|
SOMMAIRE
B. PROBLEMATIQUE ET REVUES BIBLIOGRAPHIQUES 6
C. MATERIEL ET METHODE 11
C.1 MATERIEL ET EQUIPE DU TERRAIN 11
C.2 METHODOLOGIE 11
D. PRESENTATION DE LA ZONE 20
D.1 CADRE BIOGEOGRAPHIQUE 20
D.1.1 LE CLIMAT 22
D.1.2 L'HYDROGRAPHIE 29
D.1.3 LA GEOLOGIE ET LA MORPHOPEDOLOGIE 31
C.1.4 VEGETATION FLORE ET FAUNE 32
D.2 LE CADRE HUMAIN 34
D.2.1 LE PEUPLEMENT 34
D.2.2 LE PROFIL DEMOGRAPHIQUE 35
C.2.3 L'ORGANISATION SOCIOECONOMIQUE 36
D.2.4 PRESENTATION DU VILLAGE DJIRNDA 37
D.2.5 PRESENTATION DU VILLAGE SANGHAKO 39
E. RESULTATS 41
E.1 LES RESULTATS DU SITE DE DJIRNDA 41
E.1.1 Etat des plantations : 41
E.1.2 Les régressions allométriques :
44
E.1.3 Biomasses et Stocks de Carbone : 61
E.1.4 Dynamique du carbone des plantations : 64
E.2 LES RESULTATS DU SITE DE SANGHAKO 65
E.2.1 Etat des plantations : 65
E.2.2 Les régressions allométriques :
67
E.2.3 Stocks de carbone : 70
E.2.4 Dynamique du carbone des plantations : 71
F. DISCUSSIONS 72
F.1 DISCUSSION SUR L'ETAT DES PLANTATIONS : 72
F.2 DISCUSSION SUR LA SEQUESTRATION DE CARBONE ET LES REGRESSIONS
ALLOMETRIQUES : 73
F.3 DISCUSSION SUR LA DYNAMIQUE DU CARBONE DES PLANTATIONS ET LES
PERSPECTIVES DE REBOISEMENT :
74
G. CONCLUSION 76
H. SUGGESTIONS 77
H.1 SUGGESTION SUR LA REALISATION ET LE SUIVI EVALUATION DES
REBOISEMENTS : 77
H.2 SUGGESTION SUR LA SEQUESTRATION DE CARBONE ET RECHERCHE DES
REGRESSIONS ALLOMETRIQUES : 77
H.3 SUGGESTION SUR LA DYNAMIQUE DES PLANTATIONS ET LES
PERSPECTIVES D'AVENIR DES REBOISEMENTS DE MANGROVE : 78
BIBLIOGRAPHIE 79
ANNEXES 82
LISTE DES TABLEAUX
TABLEAU 1 : TABLEAU D'IMPLICATION DES ACTEURS DANS LA GESTION DES
RESSOURCES. 38
TABLEAU 2 : RECAPITULATIF DES DONNEES CARACTERISTIQUES DE L'ETAT
DES PLANTATIONS (SITE DE DJIRNDA).
43
TABLEAU 3: RECAPITULATIF DES CARACTERISTIQUES DES REGRESSIONS
RELATIVES A LA HT (DJIRNDA). 45
TABLEAU 4 :RECAPITULATIF DES EQUATIONS DES REGRESSIONS RELATIVES
A LA HT (DJIRNDA). 46
TABLEAU 5 : RECAPITULATIF DES CARACTERISTIQUES DES REGRESSIONS
RELATIVES AU DMH (DJIRNDA) 50
TABLEAU 6 : RECAPITULATIF DES EQUATIONS DES REGRESSIONS RELATIVES
AU DMH (DJIRNDA) 50
TABLEAU 7 : RECAPITULATIF DES CARACTERISTIQUES DES REGRESSIONS
RELATIVES AU DB (DJIRNDA) 54
TABLEAU 8 : RECAPITULATIF DES EQUATIONS DES REGRESSIONS RELATIVES
AU DB (DJIRNDA) 54
TABLEAU 9 : RECAPITULATIF DES REGRESSIONS ALLOMETRIQUES MULTIPLES
(DJIRNDA). 58
TABLEAU 10 : RECAPITULATIF DES CARACTERISTIQUES DES REGRESSIONS
MULTIPLES (DJIRNDA). 58
TABLEAU 11 : TABLEAU DE BIOMASSE ET DE STOCKS DE CARBONE
(DJIRNDA). 62
TABLEAU 12 : TAUX D'HUMIDITE MOYEN, COEFFICIENT DE PONDERATION
MOYEN PAR PARTIES DES PLANTES
(DJIRNDA). 63
TABLEAU 13 : RECAPITULATIF DES STOCKS DE CARBONE PAR PARTIE DES
PLANTES (DJIRNDA). 64
TABLEAU 14 : TAUX D'HUMIDITE MOYEN ET COEFFICIENT DE PONDERATION
MOYEN (SANGHAKO) 66
TABLEAU 15 : RECAPITULATIF DES DONNEES CARACTERISTIQUES DE L'ETAT
DES PLANTATIONS (SANGHAKO) 67
TABLEAU 16 : RECAPITULATIF DES EQUATIONS DES MODELES LINEAIRES
SIMPLES (SANGHAKO) 68
TABLEAU 17 : CARACTERISTIQUES DES REGRESSIONS LINEAIRES SIMPLES
(SANGHAKO). 68
TABLEAU 18 : TABLEAU DE LA REGRESSION ALLOMETRIQUE MULTIPLE DE LA
PLANTATION 1999 (SANGHAKO). . 69 TABLEAU 19 : RECAPITULATIF DES PARAMETRES DE
L'EQUATION DU MODELE DE LA PLANTATION 1999
(SANGHAKO). 69
TABLEAU 20 : TABLEAU 20 : BIOMASSES ET STOCK DE CARBONE
(SANGHAKO) 71
TABLEAU 21 : TABLEAU 20 : RECAPITULATIF DES STOCKS DE CARBONE PAR
PARTIE DES PLANTES 71
LISTE DES FIGURES
FIGURE 1: SCHEMA DU DISPOSITIF D'INVENTAIRE 13
FIGURE 2 : CARTE DE LOCALISATION DES PLANTATIONS DE DJIRNDA 18
FIGURE 3 : CARTE DE LOCALISATION DES PLANTATIONS DE SANGHAKO.
19
FIGURE 4 : CARTE DE LOCALISATION DE LA RESERVE DE BIOSPHERE
(RBDS). 21
FIGURE 5 : EVOLUTION DE LA TENDANCE GENERALE DE L'INSOLATION
MOYENNE (EN WATTS/H/M2) ANNUELLE (METEO KAOLACK, 2008) 22
FIGURE
6 : EVOLUTION DE LA TENDANCE DES TEMPERATURES MOYENNES ANNUELLES (METEO
KAOLACK, 2008).
23
FIGURE 7 : EVOLUTION DE LA TENDANCE DES ANOMALIES DES
TEMPERATURES MOYENNES ANNUELLES (METEO
KAOLACK, 2008) 24
FIGURE 8 : EVOLUTION DE LA TENDANCE DE LA PLUVIOMETRIE (METEO
KAOLACK, 2008). 25
FIGURE 9 : EVOLUTION DE LA TENDANCE DES INDICES STANDARDISES DE
LA PLUVIOMETRIE (METEO KAOLACK,
2008). 26
FIGURE 10 : EVOLUTION DE LA TENDANCE DE LA PLUVIOMETRIE (METEO
TOUBACOUTA, 2008). 26
FIGURE 11 : EVOLUTION DE LA TENDANCE DES INDICES STANDARDISES DE
LA PLUVIOMETRIE (METEO TOUBACOUTA, 2008). 27
FIGURE 12 : EVOLUTION DE LA
TENDANCE DE L'HUMIDITE DIURNE MOYENNE ANNUELLE (METEO KAOLACK,
2008). 28
FIGURE 13 : EVOLUTION DE LA TENDANCE DE L'HUMIDITE NOCTURNE
MOYENNE ANNUELLE (METEO KAOLACK,
2008). 28
FIGURE 14 : EVOLUTION DE LA TENDANCE DE L'HUMIDITE MOYENNE
ANNUELLE (METEO KAOLACK, 2008). 29
FIGURE 15 : SECHAGE DE POISSONS AU SOLEIL. FIGURE 16 : FOUR
TRADITIONNEL LE FUMAGE DE
POISSONS 39
FIGURE 17 : BOIS DE MANGROVE. FIGURE 18 : BOVIN DU VILLAGE DE
DJIRNDA AU
PATURAGE 39
FIGURE 19 : NID D'OISEAU (PLANTATION 2003 50/50). FIGURE 20 :
PROPAGULES (PLANTATION 2003
25/25). 42
FIGURE 21 : TAILLE DES PLANTS (PLANTATION 2003 25/25). FIGURE 22
: TRANSECT/PLACETTE
(PLANTATION 2006 25/25). 42
FIGURE 23 : TAILLE DES PLANTS (PLANTATION 2005 50/50) 43
FIGURE 24 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2003 ECARTEMENT 25/25. 46
FIGURE 25 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2003 ECARTEMENT 50/50. 47
FIGURE 26 : NUAGE DE POINT ET DROITE DE REGRESSION DE LA
PLANTATION 2004 ECARTEMENT 25/25. 47
FIGURE 27 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2004 ECARTEMENT 50/50. 48
FIGURE 28 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2005 ECARTEMENT 50/50. 48
FIGURE 29 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2006 ECARTEMENT 25/25. 49
FIGURE 30: NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2003 ECARTEMENT 25/25. 50
FIGURE 31 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2003 ECARTEMENT 50/50. 51
FIGURE 32 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2004 ECARTEMENT 25/25. 51
FIGURE 33 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2004 ECARTEMENT 50/50. 52
FIGURE 34 : NUAGE DE POINT ET DROITE DE REGRESSION DE LA
PLANTATION 2005 ECARTEMENT 50/50. 52
FIGURE 35 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2006 ECARTEMENT 25/25. 53
FIGURE 36 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2003 ECARTEMENT 25/25. 54
FIGURE 37 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2003 ECARTEMENT 50/50. 55
FIGURE 38 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2004 ECARTEMENT 25/25. 55
FIGURE 39 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2004 ECARTEMENT 50/50. 56
FIGURE 40 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2005 ECARTEMENT 50/50. 56
FIGURE 41 : NUAGE DE POINTS ET DROITE DE REGRESSION DE LA
PLANTATION 2006 ECARTEMENT 25/25. 57
FIGURE 42 : RESIDUS CENTRES REDUITS DE LA PLANTATION 2003
ECARTEMENT 25/25 59
FIGURE 43 : RESIDUS CENTRES REDUITS DE LA PLANTATION 2003
ECARTEMENT 50/50 59
FIGURE 44 : RESIDUS CENTRES REDUITS DE LA PLANTATION 2004
ECARTEMENT 25/25 60
FIGURE 45 : RESIDUS CENTRES REDUITS DE LA PLANTATION 2004
ECARTEMENT 50/50 60
FIGURE 46 : RESIDUS CENTRES REDUITS DE LA PLANTATION 2005
ECARTEMENT 50/50 61
FIGURE 47 : RESIDUS CENTRES REDUITS DE LA PLANTATION 2006
ECARTEMENT 25/25 61
FIGURE 48 : LIGNE DE TRANSECT (PLANTATION 2000). FIGURE 49 :
LIGNE DE TRANSECT (PLANTATION
1999). 66
FIGURE 50 : RESIDUS CENTRES REDUITS PLANTATION 1999 (SANGHAKO)
70
LISTE DES EQUATIONS
ÉQUATION 1 : COEFFICIENT DE PONDERATION (RO). 16
ÉQUATION 2 : TAUX D'HUMIDITE (HO) 16
ÉQUATION 3 : STOCK DE CARBONE (C). 16


A. INTRODUCTION
Le climat mondial n'a jamais cessé de changer. Certains
de ces changements ont des causes naturelles mais d'autres peuvent être
imputés à des activités humaines telles que le
déboisement et les émissions atmosphériques dues, par
exemples, à l'industrie et aux se moi transports, qui ont conduit
à l'accumulation de gaz et d'aérosols dans l'atmosphère
[UNFCCC, 2008]. Ces gaz dits gaz à effet de serre (GES)
piègent la chaleur et provoquent une élévation des
températures de l'air.
Le Groupe d'Experts Intergouvernemental sur l'Evolution du
Climat (GIEC, 2001) a signalé dans son troisième
rapport sur l'état du climat mondial que les observations et mesures sur
l'évolution des températures atmosphériques montraient
que, globalement, la planète était en train de se
réchauffer et ferraient également apparaître d'autres
modifications du système climatique. Selon ce rapport, les années
1990 ont très probablement été la décennie la plus
chaude à l'échelle mondiale et 1998 l'année la plus chaude
depuis que le réchauffement a été constaté au cours
des dernières décennies. En plus, de nouvelles analyses de
données indirectes pour l'hémisphère Nord indiquent que le
20ème siècle a probablement été le
siècle le plus chaud du dernier millénaire et, en outre, de
nouvelles preuves plus concluantes permettent de dire que la majeure partie du
réchauffement observé au cours des cinquante années
écoulées est due aux activités humaines. Selon ce
même rapport, l'influence humaine continuera de modifier la composition
de l'atmosphère tout au long du 21ème
siècle.
Le réchauffement de la planète a de nombreuses
conséquences sur les ressources forestières, le niveau des mers,
la couverture neigeuse, les nappes glaciaires et les précipitations
[UNFCCC, 2008], ce qui n'est pas sans conséquence pour
les mangroves de l'estuaire du Saloum caractérisées par une forte
vulnérabilité à l'élévation du niveau des
océans. Selon cette même source, la modification des
régimes climatiques régionaux, marquée par une
élévation des températures de l'air, a déjà
des effets sur les bassins versants et les écosystèmes
(forêts, prairies, pâturages et les zones littorales) dans de
nombreuses régions du monde. Pour les économies nationales, le
coût des mesures à prendre pour faire face aux
phénomènes climatiques extrêmes, aux mauvaises
récoltes et aux autres situations d'urgence relatives au climat ne cesse
d'augmenter.
Les pays à faible revenu et les ménages pauvres
des pays en développement sont tout particulièrement
vulnérables face aux effets néfastes des changements climatiques,
qui viennent s'ajouter aux difficultés créées par la
pauvreté. Le cas de nombreux petits Etats insulaires en
développement exposés à une élévation du
niveau des mers à l'exemple des îles de l'estuaire du Saloum pose
le problème de survie des habitants de ces zones. C'est leur
existence même en tant que zone habitable qui est
menacée notamment par l'élévation du niveau des mers et
l'augmentation de l'érosion côtière qui modifie tous leurs
paysages littoraux.
Réduire les émissions de GES de façon
à ce que leur concentration atmosphérique puisse se stabiliser
à un niveau convenu permettrait d'après le GIEC notamment, de
retarder et d'atténuer les effets néfastes des changements
climatiques sur les systèmes naturels et le développement des
sociétés humaines. Mais même dans ce cas, ces effets
continueront de se faire sentir longtemps après la mise en oeuvre de
mesures de réduction ou d'atténuation des émissions de
GES. Ils devraient également varier d'un pays à l'autre et
à l'intérieur d'un même pays. Les mesures visant à
aider les sociétés et les économies les plus
vulnérables à s'adapter aux effets néfastes (y compris,
dans certains cas, aux conséquences économiques des mesures
envisagées pour réduire les émissions) ou à se
prémunir contre ceux-ci sont considérées comme un
complément essentiel par rapport à la tâche globale
consistant à stabiliser les concentrations de GES dans
l'atmosphère. Pour ce faire, le GIEC a envisagé plusieurs mesures
à travers le Protocole de Kyoto (PK, 1997) parmi
celles-ci le changement d'affectation et d'utilisation des terres, la
reforestation, le développement et l'utilisation des technologies
propres ainsi que leur transfert dans les pays en développement.
Les zones côtières du Sénégal
à l'instar des zones côtières du monde entier et des pays
insulaires constituent les zones les plus exposées aux effets des
changements, de par leur exposition à la remontée du niveau des
mers, aux ouragans et aux cyclones, et à la perturbation de la
pluviométrie. C'est ainsi que plusieurs mesures d'adaptation ont
été prises par le gouvernement en collaboration avec des
organismes nationaux et internationaux à l'exemple de l'UICN, WAAME,
OCEANIUM et PAGEMAS entre autres mesures l'érection en 1981 du Parc
National du Delta du Saloum (PNDS) créé à partir de 1976
en Réserve de Biosphère du Delta du Saloum (RBDS),
L'élaboration du premier plan de gestion des ressources de la RBDS en
1997, l'érection en 2004 d'une partie de la mangrove de Bamboung par
l'OCEANIUM en Aire Marine Protégée (AMP) afin de permettre la
préservation de la mangrove et le retour des espèces de poissons
disparus ou en voie de disparition. En même temps, des institutions comme
l'UICN, WAAME, PAGEMAS et autres ont reboisé depuis 1996 des centaines
d'hectare de terres dénudées (tannes) afin de lutter contre la
dégradation des mangroves du Sénégal en
général et du Delta du Saloum en particulier.
B. PROBLEMATIQUE ET REVUES BIBLIOGRAPHIQUES
La mangrove est définie par Blasco
(1991) et cité par Véga (2000) comme une
forêt de palétuviers se développant dans la zone de
balancement des marées des régions littorales intertropicales,
pouvant se maintenir localement jusqu'à 32° Nord et 28° Sud
sous l'effet de courants marins chauds. Selon Véga
(2000), Spalding (1997) estime qu'elle colonise dans ces
régions près de 75% des côtes et deltas, avec une surface
pouvant être évaluée à 182.000 km2.
Tomlinson (1986) cité par Véga
(2000) répartit la mangrove en deux zones biogéographiques
distinctes et caractérisées par une biodiversité
végétale particulièrement faible. L'aire orientale ou
pacifique inclut la mangrove des côtes est-africaines jusqu'à
celle de l'Asie et de l'Australie [Véga, 2000]. Elle
compte environ soixante (60) espèces arborées strictement
inféodées à ce milieu. L'aire occidentale ou atlantique
dans laquelle se situe le delta du Saloum comprend selon
Véga et al. (2000), des espèces de mangrove des
côtes atlantique et de la côte pacifique américaine. Sa
richesse spécifique est moindre puisqu'elle ne regroupe que sept (07)
espèces arborées que sont : Avicennia
germinans et A. schaueriana
(Avicenniaceae), Rhizophora
mangle, R. racemosa et R.
harissonni (Rhizophoraceae)
Laguncularia racemosa et Conocarpus
erectus (Combretaceae)
[Véga, 2000].
Le développement de la mangrove est tributaire de la
température de l'air (température moyenne du mois le plus froid
supérieure à 16°C) et la présence de zones littorales
protégées et peu profondes [Arnaud, 1997 ;
Lee, 1999]. Pool (1977) et
Amarasinghe (1992) cités par
Véga (2000) ont montré que la position des
écotones entre milieu marin et terrestre de l'écosystème
mangrove fait qu'il est soumis à de nombreux facteurs limitant tels que
: la durée d'exondation et d'inondation, la salinité des eaux et
l'hydromorphie du substrat. Dans ces conditions de vie difficiles, seules les
espèces ayant développé des mécanismes d'adaptation
divers (racines échasses, pneumatophores, mécanismes de
régulation du sel) ont pu subsister [Cantera et
Arnaud, 1997] cités par Véga
(2000).
La mangrove était considérée autrefois
comme un milieu hostile et inutile, ce qui était à la cause de sa
dégradation et sa destruction sous l'effet de l'anthropisation
[Ramirez-Garcia et al. 1998] cité par
Véga (2000) mais des travaux récents ont
montré son intérêt tant au niveau socioéconomique,
de par sa grande production de bois et l'abondance de sa faune
(crustacés, poissons etc.) [Doyen et al. 1985 ;
Ndour, 2005] qu'au niveau écologique, pour son
rôle important dans les chaînes trophiques côtières,
dans la protection des côtes contre l'érosion marine, et pour sa
sensibilité aux variations du niveau des mers [Doyen et
al. 1985 et Ndour, 2005]. DIOUF (1996)
atteste que la richesse trophique de l'estuaire du Sine-Saloum est issue
principalement de la mangrove qui joue le rôle de zone de frayère,
de nourricerie, d'habitat,


de reproduction et de la promotion du développement des
poissons. Il a affirmé en même temps que la présence d'une
mangrove luxuriante favorise une richesse spécifique et des effectifs
élevés des poissons de par son rôle d'enrichissement
trophique des milieux, de la protection contre la prédation et la
diversification des types d'habitats.
De nos jours, beaucoup de programmes de recherche
s'intéressent à l'écosystème mangrove, que ce soit
en vue de sa restauration et de sa conservation ou bien de façon plus
globale, pour la compréhension de son fonctionnement tant au niveau
national qu'international.
Au Sénégal, le delta du Saloum constitue la
région la plus septentrionale occupée par une haute mangrove dans
l'ouest de l'Afrique [Dupuy et Verschuren,
1982] cités par Ndour (2005). Giffard
(1974) cité par Ndour (2005), confirme la disparition
des reliques de la végétation de mangrove signalée par
Chaudreau (1916) en Mauritanie mais, qui sont encore
signalés dans le delta du fleuve Sénégal où
Chambonneau avait noté en 1677 la présence d'une
épaisse et haute mangrove [Chamard et al. 1999].
Trochain (1940) a par ailleurs témoigné de la
disparition de la végétation de mangrove des cuvettes du N'diael,
par la découverte de pneumatophores subfossiles, confirmant ainsi
l'existence de mangrove notée par Chambonneau sur le
fleuve Sénégal [Ndour, 2005].
Les autres localités du Sénégal où
l'on rencontre encore la mangrove sont : la Somone, JoalFadiouth, l'estuaire du
Saloum et le fleuve Casamance [Ndour, 2005]. Les formations de
mangrove y sont dominées par Rhizophora
harissonii, R. mangle et R.
racemosa (Rhizophoraceae)
[Marius, 1981 ; Diop, 1986] et
Avicennia africana
(Verbénaceae). A ces deux familles qui
constituent l'essentiel des formations de mangrove s'ajoute celle des
Combrétaceae représentée par
Conocarpus erectus et Laguncularia
racemosa [Ndour, 2005]. Il apparaît ainsi
que les mangroves du delta du Saloum, à l'instar des mangroves de
l'Afrique de l'ouest sont caractérisées par une pauvreté
floristique par rapport aux mangroves de l'Afrique orientale
[Ndour, 2005].
Les espèces du genre
rhizophora et Avicennia
sont principalement rencontrées en peuplements purs dans la zone du
delta du Saloum, mais l'étendue de ces peuplements diminue de plus en
plus sous l'effet des facteurs naturels (déficit pluviométrique,
érosion, salinité, attaque par des parasites) et anthropiques
(coupes de bois, construction des routes et des barrages, pression
foncière etc.) [Ndour, 2005 ; JICA,
2004 ; UICN, 2006]. Soumaré et al.
(1992) atteste d'une diminution des surfaces de mangrove sur la base d'une
comparaison d'images satellitaires de 1976 à 1991. Cette
régression de la mangrove est liée à la salinisation et
à l'acidification des sols favorisés par le déficit
pluviométrique [Bovin et al. 1985]. Il s'ajoute
à ces principales causes de dégradation, la perturbation des
régimes hydriques
responsables de la submersion [Diop et al.
1989] et de la sédimentation [Marius et al. 1992]. Mais
aussi, la rupture de la flèche de Sangomar qui protégeait la
mangrove de la zone du delta du Saloum des perturbations mécaniques de
la houle constitue une cause relativement importante de la régression
des espaces de mangrove du Sénégal [Ndour,
2005]. Cependant, bien que la végétation de mangrove
régresse en certains endroits, une régénération
naturelle a été signalée par divers auteurs tels que :
Marius (1984), Doyen et al. (1985),
Diop et al. (1997), et Ndour (2005).
Balla Dieye (2007) a démontré d'une
régénération de 4,15 à 10,6% pour une
régression de 8.5% par étude d'images satellitaires Landsat
(1972, 1979 et 1999) et Spot (1986, 1990, 1997, 2001 et 2003). Il a
localisé cette régénération sur les bordures des
mangroves dans les tannes humides alors que la régression est
concentrée dans la périphérie nord-ouest de l'estuaire du
Saloum.
La prise de conscience de la régression des espaces de
l'écosystème mangrove du Sénégal en
général et du delta du Saloum en particulier, s'est
matérialisée au cours de ces deux dernières
décennies par diverses actions de reboisement. Kaly,
(2004) atteste que les premiers essais de reboisement réussis et connus
remontent en 1994 à Somone, avant de s'étendre dans le delta du
Saloum les années suivantes mais seulement sur une seule espèce
de palétuviers (le Rhizophora mangle).
Le reboisement de la mangrove du delta du Saloum peut
être scindé en deux catégories suivant les phases de
réalisation. La première catégorie est celle de la
première phase qualifiée d'expérimentale et
caractérisée par un manque de technicité, une mauvaise
maîtrise de la qualité du substrat, la promotion d'une seule
espèce de palétuviers avec des taux de réussite allant de
faibles à moyens pour la plupart des plantations
[Diedhiou, UICN, 2006]. La deuxième
catégorie est celle correspondant à la deuxième phase de
réalisation marquée par une bonne acquisition
d'expériences en terme de technicité et de maîtrise de la
qualité des substrats ainsi que la diversification des espèces
caractérisée par l'introduction d'Avicennia
avec des taux de réussite généralement
appréciables [Diedhiou, UICN,
2006].
JICA/JAFTA (2005) a classé pour sa
part les reboisements de mangrove du delta du Saloum en quatre (04)
catégories en fonction des moyens utilisés à la
réalisation de la plantation. Elle distingue : des plantations
réalisées à travers des campagnes des services
administratifs (Eaux et Forêts/Parcs Nationaux) principalement dans les
régions insulaires ; des plantations réalisées à
l'initiative des populations à l'exemple du groupe des femmes de Djirnda
; des plantations réalisées sous la direction des ONG à
l'exemple de FIOD qui a incité le reboisement de 240 ha dans trois (03)
villages de 1997 à 2001 par apport d'une importante
aide en nature ; et enfin des plantations
réalisées avec l'aide des Organismes comme UICN, WAAME, OCEANIUM
et CAREM depuis 1995.
Rien que pour la période de 1995 à 2001,
JICA/JAFTA (2005) a dénombré en total 420 ha de
terres reboisées par les ONG (UICN, WAAME et CAREM) dans 49 villages du
delta du Saloum. Cependant, il est à noter que les actions de ces ONG
ont été pour la plupart concrétisées grâce
à l'appui des organisations villageoises à l'exemple des
Comités de Plage créés par l'UICN à partir de la
deuxième moitié des années 1990 dans les régions
insulaires, et, les Comités Villageois pour la Réhabilitation de
la Mangrove (CVRM) créés à partir du milieu des
années 1990 par WAAME [JICA/JAFTA, 2005]. Ces CVRM sont
principalement implantés dans 35 villages du bassin du Saloum. Ces
plantations réalisées en guise de réponse aux effets des
changements climatiques ont fait l'objet de plusieurs travaux d'étude
d'impact tant au plan écologique que socioéconomique
commandités par divers institutions (UICN, OCEANIUM, JICA, WAAME etc.).
Cependant, leurs impacts en termes de séquestration de carbone
atmosphérique n'ont jamais été évalués.
Le présent rapport porte sur le suivi évaluation
de la séquestration de carbone atmosphérique par les plantations
de mangrove de l'UICN. Il assure la continuité de la recherche
d'informations sur l'utilité sociale, écologique et
environnementale des reboisements de mangrove dans la RBDS. Une première
étude réalisée par Diédhiou,
UICN (2005) a permis de montrer au plan des impacts que
quelques années après la réalisation de ces plantations,
les impacts écologique et socioéconomique sont nombreux comme en
témoigne la réapparition des espèces jadis disparues ou en
voie de disparition comme les Tympanotonus des
vasières « paco-paco », les crevettes, les huîtres et,
le développement des activités génératrice de
revenus comme l'apiculture, la cueillette des crevettes, des harches.
Cependant, en rapport avec le protocole de Kyoto
(PK, 1997), aux recommandations de Bali
(2008) et à l'importance des forêts dans la séquestration
du carbone, les impacts des plantations de mangrove du delta du Saloum en
termes de séquestration de carbone atmosphérique reste encore
à évaluer. C'est dans cette optique que, l'UICN (Bureau national
du Sénégal) s'est engagée dans l'évaluation de la
quantité de carbone stockée par ces dites plantations de
mangrove.
En somme, la confirmation de l'hypothèse d'une forte
séquestration de carbone par les plantations de mangrove, pourrait aider
à affiner et varier l'argumentaire des programmes de restauration et de
conservation des zones de mangrove. Par ailleurs, la valorisation des
informations devrait aussi être une source supplémentaire de
motivation des populations dans la restauration des formations de mangrove. A
ces deux aspects incitatifs, il s'ajoute un besoin
de démontrer la pertinence des plantations de mangrove
par rapport à la problématique d'adaptation aux changements
climatiques. C'est dans cette dynamique, qu'un contrat de stage a
été signé entre l'UICN et l'ISE pour la conduite de cette
étude. Les principaux objectifs de cette étude sont :
o la Connaissance de la dynamique des reboisements de
mangrove,
o l'Estimation de carbone de la biomasse sur pieds et
recherche des régressions aiométriques correspondantes
enfin,
o la Recherche des intérêts de
l'élaboration des projets de séquestration de carbone par le
biais des reboisements de mangrove.
L'aboutissement de ces différents objectifs d'étude
nécessite l'utilisation d'un certain nombre de méthodes et de
matériels de mesure et de calcul mais aussi de pratiques
appropriées.
C. MATERIEL ET METHODE
Le matériel utilisé pour cette étude est
constitué d'instruments de mesure, d'outils pour la coupe rase et la
pesée des biomasses de la mangrove, de pirogue etc. La
méthodologie de l'étude part de la recherche bibliographique qui
a permise l'élaboration du protocole de recherche, au choix de la
méthode de collecte des informations pour finir par le choix des outils
et méthodes de traitement et d'interprétation des
données.
C.1 MATERIEL ET EQUIPE DU TERRAIN
o 1 GPS (global positioning system) qui a permis
de relever les coordonnées géographiques des reboisements ;
o Des cordes pour matérialiser les
limites des parcelles et des transects ;
o Jalons utilisés pour délimiter
les axes de transect. Ils sont fixés aux deux extrémités
des axes de transect ;
o 1 décamètre pour la mesure de
l'écart inter placettes de cinq (05) à dix (10) mètres
selon la taille de la plantation ;
o 1 sécateur pour la coupe rase des
sujets retenus pour la mesure de biomasses ; o Des piquets en
bois nécessaires pour la délimitation des placettes de 1m×1m
; o 1 appareil photo pour illustrer les observations faites
sur le terrain ;
o 1 pied à coulis nécessaire pour
la mesure des diamètres des plants rencontrés dans les placettes
;
o 1 bâton gradué pour mesurer la
hauteur des plants rencontrés dans les placettes ;
o 1 balance électronique de
précision pour la pesée des biomasses issues des feuilles, des
branches et des tiges ;
o Des sachets pour le prélèvement
et la conservation des échantillons de biomasses ;
o 1 étuve pour le séchage des
échantillons de biomasses en vue de la déduction des taux
d'humidité et des coefficients de pondération ;
o (01) manoeuvre pour aider aux tracés
des transects et la réalisation des différentes mesures.
C.2 METHODOLOGIE
La méthodologie de recherche utilisée dans cette
étude part d'une recherche bibliographique
qui consiste à
rassembler les documentations possibles traitant des techniques
d'inventaire
forestier, de la mangrove, de la séquestration de
carbone atmosphérique par les plantes, des
caractéristiques biophysiques et climatiques du delta
du Saloum, des reboisements de mangrove et de leurs acteurs dans le bassin du
Saloum. Cette recherche a permis l'élaboration du protocole de recherche
qui définit le plan de sondage, les différentes méthodes
de collectes de données et les méthodes et outils de traitement.
Cette recherche bibliographique qui précède les travaux de
terrain est poursuivie sur le terrain jusqu'à la phase de
rédaction du document final en vue d'éventuels compléments
d'informations.
Le nombre important des villages ayant accueilli des
reboisements de mangrove dans le delta du Saloum a fait que le choix des sites
de l'étude est basé sur un certain nombre de critères de
sélection. L'échantillonnage consiste à choisir deux
villages (sites) se trouvant respectivement l'un sur la partie continentale et
l'autre sur la partie insulaire du delta dans lesquels l'UICN a initié
des reboisements de mangrove. Pour ce faire quatre villages ont
été présélectionnés
(Sanghako, Médina Sanghako,
Baout et Djirnda) en raison de leur situation
géographique dans la RBDS (partie insulaire et continentale) ainsi que
de leur proximité, mais après la visite de terrain les sites de
Sanghako1 et de Djirnda2 ont été finalement
retenus pour diverses raisons. Le choix de Djirnda à la place de Baout
est motivé par le nombre de plantations réalisées (six
à Djirnda contre trois à Baout), l'engouement des populations et
la réussite des plantations. Quant au choix de Sanghako au lieu de
Médina Sanghako, il est motivé principalement par le fait que les
plantations de Sanghako remontent de la première campagne de reboisement
que l'UICN a réalisé dans le delta du Saloum.
Sur le terrain, les plantations sont localisées
à l'aide d'un GPS et des relevées de coordonnées
géographiques ont été faites en fonction de leur forme
géométrique en vue de la détermination de leurs
superficies. L'inventaire au niveau de chaque plantation est fait à
l'aide d'un transect réalisé suivant son plus grand axe. La
collecte des données consiste à faire des mesures
dendrométriques (diamètre à la base DB,
diamètre à mi hauteur DmH et hauteur totale
HT) et de biomasses (pesée des feuilles, fûts et
branches) dans des placettes de 1m×1m disposées alternativement le
long des transects. Ces placettes sont disposées suivant un
écartement de cinq (05) à dix (10) mètres selon la taille
des plantations considérées.
1 Cf. carte de localisation des plantations de Sanghako (fig.
3).
2 Cf. carte de localisation des plantations de Djirnda (fig.
2).

13
Plantation
Transect
Placette d'inventair
Figure 1: Schéma du dispositif
d'inventaire.
La superficie, le taux de recrus naturels et la densité
sont retenus comme paramètres d'appréciation de la dynamique des
plantations. La superficie de chaque plantation est déterminée
par traitement des coordonnées des points caractéristiques de sa
figure géométrique par utilisation du logiciel de cartographie
Arc View. Ces coordonnées géographiques
sont relevées par le moyen d'un GPS Garmin pour
traitement à l'ordinateur. La collecte des données (voir les
fiches d'inventaire en annexes) relatives au taux de recrus naturels se fait
par décompte des nouvelles pousses (recrus naturels) dans chaque
placette. Le nombre total des nouvelles plantules est rapporté à
la surface totale des placettes pour donner le taux de recrus naturels
exprimé en nombre de recrus par hectare (recrus/ha). Enfin, les
données nécessaires à la détermination de la
densité de chaque plantation sont collectées par décompte
de tous les sujets vivants (recrus naturels et plants réussis confondus)
se trouvant dans les placettes disposées le long de la ligne de
transect. Le nombre total des sujets dénombrés est
rapporté à la surface totale des placettes puis extrapolé
à l'hectare pour donner la densité exprimée en nombre
d'individus par hectare.
Les mesures dendrométriques sont utilisées pour
élaborer des régressions allométriques. Le diamètre
à la base (DB) est mesuré sur tous les sujets se
trouvant dans les placettes à la limite supérieure des racines
échasses ou bien au dessus de l'hypocotyle des jeunes sujets à
l'aide d'un pied à coulisse. Le diamètre à mi hauteur
(DmH) est mesuré sur les mêmes sujets et toujours
à l'aide du pied à coulisse mais cette fois à mi hauteur.
La hauteur totale (HT) est pour sa part mesurée sur les
plants (sujets) des placettes à l'aide d'un bâton gradué.
Toutes ces
données sont ensuite traitées en vue de la
recherche des équations allométriques correspondantes au moyen
des logiciels Excel (2003/2007) et XLSTAT par
régression simple et multiple méthode progressive (step by step).
Cette méthode laisse au logiciel la possibilité de
sélectionner les équations les plus représentatives de la
distribution des données expérimentales. Les critères de
validation des régresseurs se résument par leur contribution en
termes de probabilité associée à la variable de Student
(t) et de Fischer (F).
L'estimation de la biomasse se situe dans la perspective de
tirer une bonne connaissance de la productivité végétale
des arbres voire des écosystèmes forestiers afin de mieux
comprendre leur mode de fonctionnement en vue de leur meilleure gestion
[Rondeux, 1999]. En foresterie il existe deux principales
méthodes de mesure de biomasse : la méthode directe ou de
pesée et la méthode indirecte ou de multiplication qui consiste
à multiplier le volume des arbres par leur masse volumique. La
méthode directe très pratiquée aux USA dans le but de
rationaliser la détermination de la quantité de bois ronds et
empilés, présente plusieurs avantages entre autres son
procédé simple et rapide menant à une mesure objective de
la quantité de matière, sa possibilité d'automatisation
des mesures par le moyens des régressions allométriques et enfin,
sa détermination directe, contrairement au volume toujours sujet aux
irrégularités de forme mais nécessite cependant, un plan
de sondage statistiquement acceptable.
Cependant, la méthode indirecte bien que pouvant
être rapide si le volume et la masse volumique des bois sont connus
d'avance peut devenir fastidieux au cas où il faut estimer le volume des
bois et déduire leur masse volumique. Dans le delta du Saloum, des
auteurs comme Doyen (1985), Ndour (2005) et
JICA/JAFTA (2006) ont eu à faire recours aux techniques
de mesure de biomasse pour étudier divers aspects de
l'écosystème mangrove. Doyen a utilisé la méthode
directe pour déduire la quantité de matière
nécessaire pour la production de charbon de bois, de bois de chauffe et
de bois d'oeuvre en termes d'impact socioéconomique des massifs de
mangrove de la RBDS. Ndour (2005) a quant à lui fait
usage de cette même méthode de pesée pour calculer les taux
d'humidité et les coefficients de pondération des bois de
mangrove du delta du Saloum. Cependant, JICA/JAFTA (2006) a
fait usage de la méthode indirecte ou de multiplication pour estimer la
productivité de la mangrove de la RBDS mais aussi leur impact en termes
de séquestration au plan d'impact économique. En dépit des
nombreuses mesures de biomasse que la mangrove du delta du Saloum a fait objet,
les reboisements de mangrove n'ont jamais fait objet ni de mesure de
biomasse ni d'étude d'impact en termes de
séquestration de carbone atmosphérique et d'adaptation aux effets
des changements climatiques.
Ainsi, afin d'éviter les mesures de volume et de masse
volumique trop fastidieuses et parfois incertaines, dans cette étude la
méthode directe a été préférée
à la méthode indirecte et se structure comme suit :
o La mesure des paramètres dendrométriques
consiste premièrement à marquer sur chaque plants des placettes
et à l'aide d'un marqueur indélébile les
numéros3 de placette et d'observation de chaque individu
inventorié puis à mesurer sa hauteur totale
(HT), son diamètre à mi hauteur
(DmH) et son diamètre à la base
(DB) et ce à l'aide d'un bâton gradué et
d'un pied à coulisse.
o La coupe rase consiste à couper le tronc et les
racines échasses (si elles existent) des plants ainsi mesurés
à la limite du sol. Ils sont ensuite sectionnés en branche
feuillage et tronc (fût et racines échasses) en vue d'être
peser.
o La pesée des biomasses s'est faite à l'aide
d'une balance électronique de précision (#177;1g) et consiste
à peser chaque individu inventorié partie par partie dans l'ordre
de feuillage, branche puis tronc.
o Enfin le prélèvement des échantillons
consiste pour sa part à prendre dans un sac quelques quantités
des parties (feuillage, branche et tronc) des plants déjà
pesés. Ces échantillons sont ensuite pesés afin de
connaître leur masse à l'état humide, ce qui allait
permettre la déduction du taux d'humidité et du coefficient de
pondération.
En outre, toutes ces mesures ainsi décrites sont
notées sur les fiches d'inventaire en vue des calculs et manipulations
au cours du traitement des données. Au niveau du laboratoire les
échantillons des troncs, branches et feuillages ont été
séchés à l'étuve à 60°C pendant 72
heures (pour les feuilles), 96 heures pour les branches et 120 heures pour les
troncs (fûts et racines échasses). Le choix de ces
différents temps de séchage est fait par expérimentation
qui consiste à peser quelques échantillons deux fois par jour
jusqu'à l'obtention de masse stable. Les échantillons ont permis
le calcul des coefficients de pondération4
(Ro) des taux d'humidité
(Ho) et le carbone séquestré
(C) selon les équations suivantes :
3 Le numéro 1.1 correspond à la
placette numéro 1 individu numéro 1 ; le 2.5
correspond à la placette numéro 2 individu numéro
5 etc.
4 C'est le rapport entre la biomasse sèche
M0 et la biomasse saturée M5 de
l'échantillon.
Équation 1 : Coefficient de pondération
(Ro).
Ro=Mo/Ms ;
Équation5 2 : Taux d'humidité
(Ho).
Ho(%)=[(Ms-Mo)/Mo]×100
; Équation6 3 : Stock de carbone (C).
C=mo×0,5 avec
mo=ms× Ro.
L'élaboration d'une régression
allométrique vise à modéliser la productivité
végétale (capacité de production de matières
végétales) d'un peuplement donné de manière
à éviter les mêmes mesures nécessaires à sa
détermination afin qu'elle soit directement déterminée
à partir des facteurs ou paramètres (dendrométriques et/ou
environnementaux) qui sont facilement mesurables. Rondeux
(1999) distingue trois principales méthodes quantitatives mettant en
oeuvre ces indices à savoir :
o Les méthodes dendrométriques directes dont le
critère de productivité est la production totale de
matières ligneuses ;
o Les méthodes dendrométriques indirectes dont
le critère de productivité est défini par une
caractéristique dendrométrique étroitement liée
à la production totale (en termes de volume ou de biomasse) ;
o Les méthodes combinées dont le critère
de productivité résulte de l'association de
caractéristiques dendrométriques et de diverses variables du
milieu (sol, flore, topographie, climat etc.).
Les deux premières méthodes sont essentiellement
de nature dendrométrique et s'adressent spécifiquement au
matériel ligneux, elles font intervenir des indices (paramètres)
dendrométriques alors que la troisième méthode permet, par
contre, d'associer les indices dendrométriques et les facteurs de
l'environnement sous la forme d'indices combinés. Ainsi donc, la
recherche de régressions allométriques conduit pour la plupart
des cas à l'élaboration
5 Certains auteurs comme Rondeux (1999) ont signalé des
taux allant jusqu'à 200% notamment au niveau des feuilles.
6 Dans ces équations m0 correspond
à la biomasse sèche totale de la plantation exprimée en
tonne de matière par hectare le ms
correspond à l biomasse fraîche totale exprimée tonne de
matière fraîche par hectare alors que le coefficient
0,5 correspond à la proportion de carbone contenu dans
une quantité de biomasse sèche donnée (GIEC, 2008).
des tables de productivité (tarif de cubage ou de biomasse
selon qu'on s'intéresse au volume ou à la quantité de
matière).
Dans le cadre de ce mémoire, contrairement aux
études de foresterie où la productivité se limite aux bois
d'oeuvre (fûts et grosses branches), l'analyse de la productivité
s'étant sur toute la partie pied de l'arbre du peuplement afin d'estimer
la quantité de carbone atmosphérique qui y est
séquestré. La connaissance de la productivité des
peuplements des plantations de mangrove de la RBDS faisant partie
intégrante de tout l'écosystème mangrove du delta du
Saloum nécessite une bonne connaissance de sa biogéographie, sa
morphopédologie et de sa population d'où l'importance de la
présentation de la zone d'étude.
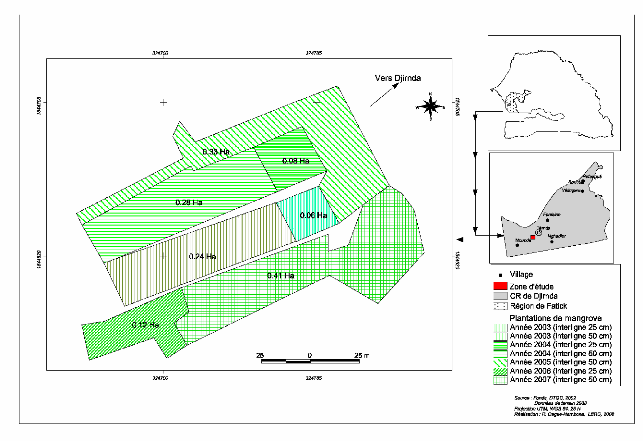
Figure 2 : Carte de localisation des plantations de
Djirnda.

Figure 3 : Carte de localisation des plantations de
Sanghako.
D. PRESENTATION DE LA ZONE
Situé au centre ouest du Sénégal dans la
région naturelle du Sine Saloum, le delta du Saloum s'étend sur
environ 500.000 ha de superficie (DIA, 2003). Il combine les
caractéristiques d'une zone humide marine, estuarienne, lacustre et
palustre, et ses différents sites remplissent les fonctions classiques
d'une zone humide. Les diverses études faites sur la zone du delta du
Saloum ont mis en exergue : un écosystème de mangrove avec ses
vasières riches en matière organique, la présence d'une
multitude de chenaux servant de tampon entre la mer et l'océan et
assurant l'interaction avec le réseau hydrographique en amont, la
diversité spécifique tant animale que végétale, la
présence d'ensembles marin, amphibie et continental.
La zone du delta du Saloum est le lieu d'un important
peuplement humain dont une partie est concentrée dans de grandes
agglomérations7 comme : Niodior, Dionewar, Bassoul, Djirnda,
Palmarin et Bétenti (DIA, 2003). Cette population vit
d'une économie basée essentiellement sur la pêche, le
prélèvement des produits de la mangrove (bois, huîtres,
arches, etc.) et, dans une moindre mesure l'agriculture et l'élevage
puis, plus récemment le tourisme (écotourisme). Le degré
de dépendance de la population du delta du Saloum aux ressources
naturelles témoigne de l'importance du rôle catalyseur que joue
son cadre biophysique en termes de son développement
socioéconomique.
D.1 CADRE BIOGEOGRAPHIQUE
BAILLON (1988) a affirmé que les
caractéristiques actuelles du milieu physique du delta du Saloum ont
été imprimées par les fluctuations eustatiques et
climatiques du quaternaire. La proximité de l'océan,
l'hydrologie, la géomorphologie et la nature des sols exercent par leur
combinaison une influence considérable sur les ressources naturelles de
la zone notamment la mangrove. Ces caractéristiques constituent un
élément déterminant de la physionomie des
écosystèmes (NOUIDEMONA, 2004).
7 Cf. carte de localisation de la RBDS (fig. 4).

Figure 4 : Carte de localisation de la réserve de
biosphère (RBDS).
D.1.1 LE CLIMAT
C'est l'un des éléments les plus
déterminants sur les ressources BAILLON (1988)
décrit le climat de la zone du delta du Saloum comme situé entre
le climat soudanien et soudano sahélien. La présentation du
climat peut être faite suivant ses composantes que sont :
l'insolation, la température, le
vent, la pluviométrie,
l'humidité relative et
l'évaporation.
L'insolation, fait partie des
paramètres climatiques importants car étant liée aux
autres paramètres climatiques par le biais de l'activité solaire
moteur de la photosynthèse et du cycle de l'eau. Elle varie en fonction
du temps d'ensoleillement mais peut être influencée par la
saisonnalité (présence des nuages pendant la saison des pluies).
L'analyse de la variation des moyennes annuelles de l'insolation au niveau de
la station météorologique de Kaolack laisse constater une petite
tendance régressive au cours de la période 1955-2004 (cf. figure
5). Cette tendance régressive pourrait être expliquée par
les effets des variabilités climatiques que connaît le monde ces
dernières décennies ce qui se caractérisent par
l'accumulation des aérosols et gaz dans l'atmosphère. Ces gaz et
aérosols absorbent une partie de l'énergie solaire incidente ce
qui traduirait cette baisse de l'insolation. Le rôle important de
principale source d'énergie et de chaleur que joue le soleil dans le
fonctionnement de la planète terre confère à l'insolation
une certaine incidence directe sur la température et donc la
végétation.

1955
1991
1981
1971
1961
1999
1997
1993
1989
1987
1983
1979
1977
1973
1969
1967
1963
1959
1957
1995
1985
1975
1965
2003
2001
300
290
280
270
260
250
Insolation moyenne
240
230
220
210
Année
Figure 5 : Evolution de la tendance
générale de l'insolation moyenne (en watts/h/m2)
annuelle (Météo Kaolack, 2008).
L'analyse des données météorologiques de
Kaolack (2008) révèle que les moyennes
mensuelles de la température varient selon que l'on se
situe à l'intérieur du continent ou dans les îles. Vers
l'océan, l'effet de l'alizé maritime et l'inertie thermique de la
mer favorisent des températures beaucoup plus basses.
Cette analyse révèle aussi une très
grande variation d'amplitude thermique (température diurne deux fois
plus supérieure à la température nocturne) sur la
période 1951-2003. La température diurne mensuelle la plus
élevée de cette même période est de 42,2° C
(mois d'avril 1987) alors que les températures diurnes moyennes
annuelles ne varient qu'aux environs de 34° C à 37° C.
La tendance généralement croissante des
températures moyennes annuelles et de leurs anomalies des années
1951 à 2003 comme en témoignent les figures N°6 et N°7
traduit une nette augmentation de la température (donc des effets des
changements climatiques) sur la zone du delta du Saloum. La température
des masses d'air caractérise les types de vents qui déterminent
les différents types de précipitations car contenant de la vapeur
d'eau libérée par évaporation à partir des
océans et des continents.

1953
1991
1981
1971
1961
1951
1999
1997
1995
1993
1989
1987
1985
1983
1979
1977
1975
1973
1969
1967
1965
1963
1959
1957
1955
2003
2001
30
29,8
29,6
29,4
29,2
29
28,8
28,6
28,4
28,2
T'moyenne annuelle
28
27,8
27,6
27,4
27,2
Année
Figure 6 : Evolution de la tendance des
températures moyennes annuelles (Météo Kaolack,
2008).
Anomalie de T°
-0,2
-0,4
-0,6
-0,8
-1,2
-1,4
-1,6
-1,8
0,8
0,6
0,4
0,2
1,8
1,6
1,4
1,2
-1
-2
0
1

1 PRI
i 96A
1 PRF,
1 P87
196c.
961
1963
19,37
1969
i 971
1'173
i 975
1977
1979
1981
1983
1985
1987
1989
1991
1993
1995
1997
1999
2001
2003
Année
Figure 7 : Evolution de la tendance des
anomalies8 des températures moyennes annuelles
(Météo Kaolack, 2008).
La zone du delta du Saloum est soumise aux trois types de
vents tels que : L'alizé maritime, l'alizé
continental appelé harmattan et la mousson.
L'alizé maritime est relativement frais.
Il balaie la côte basse et sableuse avec une direction NNW et avec un
pouvoir hygrométrique très faible au début de
l'hivernage.
L'alizé continental (harmattan) est un
vent chaud et sec. Il souffle en saison sèche, son long parcours
continental explique sa charge poussiéreuse et son pouvoir
hygrométrique quasi nul.
La mousson est de direction WSW. Après
avoir effectué un long parcours océanique elle arrive sur le
continent avec une humidité élevée de l'air qui apporte
des pluies de juillet à octobre. Le vent joue un rôle non
négligeable sur la température et la pluviométrie de la
zone du delta du Saloum à travers son rôle de vecteur de
transmission d'humidité et de chaleurs.
La pluviogénèse de la
région du Sine Saloum est située entre les isohyètes 700
mm et 900
mm entre la période de 1918 à 2006 avec une saison
des pluies qui va de juin à octobre
alternant avec une saison
sèche de sept (07) mois allant de novembre à mai
(Météo Kaolack
8 Anomalie de température : (Xi-X)/ó. Avec Xi :
température de l'année considérée, X :
température moyenne de la période et ó : l'écart
type des température de la période.
et Toubacouta, 2008). La pluviométrie
annuelle et interannuelle est très variable. La tendance
généralement décroissante de la pluviométrie et de
ses indices standardisés (voir les figures N° 8 et N° 9) selon
les données des Stations Météorologiques de Kaolack et de
Toubacouta (2008) pour la période de 1918 à 2006
(Kaolack) et de 1957 à 2006 (Toubacouta), témoigne d'une baisse
sensible de la pluviométrie par rapport aux moyennes annuelles
interannuelles de la zone du delta du Saloum pour ces mêmes
périodes. Cette évolution régressive influence
négativement la composition et la dynamique des ressources de la
mangrove.
1918 1920 1922 1924 1926 1928 1930 1932 1934 1936 1938 1940
1942 1944 1946 1948 1950 1952 1954 1956 1958 1960 1962 1964 1966 1968 1970 1972
1974 1976 1978 1980 1982 1984 1986 1988 1990 1992 1994 1996 1998 2000 2002 2004
2006

1300
1250
1200
1150
1100
1050
1000
950
Pluviometrie (mm)
900
850
800
750
700
650
600
550
500
450
400
350
300
Année
Figure 8 : Evolution de la tendance de la
pluviométrie (Météo Kaolack, 2008).

Indice standardis6
-0,1
-0,2
-0,3
-0,4
-0,5
-0,6
0,8
0,7
0,6
0,5
0,4
0,3
0,2
0,1
0
191S
1920
1922 1924 1926 1928 1930 1932 1934 1936
1938
1949
1942
1944
1 948 1950 1952 1954 1956 1958
1969
1962
1964
1966
19E4:
1973
11, 74
.47^
I 960
1952
1954
1956
1955
C....17
1=414
1995
2000
?nn:
2994
2006
Année
Figure 9 : Evolution de la tendance des indices
standardisés9 de la pluviométrie (Météo
Kaolack, 2008).

1957
1991
1981
1971
1961
1999
1997
1995
1993
1989
1987
1985
1983
1979
1977
1975
1973
1969
1967
1965
1963
1959
2003
2001
2005
1500
1450
1400
1350
1300
1250
1200
1150
1100
1050
1000
950
900
850
800
Pluviometrie (mm)
750
700
650
600
550
500
450
400
350
Année
Figure 10 : Evolution de la tendance de la
pluviométrie (Météo Toubacouta, 2008).
9 Indices standardisés : (Xi-X)/X. Xi :
pluviométrie de l'année considérée et X :
pluviométrie moyenne de la période.
Indice standardis6
-0,1
-0,2
-0,3
-0,4
-0,5
-0,6
0,9
0,8
0,7
0,6
0,5
0,4
0,3
0,2
0,1
0
1

1957
1959
19131
1963
1965
1967
1969
1971
1973
1975
1977
1979
lggi
Pri3
1485
.1987
1989
ggi
ggg
1995
1997
1999
2001
2003
2005
Année
Figure 11 : Evolution de la tendance des indices
standardisés de la pluviométrie (Météo Toubacouta,
2008).
L'humidité relative et l'évaporation fortement
tributaires de l'activité solaire et de la pluviométrie
constituent un des maillons importants du cycle de l'eau dans le delta du
Saloum.
L'humidité relative dépend
principalement de l'insolation et du vent mais aussi de la saisonnalité
climatique. Les valeurs les plus élevées sont enregistrées
pendant la saison des pluies à la faveur des masses d'air humides de
mousson et pouvant avoisiner 100% le jour (99,81% en octobre 2003) alors
qu'elles ne dépassent pas 70% la nuit (70,87% en septembre 2003)
(Météo Kaolack, 2008). L'analyse tendancielle de
l'humidité relative moyenne diurne et nocturne au cours de cette
période (1951-2003) laisse apercevoir une régression de
l'humidité relative diurne alors que l'humidité relative nocturne
augmente (voir les figures N° 12 et N° 13). Cependant, la tendance
quasi constante de l'humidité relative moyenne obtenue en faisant la
moyenne de l'humidité (diurne et nocturne) de chaque année,
explique qu'au cours de la période 1951-2003 les pertes de
l'humidité diurne égalent les gains de l'humidité
nocturne.
|
1951
|
1953
|
1955
|
1957
|
1959
|
1961
|
1963
|
1965
|
1967
|
1969
|
1971
|
1973
|
1975
|
1977
Année
|
1979
|
1981
|
1983
|
1985
|
1987
|
1989
|
1991
|
1993
|
1995
|
1997
|
1999
|
2001
|
2003
|

Humidit6 diume
85
84
83
82
81
80
79
78
77
76
75
74
73
72
71
70
Figure 12 : Evolution de la tendance de l'humidité
diurne moyenne annuelle (Météo Kaolack, 2008).

1953
1991
1981
1971
1961
1951
1999
1997
1995
1993
1989
1987
1985
1983
1979
1977
1975
1973
1969
1967
1965
1963
1959
1957
1955
2003
2001
41
40
39
38
37
36
35
34
Humidit6 nocturne
33
32
31
30
29
Année
Figure 13 : Evolution de la tendance de l'humidité
nocturne moyenne annuelle (Météo Kaolack, 2008).
|
1951
|
1953
|
1955
|
1957
|
1959
|
1961
|
1963
|
1965
|
1967
|
1969
|
1971
|
1973
|
1975
|
1977
Année
|
1979
|
1981
|
1983
|
1985
|
1987
|
1989
|
1991
|
1993
|
1995
|
1997
|
1999
|
2001
|
2003
|

Humidito moyenne
65
64
63
62
61
60
59
58
57
56
55
54
53
52
51
50
Figure 14 : Evolution de la tendance de l'humidité
moyenne annuelle (Météo Kaolack, 2008).
L'évaporation est cependant fonction
de la température de l'air, de son pouvoir hygrométrique et de la
vitesse du vent. Elle joue un rôle important dans le cycle de l'eau en
ravitaillant l'atmosphère en vapeur d'eau qui se condense afin de
précipiter sous forme de pluie, brouillard, rosée, neige etc. Les
valeurs les plus élevées sont enregistrées pendant la
saison sèche (Evaporation Potentielle supérieure à 200
mm), mais en saison des pluies elles ne dépassent pas 100 mm au niveau
de la station météorologique de KAOLACK (DRAME
et al. 2003). L'importance du climat se mesure pour la plupart des cas par son
influence sur l'hydrographie et donc par conséquent sur la
végétation, l'homme et les animaux.
D.1.2 L'HYDROGRAPHIE
La zone du delta du Saloum présente un ensemble
d'îles séparées par des cours d'eau dont les plus
importants sont le Saloum qui rejoint la mer par un estuaire et le Diombos
(Ba et al. 2000 ; UICN, 1999). Ce
réseau est alimenté par l'eau de mer qui peut remonter
jusqu'à Birkelane à 130 km de l'embouchure du Saloum (FALL et al.
2000). Cette dernière est délimitée par un cordon sableux
de 18 km, la flèche du littorale de Sangomar (NDOUR,
2005). Malgré l'extrême faiblesse des apports liquides en
provenance de l'amont, ce réseau a un mode de fonctionnement
hydrodynamique grâce à la faiblesse du profil en long de ses
cours d'eau, à la rétention opérée
par la mangrove et l'évaporation. Il n'en demeure pas moins que la
marée essentiellement de type diurne reste le principal facteur de
l'hydrodynamique estuarienne. Il en résulte donc un fort gradient de
salinité des eaux de l'aval vers l'amont. La teneur en sel des marigots
est partout élevée (DIOP et al. 1995) ; il a
été enregistré des taux de 70g/l à
côté de la mer vers la flèche de Sangomar et dans le
Bandiala, plus en aval des taux de 100g/l ont été
enregistrés vers Sokone, soit de deux à trois fois plus que la
salinité moyenne de la mer qui est de 35 g/l (UICN,
1999). Cependant, plusieurs cours d'eau débouchent dans l'estuaire du
Saloum, ce qui témoigne de l'humidité du milieu. Ces cours d'eau
sont : Le Latmingui, le Tyikat dieri, le Tawu et le Bil (FALL
et al. 2000) ; la Néma, le Sokone, le Senghor et les marigots du versant
NW de l'estuaire (NOUIDEMONA, 2004). La péjoration
climatique qui a comme corollaire la forte diminution des apports d'eau douce a
réduit énormément les débits de ces rivières
et marigots, et a occasionné en même temps une forte destruction
des ressources de la mangrove. Aussi la rupture de la flèche de Sangomar
depuis 1987 a entraîné des modifications profondes tant dans
l'hydrodynamique que dans la sédimentologie de l'estuaire
(NDOUR, 2005). Parmi ces modifications, on peut noter la
formation des bancs de sables à proximité de Niodior
(SENE et al. 1987) et l'assèchement de la mangrove au
niveau de rupture de la flèche de Sangomar (NDOUR,
2005).
Le littoral du Saloum est soumis, pour sa part à deux
types de houle, l'une en provenance de l'Atlantique nord et de direction NW
agit pendant toute la saison sèche ; l'autre en provenance de
l'Atlantique sud et de direction SE agit pendant la saison des pluies
(NOUIDEMONA, 2004). La houle du nord a une action non
négligeable car elle est responsable d'une dérive littorale qui
est responsable à son tour de la dynamique des cordons littoraux. Elle
s'est établie pendant la régression marine il y a 4.000 BP
(UICN, 1999). Elle aborde la côte avec une incidence
oblique et a entraîné par des dépôts de sable, la
formation des cordons sableux et le fonctionnement lagunaire de l'estuaire qui
a favorisé l'installation de la mangrove. Ainsi les limons et argiles
transportés par les cours d'eau se sont décantés pour
donner des vasières fixées par une végétation
adaptée à la salure : la mangrove. L'hydrographie est donc un
facteur important dans le transport des matériaux dans le delta du
Saloum et constitue à cet effet un élément non
négligeable dans l'analyse de la pédologie, la géologie et
la géomorphologie de cette zone.
D.1.3 LA GEOLOGIE ET LA MORPHOPEDOLOGIE
Les formes du relief de la zone du delta du Saloum sont
liées à son histoire géologique laquelle est directement
associée à la dynamique marine (UICN, 1999). Les
îles du Saloum sont constituées de vase associée
tantôt avec du sable dont le dépôt remonte du Flandrien
(UICN, 1999). Les tannes correspondent à d'anciennes
vasières, alors que les vases nues sont de la période actuelle.
La transgression marine du Nouakchottien (5.000 BP) a provoqué des
dépôts sableux qui forment des terrasses en bordure du plateau
continental (UICN, 1999) se transformant dans certains cas
à des îles. Toujours selon la même source, après
cette transgression marine, la houle du NW a mis en place des cordons littoraux
successifs qui ont fini par fermer partiellement le golfe à
l'intérieur duquel il s'est opéré un dépôt de
vases. A la suite de la régression marine, d'anciennes vasières
exondées se sont transformées en tannes dépourvues de
végétation (tannes nues), et cette évolution s'est
poursuivie jusqu'à la période actuelle (UICN,
1999). Les terres laissées par le retrait de la mer ont
été occupées il y a longtemps par des populations qui se
nourrissaient d'arches et d'huîtres. Les dépôts de coquilles
consécutifs à cette consommation ont été à
la base de la formation des amas coquilliers (UICN, 1999)
colonisés principalement par les Baobab (Adansonia
digitata).
Au plan morphologique, il a été noté une
dissymétrie entre les deux rives du Saloum (MARIUS,
1977). Cette dissymétrie est à mettre en relation avec
l'incidence des flux d'eau marine à l'intérieur des bolons. A
cause de la flèche de Sangomar, le flot n'entrait pas directement dans
les marigots de la rive droite du Saloum, alors qu'il pénétrait
en profondeur dans les marigots du Diombos et du Bandiala. La
conséquence est la constitution d'un réseau de chenaux moins
stable et moins dense dans la partie nord de l'estuaire, et d'un milieu
véritablement insulaire soumis aux remaniements d'un réseau de
chenaux directement ouverts sur la mer.
La pédologie du delta du Saloum est directement
liée à ses facteurs morphologiques. Sans revenir sur la
pédogenèse de la zone, il convient simplement de rappeler les
grandes formations sableuses du delta avec leurs unités morphologiques.
La carte pédologique des îles du Saloum élaborée par
MARIUS (1977) présente deux grands types de formations
sableuses.
Les sols sableux des cordons qui forment au
plan morphologique deux bandes parallèles
alignées NNW/SSE
dont l'un est situé à côté de la mer, allant de
Djinak au sud à Dionouar au
nord en passant par Bétenti ; le
second est localisé à l'intérieur du continent et couvre
une
grande partie de l'île Bétenti tout en formant un
escarpement au niveau de Ndangane Touti et Sipo.
Les sols minéraux et les sols
peu évolués qui occupent les terrasses
marines sont bien représentés au nord du Diombos, au
niveau des îles Gandoul, de Ngadior à Bakalon en passant par
Bassoul, et du bolon Guilor aux formations du Continental Terminal. Selon le
même auteur ces grands ensembles peuvent être éclatés
: en sols minéraux bruts qui occupent l'essentiel des plages sableuses
de la côte, en sols peu évolués bien
représentés au niveau des îles Bétenti, en sols
halomorphes localisés au niveau des tannes vifs (souvent
désignés sous le terme de sols sulfatés acides), en sols
hydromorphes sur les tannes herbacés, et calcimagnésiques
localisés à l'emplacement des amas artificiels de coquilles. Ces
sols, sous l'effet des déficits hydriques que connait la zones du delta
du Saloum ces dernières décennies, sont soumis à la
salinisation ainsi qu'à l'érosion hydrique et éolienne
dont les effets ont été renforcés par la
dégradation du couvert végétal notamment celui de la
mangrove.
C.1.4 VEGETATION FLORE ET FAUNE
Au plan phytogéographique, le delta du Saloum est
situé entre les zones soudano guinéenne et sahélo
soudanienne. La végétation est présente
sur deux grands types de milieux : les zones submersibles par les marées
et les zones non submersibles.
La végétation des zones
submersibles est formée par la mangrove et les prairies
à halophytes (MARIUS, 1977).
La mangrove est une végétation
caractéristique des estuaires et deltas des régions tropicales
soumises à l'influence de la marée : hauteur de marnage comprise
entre 2 m à 3 m (ISME, 1995). Cinq espèces
floristiques réparties en deux familles constituent la
végétation de cette zone estuarienne.
La famille des Rhizophoraceae est
représentée par trois espèces du genre Rhizophora
ou palétuvier rouge. Rhizophora racemosa (environ 10 m de haut)
colonise les chenaux généralement argileux. Rhizophora
mangle, de moindre taille, forme les peuplements les plus étendus.
Rhizophora harrisonii est plutôt rare. Ces espèces du
genre Rhizophora sont caractérisées par leurs racines
échasses et leur mode de reproduction. Les nombreuses propagules qui
pendent sur les branches des sujets adultes sont dispersées par le
courant. Elles
s'implantent par la suite sur les substrats vaseux. Les
espèces du genre Rhizophora constituent ainsi des plantes
pionnières des vases nues puisqu'elles sont les premières
à les coloniser.
La famille des Verbénaceae n'est
représentée que par une seule espèce, Avicennia
africana ou palétuvier blanc. Cette espèce est
caractérisée par la présence de racines aériennes
ou pneumatophores qui lui permettent d'absorber l'oxygène
atmosphérique.
La famille des Combrétaceae moins abondante
est représentée par Laguncularia racemosa et
Conocarpus erectus. Cependant, au niveau du delta du Saloum on note la
présence des Baobab (Adansonia digitata) isolés ou en
petit peuplement dont le développement est favorisé par la
présence des amas coquilliers.
Les prairies à halophytes sont connues
sous le nom de tannes herbues par opposition aux tannes vives
qui sont des étendues sursalées, dépourvues de
végétation. Sur les tannes herbues poussent des espèces
comme Sesuvium portulacastrum, Philoxerus vermicularis,
Sporobolus robustus, Schizachirium compressa, Paspalum
vaginatum. Dans les mêmes zones on rencontre couramment aussi
Tamarix senegalensis.
La végétation des zones non
submersibles sont en général des savanes arbustives
à boisées avec des espèces soudaniennes et
guinéennes.
Au niveau du littoral, on peut noter la
présence d'espèces floristiques telles que Ipomea
pescaprae, Cyperus maritimus, Imperata cylindrica,
Calotropis procera, Phoenix reclinata, Cocos
nucifera.
Les cordons sableux des îles sont
occupés par Elaeis guineensis, Acacia seyal,
Acacia ataxacantha, Neocarya excelsa. Le baobab
(Adansonia digitata) se développe sur les sols riches en calcaire,
en particulier sur les amas coquilliers.
Sur le continent la zone présente de
nombreuses espèces floristiques telles que
Daniellia
oliveri, Cordyla pinnata,
Pterocarpus erinaceus, Bombax costatum, Lannea
acida, Khaya
senegalensis, Parkia biglobosa,
Sclerocarya birrea, Stereospermum kunthianum,
Terminalia
macroptera, Terminalia avicennioides,
Tarmarindus indica. Au niveau des zones de terroir
oüse déroulent les activités agricoles et
pastorales, Cordyla pinnata est l'espèce ligneuse
dominante à cause de son rôle dans les agro
systèmes. La richesse en termes de diversité floristique et la
présence de nombreuses aires protégées font de la zone du
delta du Saloum, un habitat favorable au développement de la faune.
La faune du delta du Saloum présente
une dominance des petits mammifères dans la partie continentale, surtout
dans la forêt de Fathala.
On note la présence de beaucoup de Phacochères
(Phacocherus aethiopicus), de Guib harnaché (Tragelaphus
scriptus), de l'Hyène tachetée (Crocuta crocuta),
quelques chacals à flanc rayé. Les grands mammifères comme
le Guib harnaché ne sont présents qu'à l'intérieur
de la Réserve de Biosphère.
Dans le parc national on note la présence de quatre
espèces de primates notamment : le colobe bai de terminick (Colobus
badius termminckii) dont la forêt de Fathala constitue le domaine de
prédilection, le singe vert (Cercopithecus aethiops sabeus), le
singe rouge (Erythrolepus patas) et le Lémurien galago du
Sénégal (Galago senegalensis). On remarque aussi la
présence de rongeurs (Rat, Porc-épic), des reptiles (Python de
seba, Cobra...) (SOUMARE et al. 1992) cité par
NOUIDEMONA (2004).
Dans le domaine aquatique la faune sauvage est
constituée de dauphins (Delphinus delphis), de baleines, de
cachalots, de varans du Nil. La faune ichtyologique du Saloum est
marquée par la présence des espèces de poissons
démersales comme les Mérous et les Soles, de même que les
espèces pélagiques comme le Thon.
Les oiseaux constituent, au plan es effectifs la faune la plus
importante du parc. On peut citer entre autre : l'aigrette à gorge
blanche (Egretta galaris), la sterne royale (Sterna maxima),
le héron à dos vert (Butorides striatus), le balbuzard
fluviatile (Pandion haliaetus), etc.
La faune domestique est constituée quant à elle
par de gros bétail les petits ruminants et les volailles. L'importance
des ressources végétales et animales dans le contexte de
développement socioéconomique a fait de cette zone un lieu
d'important peuplement humain riche au plan diversité ethnique et
culturelle. Ces conditions biophysiques en équilibre expliquent la
présence de la mangrove. Leur perturbation récente a
entraîné la dégradation de tout
l'écosystème.
D.2 LE CADRE HUMAIN
D.2.1 LE PEUPLEMENT
BAILLON (1988) cité par
NOUIDEMONA (2004) situe l'implantation des villages du
delta
du Saloum entre la fin du 18ème siècle et le
début du 19ème siècle. Historiquement les
différentes communautés rurales de la zone
faisaient partie du territoire de Kaolack qui correspond au royaume du Saloum
et quelques états indépendants ou vassaux. Le royaume
s'étendait de Kaolack à Kaffrine et de Kaël au nord jusqu'au
Rip au sud (DPS/RGPH, 1992). Le découpage administratif
survenu en 1983 a scindé l'ancienne région du Sine Saloum en deux
: Kaolack et Fatick.
Le choix de la zone pour la fondation des villages
était en grande partie lié à la disponibilité des
ressources naturelles indispensables au développement des
activités socioéconomiques (NOUIDEMONA,
2004).
Les populations du delta du Saloum vivent principalement des
retombées de l'agriculture, de la pêche, de l'élevage, mais
aussi de la cueillette des produits de la mangrove (Arches, Bois, Huîtres
etc.).
La composition ethnique principale de la zone est de 55% de
Sérer, 30% des Wolof, 9,2% des Peul, 2,1% des Socé et 1,3% des
Bambara (MEF, DPS 1992). Toutefois, il faut relever que la
plupart des ethnies constitutives du Sénégal sont
présentes dans la zone. La particularité du peuplement du delta
du Saloum en termes de diversité ethnique et culturelle qu'en termes de
dépendance vis-à-vis des ressources naturelles, confère
à cette région un profil démographique très
particulier comparativement aux autres régions du territoire
national.
D.2.2 LE PROFIL DEMOGRAPHIQUE
La région administrative de Fatick couvre une
superficie de 7.935 km2, soit 4,03% de la superficie du territoire
national. Sa population était estimée en 1988 à 509.702
habitants (DPS/RGPH 1988). Au cours de la période
1976-1988, le taux moyen d'accroissement annuel était de 1,7%. Ensuite
Sa population est passée de 610.520 habitants en 1997 (EDSIII), à
628.968 habitants en 2000, soit 6,9% de la population nationale. La
densité est de 79 habitants par km2 en 2000 pour une moyenne
nationale de 35 hab/km2. La population en majorité rurale est
inégalement repartie suivant la distribution géographique des
ressources naturelles.
L'analyse des résultats du RGPH de
1988 révèle une population très jeune. Environ 57,8% ont
moins de 20 ans, 35,7% ont entre 20 et 59 ans et 6,5% ont plus de 60 ans. Ces
jeunes ont tendance à migrer à cause de faibles perspectives
qu'offre la gestion des ressources de la zone. Ces mêmes résultats
situent le taux des femmes en âge de procréer (15-49 ans)
à
41,5% de la population féminine de la région. Le
taux d'accroissement moyen annuel de l'ordre de 1,7% est inférieur
à la moyenne nationale estimée à 2,9%.
La dynamique de la population repose sur deux
déterminants essentiels que sont les mouvements naturels et la
migration. L'organisation socio-économique de la population du delta du
Saloum à l'instar d'autres populations des zones rurales du
Sénégal est fortement tributaire des ressources naturelles et de
la clémence du climat.
C.2.3 L'ORGANISATION SOCIOECONOMIQUE
Les populations du delta du Saloum sont à dominance
rurales et pratiquent une économie de subsistance de type agraire dans
leur majorité, avec des systèmes d'exploitation et de mise en
valeur basés sur des techniques souvent élaborées mais
reposant sur la force humaine (NOUIDEMONA, 2004).
Dans les villages les carrés familiaux sont des grandes
concessions, placées chacune sous l'autorité d'un patriarche ou
d'un aîné plus âgé en cas de décès des
parents (NOUIDEMONA, 2004). Mais à chaque ethnie
correspond une organisation sociale spécifique. Les rôles sur la
gestion des ressources sont attribués en fonction du genre. Ainsi, la
cueillette des produits de la mangrove (bois, arches, huîtres etc.), la
transformation des produits halieutiques, et le jardinage sont
réservés aux femmes tandis que la pêche, l'élevage
et l'agriculture sont destinés pour la plupart des cas aux hommes
(ISE 29ème Promotion, 2007).
Les principales activités de ces populations sont la
pêche, l'agriculture et l'élevage. Les populations des îles
pratiquent une pêche commerciale de subsistance. L'agriculture et
l'agroforesterie sont pratiquées dans les îles et sur la partie
continentale de la Réserve de Biosphère. Parmi les
spéculations on note des cultures de subsistance comme le mil et le riz
et des cultures de rente comme l'arachide ainsi que des cultures
fruitières dont l'anacardier (Anacardium occidentale) est
l'espèce la plus utilisée (UICN, 1999).
Dans cette zone l'élevage est du type extensif.
Cependant, le bétail est exposé à l'agression des mouches
(tsé-tsé) au niveau des îles. Les forêts
classées de Fathala, Sangakho, Keur Sambel et Djilor,
représentent dans la partie Est de la Réserve d'importantes zones
de pâturage pour le bétail.
D.2.4 PRESENTATION DU VILLAGE DJIRNDA
Le village de Djirnda chef lieu de la communauté rurale
de Djirnda fait partie des îles du Saloum et peuplé principalement
des sérères côtoyés par des marchands
guinéens qui travaillent sur les produits halieutiques localement
appelés « sousou » du nom de leur ethnie. C'est marchands
constituent les principaux acteurs étrangers de l'économie du
village et sa deuxième source de devise étrangère
après les travailleurs immigrés ressortissants du village se
trouvant en Europe notamment Espagne et France. L'immigration est
considérée comme une forme réponse donc d'adaptation des
villageois aux différents défis environnementaux comme la perte
de la productivité des sols, la diminution des ressources halieutiques
et la baisse de la pluviométrie que connaît le village.
L'observation des activités menées dans le
village révèle que la pêche, la cueillette des fruits de
mer et la transformation des produits halieutiques constitue les principales
activités génératrices de revenus. L'élevage est
encore pratiqué mais de manière artisanale alors que
l'agriculture est abandonnée depuis quelques décennies pour cause
d'augmentation de la salinité des sols due à la remontée
du sel par capillarité sous l'effet de la sécheresse. La
pêche est pratiquée par les hommes tandis que la cueillette et la
transformation des produits halieutiques sont réservées aux
femmes qui se sont organisées en association (Association des Femmes
Transformatrices pour le Reboisement de la Mangrove : AFTRM) qui gère
aussi la finance du village par la création d'une caisse
d'épargne et de crédit. Certaines activités subsidiaires
comme l'apiculture et le ramassage de bois de mangrove sont aussi
pratiquées mais avec des acteurs mixtes.
La connaissance du rôle important que joue la mangrove
dans la chaîne trophique côtière serait à l'origine
de la prise de conscience de la population en générale et des
femmes en particulier. La discussion avec les femmes a d'ailleurs
confirmé cette hypothèse. Créée dans le
début de l'année 2003 dans l'idée de permettre aux femmes
du village de s'unir afin d'augmenter la rentabilité de leurs
activités, l'AFTRM a reçu le soutien de l'UICN et d'autres
organismes comme JICA/JAFTA et PAGEMAS en termes de formation aux techniques de
reboisement et de transformation des produits halieutiques, en apiculture en
ostréiculture et au micro finance. L'objectif principal de cette
association en termes de reboisement selon les femmes, c'est de remplir toutes
les tannes humides de la zone en reboisant chaque année, c'est dans
cette optique qu'à partir du mois d'août de chaque année,
les femmes du village se réunissent pour reboiser la mangrove.
En termes de prise de décision, l'AFTRM exerce un
pouvoir certain sur la cueillette des fruits de mer et les plantations de
mangrove mais n'a aucun pouvoir sur la gestion des autres formes de ressources
qui revient au Comité de Plage (CP) et le conseil communautaire.
Chargé de faire respecter le plan de gestion des ressources du village,
le CP se trouve souvent confronté à de sérieux
problèmes de leadership et d'autorité conduisant de fois à
l'impunité des sujets pris en flagrant délit de non respect des
termes du plan de gestion des ressources du village.
Cependant, la population de Djirnda n'a pas bonne connaissance
des changements climatiques ni de ses effets à part l'avancée de
la mer et ignore l'existence du marché de carbone et le Mécanisme
pour un Développement Propre (MDP) mais, manifeste malgré tout le
vif désir d'en savoir plus.
Tableau 1 : Tableau d'implication des acteurs dans la
gestion des ressources.
|
RESSOURCES
|
ACTEURS
|
|
AFTRM
|
CP
|
|
Formation naturelle de Mangrove
|
Non
|
Oui
|
|
Reboisement mangrove
|
Oui
|
Non
|
|
Fruits de mer
|
Oui
|
Non
|
|
Autres
|
Non
|
Oui
|

39
Figure10 15 : Séchage de poissons au
soleil. Figure 16 : Four traditionnel le fumage de poissons.
Figure 17 : Bois de mangrove. Figure 18 : Bovin du
village de Djirnda au pâturage.
D.2.5 PRESENTATION DU VILLAGE SANGHAKO
Faisant partie des villages de la communauté rurale de
Toubacouta, le village de Sanghako
contigu à celui de Médina
Sanghako est situé à environ 15km de Sokone au bord d'un
grand
axe routier menant jusqu'à Gambie Il est peuplé à
majeur partie des Serrères et Mandingues.
10 Les photos des figures N° 15 à 18 sont prises par
R. Dégué-Nambona et Cheikh Ndiaye en août 2008 au village
Djirnda. Elles illustrent l'usage multiple de la mangrove en termes de
pâturage, de source d'énergie par sa production de bois de chauffe
et d'oeuvre etc.
Sa localisation au bord d'un grand axe routier
détermine son économie principalement basée sur
l'agriculture et l'élevage.
L'analyse par observation des activités
pratiquées dans le village de Sanghako laisse apercevoir que
l'agriculture constitue l'activité principale, elle est secondée
par la pèche et la cueillette des fruits de mer. Certaines
activités comme l'élevage, l'apiculture, l'ostréiculture
et le commerce sont pratiquées dans la localité mais avec une
importance moindre.
Le choix du mois d'août pour la réalisation de
l'étude de terrain a été à l'origine du manque de
cadre de discussion avec la population pour la simple raison que ça
coïncide au début de l'hivernage qui fait que toute la population
est préoccupée par les activités agricoles. Ceci
témoigne de l'importance de l'agriculture en temps qu'activité
génératrice de revenus au sein de ce village. Le manque de cadre
de discussion est aussi à l'origine de la faible disposition
d'informations sur les conditions de réalisation des reboisements mais,
selon l'UICN la réalisation de ces reboisements ne constitue qu'une
activité complémentaire des activités d'un grand projet de
formation de la population à l'ostréiculture et à
l'apiculture.
Selon certains villageois rencontrés, il existe deux
organisations principales chargées de la gestion des ressources :
l'association des femmes qui travaille avec JICA/JAFTA sur le reboisement de la
mangrove et l'ostréiculture et les écogardes qui veillent sur
l'application des textes régissant l'utilisation des ressources et la
protection de l'AMP de Bamboung contre les braconniers.
E. RESULTATS
L'analyse des données collectées au cours de
cette étude et leur traitement ont conduit à l'obtention de ces
résultats regroupés en trois grandes parties : l'état des
plantations, les régressions allométriques et la
séquestration de carbone enfin la dynamique du carbone en rapport avec
les plantations. Cette partie de l'étude consiste à
présenter ces résultats site par site afin de les discuter.
E.1 LES RESULTATS DU SITE DE DJIRNDA
E.1.1 Etat des plantations :
Les plantations de mangrove de Djirnda occupent une tanne
à environ 2 km au SW du village et s'étendent
sur une superficie totale de plus de 1,5 ha. Elles sont constituées de
deux espèces de palétuviers : le Rhizophora sp
et l'Avicennia sp. Cette étude
porte uniquement sur les plantations de Rhizophora sp
qui constituent l'essentiel des plantations du site de Djirnda.
La réalisation de la première plantation remonte au début
de la deuxième moitié de l'année 2003, puis elle s'est
poursuivie par une série de plantations réalisées chaque
année à partir du mois d'août. Selon les femmes
transformatrices des produits halieutiques cette initiative est née d'un
projet de l'UICN. Elles ont aussi expliqué que la promotion de
l'espèce Rhizophora sp à la place
d'Avicennia sp est due au fait que la
réalisation des plantations de Rhizophora sp
n'exigent pas de grands travaux de préparation car il suffit de ramasser
les propagules mûres et de les enfoncer dans la vase alors que pour
l'Avicennia sp, il faut préparer bien avant
les pépinières mais en plus de cela les graines se font rares
dans la zone et de fois, il faut aller jusqu'aux villages voisins pour en
chercher. En outre, elles ont avoué ne pas savoir laquelle des deux
techniques de reboisements (écartement 25/25 et 50/50) convient le mieux
si bien qu'elles alternent les deux techniques dans une même campagne de
reboisement (2003 et 2004) ou alors dans des campagnes différentes
(2005, 2006 et 2007).
Au plan des densités, les plantations présentent
pour la plupart des cas de bonnes densités (voir le tableau N°2)
proches des valeurs théoriques minimales (160.000 pieds/ha pour 25/25 et
40.000 pieds/ha pour 50/50) définies en supposant que 1m2
d'une plantation avec un écartement de 25/25 (respectivement 50/50)
contient au minimum 16 plants (respectivement 4 plants) s'il n'y pas eu de
mortalité. Ces densités témoignent donc d'un faible taux
de mortalité doublé d'une bonne dynamique interne qui serait
favorisée par un bon taux de régénération
naturelle. La plantation de 2004 d'écartement 50/50 va même au
delà de la valeur théorique minimale qui est de 40.000 pieds/ha,
ce qui témoigne de son bon état survie.
Cependant, la dynamique spatiale est tributaire du climat, des
conditions écologiques et des actions anthropiques car, l'accroissement
spatial de ces plantations est dû principalement aux travaux de
reboisement que font tous les ans les femmes du village.
En outre, la présence dans les plantations des
abeilles, crevettes, huîtres et d'un nid d'oiseau ainsi que celle des
fleurs et propagules observées sur beaucoup de plantes (voir figures
N° 19 à 23) témoigne des bonnes conditions
écologiques du site voire du village à abriter les plantations de
mangrove.

Figure11 19 : Nid d'oiseau (plantation 2003
50/50). Figure 20 : Propagules (plantation 2003 25/25).

Figure 21 : Taille des plants (plantation 2003 25/25).
Figure 22 : Transect/placette (plantation 2006 25/25).
11 Les photos des figures N° 19 à23 sont prises en
août 2008 à Djirnda Par R. Dégué-Nambona
et
Cheikh Ndiaye. Elles illustrent la taille et le développement des
plantations et leur diversité.

Figure 23 : Taille des plants (plantation 2005
50/50).
Tableau 2 : Récapitulatif des données
caractéristiques de l'état des plantations (Site de
Djirnda).
|
Année
|
Nbre d'ind.
mesurés
|
Ecartement
(cm)
|
DB (cm)
|
DmH
(cm)
|
HT (m)
|
Densité
(pieds/ha)
|
Recrus
naturels
(pieds/ha)
|
Superficie
(ha)
|
|
2003 N°1
|
42
|
25/25
|
[0,9 ; 2,7]
|
[0,4 ; 1]
|
[0,99 ; 1,90]
|
140.000
|
3.333
|
0,06
|
|
2003 N°2
|
27
|
50/50
|
[1,2 ; 2,7]
|
[0,5 ; 1,5]
|
[0,75 ; 1,83]
|
33.750
|
1.250
|
0,24
|
|
2004 N°1
|
33
|
25/25
|
[1 ; 2,2]
|
[0,4 ; 1,2]
|
[0,58 ; 0,90]
|
110.000
|
0
|
0,08
|
|
2004 N°2
|
32
|
50/50
|
[0,9 ; 2]
|
[0,3 ; 1,2]
|
[0,5 ; 1]
|
41.250
|
0
|
0,28
|
|
2005
|
19
|
50/50
|
[1 ; 2]
|
[0,7 ; 1,3]
|
[0,51 ; 0,75]
|
38.000
|
0
|
0,33
|
|
2006
|
36
|
25/25
|
[0,6 ; 0,8]
|
[0,2 ; 0,5]
|
[0,4 ; 0,71]
|
90.000
|
0
|
0,12
|
|
2007
|
N.I12
|
50/50
|
N.I
|
N.I
|
N.I
|
N.I
|
N.I
|
0,41
|
|
Total
|
1,62
|
Les valeurs des densités des plantations
présentées dans le tableau ci-dessus laissent apercevoir deux
classes des plantations selon les types d'écartement (25/25 et 50/50).
Les plantations d'écartement 25/25 de 2003 à 2004
présentent de bonnes densités ce qui témoignent de leur
bon taux de survie alors que celle de 2006 présente une faible
densité (90.000pieds/ha pour une valeur théorique minimale
160.000pieds/ha) ce qui démontre d'un taux de survie moyen. Cette
faiblesse de densité pourrait être expliquée par la
qualité de son substrat trop vaseux par rapport aux autres de 2003 et
2004. Cependant les plantations d'écartement 50/50 présentent
toutes de bonnes densités ce qui démontrent de leur bonne
12 N.I : Non inventoriée.
survie. En outre, au plan de recru naturels, seules les
plantations réalisées en 2003 présentent des cas de
régénération. Cependant, cela ne signifie pas qu'il n'y a
pas eu effectivement de régénération au niveau des autres
plantations mais c'est la faiblesse de leur fréquence qui est ainsi
démontrée.
E.1.2 Les régressions allométriques :
L'idée de la recherche de régressions
allométriques est de voir s'il existe des équations
mathématiques pouvant mettre en relation et de façon
significative les paramètres dendrométriques dits variables
explicatives ou régresseurs, à la productivité
végétale dite variable expliquée afin que
dorénavant, au lieu de refaire les mêmes mesures de biomasses pour
la détermination de la productivité, qu'elle soit directement
calculée à l'aide de ces régressions à partir des
mesures de certains paramètres (HT, DmH et DB). Cependant, pour
faciliter l'interprétation des résultats tout en se
prononçant sur la contribution de chaque régresseur aux
différents modèles de régression, les régressions
linéaires simple et multiple ont été
préférées aux autres types de régressions
conseillés par divers auteurs entre autre Rondeux
(1999). C'est ainsi que, pour chaque plantation il sera présenté
le modèle de régression, l'analyse de la variation des variances
et les résidus obtenus par traitement de données initiales
à l'aide du logiciel XLSTAT méthode progressive (step by step)
afin de juger de la pertinence des régresseurs et de leurs
modèles.
Les équations des modèles mettent en relation la
productivité végétale et les régresseurs qui
contribuent de manière significative à la régression en
termes de probabilité de la variable de Student (t) et de Fischer (F)
qui sont leurs estimateurs globaux. Donc, tout régresseur dont la
probabilité de la variable de Student (Pr>t) ainsi que celle de
Fischer (Pr>F) dépasse le seuil de 5% est automatiquement mis
à l'écart par le logiciel. Ceci signifie qu'au-delà du
seuil de 5%, le régresseur ne contribue pas de manière
significative à l'explication du modèle par la régression
ainsi définie.
Les équations des modèles mettent aussi en
relation la somme des contributions des régresseurs retenus et
pondérées chacune d'un coefficient, le tout corrigé d'un
autre coefficient constant.
L'importance des régresseurs (ici les paramètres
dendrométriques) varie d'une plantation à une autre mais aussi
d'un type de régression à un autre. L'analyse des
caractéristiques des régressions linéaires simples laisse
apercevoir en général et dans l'ordre une meilleure contribution
de la HT puis du DmH par rapport à celle du DB.
Au plan des régressions relatives à la HT, les
plantations de 2003 écartement 25/25 (40 individus inventoriés)
et de 2005 écartement 50/50 (17 individus inventoriés)
présentent d'assez bonnes caractéristiques en termes de
coefficient de détermination (0,706 en 2003 et 0,668 en 2005), de
statistique de Durbin-Watson partout proche de sa valeur normale de deux (1,951
en 2003 et 2,368 en 2005) et de la probabilité de la variable de Fischer
très inférieure au seuil de 5%. La statistique de Durbin-Watson
(d), si elle est proche de sa valeur normale (02) traduit une
indépendance de l'ordre d'observation des individus pendant
l'inventaire, ce qui signifie que l'ordre d'observation des individus pendant
l'inventaire ne joue pas sur les résultats obtenus. La
probabilité de la statistique de Fischer (Pr>F)
traduit quant à elle le degré de liaison des régresseurs
au modèle de régression considéré,
c'est-à-dire que si elle est inférieure ou égale à
son seuil de 5% il y a alors une bonne liaison entre les régresseurs et
le modèle. Cependant, alors que les caractéristiques de la
plantation de 2003 (voir les tableaux N° 6 et 7 et les figures N° 26
à 31) écartement 50/50 (27 individus inventoriés)
présentent un coefficient de détermination moyen (0,528) et de
bonne probabilité de la statistique Fischer (très
inférieure au seuil de 5%) et une bonne statistique de Durbin-Watson
(2,049), celles des plantations de 2004 écartement 25/25 et 50/50 et de
2006 écartement 25/25 (respectivement 31, 30 et 36 individus
inventoriés) présentent de mauvais coefficients de
détermination allant de inférieur à très
inférieur à la moyenne (0,453 en 2004 écartement 25/25,
0,193 en 2004 écartement 50/50 et 0,014 en 2006) mais de bonnes
probabilités de la statistique de Fischer ainsi que de bonnes
statistiques de Durbin-Watson proche de sa valeur normale (02) sauf en 2006
où elles sont sorties de leur valeur normale. En somme, la HT
apparaît en général comme un assez bon régresseur
malgré la faiblesse des coefficients de détermination
observée en 2004 et 2006.
Tableau 3: R écapitulatif des
caractéristiques des r égressions relatives a la HT
(Djirnda).
|
Année
|
Régresseur
|
Pr Stat de Fischer
|
Stat de Durbin-Watson
|
Coef. de dét. (R2)
|
|
2003 (25/25)
|
HT
|
0,0001
|
1,951
|
0,706
|
|
2003 (50/50)
|
HT
|
0,0001
|
2,049
|
0,528
|
|
2004 (25/25)
|
HT
|
0,0001
|
2,304
|
0,453
|
|
2004 (50/50)
|
HT
|
0,012
|
2,286
|
0,193
|
|
2005
|
HT
|
0,0001
|
2,368
|
0,668
|
|
2006
|
HT
|
0,485
|
1,411
|
0,014
|
Tableau 4 :Récapitulatif des equations des
regressions relatives a la HT (Djirnda).
|
Année
|
Equations des modèles
|
|
2003 (25/25)
|
Biom sèche(kg) = -0,750 + 0,656*HT
(m)
|
|
2003 (50/50)
|
Biom sèche (kg) = -0,356 + 0,640*HT
(m)
|
|
2004 (25/25)
|
Biom sèche (kg) = -0,129 + 0,290*HT
(m)
|
|
2004 (50/50)
|
Biom sèche (kg) = -7,801E-02 + 0,296*HT
(m)
|
|
2005
|
Biom sèche (kg) = -0,162 + 0,386*HT
(m)
|
|
2006
|
Biom sèche (kg) = 1,179E-02 + 1,507E-02*HT
(m)
|

0,7
0,6
Observations et droite de régression
-0,1
0,5
0,4
0,3
0,2
0,1
0
0 0,5 1 1,5
-0,2
HT (m)
Figure 24 : Nuage de points et droite de regression de la
plantation 2003 écartement 25/25.
Observations et droite de régression
0 0,5 1 1,5 2
HT (m)
Figure 25 : Nuage de points et droite de
régression de la plantation 2003 écartement 50/50.
Observations et droite de régression
|
0,2 0,18 0,16 0,14
|
|
|
|
|
0,12 0,1 0,08 0,06 0,04 0,02
0
|
0 0,2 0,4 0,6 0,8 1
HT (m)
Figure 26 : Nuage de point et droite de r
égression de la plantation 2004 écartement 25/25.
Observations et droite de régression
|
0,4 0,35 0,3
|
|
|
0,25
|
|
0,2 0,15 0,1 0,05
0
|
0 0,2 0,4 0,6 0,8 1 1,2
HT (m)
Figure 27 : Nuage de points et droite de
régression de la plantation 2004 écartement 50/50.
Observations et droite de régression
|
0,18 0,16 0,14 0,12
|
|
|
|
0,1 0,08 0,06 0,04 0,02
0
|
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8
HT (m)
Figure 28 : Nuage de points et droite de
régression de la plantation 2005 écartement 50/50.
Observations et droite de régression
|
0,045 0,04 0,035 0,03
|
|
|
|
0,025 0,02 0,015 0,01 0,005
0
|
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8
HT (m)
Figure 29 : Nuage de points et droite de
régression de la plantation 2006 écartement 25/25.
Au plan des régressions relatives au DmH (voir les
tableaux N° 8 et 9), les plantations de 2003 écartement 25/25 et
2005 écartement 50/50 présentent des coefficients de
détermination moyens (0.64 en 2003 et 0.63) pour de bonnes
probabilités de la statistique de Fischer (très inférieure
au seuil de 5%) et de bonnes statistiques de Durbin-Watson (1.91 en 2003 et
2.13 en 2005). Cependant, les autres plantations présentent des
coefficients de détermination allant de faible à très
faible (0.29 en 2003 écartement 50/50, 0.04 et 0.07 en 2004
écartement 25/25 et 50/50 et 0.30 en 2006 écartement 25/25) pour
de bonnes probabilités de la statistique de Fischer en 2003
écartement 50/50 et en 2006 écartement 25/25 ainsi que de bonnes
statistiques de Durbin-Watson. Le DmH apparaît comme un mauvais
régresseur en raison de sa faible contribution en termes de coefficient
de détermination ce qui ne favorise pas l'explication des observations
par les modèles ainsi définis. En outre, la faiblesse des DmH et
le degré de précision des instruments de mesure utilisés
ont fait que, les nuages de points présentent des répartitions
verticalement superposées en classes de même DmH, ce qui ne
favorise pas beaucoup l'interprétation des observations (voir les
figures N° 32 à 37).
Tableau 5 : R écapitulatif des caract
éristiques des r égressions relatives au DmH
(Djirnda).
|
Année
|
Régresseur
|
Pr Stat de Fischer
|
Stat de Durbin Watson
|
Coef. de dét. (R2)
|
|
2003 (25/25)
|
DmH
|
0,0001
|
1,912
|
0,637
|
|
2003 (50/50)
|
DmH
|
0,003
|
1,716
|
0,295
|
|
2004 (25/25)
|
DmH
|
0,237
|
2,165
|
0,045
|
|
2004 (50/50)
|
DmH
|
0,144
|
1,896
|
0,070
|
|
2005
|
DmH
|
0,0001
|
2,123
|
0,631
|
|
2006
|
DmH
|
0,001
|
1,691
|
0,298
|
Tableau 6 : R écapitulatif des équations
des r égressions relatives au DmH (Djirnda).
|
Année
|
Equations des modèles
|
|
2003 (25/25)
|
Biom. sèche (kg) = -0,190 + 0,673*DmH
(cm)
|
|
2003 (50/50)
|
Biom. sèche (kg) = -1,810E-02 + 0,521*DmH
(cm)
|
|
2004 (25/25)
|
Biom. sèche (kg) = 6,041E-02 + 3,976E-02*DmH
(cm)
|
|
2004 (50/50)
|
Biom. sèche (kg) = 6,819E-02 + 5,545E-02*DB
(cm)
|
|
2005
|
Biom. sèche (kg) = -6,684E-02 + 0,163*DmH
(cm)
|
|
2006
|
Biom. sèche (kg) = -1,420E-03 + 6,063E-02*DmH
(cm)
|
Observations et droite de régression
|
0,7
0,6
0,5
|
|
|
|
|
0,4 0,3 0,2 0,1 0
|
0 0,2 0,4 0,6 0,8 1 1,2
DmH (cm)
Figure 30: Nuage de points et droite de r
égression de la plantation 2003 écartement 25/25.
Observations et droite de régression
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6
DmH (cm)
Figure 31 : Nuage de points et droite de
régression de la plantation 2003 écartement 50/50.
Observations et droite de régression
|
0,2 0,18 0,16 0,14
|
|
|
|
|
0,12 0,1 0,08 0,06 0,04 0,02
0
|
0 0,2 0,4 0,6 0,8 1 1,2 1,4
DmH (cm)
Figure 32 : Nuage de points et droite de
régression de la plantation 2004 écartement 25/25.
Observations et droite de régression
|
0,4 0,35 0,3
|
|
|
0,25
|
|
0,2 0,15 0,1 0,05
0
|
0 0,2 0,4 0,6 0,8 1 1,2 1,4
DmH (cm)
Figure 33 : Nuage de points et droite de
régression de la plantation 2004 écartement 50/50.
Observations et droite de régression
|
0,18 0,16 0,14 0,12
|
|
|
|
0,1 0,08 0,06 0,04 0,02
0
|
0 0,2 0,4 0,6 0,8 1 1,2 1,4
DmH (cm)
Figure 34 : Nuage de point et droite de régression
de la plantation 2005 écartement 50/50.
Observations et droite de régression
|
0,045 0,04 0,035 0,03
|
|
|
|
0,025 0,02 0,015 0,01 0,005
0
|
0 0,1 0,2 0,3 0,4 0,5 0,6
DmH (cm)
Figure 35 : Nuage de points et droite de
régression de la plantation 2006 écartement 25/25.
Enfin, au plan des régressions relatives au DB (voir
les tableaux N° 10 et 11), toutes les plantations présentent de
faible à très faible coefficients de détermination pour
des statistiques de Fischer supérieures à la valeur seuil de 5%
sauf au niveau des plantations de 2004 écartement 25/25 et 2005
écartement 50/50 où elles sont au-dessous de la valeur seuil de
5%. Cependant, les statistiques de Durbin-Watson sont partout
appréciables et proches de la valeur normale sauf en 2003
écartement 50/50 et 2006 écartement 25/25. Les
caractéristiques des régressions laissent constater que le DB ne
constitue pas un bon régresseur de par ses coefficients de
détermination partout très faible mais aussi par ses
probabilités de la statistique de Fischer très supérieures
et traduisant sa non liaison aux modèles définis. Aussi, pareille
pour le DmH, l'existence des classes de même DB a fait que les nuages de
points présentent des observations verticalement superposées
comme le montrent les figures N° 38 à 43.
Tableau 7 : R écapitulatif des caract
éristiques des r égressions relatives au DB
(Djirnda).
|
Année
|
Régresseur
|
Pr Stat de Fischer
|
Stat de Durbin Watson
|
Coef. de dét. (R2)
|
|
2003 (25/25)
|
DB
|
0,575
|
1,855
|
0,008
|
|
2003 (50/50)
|
DB
|
0,071
|
1,360
|
0,124
|
|
2004 (25/25)
|
DB
|
0,004
|
1,956
|
0,237
|
|
2004 (50/50)
|
DB
|
0,313
|
1,862
|
0,034
|
|
2005
|
DB
|
0,035
|
1,417
|
0,237
|
|
2006
|
DB
|
0,570
|
1,467
|
0,010
|
Tableau 8 : R écapitulatif des équations
des r égressions relatives au DB (Djirnda).
|
Année
|
Equations des modèles
|
|
2003
|
(25/25)
|
Biom sèche(kg) = 0,220 + 3,553E-02*DB
(cm)
|
|
2003
|
(50/50)
|
Biom sèche (kg) = -6,614E-02 + 0,288*DB
(cm)
|
|
2004
|
(25/25)
|
Biom sèche (kg) = 0,164 -0,051*DB
(cm)
|
|
2004
|
(50/50)
|
Biom sèche (kg) = 6,819E-02 + 5,545E-02*DB
(cm)
|
|
2005
|
Biom sèche (kg) = -3,505E-02 + 7,916E-02*DB
(cm)
|
|
2006
|
Biom sèche (kg) = 1,451E-02 + 9,119E-03*DB
(cm)
|
Observations et droite de régression
|
0,7
0,6
0,5
|
|
|
|
|
0,4 0,3 0,2 0,1 0
|
0 0,5 1 1,5 2 2,5 3
D B (cm)
Figure 36 : Nuage de points et droite de
régression de la plantation 2003 écartement 25/25.
Observations et droite de régression
1 1,5 2 2,5 3
DB (cm)
Figure 37 : Nuage de points et droite de
régression de la plantation 2003 écartement 50/50.

0,08
0,06
0,04
0,02
0,18
0,16
0,14
0,12
0,2
0,1
0
1 1,5 2 2,5 3
DB (cm)
Observations et droite de régression
Figure 38 : Nuage de points et droite de
régression13 de la plantation 2004 écartement
25/25.
13 La tendance régressive de la variation de
biomasse par rapport aux DB n'est pas normale, elle témoigne d'un
certain nombre d'anomalies. Elle pourrait avoir pour cause des erreurs de
mesure des DB, de la pesée des échantillons ou bien leur
séchage.
Observations et droite de régression
|
0,4 0,35 0,3
|
|
|
0,25
|
|
0,2 0,15 0,1 0,05
0
|
0 0,5 1 1,5 2 2,5
DB (cm)
Figure 39 : Nuage de points et droite de
régression de la plantation 2004 écartement 50/50.
Observations et droite de régression
|
0,18 0,16 0,14 0,12
|
|
|
|
0,1 0,08 0,06 0,04 0,02
0
|
1 1,5 2 2,5 3
DB (cm)
Figure 40 : Nuage de points et droite de
régression de la plantation 2005 écartement 50/50.
Observations et droite de régression
|
0,045 0,04 0,035 0,03
|
|
|
|
0,025 0,02 0,015 0,01 0,005
0
|
0 0,2 0,4 0,6 0,8 1
D B (cm)
Figure 41 : Nuage de points et droite de
régression de la plantation 2006 écartement 25/25.
L'analyse des régressions multiples (voir les tableaux
N° 12 13) laisse apercevoir de bonnes caractéristiques par rapport
aux régressions linéaires simples, ce qui s'explique par le fait
que les régressions multiples cumulent la contribution de tous les
paramètres dendrométriques alors que les régressions
simples ne se contentent que de celle de leur paramètre
dendrométrique considéré.
Les plantations de 2003 écartement 25/25 et de 2005
écartement 50/50 présentent de très bon coefficient de
détermination partout supérieur à 0,70 pour de très
bonnes probabilités de la statistique de Fischer très
inférieures au seuil de 5% et de bonnes statistiques de Durbin-Watson
partout proches de la valeur normale (02), ce qui témoigne d'une bonne
contribution des régresseurs aux modèles mais aussi de
l'indépendance de leurs observations.
Les plantations de 2003 écartement 50/50 et 2004
écartement 25/25 présentent d'assez bons coefficients de
détermination supérieurs à 0,50 pour de bonnes
probabilités de la statistique de Fischer inférieures au seuil de
5% ainsi que de bonnes statistiques de Durbin-Watson proches de la valeur
normale (02). Cependant, seules les plantations de 2004 écartement 50/50
et 2006 écartement 25/25 présentent de faibles coefficients de
détermination inférieurs à 0,50 pour une bonne
probabilité de la statistique de Fischer inférieure au seuil de
5% en 2006 alors qu'elle dépasse ce dit seuil en 2004 ce qui traduit la
non liaison des régresseurs au modèle
ainsi défini. Cependant leur statistique de
Durbin-Watson proche de la valeur normale (02) traduit une indépendance
entre les observations des individus lors de l'inventaire. Les
régressions linéaires multiples par leur particularité
à combiner la contribution de tous les régresseurs
utilisés et leurs caractéristiques partout
généralement bonnes constituent le meilleur type de
régression applicable à notre étude car expliquant de
manière significative la productivité végétale au
niveau du site de Djirnda.
Tableau 9 : récapitulatif des régressions
allom étriques multiples (Djirnda).
|
Année
|
Equations des modèles
|
|
2003
|
(25/25)
|
Biom sèche(kg) = -0,585 + 0,329*DmH (cm)
-3,847E-02*DB (cm) + 0,441*HT (m)
|
|
2003
|
(50/50)
|
Biom sèche (kg) = -0,498 + 5,671E-02*DmH (cm) +
9,377E-02*DB (cm) + 0,572*HT (m)
|
|
2004
|
(25/25)
|
Biom sèche (kg) = -4,138E-02 + 0,035*DmH (cm)
-4,398E-02*DB (cm) + 0,222*HT (m)
|
|
2004
|
(50/50)
|
Biom sèche (kg) = -8,302E-02 + 0,128*DmH (cm)
-6,896E-02*DB (cm) + 0,298*HT (m)
|
|
2005
|
Biom sèche (kg) = -0,170 + 0,100*DmH (cm)
-3,289E-03*DB (cm) + 0,259*HT (m)
|
|
2006
|
Biom sèche (kg) = -6,629E-03 + 6,210E-02*DmH
(cm) + 1,075E-02*DB (cm) -4,341E-03*HT (m)
|
Tableau 10 : R écapitulatif des caract
éristiques des r égressions multiples (Djirnda).
|
Année
|
Régresseurs
|
Pr Stat de Fischer
|
Stat de Durbin Watson
|
Coef. de dét. (R2)
|
|
2003 (25/25)
|
DmH DB et HT
|
0,0001
|
1,820
|
0,778
|
|
2003 (50/50)
|
DmH DB et HT
|
0,0003
|
1,973
|
0,544
|
|
2004 (25/25)
|
DmH DB et HT
|
0,0001
|
2,231
|
0,593
|
|
2004 (50/50)
|
DmH DB et HT
|
0,057
|
2,400
|
0,232
|
|
2005
|
DmH DB et HT
|
0,0001
|
3,071
|
0,825
|
|
2006
|
DmH DB et HT
|
0,0001
|
1,703
|
0,312
|
En outre, les résidus centrés réduits
obtenus en divisant les résidus centrés par l'écart type
résiduel et toujours théoriquement de moyenne nulle, permettent
de mettre en évidence d'éventuelles anomalies existant dans les
données initiales. Dangélie (1992) a
attesté que pour une distribution normale ou approximativement normale
les résidus centrés réduits inférieurs à -3
ou -2 ou supérieurs à +2 ou +3 traduisent une éventuelle
anomalie au niveau des données initiales nécessaire à
découvrir. Ainsi, l'observation de divers résidus centrés
réduits ci-dessous (voir les figures N° 44 à 49) laisse
apercevoir quelques anomalies en 2003 écartement 25/25
(29ème individu), en 2004 écartement 25/25
(21ème individu) et en 2004
écartement 50/50 (2ème 6ème et
22ème individus), pouvant être attribuées aux
erreurs de mesures dues à l'inadvertance de l'opérateur qui
à la charge des mesures.

Ind40
Ind37
Ind34
Ind28
Ind25
Ind22
Ind31
Ind19
Ind16
Ind13
Ind10
Ind7
Ind4
Ind1
-3 -2 -1 0 1 2 3
Résidus centrés-réduits
Résidus centrés-réduits
Figure 42 : R ésidus centrés r
éduits de la plantation 2003 écartement 25/25.

Ind27
Ind25
Ind23
Ind21
Ind19
Ind17
Ind15
Ind13
Ind11
Ind9
Ind7
Ind5
Ind3
Ind1
-2 -1,5 -1 -0,5 0 0,5 1 1,5 2
Résidus centrés-réduits
Résidus centrés-réduits
Figure 43 : R ésidus centrés r
éduits de la plantation 2003 écartement 50/50.

Ind28
Ind25
Ind22
Ind31
Ind19
Ind16
Ind13
Ind10
Ind7
Ind4
Ind1
-4 -3 -2 -1 0 1 2 3 4
Résidus centrés-réduits
Résidus centrés-réduits
Figure 44 : R ésidus centrés r
éduits de la plantation 2004 écartement 25/25.

Ind28
Ind25
Ind22
Ind31
Ind19
Ind16
Ind13
Ind10
Ind7
Ind4
Ind1
-3 -2 -1 0 1 2 3
Résidus centrés-réduits
Résidus centrés-réduits
Figure 45 : R ésidus centrés r
éduits de la plantation 2004 écartement 50/50.

61
Ind19
Ind17
Ind15
Ind13
Ind11
Ind9
Ind7
Ind5
Ind3
Ind1
Résidus centrés-réduits
-2 -1,5 -1 -0,5 0 0,5 1 1,5 2
Résidus centrés-réduits
Figure 46 : R ésidus centrés r
éduits de la plantation 2005 écartement 50/50.

Ind34
Ind28
Ind25
Ind22
Ind31
Ind19
Ind16
Ind13
Ind10
Ind7
Ind4
Ind1
-2,5 -1,5 -0,5 0,5 1,5 2,5
Résidus centrés-réduits
Résidus centrés-réduits
Figure 47 : R ésidus centrés r
éduits de la plantation 2006 écartement 25/25.
E.1.3 Biomasses et Stocks de Carbone :
La capacité des mangroves à se développer
dans des milieux difficiles avec des conditions écologiques et
phytosociologiques (compétition entre les espèces à cause
de la forte densité) particulières, serait à l'origine de
leur forte productivité végétale qui témoigne de
leur grande capacité à fixer le carbone atmosphérique
à travers la photosynthèse.
Les résultats de cette étude ne se portent que
sur les plantations de 2003 jusqu'à 2006, celle de 2007 n'a pas
été inventoriée pour en raison de leur jeune âge ce
qui fait qu'une éventuelle série de coupes rases sur ses sujets
aurait compromis son développement.
Cependant, en dehors des séquestrations à
l'hectare, les séquestrations nettes par plantation ne dépassent
pas deux (02) tonnes de carbone (1.936 tonnes en 2003 N°2) avec un total
cumulé de 4,968 tonnes. Cette faiblesse de séquestration totale
par plantation est due à la faible superficie des plantations
(superficie totale de 1,21 ha excepter celle de 2007). L'écart assez
considérable entre la séquestration de la plantation de 2003
écartement 25/25, plus de trois fois supérieure à celle de
2004 pourrait être expliqué par leur différence d'âge
mais aussi la variabilité du substrat voire du micro climat au niveau du
site (voir tableau N° 3 ci-dessous).
Tableau 11 : Tableau de biomasse et de stocks de carbone
(Djirnda).
|
Année
|
Biomasses fraîches
(t/ha)
|
Biomasses sèches
(t/ha)
|
C séquestré (t/ha)
C=Biomasse
sèche×0,5
|
C tot. Séquestré/plantation
(t)
|
|
2003 N°1
|
71,386
|
38,187
|
19,094
|
1,146
|
|
2003 N°2
|
30,432
|
16,136
|
8,068
|
1,936
|
|
2004 N°1
|
21,426
|
9,943
|
4,971
|
0,400
|
|
2004 N°2
|
12,350
|
6,137
|
3,070
|
0,900
|
|
2005
|
6,460
|
2,488
|
1,244
|
0,475
|
|
2006
|
4,425
|
1,856
|
0,928
|
0,111
|
|
Total
|
4,968
|
L'analyse de la productivité nette des plantations
montre qu'elle évolue avec l'âge et selon le type des plantations
(écartement de 25/25 ou de 50/50). Elle est moins significative entre
deux (02) à quatre (04) ans mais acquiert très vite un niveau
significatif à partir de quatre (04) à cinq (05) ans. Pour la
période de 2003 à 2004 elle est de 28,244 t/ha/an au niveau des
plantations de 25/25 et de 9,999 t/ha/an pour celles de 50/50 alors qu'elle
n'est que de 4,043 t/ha/an et de 3,649 t/ha/an pour les plantations de 25/25 et
de 50/50 entre les périodes de 2004 à 2006. Les plantation du
type 25/25 possèdent donc une bonne capacité de production de
matière végétale comparativement à celles de 50/50,
ceci peut s'expliquer par leur différence de densité car avec un
écartement de 25/25 il y a plus de plantes à l'hectare qu'avec un
écartement de 50/50.
63 L'analyse du taux d'humidité montre qu'il varie
d'une partie des plantes à une autre (voir le tableau N° 4
ci-dessous), ainsi les feuilles présentent en général de
fort taux d'humidité par rapport aux branches et aux troncs. La forte
teneur en eau des feuilles fait que malgré leur biomasse à
l'état humide, elles possèdent des coefficients de
pondération très faibles ce qui réduit
considérablement leur biomasse sèche et par conséquent
leur taux de séquestration de carbone. Les plantations de Djirnda
malgré leur jeune âge (3,5 ans en moyenne) et leur faible
superficie totale d'environ 1,21 ha (sans celle de 2007) séquestrent en
totale 4,968 tonnes de carbone, ce qui devrait s'accroître de
façon exponentielle avec l'âge jusqu'à un certain niveau de
maturité.
Tableau 12 : taux d'humidite moyen, coefficient de
ponderation moyen par parties des plantes (Djirnda).
|
Année
|
Taux d'humidité moy. par partie des plantes
Ho(%)
|
Coefficient de pondération moy./partie des
plantes.
|
|
feuillages
|
Branches
|
Troncs
|
Feuillages
|
Branches
|
Troncs
|
|
2003 N°1
|
157,679
|
73,941
|
91,634
|
0,4
|
0,586
|
0,559
|
|
2003 N°2
|
140,135
|
60,152
|
76,572
|
0,42
|
0,632
|
0,572
|
|
2004 N°1
|
163,037
|
93,167
|
108,403
|
0,386
|
0,541
|
0,489
|
|
2004 N°2
|
154,088
|
80,476
|
93,005
|
0,403
|
0,568
|
0,528
|
|
2005
|
150,117
|
131,090
|
117,021
|
0,404
|
0,449
|
0,469
|
|
2006
|
173,809
|
100
|
137,912
|
0,396
|
0,5
|
0,432
|
L'analyse de la variation des séquestrations en
fonction de l'âge et du type de reboisement (25/25 et 50/50)
révèle que les séquestrations varient de manière
exponentielle avec l'âge et ce avec un coefficient de
détermination (R2) de 1. Ceci justifie la thèse de
croissance exponentielle des jeunes organismes énoncée par
certains auteurs à l'exemple de Dagnelie (1992).
Cependant, le faible nombre des observations à savoir trois classes
d'âge (2003, 2004 et 2006 pour les pour les reboisements de 25/25 et
2003, 2004 2005 pour les reboisements de 50/50) a fait que l'élaboration
d'un modèle allométrique en fonction de l'âge ne serrait
pas nécessaire.
Ainsi, en termes de séquestration par partie des
plantes, les troncs constituent le principal
réservoir de carbone car
ils séquestrent plus que les branches et les feuilles confondues
(voir
tableau N° 5). La faiblesse de biomasses des branches est due au
jeune âge des plantations
mais, avec le temps leurs sujets développeront davantage
de branches et donc séquestreront aussi davantage de carbone
atmosphérique.
En outre, l'analyse des séquestrations par hectare et
par âge révèle une variation croissante de la
séquestration en fonction de l'âge et selon le type de reboisement
(50/50 ou 25/25). Ainsi pour les plantations d'écartement 25/25 la
séquestration varie de 0,464 t/ha/an à deux ans (2006) à
1,243 t/ha/an à quatre ans (2004) pour atteindre 3,819 t/ha/an à
cinq ans (2003). Cependant, pour les plantations d'écartement 50/50 elle
varie de 0,415 t/ha/an à trois ans (2005) à 0,767 t/ha/an
à quatre ans (2004) pour finir à 1,614 t/ha/an à cinq ans
(2003).
Tableau 13 : R écapitulatif des stocks de carbone
par partie des plantes (Djirnda).
|
ANNEES
|
FEUILLES t de carbone/ha
|
BRANCHES t de carbone/ha
|
FUTS t de carbone/ha
|
|
2003 N° 1
|
3,686
|
2,760
|
12,649
|
|
2003 N°2
|
2,103
|
1,493
|
4,472
|
|
2004 N°1
|
1,396
|
0,791
|
2,786
|
|
2004 N°2
|
0,965
|
0,535
|
1,569
|
|
2005
|
0,616
|
0,202
|
0,621
|
|
2006
|
0,443
|
0,009
|
0,477
|
E.1.4 Dynamique du carbone des plantations :
La dynamique du carbone des plantations de Djirnda part d'un
scénario de production de biomasse nulle à celui d'une grande
productivité de matière végétale et par
conséquent d'une grande capacité de séquestration de
carbone atmosphérique. Cependant, la dynamique du carbone des
plantations de Djirnda est tributaire de leur productivité
végétale, de leur âge, de leur superficie et des conditions
du milieu. L'analyse de la variation des séquestrations de carbone des
plantations de Djirnda révèle qu'elle évolue de
manière exponentielle avec l'âge et cela, avec des
prévisions d'une séquestration totale de 1,420E13
tonnes pour l'ensemble des plantations d'écartement 25/25, contre une
séquestration totale de 8,293E09 tonnes au niveau des
plantations d'écartement 50/50 à trente (30) ans d'âge. En
outre, l'engouement des femmes de Djirnda à poursuivre les campagnes de
reboisement jusqu'au remplissage totale des tannes humides de la zone
présage un bon avenir pour la séquestration de carbone qui
contribuerait pour sa part à la réduction des changements
climatiques. En plus, le degré d'organisation des populations de Djirnda
notamment celle des femmes réunies dans l'AFTRM et leur grande
dépendance aux ressources halieutiques et aux produits de mangrove
65
apparaîtrait comme des critères de
prédisposition du village à accueillir des projets MDP. Ces
critères de prédisposition des populations de Djirnda à
accueillir les projets MDP ne sont pas contraires aux deux principales
conditions d'éligibilité définies par le UNFCCC à
savoir : la réduction des GES par rapport au scénario de base et
l'éligibilité du pays hôte (le Sénégal) en
temps que pays en développement ayant ratifié la convention.
E.2 LES RESULTATS DU SITE DE SANGHAKO
E.2.1 Etat des plantations :
Les plantations de Sanghako sont localisées à
environ 1km au SW du village dans une tanne et
s'étendant sur une superficie totale de 0,14 ha. Elles sont
constituées d'une seule espèce de palétuviers : le
Rhizophora sp. Leur réalisation remonte
à 1999 et 2000 dans le cadre d'un projet de l'UICN de formation et
renforcement de capacité des populations locales en ostréiculture
et en apiculture. Ces plantations correspondent à la première
campagne de reboisement de mangrove de l'UICN dans la RBDS et sont
marquées par un manque de technicité en termes du choix des
propagules et des types de substrats favorables au développement de la
mangrove, mais aussi le choix de la période favorable à la
réalisation des reboisements.
Au plan de survie, les reliques de plants morts trouvés
un peu partout dans les plantations surtout dans celle de 2000
témoignent d'une mortalité considérable. Les
densités (voir tableau N° 15) varient de faible à
très faible avec des valeurs (23.333 pieds/ha en 1999 et 6.000 pieds/ha
en 2000) très au dessous de la densité théorique minimale
d'une plantation réalisée avec un écartement de 50/50 qui
est de 40.000 pieds/ha. Cette densité théorique est tirée
de l'extrapolation à l'hectare de l'hypothèse qu'une placette de
1m2 dans une plantation d'écartement 50/50 contient au
minimum quatre (04) plants s'il n y a pas eu de mortalité. Cette faible
densité s'expliquerait par une imbrication des causes de la
mortalité comme la faible régénération naturelle,
l'attaque des crabes, des insectes et la péjoration climatique. La
dynamique spatiale est pour sa part régressive car en plus de la
mortalité non négligeable, aucune action visant à
poursuivre le reboisement ne s'était entreprise dans le village
jusqu'à ce jour.
Cependant, quelques caractéristiques symboles d'une
bonne adaptation aux conditions du milieu sont notées au niveau de la
plantation de 1999 entre autres la présence des ruches d'abeilles, des
fleurs, propagules et racines échasses ainsi que la haute taille (HT
moyenne supérieure à 2.42 m) des sujets. Ceci témoignerait
d'une certaine adaptation de la plantation
aux conditions de la station malgré l'effet néfaste
du déficit hydrique que connaît la zone depuis quelques
décennies.
Le taux d'humidité (voir le tableau N° 14) varie
d'une partie des plantes à une autre, elle est plus élevée
au niveau des feuilles mais n'atteignant pas 200% avec des moyennes de 131.86%
pour la plantation de 1999 et de 70.85% pour celle de 2000. Au niveau des
branches les taux d'humidité tournent autour d'une moyenne de 66.75%
(1999) et de 52.74% (2000) pas très loin de celle des troncs (fûts
et racines échasses) qui est de 64.34% (1999) et de 53.27% (2000).
Cependant, les coefficients de pondération (voir tableau N° 14
ci-dessous) varient inversement aux taux d'humidité, ils sont plus
faibles au niveau des feuilles que des branches et troncs (fûts et
racines échasses). En outre, la proportionnalité entre la
biomasse fraîche et la biomasse sèche a fait que les parties des
plantes à fort taux d'humidité présentent de faible
biomasse sèche et donc séquestrent moins à l'exemple des
feuilles.
Tableau 14 : taux d'humidit é moyen et coefficient
de pond ération moyen (Sanghako).
|
Année
|
Taux d'humidité moy. par partie des plantes
Ho(%)
|
Coefficient de pondération moy./partie des
plantes.
|
|
feuillages
|
Branches
|
Troncs
|
Feuillages
|
Branches
|
Troncs
|
|
1999
|
131,859
|
66,752
|
64,340
|
0,433
|
0,615
|
0,615
|
|
2000
|
118,095
|
90,278
|
88,782
|
0,307
|
0,523
|
0,534
|

Figure 48 : Ligne de transect (plantation 2000). Figure
49 : Ligne de transect (plantation 1999).
67 Tableau 15 : R écapitulatif des donn ées
caractéristiques de l' état des plantations
(Sanghako).
|
Année
|
Nbre ind.
observés
|
Ecartement
(cm)
|
DB (cm)
|
DmH (cm)
|
HT (m)
|
Densité
(pieds/ha)
|
Recrus
naturels
|
Superficie
(ha)
|
|
1999
|
07
|
N.I14
|
[0,2 ; 2,8]
|
[1,1 ;
|
1,7]
|
[2,18
|
; 2,64]
|
23.333
|
0
|
0,12
|
|
2000
|
03
|
N.I
|
[1,2 ; 2,6]
|
[0,9 ;
|
1,8]
|
[1,22
|
; 1,73]
|
6.000
|
0
|
0,02
|
|
Total
|
|
|
0,14
|
E.2.2 Les régressions allométriques :
La dégradation des plantations et la présence
des trois ruches d'abeilles disposées dans les plantations de Sanghako
ont fait que le nombre d'individus prélevés (07 en 1999 et 03 en
2000) au cours de cette étude est très limite, ce qui compromet
l'utilisation de la régression linéaire multiple au niveau de la
plantation de 2000 qui nécessite un nombre de prélèvement
supérieur au double de l'effectif des régresseurs.
Les équations des modèles mettent en relation la
productivité végétale (biomasse sèche) avec les
paramètres dendrométriques les plus significatifs par
méthode progressive, c'est-à-dire que seuls les paramètres
dendrométriques dont la probabilité de la variable de Student
(Pr>t) et de Fischer (Pr>F) n'atteignant pas le seuil de 5% sont
conservés dans l'équation du modèle.
Dans les développements suivants seront
présentés pour chaque plantation les équations des
modèles, l'analyse du tableau de variances, les résidus et la
statistique de Durbin-Watson (d) afin de juger de la pertinence des
régresseurs utilisés.
Au plan de régression linéaire simple, la
plantation de 1999 ne présente pas de bonnes caractéristiques de
régression en fonction de ses paramètres dendrométriques
(DB, DmH et HT). Les coefficients de détermination (R2)
varient de faible à très faible voire nul (voir les tableaux
N° 18 et 19) selon le type de régresseur (paramètre
dendrométrique) considéré. Ainsi, alors que pour le DB
dont le coefficient de détermination de 0,371 traduit
l'incapacité du modèle à expliquer ne fut ce que la
moitié des observations, ceux des autres régresseurs (DmH et HT)
variant de 0,008 à 0,000 ne pouvant apporter d'explication aux
modèles ainsi définis. En outre, la statistique de Durbin-Watson
pour chacun des régresseurs loin de la valeur normale de deux (02)
traduit une certaine dépendance entre les observations (voir les
14 N.I : Non inventorié.
tableaux N° 18 et 19). Enfin, la probabilité de la
statistique de Fischer au niveau de chacune des régressions est
très au-dessus de la valeur seuil de 5% ce qui traduit un manque de
liaison entre le régresseur considéré et la
productivité végétale.
Cependant, au niveau de la plantation de 2000 le coefficient
de détermination est supérieur à 0,80 pour les
régresseurs tels que le DmH et DB mais peu supérieur à
0,50 pour la HT ce qui traduit une bonne explication des observations par les
modèles. En outre, en termes de statistique de Durbin-Watson (d) seule
la régression avec la HT traduit une indépendance entre ses
observations avec une valeur proche de la valeur normale de deux (02). Enfin,
la probabilité de la statistique de Fischer partout très
supérieurs à sa valeur seuil de 5% laisse apercevoir un manque de
liaison entre le régresseur considéré et son
modèle.
Tableau 16 : R écapitulatif des équations
des modèles lin éaires simples (Sanghako).
|
Année
|
Equations des modèles
|
|
1999
|
Biom sèche(kg) = -0,585 + 0,329*DmH (cm)
-3,847E-02*DB (cm) + 0,441*HT (m)
|
|
Biom sèche (kg) = -0,498 + 5,671E-02*DmH (cm) +
9,377E-02*DB (cm) + 0,572*HT (m)
|
|
Biom sèche (kg) = -4,138E-02 + 0,035*DmH (cm)
-4,398E-02*DB (cm) + 0,222*HT (m)
|
|
2000
|
Biom sèche (kg) = -8,302E-02 + 0,128*DmH (cm)
-6,896E-02*DB (cm) + 0,298*HT (m)
|
|
Biom sèche (kg) = -0,170 + 0,100*DmH (cm)
-3,289E-03*DB (cm) + 0,259*HT (m)
|
|
Biom sèche (kg) = -6,629E-03 + 6,210E-02*DmH
(cm) + 1,075E-02*DB (cm) -4,341E-03*HT (m)
|
Tableau 17 : Caractéristiques des
régressions lin éaires simples (Sanghako).
|
Année
|
Régresseurs
|
Pr Stat de Fischer
|
Stat de Durbin Watson
|
Coef. de dét. (R2)
|
|
1999
|
DB
|
0,147
|
1,372
|
0,371
|
|
DmH
|
0,973
|
0,859
|
0,000
|
|
HT
|
0,852
|
0,844
|
0,008
|
|
2000
|
DmH
|
0,267
|
1,393
|
0,834
|
|
DB
|
0,256
|
1,365
|
0,847
|
|
HT
|
0,484
|
2,028
|
0,526
|
Au plan de régression linéaire multiple et au
niveau de la plantation de 1999 l'équation de régression (voir
tableau N° 20) met en relation la biomasse sèche avec tous les
paramètres dendrométriques (HT, DmH et DB), cela montre que tous
les paramètres dendrométriques contribuent de manière
significative au modèle de la régression. L'équation
explique aussi
69 que la séquestration du carbone au niveau de la
plantation de 1999 est la somme des contributions des différents
paramètres dendrométriques toutes pondérées chacune
d'un certain coefficient de pondération (+2.26 pour DmH, -0.78 pour DB
et -0.35 pour la HT) et corrigées d'un facteur constant de 2.26.
L'analyse du tableau des variances (voir tableau N° 21)
révèle une statistique de Fischer (F) avec une probabilité
associée supérieure à 5%, ce qui justifie
l'hypothèse de non liaison entre la séquestration de carbone et
les variables dendrométriques. Ceci signifie que la séquestration
de carbone n'est pas liée avec les paramètres
dendrométriques mesurés d'où la nécessité
d'envisager de nouveaux régresseurs.
En outre, le coefficient de détermination du
modèle qui est de 0.48 montre que moins de la moitié des
observations ne puissent être interprétées par le
modèle c'est-à-dire que le modèle n'explique même
pas la moitié des observations.
Enfin, la statistique de Durbin-Watson (d=1.40) un peu loin de
la valeur normale (2) témoigne d'une dépendance des observations,
c'est-à-dire que l'ordre des observations pourrait jouer sur les
résultats du modèle.
Tableau 18 : Tableau de la regression
allométrique multiple de la plantation 1999 (Sanghako).
|
Plantation
|
Equations du modèle
|
|
1999
|
Biom sèche (kg) = 2,259 + 1,722*DmH (cm)
-0,785*DB (cm) -0,347*HT (m)
|
Tableau 19 : Recapitulatif des parametres de
l2 equation du modele de la plantation 1999 (Sanghako).
|
Année
|
Régresseurs
|
Pr Stat de Fischer
|
Stat de Durbin-Watson
|
Coef. de détermination
|
|
1999
|
DB, DmH et HT
|
0,526
|
1,39
|
0,48
|
L'analyse des résidus centrés réduits
(voir la figure N° 59) montre qu'il n'existe pas des données
suspectes en termes des mesures des paramètres dendrométriques
faites sur les individus inventoriés. Ceci est justifié par les
valeurs des résidus centrés réduits toutes comprises entre
l'intervalle ]-1,5 ; +1,5[.

70
Ind7
Indio
Ind5
Ind4
Ind3
Ind2
Ind1
Résidus centrés-réduits
-1,5 -1 -0,5 0 0,5 1 1,5
Résidus centrés-réduits
Figure 50 : R ésidus centrés r
éduits plantation 1999 (Sanghako).
E.2.3 Stocks de carbone :
La séquestration de carbone à l'hectare au
niveau des plantations de Sanghako est appréciable malgré leur
dégradation, elle est tributaire des quantités de biomasse
sèche des plantations mais ne traduit pas leur séquestration
exacte car dépendant de la superficie réelle des plantations. En
outre, les quantités de biomasse varient d'une plantation à une
autre et dépendent de plusieurs facteurs entre autre l'âge,
l'écologie des milieux et les conditions climatiques de la zone.
L'analyse comparative des quantités de biomasse (voir tableau N°
16) par hectare des plantations révèle que la plantation de 1999
présente une biomasse sèche d'environ deux fois supérieure
à celle de 2000 qui est de 26.9 t/ha. En raison de l'étroite
relation de proportionnalité entre le carbone atmosphérique
séquestré et la quantité de biomasse sèche, la
plantation de 1999 séquestre environ deux fois la quantité de
carbone séquestré à l'hectare en 2000.
Cependant, la différence de superficies des plantations
a fait que celle de 1999 présente une quantité de biomasse nette
six fois supérieure à celle de 2000 qui est de 0.538 tonnes, ce
qui conduit indubitablement à une séquestration nette de carbone
atmosphérique aussi six fois supérieure à celle de 2000
(0.269 tonnes de carbone). La faible quantité de biomasse nette de la
plantation de 2000 notamment sa faible capacité de séquestration
de carbone par rapport à celle de 1999 pourrait s'expliquer par son
âge mais surtout son état de dégradation.
71
Tableau 20 : Tableau 20 : Biomasses et Stock de carbone
(Sanghako).
|
Années
|
Biomasses fraiches t/ha
|
Biomasses sèches t/ha
|
carbone séquestré
t/ha
|
carbone net
séquestré
(tonnes)
|
|
1999
|
91,570
|
51,103
|
25,551
|
3,066
|
|
2000
|
52,740
|
26,900
|
13,450
|
0,269
|
|
Total
|
3,335
|
Au plan de séquestration par partie des plantes (voir
le tableau N° 17 ci-dessous), les troncs (fûts et racines
échasses) séquestrent plus de carbone que les feuilles et
branches confondues. Au niveau de la plantation 2000, les troncs
séquestrent trois fois plus de carbone que les feuilles et branches
confondues. Les feuilles séquestrent quant à elles plus de
carbone que les branches malgré leur taux d'humidité
supérieur. Les branches apparaîtraient donc comme la partie des
plantes la moins contribuable en termes de séquestration de carbone
atmosphérique. Cependant, cette tendance pourrait être
inversée avec l'âge vu le mode de développement des
palétuviers notamment des Rhizophora sp, qui
développent beaucoup de branches chargées de racines
échasses à l'âge de maturité.
Tableau 21 : Tableau 20 : R écapitulatif des
stocks de carbone par partie des plantes.
|
PLANTATIONS
|
FEUILLES t carbone/ha
|
BRANCHES t carbone/ha
|
FUTS t carbone/ha
|
|
1999
|
5,583
|
4,541
|
15,427
|
|
2000
|
2,001
|
1,846
|
9,586
|
E.2.4 Dynamique du carbone des plantations :
Les plantations de Sanghako présentent un
scénario de base d'une production de biomasse presque nulle en raison du
fait qu'elles sont implantées dans une tanne ne contenant quasiment pas
de biomasse au départ. La dynamique du carbone des plantations de
Sanghako n'est pas très appréciable en raison de leur
altération par les mauvaises conditions de la station telles que :
l'attaque des insectes qui accroît la mortalité des sujets et les
aléas climatiques qui augmentent la salinité de l'eau et de la
vase. En outre, le manque d'analyse du degré d'organisation des
populations du village et l'abandon de la poursuite et de suivi des
reboisements de mangrove ne peuvent encourager le développement
d'éventuels projets MDP. Cependant, l'intervention de JICA/JAFTA dans la
zone notamment à Médina Sanghako peut présager une prise
de conscience des populations en matière de reboisement et de lutte
contre les effets des changements climatiques.
F. DISCUSSIONS
Les discussions portent sur l'état des plantations, la
séquestration de carbone et la dynamique du carbone des plantations. A
l'issue de ces discussions seront comparés les résultats du site
de Djirnda à ceux de Sanghako avant de les rapprocher aux
résultats obtenus au cours d'autres études antérieures
afin d'apprécier la contribution des résultats de cette
étude en termes de production de matière végétale
et de séquestration de carbone.
F.1 Discussion sur l'état des plantations :
Le site de Djirnda tout comme celui de Sanghako sont
implantés non loin des villages (environ 2 km pour Djirnda et 1 km pour
Sanghako). Les plantations présentent des superficies différentes
les unes des autres. La superficie totale des plantations de Djirnda (1,62 ha y
compris celle de 2007) est onze fois plus grande que celle de Sanghako (0.14
ha), ce qui fait que les plantations de Djirnda malgré leur jeune
âge (âge moyen de 3,5 ans) séquestrent plus de carbone
atmosphérique que celles de Sanghako âgées en moyenne de
7,5 ans.
Les principales caractéristiques de la dynamique comme
la densité et la régénération naturelle
révèlent que les plantations de Djirnda sont plus dynamiques que
celles de Sanghako. Aussi, certaines caractéristiques de
l'écologie du milieu comme la présence dans les plantations
d'autres formes de vie (crevettes, huîtres, nids d'oiseaux, abeilles
etc.), ainsi que la présence sur la plupart des plantes des propagules,
des racines échasses et des fleurs montrent qu'il y a une tentative de
mise en place d'un climax au niveau de ces sites.
Comparativement aux résultats de l'étude de
Diédhiou (2006) sur le suivi évaluation des
reboisements de mangrove de la RBDS commandité par l'UICN, les
résultats sur l'état des plantations de djirnda et de Sanghako
s'accordent plus ou moins bien malgré leur différence en termes
d'approche, de démarche et d'objectif poursuivi. Ceci confirme
l'exactitude des résultats et rassure sur le choix des démarches
et des méthodes retenues au cours de cette étude. Mais aussi, un
rapprochement avec les résultats de Ndour (2005) au
plan de la dynamique des peuplements naturels de mangrove de la RBDS montre que
les reboisements présentent des avantages significatifs au plan de
densité vis-à-vis des massifs naturels (densité comprise
entre 1500 à 3600 pieds/ha). Ceci pourrait s'expliquer par le fait que
l'écartement des plants des reboisements est fixé par l'homme
alors que celui des massifs naturels est
73
commandé par la loi des marrées mais aussi les
conditions du milieu qui déterminent la croissance développement
des plantes.
F.2 Discussion sur la séquestration de carbone
et les régressions allométriques :
En raison de l'étroite relation entre la production de
matière végétale d'un écosystème et sa
séquestration de carbone, l'analyse de la productivité des
reboisements de mangrove des sites de Djirnda et Sanghako est directement
liée à l'analyse de leur séquestration de carbone à
travers la relation de séquestration du GIEC (2008) qui
est : C=biom.(sèche)×0,5.
Cependant, l'âge la superficie et l'état des
plantations constituent des facteurs important de la productivité
végétale et donc de la séquestration de carbone
atmosphérique. Ceci explique la faiblesse de la séquestration des
plantations de Sanghako par rapport à celles de Djirnda.
Les séquestrations nettes au niveau des plantations de
Djirnda et Sanghako ne sont pas loin de la séquestration nette annuelle
par unité de surface des peuplements naturels de mangrove de la RBDS
calculée par JICA/JAFTA (2005) qui est d'environ 1
tonne/ha/an. Ceci témoigne de la grande productivité
végétale des plantations de mangrove malgré leur faible
superficie et leur jeunesse.
La recherche des régressions est beaucoup plus
réussie au niveau des plantations de Djirnda qu'au niveau de celles de
Sanghako en raison de l'effectif des individus prélevés (plus de
150 observés à Djirnda contre 10 à Sanghako), de la
superficie des plantations, de leur âge et de leur degré de
dégradation.
En terme de régression linéaire simple, les
modèles de Sanghako présentent en général de bons
coefficients de détermination surtout au niveau de celle de 2000 sauf en
1999 où le coefficient le plus élevé est inférieur
à 0,50. Cette tendance pourrait être inversée avec
l'augmentation du nombre des observations au niveau de la plantation de 2000,
car une régression avec trois observations ne puisse être
significative au sens statistique des termes. Cependant, les régressions
linéaires simples au niveau du site de Djirnda ne présentent pas
de bonnes caractéristiques notamment en termes de coefficients de
détermination sauf la HT qui apparaît comme un assez bon
régresseur par rapport aux autres, notamment au niveau des plantations
2003 écartement 25/25 et 2005 écartement 50/50.
En termes de régressions linéaires multiples, les
modèles de Djirnda contrairement à ceux de
Sanghako
présentent tous une bonne liaison entre la productivité
végétale et les paramètres
dendrométriques
(régresseurs) mais, leur degré d'explication varie d'une
plantation à une
autre et n'atteignant pas 90%. Ceci rend critique la validation
de certains modèles mais témoigne de la singularité des
peuplements de mangrove.
Au plan d'inventaire, l'ordre d'observation des individus
n'enfreigne en rien sur les résultats de l'étude et ce pour la
plupart des plantations de Djirnda et de Sanghako, comme en témoigne
leur valeur de la statistique de Durbin-Watson, ce qui laisse quasiment sans
biais leur plan de sondage.
D'autre part, une étude comparative en termes
d'explication des modèles de régression avec les résultats
de l'étude de Moussa Na Abou (2004) traitant de la
séquestration de carbone et des régressions allométriques
des haies vives des systèmes agroforestiers de Sébikotane,
révèle qu'il ne peu avoir de bonne explication aux
régressions curvilinéaires notamment aux régressions
polynomiales. En d'autres termes, ceci signifie qu'alors que tous les termes
des équations des modèles linéaires ont une signification,
ceux des modèles non linéaires notamment polynomiale n'ont pas
nécessairement de signification à l'exemple du choix des
degrés des polynômes qui malgré tout joue
considérablement sur la valeur des coefficients de détermination.
Enfin, cette difficulté d'interprétation des équations de
Sébikotane laisse comprendre que le choix d'un type de régression
pour une étude donnée ne doit pas être orienté
uniquement par la recherche de grande valeur de coefficient de
détermination mais plutôt par sa facilité à se
prêter à l'interprétation car l'objectif principal de la
recherche des régressions est de montrer clairement la contribution de
chaque régresseur au modèle défini.
F.3 discussion sur la dynamique du carbone des
plantations et les perspectives de
reboisement :
La dynamique du carbone des plantations est mieux
appréciée au niveau de Djirnda que de Sanghako en raison de
l'effectif des plantations (sept à Djirnda contre deux à
Sanghako). La dynamique du carbone des plantations de Djirnda croit de
façon exponentielle avec l'âge, c'est-à-dire qu'avec
l'âge la séquestration nette par année augmente. En plus,
elle varie d'un type de plantation à un autre ce qui fait que la
séquestration nette annuelle des plantations d'écartement 25/25
est supérieure à celle d'écartement 50/50 d'où une
prévision de séquestration de l'ordre des millions de tonnes
à l'âge mature (ici fixé à 30 ans).
Les principales activités génératrices de
revenus au niveau de Djirnda sont la pêche la
cueillette des fruits de
mer, la transformation des produits halieutiques et
l'élevage.
Cependant à Sanghako les populations vivent
principalement de l'agriculture, de la pêche et
75
de l'élevage. Toute fois, quelques activités
subsidiaires comme le ramassage des bois, l'apiculture et autres sont
pratiquées tant à Sanghako qu'à Djirnda.
A Djirnda la gestion des ressources du village est
assurée par l'AFTRM, le comité de plage et le conseil
communautaire alors que le manque de cadre de discussion avec la population de
Sanghako a fait que ses principaux acteurs en termes de gestion des ressources
n'ont pas été identifiés. Les résultats de Djirnda
confirment ceux de JICA/JAFTA (2005) sur le profil
sociologique du village de Djirnda qui présentent le comité de
plage, l'association des femmes et le conseil communautaire comme les
principaux acteurs de la gestion des ressources du village. Cependant, le
degré d'influence de ces acteurs sur la gestion des ressources varie
d'une ressource à une autre, c'est le cas de l'association des femmes
qui dispose de plein pouvoir sur la gestion des reboisements de mangrove et des
fruits de mer et du comité de plage qui veille sur l'application du plan
de gestion des ressources du village sous la houlette du conseil
communautaire.
En outre, les populations ne disposent d'aucune connaissance
ni sur les changements climatiques ni sur leurs effets mis à part
l'avancée de la mer qu'elles reconnaissent tout en ignorant les causes.
De plus elles n'ont aucune connaissance de l'existence des marchés de
carbone ni du MDP mais manifestent cependant un ardent désire d'en
savoir davantage.
Cependant, le degré d'organisation de la population de
Djirnda en termes de prise de décision et de la gestion des ressources
et son engouement à reboiser et à préserver son
environnement constitue pour lui des critères
d'éligibilité aux projets MDP, ce qui n'est pas le cas à
Sanghako qui a encore des efforts à faire.
Enfin, les plantations de Djirnda et de Sanghako sont
éligibles en termes d'adéquation aux principaux critères
d'éligibilité au MDP à savoir le scénario de
référence (Base line) et du pays hôte qu'elles ont en
commun mais à la différence que celles de Djirnda conservent
l'avantage d'avoir une organisation unique chargée de la gestion et du
suivi voire de l'évaluation des ressources des plantations pouvant
abriter d'éventuels projets MDP.
G. CONCLUSION
Les plantations des sites d'étude de Djirnda et de
Sanghako sont toutes situées au SW de leur village
respectif avec des caractéristiques de la dynamique très
particulières. Alors que la dynamique est partout appréciable
à Djirnda elle est un peu critique à Sanghako pour les raisons de
mortalité et de manque de suivi.
En dépit de cette disparité en termes de la
dynamique, les plantations de Djirnda et de Sanghako ont rempli pleinement leur
rôle de puits de carbone en séquestrant malgré leur jeune
âge plus de 4 tonnes de carbone à Djirnda et plus de 3 tonnes de
carbone à Sanghako pour des superficies relativement faibles (1.21ha
à Djirnda et 0.14ha à Sanghako) à l'échelle de
celles des autres types de reboisements connus. Cette forte
séquestration témoigne d'une bonne contribution des reboisements
de mangrove de la RBDS à la séquestration de carbone
atmosphérique. Toute fois, des efforts restent à faire au plan de
superficie, de la recherche des types de substrats adéquats, de la
connaissance de la performance des différentes familles et
espèces de palétuviers et du suivi évaluation.
Les équations des modèles de régression
mettent en relation la contribution de chacun des régresseurs
utilisés, elles ont aussi révélé qu'il y a une
forte liaison entre la productivité végétale et les
régresseurs sauf à Sanghako où le contraire a
été observé ce qui s'expliquerait par le faible effectif
des individus échantillonnés.
En outre, l'analyse des paramètres statistiques des
modèles révèle qu'en général, l'ordre de
prélèvement des échantillons n'a aucun effet sur les
résultats et que les équations mathématiques ne
constituent pas un outil idéal d'estimation des stocks de carbone au
niveau des jeunes plantations, d'où la nécessité
d'accompagner les équations d'inventaires de biomasse. Ceci est
confirmé par la variation des coefficients de détermination ou
degré d'explication des équations des modèles des
différentes plantations.
Enfin, les perspectives de reboisement révèlent
que le site de Djirnda s'annonce plus apte à recevoir des projets MDP en
matière de reforestation car il remplisse certains critères
d'éligibilité du MDP à savoir le degré
d'organisation de sa population, son engagement en matière de
reboisement de mangrove et les atouts qu'il présente en termes de
projets de développement comme le tourisme (écotourisme) et la
transformation des produits halieutiques l'ostréiculture et
l'apiculture.
77
H. SUGGESTIONS
Les suggestions sont principalement axées sur la
réalisation et le suivi évaluation des reboisements, la
séquestration de carbone et les perspectives de développement des
projets MDP de reforestation.
H.1 Suggestion sur la réalisation et le suivi
évaluation des reboisements :
La réalisation des plantations d'écartement
25/25 doit être encouragée par rapport aux autres en raison de
leur forte productivité végétale par unité de
surface mais aussi de leur bonne « santé » au sens
écologique du terme surtout en termes de croissance et
développement.
Il est aussi nécessaire de promouvoir le reboisement
d'autres espèces de palétuviers comme l'Avicennia
sp afin de juger de leur productivité vis-à-vis des
Rhizophora sp mais aussi de garantir une
diversité spécifique au niveau des plantations nécessaire
à l'équilibre écologique de tout le système
mangrove.
Afin de favoriser un échange de connaissances et
d'expériences en matière de reboisement et de gestion des
ressources de mangrove dans la RBDS, il est bon de créer un cadre
d'échange et de discussion entre les principaux acteurs des
différents villages tout en encourageant l'implication des hommes
toujours réticents jusqu'à aujourd'hui à s'investir
pleinement dans le reboisement de mangrove.
H.2 Suggestion sur la séquestration de carbone
et recherche des régressions
allométriques :
Afin d'augmenter le degré d'explication des
modèles de régression c'est-à-dire leurs coefficients de
détermination (R2), il est souhaitable de repenser le choix
des régresseurs en envisageant l'utilisation de nouveaux
régresseurs comme le nombre de branches, la surface foliaire voire le
diamètre de houppiers.
La forte productivité des parties racinaires des
palétuviers deux à trois fois supérieure à celle
des parties aériennes nécessite d'étendre
dorénavant l'évaluation de la séquestration de carbone
jusqu'au niveau des racines afin de permettre une évaluation plus exacte
de la séquestration des peuplements des plantations.
En outre, la particularité du mode d'enfouissement de
la matière organique dans le sol des mangroves a fait qu'il est
nécessaire d'évaluer aussi la séquestration de carbone du
sol en vue de rendre plus complète l'analyse de la contribution de ces
reboisements de mangrove à la séquestration de carbone.
Enfin, pour permettre une bonne manipulation des logiciels et
d'éviter les difficultés de traitement et d'interprétation
comme rencontrées au niveau du site de Sanghako, il est souhaitable de
prélever au minimum un échantillon de trente individus par
plantation.
H.3 Suggestion sur la dynamique des plantations et les
perspectives d'avenir des
reboisements de mangrove :
Les perspectives d'avenir en termes de reboisement de mangrove
au niveau du site de Djirnda sont d'encourager l'association des femmes
à continuer sur la même lancée en les aidant à
développer des projets MDP de reforestation, afin de
bénéficier des renforcements de capacité dans leurs
diverses activités génératrices de revenus.
Cependant, pour le site de Sanghako il est nécessaire
de faire bénéficier la population surtout les femmes du village
d'un voyage d'imprégnation à Djirnda, afin de leur permettre
d'échanger avec les femmes de l'AFTRM sur les techniques de
réalisation mais aussi sur le suivi des reboisements afin de
réduire la mortalité enregistrée au sein de leurs
plantations avant d'envisager un quelconque développement
d'éventuels projets MDP.
Enfin, pour encourager à la reconstitution de tout le
couvert végétal de la mangrove de la RBDS par le biais des
reboisements, il est bon de disséminer les bonnes expériences de
réussite enregistrer au niveau des sites pilotes de Djirnda et
Sanghako.
79
BIBLIOGRAPHIE
Ba Amadou T, Sambou B, Goudiaby A, Ndour N, Mbow C, et Camara
A. 1999. Flore et végétation ligneuse de la réserve de
biosphère du delta du Saloum. Etat actuel, tendances évolutives
et facteurs structurants. Rapport de consultation pour l'élaboration du
plan de gestion de la réserve de biosphère du delta du Saloum
(RBDS), UICN.
Baillon F. (1988). Le complexe hydro-cotier du Sine Saloum :
Synthèse des paramètres écologiques et
socio-économiques de la zone nord du delta du Saloum. ORSTOM. 72
pages.
Balesdent J. 2002. Cycle et temps de résidence du
carbone dans les écosystèmes terrestres, approches isotopiques.
Laboratoire d'écologie microbienne de la rhizosphère Cadarache
France. 26 pages.
Banque Africaine pour le Développement (BAD) et Fonds
Africains de développement (FAD). 1997. Sénégal, profil
environnemental : « Etat de l'environnement du Sénégal
». Revue d'information et de sensibilisation. 41 pages.
Capagna M. 1996. Le cycle du carbone et la forêt : De la
photosynthèse aux produits forestiers. Direction de l'environnement
forestier, Service de l'évaluation environnementale Québec. 56
pages.
Centre de suivi écologique (CSE). 2005. Rapport sur
l'état de l'environnement au Sénégal. Revue d'information
et de sensibilisation.
Dagnélie P. 1992. Statistique théorique et
appliquée (tome 1). Faculté des sciences agronomiques de Gembloux
(Belgique). 492 pages.
Dia M. 2003. Elaboration et mise en oeuvre d'un plan de gestion
intégrée pour la réserve de biosphère du Delta du
Saloum (Sénégal). Union mondiale pour la nature (UICN). 145
pages.
Direction de l'environnement et des établissements
classés (DEEC). 1999. Stratégie nationale initiale de mise en
oeuvre (SNMO) de la convention cadre des nations unies sur les changements
climatiques (CCNUCC). Ministère de l'environnement et protection de la
nature (MEPN). 53 pages.
Doyen A. 1985. La mangrove à usages multiples de
l'estuaire du Saloum (Sénégal). Rapport de consultation. 144
pages.
Gounot M. 1969. Méthodes d'étude quantitative de la
végétation. Livre d'enseignement en inventaire forestier. 305
pages.
Jayaraman K. 1999. Manuel de statistique pour la recherche
forestière. Kerala forest researche institute Peechi, Thrissur, Kerala
(Inde). 239 pages.
Marius C. 1977. Propositions pour une classification et
cartographie des sols de mangroves tropicales. ORTSTOM.
Ministère de l'environnement et de protection de la nature
(MEPN). 1999. Rapport national sur la biodiversité. Rapport de
consultation. 94 pages.
Moussa Na Abou M. 2004. Justification de critères de
durabilité des projets de séquestration du carbone dans la lutte
contre les changements climatiques : Cas des systèmes agro forestiers de
Sébikotane. Mémoire de DEA. Institut des Sciences de
l'Environnement (ISE). 101 pages.
Ndour N. 2005. Caractérisation et étude de la
dynamique des peuplements de mangrove de la réserve de biosphère
du Delta du Saloum (Sénégal). Thèse de doctorat de
troisième cycle. Institut des Sciences de l'Environnement (ISE).170
pages.
Nouidemona J. 2004. Problématique des activités
humaines dans les aires classées : Cas du parc national du delta du
Saloum. Mémoire de DEA. Institut des Sciences de l'Environnement (ISE).
86 pages.
Office des Nations Unies à Genève (ONUG) et le
Secrétariat des Nations Unies sur les Changements Climatiques (UNFCCC).
2008. Protocole de Kyoto à la Convention-Cadre des Nations Unies sur les
Changements Climatiques. Revue d'information et de sensibilisation. 35
pages.
Par l'Office des Nations Unies à Genève (ONUG)
et le Secrétariat des Nations Unies sur les Changements Climatiques
(UNFCCC). Convention Cadre des Nations Unies sur les Changements Climatiques.
Revue d'information et de sensibilisation. 37 pages.
Proisy C. 1999. Apport des données radar à
synthèse d'ouverture pour l'étude de la dynamique des
écosystèmes forestiers. Thèse présentée pour
l'obtention du titre de Docteur de l'Université Paul Sabatier Toulouse 3
spécialités télédétection spatiale de la
biosphère continentale. 103 pages.
Roberts-Pichette P et Gillespie L. 1999. Réseau
d'évaluation et de surveillance écologique « Protocole de
suivi de la biodiversité végétale terrestre ».
Collection des publications hors série de RESE rapport N° 9 Bureau
de coordination du RESE 1999, pour le compte du groupe de travail sur les
protocoles de suivi des végétaux du conseil scientifique de la
biodiversité canadienne. 138 pages.
Secrétariat de la convention Cadre des Nations Unies
sur les Changements Climatiques (UNFCCC). 2008. Guide de la convention-cadre
des nations unies sur les changements climatiques. Revue d'information et de
sensibilisation. 221 pages.
Société internationale pour l'étude des
écosystèmes de mangrove (ISME). 1995. Faite connaissance avec la
mangrove. Revue d'information et de sensibilisation.
Tomassone R. 1989. Comment interpréter les
résultats d'une régression linéaire ? Institut Technique
des Céréales et des Fourrages (ITCF). 55 pages.
Touré A. 2002. Contribution à l'étude de
l'évolution des réservoirs de carbones en zones soudaniennes au
Sénégal. Thèse N° 2585 (2002) présentée
à la faculté environnement naturel, architectural et construit
(ENAC) Ecole polytechnique fédérale de Lausanne (EPEL) pour
l'obtention du grade de docteur ès sciences dans le domaine des sciences
et ingénierie de l'environnement.
UICN bureau régionale pour l'Afrique de l'ouest. 2006.
En route pour la « charte mangroves ». Consultation en
Guinée-Bissau dans le cadre de la préparation d'un accord
sous-régional avec le Sénégal et la Gambie. 2 pages.
Union mondiale pour la nature (UICN). 2002. Programme de
l'UICN pour la conservation des forêts collection vies et paysages
N°1 « Le carbone, les forêts et l'homme » vers une gestion
intégrée du piégeage du carbone, de l'environnement et de
moyens d'existence durables. 50 pages.
www.unfccc.int Site du secrétariat des nations
unies sur la CCNUCC [octobre 2007].
81
www.denv.gouv.sn Site de la
direction de l'environnement du Sénégal [fin septembre et octobre
2007].
http://www.unep.org ou
http://www.unep.net Sites du programme
des nations unies pour l'environnement (PNUE) [septembre 2007].
http://www.epa.gov Site de l'agence
américaine chargée de la gestion de l'environnement [septembre et
octobre 2007].
http://europa.eu.int/comm/environment/
Site de la commission européenne pour
l'environnement [septembre début octobre 2007].
http://www.gtz.de Site de l'agence
allemande pour la coopération technique [septembre 2007].
http://www.ipcc.ch Site du groupe
d'experts intergouvernemental pour le climat (GIEC) [septembre octobre et
début novembre 2007].
ANNEXES
FICHE D'INVENTAIRE
Date Localité Coordonnées
géographiques de la plantation: Latitude ; Longitude:
Numéro de la plantation : .. ; Temps de collecte
des données: début opération: ; fin
opération:
Transect n° :
Code : 1 pour plants réussis ; code 2: pour plants
intrus à noter dans le colonne espèces ; Les impacts sont
notées dans la colonne observations (formes de vie...)
BIOMASSE TOTALE DE LA PLANTATION PAR ESPECE
Date : Localité :
Numéro de la plantation :
:
Transect n° :
BIOMASSE DES ECHANTILLONS PAR PLANTATION
Date : Localité :
Numéro de la plantation : .:
|
N° placettes
|
Espèces
|
Echantillons n° :
|
Bois( g)
|
Feuilles, ... (g)
|
Troncs : fûts &
racines éch.
(g)
|
|
|
Biomasse à l'état frais
|
|
|
|
|
Biomasse à l'état sec
|
|
|
|
|
N° placettes
|
Espèces
|
Echantillons n° :
|
Bois( g)
|
Feuilles, ... (g)
|
Troncs : fûts &
racines éch.
(g)
|
|
|
Biomasse à l'état frais
|
|
|
|
|
Biomasse à l'état sec
|
|
|
|
|
N° placettes
|
Espèces
|
Echantillons n° :
|
Bois( g)
|
Feuilles, ... (g)
|
Troncs : fûts &
racines éch.
(g)
|
|
|
Biomasse à l'état frais
|
|
|
|
|
Biomasse à l'état sec
|
|
|
|
|
N° placettes
|
Espèces
|
Echantillons n° :
|
Bois( g)
|
Feuilles, ... (g)
|
Troncs : fûts &
racines éch.
(g)
|
|
|
Biomasse à l'état frais
|
|
|
|
|
Biomasse à l'état sec
|
|
|
|
|
N° placettes
|
Espèces
|
Echantillons n° :
|
Bois( g)
|
Feuilles, ... (g)
|
Troncs : fûts &
racines éch.
(g)
|
|
|
Biomasse à l'état frais
|
|
|
|
|
Biomasse à l'état sec
|
|
|
|
|
N° placettes
|
Espèces
|
Echantillons n° :
|
Bois( g)
|
Feuilles, ... (g)
|
Troncs : fûts &
racines éch.
(g)
|
|
|
Biomasse à l'état frais
|
|
|
|
|
Biomasse à l'état sec
|
|
|
|
|
|


