|
1
Université de Droit, d'Economie et des Sciences
d'Aix-Marseille III
FACULTE DES SCIENCES ET TECHNIQUES DE
SAINT-JEROME
Institut Méditerranéen d'Ecologie et de
Paléoécologie
Laboratoire de Botanique Historique et
Palynologie
D.E.A. BIOSCIENCES DE L'ENVIRONNEMENT, CHIMIE ET
SANTE
Option Milieux continentaux
RECHERCHE DE L'IMPACT DE LA POLLUTION PAR L'OZONE
SUR LA
CROISSANCE RADIALE DU PIN CEMBRO -Pinus cembra
L.-
DANS LE MASSIF DU MERCANTOUR (ALPES MARITIMES,
FRANCE).
APPROCHE DENDROCLIMATOLOGIQUE.

Mémoire réalisé par Florent
FOURNIER
Sous la direction de M. Jean-Louis EDOUARD
Année universitaire 2000-2001
2
REMERCIEMENTS
Je tiens à remercier M. Jean-Louis EDOUARD pour sa
disponibilité, son aide importante et ses conseils précieux tout
au long de ce stage.
J'adresse également mes vifs remerciements à M.
Frédéric GUIBAL pour m'avoir consacré un peu de son temps
ainsi que M. Thierry KELLER pour sa patience et ses conseils
éclairés.
Je remercie M. André THOMAS et Mme Nicole DENELLE pour
leur aide technique et leurs remarques pertinentes ainsi que Mme Claude
GADBIN-HENRY pour son apport au travers de discussions enrichissantes. Enfin,
je suis reconnaissant envers M. Jacques-Louis de BEAULIEU pour m'avoir
accueilli au sein du LBHP.
3
SOMMAIRE
1. INTRODUCTION 4
2. MATERIELS ET METHODES 6
2.1. Le Pin cembro 6
2.2. Les sites d'étude et leurs
caractéristiques 7
2.2.1. Localisation et contexte géomorphologique 7
2.2.2. Le climat 9
2.3. L'échantillonnage 11
2.4. Acquisition des données et traitement des
données cernes 11
2.4.1. Méthodes de mesure 11
2.4.2. Chronologies élémentaires, individuelles
et maîtresses 13
2.4.3. La standardisation 14
2.5. Les relations cerne/climat : la fonction de
réponse 15
2.5.1. Données climatiques 16
2.5.2. Méthodes de calcul : 16
2.5.3. La signification globale : 17
2.5.4. Procédure de calcul 17
2.6. Recherche des perturbations temporelles de la croissance
radiale dues à la pollution
atmosphérique sur la base d'une modélisation de
la croissance 18
3. RESULTATS 20
3.1. Chronologies maîtresses 20
3.1.1. Séries des largeurs brutes 20
3.1.2. Séries des surfaces des cernes 21
3.1.3. Séries des largeurs indicées 21
3.1.4. Largeur du bois initial 22
3.1.5. Largeur du bois final 22
3.1.6. Densité moyenne du bois initial 23
3.1.7. Densité moyenne du bois final 23
3.2. Fonctions de réponse 24
3.2.1. Modèle sur données
standardisées/Modèle ARMA 24
3.2.2. Fonction de réponse calculée sur la
période (1932-1966) pour les largeurs résidus de la
modélisation ARMA confrontés au couple PTmoy des
stations météorologiques 24
3.3. Recherche de l'influence de la pollution par l'ozone sur la
croissance radiale 28
3.3.1. Analyse des résidus 28
3.3.2. Estimation des largeurs des cernes 29
3.3.3. Récapitulatif 32
4. DISCUSSION 33
4.1. Les chronologies 33
4.2. Les relations cernes/climat 33
4.2.1. Précipitations et températures moyennes
hivernales 33
4.2.2. Les températures de mars, avril et mai 35
4.2.3. Les précipitations et les températures
estivales 35
4.3. Quelle influence de la pollution par l'ozone sur la
croissance radiale ? 36
5. CONCLUSION 39
4
1. Introduction
Depuis le début des années 1980, dans les Alpes
Maritimes et plus particulièrement dans le massif du Mercantour,
l'état de santé du pin cembro est l'objet d'une attention
soutenue. En effet, des jaunissements et défoliations de la partie
supérieure des houppiers ont été signalés sur des
arbres, par les agents du Parc National, dans la vallée du Boréon
(Vésubie).
Une concentration élevée de polluants
atmosphériques (l'ozone en particulier) dans le massif est actuellement
incriminée. Ce gaz s'accumule sur la barrière montagneuse
située dans l'arrière-pays des agglomérations urbaines de
Nice en France et Cuneo-Turin en Italie. L'ozone et ses précurseurs sont
transportés depuis le littoral niçois par exemple. Il est ainsi
constaté dans le sud-ouest du Mercantour l'arrivée d'un front
d'ozone du littoral le jour et la persistance de concentrations
élevées la nuit (Dalstein-Richier, 1997). Cette situation est
classique en montagne : les substances réductrices (NO) qui
détruisent l'ozone la nuit en ville sont absentes car elles restent
confinées dans les vallées du fait des inversions thermiques.
Pour l'Europe, la commission économique des Nations Unies a
défini les « niveaux critiques » en ozone pour la forêt
comme le cumul des dépassements horaires au dessus du seuil de 40 ppb
(ou 80 jig/m3). Cet indice d'exposition appelé AOT 40 correspond
à la somme des différences entre les concentrations horaires et
la valeur de 40 ppb pour chaque heure pendant laquelle la concentration
excède 40 ppb. Cette exposition cumulative est calculée pour les
heures ensoleillées durant une période de 6 mois à partir
du 1er avril. Un seuil critique de 10000 ppb.h au-dessus de 40 ppb
pour cette période a été adopté. Pour la station
expérimentale d'Adrechas-Mercantour (vallée de Mollières,
Tinée), la valeur de l'AOT 40 est dépassée de plus du
double en 2 mois de saison de végétation. En Europe de l'Ouest,
le taux d'ozone a plus que doublé depuis les années 1930 (figure
1).

Figure 1 : Evolution de l'ozone dans l'atmosphère libre
au dessus de l'Europe de l'Ouest durant le XXème siècle,
déduite des mesures effectuées au Pic du Midi et dans diverses
stations européennes d'altitude.
5
L'objectif de l'étude est de déterminer si les
symptômes (jaunissements, défoliation) observés
s'accompagnent d'une perturbation de la croissance des arbres. Il s'agira donc
de rechercher un changement dans la croissance radiale annuelle, plus
précisément de rechercher un éventuel déficit de
croissance significatif des arbres des Cembraies du Mercantour. La croissance
annuelle en diamètre pourrait être considérée comme
un bon indicateur de l'état de santé des arbres. Cependant, les
variations inter-annuelles sont influencées par les fluctuations des
paramètres climatiques. Il est donc nécessaire de séparer
dans les séquences de cernes annuels, la part de variation attribuable
au rythme interne de la croissance et celle liée aux variations
inter-annuelles du climat. En d'autres termes, il est indispensable de mieux
discerner le rôle du climat du rôle éventuel de la pollution
atmosphérique
Une analyse dendroclimatologique a donc été
réalisée pour avoir une image plus complète des variations
des caractéristiques de la croissance radiale et de la
sensibilité climatique du pin cembro au cours du temps. La
première étape a été d'établir les relations
cernes/climat sur une période dite « non polluée »
(1932-1966) pour mieux cerner l'écologie du pin cembro. La
méthodologie mise en oeuvre conduit à l'établissement d'un
modèle statistique des relations cerne/climat basé sur les
variations de haute fréquence. L'analyse porte sur plusieurs
paramètres descripteurs du cerne : l'épaisseur totale du cerne,
l'épaisseur du bois initial et final, la surface des cernes, les
densités moyennes du bois initial et final. La deuxième
étape a été de rechercher un impact de la pollution
atmosphérique (ozone en particulier) sur la croissance radiale pour une
période dite « polluée » (1967-1998) avec comme
période de calibration la période « non polluée
». Une telle méthode est une adaptation de celle employée
auparavant pour étudier l'impact de la pollution fluorée sur la
croissance radiale des conifères en Maurienne par Tessier et
al. (1989).
Cette étude s'inscrit dans un programme de recherche
financé par la Direction Santé des Forêts de l'Office
National des Forêts (2000-2001).
6
2. Matériels et Méthodes
2.1. Le Pin cembro
Existant en Sibérie au Tertiaire, le pin cembro
(Pinus cembra L.), appelé couramment Arolle (en Savoie) ou
Auvier (Briançonnais) est une essence intra-alpine venue de l'est qui
s'est développée aux époques interglaciaires.
L'aire de répartition du pin cembro est restreinte et
morcelée. Elle est principalement située dans les Alpes internes
et intermédiaires. Les plus beaux peuplements de pin cembro se
rencontrent en Autriche, notamment dans la province du Tyrol (Fourchy, 1968).
L'aire de répartition dans les Alpes françaises est très
réduite qui couvre seulement 12500 ha environ est morcelée, les
secteurs les plus étendus sont situés dans le
Briançonnais, le Queyras, et le Mercantour.
Le pin cembro est un arbre très longévif qui
peut vivre jusqu'à 1000 ans (Oswald, 1995). Il présente une
croissance lente et atteint rarement plus de 25 m de hauteur. Monoïque, il
fleurit de juin à juillet. Le rôle du casse-noix est essentiel
dans la régénération du pin cembro. Les graines sont
lourdes mais leur dispersion est assurée par le casse-noix
moucheté (Nucifraga cryocatactes) qui s'en nourrit et qui
constitue des réserves en enfouissant les graines dans des caches
(anfractuosité de rochers, souches, etc). Cet arbre n'est donc pas une
essence colonisatrice mais au contraire une essence apte à former des
forêts stables de longue durée.
Le pin cembro forme des peuplements peu denses, dans la partie
supérieure de l'étage subalpin, en limite supérieure des
forêts des Alpes internes, entre des altitudes échelonnées
entre 1800 et 2500 m. Il est souvent en association avec le
mélèze, plus rarement avec le pin à crochets. Les
peuplements purs, les cembraies, sont assez rares (bois des Ayes dans le
Briançonnais, Col de Salèse dans le Mercantour, par exemple). En
allant vers les Alpes internes, il est en mélange avec
l'épicéa commun et/ou le sapin pectiné. Depuis la fin du
siècle dernier marquée par la déprise agricole en
montagne, le pin cembro est en pleine expansion. Il colonise les espaces
autrefois pâturés dans la zone ouverte entre la limite de la
forêt et la limite supérieure des arbres.
Son métabolisme est adapté aux zones de hautes
altitudes et lui permet de résister aux conditions extrêmes
rencontrées : c'est une espèce à affinité
continentale, qui ne craint pas les écarts de températures
élevés et brutaux, une luminosité vive, une
hygrométrie peu élevée et des vents violents (Contini et
Lavarelo, 1982). Mais le pin cembro a aussi une grande tolérance
à l'égard de l'humidité de l'air et une certaine
répulsion à l'égard d'un renforcement des
caractères méditerranéens du climat tels que la
sécheresse d'été (Fourchy, 1968).
7
Son système racinaire est traçant ce qui lui
permet de s'installer sur des sols très minces- soit parce que la
roche-mère est à faible profondeur, soit parce que le sol «
utile » est peu épais, en raison de la présence d'horizons
pédologiques très lessivés (Fourchy, 1968). A haute
altitude, le cembro est relativement indépendant de l'orientation s'il
est dans son aire de répartition naturelle. Le pin cembro peut occuper
indifféremment toutes les expositions mais avec une prédilection
très nette pour les expositions fraîches (nord et voisines), sans
toutefois qu'on puisse faire la part exacte de l'écologie et de
l'influence humaine qui a été beaucoup plus forte aux expositions
chaudes dans cette répartition. L'homme a aussi longtemps
favorisé le mélèze qui présentait des
qualités plus intéressantes que celles du pin cembro pour
l'élevage et la construction.
2.2. Les sites d'étude et leurs
caractéristiques
2.2.1. Localisation et contexte
géomorphologique
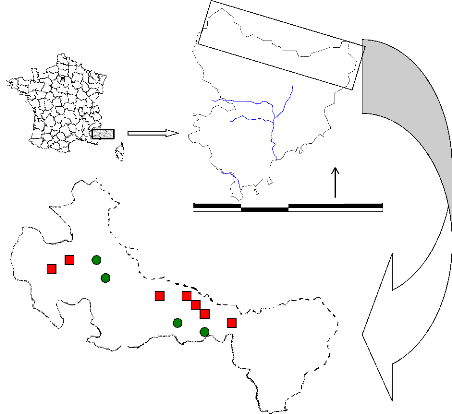
Dans le cadre de cette étude, 7 sites ont
été retenus dans le massif du Mercantour où 7 populations
ont été échantillonnées, respectivement du
nord-ouest au sud-ouest (figure 2) :
> Le vallon de la Braïsse (1) et le Bois de
Sestrière (2), en amont de Saint Dalmas le Selvage, correspondant
à deux placettes d'observation ONF, (dans la vallée de la
Haute-Tinée).
> Le vallon de Chastillon (Isola 2000) (Tinée),
> le versant nord du col de Salèse (1), le versant
sud du col de Salèse (2), le vallon de Salèse (3) (sites de la
Haute vallée de la Vésubie).
> Le vallon de Prals plus au Sud (vallée de la Madone
de Fenestre).
Les altitudes des populations prélevées sont
comprises entre 1900 et 2250 m. 5 populations sont en exposition nord et 2 en
exposition sud sur des versants de pente moyenne, cependant plus forte en
versant nord qu'en versant sud.
Les sites de Salèse, d'Isola et de Prals sont
localisés sur un substrat cristallin (magmatique). Les sites du vallon
de la Braïsse sont sur un substrat gréseux
(sédimentaire).
Quelques très jeunes arbres présentent des
jaunissements et des signes de dessèchement importants. Les arbres les
plus gros, peut-être les plus vieux ont souvent une cime avec des
branches défoliées, et un jaunissement des
extrémités des touffes d'aiguilles sommitales. Les arbres plus
jeunes bien développés présentent un ourlet jaunissant
vers les sommet de la cime. L'observation rapprochée des rameaux de
très nombreux de tous les secteurs visités permet de constater la
présence sur les aiguilles de petites nécroses plus ou moins
nombreuses, jaunes, avec parfois un point marron au centre de la tache.
Alpes de Haute-Provence
0 3 5
7 0
Tinée
Alpes-Maritimes
N
kilometres
St-Dalmas
Auron
ITALIE
N
Braisse
Sestrière
Isola
Salèse
!
1
2
3
Vallon de Prals
Valdeblore
St-Martin-Vésubie
Alpes-Maritimes
8
Figure 2 : Localisation des sites (rouge) et des stations
météorologiques (vert) dans le Mercantour.
Le tableau 1 rassemble les principales caractéristiques
des 5 sites.
Tableau 1 : Caractéristiques principales des sites
échantillonnés.
|
longitude
|
latitude
|
altitude
|
exposition
|
pente
|
substratum
|
végétation
|
|
Salèse 1
|
7°14' E
|
44°08' N
|
2030 m
|
N
|
35°
|
cristallin
|
Cembraie
|
|
Salèse 2
|
7°14' E
|
44°08' N
|
2010 m
|
SW
|
30°
|
cristallin
|
Cembraie
|
|
Salèse 3
|
7°15' E
|
44°08' N
|
1900 m
|
SW
|
20°
|
cristallin
|
Cembro-Larix
|
|
Braisse 1
|
6°48' E
|
44°17' E
|
2230 m
|
NNE
|
25°
|
grès
|
Cembro-Larix
|
|
Sestrière 1
|
6°49' E
|
44°17' E
|
2000 m
|
N
|
25°
|
grès, calcaire
|
Cembro-Larix
|
|
Isola 1
|
7° 09 E
|
44° 11' N
|
2100- 2170 m
|
NE
|
25°-30°
|
granite
|
Cembro-Larix
|
|
Vallon de
Prals
|
7° 21' E
|
44° 04' N
|
2050 m
|
NE
|
25°
|
granite
|
Cembro-Larix
|
9
2.2.2. Le climat
a) Angles de Gams
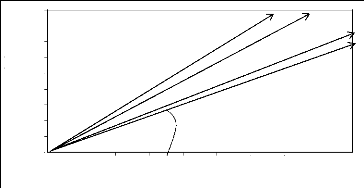
L'angle de Gams (Michalet, 1991), est un descripteur du climat
qui permet une différenciation climatique des sites alpins. Il permet de
s'affranchir de l'effet de l'altitude pour comparer les précipitations
(G est d'autant plus élevé que les précipitations sont
faibles à une même altitude).
Cotg G = P(mm)/A(m)
G=angle de Gams
P=précipitations annuelles (mm) A= altitude (m)
Suivant la valeur de l'angle G, on distingue 3 domaines : >
G<45°: domaine externes des Alpes humides ,
> 45°<G<55°:domaine intermédiaire,
> G>55°: domaine interne « continental ».
Altitude (m)
1800
1600
1400
1200
1000
400
800
600
200
0
0 200 400 600 800 1000 1200 1400 1600 1800
Précipitations (mm)
Auron =57.7 °
St Dalmas =53.6 °
St Martin - Vésubie = 41.5° St Dalmas
Valdeblore= 44.6° Dalmas
angle alpha
Auron
Valdeblore
St Martin - Vésubie
Figure 3 : Angles de Gams calculés
à partir de 4 stations du Mercantour.
Ainsi est constaté un gradient décroissant de
continentalité du nord-ouest au sud-ouest du Mercantour dans lesquelles
se rangent 2 groupes de stations météorologiques : Auron et St
Dalmas au nord-ouest qui sont des stations plus sèches en domaine
interne, St Martin Vésubie et Valdeblore au sud-ouest qui sont des
stations plus humides à la limite Alpes intermédiaires/Alpes
externes. Les stations du sud-ouest sont en effet influencées par la
proximité de la mer.
L'application saisonnière de l'angle de Gams plus
adaptée à l'étude de la répartition de la
végétation (tableau 3) permet d'exprimer l'aridité
estivale :
10
> Ge (G estival) (juin, juillet, août) exprime
l'aridité estivale. A altitude constante, l'aridité estivale est
d'autant plus forte que Ge est élevé et donc que les
précipitations estivales sont faibles.
> Gh (G hivernal) (décembre, janvier,
février) exprime la continentalité hydrique. A altitude
constante, la continentalité hydrique est d'autant plus forte que Gh est
élevé et que les précipitations hivernales sont
faibles.
Tableau 2 : Angles de Gams globaux et
saisonniers.
|
Auron
|
St Dalmas
|
St Martin
|
Valdeblore
|
|
G (°)
|
57.70
|
53.60
|
41.50
|
44.60
|
|
Ge (°)
|
62.11
|
60.77
|
46.91
|
49.66
|
|
Gh (°)
|
60.37
|
56.45
|
45.92
|
50.10
|
Cela permet de différencier nettement la Tinée
(stations Auron et St Dalmas), plus sèche en hiver et en
été, de la Vésubie.
b) Diagrammes ombrothermiques des stations
météorologiques du Mercantour

P (mm)
P (mm)
140
120
100
140
120
100
40
20
80
60
40
20
80
60
0
0
JAN MAR MAY JUL SEP NOV
AN MAR MAY JUL SEP NOV
Auron
aldebloreValdeblore
40
20
70 T°(C)
60
50
30
0
10
40
20
70
T°(C)
60
50
30
10
T°moy
-écart-type
+écart-type
P
P (mm)
Tmoy
-écart-type
+écart-type
P (mm)
P (mm)
140
120
100
140
120
100
40
20
80
60
40
20
80
60
0
0
JAN MAR MAY JUL SEP NOV
AN MAR MAY JUL SEP NOV
St Dalmas
St Martin de Vésubie
40
20
70
60
T°C
50
30
0
10
40
20
70
T°(C)
60
50
30
0
10
Tmoy
-écart-type
+écart-type
Pmoy
P moy T moy
-écart-type
+écart-type
Figure 4 : Diagrammes ombrothermiques des stations
météorologiques du Mercantour.
Cependant, aucune région étudiée ne
présente de mois sec au sens de Gaussen (Pmm<2T) et seule la station
de St Dalmas présente un mois sub-sec (juillet) défini par
2T<Pmm<3T (figure 4).
11
Le massif du Mercantour, situé dans les Alpes internes,
est sous l'influence du climat Méditerranéen. Les pluies sont
abondantes en automne et au printemps, l'été est la
période la plus sèche. Les précipitations
élevées (toujours plus de 1000 mm par an) résultent de
l'influence directe de la mer. Mais, les températures sont
caractéristiques d'un climat de montagne avec des températures
moyennes proches de 0°C l'hiver. Les angles de Gams montrent un gradient
croissant de continentalité du sud-est vers le nord-ouest.
2.3. L'échantillonnage
Sur chaque site, 15 à 20 arbres ont été
prélevés, à raison de 3 carottes par arbre. Les arbres ont
été identifiés sur place par des numéros. Les
arbres présentent tous un état similaire en terme de jaunissement
et de défoliation sans qu'il ait été possible de
définir des classes différentes.
Les sondages ont été effectués à
la tarière de Pressler. Les carottes ont été
prélevées à 120° les unes des autres, si possible
à hauteur de poitrine (1,30 m environ), sinon à des hauteurs
variables en fonction de la pente, de la configuration du terrain et des
anomalies liées à la ramification du tronc ou les blessures.
2.4. Acquisition des données et traitement des
données cernes
2.4.1. Méthodes de mesure
a)Interdatation
Après séchage, les carottes sont poncées
afin que les cernes soient parfaitement visibles. Les cernes sont ensuite
comptés et interdatés. L'interdatation consiste à
attribuer à chaque cerne l'année de sa formation. Le comptage
commence par le cerne directement en contact avec l'écorce ; il s'agit
alors du cerne formé durant l'année en cours. Cette
opération s'effectue par observation comparée des
séquences de cernes, d'abord sur les carottes recueillies sur le
même individu, puis sur l'ensemble des carottes d'une même
population. Pour éviter toute erreur, on utilise les cernes
caractéristiques (cernes très minces) ou des séries de
cernes caractéristiques de l'arbre, synchrones sur l'ensemble des
individus d'une même population (Schweingruber, 1988). Cela permet de
contrôler l'exactitude de la date de chaque cerne. En remontant
jusqu'à la moelle, on peut ainsi attribuer à chaque individu son
âge, du moins son âge minimal si la carotte n'atteint pas la
moelle.
b) 12
Mesure de la largeur des cernes
La mesure de la largeur des cernes s'effectue sous loupe
binoculaire avec une table à mesurer (mesureur d'Eklund), reliée
à un ordinateur sur lequel les mesures sont enregistrées (en
centième de millimètre). Chaque carotte mesurée fournit
une chronologie élémentaire.
c) Mesure densitométrique
Une deuxième méthode permet non seulement de
mesurer la largeur des cernes mais aussi de mesurer des variations
intrannuelles de la densité des cernes.
La structure générale du cerne, chez les
conifères (homoxylés), présente schématiquement
deux zones : le bois initial, d'aspect clair et le bois final d'aspect plus
sombre. Dans les régions à saisons climatiques marquées,
les arbres subissent chaque année une période de repos et une
période d'activité cambiale. Ainsi, de mars à juin
(début de la période d'activité), on observe la formation
du bois initial (zone claire du cerne) correspondant à des
trachéides larges et à parois minces avec un lumen important. La
fin de la période d'activité (de juin à octobre) conduit
à l'élaboration du bois final (foncé), où les
éléments conducteurs sont plus petits et présentent une
paroi plus épaisse (Schweingruber, 1988, Nicault, 1999).
L'analyse densitomérique implique une
préparation spécifique des échantillons conduisant
à l'obtention d'un cliché radiographique des carottes.
Après avoir été débarrassées des traces de
résines par un passage dans un soxhlet à alcool, les carottes
sont débitées en éprouvettes calibrées (lames de
bois) d'1 mm d'épaisseur, perpendiculairement aux trachéides
(Schweingruber, 1988). Seules les carottes dont les trachéides ne
divergeaient de la perpendiculaire de l'axe de la carotte que de moins de
5° sont conservées. Cette sélection sévère est
indispensable pour avoir des radiographies correctes des éprouvettes
c'est à dire pour que les limites des cernes soient parfaitement nettes,
condition nécessaire pour des mesures de densité fiables. Les
lames de bois sont exposées sous une source de rayon X distante de 2.5 m
et pendant une heure à une heure et demie selon l'espèce et
l'épaisseur des échantillons. Les films obtenus sont des
négatifs dont les niveaux de gris correspondent à des
densités plus ou moins élevées du bois. La densité
du bois correspond à la densité absolue de la matière
cellulaire : parois cellulaires (cellulose et lignine), vides (lumen) et
proportion cellulose/lignine. L'analyse densitométrique consiste donc
à mesurer les variations des niveaux de gris (densité optique) et
à les convertir, grâce à une calibration préalable,
en densité. Il est obtenu au final, dessiné sur l'écran de
l'ordinateur auquel est couplé le microdensitomètre, le profil
détaillé des variations de densité du cerne.
Pour chaque éprouvette, 7 séries de
données-cerne annuelles sont fournies par le densitomètre, ce
sont :
13
- la largeur totale du cerne complet, la largeur du bois initial
et celle du bois final exprimées en microns. - les densités
maximale et minimale exprimées en kg/m3
- les densités moyennes du bois initial et du bois final
exprimées en kg/m3.
La structure du cerne est donc appréhendée ci
à travers l'analyse densitométrique. La limite bois initial /
bois final est définie, pour chaque cerne, en fonction de sa
densité minimale et maximale (figure 5).
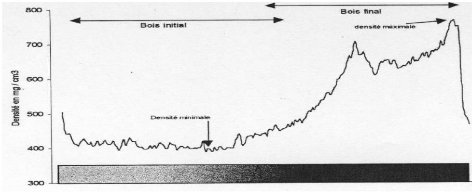
Figure 5 : Variations intrannuelles de densité d'un cerne
de croissance.
Les mesures de densité ont été faites
pour 5 populations. Mais le traitement des données ne porte dans ce
travail que sur 3 populations.
2.4.2. Chronologies élémentaires,
individuelles et maîtresses a) Définitions
A partir des mesures de la largeur des cernes des carottes,
trois types de chronologies sont
construites :
> Les chronologies élémentaires :
Elles représentent la séquence chronologique des
largeurs de cernes de chacune des carottes.
> Les chronologies individuelles : Elles
représentent la croissance moyenne d'un individu en fonction du temps et
sont construites en faisant la moyenne des séries
élémentaires d'un même individu.
> Les chronologies maîtresses :
Elles sont réalisées à partir de la moyenne de
toutes les séries élémentaires d'une population. Elles
représentent le patron général de la croissance de la
population du site étudié. Ces chronologies suppriment ou, du
moins, minimisent les variations atypiques propres à chaque arbre et
dues à des phénomènes particuliers, le plus souvent
micro-stationnels. Les variations inter-annuelles des cernes observées
sont dues essentiellement au climat (Fritts, 1976, Nefaoui, 1996). On peut
également construire des chronologies en utilisant les
échantillons de plusieurs populations
14
situées dans un site géographique donné.
On obtient alors une chronologie maîtresse régionale où
ressortent les variations communes à toutes les populations. Il est
alors possible de voir si certaines variations sont à l'échelle
régionale ou stationnelle.
b) Recherche du synchronisme entre les
chronologies
La superposition des chronologies élémentaires
d'un même individu permet une première vérification
visuelle de l'interdatation et donc du bon synchronisme des chronologies. Des
corrections pourront être apportées si, par exemple, des cernes
sont manquants sur une partie de la circonférence. Ainsi en
théorie, la largeur des cernes est identique pour une année
donnée, dans une population d'arbres soumis aux mêmes conditions
environnementales
Une autre méthode pour vérifier la bonne
synchronisation des chronologies ainsi construites est de calculer le
pourcentage de coïncidence ou d'effectuer un test de Student sur des
valeurs transformées (test de Baillie-Pilcher). Ce test permet
d'apprécier l'homogénéité de comportement des
arbres échantillonnés.. Le pourcentage de coïncidence se
définit comme le pourcentage de cas où la variation entre deux
cernes consécutifs se fait dans un sens identique pour les deux courbes.
Ce test ne tient donc compte que du sens de variation de l'épaisseur des
cernes d'une année sur l'autre ou pente des écarts inter-annuels.
Ce calcul exprime donc la concordance des variations inter-annuelles des deux
chronologies. Ces tests sont exécutés avec un sous-programme du
logiciel de dendrochronologie TSAP.
2.4.3. La standardisation
La standardisation est effectuée pour éliminer
des variations inter-annuelles l'effet à long terme de l'âge et
l'ensemble des bruits d'origines différentes pour ne conserver que la
part de la variance de haute fréquence principalement due aux facteurs
climatiques. Ceci permet aussi la comparaison de courbes issues d'arbres
d'âges différents comme c'est le cas dans les populations
étudiées. Deux méthodes de standardisation ont
été utilisées : celle des indices (ou indexation) et celle
des résidus (modélisation ARMA).
a) L'indexation
La standardisation utilisée ici consiste à
convertir les épaisseurs brutes en indices. La méthode
employée a consisté à utiliser un filtre, avec une
fenêtre permettant de conserver les fréquences souhaitées.
Un filtre passe bas assorti d'une fenêtre de 10 ans a été
adopté, pour conserver les variations de haute-fréquence
principalement dues aux variations inter-annuelles du climat. Puis les indices
ont été obtenus en substituant à chaque largeur de cerne
mesurée le quotient de la largeur annuelle observée du cerne par
la valeur filtrée de celui-ci.
15
b) La modélisation ARMA (Auto Regressive
Moving Average)
Cette méthode de standardisation a été
privilégiée dans cette étude car le signal
aléatoire climatique est plus fort que celui fournit par l'indexation.
Le processus ARMA est basé sur la décomposition de la croissance
Ct en une partie aléatoire at (ou résidu) et une partie,
prédéterminée, fonction de la croissance antérieure
aux temps t-1, t-2,...t-p. On peut écrire :
Ct = f(Ct-1, ..., Ct-p)+at
Ct : grandeur brute (épaisseur ou surface) du cerne au
temps t
f (Ct-1,..., Ct-p) : part du cerne de l'année t dont la
construction est prédéterminée avant même que
n'intervienne le facteur climatique de l'année t
La partie prédéterminée peut s'exprimer
sous la forme d'un processus autorégressif d'ordre p noté AR(p)
qui prend en compte la grandeur des cernes des années t-1, t-2,...,t-p,
soit :
Ct = Ö1Ct-1+Ö2C t-2+...+ÖpCt-p+at avec Öi :
coefficients « autorégressifs »
La modélisation ARMA prend en compte les deux facteurs
intrinsèque et extrinsèque majeurs de la croissance que sont la
tendance d'âge et le climat.
Chaque série moyenne de cernes est d'abord
modélisé par un processus de type ARMA et adapté à
l'analyse des séries temporelles que constituent les cernes (Guiot
et al., 1982). Le programme CALARE du logiciel 3Pbase a
été utilisé. Un modèle simple
autorégressif 1-0 (autocorrélations calculées
avec le programme GALOTO du logiciel 3PBASE), dans la plupart des cas
étudiés ici, suffit à expliquer la presque totalité
de la variance liée à l'autocorrélation des cernes.
L'épaisseur du cerne au temps t est donc toujours exprimée sous
la forme :
Ct = Ö1 Ct-1 + at
Le premier terme, lié à l'autocorrélation
des cernes, exprime le degré d'inertie de la réponse des arbres
aux facteurs externes. Le dernier terme (at) aléatoire
appelé résidu, lié à l'année
où se construit le cerne, exprime la réponse des arbres aux
facteurs externes et en particulier au climat. Les séries de
résidus sont ainsi isolées par la modélisation ARMA.
2.5. Les relations cerne/climat : la fonction de
réponse
Les arbres sont des enregistreurs plus ou moins fidèles
de la variabilité des facteurs externes dont le climat. L'objectif est
de trouver une équation statistique traduisant au mieux la relation
entre les différents paramètres climatiques
(précipitations et températures essentiellement) et la croissance
des individus représentée par les séries temporelles des
largeurs de cernes : c'est la fonction de réponse. Elle
16
est un moyen d'apprécier l'aptitude d'un modèle
statistique à prédire la part de variation de la largeur des
cernes attribuable aux facteurs environnementaux dont le facteur climatique.
Une méthode a été utilisée : celle
de la régression orthogonalisée à laquelle a
été appliquée la procédure Bootstrap.
2.5.1. Données climatiques
L'analyse de la relation cerne/climat nécessite
l'utilisation de données météorologiques locales
complètes pour la période sur laquelle l'analyse est faite.
Le tableau 2 réunit les stations
météorologiques utilisées. Les séries de
température (maximales et minimales) couvrent une période
d'environ 40 ans. Mais les stations du nord du massif étant
significativement corrélées avec la station de Gap et d'autres
stations proches, les données manquantes ont pu être
reconstituées sur la base de corrélations multiples. La
température moyenne est obtenue en faisant (Tmin+Tmax)/2.
Tableau 3 : Postes météorologiques.
|
poste
|
longitude
|
latitude
|
altitude
|
Précipitations
|
T°min
|
T°max
|
|
Gap
|
6° 05' E
|
44° 34' N
|
750 m
|
1857-1998
|
1878-1998
|
1878-1998
|
|
St Dalmas le Selvage
|
6° 51' E
|
44° 17' N
|
1510 m
|
1931-1998
|
1951-1981
|
1951-1981
|
|
Auron
|
6° 56' E
|
44° 13' N
|
1610 m
|
1950-1998
|
1951-1998
|
1951-1998
|
|
St Martin Vésubie
|
7° 15' E
|
44° 04' N
|
1000 m
|
1952-1998
|
1959-1998
|
1959-1998
|
|
Valdeblore
|
7° 10' E
|
44° 04' N
|
1000 m
|
1931-1998
|
1954-1998
|
1954-1998
|
|
Peira Cava
|
7° 22' E
|
43° 56' N
|
1480 m
|
1954-1997
|
1956-1997
|
1956-1997
|
|
St Dalmas-de-Tende
|
7° 35' E
|
44° 03' N
|
650 m
|
1938-1998
|
1951-1998
|
1951-1998
|
2.5.2. Méthodes de calcul :
Le programme CALROB (logiciel PPPhalos) est utilisé
pour calculer les fonctions de réponse. Les différentes
étapes de calcul sont les suivantes :
> La régression orthogonalisée : La
variable dépendante est le cerne exprimé par la largeur ou la
densité. Les variables explicatives (ou régresseurs) sont les
paramètres climatiques mensuels: précipitations et
températures. L'analyse est une régression multiple se faisant
sur les composantes principales des régresseurs, afin d'éliminer
la corrélation présente entre les paramètres climatiques.
Cette régression est dite orthogonalisée.
> La méthode Bootstrap : Cette méthode
(Guiot, 1991) permet de tester la fiabilité de la fonction de
réponse sans utiliser les tests d'hypothèse classiques. Elle
estime les erreurs des coefficients de régression de la fonction de
réponse en étudiant la variabilité des estimations faites
à partir de sous-ensembles du jeu de données initiales. Ces
échantillons sont obtenus par tirage aléatoire avec remise parmi
les observations initiales (Keller, 1999). Ici, le calcul de la
régression a été répété cinquante
17
fois (50 tirages) sur le même ensemble de variables. Les
fonctions de réponse sont calculées sur ces années dites
de calibration et la relation obtenue est vérifiée avec les
années non tirées au sort (années de
vérification).
2.5.3. La signification globale :
Celle-ci est évaluée sur la base de deux
paramètres : la valeur moyenne des coefficients de corrélation
multiple relatifs aux années de calibration (RMC) et celle des
coefficients de corrélation multiple relatifs aux années de
vérification (RMV). La valeur du RMV au carré exprime le
pourcentage de variance expliquée par la régression, tandis que
la valeur Rv, donnée par le rapport entre RMV et son écart type,
permet d'apprécier la signification de la fonction de réponse. Si
l'on suppose que Rv présente une distribution normale, son degré
de signification est établi selon les seuils suivants :
1.65 < Rv < 1.96 significatif à 90 %
1.96 < Rv < 2.58 significatif à = 95 %
2.58 < Rv < 3.29 significatif à = 99 %
Rv > 3.29 significatif à > 99 %
De la même façon, le rapport entre chaque
coefficient de régression partiel moyen et son écart-type (r/s)
précise le niveau de signification de la relation (directe ou inverse,
selon le signe positif ou négatif) entre la croissance et le
paramètre climatique mensuel considéré. Une relation
directe (+) associe une augmentation de l'épaisseur du cerne à
une augmentation de la variable climatique et inversement.
2.5.4. Procédure de calcul
L'analyse en composante principale et matrice de
corrélations sur les mesures indicées, à l'aide du
programme GLCPC du logiciel 3Pbase et du logiciel SPAD a montré que les
carottes étaient homogènes et qu'il n'y avait pas d'erreurs
d'interdatation.
La procédure des calibrations cernes/climat comporte
deux phases :
Tout d'abord les fonctions de réponse ont
été calculées avec une combinaison de 24
régresseurs (12 précipitations et 12 températures) sur une
période allant du mois d'octobre de l'année t-1 au mois de
septembre de l'année t (année « biologique » couvrant
la phase antérieure à l'élaboration du cerne annuel,
d'octobre t-1 à juin de l'année t de fonctionnement cambial, et
la phase de construction du cerne de l'année t de juin à
septembre) et sur toutes les séries chronologiques individuelles
constituant
18
chacune des populations. Le couple de régresseurs est
constitué par les précipitations et les températures
moyennes mensuelles.
En raison de l'homogénéité de
comportement des individus de chaque population, la suite des calculs a
été effectuée directement sur la chronologie
maîtresse de chaque population. Puis dans un second temps, le
modèle est optimisé en réduisant le nombre de
régresseurs (Keller, 1999). Les paramètres climatiques sont
regroupés en respectant à la fois, la phénologie de
l'essence, telle qu'elle peut être traduite en termes saisonniers et le
sens des relations dégagées entre le cerne et les
paramètres explicatifs.
2.6. Recherche des perturbations temporelles de la
croissance radiale dues à la pollution atmosphérique sur la base
d'une modélisation de la croissance
Deux périodes ont été définies,
d'égale durée, la première (1932-1966) étant prise
comme période de référence, au cours de laquelle on peut
considérer que le niveau très bas de pollution n'a eu qu'un effet
minimal sur la production ligneuse annuelle (période « non
polluée »), la seconde (19671998) correspondant à la
période marquée par un maximum de pollution, au cours de laquelle
la production ligneuse peut avoir été affectée
significativement (période « polluée »).
Dans des conditions « normales », hors pollution
(période 1932-1966), l'inertie des arbres aux facteurs externes
(exprimée par le coefficients Ö1) est au moins partiellement
attribuée à la géométrie du cerne et au potentiel
biologique de l'arbre dans son biotope, lui même lié à
l'âge. La composante climatique est exprimée par la série
des résidus at découlant du modèle
précédent.
Dans la période avec pollution (1967-1998), l'inertie
des arbres aux facteurs externes peut-être considérée comme
modifiée par les perturbations vraisemblablement induites par la
pollution : jaunissement des aiguilles, défoliation, modification
progressive de la biologie de l'arbre. Par voie de conséquence
l'intervention du climat au temps t (exprimée par les résidus at)
est également modifiée.
La relation cerne-climat est ensuite cherchée sur
chacune des 2 périodes par le calcul d'une fonction de réponse.
La variable dépendante est la série des résidus fournie,
par la modélisation ARMA réalisée à la
première étape. Le calcul donne donc, une série de 24
coefficients de régression, différents d'une période
à l'autre puisque, malgré une action du climat que l'on peut
considérer comme stable au cours des deux périodes prises en
compte, les résidus a't de la deuxième période expriment
une relation de « l'enregistreur » arbre à ce climat,
perturbée par la pollution. L'ultime étape consiste à
comparer, sur l'ensemble de la période 1932-1998, la série brute
des épaisseurs de cerne mesurées avec une série
estimée reconstruite (programme FILARE du logiciel 3Pbase) sur la base
des coefficients calculés pour
19
la période « non polluée » de
calibration, tant au niveau de la fonction de réponse qu'à celui
de la modélisation ARMA. (Tessier et al., 1990, Gandolfo et
al., 1994).
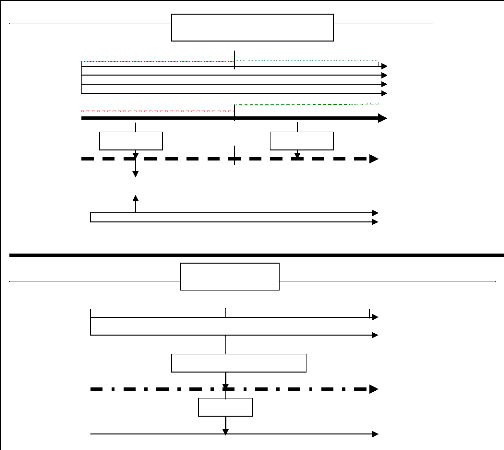
Modélisation ARMA (1-0), programme CALARE
Régression sur le climat, programme CALROB
Interdatation mesure
Moyenne des épaisseurs observées
Reconstruction
C1 à partir du climat
Reconstruction
à partir du modèle
1932 1966 1998
1932 1966 1998
24 coefficients de régression
Ö1
Séries météo,12 mois P, 12 mois T.
24 coefficients de régression
Reconstruction
Analyse et calibration
Ö1
Ö'1
Séries météorologiques 12 mois P 12 mois
T.
Chronologie des résidus estimés
Chronologies élémentaires
Chronologie de synthèse
Chronologie des épaisseurs estimées
Figure 6 : Organigramme du traitement des
séries chronologiques de cernes utilisés pour mettre en
évidence les variations de la croissance radiale annuelle attribuables
à la pollution. (d'après Tessier et al., 1990,
modifié).
20
3. Résultats
3.1. Chronologies maîtresses
L'analyse dendroécologique pratiquée ici porte sur
des arbres âgés d'au moins 100 ans. Les pins cembro du vallon de
la Braisse sont très vieux (certains ont plus de 600 ans), ceux de
Sestrière, du col de Salèse (1 et 2), d'Isola et du vallon de
Prals ont plus de 250 ans alors que ceux de Salèse 3 sont plus jeunes,
moins de 200 ans.
Les chronologies ont été reconstruites sur la
période 1870-2000.
3.1.1. Séries des largeurs brutes

Largeurs (1/100e mm).
400
350
300
250
200
150
100
50
0
1870 1875 1880 1885 1890 1895 1900 1905 1910 1915 1920 1925 1930
1935 1940 1945 1950 1955 1960 1965 1970 1975 1980 1985 1990 1995
Figure 7 : Chronologies moyennes des largeurs
de cerne, populations de Salèse (1-2-3), Sestrière, Braisse,
Isola, Prals.
L'allure de ces différentes courbes est sensiblement la
même pour les différentes populations sur une période
identique, les populations étant toutes sauf une composées
d'arbres de plus de 200 ans. En outre, la variable largeur est clairement
synchronisée sur le signal de haute fréquence. Seule
Salèse 3 se distingue par une croissance plus forte, les arbres
étant plus jeunes.
Tableau 4 : Interdatation des chronologies moyennes des 7
populations de pin cembro (% de coïncidence, test de Baillie-Pilcher).
|
Sestrière
|
Braisse
|
Salèse 1
|
Salèse 2
|
Salèse 3
|
Isola 1
|
|
Prals
|
|
Sestrière
|
|
-
|
|
|
|
|
|
|
|
|
|
|
|
|
Braisse
|
83%,
|
6.80
|
|
-
|
|
|
|
|
|
|
|
|
|
|
Salèse 1
|
68 %,
|
4.84
|
79%,
|
5.56
|
|
-
|
|
|
|
|
|
|
|
|
Salèse 2
|
71%,
|
5.81
|
73%,
|
4.67
|
79%,
|
8.3
|
|
-
|
|
|
|
|
|
|
Salèse 3
|
67%,
|
4.46
|
71%,
|
4.2
|
77%,
|
6.36
|
81%,
|
7.67
|
-
|
|
|
|
|
|
Isola 1
|
70%,
|
6.8
|
74%,
|
9.1
|
80%,
|
8.9
|
71%,
|
8.3
|
71%,
|
6.2
|
-
|
|
|
|
Prals
|
65%,
|
5.9
|
69%,
|
4.6
|
78%,
|
6.8
|
77%,
|
6.9
|
69%,
|
7.5
|
77%,
|
6.5
|
-
|
21
Le pourcentage de coïncidence entre les 7 chronologies
est toujours supérieur à 65%. Tous les coefficients
Baillie-Pilcher calculés pour les populations du Mercantour sont
significatifs à 99%. Ces différentes populations
présentent donc un niveau élevé de synchronisme.
3.1.2. Séries des surfaces des
cernes
Surfaces (mm2)
4500
4000
3500
3000
2500
2000
1500
1000
500
0
1870 1875 1880 1885 1890 1895 1900 1905 1910 1915 1920 1925 1930
1935 1940 1945 1950 1955 1960 1965 1970 1975 1980 1985 1990 1995
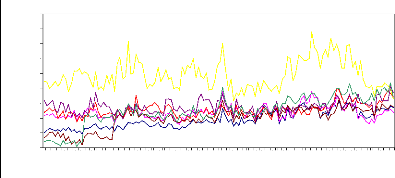
Figure 8 : Chronologies moyennes des surfaces
de cerne, populations de Salèse (1-2-3), Sestrière, Braisse,
Isola, Prals.
Les chronologies de surface des cernes des populations
d'arbres vieux témoignent d'une tendance légèrement
croissante sur les dernières 130 années. Au cours de la
période des 20 dernières années, on observe une
décroissance marquée (minimum vers 1990), plus accentuée
pour la population des arbres plus jeunes, suivie d'une reprise.
3.1.3. Séries des largeurs indicées
Indices
4.5
4.0
5.0
3.5
3.0
2.5
2.0
0.5
0.0
1.5
1.0
1870 1875 1880 1885 1890 1895 1900 1905 1910 1915 1920 1925 1930
1935 1940 1945 1950 1955 1960 1965 1970 1975 1980 1985 1990 1995
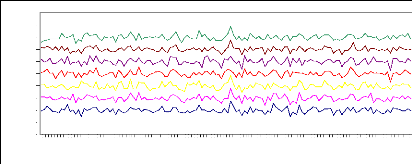
Figure 9 : Chronologies moyennes des largeurs
standardisées, populations de Salèse (1-2-3), Sestrière,
Braisse, Isola, Prals.
Le synchronisme des chronologies maîtresses
indicées des 7 populations est visuellement très fort. Les
populations présentent les mêmes fluctuations de largeurs de
cernes et ce, tout au long de leur chronologie.
22
3.1.4. Largeur du bois initial

Largeur du bois initial (1/100 mm).
140
120
100
40
80
60
20
0
1870 1880 1890 1900 1910 1920 1930 1940 1950 1960 1970 1980
1990
Figure 10 : Chronologies moyennes des largeurs du bois
initial, populations de Salèse (1), Sestrière et Braisse.
Le synchronisme entre les populations est aussi fort. Les
populations Salèse 1 et Sestrière ont des fluctuations de haute
et basse fréquence très similaires Les amplitudes inter-annuelles
sont moins marquées pour la population vallon de la Braïsse,
constituée de très vieux arbres Cette dernière population
montre des fluctuations de moyenne fréquence pluridécennales
nettes.
3.1.5. Largeur du bois final

Largeur du bois final (1/100 mm)
45
40
50
35
30
25
20
15
10
5
0
1870 1880 1890 1900 1910 1920 1930 1940 1950 1960 1970 1980
1990
Figure 11 : Chronologies moyennes des largeurs du bois final,
populations de Salèse (1) Sestrière et Braisse.
L'épaisseur du bois final ne montre pas de tendance
bien nette même si on observe une légère augmentation
depuis 1940. Il est constaté un bon synchronisme entre les populations.
Les valeurs des variations des largeurs ont une forte amplitude inter-annuelle
(allant de 10 à 50 centièmes de mm).
23
3.1.6. Densité moyenne du bois initial
densité moy du bois initial (kg/m3).
440
420
400
380
360
340
320
300
1870 1880 1890 1900 1910 1920 1930 1940 1950 1960 1970 1980
1990
Figure 12 : Chronologies moyennes des densités moyennes du
bois initial, populations de Salèse (1) Sestrière et Braisse.
Le synchronisme visuel est satisfaisant et la densité
moyenne du bois initial de la population Salèse 1 est significativement
plus élevée que celle des deux autres populations.
3.1.7. Densité moyenne du bois final
densité moyenne du bois final (kg/m3)
450
400
700
650
600
550
500
Figure 13 : Chronologies moyennes des densités moyennes du
bois final, populations de Salèse (1) Sestrière et Braisse.
Il est constaté un bon synchronisme entre les courbes
de densité moyenne du bois final. Les fluctuations inter-annuelles ont
moins d'amplitude que celle pour la densité moyenne du bois initial. La
densité du bois final de la population Salèse 1 est aussi plus
élevée que celle des deux autres populations.
24
3.2. Fonctions de réponse
3.2.1. Modèle sur données
standardisées/Modèle ARMA
Les fonctions de réponse ont été
calculées en utilisant les données brutes indicées et les
résidus ARMA. L'adéquation du modèle (coefficient de
corrélation moyen des années de
vérification/écart-type) avec les données ARMA
étant meilleur, seuls les résultats avec ARMA seront
présentés (cf tableau 5).
Tableau 5 : Coefficients de
corrélation multiple de calibration et de vérification et
signification globale des fonctions de réponse (exemple pour la
population de Sestrière).
|
Coefficient de
corrélation
calibration
RMC
|
Coefficient de
corrélation
vérification
RMV
|
Ratio
RMV/ETV
=Rv
|
Signification
globale
|
|
Sestrière ARMA,
épaisseur
|
0.725
|
0.644
|
8.02
|
> 99 %
|
|
Sestrière
standardisation,
épaisseur
|
0.78
|
0.43
|
2.68
|
=99%
|
3.2.2. Fonction de réponse calculée
sur la période (1932-1966) pour les largeurs résidus de la
modélisation ARMA confrontés au couple PTmoy des stations
météorologiques
a) Calculs avec 24 régresseurs pour les
paramètres largeur et surface Dans l'ensemble, les
réponses des populations sont homogènes (Figures 14 et 15).
> Les précipitations : la largeur des cernes et la
surface montrent une relation directe avec les précipitations des mois
d'hiver. La relation avec les précipitations de juin est inverse.
> Les températures moyennes : la largeur est en
relation inverse avec les températures des mois de février, mars.
La surface est en relation inverse avec les températures de mars, avril.
La largeur et la surface des cernes ont une relation directe avec les
températures de juin.
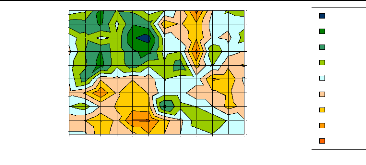
r/s
OCT NOV DEC JAN FEV MAR AVR MAI JUN JUL
AOUT SEP
sa1-P
ses-P
sa1-T
ses-T
Prals-P
iso-P
br-P
Prals-T
iso-T
br-T
3.2-4 2.4-3.2 1.6-2.4 0.8-1.6 0-0.8 -0.8-0 -1.6--0.8 -2.4--1.6
-3.2--2.4
Figure 14 : Signification globale des fonctions de
réponses pour les largeurs sur la période octobre t-1 - septembre
t pour chaque population du Mercantour calculées sur le
couple de paramètres précipitations et températures
moyennes.
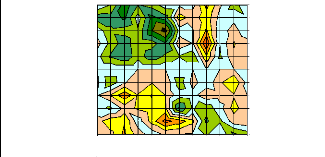
r/s
OCT NOV DEC JAN FEV MAR AVR MAI JUN JUL AUT SEP
sa1-P
ses-P
sa1-T
ses-T
prals-P
iso-P
br-P
prals-T
iso1-T
br-T
4-4.8 3.2-4 2.4-3.2 1.6-2.4 0.8-1.6 0-0.8 -0.8-0 -1.6--0.8
-2.4--1.6 -3.2--2.4
Figure 15 : Signification globale des fonctions de
réponse pour les surfaces sur la période octobre t-1 - septembre
t pour chaque population du Mercantour calculées sur le
couple de paramètres précipitations et températures
moyennes.
b) Calculs avec 24 régresseurs pour les
paramètres densitométriques
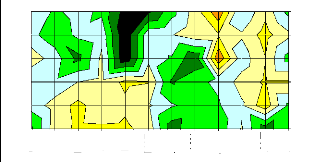
r/s
OCT
NOV
DEC
JAN
FEV
MAR
AVR
MAI
JUIN
JUIL
AUT
SEP
sal1P
sesP
sal1T
sesT
brP
brT
2.4-3.2 1.6-2.4 0.8-1.6 0-0.8 -0.8-0 -1.6--0.8 -2.4--1.6
-3.2--2.4
25
Figure 16 : Signification globale des fonctions de
réponses de la largeur du bois initial sur la période octobre t-1
- septembre t pour chaque population du Mercantour calculées
sur le couple de paramètres précipitations et températures
moyennes.
On constate une relation inverse avec les températures
d'hiver pour toutes les populations, une relation directe avec les
températures moyennes d'avril, mai et août pour la population de
Sestrière seulement ainsi qu'une relation inverse avec les
températures d'août (pour Braisse et Salèse 1). La largeur
du bois initial est positivement corrélée avec les
précipitations d'hiver et négativement avec les
précipitations de juin.

r/s
OCT NOV
DEC
JAN
FEV
MAR
AVR
MAI JUN
JUL
AOUT
SEP
sal1P
sesP
sal1T
sesT
brP
brT
1.6-
2.4
0.8-
1.6
0-0.8
-0.8-0
-1.6--
0.8
-2.4--
1.6
-3.2--
2.4
-4--3.2
26
Figure 17 : Signification globale des
fonctions de réponse de la largeur du bois final sur la période
octobre t-1 - septembre t pour chaque population du Mercantour
calculées sur le couple de paramètres précipitations et
températures moyennes.
Les variations inter-annuelles du paramètre largeur du
bois final sont très peu influencées par les variations du climat
ce qui confirme les résultat constaté aussi sur une autre essence
forestière, le pin d'Alep en Provence calcaire à plus basse
altitude (Nicault, 1999).

r/s
OCT NOV
DEC
JAN
FEV
MAR
AVR
MAI JUN
JUL
AOUT
SEP
sal1P
sesP
sal1T
sesT
brP
brT
2.4-3.2 1.6-2.4 0.8-1.6 0-0.8 -0.8-0 -1.6--0.8 -2.4--1.6
Figure 18 : Signification globale des
fonctions de réponse de la densité moyenne du bois initial sur la
période octobre t-1 - septembre t pour chaque population du
Mercantour calculées sur le couple de paramètres
précipitations et températures moyennes.
L'influence des précipitations sur les variations
inter-annuelles de la densité du bois initial est toujours inverse. Les
températures moyennes ont peu d'influence sur les variations de la
densité du bois initial. Seules les températures de
décembre et juillet semblent avoir un poids : la densité
réagit indirectement avec décembre et directement avec juillet,
en particulier pour la population de Sestrière.

r/s
OCT NOV DEC JAN FEV MAR AVR MAI JUIN JUIL AUT SEP
Sal1P
SesP
Sal1T
SesT
BrP
BrT
2.4-3.2 1.6-2.4 0.8-1.6 0-0.8 -0.8-0 -1.6--0.8 -2.4--1.6
-3.2--2.4
27
Figure 19 : Signification globale des
fonctions de réponse de la densité moyenne du bois final sur la
période octobre t-1 - septembre t pour chaque population du
Mercantour calculées sur le couple de paramètres
précipitations et températures moyennes.
La densité moyenne du bois final est
corrélée positivement avec les températures
d'été, négativement mais plus faiblement avec les
températures avec de mars et avril. La densité est inversement
corrélée avec les précipitations de mai-septembre en
particulier pour la population de Sestrière.
Les variations inter-annuelles de densité du bois final
semblent assez faiblement influencées par le climat ce qui a
été aussi constaté par Nicault (1999) à propos du
Pin d'Alep en basse Provence.
c) Regroupements de régresseurs pour les
paramètres largeur, surface et densité
L'utilisation simultanée des 24 régresseurs
climatiques ne permet pas d'obtenir une relation cerne/climat simple. Bien que
le coefficient de corrélation obtenu sur le sous-ensemble de calibration
soit élevé (proche de 0.7 souvent), le coefficient de
corrélation sur la vérification est en général
très faible (0.3, 0.4) et non significatif au regard de son
écart-type.
A partir des résultats précédents, des
regroupements permettent d'obtenir une fonction de réponse hautement
significative, notamment pour la population la plus sensible aux conditions
climatiques (Sestrière) et dont les résultats sont
présentés ci dessous (figure 20). Ce nouveau modèle met en
évidence l'influence importante des précipitations d'hiver, du
mois de juin, des températures de la fin de l'hiver et
d'été, sur la croissance radiale (figure 20). En outre, le
paramètre largeur du bois final réagit peu puisque le climat
n'explique que 18 % de la variance (r2 ou RMV2). Pour les
autres paramètres, le climat explique jusqu'à 59 % de la
variance, ce qui est un résultat hautement significatif des fonctions de
réponse.

r/s
r/s
r/s
-1
-2
-3
-1
-2
-3
-4
-5
-6
-1
-2
-3
-4
-5
4
3
2
0
4
7
6
5
3
2
0
1
1
4
3
2
0
1
|
|
Sestrière, bois initial, Dalmas.
|
St
|
largeur
du
|
|
R=0.81 RMV=0.59 RMV/S=3.18 r2=0.35
|
|
|
|
|
TMOYAOUT PRDEC PRFEV
|
PRJUN
|
|
|
Ses, épaisseur, St Dalmas
Sestrière, densité
moyenne du
bois
initial, St Dalmas.
TMOYMAR
TMOYDEC
PRDEC
TMOYJUL_AOUT
PRFEV
R=0.67 RMV=0.57 RMV/s=4.38 r2=0.32
R=0.72 RMV=0.53 RMV/S=2.94 r2=0.28
PRJUN
PRMAI
PRJUIN
PRJUL
r/s
r/s
r/s
-1
-2
-3
-4
-1
-2
-3
-4
-5
2
3
0
1
4
2
3
0
1
-1
-2
4
2
6
5
3
0
1
Sestrière, densité
moyenne du
bois
final, St Dalmas.
TMOYFEV PRNOV PRMAI PRSEP
PRNOV_DE PRFEV_MAR
Sestrière, largeur du bois final, St
Dalmas.
TMOYDEC
Ses, surface, St Dalmas
TMOYJUL_AOUT
R=0.62 RMV=0.59 RMV/s=2.68 r2=0.35
R=0.79 RMV=0.53 RMV/S=3.79 r2=0.28
PRJUN
R=0.68 RMV=0.43 RMV/S=2.39 r2=0.18
PRJUN
PRJUL
Figure 20 : Régresseurs significatifs
dégagés par les fonctions de réponse pour les
paramètres de la croissance radiale de la population de
Sestrière.
3.3. Recherche de l'influence de la pollution par
l'ozone sur la croissance radiale
3.3.1. Analyse des résidus

-10
-20
-30
-40
40
20
50
30
10
0
1932 1936 1940 1944 1948 1952 1956 1960 1964 1968 1972 1976 1980
1984 1988 1992 1996
-sigma résidus
estimés +sigma
résidus réels
28
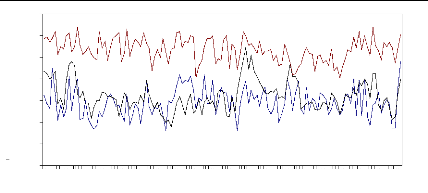
Figure 21 : Courbes de la variation des résidus obtenues
à partir de la modélisation ARMA pour la population Salèse
2.
29
Les deux lignes pointillés représentent
l'intervalle de confiance (au seuil 95 %). Il est à noter que la marge
d'erreur reste assez faible. L'échelle (axe y) est identique pour toutes
les courbes et indique des variations par rapport à la moyenne qui est
égale à 0.
Les séries de résidus, supposés
représenter au mieux l'impact des variations inter-annuelles du climat
sur la croissance, sont caractérisées par une diminution de la
variabilité inter-annuelle à partir des années 1972, 1982
et 1990. Les écarts-types suivent cette tendance et restent dans un
faible intervalle autour de la moyenne.
3.3.2. Estimation des largeurs des cernes
> Population de Sestrière :
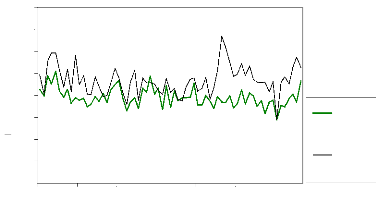
Largeurs (1/100 mm)
160
140
120
100
40
80
60
20
0
1932 1937 1942 1947 1952 1957 1962 1967 1972 1977 1982 1987 1992
1997
largeurs estimées
largeurs réelles
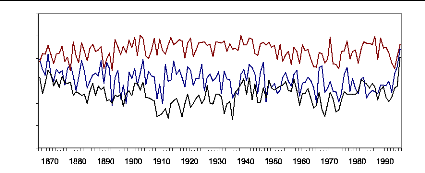
Figure 22 : Comparaison largeurs des cernes
réelles/estimées pour la population de Sestrière.
La courbe des épaisseurs réelles est constamment au
dessus des épaisseurs estimées.
Sur la période de calibration, les données
réelles et reconstruites ne se superposent pas parfaitement. Le
modèle n'est pas parfait et tend à sous-estimer la croissance. En
effet, la différence entre la courbe réelle et estimée est
significative au seuil 95% pour la période de
référence.
La période dite « polluée »
(1967-1998) est caractérisée par des largeurs de cerne largement
supérieures et significative à celle de la période non
polluée malgré la sous-etimation constatée.
Sur le plan dendroclimatique, pour les dernières
décennies quelques constatations sont possibles.
Des années exceptionnelles peuvent être
constatées, notamment 1949, 1976, 1992, caractérisées par
une forte chute de la croissance radiale.
30
> Population de Salèse 1 :
|
Largeurs (1/100 mm).
|
140 120 100 80 60 40 20
0
|
|
épaisseurs estimées
épaisseurs réelles
|
|
1932
1937
|
1942
|
1947
|
1952
|
1957
|
1962
|
1967
|
1972
|
1977
|
1982
|
1987
|
1992
|
1997
|
|
Figure 23 : Comparaison largeurs des cernes
réelles/estimées pour la population de Salèse 1.
La différence entre les largeurs réelles et
estimées n'est pas significative pour la période polluée.
Les valeurs estimées par le modèle sont assez bien
reconstituées en terme de variation et sont stables sur toute la
période.
> Population de Salèse 2 :

Largeurs (1/100 mm)
160
140
120
100
40
80
60
20
0
1932
1937
1942
1947
1952
1957
1962
1967
1972
1977
1982
1987
1992
1997
largeurs estimées
largeurs réelles
Figure 24 : Comparaison largeurs des cernes
réelles/estimées pour la population de Salèse 2.
La croissance estimée est légèrement
supérieure à la croissance réelle pour les 20
dernières années. La différence était significative
pour la période 1985-1998 au seuil 95 % (test de Student). La
différence pour la période 1964-1998 n'est pas significative.
31
> Population de Prals :
Largeur (1/100 mm)
200
180
160
140
120
100
40
20
80
60
0
1932
1937
1942
1947
1952
1957
1962
1967
1972
1977
1982
1987
1992
1997
largeurs estimées
largeurs réelles
Figure 25 : Comparaison largeurs des cernes
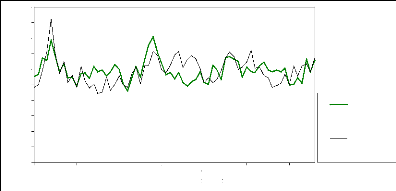
réelles/estimées pour la population de Prals.
La correspondance courbe croissance radiale réelle et
estimée n'est, là aussi, pas parfaite pour la période de
référence, il existe des phases pendant lesquelles la croissance
est surestimée, d'autres où elle est sous-estimée.
La différence est significative (au seuil de 95 %) pour
la période 1984-1992 seulement, où la croissance réelle
serait inférieure à ce qu'elle aurait due être. La
différence n'est pas significative sur la totalité de la
période « polluée » 1964-1998.
> Population d'Isola 1 :

Largeurs (1/100 mm).
120
100
40
80
60
20
0
1932 1936 1940 1944 1948 1952 1956 1960 1964 1968 1972 1976 1980
1984 1988 1992 1996
largeurs estimées
largeurs réelles
Figure 26 : Comparaison largeurs des cernes
réelles/estimées pour la population d'Isola 1.
La croissance pendant la période dite polluée
est le plus souvent supérieure à celle estimée. La
différence n'est pas significative pour l'ensemble de la période
« polluée ». La différence est significative pour la
période 1980-1998 mais dans le sens réel>estimé.
32
3.3.3. Récapitulatif
Tableau 6 : Différences entre les valeurs moyennes des
largeurs mesurées et les valeurs estimées pour les
périodes « non polluées » et « polluées
».
|
Largeurs moyennes (1/100e mm)
réelles/estimées
|
|
|
1932-1966
|
1967-1998
|
Différence significative au seuil 95 %
|
|
Ses
|
92.26/80.86
|
95.25/75.44
|
Oui (les 2 périodes)
mais dans le sens
réel>estimé
|
|
Sal 1
|
81.24/79.97
|
80.15/81.94
|
non
|
|
Sal 2
|
85.91/81.70
|
77.93/80.67
|
non
|
|
Iso 1
|
71.53/72.05
|
72.65/72.77
|
non
|
|
Prals
|
116.03/120.5
|
119.67/116.24
|
non
|
Remarque : les différences sont significatives pour des
sous-périodes de la période polluée dans le sens
estimé>réel (populations de Salèse 2 et Prals, cf
§ 3.3.2.) et dans le sens réel>estimé (population de
Prals.
33
4. DISCUSSION
4.1. Les chronologies
Les chronologies du Mercantour présentent un bon
synchronisme visuel confirmé par les tests statistiques. Cela signifie
que les différentes populations réagissent de façon
homogène à un facteur externe commun qui est le climat
régional de la Tinée à la Vésubie. Celui ci semble
donc assez homogène même s'il existe un gradient de
continentalité hydrique du nord-ouest au sud-est.
4.2. Les relations cernes/climat
Les relations cernes-climat fournies par les fonctions de
réponse permettent d'avoir une idée du rôle du climat sur
la croissance des arbres. Mais une relation statistique ne signifie pas
forcement une relation simple de cause à effet : en effet les
différents paramètres climatiques n'ont pas forcément une
action directe et immédiate sur la largeur des cernes. Par exemple, les
précipitations peuvent avoir une action positive sur la croissance des
individus par l'alimentation en eau qu'elles apportent mais aussi par le fait
que, en milieu de montagne, la pluie entraîne une baisse des
températures qui, elles-mêmes, peuvent exercer un effet inverse
sur la croissance des arbres.
Afin d'obtenir une meilleure interprétation des
fonctions de réponse et pour mieux comprendre les mécanismes de
croissance des individus ainsi que l'action spécifique du climat, il est
nécessaire de replacer les différentes populations dans leur
environnement propre et d'y intégrer les connaissances disponibles sur
l'autoécologie et l'écophysiologie de l'essence.
4.2.1. Précipitations et températures
moyennes hivernales
-Les précipitations : Sur la
période octobre t-1-septembre t, la relation est directe entre les
précipitations hivernales et les paramètres largeur totale,
largeur du bois initial et surface des cernes. Cela peut être
interprétée comme une influence positive sur la croissance
radiale des individus, mais différée. En effet, l'arbre entre
progressivement en phase de dormance qui commence à la mi-novembre pour
être complète en décembre. L'arbre est inactif entre
décembre et mars, période pendant laquelle le bilan de CO2 est
quotidiennement négatif (Wieser, 1996).
Les précipitations hivernales ont deux rôles
primordiaux :
> Le premier rôle est de reconstituer les
réserves hydriques du sol utilisées par les arbres lors de la
reprise de l'activité photosynthétique qui aura lieu au cours du
mois d'avril en montagne (Pernin, 1999).
34
> Le deuxième rôle est un rôle de
protection vis à vis de l'arbre (Pernin, 1999). En hiver, les
précipitations tombent essentiellement sous forme de neige et la
couverture neigeuse empêche la température du sol de descendre en
dessous de 0°C (Contini, Lavarelo, 1982). Elle évite donc au sol de
geler et par conséquent évite des dégâts
irrémédiables aux plantes. Les plantes soumises à une
forte dessiccation hivernale (due au vent froid notamment) doivent puiser l'eau
des réserves du sol pour pallier les pertes causées par
l'évapo-transpiration. Le pin cembro possède de nombreuses
adaptations à la vie en conditions extrêmes dont la faculté
de réguler la fermeture des stomates grâce à une
augmentation d'une phytohormone, l'ABA (acide abscissique), dans les aiguilles
(Christmann et al., 1999) pour minimiser les pertes d'eau vers
l'atmosphère. Malgré cela, l'épaisseur de sa cuticule est
mince et entraîne des pertes d'eau importantes qui doivent être
compensées. Si le sol est gelé, l'absorption d'eau du sol et le
transport dans les vaisseaux est interrompu. Les pertes dues à la
transpiration peuvent entraîner des dessèchements mortels. Une
couche de quelques dizaines de centimètres suffit à
protéger le sol contre le gel. De plus la neige, facilement
piégée par les aiguilles groupées par cinq, forme un
volume capable d'isoler l'extrémité de la pousse et les bourgeons
(Tranquilini, 1978).
-Les températures : Les
températures moyennes de décembre et février sont en
relation inverse avec l'épaisseur des cernes, c'est à dire que
plus les températures sont basses (jusqu'à - 2°C) plus les
cernes sont épais.
Des études écophysiologiques montrent que la
dormance dans laquelle l'arbre se trouve en hiver n'est pas sous la dominance
de la photopériode réduite mais serait plutôt d'origine
endogène (Contini, Lavarelo, 1982). Toutefois, le pin cembro peut
continuer à avoir une assimilation notable en hiver dans des conditions
favorables d'ensoleillement (Keller, 1970). Or des températures
élevées en hiver, liées à un fort ensoleillement
peuvent entraîner une hausse de la respiration et de la transpiration qui
mobiliseraient une partie des réserves nécessaires lors de la
période végétative suivante. Cet ensoleillement est
également renforcé par l'action de la couverture neigeuse qui
peut doubler l'énergie reçue (Tranquilini, 1978).
Des températures froides hivernales sont donc
nécessaires à la mise en place du cerne initial au printemps et
agissent comme une véritable « vernalisation ».
Le paramètre largeur du bois final est peu
influencé par les variations inter-annuelles. La largeur du bois final
est en effet très faible par rapport à celle du bois initial et
sa mise en place est rapide ; il est donc difficile de mettre en
évidence d'éventuelles variations inter-annuelles.
35
4.2.2. Les températures de mars, avril et
mai
L'effet inverse des températures de mars et avril
montre que la couverture neigeuse a un rôle de protection très
important. En effet, les températures qui restent basses assurent le
maintien de cette couverture neigeuse et donc la protection des individus
contre d'éventuels gels tardifs très fréquents à
cette période et à cette altitude. De plus, la présence de
la neige tardivement dans la saison n'empêche en rien la reprise de
l'activité photosynthétique car le pin cembro possède la
particularité de reprendre son activité à basse
température (à partir de - 4°C) (Aussenac, 1994). De ce
fait, le pin cembro peut profiter d'une période de
végétation relativement étendue, d'avril-mai à
novembre, contrairement à d'autres espèces de conifères
tels que le mélèze qui, pendant cette période, ne
possède pas encore d'aiguilles.
Seule la largeur du bois initial est corrélée
positivement avec les températures de mars et avril, cette
période correspondant au début de la croissance du cerne.
De même, seul le paramètre largeur du bois
initial est corrélé positivement avec les températures du
mois de mai. L'épaisseur des parois cellulaires est en effet un meilleur
enregistreur des conditions climatiques que la largeur des cernes. Le pin
cembro est l'espèce la plus sensible à la chaleur (par rapport au
mélèze et au sapin notamment) et utilise le plus efficacement la
chaleur pour sa croissance (Kronfuss, 1992). Cette espèce démarre
sa croissance en premier au mois de mai.
4.2.3. Les précipitations et les
températures estivales
Les précipitations du mois de juin ont un effet inverse
sur la croissance des arbres tandis que les températures ont, elles, un
effet direct. Le pin cembro est perturbé par ces fortes
précipitations qui signifient des températures basses. Le mois de
juin correspond à la pleine période végétative. Les
premières chaleurs d'été sont nécessaires au pin
cembro qui est génétiquement programmé pour utiliser
efficacement la chaleur de l'air et terminer le plus tôt possible la
croissance. Le pin cembro est l'espèce la plus sensible, la plus
réactive à la chaleur (par rapport au mélèze et au
sapin notamment) (Kronfuss, 1992).
Inversement, les températures élevées du
mois de juillet, qui est le mois le plus chaud, entraînent une forte
évapo-transpiration qui n'est compensée ni par les
réserves en eau accumulée les mois précédents, ni
par les faibles précipitations qui tombent durant cette période.
Par conséquent, une baisse dans l'activité
photosynthétique des arbres peut se traduire par une diminution de la
largeur des cernes pendant cette période. Les pins cembro sont sensibles
à la chaleur (Kronfuss, 1992), c'est pourquoi les pluies du mois de
juillet, qui jouent un rôle positif sur la croissance des individus, sont
nécessaires pour alimenter les arbres en eau et limiter les pertes en
eau par transpiration grâce aux nébulosités
associées.
36
De plus, de par leur appareil racinaire très
développé mais traçant qui leur permet de s'installer sur
des substrats de types éboulis, crêtes, l'alimentation en eau doit
être importante ou du moins régulière (Fourchy, 1968).
Les températures trop élevées
d'été (juin, juillet notamment) ont une influence négative
sur la croissance du bois initial. La grande sensibilité du pin cembro
est confirmée.
Le paramètre largeur du bois initial est
corrélé positivement avec les températures du mois
d'août, contre toute attente, pour une population, celle de
Sestrière. Comme cette station est un milieu plus fermé,
forestier, l'humidité y est plus élevée et la chaleur de
l'été peut entraîner une reprise de la croissance. Le
paramètre densité du bois initial est sous la dépendance
de la disponibilité en eau du début de l'été.
Globalement, on constatera dans toutes les fonctions de
réponse l'influence dominante des précipitations qui sont le
facteur limitant en région méditerranéenne par rapport aux
températures.
4.3. Quelle influence de la pollution par l'ozone sur
la croissance radiale ?
La difficulté est grande d'isoler le facteur pollution
par l'ozone des autres facteurs susceptibles d'intervenir sur la croissance
radiale annuelle des arbres.
La croissance radiale estimée des cernes étant
significativement inférieure à la croissance réelle durant
la période « polluée » pour 2 populations sur 5, il
semble difficile de relier catégoriquement une éventuelle
diminution des largeurs à une augmentation du niveau de pollution
atmosphérique (ozone en particulier).
Cette difficulté tient au fait que la croissance des
cernes subit les effets de multiples influences :
> L'âge, regroupant des facteurs internes propres
à chaque individu. Les mécanismes physiologiques normaux de
vieillissement des arbres s'accompagne souvent d'une dégradation de
l'état du houppier. (Dalstein, 1999) ;
> La compétition interspécifique (vis
à vis du mélèze notamment) ;
> L'environnement local (conditions stationnelles). Le
micro-environnement où pousse chaque arbre est plus ou moins favorable
à sa croissance, dès sa naissance et tout au long de sa vie.
L'état de santé des houppiers pourrait traduire ces
différences stationnelles. La pauvreté des sols type Ranker sur
un substratum cristallin (carences en éléments nutritifs, tels
que magnésium, azote et potassium), pourraient constituer un facteur
prédisposant local. Les jaunissements pour les populations
étudiées ici sont visibles alors que le substratum est de type
cristallin ou gréseux. Tout se passe donc comme si les
difficultés d'alimentation jusqu'ici chroniques étaient
brusquement mises au grand jour sous l'influence d'un facteur nouveau. Le
jaunissement des
37
aiguilles serait déclenché et aggravé par
des conditions climatiques défavorables de ces dernières
années (faibles enneigements hivernaux, enneigements tardifs au
printemps, fortes chaleurs et sécheresses estivales), et par les
concentrations croissantes d'ozone dans l'atmosphère durant
l'été. De plus, les dépôts acides pourraient induire
l'apparition de difficultés d'alimentation en magnésium et
calcium sur les sols pauvres, extériorisés par les
jaunissements.
D'autre part, l'augmentation du taux de CO2
atmosphérique depuis le début de l'ère industrielle
pourrait masquer l'effet négatif de l'ozone sur la croissance. Il a
été constaté une augmentation significative de la largeur
des cernes (pour une même classe d'âge) d'un millimètre par
an depuis le milieu du siècle dernier pour une population de pin cembro
de l'étage subalpin du Tyrol Autrichien (Nicolussi et al.,
1995). De plus, l'augmentation continuelle du CO2 dans l'atmosphère, en
entraînant une fermeture partielle des stomates, pourrait limiter
l'impact de l'ozone sur les arbres.
Les entrées azotées atmosphériques
(nitrates et ions ammonium) contenues dans la pluie ou les dépôts
secs azotés, en constante augmentation ces 20-30 dernières
années, pourraient également constituer un stimulant de la
croissance. (Nicolussi et al., 1995).
Il existe cependant de fortes présomptions pour que les
symptômes observés (atteintes des membranes cellulaires des
aiguilles) soient dus à l'ozone (Dalstein, 1997). De plus, la diminution
de la variabilité inter-annuelle des résidus à partir des
années 1970 pour la population Salèse 2 (la plus touchée)
pourrait signifier un impact de plus en plus important des taux d'ozone sans
cesse croissants. En effet, l'augmentation du taux d'ozone s'est
accélérée ces 15-20 dernières années en
passant de +1.6 % à + 2.4% par an (Le Cloirec, 1998) (figure 1). En ce
qui concerne l'analyse des largeurs des cernes, pour les populations
Salèse 2 et Prals, la croissance réelle a été
significativement inférieure à la croissance potentielle pour la
période 1984-1985 jusqu'en 1992 (Prals) et 1998 (Salèse 2).
L'hypothèse forte que l'on peut émettre est qu'il existe une
interaction entre les effets du climat et l'impact de l'ozone. Les
dépérissements, les défoliations, caractériseraient
donc en partie au moins, des « réajustements » parfois brutaux
mais normaux en réponse à des stress climatiques. Les effets de
l'ozone et des dépôts acides humides, occultes (nuages, etc) et
secs entreraient en interaction avec l'influence à moyen terme (quelques
années) et à long terme des cycles d'années sèches
(puisque l'on a vu l'influence prédominante des précipitations
sur la croissance au travers des fonctions de réponse). Les effets de la
pollution atmosphérique viendraient s'ajouter à ceux du climat et
pourraient même agir comme un déclencheur du
dépérissement. Des divergences de croissance entre arbres sains
et arbres défoliés existent dans les plus vieux peuplements de
Forêt-Noire dès la fin du siècle dernier ce qui peut
indiquer que le climat a toujours eu une influence primordiale. On notera ainsi
que les années exceptionnelles constatées par exemple dans la
figure 22 sont à mettre en relation avec des accidents
38
climatiques. La chute spectaculaire de la croissance en 1992 a
lieu suite à un mois de juin très froid (inférieur
à la moyenne-2 fois l'écart-type) et très pluvieux alors
que l'hiver a été très sec (non reconstitution des
réserves hydriques du sol). On remarquera l'effet des grandes
sécheresses de 1949 et 1976. La chute de croissance de 1954 fait suite
à un hiver (mois de janvier et février) plus froid que la normale
et à un mois de juin plus chaud.
Cependant, l'étendue et l'intensité du
dépérissement des années 1980 et 1990 dépasse celle
des phénomènes anciens et appelle la mise en jeu d'un autre
facteur perturbateur.
Enfin, une conduite sylvicole ancienne localement
négligée peut avoir des conséquences néfastes
à long terme. Elle peut induire une compétition très
accrue pour l'eau et l'engagement d'un processus d'affaiblissement
irréversible, accélérés par les sécheresses
de 1959, 1962, 1964 puis 19731976. (Becker et al., 1994).
39
5. CONCLUSION
Les 7 populations du Mercantour étudiées,
produisent les mêmes types de réponse croissance/climat que les
populations du Queyras à des divergences stationnelles près. Le
climat régnant sur les stations étudiées semble ainsi
exercer une influence suffisamment importante pour que la variabilité
inter-annuelle soit semblable. Cependant, le climat n'explique que 59 % de la
variance au maximum et laisse supposer une influence importante d'autres
facteurs sur la croissance radiale. Il faut donc conserver à l'esprit
que les conditions stationnelles comme l'exposition, la topographie,
l'édaphisme, les phénomènes de compétition, etc.
sont également des facteurs à prendre en compte pour expliquer la
croissance des individus. Les paramètres largeur totale, surface,
largeur du bois initial, densité moyenne du bois final se
révèlent les plus sensibles vis à vis des facteurs
climatiques. Le paramètre largeur du bois final n'est pas un bon
descripteur des variations inter-annuelles de la croissance radiale,
résultat en accord avec Nicault (1999). Les relations entre les
différents paramètres climatiques révèlent
l'importance du rôle inverse des températures de l'hiver, du
printemps de l'année t et du rôle direct des précipitations
hivernales sur la croissance des individus.
Concernant l'influence d'une pollution par l'ozone, des
études antérieures ont montré que « la dynamique de
croissance radiale des jeunes arbres jaunissants ou défoliés
n'est pas incompatible avec l'hypothèse d'un effet des polluants
atmosphériques puisqu'une augmentation du décalage de croissance
radiale est constatée entre les arbres verts et jaunissants depuis une
quinzaine d'année ». (Dalstein, 1999). La modélisation ARMA
et l'estimation d'une croissance radiale potentielle pour la période
« polluée » ne permet ni de confirmer ni d'infirmer le
rôle de l'ozone sur l'état de santé du pin cembro dans le
Mercantour. Cela signifie que le climat, facteur pris en compte dans les
modèles de croissance, n'influence qu'en partie mais pas en
totalité la croissance radiale. Un déficit de croissance par
rapport à une croissance potentielle modélisée
apparaît comme significatif pour les 20 dernières années,
en particulier chez la population du site de Salèse 2. Cette
évolution de la croissance peut, outre l'ozone, avoir des causes
multiples avec en premier lieu des accidents climatiques de plus en plus
fréquents ces deux dernières décennies comme des
étés très secs (stress hydrique) ou des fins de printemps
très froids. L'âge élevé des arbres, des carences en
ressources minérales locales ou la sensibilité du pin cembro aux
excès d'eau (ce n'est pas le cas des sites étudiés, bien
drainés du fait d'une forte pente) peuvent jouer aussi un rôle
important. L'impact négatif de l'ozone sur la croissance peut être
masqué par l'augmentation du taux de CO2 ou l'apport
d'éléments azotés atmosphériques.
La sensibilité de la méthode, satisfaisante dans
le cas des études déjà citées (Tessier et
al., 1990, Gandolfo et al., 1994) ne met pas en évidence
un effet négatif. Il est possible que cette pollution
atmosphérique soit masquée par les effets fertilisants
évoqués plus haut. Enfin, peut-être que le taux
40
d'ozone est encore insuffisant pour influer significativement
sur la croissance radiale et que les atteintes ne sont encore pas trop intenses
mais pas globale à l'échelle d'une région malgré
les symptômes visibles sur tous les arbres.
Cependant, il semble aujourd'hui qu'il faille prendre en
compte l'influence de la pollution par l'ozone en raison de la destruction des
tissus qu'elle provoque. Ces jaunissements encore localisés (pins
cembros dans le Mercantour, pins parasols en zones urbaines et suburbaines des
Alpes-Maritimes et des Bouches-du-Rhône) sont-ils les signes
avant-coureurs des problèmes qui attendent les forêts à
venir ? La concentration d'ozone dans l'atmosphère moyenne de l'Europe
augmente de 1 ppbv par an, et tous les écosystèmes forestiers
vont se trouver confrontés à des concentrations d'ozone
croissantes. Cependant, il faut garder à l'esprit que les
émissions annuelles naturelles (1 million de tonnes / an) de COV
précurseurs de l'ozone seraient d'environ la moitié des
émissions anthropiques. Dans les forêts de pins des régions
méditerranéennes, les émissions naturelles (sous forme
d'isoprène et de monoterpènes essentiellement) peuvent largement
dépasser durant certaines périodes les émissions
anthropiques (Luchetta et al., 2000).
Au final, pour mieux discerner la part de la pollution
atmosphérique (ozone en particulier) dans l'influence sur la croissance
radiale (par rapport au climat et à l'augmentation du taux de CO2
notamment), il conviendrait d'étendre les études spatialement
à d'autres populations, ainsi qu'à d'autres espèces
touchées (sapin par exemple) pour dégager une tendance à
plus grande échelle. Une perspective lourde à mettre en place,
parce que nécessitant de travailler sur de gros effectifs, consisterait
à chercher des différences significatives de la croissance
moyenne d'arbres de même âge mais à différentes
époques.
41
BIBLIOGRAPHIE
Aussenac G., Guehl J.M., 1994.
Dépérissements et accidents climatiques. Rev. For. Fr. 5
: 458-469. Becker et al., 1994. Conjonction des
facteurs naturels et des pratiques sylvicoles dans le dysfonctionnement des
écosystèmes forestiers. Revue forestière
française, 5 : 572-577.
Camaret Sylvaine et al., 1998. Mise
en évidence de l'impact du casse-noix moucheté (Nucifraga
caryocatactes L.) sur la distribution spatiale de la
régénération du Pin cembro (Pinus cembra L.).
Bull. Soc. Zool. Fr., 123(4) : 383-392.
Christmann et al., 1999. Seasonal
variation of abscisic Acid in needles of Pinus cembra L. at the Alpine
timberline and possible relations to frost resistance and water status.
Phyton (Austria), special issue «Eurosilva».
Contini L., Lavarelo Y.,
1982. Le pin cembro -répartition, écologie, sylviculture et
production-. INRA publication, 197 p.
Dalstein-Richier Laurence, 1997. Etat
phytosanitaire des cembraies du Mercantour et du Queyras en liaison avec la
pollution atmosphérique et les facteurs écologiques.
Thèse : Université de Droit, d'Economie et des Sciences
d'Aix-Marseille III. Faculté des Sciences et Techniques de Saint
Jérôme, IMEP. 273 p.
Eschbach Walter et al, 1995.
Technical advances in the radiodensitometrical determination of wood
density. Dendrochronologia, 13 : 155-168.
Fourchy Pierre, 1968. Notes sur le pin cembro
(Pinus cembra L.) dans les Alpes françaises. Revue
forestière française, 2 : 77-94.
Fritts HC., 1976. Tree ring and
climate. Academic press. New York, 567 p.
Gadbin-Henry Claude, 1994. Etude
dendroécologique de Pinus pinea L. Aspects méthodologiques.
Thèse : Université de Droit, d'Economie et des Sciences
d'Aix-Marseille III. Faculté des Sciences et Techniques de Saint
Jérôme, IMEP, 80 p.
Ganfoldo C., Tessier L.,
1994. Analyse dendroécologique sur des sapins (Abies alba
Mill.) et des épicéas (Picea abies (L.) Karst) de
différentes classes de défoliation dans la forêt de Vigo di
Ton (Trento, Italie). Ann. Sci. For., 51 : 391-406.
Garrec et Dalstein, 1997. Quand
l'ozone attaque les forêts de Provence. La Recherche n° 298
: 34-35. Guibal Frédéric, 1984. Contribution
dendroclimatologique à la connaissance de la croissance du Cèdre
de l'Atlas dans les reboisements du Sud-Est de la France. Thèse :
Université de Droit, d'Economie et des Sciences d'Aix-Marseille III, 123
p.
Guiot J., 1991. The bootstrap reponse function.
Tree-ring Bulletin, 51 : 39-41.
42
Keller T., Guiot J.,
Tessier L., 1997. Climatic effect of atmospheric CO2 doubling
on the radial tree growth in south eastern France. Journal of
biogeography, 24 : 857-864.
Keller T., 1999. Variabilité
climatique inter-annuelle quantifiée à partir de la
densité et des isotopes stables de la cellulose du bois. Thèse,
université des sciences d'Aix-Marseille III, 187 p.
Kronfuss Herbert, 1992. Height Growth in
Cembran Pine as a factor of air Temperature. Proceedings-International
Worshop on subalpine Stone Pines and their environment : the Status of Our
Knowledge. St. Moritz, Switzerland, September 5-11. p. 99-104.
Lambert et al., 1988. Pratique de la
dendrochronologie. Histoire et mesure, III-3 : 279-308.
Landmann G., 1998. La santé des
forêts françaises en 1996. Rev. For. Fr. L-1 : 21-31.
Le Cloirec Pierre, 1998. Les
composés organiques volatils dans l'environnement. Editions
Lavoisier Tec et Doc. 734 p.
Luchetta et al., 2000. Emission des
principaux composés organiques volatils biogéniques en France.
Pollution atmosphérique, 167 : 387-427.
Mégie Gérard, 1989. Ozone,
l'équilibre rompu. Presses du CNRS. 148 p.
Michalet R., 1991. Une approche
synthétique biopédoclimatique des montagnes
méditerrannéennes : exemple du Maroc septentrional.
Thèse, UJF Grenoble I, 238 p. + annexes.
Mouvier Gérard, 1994. La pollution
atmosphérique. Collection Dominos, Flammarion. 77 p.
Munaut, 1969. La dendrochronologie.
Bulletin de l'association française pour l'étude du
quaternaire : 65-71.
Nefaoui Mohamed, 1996.
Dendroécologie, productivité et dynamique de la croissance
radiale du pin maritime naturel au Maroc. Tome 2 : Typologie,
productivité et dynamique de la croissance radiale. Thèse :
Université de Droit, d'Economie et des Sciences d'Aix-Marseille III.
Faculté des Sciences et Techniques de Saint Jérôme, IMEP,
157 p.
Nicault Antoine, 1999. Analyse de
l'influence du climat sur les variations inter et intra-annuelles de la
croissance radiale du pin d'Alep(Pinus halepensis Mill.) en Provence
calcaire. Thèse : Université de Droit, d'Economie et des
Sciences d'Aix-Marseille III. Faculté des Sciences et Techniques de
Saint Jérôme, IMEP, 256 p.
Nicolussi Kurt, 1995. Increase in tree-ring
width in subalpine Pinus cembra L. from the central Alps that may be
CO2-related. Trees, 9 : 181-189.
Oswald H., 1995. Evolution des peuplements
forestiers dans l'étage subalpin: les forets à base de Pin cembro
(Pinus cembra L.). Ecologie, 29, 1-2, 239-246.
Pernin Céline, 1999. Approche
dendroécologique de l'impact des changements climatiques sur la
croissance du pin cembro en limite supérieure de la forêt dans le
Queyras (Hautes-Alpes). Mémoire de DEA : Université de
Droit, d'Economie et des Sciences d'Aix-Marseille III. Faculté des
Sciences et Techniques de Saint Jérôme, IMEP., 40 p.
43
Petitcolas Véronique, 1998.
Dendroécologie comparée de l'épicéa, du
Mélèze, du pin cembro et du pin à crochets en limite
supérieure de la forêt des Alpes françaises : influence de
la variabilité macro-écologique. Thèse : UJF-Grenoble
I, Laboratoire des Ecosystèmes Alpins. 66 p + annexes.
Rameau J.C. et al, 1993. Flore
forestière française, guide écologique
illustré. Tome 2 : montagnes. Institut pour le développement
forestier. 877 p.
Sandoz Henri, 1987. Recherches
taxonomiques, biogéographiques et phytoécologiques sur les
principaux conifères subalpins des Alpes : mélèze
d'Europe, pin cembro, pin à crochets et pin mugho. Etudes de terrain
centrées sur les Alpes maritimes et ligures (région du parc
national du Mercantour). Thèse : Université de Droit,
d'Economie et des Sciences d'Aix-Marseille III. Faculté des Sciences et
Techniques de Saint Jérôme, 647 p.
Schweingruber Fritz Hans, 1988. Tree
Rings, Basics and Applications of Dendrochronology. Ed. Reidel Publ. Comp.
Dordrecht, Holland. 276 p.
Tessier Lucien, 1984. Dendroclimatologie
et écologie de Pinus sylvestris L. et Quercus pubescens Willd. dans le
Sud-Est de la France. Thèse : Université de Droit,
d'Economie et des Sciences d'Aix-Marseille III, 257 p.
Tessier L., Serre-Bachet F.,
Guiot J., 1990. Pollution fluorée et croissance radiale
des conifères en Maurienne (Savoie, France). Ann. Sci. For., 47
: 309-323.
Tranquillini W., 1978. Physiological ecology
of the alpine timberline. Ecological studies 31. 137 p.
Wieser G, 1997. Carbon dioxyde gas exchange of cembrane pine
at the timberline during winter. Tree Physiology 17: 473-477.
| 

