérieur.
Espèce : Whiteinella baltica DOUGLAS
& RANKIN, 1969
Pl.4, fig.10-11
Synonymie
1969 - Whiteinella baltica DOUGLAS & RANKIN, fig.9,
p.198
1979 - Whiteinella baltica DOUGLAS & RANKIN,
E.W.G.P.F.1,pl.35,Fig.1-5; pl.36,fig.1-2;p169, p.174
1985 - Whiteinella baltica DOUGLAS & RANKIN, Caron,
fig.37; 1-3, p. 79
1990 - Whiteinella baltica DOUGLAS & RANKIN,
Robaszynski et al., pl. 27 fig.1,8, p.328
1994 - Whiteinella baltica DOUGLAS & RANKIN, Premoli
Silva & Sliter, pl.11, fig.11
2000 - Whiteinella baltica DOUGLAS & RANKIN,
Georgescu, pl. 1, fig. 6-7, p. 165
2001 - Whiteinella baltica DOUGLAS & RANKIN, Keller
et al., fig. 9, 1-3, p. 406
Caractères généraux :
Le test de cette espèce est constitué au dernier
tour le plus souvent par quatre loges très globuleuses. Elles sont
toutes à surface rugueuse à contour distinctement lobé
montrant une croissance nettement rapide. Les sutures sont radiales et
typiquement déprimées. L'ouverture principale est
extraombilicale-ombilicale, bordée par un porticus recouvrant une partie
de l'ombilic. En profil cette espèce montre une trochospire basse et
symétrique.
Répartition stratigraphique :
Cette espèce est signalée dans le
cénomanien supérieur jusqu'à la limite coniacien -
Santonien. Dans la coupe CES, cette espèce est présente
déjà dans la sous-zone à D. algeriana de la zone
à Rotalipora cushmani du Cénomanien supérieur et
dans les niveaux sus-jascents. Dans la coupe COK, elle est plutôt
présente dans les premiers niveaux correspondant à la sous-zone
à R. greenhornensis.
115
Espèce: Whiteinella brittonensis
(LOEBLICH & TAPPAN, 1961)
Pl.8, fig.5
Synonymie :
1961 - Hedbergella brittonensis LOEBLICH et TAPPAN, pl.
4, fig. 1-8, p. 274
1979 - Whiteinella brittonensis (LOEBLICH et TAPPAN),
E.W.G.P.F.1, pl. 37, fig.1-2, p.177, fig.1-2, p. 179
1979 - Hedbergella brittonensis LOEBLICH et TAPPAN,
Chitta, p.89
1980 - Hedbergella brittonensis LOEBLICH et TAPPAN,
Salaj, pl. 7, fig.4
1994 - Whiteinella brittonensis (LOEBLICH et TAPPAN),
Premoli Silva & sliter, pl.10,11, fig.2,8
2000 - Whiteinella brittonensis (LOEBLICH et TAPPAN),
Georgescu, p. 164.
2001 - Whiteinella brittonensis (LOEBLICH et TAPPAN),
Keller et al., fig. 9, 5, p. 408
Caractères généraux :
Cette espèce montre un test composé au dernier
tour le plus souvent par six loges globuleuses à croissance lente et
à surface granuleuse. Le contour est lobé subcirculaire. Les
sutures sont déprimées et radiales. L'ouverture principale est
extraombilicale-ombilicale, n'atteignant pas la périphérie, elle
est bordée par un porticus. En profil, W. brittonensis se
distingue des autres Whiteinelles par une trochospire moyennement
élevée déterminant un profil dissymétrique.
Répartition stratigraphique :
W. brittonensis est signalé dans le
Cénomanien supérieur et dans le reste du Crétacé
supérieur. Dans la coupe CES, cette espèce est présente
déjà dans la sous-zone à D. algeriana de la zone
à Rotalipora cushmani du Cénomanien supérieur et
dans les niveaux sus-jascents. Dans la coupe COK, elle est plutôt
présente dans les premiers niveaux correspondant à la sous-zone
à R. greenhornensis.
Espèce: Whiteinella paradubia (SIGAL,
1952)
Pl.1, fig.8-9
Synonymie :
1952 - Globigerina paradubia SIGAL, fig. 28, p. 28
1979 - Whiteinella paradubia (SIGAL), E.W.G.P.F.1, pl.
39, fig.1-2, p.183 1994 - Whiteinella paradubia (SIGAL), Premoli Silva
& Sliter, pl. 10, fig. 1,5 2000 - Whiteinella paradubia (SIGAL),
Georgescu, pl. 1, fig. 9-10, p. 165 2001 - Whiteinella paradubia
(SIGAL), Keller et al., fig.9, 6 ; p.406
Caractères généraux :
W. paradubia se distingue par une très haute
trochospire déterminant un profil nettement dissymétrique. Le
test de cette espèce est constitue le plus souvent par sept loges
116
globuleuses au dernier à surface granuleuse.
L'ouverture primaire est extraombilicale-ombilicale bordée par un
porticus. Les loges dessinent un contour lobé subcirculaire
séparées par des sutures sont radiales et
déprimées. Le seul critère distinctif de cette
espèce par rapport à W. brittonensis est la hauteur de
la spire.
Répartition stratigraphique :
W. paradubia se répartie depuis le
Cénomanien supérieur et continue jusqu'au crétacé
supérieur. Dans la coupe CES comme la coupe COK, cette espèce est
présente déjà dans la sous-zone à D. algeriana
appartenant à la zone à Rotalipora cushmani du
Cénomanien supérieur mais qui marque des présences
sporadiques dans les niveaux sus-jascents.
Espèce: Whiteinella praehelvetica
(TRUJILLO, 1960)
Pl.4, fig.6-8
Synonymie
1960 - Rugoglobigerina praehelvetica TRUJILLO, pl.49,
fig.6 p.340
1979 - Praeglobotruncana praehelvetica (TRUJILLO),
E.W.G.P.F.2, pl. 49, fig.1-2, p.43
1983 - Whiteinella praehelvetica (TRUJILLO), Bellier,
pl. 12, fig. 10-12, p.54
1985 - Helvetoglobotruncana praehelvetica (TRUJILLO),
Caron, fig.30; 9-10, p. 68
1990 - Praeglobotruncana praehelvetica (TRUJILLO),
Robaszynski et al. pl. 42, fig.4 1993 - Whiteinella praehelvetica
(TRUJILLO), Balla et al., fig. 5 ; 5,11 p.436 1998 - Whiteinella
praehelvetica (TRUJILLO), Rami, pl.11, fig. 2-4
2001 - Helvetoglobotruncana praehelvetica (TRUJILLO),
Keller et al., p. 407
Caractères généraux :
W. praehelvetica est caractérisée par
des loges à section trapézoïdales, subaplaties à la
face spirale. Le contour du test est lobé. Les sutures sont radiales et
légèrement déprimées. L'ouverture principale est
extraombilicale-ombilicale bordée par une lèvre bien
développée pouvant atteindre la taille d'un porticus, l'ombilic
est relativement étroit (1/4 environ du diamètre maximal). En
profil, cette espèce est nettement dissymétrique, ne
présentant pas de véritable carène mais plus tôt une
ligne de pustules. L'absence d'une véritable carène et la
présence d'un portici pourraient placer l'espèce
praehelvetica dans le genre Whiteinella.
Répartition stratigraphique :
L'espèce praehelvetica est signalé au
sommet du Cénomanien jusqu'au Turonien moyen c'est-à-dire depuis
la partie terminale de la zone de Rotalipora cushmani jusqu'à
la partie
117
moyenne à supérieure de la zone de
Helvetoglobotruncana helvetica. Dans notre matériel, elle est
présente dans la partie terminale de Rotalipora cushmani. Dans
l'intervalle de la sous-zone à D. algeriana de la coupe CES
dés l'échantillon CES-12 et elle continue jusqu'au dernier
échantillon tout en étant associée à H.
helvetica. Dans la coupe COK, elle apparaît dans
l'échantillon COK-52 de la sous-zone à Dicarinella hagni
de la zone à W. archaeocretacea.marquant le passage
Cénomanien Turonien.
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958
FAMILLE: HEDBERGELLIDAE
LOEBLICH & TAPPAN, 1961 SOUS-FAMILLE :
ROTUNDININAE BELLIER & SALAJ, 1977 GENRE
: Praeglobotruncana BERMUDEZ, 1952
Espèce - Type: Globorotalia
(=Praeglobotruncana) delrioensis, PLUMMER, 1931
Synonymie du genre :
1931 - Globorotalia PLUMER, p. 129
1949 - Globotruncana CUSHMAN, Gandolfi p. 609 1953 -
Rotundina SUBBOTINA,
1959 - Praeglobotruncana BERMUDEZ, Klaus, p. 793 1966 -
Praeglobotruncana BERMUDEZ, Caron, p. 72 1979 - Praeglobotruncana
BERMUDEZ, E.W.G.P.F.2, p. 15 1985 - Praeglobotruncana BERMUDEZ,
Caron, p. 65 2000 - Praeglobotruncana BERMUDEZ, Georgescu, p. 164
Diagnose du genre :
Le genre Praeglobotruncana est
carctérisé par un test trochospiralé biconvexe à
planoconvexe avec un contour peu lobé. L'ouverture principale est
extraombilicale-ombilicale n'atteignant pas la périphérie avec un
ombilic étroit (ne dépassant 1/4 du diamètre maximal),
bordée par les extensions lamelleuses plus ou moins recourbées
des loges successives. Seules les formes évoluées comme P. gibba
possèdent des portici. En profil, le test montre une accumulation de
pustules qui tendent à s'aligner parallèlement au niveau de la
périphérie même des premières loges du dernier tour,
donnant l'impression d'une ou de deux carènes pas
régulièrement espacées.
Espèce : Praeglobotruncana delrioensis
(PLUMMER, 1931)
Pl.5, fig.11-12
118
Synonymie
1931 - Globorotalia delrioensis PLUMMER, p.199, pl.13,
fig.2
1946 - Globorotalia marginaculata LOEBLICH & TAPPAN,
p.257, pl.37, fig.19-21 1952 - Praeglobotruncana delrioensis
(PLUMMER), Bermudez, p.52 pl.7, fig.1 1961 - Praeglobotruncana
delrioensis (PLUMMER), Loeblich et Tappan, pl.6 fig.9-12 1970 -
Praeglobotruncana stephani (GANDOLFI), Eicher et Worstell, pl.10,
fig.9 1979 - Praeglobotruncana delrioensis (PLUMMER), EWGPF, p.29, 32,
pl.43, fig.1-2 1979 - Praeglobotruncana delrioensis (PLUMMER), Chitta,
P.99-100,pl.13, fig.1-6 1984 - Praeglobotruncana delrioensis
(PLUMMER), Leckie, p.600, pl.12, fig.1-8 1994 - Praeglobotruncana
delrioensis (PLUMMER), Premoli Silva et Sliter, pl.9, fig.5 2000 -
Praeglobotruncana delrioensis (PLUMMER), Georgescu, pl.2, fig. 1-2
Caractères généraux :
Le test de cette espèce est trochospiralé à
spire basse, biconvex à périphérie subcirculaire,. Les
premiers tours montre une carène qui s'efface dans les dernières
loges. Les sutures sur la face spirale sont courbées alors qu'elles sont
plus épaisses, radiales et déprimées sur la face
ombilicale. L'ouverture est obilicale-extraombilicale en forme d'arc qui peut
s'étendre vers la périphérie. Elle est bordée par
une lèvre.
Répartition stratigraphique
Cette espèce a été confondue avec des
espèces de P. stephani en stade juvenile. Nous avons reconnu cette
espèce dans les deux coupes étudiées. Dans la coupe CES,
cette espèce n'est présente que dans les deux premiers
échantillons. Alors que dans la coupe COK, elle s'etend jusqu'à
l'échantillon COK-36 correspondant à la sous-zone à D.
algeriana.
Espèce : Praeglobotruncana gibba KLAUS,
1960
Pl.5, fig.9-10
Synonymie
1960 - Praeglobotruncana stephani var. gibba
KLAUS, p. 301
1966 - Praeglobotruncana stephani gibba KLAUS, Caron,
pl. 2, fig. 4, p. 73
1979 - Praeglobotruncana gibba KLAUS, E.W.G.P.F.2, pl.
44,fig.1-2, p. 33; pl.45, fig. 1-2, p. 38
1979 - Praeglobotruncana gibba KLAUS, Chitta, p.101,
pl.13, fig.11-12
1980 - Praeglobotruncana gibba KLAUS, Salaj, pl. 7, fig.
11-12
1985 - Praeglobotruncana gibba KLAUS, fig. 30, 1-2
1987 - Praeglobotruncana gibba KLAUS, Ben Haj Ali, pl.
8, fig. 7-8, 15-16, p. 105
1993 - Praeglobotruncana gibba KLAUS, Balla et al., fig.
5, 3, p. 436
1993 - Praeglobotruncana gibba KLAUS, Caus et al, fig.
7, 5a-c, p.541
1994 - Praeglobotruncana gibba KLAUS, Premoli Silva
& Sliter, pl. 9, fig. 6
2000 - Praeglobotruncana gibba KLAUS, Georgescu,
p.166
2001 - Praeglobotruncana gibba KLAUS, Keller et al.,
119
Caractères généraux :
Le test de cette espèce contient au dernier tour le
plus souvent huit loges à section triangulaire ou trapézoïde
à surface légèrement bombée et granuleuse qui sont
à croissance modérée et séparées par des
sutures arquées et marquées par des pustules. L'ombilic est
relativement étroit il constitue 1/4 du diamètre maximal.
L'ouverture principale est extraombilicale-ombilicale bordée par une
lèvre bien développée émettant un prolongement vers
l'ombilic
Elle se distingue de Praeglobotruncana stephani par
une face spirale plus haute, ce qui fait que le test présente un aspect
conique, et par un contour moins lobé.
Répartition stratigraphique :
La répartition de gibba a été
discutée par plusieurs auteurs, puisqu'elle est considérée
comme l'espèce la plus évoluée parmi les
Praeglobotruncana, la plupart de certains auteurs la
considèrent qu'elle débute dans le Cénomanien moyen et
continue au Turonien supérieur et plus précisément elle
apparaît dans la partie terminale de la Zone à Rotalipora
brotzeni et disparaît dans la zone à Helvetoglobotruncana
helvetica.
Dans la coupe CES, elle est présente depuis le premier
échantillon correspondant à la sous-zone à D. algeriana et
elle disparaît au-delà de l'échantillon CES-50. dans la
coupe COK, cette espèce coexiste avec R. appenninica dans la sous-zone
à R. greenhornensis et disparaît dans l'échantillon
COK-66.
Espèce : Praeglobotruncana stephani
(GANDOLFI, 1942)
Pl.1, fig.1-2
Synonymie
1942 - Globotruncana stephani GANDOLFI, p.130, pl.3,
fig. 4, pl.4, fig.36-37, 41-44, pl.6,
fig.4, pl.9, fig.5,8, pl.14, fig.2
1957 - Praeglobotruncana stephani (GANDOLFI), Bolli,
Loeblich et Tappan, p.39, pl.9, fig.2
1961 - Praeglobotruncana stephani (GANDOLFI), Loeblich
et Tappan, pl.6, fig.1-8
1970 - Praeglobotruncana stephani (GANDOLFI), Eicher et
Worstell, pl.10, fig.9; pl.11, fig.2-3
1979 - Praeglobotruncana stephani (GANDOLFI),
EWGPF,p.47, 50, pl.48, fig. 1-3
1979 - Praeglobotruncana stephani (GANDOLFI), Chitta,
p.101-102, pl.13, fig.7-10
1984 - Praeglobotruncana stephani (GANDOLFI), Leckie,
p.600, pl.12 fig.1-12
1985 - Praeglobotruncana stephani (GANDOLFI), Caron,
p.65, fig.30 (3-4)
1994 - Praeglobotruncana stephani (GANDOLFI), Premoli
Silva et Sliter, pl.9, fig.1-3,8
2000 - Praeglobotruncana stephani (GANDOLFI), pl.2;
fig.7-8
Caractères généraux :
Le test de cette espèce est trochospiralé, la
face spirale est fortement convex, le dernier tour comporte
généralement de cinq à 7 loges, la face ombilicale est
applatie et parfois peu convex. Les premières loges sont subarrondies.
L'ombilic est étroit. Sur la face spirale, les sutures sont
déprimées et radiales sur la face ombilicale. En profile, cette
espèce est marquée par une carène qui s'efface sur les
deux ou trois dernières loges. L'ouverture est intermarginale en forme
d'arc bordée par une lèvre spatulée.
Répartition stratigraphique
Elle est connue de l'Albien supérieur (Zone à
R. apenninica) au Turonien inférieur (H. helvetica).
Dans la coupe CES, cette espèce « disparaît » dans les
premiers niveaux CES-9 (sous-zone à G. bentonensis). Dans la
coupe COK, elle est bien présente depuis le premier échantillon
correspondant à la sous-zone à R. greenhornensis (Zone
à R. cushmani du Cénomanien supérieur)
jusqu'à l'échantillonCOK-68 de la sous-zone à
(acmé) H. moremani (partie supérieure de la Zone
à Whiteinella archaeocretacea du passage C-T).
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958
FAMILLE: HEDBERGELLIDAE
LOEBLICH & TAPPAN, 1961
SOUS-FAMILLE : HELVETOGLOBOTRUNCANINAE
LAMOLDA, 1976 GENRE : Dicarinella
PORTHAULT, 1970
Espèce - Type : Globotruncana (=Dicarinella)
indica JACOB & SASTRY, 1950 synonyme de Praeglobotruncana
(=Dicarinella) hagni SCHEIBNEROVA, 1962.
Synonymie du genre :
1950 - Globotruncana CUSHMAN, Jacob et Sastry, p. 267
1967 - Marginotruncana HOFKER, Pessagno, p.
1970 - Praeglobotruncana (Dicarinella) PORTHAULT, p.
1975 - Dicarinella PORTHAULT, Longoria et Gamper, p. 1976 -
Dicarinella PORTHAULT, Lamolda, p.
1979 - Dicarinella PORTHAULT, E.W.G.P.F., p. 51 1985 -
Dicarinella PORTHAULT, Caron, p. 22
2000 - Dicarinella PORTHAULT, Georgescu, p. 166
120
Diagnose du genre :
121
Le test est trochospiralé biconvexe à contour
lobé. En profil, les loges sont soulignées par deux
carènes surtout sur les loges du dernier tour séparées par
un mince bandeau périphérique imperforé. L'ouverture est
extraombilicale-ombilicale. L'ombilic est relativement large, il atteint par
fois 1/3 du diamètre et partiellement recouvert par les portici
ménageant des ouvertures accessoires infralaminales. Les sutures sont
radiales, déprimées et arquées sur le côté
ombilical. Sur le côté spiral elles sont en relief.
Dicarinella se rapproche de Praeglobotruncana par la
position de l'ouverture principale extraombilicale-ombilicale et les sutures
radiales déprimées sur le coté ombilical. Toutefois, elle
se distingue par la présence de portici et de deux carènes
nettement séparées par un bandeau périphérique
mince imperforé. Avec ces deux derniers caractères elle se
rapproche plutôt à Marginotruncana.
Espèce : Dicarinella algeriana (CARON,
1966)
Pl.5, fig.2-4
1966 - Praeglobotruncana algeriana CARON, p.74
1979 - Dicarinella algeriana (CARON), EWGPF, p.57, 60,
pl.50, fig. 1-2
1979 - Praeglobotruncana algeriana CARON, Chitta,
p.98
1994 - Dicarinella algeriana (CARON), Premoli Silva et
Sliter,pl.12, fig.5,7;9-10
2000 - Dicarinella algeriana (CARON), Georgescu,
p.166
2001 - Dicarinella algeriana (CARON), Keller et al.,
p.408, fig.9
2004 - Dicarinella algeriana (CARON), Keller et Pardo,
fig.8 ;11, p.99
Caractères généraux :
Test trochospiral, biconvexe, de contour est lobé, le
dernier tour comporte cinq à sept loges pétaloïdes
séparées par des sutures arquées en relief sur la face
spirale et déprimées sur la face ombilicale. L'ombilic est large.
La surface du test est pérforée. Elle est pustuleuse sur la face
ombilicale de nombreuses loges. En profile, sur la périphérie on
observe deux carènes. La dernière loge peut porter une seule.
Répartition stratigraphique :
Cette espèce est connue dans le Cénomanien
supérieur (Zone à R. cushmani). Son apparition determine
la dernière sous-zone de la zone à R. cushmani. Elle
disparaît au Turonien inférieur à moyen. Dans la coupe CES
cette espèce est présente depuis le premier échantillon
jusqu'à l'échantillon CES-65 correspondant à la zone
à H. helvetica.
122
Dans la coupe COK, elle apparaît dans
l'échantillon COK-8 et continue jusqu'à l'échantillon
COK-99 correspondant à la zone à H. helvetica.
Espèce : Dicarinella hagni
(SCHEIBNEROVA, 1962)
Pl.6, fig.6
Synonymie
1962 - Praeglobotruncana hagni SCHEIBNEROVA, fig. 6, p.
219
1966 - Globotruncana roddai MARIANOS & ZINGULA, pl.
39, fig. 5, p. 340
1979 - Dicarinella hagni (SCHEIBNEROVA), E.W.G.P.F.2,
pl.56, fig.1-2,pl.57, fig. 1-2, p. 81, 83
1980 - Praeglobotruncana hagni SCHEIBNEROVA, Salaj,pl.
9, fig. 7
1985 - Dicarinella hagni (SCHEIBNEROVA), Caron, fig. 18,
1-3
1993 - Dicarinella hagni (SCHEIBNEROVA), Balla et al.,
fig. 5.
2001 - Dicarinella hagni (SCHEIBNEROVA), Keller et al.,
fig. 10. 12, p. 408
Caractères généraux :
Dicarinella hagni se caractérise par un
contour modérément lobé. Les loges sont
pétaloïdes, planes et faiblement bombées à croissance
de spire lente. L'ouverture est extraombilicale-ombilicale recouverte par des
portici s'imbriquant au tour de l'ombilic qui est large et ne dépasse
pas le 1/3 du diamètre. Les sutures sont arquées et en relief sur
la face spirale. Elles sont radiales et déprimées sur la face
ombilicale. En profil, le test est biconvexe à trochospire basse, plus
ou moins dissymétrique. Les loges globuleuses, du coté ombilical
sont ornées par deux carènes assez rapprochées.
Répartition stratigraphique :
Dicarinella hagni est présente depuis la zone
à Whiteinella archaeocretacea, la partie supérieure du
Cénomanien supérieur, et s'éteint au Turonien
supérieur.
Espèce : Dicarinella imbricata (MORNOD,
1949)
Synonymie
1949 - Globotruncana imbricata MORNOD, pl. 15, fig.
21-34, p. 589
1966 - Praeglobotruncana imbricata (MORNOD), Caron, pl.
6 , fig. 4, p. 76
1976 - Dicarinella imbricata (MORNOD), Caron,
pl.3,fig.1-6, pl.4,fig. 1-6, pl. 5, fig. 1-6, p. 332 1979 - Dicarinella
imbricata (MORNOD), E.W.G.P.F.2, pl. 58, fig. 1-2, pl. 59, fig. 1-2, p.
87, 92 1980 - Praeglobotruncana imbricata (MORNOD), Salaj, pl. 9, fig.
2
123
1985 - Dicarinella imbricata (MORNOD), Caron, fig. 18,
4-5
1992 - Dicarinella imbricata (MORNOD), Wonders, pl. 5,
fig. 1
1994 - Dicarinella imbricata (MORNOD), Premoli Silva
& Sliter, pl. 12, fig.8 2000 - Dicarinella imbricata (MORNOD),
Georgescu, pl. 2, fig. 3, p. 166 2001 - Dicarinella imbricata
(MORNOD), Keller et al., fig. 10. 8, p. 408
Caractères généraux :
Le test est composé de loges pétaloïdes
à surface plane ou bombée qui peuvent être granuleuses,
séparées, sur le coté spiral par des sutures
arquées soulignées par le prolongement septal de la carène
spirale. L'ouverture principale est extraombilicale-ombilicale bordée
par une lèvre mince visible sur les deux dernières loges sans la
présence de portici décelables. L'ombilic est relativement
étroit, il atteint 1/5 du diamètre maximal. En profil,
l'espèce imbricata est caractérisée par une trochospire
basse avec un test concavo-convexe. Les deux dernières loges sont
globuleuses, pincées et non carénées, se déversant
vers l'ombilic sur les loges précédantes. Les deux carènes
sont séparées par un bandeau périphérique
imperforé. Cette espèce diffère de D. algeriana
par les caractères de ses deux dernières loges se
déversant vers l'ombilic et à carènes effacées et
surtout par l'aspect imbriqué des loges successives. Elle diffère
aussi de D. hagni par ses loges qui paraissent décalées
les unes par rapport aux autres et par son ombilic plus profond. Elle
diffère des autres Dicarinella toujours par l'aspect
imbriqué.
Répartition stratigraphique :
Cette espèce se répartie depuis la partie
inférieure de la zone à Whiteinella archaeocretacea , le
cénomanien - Turonien, jusqu'au Coniacien moyen.
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958
FAMILLE: HEDBERGELLIDAE
LOEBLICH & TAPPAN, 1961
SOUS-FAMILLE: HELVETOGLOBOTRUNCANINAE
LAMOLDA, 1976
GENRE: Helvetoglobotruncana
REISS, 1957
Espèce - Type : Globotruncana
(=Helvetoglobotruncana) helvetica BOLLI, 1945
Synonymie du genre :
1945 - Globotruncana CUSHMAN, Bolli, p. 226 1957 -
Helvetoglobotruncana REISS, p. 137
124
1966 - Praeglobotruncana BERMUDEZ, Caron, p. 74
1979 - Praeglobotruncana BERMUDEZ, E.W.G.P.F.2, p. 15
1985 - Helvetoglobotruncana REISS, Caron, p. 23
2000 - Helvetoglobotruncana REISS, Georgescu, p. 169
Diagnose du genre :
Ce genre possède un test plano-convexe
trochospiralé. Le contour peut être lobé ou peu
lobé. Le profil est souligné par une carène nette.
L'ouverture principale est extraombilicale-omlicale avec un ombilic recouvert
de portici relativement peu large (1/3 du diamètre). A la face spirale
on distingue des sutures bien marquées en relief et arquées alors
qu'au coté ombilical elles sont radiales et déprimées. Le
test comporte généralement sept loges fortement renflées
et rugueuses
Espèce : Helvetoglobotruncana helvetica
(BOLLI, 1945)
Pl.1, fig.15
Synonymie :
1945 - Globotruncana helvetica BOLLI, pl. 9, fig. 1, p.
227
1952 - Globotruncana helvetica BOLLI, Sigal, fig. 32, p.
31
1955 - Globotruncana helvetica BOLLI, Schisfma
1957 - Globotruncana helvetica BOLLI, Bolli, pl. 13,
fig. 1, p.56
1966 - Globotruncana helvetica BOLLI, Caron, pl. 3, fig.
2, p. 74
1979 - Praeglobotruncana helvetica (BOLLI), E.W.G.P.F.2,
pl. 46, figs 1-2, p. 39
1979 - Praeglobotruncana helvetica (BOLLI), Chitta, fig.
4-6, pl. 14 p.128
1980 - Helvetoglobotruncana helvetica (BOLLI), Salaj,
pl. 8, fig. 3
1983 - Helvetoglobotruncana helvetica (BOLLI), Bellier,
pl. 13, fig. 1-3
1990 - Praeglobotruncana (Helvetoglobotruncana) helvetica
(BOLLI),Robaszynski et al, pl. 42, fig. 5-8,
1992 - Helvetoglobotruncana helvetica (BOLLI), Wonders,
pl. 4, fig. 7
1994 - Helvetoglobotruncana helvetica (BOLLI), Premoli
Silva & Sliter, pl. 11, fig. 9-10
1998 - Helvetoglobotruncana helvetica (BOLLI), Rami,
pl.3, pl.11, fig. 2, fig.5-7
2000 - Helvetoglobotruncana helvetica (BOLLI),
Georgescu, pl. 2, fig. 9, p.166
2001 - Helvetoglobotruncana helvetica (BOLLI), Keller et
al, fig. 10. 14, p. 408
Caractères généraux :
L'ouverture principale extraombolicale-ombilicale est
bordée par une lèvre se prolongeant sur l'ombilic en porticus. A
la face spirale les loges sont pétaloïdes et aplaties
séparées par des sutures en relief. La croissance de la spire est
lente. Le profil est souligné par une carène
périphérique, certains spécimens peuvent présenter
une imbrication des loges sur la face spirale donnant au profil un aspect en
escalier. Chez cette espèce l'ombilic est assez large, il peut atteindre
1/3 du diamètre du test. La présence de la carène unique
exclut helvetica du genre Whiteinella.
125
Répartition stratigraphique :
Cette espèce représente une zone d'extension totale
: zone à Helvetoglobotruncana helvetica du Turonien
inférieur à moyen.
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958 FAMILLE: ROTALIPORIDAE
SIGAL, 1958
SOUS-FAMILLE: ROTALIPORINAE
SIGAL, 1958 GENRE: Anaticinella
EICHER, 1972
Espèce-type : Globorotalia? (=Anaticinella)
multiloculata Morrow, 1934
Synonymie du genre :
1950 - Ticinella REICHEL, p. 600
1961 - Ticinella REICHEL, Loeblich et Tappan, p. 290
1969 - Ticinella REICHEL, Eicher et Worstell, p. 312 1972 -
Anaticinella EICHER, p. 256
Espèce : Anaticinella multiloculata
(Morrow, 1934)
Pl.1, fig.10/Pl.5, fig.7-8
Synonymie :
1934 - Globorotalia? multiloculata MORROW, pl. 31, gig.
3-5, p. 200
1946 - Globorotalia? multiloculata MORROW, Cushman, pl.
62, fig. 10-11, p. 153
1952 - Rugoglobogerina multiloculata (MORROW), Bermudez,
pl.22, fig.1
1956 - Thalmanninella multiloculata (MORROW), Bronnimann
et Brown. Pl. 20, fig. 1-3, p. 534
1961 - Ticinella multiloculata (MORROW), Loeblich et
Tappan, pl. 6, fig. 13, p. 292
1969 - Ticinella multiloculata (MORROW), Eicher et
Worstell, pl. 13 , Fig.2, 4, p. .312
1972 - Anaticinella multiloculata (MORROW), Eicher,
pl.11, fig2,5, p.302
2001 - Anaticinella multiloculata (MORROW), Keller
et al., fig.10, 10-11, p.408
2003 - Anaticinella multiloculata (MORROW), Desmares et
al., 2003, Fig.2.2a-c p.592
2004 - Anaticinella multiloculata (MORROW), Keller et
Pardo, pl.2, fig.1-5, p.98
Caractères généraux :
Test relativement large, trochospirale à face spirale
convexe composée de deux à trois tours, àloges
globuleuses. Sa face ombilicale composée de six à huit loges
globuleuses présente un ombilique large. Cette espèce
présente un contour lobé, les loges globuleuses augmentent
graduellement de taille. Les sutures sont distinctes. Elles sont
déprimées, radiales et légérement obliques ou
légérement courbées. La surface du test est finement
perforée et rugueuse. L'ornementation la plus développée,
étant dans la face ombilicale prés de l'ombilic. L'ouverture
principale est en forme d'arc extraombilical-oblical, bordée par une
lèvre étroite. Les ouvertures supplementaires sont très
larges et situées presque en bordure de l'ombilic.
126
Remarque : C'est la première fois que cette
espèce soit décrite en Tunisie. Cette espèce a
été rapportée par plusieurs auteurs à d'autres
genres (Globorotalia, Rugoglobigerina, Thalmanninella, Ticinella),
elle diffère de Rotalipora par sa périphérie non
carénée et de Ticinella par ces ouvertures
supplémentaires qui sont situées sur la marge ombilicale des
loges et non pas sur les sutures.
SUPERFAMILLE: ROTALIPORACEA
SIGAL, 1958 FAMILLE: ROTALIPORIDAE
SIGAL, 1958
SOUS-FAMILLE: ROTALIPORINAE
SIGAL, 1958 GENRE: Rotalipora
BROTZEN, 1942
Espèce-type: Rotalipora turonica BROTZEN,
1942
Synonymie du genre :
1942 - Rotalipora BROTZEN, p. 32.
1948 - Thalmanninella SIGAL, p. 101.
1942 - Anaticinella EICHER, p.
1973 - Pseudoticinella LONGORIA, p.
1978 - Pseudothalmanninella WONDERS. p. 125.
1979 - Rotalipora BROTZEN, Chitta, p. 103.
1979 - Rotalipora BROTZEN, E.W.G.P.F., p. 51-53.
1983 - Pseudorotalipora ION, p. 95.
1985 - Rotalipora BROTZEN, Caron, p. 24.
1988 - Rotalipora BROTZEN, Loeblich & Tappan, p.
467.
1998 - Rotalipora BROTZEN, Rami, p. 171
2000 - Rotalipora BROTZEN, Georgescu, p. 169
Diagnose du genre:
Le test est trochospiralé, de convexité variable
à loges anguleuses et rhomboédriques, parfois fortement
bombées. Le contour est peu lobé a lobé. En profil on
distingue une seule carène se prolongeant le long des sutures spirales,
parfois aussi le long des sutures ombilicales. L'ouverture principale
extraombilicale-ombilicale, atteint parfois la périphérie. Les
ouvertures supplémentaires sont ombilicales et suturales, situées
contre la paroi adombilicale des loges successives ou contre les sutures
(distale). L'ombilic est étroit ou large, parfois bordé par des
bourrelets périombilicaux. Les sutures spirales ombilicales sont
radiales ou arquées, déprimées ou en relief.
Certains auteurs ont confondu le genre Rotalipora
avec le genre Globotruncana en se basant sur l'existence d'une
seule carène et des ouvertures supplémentaires ombilicales
(Gandolfi, 1942 ; Reichel, 1949 ; et d'autres), d'autres distinguent le
genre
127
Thalmanninella lorsque les ouvertures
supplémentaires sont ombilicales et considèrent que le genre
Rotalipora est limité aux espèces à ouvertures
supplémentaires suturales (Sigal, 1948 ;1958 ; Bellier, 1983) ensuite
séparés par Maslakova (1963) sur le plan phylétique et par
Longoria (1973) sur le plan morphologique.
Nous allons suivre dans ce mémoire Sigal (1958) et
E.W.G.P.F (1979) en incluant ce genre dans Rotalipora.
Espèce : Rotalipora cushmani (MORROW,
1934)
Pl.1, fig.16-17/Pl.5, fig.1
Synonymie:
1934 - Globorotalia cushmani MORROW, pl. 31, fig. 2, 4,
p. 199,
1942 - Rotalipora turonica BROTZEN, fig. 10-11, p. 32
1946 - Globorotalia cushmani MORROW, pl. 62, fig. 9a-c,
p. 152
1948 - Rotalipora cushmani (MORROW), Sigal, p1.1, fig.
2, pl. 2, fig. 1, p. 96-100
1959 - Rotalipora (Rotalipora) turonica BROTZEN et
TAPPAN, pl. 8, fig. 1-10, p.297
1967 - Rotalipora cushmani (MORROW), Pessagno, pl. 51,
fig. 6-9
1979 - Rotalipora cushmani (MORROW), E.W.G.P.F, pl. 7,8,
fig. 1,1-2, p71, 73.
1979 - Rotalipora cushmani (MORROW), Chitta, p.108-109,
pl.11, fig1-6
1983 - Rotalipora cushmani (MORROW), Bellier, pl. 5,
fig. 5, 8, 10, 14. p. 47-49
1985 - Rotalipora cushmani (MORROW), Caron, fig.31, 8-11
; p. 70
1987 - Rotalipora cushmani (MORROW), Ben Haj Ali, pl.8,
fig.17-21, p. 105
1993 - Rotalipora cushmani (MORROW), Caus et al, fig.8,
1a-c, p. 542
1998 - Rotalipora cushmani (MORROW), Rami, pl. 2, fig.
4, 9; pl. 9, fig. 5-7
2001 - Rotalipora cushmani (MORROW), Keller et al., fig.
10, 5-7, p. 408
Caractères généraux :
L'espèce cushmani est
caractérisée par un contour lobé et par la présence
le plus souvent de quatre à cinq loges au dernier tour à
croissance rapide. Du côté ombilical, les loges sont à
section triangulaire, renflées, pincées latéralement et
sont marquées par des sutures déprimées, montrant des
ouvertures supplémentaires suturales. Du coté spiral, elles sont
à section semi-circulaires et séparées par des sutures en
relief.
Plusieurs auteurs admettent que Rotalipora cushmani
se proche de Rotalipora montsalvensis , dont elle s'en distingue
par un renflement plus important de ses loges, sa carène plus saillante,
ses sutures en relief du côté spiral et un pincement
caractéristique de la portion adomblicale des loges. Selon certains
auteurs (E.W.G.P.F, 1979 ; Bellier, 1983), Rotalipora cushmani
dérive probablement de R. montsalvensis.
Répartition stratigraphique :
L'extension totale de l'espèce définit la zone
à R. cushmani. Elle indique le Cénomanien
supérieur en abondance. Dans le matériel étudié de
la coupe CES, nous avons reconnus la partie supérieure de cette zone
marquée par la sous-zone à D. algeriana. L'espèce
index disparaît dans l'échantillon CES-7. Toutefois, dans la coupe
COK, nous avons reconnus la partie sommitale de la sous-zone à R.
greenhornensis. Dans cette coupe R. cushmani disparaît dans
l'échantillon COK-38.
Espèce : Rotalipora greenhornensis
(MORROW, 1934)
Pl.1, fig.18-20
Synonymie:
1934 - Globorotalia greenhornensis MORROW, p1. 31,
fig.1, p. 199
1946 - Planulina greenhornensis CUSHMAN, pl. 65, figs.
3a-c, p. 159
1948 - Rotalipora globotruncanoides SIGAL, p.100
1956 - Thalmanninella greenhornensis (MORROW),
Brönnimann et Brown, pl. 20, fig.7-9
1959 - Rotalipora (Thalmanninella) greenhornensis KLAUS,
pl. 2, fig. 3a-c, p. 805
1964 - Rotalipora greenhornensis (MORROW), Loeblich et
Tappan, fig. 4a-c,p. c661.
1979 - Rotalipora greenhornensis (MORROW), Chitta, pl.
12, fig. 1-6
1979 - Rotalipora greenhornensis (MORROW), E.W.G.P.F,
fig 1-2; pl. 13, fig 1-2, p.85, 112
1983 - Rotalipora greenhornensis (MORROW),
Gargouri-Razgallah, p.142, pl.24, fig.7
1985 - Rotalipora greenhornensis (MORROW), Caron, fig.
32, 1-2, p. 71
1987 - Rotalipora greenhornensis (MORROW), Ben Haj Ali,
pl. 7, fig. 1-3, 7-9, p. 103
1993 - Rotalipora greenhornensis (MORROW), Caus et al.,
fig. 8, 3a-c, p.542
1993 - Rotalipora greenhornensis (MORROW), Balla et al.,
fig. 5, 1-2, p. 436
1998 - Rotalipora greenhornensis (MORROW), Rami, pl. 9,
fig. 8-9; pl. 10, fig. 1
2001 - Rotalipora greenhornensis (MORROW), Keller et
al., fig. 10, 3-4, p. 408
Caractères généraux :
Rotalipora greenhornensis présente un test
formé par un grand nombre de loges pétaloïdes, la plus part
du temps le dernier tour comporte sept loges, séparées par des
sutures spirales et ombilicales arquées et en relief soulignées
par un bourrelet. Les ouvertures supplémentaires sont intra-ombilicales
et parfois suturales entre les deux dernières loges. En profil,
l'espèce greenhornensis, se distingue par une convexité
plus forte du côté ombilicale. La face spirale a tendance à
s'aplatir. Rotalipora greenhornensis diffère de R. brotzeni
par un profil plus nettement dissymétrique, un nombre de loges au
dernier tour plus élevé (pouvant atteindre 10 loges chez les
formes évoluées) et un ombilic plus large.
128
Répartition stratigraphique:
129
Rotalipora greenhornensis se trouve en association
avec Rotalipora cushmani qui est de même extension
stratigraphique, c'est-à-dire du Cénomanien supérieur.
Elle est très répandue dans le domaine téthysien. Certains
pensent qu'elle aboutit au Turonien inférieur (Saint-Marc, 1974)
Espèce : Rotalipora montsalvensis
(MORNOD, 1949)
Pl.1, fig.3-4
1949 - Globotruncana (Rotalipora) montsalvensis MORNOD,
p. 585, fig. 4,7 1978 - Rotalipora montsalvensis (MORNOD), Wonders, p.
135, pl. 4, fig. 1-2. 1979 - Rotalipora montsalvensis (MORNOD), EWGPF,
p. 95, 98, pl. 15, fig. 1-2. 1994 - Rotalipora montsalvensis (MORNOD),
Premoli Silva et Sliter, pl. 6, fig. 8, pl.8, fig.5 1998 - Rotalipora
montsalvensis (MORNOD), Rami, pl. fig.
2001 - Rotalipora montsalvensis (MORNOD), Keller et al.,
fig.10, 1-2, p.408
2004 - Rotalipora montsalvensis (MORNOD), Keller et
Pardo, pl.1, fig.9 et 11, p.97
Caractères généraux :
Cette espèce diffère de R. cushmani par
des sutures plus renflées sur la face spirale.
Répartition stratigraphique:
R. montsalvensis est connue du Cénomanien
moyen. Toutefois elle disparaît peu avant R. cushmani et R.
greenhornensis. Dans la coupe CES, cette espèce disparaît
au-delà de l'échantillon CES-2 et dans la coupe COK, elle
disparaît au-delà de l'échantillon COK-
SUPERFAMILLE: GLOBOTRUNCANACEA
BROTZEN, 1942 FAMILLE: GLOBOTRUNCANIDAE
BROTZEN, 1942
SOUS-FAMILLE : GLOBOTRUNCANINAE
BROTZEN, 1942 GENRE : Marginotruncana
HOFKER, 1956
Espèce - Type : Rosalina
(=Marginotruncana) marginata REUSS, 1845 Synonymie du genre
:
1956 - Marginotruncana HOFKER, p. 319
Diagnose du genre :
Les représentants de ce genre présentent un test
trochospiralé qui peut être plano-convexe ou biconvexe en ayant un
contour généralement subcirculaire ou lobé. L'ouverture
principale est extraombilicale-ombilicale où l'ombilic est relativement
assez large (1/3 du
130
diamètre maximal du test). Les loges sont
marquées par des ouvertures successives bordées par des portici
de forme triangulaire formant des ouvertures accessoires infralamellaires. En
profil, les loges sont soulignées par deux carènes
généralement séparées par un bandeau
périphérique imperforé. Les sutures sont le plus souvent
sigmoïdes et en relief sur la face ombilicale. Sur la face spirale elles
sont arquées et en relief. Ainsi Marginotruncana se rapproche
de Dicarinella par ses deux carènes et la position de
l'ouverture principale et de Globotruncana par sa double
carène. Elle diffère de la première par la présence
de plusieurs sutures ombilicales sigmoïdes et en relief et de la
deuxième par son système ombilical qui est à portici et
non à tegilla.
Espèce - Type : Marginotruncana renzi
(GANDOLFI, 1942)
Synonymie :
1942 - Globotruncana renzi GANDOLFI, pl. 3, fig. 1
1966 - Globotruncana renzi GANDOLFI, Caron, pl. 4, fig.
5a-c, p. 78
1979 - Marginotruncana renzi (GANDOLFI), EWGPF.2, pl.
69, fig. 1-2
1994 - Marginotruncana renzi (GANDOLFI), Premoli Silva
& Sliter, pl. 13, fig. 1,7
2000 - Marginotruncana renzi (GANDOLFI), Georgescu, p.
172
Caractères généraux :
L'espèce renzi est caractérisée
par une trochospire basse à test biconvexe généralement
symétrique en profil. Le profil est souligné par la
présence de deux carènes étroitement accolées dont
une seule persiste sur les deux dernières loges ou seulement sur la
dernière. Sur la face ombilicale, les loges sont à section
trapézoïdales, allongées dans le sens de l'enroulement et
faiblement bombées séparées par des sutures en forme de U
qui sont soulignées par un bourrelet sigmoïde saillant se
prolongeant au bord de l'ombilic. Sur la face spirale, les sutures sont en
revanche courbes, obliques entre les premières loges et
soulignées par un bourrelet saillant. L'ouverture principale est
extraombilicale-ombilicale où l'ombilic est recouvert par des
portici.
Répartition stratigraphique :
L'espèce renzi apparaît avec
Helvetoglobotruncana helvetica du Turonien inférieur à
moyen. Elle disparaît au Santonien basal. Dans la coupe CES comme la
coupe COK, elle apparit en même temps que H. helvetica.
131
CONCLUSION GÉNÉRALE
Cette étude nous a permis d'aboutir aux conclusions
suivantes : Sur le plan faciès :
1) Les dépôts de l'intervalle du passage
Cénomanien Turonien affleurant dans la région de Jerissa (Oued Es
Smara) sont caractérisés par des marnes et des calcaires argileux
noirâtres, en plaquettes et riches en matière organique (Layeb,
1990), donc typiques de la Formation Bahloul. Ce faciès est fort
semblable à celui de Oued Bahloul (Burollet, 1956). Il est
également semblable à celui de Kalâat Es Senan en Tunisie
centrale (Robaszynski et al., 1990), à celui de Gun Gardens
à Eastbourne en Angleterre là où il est constitué
de marnes, calcaires noirâtres en plaquettes ou noduleux et riches en
matière organique. En outre ce faciès est semblable à
celui de Pueblo située dans le bassin de Colorado là où Il
est formé de deux membres, celui de « Hartland Shale » marneux
et celui de « Bridge Creek Limestone » calcaire. L'ensemble y est
riche en matière organique.
2) Le même intervalle du passage Cénomanien -
Turonien affleurant dans la région de Bargou (Oued El Kharroub) est
plutôt siliceux. Ce faciès est comparable avec le niveau Bonarelli
en Italie.
Sur le plan taxinomique :
Dans les dépôts de l'intervalle Cénomanien
Turonien, nous avons reconnu 33 espèces de Foraminifères
planctoniques se répartissant en 12 genres et 6 familles. Parmi ces
espèces, quatre sont signalées pour la première fois en
Tunisie. Il s'agit de : Guembelitria cenomana (KELLER, 1935),
Guembelitria albertensis STELCK & WALL, 1954 ; Anaticinella
multiloculata (MORROW, 1934) et Globigerinelloides bentonensis
(MORROW, 1934).
132
Sur le plan Biostratigraphie :
En adoptant la méthodologie de la biostratigraphie de
haute résolution, nous avons pu préciser les dépôts
de 3 Zones que nous avons subdivisées en 5 sous-zones :
1) Zone à Rotalipora cushmani du
Cénomanien supérieur avec les deux sous-zones qui sont de la plus
ancienne à la plus récente :
- La Sous-zone à Rotalipora greenhornensis
- La Sous-zone à Dicarinella algeriana
2) Zone à Whiteinella archaeocretacea du passage
Cénomanien Turonien
englobabt de bas en haut
- La sous-zone à Globigerinelloides
bentonensis
- La sous-zone à Dicarinella hagni
- La sous-zone à acmé de Heterohelix
moremani
3) Zone à Helvetoglobotruncana helvetica du
Turonien inférieur à moyen
Les épaisseurs de ces différentes zones et
sous-zone ce sont revélées inégales d'une région
à une autre impliquant des taux de sédimentation et/ou de
subsidence différentielle.
Sur le plan paléoécologie :
Par l'étude paléoécologique que nous
avons accomplie, qui consiste en l'analyse des morphotypes et des
fréquences des différentes espèces de Foraminifères
planctoniques, nous a conduit de mettre en évidence quatre phases
successives dans le comportement des Foraminifères planctoniques en
relation avec le développement de l'événement anoxique du
passage Cénomanien Turonien « EAO-2 »
- Une première phase coincidant avec la partie
supérieure de la zone à Rotalipora cushmani durant
laquelle l'association très diversifiée est de stratégie
K.
133
- Une deuxième phase coincidant avec la partie
terminale de la zone à Rotalipora cushmani durant laquelle les
Rotalipores sont encore présentes mais la faune est dominée par
les Hétérohélicidés. Cela évoque
l'installation de la ZOM.
- Une troisième phase après l'extinction des
Rotalipores. L'association devient de stratégie r ; elle est
dominée par les Guembelitriidés. Ces taxons qualifiés
d'opportunistes atteignent une fréquence de 30 % de l'association.
- Une quatrième phase coincidant avec la zone à
Helvetoglobotruncana helvetica évoquant la tendance des
conditions initiales.
Vu l'intérêt économique de la Formation
Bahloul en tant que Roche mère de pétrole, cette dernière
mérite d'être travaillée sur plusieurs plans et suivant le
modèle de la biostratigraphie de haute résolution. Ceci
permettrait de mieux comprendre le déroulement des différentes
phases ayant abouti aux conditions eutrophiques et anoxiques notamment de la
période du passage Cénomanien-Turonien. De ce fait, une
étude des isotopes stables C13 et O18 serait
bénéfique. En outre, les différents faciès riches
en matière organique de la Formation Bahloul, notamment le faciès
siliceux, devrait être étudié d'avantage et comparé
avec le Livello Bonarelli en Italie sur tous les plans. Une
cartographie numérique en 3D des faciès de l'intervalle
Cénomanien - Turonien et leurs environnements de dépôt
devrait être aussi élaborée.
134
Références Bibliographiques
ABDALLAH H. (1995) - La limite Cénomanien - Turonien en
Tunisie du Centre-Sud : Biostratigraphie et paléoenvironnement.
Résumé p.1. 3ème congrès national
des Sciences de la Terre. Tunis.1995
ABDALLAH H. (2000) - L'événement Anoxique de la
limite Cénomanien - Turonien. Thèse Doc Etat,
Université de Tunis El Manar, Faculté des Sciences de Tunis
pp.11-56 (Annexe).
ACCARIE M., ROBASZYNSKI F., AMEDRO F., CARON M. et ZAGRARNI M.
F. (1999) - Stratigraphie événementielle au passage
Cénomanien - Turonien dans le secteur occidental de la plateforme de
Tunisie centrale (Formation Bahloul, région Kalaat Senen), Annales
des mines et de la géologie N°40 - les septièmes
journées de la géologie tunisienne - Tunis (2-3 Décembre
1999) - pp, 63-80
ADATTE T., STEINMANN P., MORT, H., AFFOLTER S., JACQUAT O.,
PARATTE G., DUCOMMUN P. KELLER G. BERNER Z. and STÜBEN D. (2003) - The
Cenomanian Turonian Anoxic Event In Northern Italy (Southern Alps And Central
Apennines) Correlation with The Penibetic Zone (South Spain) Séance
spécialisée de la Société géologique de
France Paris, 10/11 juillet 2003 Paléocéanographie du
Mésozoïque Mesozoic paleoceanography Volume des
résumés Abstract volume internet
AMEDRO F et ROBASZYNSKI F (1993) - La zone Actinocamax
plenus dans le domaine boréal. Éléments de
comparaison de la limite Cénomanien -Turonien entre le domaine
boréal et téthysien, pp487-494, Cretaceous research, Vol.
14,N°4/5 August/October 1993 - special issue : Cenomanian - Turonian
Boundary Events
ANDERSON D.L. (1994) - Superplumes or supercontinents?
Geology, 22 (1): 39-42. internet
ANDREU B, (1993) - Association d'Ostracodes et
paléoenvironnements au passage Cénomanien - Turonien dans les
régions d'Agadir (Haut Atlas) et de Boulmane (Moyen Atlas),
Maroc,pp409-430, Cretaceous research, Vol. 14,N°4/5 August/October
1993 - special issue : Cenomanian-Turonian Boundary Events
ARTHUR, M. (2003) - The Cretaceous Carbon Cycle: Interpreting
the Carbon Isotope Record. ccod_abstracts. p3. internet.
ARTHUR M. A. and PREMOLI SILVA I. (1982) - Development of
widespread organic carbon-rich strata in the Mediterranean Tethys. In nature
and origin of Cretaceous carbon-rich facies, pp. 7-54. Internet
ARTHUR, M.A., DEAN, W.E., and PRATT, L. M. (1988) -
Geochemical and climatic effects of increased marine organic carbon burial at
the Cenomanaian/Turonian boundary: Nature, 335, 714-717. internet
ARTHUR, M.A., JENKYNS H C. BRUMSACK H. J. and SCHLANGER S. O.
(1990) - Stratigraphy, geochemistry, and paleoceanography of organic
carbon-rich cretaceous sequences. Cretaceous research, Events and Rhythms.
pp.75-119. internet
AUBERT F. (1892) - Explication de la carte géologique
provisoire de la Tunisie. H. Barrère, éd., paris, vol.
n°8, III+91pp.
BAGGA M. A. O., TLATLI M. S. et ABDELJAOUAD S. (1995) - Le
récif Cénomano-Turonien de Borj Cédria, aspects
sédimentologiques diagénétiques et pétrophysiques.
Résumé P.18. 3ème congrès
national des Sciences de la Terre. Tunis.1995
135
BALLA Z and BODROGI I (1993) - The Vékény Marl
Formation of Hungary, pp431-448, Cretaceous research, Vol. 14,N°4/5
August/October 1993 - special issue : Cenomanian-Turonian Boundary
Events.
BARR F. T. (1972) - Cretaceous biostratigraphy and planktonic
Foraminifera of Lybia. Micropaleontology. Vol.18, n°1, pp.1-46
BASSOULLET J. P. et DAMOTTE R. (1971) - Quelques Ostracodes
nouveaux du Cénomano-Turonien de l'Atlas saharien Occidental
(Algérie), Rev, Micropal, Vol. 12. n° 3. pp. 130-144.
BATIK P., DONZE P., GHALI A., MAAMOURI A. L. et MEMMI L.
(1987) - Les dépôts crétacés dans le secteur du
Jebel Trozza. Note service géologique de Tunisie, n°54 pp.
5-24
BAUDIN F., FIET N., COCCIONI R., and GALEOTTI S. (1998) -
Organic matter characterization of the Selli Level (Umbria-Marche Basin,
central Italy): Cretaceous Research, v. 19, p. 701-714.
BÉ A. W. H. (1982) - Biology of planctonic Foraminifera
In: Broadhead T. W. Ed, Foraminifera, notes for a short course. University
of Tennessee. Studies in Geology 6. 51-89, internet
BELLIER J.P. (1983) - Foraminifères planctoniques du
crétacé de Tunisie septentrionale : Systématique,
biozonation utilisation stratigraphique de l'Albien au Maastrichtien,
Thèse Doct, Etat Paris,
BELLIER J.-P., MOULLADE M. and HUBER B.T. (2000) -
Mid-Cretaceous planktonic foraminifers from Blake Nose: revised
biostratigraphic framework. In Kroon, D., Norris, R.D., and Klaus, A.
(Eds.), Proc. ODP, Sci. Results, 171B [Online]. Available from World Wide Web:
<
http://www-odp.tamu.edu/publications/171B_SR/chap_03/chap_03.htm>.
[Cited YYYY-MM-DD] internet.
BEN AYED N. (1986) - Évolution tectonique de l'avant-pays
de la chaine alpine de la Tunisie du début du Mésozoïque
à l'actuel. Thèse d'État Univ. De paris, sud-centre
d'Orsey. pp.1-327
BEN FERJANI A., BUROLLET P.F, MEJRI F. (1990) - Petroleum geology
of Tunisia pp65-135
BEN HAJ ALI N. RAZGALLAH S. BEN HAJ ALI M. et KENNEDY J. W.
(1994) - La Formation Bahloul dans sa localité type, précisions
stratigraphiques basées sur les Ammonites et les Foraminifères
planctoniques. Notes du Serv. Géol. N°60 pp35-68
BEN HAJ ALI N. (1987) - Étude biostratigraphique du
crétacé (Aptien à Cénomanien) de la région
de Téboursouk (Tunisie septentrionale) sur la base des
foraminifères planctoniques. Notes de service géologique de
Tunisie n°54 pp. 75-105
BEN YOUSSEF M. (1981) - Répartition des microfaunes
dans le Crétacé du Jebel Koumine (Tunisie Centrale) Actes du
1er congrès National des Sciences de la Terre (Tunis,
Septembre 1981) n°1, pp, 43-49
BENGSTON P. (1996) - The Turonian stage and substage boundaries.
Bull. Inst. Royal. Sci. Nat. Belgique, Sci. Terre, 66, 69-79.
Internet.
BENZARTI-SAID R. (1981) - Étude biostratigraphique du
crétacé supérieur du sondage Djebiniana 1 (DJB.1) (Sahel
tunisien) et corrélations avec Kerkennah 1 (KK.1) et Reshef 1 (RHF.1)
(Golfe de Gabès) Actes du 1er congrès National des
Sciences de la Terre (Tunis, Septembre 1981) n°1, pp, 51-68
BERTHE (1949) - Stratigraphie du Crétacé moyen et
supérieur de la Tunisie, rapport inédit SEREPT
BISMUTH H, et MAHJOUB M, N, (1981) - Le Crétacé
moyen du Jebel Chambi (Tunisie du Centre-Nord) Aperçu biostratigraphique
et sédimentologique, Actes du 1er congrès National
des Sciences de la Terre (Tunis, Septembre 1981) n°2, pp, 37-54
136
BISMUTH H., BONNEFOUS J.et DUFAURE Ph., (1962)- Mezozoic
microfacies of Tunisia 173 pp. 27p1.
BOLLI H. (1945)- Zur stratigraphie der Obern Kreide in der
hohern helvetischen Decken. Ecl. Gol. Helv., vol. 37, n° 2, pp.
217-228.
BOLTENHAGEN C. (1981) - Les séquences de
sédimentation du Crétacé moyen en Tunisie centrale.
Actes du 1er congrès National des Sciences de la Terre
(Tunis, Septembre 1981) n°2, pp. 55-71
BRALOWER T. J., FULLAGAR, P. D., PAULL C. K. DWYER G. S. and
LECKIE R. M. (1987) - Mid-Cretaceous strontium-isotope stratigraphy of deep-sea
sections. Geol. Soc. Am. Bull., 109:1421-1442. Internet.
BRALOWER, T. J., BROWMAN A. and WRIGHT J. (2003) - Detailed
C-isotope analysis of the Cenomanian-Turonian Boundary Oceanic Anoxic Event and
thepotential link to methane hydrate dissociation. ccod_abstracts. p9.
internet
BROTZEN F. (1934)- Foraminifern aus dem Senon Palastinas.
Zeitschr. deutsch. Verein Palastinas (Leipzig), 57 (1), pp. 28-72, 4
pl.
BROTZEN F. (1936)- Foraminifern aus dem schwedischen untersten
Senon von EriKsdal in Schonen. Sverig Geol. Unders., Ser. C, 396, 30 (3),
206p.,
BUROLLET P.F. DUMESTRE A., KEPPEL D. et SALVADOR A. (1954) -
Unités stratigraphiques en Tunisie centrale. pp. 1 ; abstract.
19ème congrès géol. Alger. Internet.
BUROLLET P.F., DARDEL R., GOTTIS Ch, SAINFELD (1953-1954) -
Carte géologique de Tajerouine n°51 BUROLLET P.F., SAINFELD (1956)
- Carte géologique de Tajerouine n°51 et Notice explicative
BUROLLET P.F. (1956) - Contribution a l'étude
stratigraphique de la Tunisie centrale. Ann. Mines Geol., Tunis, n°
18, 350p. IVpl.
BUROLLET P. F. et ROBASZYNSKI F. (1991) - les
événement de la limite Cénomanien - Turonien en Tunisie
centrale : La Formation Bahloul, Géol alpine, n°67,
pp3-42.
BUSSON G. et CORNÉE A., (1996) -
L'événement océanique anoxique du Cénomanien
supérieur-terminal. Soc. Géol. Nord. Publ. 23. 143
pp.
BUSSON G., DHONDT A., AMEDRO F., NERAUDO D. et CORNÉE
(1999) - La grande transgression du Cénomanien supérieur-Turonien
inférieur sur la Hamada de Tinrhert (Sahara algérien): datations
biostratigraphiques, environments de dépot et comparaison d'un
témoin épicratonique avec les séries contemporaines
à matière organique du Maghreb. Cretac. Res. 20 (1999), pp.
29-46.
BUTT A. (1982) - Micropaleontological bathymetry of the
cretaceous of western Morocco.
Paleogeography, Paleoclimatology, Paleoecology. Vol.37,
pp235-275. internet
CARON M. (1966)- Globotruncanidae du Crétacé
supérieur du synclinal de la Gruyère (Préalpes
médianes, Suisse). Rev. Micropal., vol. 9, n° 2, pp.
68-93.
CARON M. (1972)- Planktonic foraminifera from the upper
Cretaceous of site 98, leg II. Init. Rel). Deep sea Drilling project., Vol.
11, pp.551-559
CARON M. (1985) - Cretaceous planktic foraminifera.
Plankton stratigraphy. Edited by H. BOLLI, I.B. SAUNDERS, K. PERCWIELSEN.
Cambridge University Press. pp 17-79.
CARON M. and HOMEWOOD P., (1983) - Evolution of early
planktonic foraminifers. Marine Micropaleontology 7, 453-462.
CARON M. ROBASZYNSKI F. AMEDRO F. BAUDIN F. DECONINCK J. F.
HOCHULI P. SALIS-PERCH NIELSEN K (von) et TRIBOVILLARD N. (1999) - Estimation
de la durée de l'événement anoxique global au passage
Cénomanien Turonien. Approche cyclostratigraphique dans la Formation
Bahloul en Tunisie centrale. Bull. soc. Géol. Fr.,
CARSEY D. O. (1926)- Foraminifera of the Cretaceous of Central
Texas. Texas univ. Bull., 2612, pp. 1-56.
CASTANY G. (1951d) - Paléogéographie et
orogenèse de la dorsale tunisienne. CR. Acad. Sc., paris,
t.232,1,p.88-90.
CAUS E., GOMEZ-GARRIDO A., SIMÓ A. and SORIANO K.
(1993) - Cenomanian - Turonian platform to basin integrated stratigraphy in the
south Pyrenees (Spain). pp 531 - 552, Cretaceous research, Vol.
14,N°4/5 August/October 1993 - special issue : Cenomanian-Turonian
Boundary Events
CHAABANI F. RAZGALLAH S. DONZE P. et BELAYOUNI H. (1994) -
L'épisode anoxique du passage Cénomanien - Turonien au J. Berda
(Tunisie centro-méridionale). Notes du service Géologique
N°60, pp.21-33.
CHARRÈRE A., ANDREU B., CISZAK R., KENNEDY J. W., ROSSI
A. et VILLA J-M (1998) - La transgression du Cénomanien supérieur
dans la Haute Moulouya et le Moyen Atlas Méridional. Maroc. Geobios
31 5 (1998), pp. 551-569.
CHIKHAOUI M. (2002) - La zone des diapirs en Tunisie : cadre
structural et évolution géodynamique de la sédimentation
méso-cénosoïque et géométrie des corps
triasiques. Thèse d'état université Tunis II.
Faculté des sciences de tunis pp.1-323
CHITTA N. (1979) - Contribution à l'étude
micropaléontologique du Crétacé (Aptien è
Cénomanien) de la region de Teboursouk (Tunisie septentrionale).
Thèse troisième cycle, université paris 6
CLARKE, L. (2003) - Globally synchronous long-term climate
changes during the mid- to Late Cretaceous? ccod_abstracts. p14.
internet
COCCIONI R. (2003) - Cretaceous EAOs : the Italian records
Séance spécialisée de la Société
géologique de France Paris, 10/11 juillet 2003
Paléocéanographie du Mésozoïque Mesozoic
paleoceanography Volume des résumés Abstract volume
internet
COCCIONI R. and LUCIANI V. (2003) - Reading palaeoceanography
from the microfossils : the planktonic Foraminifera across the Selli (EAO-1:
Late early Aptian) and the Bonarelli (EAO-2, Latest Cenomanian) Events.
Séance spécialisée de la Société
géologique de France Paris, 10/11 juillet 2003
Paléocéanographie du Mésozoïque Mesozoic
paleoceanography Volume des résumés Abstract volume
internet
COFFIN M., and ELDHOLM, O. (1994) - Large igneous provinces:
crustal structure, dimensions, and external consequences. Review of
Geophysics, 32, 1-36. internet abstract. pp. 64-79
CRUMIÈRE J-P (1989) - Crise anoxique à la limite
Cénomanien - Turonien dans le sud-est de la France. Relations avec
l'eustatisme. Geobios. Lyon. «Événements de la partie
moyenne du Crétacé» Mém sp. 11. pp.189-203.
internet
CUSHMAN J. A. and TEN DAM A. (1948) - Globigerinelloides
a new genus of Globigerinidae. Contr. Cushm. Lab. Foram. Res., vol.
24, part. 2, p. 42-43, fig. 4-6 pl. 8.
CUSHMAN J. A. (1926) - Some Foraminifera from the Mendez Shale
of Eastern Mexico. Contr. Cushman Lab. Foram. Res., 2(1), pp.
16-26.
CUSHMAN J. A. (1927) - Some new genera of the
Foraminifera. Contr. Cushman Lab. Foram. Res., 4(1),8p.
CUSHMAN J. A. (1933) - Some new Foraminiferal genera.
Contr. Cushm. Lab. Foram. Res., vol. 9, part 2, pp. 32-38, pl. 3-4
76
137
138
CUSHMAN J. A. (1938) - Cretaceous species of Guembelina
and related genera. Contr. Cushman Lab. Foram. Res.;14 (1) pp. 2-28,
1pl.
DALBIEZ F. (1955) - The Genus Globotruncana in Tunisia.
Micropaleontology, vol. n° 2, p. 161-171.
DALBIEZ F. (1956) - Étude sommaire des microfaunes de
la région du Kef. In : Notice explicative de la feuille n°
44, le Kef , carte géologique de la Tunisie au 1/50.000.
DALI - ROSSOT M, D, (1987) - Les calcisphaerulidae des
terrains albiens à maastrichtien de Tunisie Centrale (J, Bireno et J,
Bou El Ahneche) : Intérets systématique, stratigraphique et
paléogéographique, Thèse, 3è cycle,
DAMOTTE R, et GRSDIDIER E, (1963) - Quelques Ostracodes du
Crétacé de la Champagne Humide 1 - Albien -
Cénomanien, Rev, Micropal, Vol, 6, n° 1, pp, 51-66
DELAGE Y. et HEROUARD E. (1896) - Traité de zoologie
concrète. vol. 1, la cellule et Ies protozoaires. Paris: Schleicher
freres.
DE WEVER P., DUÉE G., EL KADIRI K. (1985) . Les
séries stratigraphiques des klippes de Chrafate (Rif septentrional,
Maroc) témoins d'une marge continentale subsidente au cours du
Jurassique Crétacé. Bull. Soc. Geol. Fr. (8), (13), 363-379.
Internet.
DESMARES D., GROSHENY D., BEAUDOIN B. et GARDIN S. (2003) -
Enregistrement à haute résolution des modifications
environnementales inscrites dans un cadre téphrochronologique (bassin de
Western Interior, USA). Séance spécialisée de la
Société géologique de France Paris, 10/11 juillet 2003
Paléocéanographie du Mésozoïque Mesozoic
paleoceanography Volume des résumés Abstract volume
internet
DONZE P., PORTHAULT B., THOMEL G. et DE VILLOUTREYS O. (1970)
- Le sénonien inférieur de Puget Theniers et sa microfaune
Géobios n° 3, fasc.2.
EGGER J. G. (1899) - Foraminiferen und Ostrakoden aus den
Kreide-mergein der oberbayerischen Alpen. Abh. bKönigl. Bayerischen
Akad. Wissensch., Münschen, mathen-naturwiss. Cl., Vol. 21 (1902), pt.1
(1899), p. 3-230, 27pl.
EHERENBERG C. G. (1840) - Uber die Bilding der Kreidefelsen
and des Kreidemergels durch unischtbare Organismen. Abh. Konigi. Preussischen
Akad. Wussensch., Berlin, physik. Kl. (Jahrg. 1838). pp. 59-147.
EHERENBERG C. G. (1843) - Verbreintung und Einfluss des
mikroskopischen Lebens in Süd-und Nord Amerika. Abh. Königl.
Preussischen Akad. Wissensch., Berlin, Physik. Kl. (jahrg. 1838), pp. 59147;
4pl.
EHERENBERG C. G. (1844) - uberzwei ue Larger von Gebirgsmassen
Aus in fusorien als Meeres- Absatz in nord- Amerika and eine vergieichung der
selben mit den organischen kreide-Gebilden in Europa and Africa. Bericht Verh.
konigl. preussischen Akad. Wissensch., Berlin. 57-98.
EICHER D. L. and WORSTELL P. (1970) - Ceno,anian and Turonian
Foraminifera from the Great Plains, United States, micropal. Vol. 16,
N°3. pp. 269-324, pl. 1-13, july 1970, Internet
EICHWALD C. E. (1830)- Zoologia specialis. vol. 2, Vilnae: D. E.
EICHWALD. pp. 1-323.
EL-AMRI Z. (2001) - Les Foraminifères du Coniacien
Supérieur-Maastrichtien inférieur de la région d'Elles
(Tunisie Nord-Occidentale) : Biostratigraphie et Paléoécologie.
DEA. Fac. Sc. de Tunis. pp. 3870.
EL OUERDI H. (1996) - Halocinèse et rôle des
décrochements dans l'évolution géodynamique de la partie
médiane de la zone des dômes (Tunisie septentrionale).
Thèse doctorat. Fac. Sc. Tunis, 242p.
139
ERBACHER J. (1994) - Entwicklung und Palaeoozeanographie
mittelk-retazischer Radiolarien der westlichen Tethys (Italien) und des
Nordatlatiks: Tuebinger Mikropalaeontologische Mitteilungen, vol.12, 120 pp.
ERBACHER J. and THUROW, J. (1997) - Influence of oceanic
anoxic events on the evolution of mid-Cretaceous radiolaria in the North
Atlantic and western Tethys: Marine Micropaleontology, vol. 30, p. 139-158.
ETTACHFINI EL M., SOUHEL A., ANDREU B. et CARON M. (2004) - La
limite Cénomanien Turonien dans le Haut Atlas central, Maroc.
Geobios 38 (2005) 57-68. internet
FERRY S., GROSHENY D. et MALARTRE F., (2003) - Contexte
séquentiel de la crise de la limite Cénomanien - Turonien, marge
subalpine française et autres bassins comparés. Poster.
Internet
FISHER J, K,, PRICE G, D,, HART M, B, and LENG M, (2003) -
Isotopic and foraminiferal analysis of the Cenomanian-Turonian boundary event
of the Indian Ocean, Geophysical Research Abstracts, Vol. 5, 09131, 2003,
Abstract,
FORSTER A., BAAS M. and SINNINGHE DAMSTÉ J. S. (2002) -
Black shale deposition during the Cenomanian - Turonian Oceanic Anoxic Event:
Compound Specific ä13C-Isotope Records from different
key-Location. NAC7 Oceanography internet article
FOURCADE E,, PICCINI L,, ESCRIB A. J, and ROSSELO E, (1999) -
Cretaceous stratigraphy and palaeoenvirenments of the southern Ptén
Basin, Guatemala, Cretaceous Research (1999) 20, pp, 793-811,
FOURNIÉ D. (1978) - Nomenclature lithostratigraphique
des séries du Crétacé supérieur au Tertiaire de
Tunisie. Bull. Centre Rech. Expl. Prod. Elf Aquitaine, Pau, Vol.2,
n°1,pp.97-148.
GALE A. S., JENKYNS H. C., KENNEDY W. J. and CORFIELD R. M.
(1993) - Chemostratigraphy versus biostratigraphy: data from around the
Cenomanian Turonian boundary. Journal of the geological society. Abstract,
Internet
GANDOLFI R. (1955)- A Globotruncana fauna from the Pecan Gap
Chalk of Texas. Micropal.. vol. 1, n°3, pp. 257-259.
GARCÍA A. SEGURA M. CARENAS B. and PÉREZ P.
(1987) - Transgression, discontinuités, eustatisme et tectonique dans le
crétacé moyen du secteur central de la chaïne
Ibérique (Espagne). Mem Geol Univ de Dijon N°11 pp 81-89,
internet
GARGOURI S. (1981) - Mise au point et données nouvelles
sur Thomasinella punica (SCHLUMBERGER), foraminifère
Lituolidé du Cénomanien, répartition stratigraphique et
géographique. Actes du 1er congrés Nat. Des Sc. De la Terre
(Tunis, Septembre 1981). pp209-221
GARGOURI - RAZGALLAH S (1983) - Le Cénomanien de la
Tunisie centrale. Étude paléontologique, stratigraphique,
micropaléontologiques et paléoécologique. Thèse
doctorat d'état, université Claude Bernard Lyon1, 214p,
39pl.
GENDROT C, (1963) - Quelques Foraminifères nouveaux du
Sénonien inférieur des Martigues (Bouches-du-Rhône),
Rev, Micropal, Vol, 6, n° 1, pp, 67-72
GEORGESCU M, (1995) - Upper Cretaceous Heterohelicidae in the
Romanian Black Sea offshore. Revista Española de Micropaleontologia,
27, pp.91-106.
GEORGESCU M, (2000) - Late Albian-Turonian planktonic
foraminifera in the Romanian western black sea offshore, Revista
Española de Micropaleontologia, 32(2), 2000, pp, 157-173,
140
GROSHNEY D. et TRONCHETTI G (1993) - La crise
Cénomanien - Turonien : Réponse comparée des assemblages
de foraminifères benthiques de plate-forme carbonatée et de
bassin dans le SE de la France, pp397-408, Cretaceous research, Vol.
14,N°4/5 August/October 1993 - special issue : Cenomanian-Turonian
Boundary Events
GUIRAND P. (1968) - Étude stratigraphique et tectonique
du secondaire dans la bordure orientale des masiifs tunisiens, Thèse
Univ. Bordeaux, 235p.
HANCOCK J. M. (1993) - Sea level changes around the Cenomanian
- Turonian boundary. pp 553 - 562. Cretaceous research, Vol. 14,N°4/5
August/October 1993 - special issue : Cenomanian-Turonian Boundary
Events
HAQ B U. and HARDENBOL J. (1987) - The new chronostratigraphic
basins of Cenozoic Mesozoic. Sea Level Cycles - Cushman found. Foram
research. Spec. Pub. Vol. 24 pp. 7-13
HAQ B. U., HARDENBOL J. et VAIL P. R., (1987)- Chronology of
fluctuating sea levels, since the Triassic. Science, Washington, pp.
1156-1165. internet
HARDENBOL, J., CARON, M., AMEDRO, F., DUPUIS, C. and
ROBASZYNSKI, F., (1993) - The Cenomanian-Turonian boundary in central Tunisia
in the context of a sequence-stratigraphic interpretation. Cretaceous
Research, n°14, pp, 449-454.
HARDENBOL, J., THIERRY J., FARLEY M. B., JACQUIN T., DE
GRACIANSKY P-C. and VAIL P-R, (1998) - Cretaceous sequence chronostratigraphy.
In: P.-C. De Graciansky, J. Hardenbol, T. Jacquin and P.R. Vail,
Editors, Mesozoic and Cenozoic Sequence Stratigraphy of European BasinsSoc.
Econ. Paleontol. Mineral. Spec. Publ. vol. 60 (1998) Chart 4; Tulsa .
HARRIES P. J. (1993) - Dynamics of Survival following the
Cenomanian - Turonian (Upper Cretaceous) mass extinction event. pp 563 -
584. Cretaceous research, Vol. 14,N°4/5 August/October 1993 -
special issue : Cenomanian-Turonian Boundary Events
HART M.B, DODSWORTH P and DUANE A.M (1993) - The late
Cenomanian Event in eastern England, pp495-509, Cretaceous research, Vol.
14,N°4/5 August/October 1993 - special issue : Cenomanian-Turonian
Boundary Events
HART, M.B. and BAILEY, H.W. (1979) - The distribution of
planktonic foraminiferida in the mid-Cretaceous of NW Europe. In: Wied-man,
J. (Ed.), Aspekte der Kreide Europas. International Union of Geological
Sciences Series A, vol. 6, pp. 527- 542 internet.
HEMLEBEN C. and SPINDLER M. (1983) - Recent advances in
research on living planctonic Foraminifera. Utrecht Micropaleontological
Bulletins 30, 141-170, internet
HERBIN J.P., MONTADERT L., MULLER C., GOMEZ R., THUROW J. and
WIEDMANN J. (1986). Organic-rich sedimentation at the Cenomanian - Turonian
boundary in oceanic and coastal basins in the North Atlantic and Tethys.
Geological Society of London, Special Publication, abstract internet 21,
389-422.
HOFKER. (1956)- Die Globotruncanen von Nord-West-Deutschland
and Holland. Neuues Jb. Geol. u. Pal. Abh., 103(3),pp. 312-340,
26-28.Internet.
HUBER B.T., LECKIE R.M., NORRIS R.D., BRALOWER T.J. and COBABE
E. (1999) - Foraminiferal assemblage and stable isotopic change across the
Cenomanian-Turonian boundary in the subtropical North Atlantic. J.
Foraminiferal Res., 29:392-417. ODP report abstract internet.
IÇAME N. (1994) - Sedimentologie, Stratigraphie
sequentielle et diagenese carbonatée des faciés du
Crétacé moyen du bassin d'Essaouira (Haut Atlas, Maroc).
Thèse, Université de Tunis II, Tunis.
141
JAILLARD E. and ARNAUD-VANNEAU A. (1993) - The Cenomanian -
Turonian transition on the Peruvian margin. pp 585 - 606. Cretaceous
research, Vol. 14,N°4/5 August/October 1993 - special issue :
Cenomanian-Turonian Boundary Events
JALAL M., BLAVOUX B., BAHIR M., BELLION Y., LAFTOUHI N. E.,
PUIG J. M., MENNANI A. et DANIEL M. (2002) - Étude du fonctionnement du
système aquifère karstique cénomano-Turonien de l'Oued
Igrounzar, Bassin d'Essaouira, Maroc. Journal of African Earth sciences vol.32
n°4, pp803-817
JARVIS I., CARSON G., COOPER M., HART M., LEARY P., TOCHER B.,
HORNE D., and ROSENFELD A. (1988) - Microfossil assemblages and the Cenomanian
Turonian (Late Cretaceous) OAE-Cretaceous Research 9, pp.3-103.
JATI M., GROSHENY D., FERRY S., GAUTHIER-LAFAYE F. et
EL-KAMALI N. (2001) - Le passage Cénomanien - Turonien sur la marge
Atlantique (Agadir, Maroc). Abstract, internet. 2p.
JAUSEIN A. (1967) - Contribution à l'étude
géologique des confines de la dorsale Tunisienne (Tunisie
septentrionale). 329p. 69fig. Annale des mines et de la
géologie N° 22
JENKYNS H.C. (1980). Cretaceous anoxic events: from continents
to oceans. Journal of the Geological Society, London, 137, 171-188.
abstract internet
JENKYNS, H.C., GALE, A.S., and CORFIELD, R.M., (1994) - Carbon
and oxygen isotope stratigraphy of the English Chalk and Italian Scaglia and
its palaeoclimatic significance: Geological Magazine, 131, 1-34.
internet
JONES C E and JENKYNS H C (2001) - SEAWATER STRONTIUM
ISOTOPES, OCEANIC ANOXIC EVENTS, AND SEAFLOOR HYDROTHERMAL ACTIVITY IN THE
JURASSIC AND CRETACEOUS [American Journal of Science, Vol. 301, February, 2001,
P. 112-149]
KASSAA S., MIADI F. et BOUHLEL S. (1995) - Sulfures de Zn-Fe
syn-diagénétiques dans les sédiments cénomaniens et
turoniens de Garn Halfeya (Zone des dômes). Résumé
p.103. 3ème congrès national des Sciences de la Terre.
Tunis.1995
KELLER G., LI L. and MACLEOD N. (1995) - the
Cretaceous/Tertiary boundary stratotype sction at El Kef, Tunisia. How
catastrophic was the mass extinction. Palaeogeography, Palaeoclimatology,
Palaeoecology.
KELLER G., HAN Q., ADATTE T. and BURNS S. (2001) -
Palaeoenvironment of the Cenomanian-Turonian transition at Eastbourne, England
Cretaceous Research (2001) 22, 391-422 doi:10.1006/cres.2001.0264,
available online at
http://www.idealibrary.com
on. Internet.
KELLER G and PARDO A (2004a) - Age and paleoenvironment of the
Cenomanian-Turonian global stratotype section and point at Pueblo,
Colorado. Marine Micropaleontology 51 (2004) pp95- 128
KELLER G., STUEBEN D., ZSOLT B. and ADATTE T., (2004) -
Cenomanian-Turonian sea level and salinity variations at Peublo, Colorado.
Palaeogeography, Palaeoclimatology, Palaeoecology.
KHAYATI - AMMAR H. (1996) - Stratigraphie et
micropaléontologie de la série crétacée
inférieur des massifs Serj-Bargou (Tunisie centgre nord). DEA, Fac.
Sc. Tunis.
KLASZ I. (1953) - On the foraminiferal genus Gublerina KIKOINE.
Geol. Bavarica, 17, pp. 245-251, pl. 8.
KLAUS J. (1959) - Le « Complexe schisteux
intermédiaire » dans le synclinal de la Gruyère
(Préalpes médianes). Stratigraphie et Micropaléontologie,
avec étude spéciale des Globotruncanidés de l'Albien, du
Cénomanien et du Turonien. Ecl. Helv., vol. 52, n°2, pp.
753-851.
KUYPERS M. M. M., PANCOST R. D., SINNINGHE DAMSTÉ J.S
(2000) - A large abrupt fall in atmospheric CO2 concentration during Cretaceous
times, Nature . Abstract pp. 342 - 345. Internet
142
KUYPERS M. M. M., RIJPSTRA W. I. C., PANCOST R. D., NIJENHUIS
I. A., SINNINGHE DAMSTÉ J.S (2002) - Orbital forcing of organic carbon
accumulation in the proto-North Atlantic during the Cenomanian Turonian oceanic
anoxic event. Abstract pp. 43 - 64. Internet
KUHNT W., THUROW T., WIEDMANN J. and HERBIN J.P. (1986) -
Oceanic anoxic conditions around C/T boundary at the response of the biota -
Mitt. Geol-Paläont. Int. SCOPE/UNEP. 60, pp205-246.
KUHNT W., HERBIN J.P., THUROW T. and WIEDMANN J. (1990) -
Distribution of Cenomanian-Turonian organic facies in the western Mediterranean
and along the adjacent Atlantic margin. In Depositional of organic facies,
Huc, A. Y. (ed.) AAPG studies in Geology, 30, p. 133-160. internet
abstract.
LAMOLDA M., GOROSTIDI A., MARTINEZ R., LOPEZ G. and PERYT D.
(1997) - Fossil occurrences in the upper Cenomanian - lower Turonian at Ganuza,
northern Spain: an approach to C/T boundary stratigraphy. Cretaceous
Research. Vol18, 3, juin 1997, pp331-353.
LAYEB M, (1990) - Étude géologique,
géochimique et minéralogique, régionale, des faciès
riches en matière organique de la formation Bahloul d'âge
Cénomano-Turonien dans le domaine de la Tunisie Nord-Centrale,
Thèse, Doct, Tunis.
LAYEB M, et BELAYOUNI H, (1989) - La formation Bahloul au
Centre et au Nord de la Tunisie un exemple de bonne Roche mère de
pétrole à fort intérêt pétrolier,
mémoires de l'ETAP, n°3, Actes des II ème
journées de géologie Tunisienne appliquée à la
recherche des hydrocarbures (Tunis, Nov, 1989), pp, 489-503
LAYEB M, et BELAYOUNI H, (1991) - Géologie de la
Formation Bahloul (Cénomanien - Turonien) en Tunisie. Note de l'ass.
Tun. étu. Int. Géol. pp.12-14.
LAYEB M, et BELAYOUNI H, (1999) -
Paléogéographie de la Formation Bahloul (passage
Cénomanien - Turonien), Annales des mines et de la géologie
N°40 - les septièmes journées de la géologie
tunisienne - Tunis (2-3 Décembre 1999) - pp, 21-44
LECKIE R. M, YURITICH R.F., WEST L.O.L., FINKELSTEIN D.,
SCHMIDT M. (1998) - Paleoceanography of the southwestern Interior Sea during
the time of the Cenomanian - Turonian boundary (late Cretaceous). In: Dean,
W.E., Arthur, M.A. (Eds.), Concepts in Sedimentology and Paleontology, vol. 6.
SEPM, USA, pp. 101- 126. internet
LECKIE R. M. BRALOWER T. J. CASHMAN R. (2002) - Oceanic anoxic
events and plankton evolution: Biotic response to tectonic forcing during the
mid-Cretaceous, Paleoceanography, 17, NO. 3, 10.1029/2001PA000623. internet
abstract.
LECKIE R. M. (1984) - Mid Cretaceous Planktonic Foraminiferal
biostratigraphy of Central Morrocco, Deep Sea Drilling Project leg 79,
Sites 545 and 547. initial Reports of the DSDP, 79, 579-620. internet
LECKIE R. M. (1987) - Paleoecology of mid Cretaceous
Planktonic Foraminifera : A comparison of open ocean and epicontinental sea
assemblages. mcropal. Vol. 33, n°2, pp164-176. internet.
LEHMANN R. (1962) - Etude des Globotruncanides du Cretace
superieur de la province de Tarfaya (Maroc occidentale). Notes serv.
Géol. Maroc, t. 21, n° 156, pp. 133-159.
LEHMANN R. (1963)- Résultats d'une étude des
Globotruncanides du Crétacé supérieur de la province de
Tarfaya (Maroc occidentale). Coll. Inter. Micropal., Dakar,
Man. B. R. G. M., N° 32, pp. 113-117.
LIPPS J. H. (1979) - Ecology and Paleoecology of planctonic
Foraminifera. In Lipps et al. (Eds). Foraminiferal ecology and
paleoecology. Society of Paleontologists and Mineralogists. Short course
n° 6 (Houston), 62-104 internet
143
LOEBLICH J. R. and TAPPAN H. (1961) - Cretaceous planktonic
Foraminifera : Part I - Cenomanian. Micropaleontology, vol.7, n°3.
pp.257-304, pl.1-8. internet
LOEBLICH J. R. and TAPPAN H. (1964a) - Protista 2 (Sarcodina:
«Thecamoebians and Foraminiferida») In Moore R. C. Treatise on
invertebrate paleontology. Univ. Kansas Press, Part C : Vol.1/Vol.2 pp.
C1-C900.
LOEBLICH J. R. and TAPPAN H. (1988) - Foraminiferal genera and
their classification. New York : Van Nostrand Reinhold Company, 970 p. 847
pl.
LOEBLICH J. R. (1951) - Coiling in the Heterohelicidae contr.
Cushman Found. Foram. Res. , 2(3), pp. 106-110.
LÜNING S. (2001) - An integrated depositional model for
the Cenomanian-Turonian organic-rich strata in North Africa. North Africa
Research Group GeoNet: Knowledgebase, Contacts, Cooperation. North Africa
Research Workshop 4th-5th September 2001. Abstracts for Talks. Internet
LÜNING S., KOLONIC S, BELHAJ E M, BELHAJ Z, COTA L, BARIC
G and WAGNER T (2004) - An integrated depositional model for the
Cenomanian-Turonian organic-rich strata in North Africa. Earth Science
reviews, Vol 64, Issues 1-2. pp51-117, full text+Link, internet.
MAAMOURI A. L., ZAGHBIB-TURKI D., MATMATI M. F., CHIKHAOUI M.
et SALAJ J. (1994) - La formation Bahloul en Tunisie centro-septentrionale :
variation latérales nouvelle datation et nouvelle interprétation
en terme de stratigraphie séquentielle. Journal of African Earth
Sciences Vol. 18, N°1 , pp. 37-50, 1994.
MACLEOD N, and CARTER J, L, (1984) - A method for obtaining
consistent specimen orientations for use in microfossil biometric studies,
micropalentology, vol, 30, no, 3, pp, 306-310, 1984,
MANIVIT H, (1969) - Nannofossiles calcaires du Turonien et du
Sénonien, Rev Micropal, Vol, 10, n° 4, pp, 277-286,
MARIE J., TROUVÉ P., DESFORGES G., et DUFAURE P. (1984)
- Nouveaux éléments de paléogéographie du
Crétacé de Tunisie. Notes et Mém. N°19 CFP,
pp7-37.
MARTIN-CHIVELET J et GIMÉNEZ R (1993) -
Évolutions sédimentaires et tectoniques des plateformes du SE de
l'Espagne au cours du Cénomanien supérieur - Coniacien
inférieur, pp509-518, Cretaceous research, Vol. 14,N°4/5
August/October 1993 - special issue : Cenomanian-Turonian Boundary
Events
MARTINEZ C., MAHJOUBI H. et BEN SAID H. (1984) - Structure et
évolution de Jebel Chambi (Tunisie centrale) 10ème
RAST, Bordeaux, p.381
MATMATI M. F., MAAMOURI A. L., BEN HAJ ALI M., SALAJ J. et
DONZE P. (1991) - Étude stratigraphique et micropaléontologique
dans le crétacé supérieur et le paléocène de
la chaîne Faid-Chaabet El Attaris (Axe Nord-Sud, Tunisie Centre-est).
Notes de service géologique de Tunise, 1991, n°57 pp.
35-69.
MORNOD L. (1949) - Les Globotruncanidés du
Crétacé Supérieur du Montsalvens (Préalpes
Fribourgeoises). Ecl. Geol. Helv., Vol. 42, pp. 573 -
596
MORROW A. L. (1934) - Foraminifera ans Ostracoda from the
upper cretaceous of Kansas. Journ. Paleont. Vol. 8 N°2 pp. 186 -
205.
MORT H., ADATTE T., STEINMANN P., KELLER G., STUEBEN D.,
BERNER Z. and PARDO A. (2002) - Palaeoenvironmental constraints on productivity
at the Cenomanian - Turonian boundary : an example from the Pueblo stratotype,
Colorado. Internet
144
MORT H., ADATTE T., STEINMANN P., KELLER G., STUEBEN D. and
BERNER Z. (2003) - Productivity and continental weathering in the western
interior during oceanic anoic event 2 ; an example from Pueblo stratotype,
Colorado. Séance spécialisée de la
Société géologique de France Paris, 10/11 juillet 2003
Paléocéanographie du Mésozoïque Mesozoic
paleoceanography Volume des résumés Abstract volume
internet.
MÜLLER C., HERBEN J. P., MONTADERT L., GOMEZ R., THUROW
J., and WIEDMANN J. (1984) - Biostratigraphy of the Organic-rich sedimentation
at the Cenomanian-Turonian boundary in oceanic and coastal basins in the North
Atlantic and Tethys. Geological Society of London, Special Publication, 21,
389-422. ODP report internet
MUSAVU-MOUSSAVOU B., DANELIAN T and COCCIONI R (2003) - A
high-resolution study of radiolarian assemblages from within and around
Bonarelli horizon (Umbria Marches Italy). Séance
spécialisée de la Société géologique de
France Paris, 10/11 juillet 2003 Paléocéanographie du
Mésozoïque Mesozoic paleoceanography Volume des
résumés Abstract volume internet
MURRAY J. W. (1976) - A method of determining proximity of
marginal seas to an ocean Marine geologie. Vol.22, pp103-119.
internet.
M'RABET A,, MEJRI F,, BUROLLET P, F,, MEMMI L, et CHANDOUL H,
(1995) - Cretaceous - Catalog of type sections in Tunisia, ETAP -
mémoire n ° 8A.
NAILI H,, BELHADJ Z,, ROBASZYNSKI F,, CARON M, et DEPUY C,
(1995) - Présence de roche mère "Bahloul" au passage
Cénomanien-Turonien dans la région de Tebessa
(Algérie), Proceeding of
The 4th Tunisian Petroleum Exploration Conference (Tunis, May
1994) pp, 167-168
NEDERBRAGT A. J. (1989a)- Chamber proliferation in the
Cretaceous planktonic foraminiferal Heterohelicidae. j. Foram. Res., 19
(2), pp.105-114.
NEDERBRAGT A. J. (1989b)- Maastrichtian Heterohelicidae
(planctonic Foraminifera) from the NW Atlantic. J. Micropal., 8 (2) pp.
183-206.
NEDERBRAGT A. J. (1990)- Late cretaceous biostratigraphy and
development of Heterohalicidae (plancton foraminifera) in biostratigraphie and
paleooceanographic potential of cretaceous planctik foraminifera
heterohelicidae, central Huisdruk Kerj Vrije universiteit Amesterdam pp.
61-125.
NEDERBRAGT A. J. and FIORENTINO A. (1999) - Stratigraphy and
paleoceanography of the Cenomanian-Turonian Boundary Event in Oued Mellegue,
north-western Tunisia. Cretaceous Research, vol. 20, pp. 47-62.
NEGRA M. H. (1994) - Les dépôts de plate-forme
à bassin de Crétacé supérieur en Tunisie
centro-septentrionale (Fm Abiod et faciès associés)
stratigraphie, sédimentologie, diagenèse et intérêt
pétrolier. Thèse doct. Es-Sc. Univ Tunis II, Fac. Sc. De
Tunis. Pp.1-634.
NEUMANN M. (1967) - Manuel de Micropaléontologie des
Foraminifères (systématique - stratigraphie),
Paris-Gauthier-villars, pp9-273
NZOUSSI - MBASSANI P (2003) - Le Cénomanien - Turonien
de l'Atlantique Nord (Bassin de Sénégal) : environnement de
dépôt et évolution diagénétique. Implication
pétrolière Thèse de Doctorat. pp1-254.
internet
OUERTANI A. (2004) - Étude de la série
sédimentaire des structures et des substances utiles de la région
de Jebel Hout-jerissa. Mémoire de mastère. Univ. De Tunis El
Manar, Fac. De Sc. De Tunis. 199pp.
OLSSON R.,K., HEMLEBEN C., BERGGREN W.A. and HUBER B.T. (1999)
- .Atlas of Paleocene Planktonic Foraminifera. Smithsonian
Contr.Paleobiol.85, Smithonian Institution Press,Washington, DC,252 pp.
internet
145
ORGEVAL J. J., GIOT D., SAHLI R., LENINDRE Y. M., GARBI M. et
MONCIARDINI C. (1986) - Caractérisation des concentrations
sulfurées associées au faciès calcaire « Bahloul
» (Cénomanien supérieur-Turonien basal) du secteur de J.
Bougrine (Gouvernorat du Kef, Tunisie). 148p.
O'DOGHERTY L.. (1994) - Biochronology and paleoecology of
Mid-Cre-taceous radiolarians from Northern Apennines (Italy) and Betic
Cordillera (Spain). Mémoires de Géologie (Lauisianne), v. 21,
415 pp. internet.
PAUL C.R.C., LAMOLDA M.A., MITCHEL S.F., VAZIRI M.R.,
GOROSTIDI A. and MARSHALL J.D. (2000) - The Cenomanian- turonian boundary at
Eastbourne (Sussex, UK): a proposed European reference section.
Palaeogeography, Palaeoclimatology, Palaeoecology 150, 83- 121.
PEDERSON T. F. and CALVERT S. E. (1990) - Anoxia vs
productivity: what controls the formation of organic-carbon-rich sediments and
sedimentary rocks ? Amer. Assoc. Pet. Geol. Bull. N° 74. pp
454-466
PERVINQUIÉRE L. (1903) - Étude géologique de
la Tunisie centrale. Thèse de doc Rud paris. pp 1-360.
PESSAGNO E. A. (1967) - Upper Cretaceous Planktonic Foraminifera
from the western Gulf Coastal Plain. Pal. Amer., Vol. 5, N° 37, pp.
243-244.
PHILLIP J (1981) - Les Formations à rudistes du
Crétacé de Tunise. Une revue. Actes du premier Congrès
National des Sciences de la Terre (Tunis, Septembre 1981).
PHILLIP J (1982) - Paléogéographie des Rudistes
et géodynamique des marges mésogéennes. Bull. Soc. de Fr.
(7T, 24, n°5-6, pp. 995-1006)
PREMOLI SILVA I. and SLITER W. V. (1994) - Cretaceous
planktonic foraminiferal biostratigraphy and evolutionary trends in the
Bottaccione section, Gubbio, Italy. Paleontographia Italica, 82, pp. 189,
Internet
PREMOLI SILVA I. ERBA E. SALVINI G. LOCATELLI C. and VERGA D.
(1999) - Biotic changes in cretaceous oceanic anoxic events of the Tethys.
Journal of Foraminiferal research. Vol.29 n°4, pp.352-370
PRICE G.D. and HART M.B. (2002) - Isotopic evidence for early
to mid-Cretaceous ocean temperature variability. Marine Micropaleon-tology
vol.46, pp45- 58. internet
RAMI A, (1998) - Stratigraphie, Micropaléontologie et
environnement de dépôt du Crétacé supérieur
de la Tunisie centro-septentrionale, Thèse, Doct PP. 1-243
RAMI A,, ZAGHBIB-TURKI D, et EL OUARDI H, (1997) -
Biostratigraphie (foraminifères) et contrôle
tectono-sédimentaire du crétacé
supérieur dans la région de Mejez El Bab (Tunisie
Septentrionale), Géologie
Méditerranéenne, Tome XXIV n° 1-2 1997, pp 101-123
RAZGALLAH S,, PHILIP J,, THOMEL G,, ZAGHBIB-TURKI D,, CHAABANI
F,, BEN HAJ ALI N, et M'RABET A, (1994) - La limite Cénomanien-Turonien
en Tunisie centrale et méridionale : biostratigraphie et
paléoenvironnements, Cretaceous research (1994) 15, 507-533,
ROBASZYNSKI F, CARON M and the European working group of
Planktonic Foraminifera (1979.1) - Description des genres et des
espèces, Cahiers de Micropaléontologie, pp41-181
ROBASZYNSKI F, CARON M and the European working group of
Planktonic Foraminifera (1979.2) - Description des genres et des espèces
(suite), Cahiers de Micropaléontologie, pp13-179
ROBASZYNSKI F, et CARON M, (1995) - Foraminifères
planctoniques du Crétacé : Commentaire de la zonation
Europe-Méditerranée, Bull, Soc, géol, France, 1995, t,
166, n° 6, pp, 681-692, Biostratigraphie mésocenozoïque, Ouest
Europe,
146
ROBASZYNSKI F. and GALE A. S. (1993) - The Cenomanian -
Turonian boundary: a discussion held at final session of the colloquium on
Cenomanian - Turonian events. Grenoble, 26th May 1991 France. pp
607 - 611. Cretaceous research, Vol. 14,N°4/5 August/October 1993 -
special issue : Cenomanian-Turonian Boundary Events
ROBASZYNSKI F., AMEDRO, F. & CARON, M., (1993) - La limite
Cénomanien-Turonien et la Formation Bahloul dans quelques
localités de Tunisie Centrale. Cretaceous Research, n°14, pp,
477-486.
ROBASZYNSKI F., CARON M., DUPUIS C., AMEDRO F.,
GONZALEZ-DONOSO J.M., LINARES-RODRIGUEZ D., HARDENBOL J., GARTNER J., CALANDRA
F. & DELOFFRE R., (1990) - A tentative integrated stratigraphy in the
Turonian of Central Tunisia : Formations, zones and sequential stratigraphy in
the Kalaat Senan area. - Bull. Centres Rech. Explor. Prod. Elf-Aquitaine,
14 / 1, 213-384.
ROBASZYNSKI F., CARON M., GONZALEZ DONSO J. M., WONDER A. A.
H. EDITORS and the European working group of Planktonic Foraminifera (1984) -
Atlas of late Cretaceous Globotruncanids, Revue de
Micropaléontologie vol. 26, n° 3-4, pp. 145-305.
ROTTMAN M. (1978) - Species associations of planctonic
Foraminifera and zooplancton in south China and Java seas. Journal of
planctonic Foraminifera Research. Vol.8, pp350-359. internet
RYAN W.B.F. and CITA M.B. (1977) - Ignorance concerning
episodes of ocean-wide stagnation. Mar. Geol., 23, 197-215. Abstract.
Internet. pp1-2.
SAINT-MARC P. (1974) - Étude stratigraphique et
micropaléontologique de l'Albien, Cénomanien et du Turonien du
Liban. Notes Mém. Moy Orient 13, 342p., 20pl. 58 fig, 4tab
SALAJ J. (1976) - Contribution à la
microbiostratigraphie du Mésozoïque et du Tertiaire de Tunisie
septentrionale. Note de Service géologique, n°42,
pp.29-69.
SALAJ J. (1980) - Microbiostratigraphie du
crétacé et du paléogène de la Tunisie
septentrionale et orientale (hypostratotypes tunisiens). Congrès
géologique international 26ème session - 1er centenaire - Paris
France 1980.
SALVINI G. and MARCUCCI PASSERINI M. (1998) - The Radiolarian
assemblages of « the Bonarelli horizon» in the Umbria Marche
apennines and southern Alps (Italy). Cretaceous Research vol19,
pp.777-804.
SCHIJFSMA E. (1955) - La position stratigraphique de
Globotruncana helvetica BOLLI en Tunisie. Micropal. Vol. 1. pp.
321-334.
SCHLANGER S. O., ARTHUR M A., JENKYNS H C. and SCHOLLE (1987)
- The Cenomanian - Turonian Oceanic Event, I. Stratigraphy and distributions of
organic-rich beds and the marine 13C excursion. Spec. pub. of Geol. Soc. Of
London 26, 371 - 399.
SCHLANGER S. and JENKYNS H C. (1976) - Cretaceaous oceanic
anoxic events: causes and consequences. Abstract internet pp1-2
SCHOLLE P. A. and ARTHUR M. A. (1980) - Carbon isotop
fluctuation in cretaceous pelagic limestones: potential stratigraphic and
petroleum exploration tool. Pet. Geol. Bull. N° 64, pp 67-87
SEGURA M, GARCIA A, GARCIA-HIDALGO F and CARENAS B (1993) -
The Cenomanian-Turonian transgression in the Iberian Ranges (Spain) :
depositional sequences and the location of Cenomanian-Turonian boundary,
pp519-610, Cretaceous research, Vol. 14,N°4/5 August/October 1993 -
special issue : Cenomanian-Turonian Boundary Events
SHACKLETON N.J., HALL M.A. and BOERSMA A. (1994) - Oxygen and
car-bon isotope data from Leg 74 foraminifers. Initial Rep. Deep Sea Drill.
Proj. 74, 599-612. internet
147
SIGAL J. (1977) - Essai se zonation du Crétacé
méditerranéen à l'aide des Foraminifères
planctoniques. Rev. Géol. Médit. Marseille , t, IV, n°2,
49-108, 1tab.
SIGAL J. (1987) - Une échelle zonale du
Crétacé méditerranée et quelques réflexions
suscitées par son établissement, particulièrement à
propos du Danien. Revue Micropal. Vol.30, n°1, pp.32-51.
SMITH A. B., GALE A. and MONKS N. E. A. (2001) - Sea-level
change and rock-record bias in the Cretaceous: a problem for extinction and
biodiversity studies. Paleobiology, 27(2), 2001, pp. 241-253.
internet.
SRARFI D. (1999) - La Formation Bahloul (passage
Cénomanien Turonien) aux Jebels Sekarna et Bargou : stratigraphie et
sédimentologie Tunisie centro sptentrionale. DEA, Fac. Sc.
Tunis
STELCK C. R. and WALL J. H. (1954) - Kaskapau Foraminifera
from Peace River area of Western Canada. Alberta Research Council Report,
N°68, pp. 2-38, Pl. 1-2.
SUBBOTINA N. N. (1949) - Microfauna of the Cretaceous of the
southern slope of the Caucasus (Russian). Leningrad, Vses Neft.
Nauchno-Issled. Geol. Razved Inst. Vnigri. (All-Union Petro. Scient. Res. Geol.
Prosp. Inst). Microfauna of the oil field of the USSR, sbornik 2, Trudy, new
series, vypusk 34, p. 33.
SÜKRÜ YURTSEVER T., KAGAN TEKIN U. and HAKKI DEMIREL
I (2003) - First evidence of the Cenomanian/Turonian boundary event (CTBE) in
the Alakirçay Nappe of the Antalya Nappes, southwes Turkey. Cretaceous
research, 24, pp.41-53, internet
TAPPAN H. (1940) - Foraminifera from Grayson Formation of the
northern Texas. J. Paleon., 14 (2), pp. 93-126, 14 pl.
TARDUNO J.A., BRINKMAN D.B., RENNE P.R., COTTERELL R.D., SCHER
H., and CASTILLO P. (1998) - Evidence for extreme climatic warmth from Late
Cretaceous Arctic vertebrates, Science, 282, 2241-2244. internet
THOMEL G (1993) - Biostratigraphie et faunes d'Ammonites du
Cénomanien et du Turonien inférieur dans le SE de la France,
pp455-476, Cretaceous research, Vol. 14,N°4/5 August/October 1993 -
special issue : Cenomanian-Turonian Boundary Events
THUROW J. and KUHNT W. (1986) - Mid-Cretacrous of the
Gibraltar Arch Area In : Summerhayes C. P. and Shackleton N. J. Eds North
Atlantic Paleoceanography, Geological Society, Special Publication, 21 Oxford
(Blackwell), pp423-445
TLATLI M. S., MAURIN A. et DUFAURE P. H. (1977) - Le complexe
récifal de Potinville, étude sédimentologique. Rapport
inédit TOTAL.
TROUDI H,, NEGRA M, H, et M'RABET A, (1999) - Predictive
evaluation of lower middle Turonian carbonate reservoir in central Tunisia
through integration of deposition environment and diagenesis, Annales des
mines et de la géologie N°40 - les septièmes journées
de la géologie tunisienne - Tunis (2-3 Décembre 1999) pp,
81-100
TURKI M. M. (1975) - Étude géologique du massif
Serj-Bargou (Atlas tunisien central). Thèse troisième cycle,
Univ. Paris VI.
TURKI M. M. (1985) - Polycinématique et contrôle
sédimentaire associé sur la cicatrice de Zaghouan-Nabhena.
Thèse Doct. Es-Science. Fac. De Sc. De Tunis et Rev. Science de Terre
Tunis CST-INRST.7 (1988).
VAN HINTE J. E. (1972) - The Cretaceous time scale and
planctonic foraminiferal zones proceedings. Series B. 75, n°1, pp1-8
148
WHITE M. P. (1928) - Some index Foraminifera of the Tampico
Embayement area of Mexico. Journ. Paleon., vol. 2, n° 3,pp. 177-215;
n° 4, pp.280-317.
WONDERS A. A. H. (1980) - Middle and late Cretaceous planctonic
Foraminifera of western Mediteranean area. Utrech Micropal. Bull.
N°24, 136p.
YASUKOCHI T. and TADA R. (2001) - Sea-level changes across the
Cenomanian/Turonian boundary and their relation with orbitally driven turbidite
cycles in Hokkaido. L023-021 - Internet.
ZAGHBIB-TURKI D., KAROUI-YAAKOUB N., ROCCHIA R., ROBIN E. and
BELAYOUNI H. (2000) - Characteristic events record of the K/T boundary in the
Elles section (Tunisia). Ingenta connect inernet.
ZAIER A. (1999) - Évolution tectono-sédimentaire
du bassin phosphaté du centre-ouest de la Tunisie. Minéralogie,
pétrographie, géochimie et genèse des phosphates.
Thèse d'État, 370p.
ZARGOUNI F. (1985) - Tectonique de l'Atlas méridional
de Tunisie : évolution géométrique et cinématique
des structures en zone de cisaillement. Thèse doctorat es-Sciences
Nat- Inst. De Geol. Univ. de Louis Past. De Str pp.3-304.
ZEBIDI H. (1963) - Contribution à l'étude du
bilan hydrogéologique du Jebel Bargou. Thèse de
3ème cycle, Paris.
ZHAO X, ANTRETTER M., RIISAGER P. and HALL S. (2004) - Rock
magnetic results from Ocean Drilling Program Leg 192: implications for Ontong
Java Plateau emplacement and tectonics of the Pacific. pp. 1-21,
internet
149
Planche 1
1-2 : Praeglobotruncana stephani (GANDOLFI),
Cénomanien supérieur, échantillon CES-2
3-4 : Rotalipora montsalvensis, Cénomanien
supérieur, échantillon CES-1
5-7 : Rotalipora apenninica, Cénomanien moyen,
éch COK-2
8-9 : Whiteinella paradubia, Cénomanien
supérieur, échantillon CES-1
10 : Anaticinella multiloculata, Passage C-T,
échantillon COK-51
11-12 : Globigerinelloides bentonenis, passage C-T,
échantillon COK-41
13 : G. ultramicra, échantillon passage C-T,
CES-46
15 : Helvetoglobotruncana helvetica, Turonien
inférieur, échantillon COK-96
16-17: R. cushmani, Cénomanien supérieur,
échantillon CES-2
18-20 : R greenhornensis, Cénomanien
supérieur, échantillon COK-22
21 : W. archaeocretacea, Cénomanien
supérieur, échantillon COK-36
150
Planche 1

151
Planche 2
1-3 : Guembelitria cenomana (KELLER), échantillon
CES-31
4-6 : Guembelitria albertensis STELCK & WALL,
échantillon CES-31 7-8 : Guembelitria sp, échantillon
CES-42
9 : Guembelitria cretacea (CUSHMAN), Maastrichtien
152
Planche 2

153
Planche 3
1-2 : Heterohelix reussi (CUSHMAN), passage
Cénomanien Turonien, échantillon CES-32
3-4 : Heterohelix navarroensis LOEBLICH, passage
Cénomanien Turonien, échantillon CES-32
5-6 : Heterohelix moremani (CUSHMAN), passage
Cénomanien Turonien, échantillon CES-32
7-8 : Heterohelix pulchra (BROTZEN), passage
Cénomanien Turonien, échantillon CES-32
154
Planche 3

155
Planche 4
1-3 : Hedbergella delrioensis (CARSEY),
Cénomanien supérieur, échantillon COK-26
4-6 : Clavihedbergella simplex (MORROW),
Cénomanien supérieur, échantillon COK-26
6-8 : Whiteinella praehelvetica (TRUJILLO), passage
Cénomanien Turonien, échantillon, CES-56
9 : Whiteinella paradubia (SIGAL), Cénomanien
moyen, échantillon COK-13
10-11 : Whiteinella baltica DOUGLAS & RANKIN,
Cénomanien moyen, échantillon CES-1
156
Planche 4

157
Planche 5
1 : Rotalipora cushmani (MORROW), Cénomanien
moyen échantillon COK-19
2-4 : Dicarinella algeriana (CARON), Cénomanien
moyen échantillon COK-29
5 : Whiteinella aprica (LOEBLICH & TAPPAN),
Cénomanien moyen, échantillon CES-2
6 : Globigerinelloides ultramicra (SOBBOTINA), passage
Cénomanien Turonien, échantillon COK-42
7-8 : Anaticinella multiloculata (MORROW), passage
Cénomanien Turonien, échantillon COK-51
9-10 : Praeglobotruncana gibba KLAUS, Cénomanien
moyen, échantillon CES-2
11-12 : Praeglobotruncana delrioensis (PLUMMER),
Cénomanien moyen échantillon COK-18
158
Planche 5

159
Planche 6
1-2 : Whiteinella aumalensis (SIGAL), Cénomanien
moyen échantillon COK-19
3 : Hedbergella planispira (TAPPAN), Cénomanien
supérieur échantillon COK-44
4-5 : Whiteinella inornata (BOLLI) Cénomanien
supérieur échantillon COK-44
6 : Dicarinella hagni (SCHEIBNEROVA), Turonien
inférieur, échantillon CES-63
Les Radiolaires
7, 8 : Pseudodictyomitra pseudomacrocephala (SQUINABOL),
passage Cénomanien Turonien, échantillon COK-38
160
Planche 6
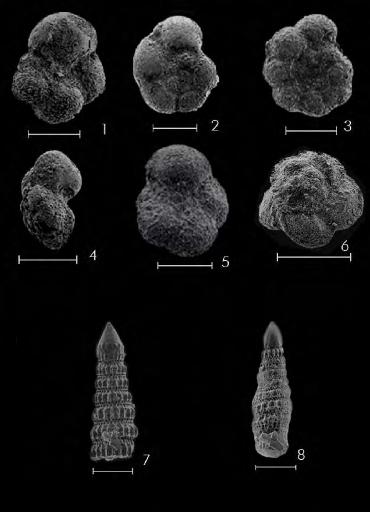
161
Planche 7
1 : Filaments longs et minces à disposition
imbriquée et enchevêtrée enveloppant une vésicule
à remplissage sparitique que nous rapportant à des débris
de coquille de lamellibranche pélagique. (x80), échantillon
COK-28
2 : Globigerinelloides bentonensis en section
équatoriale, associé à une faune benthique et des
calcisphaerulidae (x80), COK-29
3 : Dicarinella algeriana associée à des
calcisphaerulidae (x80), COK-33
4 : Whiteinella archaeocretacea, Hedbergella delrioensis,
Heterohelix sp (x80), COK-29
162
Planche 7

163
Planche 8
1 : Rotalipora cushmani : c et R. greenhornensis
: g (x80), COK-24
2 : Rotalipora greenhornensis (x80), COK-24
3 : Dicarinella algeriana (x80), COK-33
4 : Praeglobotruncana gibba (x80), COK-24
5 : Whiteinella brittonensis (x80), CES-5
6 : Whiteinella aprica(x80), CES-5
7 : W. archaeocretacea COK-29
8 : Globigerinelloides bentonensis (x80), COK-24
164
Planche 8

165
Planche 9
1 - Hedbergella delrionsis (x80) COK-33
2,3 - Clavihedbergella simplex (x80) : 2-
section subspirale, 3- section subombilicale (dans les deux cas le contour du
test est fortement lobé et la dernière loge est allongée
radialement). COK-24
4,5,6 - Guembelitria cenomana : 4 - section subaxiale
(x100) ; 5 - section terminale dans laquelle on observe les trois
dernières loges (x400) ;
6 - section subaxiale (x400). CES-5
7- Faciès bréchique à faune
benthique cassée, à Textularia sp., radiole
d'Echinide (x80), COK-37
8 : Gaudryina sp., (x80), CES-3
166
Planche 9

167
Planche 10
Les Radiolaires
1,2,5 : Sphaerellaria associée à des
filaments et des Calcisphaerulidae (Pithonella ovalis et P.
sphaerica) (x80), COK-34
3,4,6 : Nassellaria associée à des
filaments et des Calcisphaerulidae (Pithonella ovalis et P.
sphaerica) (x80), COK-34
168
Planche 10


169
INDEX
LES FORAMINIFÈRES PLANCTONIQUES
Heterohelix moremani (CUSHMAN, 1938) Heterohelix
navarroensis LOEBLICH, 1951 Heterohelix pulchra (BROTZEN, 1936)
Heterohelix reussi (CUHMAN, 1938) Guembelitria cenomana
(KELLER, 1935) Guembelitria albertensis STELCK et WALL, 1954
Globigerinelloides bentonensis (MORROW, 1934) Globigerinelloides
ultramicra (SUBBOTINA, 1949) Clavihedbergella simplex (MORROW,
1934) Hedbergella delrioensis (CARSEY, 1926) Hedbergella
planispira (TAPPAN, 1940) Whiteinella aprica (LOEBLICH &
TAPPAN, 1961) Whiteinella archaeocretacea PESSAGNO, 1967
Whiteinella aumalensis (SIGAL, 1952) Whiteinella baltica
DOUGLAS & RANKIN, 1969 Whiteinella brittonensis (LOEBLICH
& TAPPAN, 1961) Whiteinella inornata (BOLLI, 1957) Whiteinella
paradubia (SIGAL, 1952) Whiteinella praehelvetica (TRUJILLO,
1960) Praeglobotruncana delrioensis (PLUMMER, 1931)
Praeglobotruncana gibba KLAUS, 1960 Praeglobotruncana stephani
(GANDOLFI, 1942) Dicarinella algeriana (CARON, 1966)
Dicarinella hagni (SCHEIBNEROVA, 1962) Dicarinella imbricata
(MORNOD, 1949) Helvetoglobotruncana helvetica (BOLLI, 1945)
Anaticinella multiloculata (MORROW, 1934) Rotalipora apenninica
(RENZ, 1936) Rotalipora cushmani (MORROW, 1934) Rotalipora
greenhornensis (MORROW, 1934) Rotalipora montsalvensis (MORNOD,
1949) Marginotruncana renzi (GANDOLFI, 1942) Marginotruncana
sigali (REICHEL, 1950)
LES RADIOLAIRES
Novixitus mclaughlini Xitus picenus
Xitus spicularius
Pseudodictyomitra carpatica, Pseudodictyomitra pentacolensis
Pseudodictyomitra pseudomacrocephala Pseudodictyomitra tiara Stichomitra
communis
| 

