|


MINISTERE DE L'ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE
SCIENTIFIQUE
************************
UNIVERSITE TOLIARA
**************
DOMAINE SCIENCES ET TECHNOLOGIES
**************
MENTION : SCIENCE DE LA VIE
***************
PARCOURS : BIODIVERSITE ET
ENVIRONEMENT
***************
SPECIALITE : BIOLOGIE
ANIMALE
***************
Mémoire en vue de l'obtention de diplôme
de MASTER II

BIOLOGIE ET ECOLOGIE DE DEUX ESPECES DE
RAPACES
SYMPATRIQUES MALGACHES, EPERVIER DE
MADAGASCAR Accipiter
madagascariensis (Verreaux j, 1833) et FAUCON A
VENTRE RAYE Falco
zoniventris (Peters, 1854) dans les Aires Protégées
:
réserve privée Reniala et forêt de Mosa, Mangily,
sud-ouest de Madagascar
Soutenu publiquement le 11 septembre 2021
Présenté par : RAKOTOSON Mauhamed Galien
Président de Jury : Monsieur KOTO Bernard, Professeur
Examinateur : Monsieur SAMA Zefania, Maitre de
Conférences
Raporteur : Monsieur RENE DE ROLAND Lily-Arison,
Professeur titulaire
Année universitaire : 2018-2019

i
REMERCIEMENTS
La réalisation de ce présent mémoire
a été possible grâce aux concours de plusieurs institutions
et personnes à qui je voudrais témoigner toute ma reconnaissance.
Certes, j'ai bénéficié l'aide de plusieurs personnes et
des organismes qui méritent mes vifs remerciements.
Je suis reconnaissant envers Monsieur FATIANY Pierre
Ruphin, Professeur, Doyen du Domaine des Sciences et Technologies,
Université de Toliara, de m'avoir autorisé à soutenir
publiquement ce mémoire en vue de l'obtention de Diplôme de MASTER
II.
Mon profond remerciement s'adresse à Madame
REJO-FIENENA Félicité, Professeur titulaire, Directeur de l'Ecole
doctorale de la Biodiversité et Environnements Tropicaux,
Université de Toliara, de nous avoir donner la plus grande
opportunité sur cette discipline et de m'avoir reçu
administrativement parmi les étudiants de cette école et d'avoir
permis à mon mémoire de bénéficier de toutes ses
corrections précieuses et pertinentes.
Il m'est particulièrement agréable
d'adresser ma respectueuse gratitude à Monsieur KOTO Bernard, Professeur
et Enseignant-Chercheur au sein de la Mention Géographie de
l'Université de Toliara, Domaine des lettres et des Sciences Humaines,
d'avoir accepté de présider cette séance soutenance de
mémoire.
Il me tient également à coeur de remercier
vivement à Monsieur l'examinateur, Docteur SAMA Zefania, Directeur de
l'Institut de l'Enseignement Supérieur de Menabe, Université de
Menabe, membre de la commission de lecture et Examinateur de ce mémoire,
qui a sacrifié son précieux temps pour améliorer ce
manuscrit
Je voudrais adresser toute ma gratitude à mon
encadreur, Monsieur RENE DE ROLAND Lily-Arison, Professeur titulaire,
Enseignant-chercheur au sein de la Mention Science de la vie dans le Domaine
des Sciences et Technologies, Université de Toliara, en même temps
Directeur National de l'ONG «The Peregrine Fund» à Madagascar,
d'avoir accepté l'encadrement de ce travail, pour son aide
précieuse, sa patience, sa disponibilité et surtout ses judicieux
conseils, qui m'a permis d'alimenter mes connaissances et d'orienter ma
réflexion.
Je remercie sincèrement l'ONG « Bel Avenir
» et l'ONG « The Peregrine Fund » pour leurs appuis
matériels et financier au cours de la réalisation de ce
présent mémoire.
ii
Toute ma gratitude aux personnels du réserve Reniala,
du Parc Mosa et à la population du village de Mangily, de nous avoir
bien accueillis et de nous avoir aidés (support morale, matériel
et techniques) durant notre étude sur terrain
Je tiens à remercier tous les personnels enseignant et
administratif de la Faculté des Sciences de l'Université de
Toliara pour les efforts investis tout au long de mon cursus
académique.
J'adresse vivement ma profonde gratitude à tous les
personnels enseignant et administratif de l'Université de Menabe, de
nous avoir accueillis pour la présentation de ce présent
mémoire
Je voudrais exprimer ma reconnaissance envers Mr RIVO Martin,
étudiant de la Faculté des sciences de l'Université
Antananarivo et Mr Gilbert Odilon, étudiant en Chimie de
l'Université de Toliara, d'être collaborant sur terrain.
J'adresse mes plus sincères remerciements à ma
famille : mes parents, mes frères, mes soeurs et tous mes proches, qui
m'ont accompagné, aidé, soutenu et encouragé tout au long
de la réalisation de ce mémoire.
Et j'ai gardé pour la fin les remerciements
exceptionnels de ceux qui ont contribué de près ou de loin
à ce mémoire. Sans vous, ce travail serait probablement encore
inachevé.
Merci à TOUS !
iii
ACRONYMES
ADN : Acide
Désoxyribonucléique
ANGAP : Association
Nationale pour la Gestion des
Aires Protégées
AP : Aire
Protégée
C : Celsius
Chap : Chapitre
CITES: Conservation on
International Trade in
Endagered Species of wild fauna and flora
DBH (anglais) : Diameter on
Breast High ou Dhp (français) :
Diamètre en haut de la poitrine
FAO : Food and
Agriculture Organization
Ha : Hypothèse
alternative
H0 : Hypothèse
nulle
fig : figure
g : gramme
GPS : Global Positioning
System
L : Longueur
l : largeur
mm : millimètre
mn : minute
T° : Température
ONG : Organisation Non
Gouvernementale
sp : espèce
P° : Précipitation
PPN : Produit de
Première Nécessité
UICN : Union Internationale
pour la Conservation de la Nature
ZICOMA : Zone d'Importance
pour la Conservation des Oiseaux à
Madagascar
iv
GLOSSAIRE
Aire Protégée : Espace
géographique clairement défini, reconnu, consacré et
géré, par tout moyen efficace, juridique ou autre, afin d'assurer
à long terme la conservation de la nature ainsi que les services
écosystémiques et les valeurs culturelles qui lui sont
associés.
Bioindicateur : espèce vivante dont la
présence renseigne sur certaines caractéristiques
physico-chimique ou biologiques de l'environnement ou sur l'incidence de
certaines pratiques
Biologie de reproduction : Science qui
étudie un phénomène par lequel une espèce engendre
une descendance qui la perpétue.
Colonie : Groupe d'oiseaux d'une même
espèce (colonie monospécifique) ou d'espèces
différentes (colonie mixte) réunis pour nicher.
Conservation (d'une espèce) : Ensemble
des mesures et actions (une large gamme de projets et programmes) entreprises
visant à préserver, protéger et sauver une espèce
de la disparition.
Croupion : extrémité
postérieure du corps d'un oiseau où sont fixées les
rectrices.
Dimorphisme sexuel : Cas pour une
espèce lorsque le mâle et la femelle ont un aspect
différent (forme, taille, couleur).
Ecologie : Science qui étudie les
conditions d'existence des êtres vivants et les interactions entre ces
êtres vivants et leur environnement (milieu abiotique et biotique).
Habitat (d'une espèce) : Lieu
caractérisé par un ensemble de conditions environnementales
comprenant des composants abiotiques (physico-chimiques) et biotiques (tels que
la présence d'autres espèces) dans lequel vit un ou plusieurs
individus d'une espèce.
Menace : Facteurs anthropiques et naturels
susceptibles de porter atteinte à la viabilité d'une
espèce et de son habitat.
Nichée : Ensemble des oisillons
nés d'une même ponte contenue par un nid
Niche écologique : Place et
spécialisation d'une espèce vivante à l'intérieur
d'un peuplement ou ensemble des conditions d'existence d'une espèce
(habitat, nourriture, conditions de reproduction, relation avec les autres
espèces).
Nid actif : Nid contenant un/des oeuf(s) ou
un/de(s) jeune(s) encore occupé par les parents.
v
Pelote de régurgitation : Sorte de
boule contenant des amas de débris alimentaires indigestes que rejettent
les oiseaux par régurgitation.
Population : Ensemble d'individus appartenant
à la même espèce et trouvés dans un endroit
particulier à un moment donné.
Photophase : Phase diurne ou phase claire du
jours ou phase de la lumière solaire, contrairement la nuit ou phase
nocturne ou la scotophase.
Population minimale viable : Plus petite
population qui pourrait avoir une forte probabilité de survivre et de
persister dans un avenir prévisible, ou nombre d'individus indispensable
pour maintenir la survie d'une espèce.
Pression : Activités humaines et
événements naturels tangibles et immédiats à
l'origine des impacts. Les impacts sont les résultats des pressions qui
réduisent la viabilité de l'espèce cible en raison de
leurs effets négatifs.
Rectrice : Grande plume rigide (de taille et
de forme variables) de la queue d'un oiseau. Rémiges :
Grandes plumes des ailes d'un oiseau.
Régime alimentaire : Aliments
consommés par un organisme. Ce terme désigne aussi la liste et
l'abondance des espèces animales ou végétales
consommées par une espèce donnée.
Scapulaire : Désigne les plumes de
l'épaule, insérées sur les bords du dos, qui recouvrent le
pli de la base de l'aile d'un oiseau.
vi
LISTE DES FIGURES
Figure n° 1 : Localisation de la
région Sud-ouest et du district de Toliara II 8
Figure n° 2 : Commune Rurale de Belalanda
et des sites étudiés 9
Figure n° 3 : Moyenne mensuelle de la
Température et de la précipitation à Mangily 11
Figure n° 4 : Forêt sèche de
Mangily, Rakotoson, 2020 13
Figure n° 5 : Les sels de Mangily,
Rakotoson, 2020 15
Figure n° 6 : Epervier de Madagascar
femelle, cliché par Rakotoson, 2018 19
Figure n° 7 : Faucon à ventre
rayé, cliché par Rakotoson, 2020 20
Figure n° 8 : Piège à
bâton 26
Figure n° 9 : Filet de Bal chatri et Bal
chatri modifié 26
Figure n° 10 : Mode de mensuration des
oeufs 27
Figure n° 11 : Schéma
d'illustration de la mensuration du nid (Stock, 2017) 29
Figure n° 12 : Technique
d'évaluation de l'habitat de nidification selon Greig-Smith, 1983 30
Figure n° 13 : Poids des poussins par
cinq jours 43
Figure n° 14 : Résultats de la
mensuration des nids 47
Figure n° 15 : Les régimes
alimentaires des oiseaux étudiés 50
Figure n° 16 : Faucons à Ventre
rayé sur le perchoir avec les proies capturées, Reniala 2020
51
LISTE DES TABLEAUX
Tableau n° 1 : Listes des individus
inventoriées 37
Tableau n° 2 : Résultats de la
mensuration et pesage des individus adultes capturés 39
Tableau n° 3 : Forme et dimension des
nids 46
Tableau n° 4 : Résultats de la
mensuration des arbres aux alentours de nid 47
vii
1
INTRODUCTION GENERALE
2
INTRODUCTION
Madagascar est une grande île de l'Océan Indien.
Elle est située à l'Est de l'Afrique et distant 400km par le
Canal de Mozambique. L'île est très connue mondialement par sa
potentialité au niveau des richesses naturelles surtout en
biodiversité. La plupart des espèces qu'elle abrite sont
endémiques. Cette endémicité atteigne 87% chez les
reptiles, 100% chez les amphibiens et les primates, 37% pour l'oiseau et 96%
des plantes vasculaires (Rakotoarinivo et al. 2014). Alors des auteurs
et des chercheurs la qualifient comme un pays à
mégadiversité et parmi les 10 hots spots mondiaux (Ganzhorn et
al, 2001, Mittermeier et al. 2004). Concernant la richesse en
avifaune, Madagascar abrite 282 espèces d'où 37% sont
endémiques de l'île (Raherilalao M. J. et Goodman M. S, 2011).
Avec des découvertes incessantes, et l'endémicité au
niveau supérieur (famille et sous-famille) présente à
Madagascar et à la région malgache, elle possède au totale
sept (07) dont cinq (05) familles (Mesithornitidae, Brachypteracidae,
Leptosomidae, Bernieridae et Vangidae) et 02 sous famille (Couinae et
Philepittinae) (Langrand 1995 ; Morris &Hawkins, 2000, Raherilalao et
Goodman S.M., 2011). Ce sont des familles composées d'espèces
largement à affinités forestières (Raherilalao &
Goodman, 2011).
Parmi ces 282 espèces d'oiseaux malagasy, 24
espèces sont constituées par des rapaces : 17 espèces
diurnes et 7 nocturnes (Collar et al, 1994 ; Fuchs et al.,
2007, Rene De Roland et Thorstrom, 2010). Parmi ces 24 espèces, 3 sur
les 12 endémiques sont classées dans les plus rares oiseaux du
monde, à savoir, le Harrier de Madagascar Circus macrosceles,
l'Aigle serpentaire de Madagascar Eutriorchis astur et l'Aigle
pêcheur de Madagascar Haliaeetus vociferoides (Collar et al.
1985 ; Collar et al., 1994, in Rasolonjatovo, 2016).
Malheureusement, ces richesses naturelles sont presque
pratiquement menacées due aux activités anthropiques dont les
actes les plus fréquentes sont la déforestation ou la
fragmentation des habitats naturels, la surexploitation, le trafic illicite.
Selon FAO, le taux de la déforestation est estimé à 1,6%
soit 200.000 ha (Meyers, 1990). Ces agressions appauvrissent les ressources
naturelles voire l'extinction des nombreuses espèces floristique et
faunistique de Madagascar.
En effet, des activités conservatives sont y
strictement souhaitables à développer afin d'éviter les
menaces qui pèsent sur les espèces sauvages malgaches. Les
forêts de Madagascar sont actuellement citées parmi l'une des
grandes priorités de conservation mondiale, non seulement par leurs
richesses biologiques qui sont uniques sur la planète, mais aussi par
leurs
3
fragilités surtout pour le cas de
l'écorégion du sud de l'île qui est strictement difficile
à restaurer en cas de dégradation (Nicoll et Langrand, 1989 ;
Ganzhorn et autres, 1997 ; ZICOMA (1999) ; Green et Susman, 1999 ;
Raselimanana, 2002).
La plupart des rapaces ou des oiseaux des proies sont
naturellement des espèces forestières strictes pour l'habitat
comme les aigles, les buses, les strigides, les accipitres et les faucons de
Madagascar mais vue l'effet de la déforestation, certaines
espèces peuvent s'adapter à s'installer dans les milieux ouverts
et dégradés.
L'Epervier de Madagascar et les Faucons à ventre
rayé qui sont nos objets d'étude sont des rapaces diurnes
endémiques de Madagascar qui préfèrent naturellement
l'habitat forestiers surtout pour la reproduction. Ces deux rapaces ont une
distribution dans le Nord et la côte Ouest de Madagascar, Parc Masoala
(René De Roland., 2000), Complexe Tsimembo Manambolomaty (Raveloson,
2017), Parc National de Marojejy (revue 2004 Parc National Marojejy).
Des études ont été déjà
faites sur ces deux rapaces dans ces régions, comme l'étude
biologique, écologique et éthologique de la famille de la
Falconidé, Faucon de newton (Falcon newton) et Faucon à
ventre rayé Falco zoniventris dans la partie occidentale de la
presqu'île de Masoala, Robenarimangason, H. 1999, étude
bio-écologique et éthologique des trois espèces
d'Accipiter sympatrique (A. francesiae, A. madagascariensis
et A. henstii) encore dans la presqu'île de Masoala
(René de Roland, 2000), la description du nid du Faucon à ventre
rayé (Falco zoniventris), son alimentation et ses
comportements, étude biologique de Faucon à ventre rayé au
Complexe Tsimembo Manambolomaty (René de Roland et al. 2005,
Raveloson, 2016). Les informations obtenues sur ces espèces ont
été seulement collectées dans les parties nord-ouest et
nord-est de l'île. Mais ces deux espèces d'oiseaux peuvent se
trouver dans la région Sud-Ouest de Madagascar, comme dans le Zombitse
National Park, dans les aires protégées Reniala et du parc Mosa
de Mangily-Ifaty Toliara, qui sont des sentiers botaniques et des
réserves ornithologiques, situés dans l'écorégion
du Sud et Sud-ouest malgache (Reniala, 2010). Ces deux dernières aires
protégées jouent les rôles importants en matière de
conservation mais aucune étude bioécologique et d'inventaire n'y
a été encore faite pour évaluer le succès de la
conservation pour ces deux espèces afin de compléter les bases de
données sur ces rapaces. C'est la raison pour laquelle qui nous pousse
à choisir notre présent sujet : « Biologie et
Ecologie de deux rapaces sympatriques malgaches, Epervier de Madagascar
(Accipiter madagascariensis (Verreaux, 1833) et Faucon à ventre
rayé (Falco zoniventris, Peter, 1984)
4
dans les Aires Protégées de Reniala et
de la forêt de Mosa, Mangily, Sud-ouest de Madagascar »
La question suivante constitue notre problématique :
Comment se présentent ces espèces dans ces sites et est-ce que
les conditions bioécologiques y sont favorables aux besoins de ces
espèces ?
Ce travail a pour objectif global de donner les informations
biologiques et écologiques conduisant la présence de ces deux
rapaces dans la forêt sèche du Sud-ouest de Madagascar afin de
compléter les bases des données et de raffiner le plan de
conservation existant dans ces sites.
Pour atteindre cet objectif global, des objectifs
spécifiques sont assignés à cette étude : >
Inventorier les populations présentes
> Recenser les couples reproducteurs dans les sites
d'étude
> Localiser et caractériser les nids de ces rapaces
dans la zone d'étude
> Etudier la biologie (morphologie et biologie de la
reproduction) de ces deux rapaces > Etudier l'écologie (régime
alimentaire, habitats, relation intra- et interspécifiques et
comportements) de ces deux espèces
> Identifier les pressions et les menaces pesant sur ces
espèces dans ce site
> Comprendre les facteurs écologiques qui
déterminent leur présence
Dans ce cadre de travail, cinq hypothèses suivantes
doivent être vérifiées : y' Hypothèse 1 : La
distribution de l'effectif des individus des deux espèces dans les zones
protégées et non protégées est homogène
y' Hypothèse 2 : L'effectif des nids et des individus
présents dans la zone d'étude pour les deux espèces est
égal.
y' Hypothèse 3 : La participation entre les mâles et
les femelles pendant la reproduction (fabrication du nid, incubation et
élevage des poussins) est homogènes.
y' Hypothèse 4 : Ces deux rapaces ont des mêmes
préférences écologiques.
Ce présent mémoire comporte trois parties.
D'abord, la première partie est destinée à la
méthodologie, qui englobe la description de la zone d'étude et la
présentation des espèces étudiées, des
matériels utilisés, des méthodes de collecte des
données suivi de traitement et de l'analyse statistique des informations
collectées. Puis, les résultats et les interprétations
sont donnés dans la deuxième partie. Tandis que, la
troisième dernière partie se rapporte à la discussion des
résultats, des recommandations et de conclusion.
PARTIE I : METHODOLOGIE
6
I.1. PRESENTATION DE LA ZONE D'ETUDE
Pour cette étude, la réserve Reniala et le parc
Mosa ont été choisis comme sites d'étude, en tant que
réserves abritant une concentration importante en avifaune,
particulièrement des rapaces selon notre étude bibliographique et
notre enquête préliminaire effectué aux responsables des
aires protégées.
Cette partie alors nous présentera les contextes :
géographiques, historiques, abiotiques, biotiques et anthropiques, de la
zone étudiée.
I.1.1. Contexte historique et localisation
géographique de la zone d'étude
Historiquement, Reniala est une aire protégée
privée, fondée en 1998 et ouvert publiquement en 2000. Elle
dispose une surface 60ha dont 15 ha constitue la zone tampon. (Andrianarivo
& Rakotovazaha. 2011, Fomenjanahary, 2014, Bothel, 2018). Par contre, la
Forêt de Mosa ou forêt de Baobab, son histoire concernant la date
de création n'est pas disponible pour cette étude, tandis que son
nom « forêt de Baobab » qui signifie en malgache « Alan'ny
Baobab » vient de la particularité de cette parcelle
forestière lors de l'abondance des pieds de Baobab.
La zone d'étude se trouve dans l'écorégion
du Sud-Ouest de Madagascar, située à 800m et à l'Est par
rapport au canal de Mozambique. Les deux sites se trouvent côte à
côte et forment généralement un bloc forestier.
Administrativement, cette zone d'étude est
située dans la Commune Rurale de Belalanda, District de Toliara II, tous
près du village de Mangily, elle est distante de 27km par rapport
à la ville de Toliara en suivant la route nationale numéro 09,
dans les coordonnées géographiques suivants : latitude
23°07'22» Sud et longitude 043° 37'14» Est (figure 1).
Ces deux aires protégées sont qualifiées
dans la catégorie différente :
? La réserve privée Reniala est qualifié
dans la catégorie IV : Réserve Naturelle ou Aire de Gestion des
habitats ou des espèces selon la classification de l'UICN (Renila,
2019). Cette catégorie englobe les aires protégées
gérées principalement à de fin de conservation avec
l'intervention de la gestion. On peut y faire des visites à titre
éducative, contrairement aux critères de la catégorie
I.
7
? La réserve Mosa est qualifiée dans la
catégorie 2 ou Parc National (PN), qui est une aire
protégée gérée principalement dans le but de
protéger les écosystèmes et à des fins
récréatives (Parc Mosa, 2019).
Ces deux aires protégées ont des principaux
objectifs de sauvegarder, de gérer et de protéger la faune
ornithologique et de la flore particulièrement les séries de la
Didieracée et de la Malavacée. Spécifiquement, la
création de ces deux AP permet de soutenir la protection de
l'environnement et surtout le développement au niveau local à
travers l'écotourisme.
Tous ces objectifs font de Reniala et de Mosa une structure
tournée vers le tourisme durable puisqu'elles prennent en compte,
à ses échelles, la dimension économique, éducative
et aussi la préservation des sites exceptionnelles
Figure n° 1 : Localisation de la région
Sud-ouest et du district de Toliara II
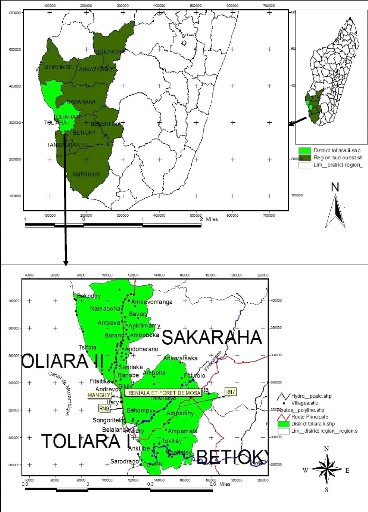
Source: RAKOTOSON, 2020
8
9
Figure n° 2 : Commune Rurale de Belalanda
et des sites étudiés
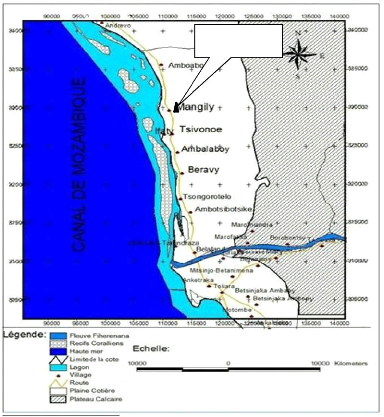
Source: RAKOTOSON, 2020
Parc Mosa et réserve Reniala
I.1.2. Milieu abiotique
Ce sous chapitre va nous donner les informations naturelles
physico-chimiques de notre zone d'étude, en parlant la situation
climatique et géo-pédologique.
I.1.2.1. Le climat
La région Sud-Ouest de Madagascar fait partie des
régions les plus ensoleillées de Madagascar. Des auteurs la
qualifient comme une zone subaride marquée par l'insuffisance et
l'irrégularité des précipitations (ANGAP, 1998). Le climat
de la région est de type tropical semi-aride et chaud avec deux saisons
bien marquées : une saison sèche d'avril à novembre et une
saison un peu pluvieuse très chaude, de décembre à mars
(Rejo-Fienena F., 1995).
10
Le vent sec dominant appelé « tioka atimo
» ou vent du sud en français, se dirige du canal de Mozambique
vers le milieu continental, plus précisément vers le Nord de
l'île, avec une vitesse moyenne entre 12 et 15km/h selon Thommasson,
1999.
Les données climatiques utilisées dans cette
étude sont obtenues à partir de la station
météorologique basée à Toliara, durant la
période 2015 à 2018.
? La précipitation
Notre zone d'étude occupe la transition entre la
côte ouest tropicale et le sud désertique avec une
précipitation faible, moins de 600 mm par an. La quantité de
pluie enregistrée durant l'année 2015 à 2018 est de 2071mm
avec un maximum de pluies enregistrés durant le premier trimestre
(novembre à janvier). La pluviométrie annuelle est 418mm. La
pluie tombe essentiellement de décembre en avril avec des
quantités variées d'une année à l'autre tandis que
la période sèche s'étend entre Mai à Novembre
(Monographie de la ville de Toliara, 2011). Comme en 2016, les pluies
abondantes sont observées au mois de Mars avec une pluviométrie
116mm (Bothel D, 2018).
b. La température
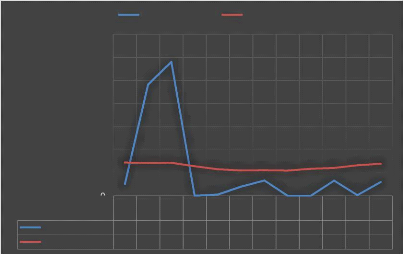
La température moyenne annuelle en 2016 est
24,95°C à Mangily, mais elle peut atteindre jusqu'à
28,8°C pendant la saison de pluie. Le mois le plus froid semble être
le mois de juillet, durant lequel la température peut descendre
au-dessous de 20°C. Par contre, les mois de novembre et décembre
sont les mois le plus chauds.
Précipitation (mm) T moyennes
140
120
100
80
60
40
20
|
0
|
Janvi er
|
Févri er
|
Mars
|
Avril
|
Mai
|
Juin
|
Juillet
|
Aout
|
Septe mbre
|
Octo bre
|
Nove mbre
|
Déce mbre
|
|
Précipitation (mm)
|
10,3
|
96,4
|
116
|
0
|
1,1
|
7,9
|
13,2
|
0
|
0
|
13,1
|
0,5
|
11,9
|
|
T moyennes
|
28,8
|
28,4
|
28,6
|
25,6
|
23
|
21,9
|
22,1
|
21,8
|
23,4
|
24,1
|
26,4
|
27,7
|
11
Figure n° 3 : Moyenne mensuelle de la
Température et de la précipitation à Mangily
Source :
Centre météorologique Toliara, 2016
Durant la saison chaude, la température varie en
moyenne entre 27 et 32°C. Les mois de Janvier et de février sont
les mois les plus chauds. Tandis que pendant la saison froide, les mois les
plus frais sont les mois de juin et de juillet avec une température
moyenne entre 27 et 29° C. Dans la nuit, elle peut descendre
jusqu'à 14° C (Collège français Toliara, 2013). La
précipitation est un peu élevée entre octobre à
mars. Le mois de février tient la valeur maximale tandis que durant les
mois restant, la valeur de la précipitation est totalement faible. Elle
peut atteindre 0 mm comme le cas des mois d'avril, d'août et de
septembre.
c. Vent
Il existe toute l'année. La période
sèche, de juillet à octobre est la période la plus
ventée (fréquence supérieure à 70 %). En
général, c'est le vent du sud (Tiokantimo). Il est
habituel observé un vent modéré de secteur N-N-E,
soufflant faiblement la nuit et le matin. Il tombe en fin de matinée
pour laisser place à un vent du Sud plus soutenu, qui atteint sa force
maximale dans l'après-midi. Ce fait est bien connu des pêcheurs
Vezo qui partent la nuit s'ils veulent voyager vers le sud,
l'après-midi, pour aller au Nord. L'existence de ces vents dominants du
Sud s'inscrit bien dans la morphologie des dunes, groupées en vastes
ensembles, le long du littoral, ainsi que dans la végétation
(orientation de l'espèce Didierea madagascariensis) (Rabevao,
2018).
12
I.1.2.2. Géo-pédologie
Selon Sourdat, 1972, les types du sol du Sud-Ouest de
Madagascar sont caractérisés par des sols minéraux bruts,
sols peu évolués d'apport alluvial, sols hydromorphes, sols
à peroxydes de fer (ou sable roux) décalcifiés, sables
roux. Ces sont les types de sol qui caractérisent notre zone
d'étude, ce qui permet l'existence et l'abondance des
Didieracées et des Euphorbiacées.
Cette zone d'étude fait partie du bassin
sédimentaire de Morondava, avec une formation post karroo. En effet,
elle est généralement constituée principalement par des
roches et des sols calcaires gréseux. Cette particularité
détermine sa diversité biologique : faunistique et floristique.
Dans cette zone, on y rencontre partiellement des sols sablonneux, couleur
rousse à affleurement calcaire, des sols ferrugineux c'est-à-dire
riches en fer et pauvre en aluminium (sol rouge) plus précisément
dans le milieu forestier.
I.1.3. Milieu Biologique
Notre zone d'étude est un site destiné à
protéger in-situ les espèces animales et végétales
sauvages dans cette région. Ces deux aires protégées ont
principalement objectif de protéger la faune ornithologique y compris
les rapaces et les végétaux supérieurs
particulièrement les Didieracées, les Euphorbiacées et les
Malvacées.
I.1.3.1. Flore et Végétation
La forêt du sud-ouest malgache est
caractérisée par une végétation sèche
souvent avec une adaptation importante comme la spinescence, l'aphyllie, la
succulence, la caducifoliée, la vivacité, etc. La
végétation de Mangily comprend une vaste pluralité de
formations végétales, qui est composée par de la
forêt dense sèche et du fourré xérophile du sud.
Elle est dominée par les cinq (05) familles des plantes telles que les
Euphorbiacées (Jatropha mahafaliensis), les Liliacées,
les Apocynacées, Malvacées et les Didiéracées dont
la plupart des espèces sont endémiques (Lloyd, 2012). Le Baobab
Fony ou Adansonia rubrostipa est une espèce
végétale faisant partie des espèces cible de conservation.
Le nom de l'espèce est la source du nom de l'aire protégée
: Foret de Baobab (ou forêt de Mosa) et Reniala.
Forêt de Baobab ou « alan'ny Baobaob » veut
dire une forêt constituée par des Baobab, c'est une espèce
autochtone et y présente depuis plusieurs années d'où le
nom Reniala ou mère de la Forêt. Dans cette aire
protégée, le Baobab tient la biomasse importante et attire
beaucoup
13
des touristes et des écotouristes. La liste des
espèces végétales dans la zone d'étude est
enregistrée dans l'annexe 7.

Figure n° 4 : Forêt sèche de
Mangily, Rakotoson, 2020
Ces photos montrent à gauche des végétaux
épineux et à droites quelque pieds de Baobab fony. Ces
végétaux sont à l'état vert après deux mois
de la précipitation élevée du deux mois avant
c'est-à-dire le mois de décembre et de Janvier.
I.1.3.2. Faune
La forêt sèche de Mangily renferme une
potentialité sur la richesse faunistique d'où la plupart des
espèces sont endémique de l'île.
Pour les invertébrés, elle possède plus
d'une vingtaine d'espèces et le nombre est inestimable surtout pendant
la saison de pluie de Novembre à Avril. Notre étude sur terrain
s'est réalisée pendant cette période, alors nous avions
rencontré une diversité spécifique et de nombre des :
papillons, des criquets (Locusta migratoria, des sauterelles, ...).
Les invertébrés sont un groupe d'animaux très important
sur l'écosystème. Ils assurent la pollinisation et sont servi
comme régime alimentaire des insectivores voire même pour certains
rapaces.
Pour les vertébrés, cet embranchement est
constitué par les groupes des poissons et des tétrapodes. Dans
notre zone d'études les poissons sont absents (sauf dans la mer) lors de
l'absence des réseaux hydrographiques tandis que tous le quatre classes
des tétrapodes sont y présents : Les amphibiens (Grenouilles),
les reptiles (lézards Chalaradon madagascariensis,
Chaméleons, serpents Boa madagascariensis, etc, des
tortues en captivités Astrocheleus arachnoïdes, Geochelon
radiata, Pyxus sp, des geckos), des 59 espèces d'oiseaux tel la
sous-
14
famille de Couinée de la forêt denses
sèches (Coua coquereli, C. gigas, C. coquereli, C.
ruficeps, C. cursor, C. cristata et C. verreauxi),
Epervier de Madagascar, Epervier de France, le faucons à ventre
rayé Falco zoniventris, Gymniophones de Madagascar, des
mammifères particulièrement les primates comme le Lemur
catta, le Propithecus verrauxi, ces deux lémuriens vivent
en captivité dans cette aire protégée. La liste des
oiseaux dans ce site est enregistrée dans l'annexe 6.
I.1.4. Population et ses activités
Les populations dans le village de Mangily-Ifaty sont
constituées par divers groupes ethniques tels que : Vezo, Masikoro,
Tanalana, Bara, Merina et d'origines étranger comme les Indiens, les
Français, les Chinois et autres. En général, les
populations autochtones sont le Masikoro et le Vezo tandis que les autres sont
des populations immigrantes. En 2012, l'effectif de la population a
été estimé 8500, (Romeo, 2014)
En effet, des activités différentes y sont
rencontrées tel que l'hôtellerie, production de sels, agronomie,
tourisme, commerce, ...
Agronomie : La culture dépend des
précipitations. Chaque orage ouvre une période de travaux sur
laquelle, les hommes, les femmes et les enfants se côtoient dans les
champs. Les cultures de manioc (Manihot esculenta), le maïs
(Zea mays), la patate douce (Ipomoea batatas), les pois du
cap (Phageolus lunatus), le haricot (Phaseolus vulgaris),
l'arachide (Arachis hypogaea), le coton (Gossypium hirsutum)
ainsi que quelques Cucurbitacées sont les plus importants.
L'élevage des zébus et le cheptel des ovins, des
caprins et des volailles sont les activités principales de la
population. Les zébus ont des valeurs culturelles dans les rites comme
les mariages, funérailles et ils font le bailleur de fonds
économique des familles. L'élevage dans cette région est
fréquemment extensif. Les troupeaux sont souvent mélangés,
libre ou semi libre sous la surveillance d'un bouvier (Mpiarak'andro)
(Rabarisoloaritefy, 2014).
Exploitation forestière :
L'espèce Aluauda procera (fatsilotra) est
exploitée par les bucherons pour fabriquer les planches vendues comme
principale bois de construction. L'exploitation est prolongée car c'est
la source d'argent assez facile et rapide. De plus, l'espèce
Cedrolopsis greveï (Katrafay) sert lieu de bois de
chauffe et est exploité pour la fabrication de charbon ainsi que pour la
médicine traditionnelle. Les Mendoravy (Albizia) sont
utilisés pour
15
la fabrication de cercueil et enfin, le Daro ou
Commiphora sp est exploité pour la menuiserie et la
médicine traditionnelle (Rabarisoloaritefy, 2014).
Pêche : La pêche est
réservée pour les Vezo. Elle est très importante, car leur
activité principale ; pourtant cette activité dépend des
conditions naturelles. La plupart d'entre eux sont des pêcheurs
traditionnels ; ils utilisent des matériels artisanaux et
archaïques tels que la pirogue, la canne à pêche, le filet
maillant et autres. Les rendements sont très faibles à cause de
marché local comme le seul lieu de liquidation pour leur marchandise
(Rabarisoloaritefy, 2014).
Production de sel : La production de sels est
une activité importante pour la population côtière de la
région Sud-ouest de Madagascar. Actuellement cette opportunité
est bien exploitée dans cette région surtout par des entreprises
de certains étrangers.

Figure n° 5 : Les sels de Mangily,
Rakotoson, 2020
Commerce et hôtellerie : Le commerce
est une activité inséparable à la société
humaine pour permettre l'échange. Comme à Mangily, le commerce se
fait irrégulier, car il s'est réalisé partout, le plus
fréquent c'est au bord de la route national n° 9 du village.
Là on trouve des légumes, des fruits de mer et des PPN.
D'ailleurs, entant qu'un village touristique,
l'hôtellerie est un secteur ou une activité très importante
pour héberger les touristes qui appelle donc des mains d'ouvre pour y
effectuer les taches dans ce domaine ainsi que des devises.
Ecotourisme : Cette activité est une
activité la plus importante car il est la source de la devise du village
donc source du développement. Comme nous avions dit ci-dessus, le
village de Mangily possède non seulement des aires
protégées qui lui entourent mais aussi située tout
près des merveilleuses plages : plage d'Ifaty, plage de Madiorano et
plage de Mangily.
16
D'ailleurs, la température est quasiment optimale
favorable au swimming (nage). Il y a une période destinée
à découvrir les baleines à bosses. En effet, des
nombreuses touristes arrivent toujours dans ce village pour découvrir
les secrets naturels de cette région.
I.2. PRESENTATION DES ESPECES ETUDIEES
La présente étude se focalise sur les deux
espèces aviaires malgaches dans le groupe des rapaces. Par
définition, les rapaces sont des oiseaux carnivores qui capturent
généralement d'autres animaux vivants à l'aide de leurs
serres (sauf les rapaces charognards), tels que les petits
vertébrés comme les micromammifères, d'autres oiseaux de
petites tailles, des caméléons, des geckos, des poissons et des
insectes (Langrand, 1995 ; Safford & Hawkins, 2013). Plusieurs institutions
et auteurs ont consacré une attention particulière sur ce groupe
qui est à la fois fascinant et difficile à étudier. Dans
le cas de Madagascar, les activités de The Peregrine Fund s'occupent
principalement sur les rapaces, en particulier la recherche sur ce groupe et sa
conservation. Langrand et Meyburg (1984) ont fait une évaluation de la
distribution, du statut de conservation des oiseaux de proies et des hiboux, en
particulier les rapaces endémiques telles que, Aviceda
madagascariensis, Accipiter henstii, Eutriorchis astur,
Buteo brachypterus, Polyboroides radiatus et Haliaeetus
vociferoides, Falco zoniventris et Accipiter
madagascariensis. Ces deux derniers sont l'objet de cette étude.
Ces sont des rapaces diurnes appartenant de deux ordres différents
suivants : ordre des Falconiformes famille de Falconidae (Faucon à
ventre rayé ou Falco zoniventris) et ordre des Accipitriformes,
famille de l'Accipitridae (Epervier de Madagascar ou Accipiter
madagascariensis).
Les ordres des Accipitriformes et des Falconiformes regroupent
les rapaces ou les oiseaux de proie diurnes tels que les aigles, les vautours,
les buses les éperviers et les faucons. Ces espèces se
caractérisent par leur court bec crochu et leurs pattes puissantes et
griffues appelées serres. Ce sont le plus souvent d'excellents chasseurs
diurnes à vue perçante. Certains, comme les vautours, se
nourrissent de charognes. Ils nichent sur les arbres, sur les rochers ou sur
les falaises. Les grandes espèces n'élèvent, en
général, qu'un à deux poussins par an. Les jeunes sont
nidicoles.
Auparavant, les Falconiformes et les Accipitriformes se sont
regroupés dans un même ordre appelé Falconiformes par leurs
ressemblances morphologiques entant que rapaces diurnes mais les
récentes études génétiques par l'hybridation d'ADN
confirment que ces groupes sont génétiquement loin
(Sibley-Ahlquist (1990, 1993), Raposo do Amaral et
17
al. (2009), David P. Mindell, J. F., Jeff A J.
(2018). Cela entraîne un bouleversement dans la taxonomie du groupe, qui
est répercutée dans la version 3.3 (2013) de la classification de
référence du Congrès ornithologique international,
c'est-à-dire, la séparation de ces groupes en deux ordres
différents :
? Les Accipitriformes sont un ordre qui
regroupe les rapaces diurnes tels que les aigles, les vautours, les buses et
les éperviers, constitue environs 220 espèces y compris
l'épervier de Madagascar ou Accipiter madagascariensis.
? Les falconiformes sont aussi un ordre de
rapaces diurnes qui ne contient plus qu'une seule famille, celle les
Falconidés, qui inclut les faucons, les caracaras, le macagua, les
carnifex, les fauconnets, les crécerelles, soit environ 60
espèces, y compris le faucon à ventre rayé ou Falco
zoniventris.
I.2.1. Position Systématique
I.2.1.1. Epervier de Madagascar (Accipiter
madagascariensis)
Règne : Animal
Embranchement : Vertébrés
Classe : Oiseaux
Ordre : Accipitriformes
Famille : Accipitridae
Genre : Accipiter
Espèce : madagascariensis (Verreaux, 1833)
Nom français : Epervier de Madagascar
Nom anglais : Madagascar sparrowhawk
Nom vernaculaire en Malgache : Firasa, Firasambalala, Pera,
Tsipara
18
I.2.1.2. Faucon à ventre rayé (Falco
zoniventris)
Selon la classification de Limnaeus (1758), cette
espèce appartient à la classification suivante :
Règne : Animal
Embranchement : Vertébrés
Classe : Oiseaux
Ordre : Falconiformes
Famille : Falconidae
Genre : Falco
Espèce : zoniventris, (Peters, 1985)
Nom français : Faucon à ventre rayé
Nom anglais : Madagascar Banded kestrel
Nom vernaculaire : Hitsikitsik'ala, Hitsikitsika
I.2.2. Description morphologique
I.2.2.1. Epervier de Madagascar (Accipiter
madagascariensis)
Cet oiseau mesure 34 à 40 cm, la femelle étant
plus grande que le mâle. Le dimorphisme sexuel porte non seulement sur la
taille mais également sur la coloration du corps. Le mâle a les
parties supérieures brun grisâtre foncé. Les plumes de la
nuque ont des bases blanches tandis que celles du croupion ont des
extrémités rousses. Les rémiges sont grises-brun
foncé sur le dessus mais plus claires en dessous avec de larges barres
brunes. La queue est grise-brun avec des bandes plus foncées avec sept
à huit bandes claires, mais elle apparait uniformément sombre.
Les parties inférieures sont blanches et nettement barrées de
gris foncé. La gorge est plutôt striée de gris foncé
tandis que le bas-ventre est blanc uni.
La tête est grise-brun foncé ou gris
ardoisé, mais les couvertures auriculaires sont striées de gris
foncé. Le bec est noir avec la cire jaune. Les yeux sont jaunes. Les
pattes et les doigts sont jaune verdâtres. La femelle adulte est plus
brune sur le dessus et les parties inférieures
19
sont plus intensément barrées. La tête et
les joues sont plus foncées. Elle est environ 18% plus grande que le
mâle. Le juvénile a une coloration brune plus clair sur le dessus
avec quelques stries sur la nuque et des liserés roux clairs sur le
dessus qui donnent un aspect légèrement écaillé.
Les parties inférieures sont blanc-crème avec des stries brunes
variables. Le dessous des ailes est barré. L'Epervier de Madagascar est
légèrement plus grand que l'Epervier de Frances, mais il semble
mince et petit comparé à l'Autour de Henst. Mâle et femelle
ressemblent à la femelle de l'Epervier d'Europe.

Figure n° 6 : Epervier de Madagascar
femelle, cliché par Rakotoson, 2018 I.2.2.2. Faucon à
ventre rayé (Falco zoniventris)
Le Faucon à ventre rayé est un petit et atypique
faucon. Son plumage varie légèrement avec l'âge. Sa taille
est comprise entre 25-35cm (Morris & Hawkins, 1998). La tête est
grosse et grise, sans moustache avec une zone périoculaire
dénudée et jaune. Les cires sont jaunes, le bec est noir et fort,
L'iris est jaune pâle. Les parties supérieures du corps et des
ailes sont grises-bleu barrées de brun noir. Les parties
inférieures du corps : la poitrine est rayée, le ventre et le
sous-caudales sont fortement barrés de brun. Les pattes sont de couleur
jaune. L'espèce possède une queue de couleur gris-bleu
barrée de 6 à 8 bandes sombres. Il n'y a pas de dimorphisme
sexuel bien évident mais quelques caractères morphologiques
peuvent les différencier, le mâle semble petit que le femelle et
plus claire. Au niveau de la tête, le male a une tête un peu plus
grande que son partenaire. Ensuite la coloration jaune au niveau du tarse, cire
et le sourcil est plus vive chez le mâle. Et enfin, l'absence de la
couleur tache blanche au niveau de 6ème et 7ème
rémiges chez la femelle est aussi une clé de détermination
du sexe, alors que ces taches sont visibles chez le mal. Le chant est aigu et
rapidement enchainé. C'est une espèce solitaire,
20
discrète et peu farouche (Langrand, 1995). Le
juvénile se diffère de l'adulte par la coloration
générale du plumage brune, ses yeux plus sombres, ses stries sur
le ventre discontinue et ses pattes grises.

Figure n° 7 : Faucon à ventre
rayé, cliché par Rakotoson, 2020
I.2.3. Distribution et statut de l'espèce
I.2.3.1. Epervier de Madagascar (Accipiter
madagascariensis)
L'Epervier de Madagascar est l'un des Accipitres
endémiques de l'île se trouvant dans les parties Nord-est, Ouest
et Est (Langrand 1990, del Hoyo et al. 1994, René de Roland
2013a). Mais cette espèce se présente dans la partie Sud-ouest de
l'île. C'est une espèce naturellement forestière.
De plus, dans le Statut du CITES, l'espèce est
listée dans l'annexe I et le Statut de conservation selon UICN confirme
qu'elle est quasi menacée, Rév. du 12/06/2013. Vue cette aire de
distribution, la taille de population de l'espèce est encore
pratiquement stable, c'est pourquoi son risque d'extinction est
évalué par UICN en 2013 comme ceci.
I.2.3.2. Faucon à ventre rayé (Falco
zoniventris)
Le Faucon à ventre rayé est le seul faucon
endémique de Madagascar. Il est rarement distribué dans
l'ensemble de l'île et surtout absente sur les hauts-plateaux. En plus,
le Faucon est listé dans l'annexe I du CITES. La taille de la population
globale de l'espèce parait encore stable, c'est pourquoi le risque
d'extinction a été évalué à
préoccupation mineure (UICN, 2015).
I.3. 22
MATERIELS UTILISES SUR LE TERRAIN ET LEURS UTILISATIONS
Afin de conduire à bien cette étude, les
matériels suivants ont été utilisés :
> Appareil photo numérique pour prendre les photos
;
> Fiche de collectes, fiche d'enquête, bloc note,
stylos, crayon pour enregistrer les données brutes ;
> Classeur aluminium : pour protéger les fiches en cas
de pluie ;
> Une paire de jumelles: pour visualiser les
espèces qui sont loin et pour l'observation des individus et du contenu
du nid ;
> Un GPS (Global Positioning System) et
téléphone Android (avec application google Earth) pour prendre
les coordonnées géographiques ;
> Une montre avec chronomètre : ceci permet de
mesurer le laps de temps lors des activités ;
> Une balance et une règle graduée pour
mesurer les poids et la taille (longueur et largeur) des oeufs, des poussins et
des individus adultes,
> Une corde nylon et un mètre ruban : pour mesurer
la hauteur du nid et Accessoire d'escalade (corde et jumar) : pour permettre
à l'observation directe à l'intérieur du nid.
> Cartes du site d'étude : pour les mises au point des
sites ;
I.4. METHODES DE COLLECTES DE DONNEES I.4.1.
Période d'étude sur le terrain
Après la recherche bibliographique dans
différents centres de documentation (bibliothèques et sites
internet), nous avons fait une descente sur terrain. Cette dernière
s'est déroulée à partir de 05 Novembre 2019 jusqu'au 15
Mars 2020 soit 04 mois et demi. Cette période correspond à la
saison de reproduction de ces deux rapaces (Hawkins, 1998 ; René De
Roland, 2006 ; Raveloson, 2016).
I.4.2. Technique de recensement pour l'inventaire des
individus et des nids
La recherche d'individu et du nid de ces espèces a
été effectué dans les aires protégées :
réserve Reniala et forêt de Mosa Mangily ainsi que leurs
alentours. Durant les collectes de données sur terrain, nous avons
adopté les méthodes suivantes : enquête et
23
observation directe, la méthode de point
d'écoute et la visite de sentier avec suivi de l'individu. Cette
activité nous permet de déterminer l'effectif des individus et du
nid présent dans le site étudié.
I.4.2.1. Enquête
Cette méthode a pour objectifs de relever tous les
endroits susceptibles d'abriter ces espèces et de comprendre leur
situation face aux diverses pressions et menaces exerçant sur cette
espèce et leur habitat (Rasolonjatovo, 2016). Pour atteindre ces
objectifs, des enquêtes individuelles ont été
effectuées au niveau du villageois aux alentours de ce site.
Durant cette étude, l'enquête envers la
population locale a été généralement faite
individuellement car c'était difficile de faire réunir la
population. De plus, nombreux gens enquêtés sont réticents
et donnent des réponses vagues ou fausses sans résultat
exacte.
Pendant la conversation, on explique d'abord nos missions puis
on décrit brièvement la morphologie des espèces
étudiées en montrant des photos, ensuite on se renseigne sur la
présence ou non des endroits susceptibles d'abriter cette espèce,
et enfin consultons les différentes sortes de menaces et des pressions
qui pèsent sur les espèces et leurs habitat naturel.
I.4.2.2. Observation directe et suivi de l'individu
Après l'enquête, la descente sur terrain a
été effectuée. Dans cette étude, nous avons
effectué le recensement dans deux sites différents : l'un est
à l'intérieur des aires protégées (Parcs Mosa et
réserve Reniala) et l'autre, aux alentours ou aux lisières du
parc. Chaque site est subdivisé en six transects.
L'ensemble des activités se repose sur la collecte des
données concernant les espèces notamment la localisation des
individus, les suivis de comportement des individus et l'étude de la
biologie de reproduction telle que les caractéristiques des nids avec
leurs contenus (oeufs ou poussins) et les activités des couples.
Pour ceux-là, l'observation directe systématique
a été la méthode adoptée. Cette méthode
consiste à explorer des endroits susceptibles d'héberger cette
espèce (Rasolonjatovo, 2016). On a fait l'observation directe
généralement à l'oeil nu mais nécessitant parfois
l'utilisation d'un télescope pour bien observer le comportement des
individus.
24
Pour le suivi de l'individu, cette méthode a
été réalisée le matin de 5h15' à 9h15' et de
15h30' à 18h30' de l'après-midi. Les individus du couple
étudiés (surtout la femelle) ont été
talonnés jusqu'à ce qu'ils nous conduisent vers leur nid, dans
les sites.
I.4.2.3. Méthode de point d'écoute et visite
de sentier
La recherche des individus et du nid a été faite
par la méthode de « point d'écoute » (Blondel et al,
1970) à part de l'observation directe. Pour ce faire, on a marché
lentement sur un transect de 1km, avec 10 points d'écoute distant de
100m. On est resté de 10 min sur chaque point d'écoute. Dans ce
site nous avons fait 12 transects de 1km : 6 à l'intérieur de
l'aire protégée et 6 aux alentours. Chaque transect est
subdivisé en 10 points d'écoute distant de 100m, est passé
3 fois dans trois jours différents afin d'avoir des données
fiables. Cette activité est particulièrement faite pour le
comptage des individus.
I.4.3. Observation du nid
Dès que les nids ont été trouvé,
l'observation a été faite jusqu'à la dispersion des
poussins. Dans cette activité, nous avons adopté deux types
d'observations :
- l'observation directe pour voir le contenu du nid, la
caractéristique du nid et de l'arbre de nidification, l'habitat
environnent...
- et l'observation directe à l'aide d'une paire de
jumelles et d'un télescope constituent les techniques appropriées
à l'étude des nids, en se positionnant à une distance
allant de 10 à 50m afin de bien suivre les mouvements et les
comportements qui se sont passés à l'intérieur du nid et
aux alentours de site nidification.
Pour chaque nid, nos investigations ont été
effectuées de 5h30 du matin jusqu'à 18h de l'après-midi
(13h d'observation). Les deux nids ont été observés
simultanément.
I.4.4. Capture et relâchement
Le but pour la méthode de capture c'est d'identifier la
morphologie des individus adultes et de permettre la mensuration individuelle.
Trois sortes de piège ont été choisies selon leur
efficacité :
? Bal chatri : c'est une sorte de cage en
grillage (L : 40cm, l : 20cm, h : 20cm) muni de plusieurs noeuds en fil de
nylon de 15cm de long à l'extérieur (Berger et Muller, 1959 ;
Thorstrom, 1996). Ces noeuds servent à coincer les serres du
prédateur quand il se pose sur la
25
cage. Pour attirer l'attention de ces rapaces, des
appâts ont été mis tels que des caméléons ou
des lézards sur la paroi extérieure de la cage. Le piège
est placé sur terre et fixé à l'aide de deux cordes sur
les deux côtés pour avoir un équilibre stable. Le
bal-chatri est le plus utilisé pour capturer les Accipitridés de
Madagascar (René de Roland, 2000).
? Bal chatri modifié où les
séries de rangées de noeuds sont étalées au-dessus
d'une plaquette de grillage en plastique (L : 60cm, l : 50cm). Ce piège
est monté soit avec des appâts posés entre les noeuds, soit
cachés seulement tout près de gros cadavres (ex : lézard)
et de façon à ne pas laisser apparaître que les noeuds. Un
individu peut être capturé par cette dernière
démarche lorsque les individus se mêlent dans la dissection du
cadavre. Ce piège a été choisi par son efficacité
lors du baguage des Pygargues de Madagascar, une rapace 17 fois plus de grande
taille que le faucon à ventre rayé et l'épervier de
Madagascar (Andriamalala, 2006). La différence repose principalement sur
le fait que le grillage est caché sous les sables ou des buissons.
? Piège à bâton : c'est
un dispositif formé d'un bâton d'un mètre de long ayant un
poids de 2kg environ et muni de deux noeuds de 40cm de long chacun. Les
appâts qui sont des intestins de volailles ou des cadavres des petits
animaux sont posés sous les noeuds au sol. Cette mode de capture peut
appliquer souvent à la surface du sol en faisant flotter l'appât
sur des buissons, souvent des poussins, gecko ou de petit lézard. Ce
type de piège a été emprunté chez la population
locale. Son avantage repose sur le nombre réduit des noeuds
utilisés, donc plus facile et rapide à fabriquer. Les
pièges sont installés à l'intérieur ou à la
périphérie du territoire de nidification du couple choisi. Il est
à noter que ces rapaces ont l'habitude de raser le sol pour saisir leur
proie (Milon et al, 1973). La mensuration des individus ainsi
capturé se fait sur place puis relâché après avoir
mis une marque (coloration ou bague) sur leurs pattes droites.

|
Figure n° 8 : Piège à
bâton (Andriamalala, 2006)
|
26
Figure n° 9 : Filet de Bal chatri et Bal
chatri modifié
I.4.5. Mensuration et pesage des adultes et des
poussins
Nous avons fait une mensuration et un pesage
systématique des poussins par 5 jours. L'activité de pesage a
été faite en absence des parents. L'objectif de cette
activité c'est de savoir la croissance pondérale et le
développement du poussin. Pour cette étude, la mensuration a
été effectuée à partir de cinquième
(5e jour) de l'éclosion jusqu'à ce qu'ils se
dispersent. En plus, le paramètre morphométrique des poussins, la
durée de leur séjour dans le nid a été aussi
notée.
Les poussins et les individus adultes capturés ont
été mis dans un sac en tissu. La balance a été mise
à zéro avec la masse du sac en tissu fixé sur la balance
même. Les mensurations ont été prises à l'aide d'un
pied à coulisse de 153mm à un centième près et
d'une balance à ressort (pesola) de 100g, 300g, 500g. Pendant cette
activité, les paramètres suivants ont été
considérés : poids, longueur du corps entier, de la queue et de
la rémige primaire. Ici, la méthode standard de Biggs et al.
(1978) n'a pas été totalement appliquée.
I.4.6. Biologie de la reproduction des espèces
Dans ce travail, divers stades de cycle de reproduction ont
été considéré pour les deux espèces
étudiées notamment la ponte et l'incubation, l'éclosion,
l'élevage des poussins dans le
27
nid suivi de l'évaluation de la productivité.
Tandis cette étude a raté la période de formation de
couple et de l'accouplement vue le retard de la période d'étude
sur terrain.
I.4.6.1 Formation du couple et Accouplement
La formation du couple est considérée lorsque
l'un des individus du couple commence à s'occuper de l'autre. De plus,
une construction ou réparation du nid par les deux conjoints constitue
la cohésion du couple. (Rasolonjatovo, 2016). Cette activité
commence par la parade nuptiale et se termine par le choix de l'emplacement du
nid.
I.4.6.2. Ponte
Dès qu'on a trouvé que les nids contiennent des
oeufs, nous avons fait la mensuration et le pesage de tous les oeufs pondus.
Cette activité se réalise dans chaque nids cibles tous le matin
à 5h30' pour savoir le nombre des oeufs et l'intervalle de ponte au
moment que les femelles s'absentent. L'oeuf pondu en premier est marqué
pour éviter la confusion avec le nouveau. Alors dans cette étude,
nous avons enregistré les résultats de la mensuration (longueur
et largeur) et du pesage (poids) des oeufs.
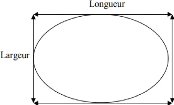
Figure n° 10 : Mode de mensuration des
oeufs
I.4.6.2. Incubation et éclosion
L'incubation est une action des volatiles ou bien des oiseaux
qui couvent leurs oeufs, de manière à les maintenir au chaud et
à permettre le développement de l'embryon. A ce stade, les
adultes commencent à rester sur le nid à partir de ce moment.
Alors après la ponte, nous avons remarqué si l'un du couple reste
sur le nid ou non. Alors, la durée de l'incubation a été
noté dans cette étude, à partir du premier jour
jusqu'à l'éclosion c'est-à-dire la fréquence ou le
temps
28
total effectué entre les deux sexes durant cette
période et aussi des comportements du couple autour du nid tels que
l'échangé de proies.
L'absence d'un couple dans le nid correspond au moment
où aucun individu n'a couvé les oeufs, par exemple si l'individu
est sorti du nid pour effectuer d'autres activités telles que la chasse,
l'auto-alimentation et la surveillance du territoire.
L'éclosion à la période de sortie des
oisillons des coquilles. Elle marque la fin de l'incubation.
I.4.6.3. Elevage et croissance des poussins
A partir de l'éclosion, notre attention
s'intéresse à noter les comportements des adultes envers les
poussins. Alors, dans cette étude nous avons l'opportunité
d'enregistrer la période d'élevage des poussins, la
fréquence de la couvaison c'est-à-dire la participation de deux
sexes relative à la couvaison des poussins, la livraison des proies, la
qualité et la quantité des proies livrées, le nourrissage
des poussins.
Tandis que la détermination de l'âge probable des
poussins se fait par l'identification des poids, du régime alimentaire,
de la morphologie des plumes, des ailes et de la queue.
I.4.6.4. Productivité
La productivité s'est définie comme le nombre de
poussins dispersés par rapport aux nombres d'oeufs éclos dans un
nid donné (Rasolonjatovo, 2016). Pour évaluer la
productivité, il est nécessaire de calculer le pourcentage de
poussins ayant pris leur envol. Le pourcentage de poussins envolés
s'obtient en divisant le nombre des poussins envolés par le nombre total
de poussins éclos puis multiplié par cent.
I.4.7. Etude écologique des espèces
L'écologie est une discipline qui s'intéresse
à étudier la relation des êtres vivants entre eux et avec
leur environnement. C'est donc une étude de condition d'existence et des
comportements des êtres vivants en fonction de l'équilibre
biologique et la survie des espèces. Dans notre cas, nous somme
penché sur l'étude de paramètres suivants : environnement
du nid, habitat des espèces et régime alimentaire.
29
I.4.7.1. Etude de nid
? Description du nid
Le nid a été décrit une fois que l'arbre
du nid a été trouvé. Mais la description complète a
été faite après l'indépendance des poussins. Ce
choix a pour objectif de ne pas perturber les oiseaux. Dans cette étude,
nous avons grimpé sur l'arbre du nid pour voir les matériaux de
construction et mesurer les nids (l'épaisseur, longueur, largeur la
profondeur) fig n°5, la hauteur du nid par rapport au sol. En outre,
l'orientation du nid et la longueur de l'arbre du nid ont été
aussi considérée pour avoir d'idée sur l'ensoleillement du
nid.
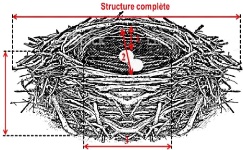
3
1 : Profondeur, 2 : Diamètre interne, 3 :
Diamètre externe ou structure complète
Figure n° 11 :
Schéma d'illustration de la mensuration du nid (Stock, 2017)
? Analyse de l'habitat
Cette partie nous permet de collecter les informations sur la
structure du territoire de la nidification des espèces
étudiées afin de savoir leur exigence écologique en tenant
compte le territoire de chasse, le perchoir et le dortoir.
b.1-Analyse de l'habitat de la nidification
La méthode adoptée pendant l'analyse de
l'habitat est la méthode de Greig-Smith (1983) et l'utilisation de GPS.
La première activité consiste à considérer le
nombre des arbres qui a un diamètre à la hauteur de la poitrine
(DHP) supérieur ou égal de 10cm. Tandis que la deuxième
sert à enregistrer les coordonnées géographiques. L'arbre
du nid est pris comme point de départ pour les quatre quadras.
Nord-ouest, Nord-est, Sud-ouest, sud-est. Un décamètre (15m) et
des rubans ont permis d'établir ces quadras. Pour chaque quadra, l'arbre
le plus proche à partir de
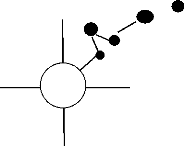
l'arbre du nid a été marqué à
l'aide de rubans et étudié. Ensuite, l'arbre à DHP
supérieur à 10cm le plus proche de ce dernier sera
étudié et ainsi de suite jusqu'à ce qu'on obtienne une
distance totale de 50m. L'arbre déjà étudié a
été marqué à l'aide d'un ruban coloré en
orange, afin de ne pas le réétudier. Le DHP et le nom
vernaculaire de tous les arbres ont été notés ainsi que
leur hauteur.
50m
: Arbre à DHP>10cm
Nord
Ouest
NID
Est
30
Sud
Figure n° 12 : Technique
d'évaluation de l'habitat de nidification selon Greig-Smith, 1983
D'autres paramètres utiles pour faire l'analyse ont été
aussi tels que :
? La distance de chaque arbre du nid par rapport au bord de la
forêt ;
? La distance de chaque nid par rapport à la construction
humaine la plus proche
(maison, route, ...)
b.2 Délimitation du territoire de chasse, perchoir
et dortoir
Le territoire de chasse désigne l'ensemble des endroits
où les individus réalisent les captures des proies. Nous avons
enregistré les coordonnées géographiques et
déterminé la distance entre les points de capture et les nids. En
outre, les données concernant la hauteur des arbres de perchoir par
rapport au sol ont été collectées.
La détermination du dortoir du mâle repose sur le
nom et les caractéristiques de l'arbre de dortoir ainsi que leur
distance par rapport à l'arbre du nid. La délimitation a
été donc estimée par rapport à l'emplacement du nid
en tenant compte les trois facteurs suivants: la localisation des perchoirs
fréquemment utilisés par le couple propriétaire pour
différents usages (bain de soleil, accouplement, transfert de proie,
repos), la distance maximale effectuée par chaque
31
élément du couple pour défendre la
surface occupée lors de la compétition intra- et
interspécifique et l'emplacement du juchoir (endroit de repos durant la
nuit) si celui-ci se situe dans la périphérie de l'arbre du
nid.
I.4.7.2. Régime alimentaire
La détermination du régime alimentaire permet
d'envisager la conservation et de sauvegarder une espèce dans leur
milieu naturel facilitant ainsi les moyens de définir sa niche
écologique et de s'informer sur son niveau trophique (René de
Roland, 2010). L'identification de l'alimentation de chaque espèce est
connue grâce à l'identification des proies vues ou
apportées par des individus, ou encore par les restes de nourritures
récoltées dans les nids ou dans les dortoirs soit aussi à
partir des pelotes de régurgitations. Les observations
directes pendant la période de reproduction ont permis d'identifier les
proies apportées par chaque individu du couple. Pour compter et
quantifier les proies, chaque apport effectué par l'individu du couple
est considéré comme une nouvelle proie. Par contre pour les
restes de proies trouvées, seul leur présence est
notée.
I.4.7.3. Evaluation des menaces et des pressions dans
l'ensemble d'habitat
Lors de notre étude sur le terrain, toutes sortes de
menace anthropique et naturelle ainsi que leurs impacts sur le mode vie de ces
deux espèces de rapaces ont été étudié comme
au niveau de la structure de l'habitat de nidification et du régime
alimentaire. Pour ceux-là, certains paramètres étaient
utilisés pour évaluer le degré des menaces et pressions
pesant sur l'espèce et son habitat. Ces paramètres sont les
suivants : la collecte des oeufs, l'enlèvement des aiglons au nid et la
chasse, le nombre des grands arbres coupés en tenant compte leurs
distances par rapport aux arbres de nidification, le défrichement, le
feu de brousse et les facteurs climatiques.
Pour combler les informations, des enquêtes ont
été effectuées auprès de la population riveraine et
des agents des sites protégés.
I.4.8. Comportements des adultes
Pour cette étude, le comportement se définit
comme étant la séquence d'une ou plusieurs activités
effectuées par un individu quelconque afin d'atteindre un but, à
un moment bien précis (Raveloson, 2016). La méthode d'observation
directe et continue est une méthode adoptée pour l'étude
de comportement (Martin & Bateson, 1993) en utilisant d'un télescope
et une paire des jumelles pour noter les activités et une montre
à chronomètres enregistrer les durées des
évènements données. Cette méthode permet donc
d'enregistrer le type d'activité et d'analyser
32
les fréquences et la durée des
évènements se sont déroulés. Dans cette
étude, le suivi de comportement des couples s'est fait surtout pendant
l'observation du nid et l'analyse de l'habitat. Alors, la relation intra- et
interspécifique des couples de ces rapaces a été
notée.
I.5. ANALYSE DE DONNEES
Cette démarche nous a permis de vérifier
statistiquement nos hypothèses. Afin d'effectuer cette analyser, nous
avons utilisé les logiciels : XLSTAT 9, version 2013 pour le test
statistique et Microsoft Excel 2013 pour la manipulation et le formatage des
données. Avant de procéder à toutes les analyses, toutes
les données collectées sont enregistrées sous forme de
base de données. Alors, ce chapitre nous exhibera la démarche a
suivi pour l'analyse des données collectés sur terrains.
I.5.1. Moyen et écart-type
Certains résultats sont présentés sous
forme de moyenne accompagnée de l'écart-type (moyenne#177;
écart-type) selon les formules suivantes :
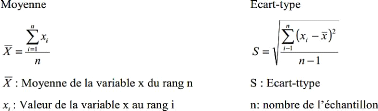
33
I.5.2. Test de Chi-deux (÷2)
Le test de ÷2 sert à tester
l'homogénéité des faits observés avec ceux à
espérer pour deux ou plus d'échantillons (Jonshon, 1992). Ce test
est le plus utilisé et conseillé pour les ornithologues (Fowler
& Cohen, 1985). Dans cette étude, il a été
utilisé afin de comparer la distribution des effectifs de population
dans les zones protégées et non protégées. Ce test
est employé pour vérifier les hypothèses suivantes :
Hypothèse 1 :
H01 : « La distribution des effectifs des populations dans
les zones protégées et non protégées est
homogène ».
Ha1 : « La distribution des effectifs des populations dans
les zones protégées et non protégées est
hétérogène ».
34
Hypothèse 2 :
H02 : L'effectif de ces deux rapaces dans la zone
étudiée est égal Ha2 : L'effectif de ces deux rapaces dans
zone étudiée est différent La valeur de ÷2 est
obtenue par la formule suivante (Jonshon, 1992) :
Où : Oi : valeur observée et Ci : valeur
calculée (théorique) Calcul de la fréquence
calculée
Les données initiales étaient groupées dans
un tableau de "l" lignes et de "c" colonnes. L'effectif
calculé d'une case est le produit du total de la ligne par le total de
la colonne, divisé par le total général.
Le degré de liberté (d.d.l) est obtenu par :
d.d.l = (Nombre de ligne - 1) (Nombre de colonne - 1).
Dans le cas où d.d.l = 1, il faudrait passer à la
correction de Yates (Fowler et Cohen, 1985) pour chaque variable
considérée dont :
(/0-C/-0,5)2
C
X2 yate =
La valeur de ÷2 à la fin de la correction de Yates
sera 2 X ÷2yate.
Pour évaluer le tes de Chi-deux, la valeur de ÷2
ainsi obtenue (÷2cal) est comparée à celle donnée par
la table (÷2tab). La probabilité de risque est de 5% avec
l'hypothèse nulle H0 : « Il n'y a pas de
différence significative entre les variables testés »,
donc :
- Si ÷2cal >÷2tab la différence est
significative donc H0 rejetée
- Si ÷2cal < ÷2tab H0 acceptée
PARTIE II : RESULTATS ET INTERPRETATIONS
37
Cette partie nous présente les résultats de
cette recherche. Elle renferme les informations des espèces
étudiées concernant : l'effectif des populations présentes
et le nombre et les caractéristiques du nid dans les zones
étudiées, la morphologie et la morphométrie des individus,
la biologie de la reproduction, l'écologie et les comportements.
II.1. NOMBRE DES INDIVIDUS ET DES NIDS TROUVES
La recherche des individus et du nid basant sur
l'enquête, l'observation directe, suivi de l'individu avec une
méthode d'écoute pendant le mois de novembre 2019, (du 5 à
20 Novembre soit 15jours) nous donne les résultats suivants :
II.1.1. Nombre des individus
Pour cette étude, nous avons trouvé au total 29
individus adultes : 11 Accipiter Madagascariensis et 18 Falco
zoniventris. Le résultat de cet inventaire est
présenté sur le tableau suivant :
Tableau n° 1 : Nombre des individus
inventoriées
|
Espèces inventoriées
|
Dans les aires protégées ou Site 1
|
Aux alentours
ou Site 2
|
Total
|
|
Accipiter Madagascariensis
(Population A)
|
09
|
2
|
11
|
|
Falco zoniventris
(Population B)
|
12
|
6
|
18
|
|
Total
|
21
|
8
|
29
|
Le tableau ci-dessus affirme que les Accipiter
Madagascariensis sont moins nombreux que les faucons à ventre
rayé. Les deux espèces fréquentent beaucoup le milieu
forestier du site 1. Ceci est prouvé par notre analyse statistique lors
du test de khi-2 :
? Pour l'effectif des individus dans deux sites :
Ici, la valeur observée est supérieure à
la valeur seuil, ÷2cal 0,468>÷2tab 0,05, alors l'hypothèse
nulle (H01) est rejetée en faveur de l'hypothèse alternative Ha1,
c'est-à-dire la distribution des effectifs des populations dans les
zones protégées et non protégée est
hétérogène, ces rapaces sont nombreux dans le site 1 par
rapport au site 2.
38
? Pour la comparaison des effectifs entre les deux rapaces dans
notre zone d'étude,
les Falco zoniventris (population B) tient l'effectif
élevé que les Accipiter madagascariensis (population A).
Ceci affirme que les effectifs de ces deux espèces ne sont pas
égaux dans notre zone d'étude. C'est qui veut dire que le H02 est
rejetée en faveur de Ha2 (Ha2 : L'effectif de ces deux rapaces dans la
zone étudiée est différent).
L'abondance relative de ces rapaces par rapport à
l'effectif total des espèces d'oiseaux présents dans ce site est
relativement très faible. Ces espèces ne sont pas colonies, ils
vivent soit seul soit en couple. Parmi les 29 individus trouvés, 2
paires des A. madagascariensis ont été en couple et 2
paires pour le F. zoniventris. Ceci nous a permis d'étudier les
paramètres de nidification dans notre zone d'étude.
II.1.2. Nombre des nids
Dès qu'on a bien identifié les individus en
couples, notre attention se concentre sur le suivi de ces individus afin de
trouver leurs nids. Cette activité s'est réalisé
évidement dans chaque site étudié. En effet, nous avons
localisé 5 nids (4 nids actifs et 1 nid abandonné) :
? 2 pour la population A, tous nids se trouvent dans le site1.
Aucun nid n'a été trouvé dans le site 2. Le nid 1 a
été trouvé le 09 Novembre 2019 sur les coordonnées
géographiques Latitude sud : 23°07'28» et Longitude Est
43°37'44,2» tandis que l'autre (nid 2) s'est trouvé le 11
Novembre 2019, distant environ 2,2km et sud Est par rapport au nid 1 sur les
cordonnées Latitude sud : 23°07'12,7» et Longitude Est
043°37'27,8».
? 3 pour la population B, l'un dans le site 1 appelé
nid 3, a été trouvé le 5
Novembre 2019 sur les
coordonnées géographiques suivants : Latitude sud :
23°07'21,8» et Longitude Est 043°37'43,4» et les autres aux
alentours du parc (un nid actif et un nid abandonné). L'autre nid actif
dans le site 2 appelé nid 4 a été trouvé en premier
le 18 Novembre 2019 sur les coordonnées suivants Latitude sud :
23°07'22» et Longitude Est 043°37'13,8»avant la
découverte du nid 5 abandonné un jour après sur les
coordonnées Latitude sud : 23°07'18,4» et Longitude Est
043°37'32,6».
Ce résultat affirme que l'habitat forestier du site 1
est indispensable à la reproduction pour la population A tandis que le
choix du site de nidification est facultatif pour la population B.
39
II.2. MORPHOMETRIE
Les résultats de cette partie sont obtenus à
partir de l'observation directe nous permettent de faire la description
morphologique et morphométrique des individus observés et
capturés. Pendant la capture, nous avons piégé 4
individus, 2 mâles et 2 femelles, respectivement 1 individu mâle et
1 femelle pour chacune de deux espèces. Les individus capturés
ont été les couples suivis.
Le couple de l'Epervier de Madagascar a été
capturé le 15 (le mâle) et 17 (la femelle) décembre 2019.
Tandis que le couple de faucon a été capturé le 26 (la
femelle) et 29 (le mâle) novembre 2019. Après la capture, la
mensuration du poids et la longueur du corps a été faite pour les
quatre individus adultes capturés. Des extraits de résultats de
cette mensuration pour la dimension corporelle obtenue sur ces deux couples
sont montrés sur le tableau ci-dessous suivant.
Tableau n° 2 : Résultats de la
mensuration et du pesage des individus adultes capturés
|
Espèces
|
Sexes
|
Longueur du
corps (cm)
|
Poids (en g)
|
Longueur de la
rémige I (cm)
|
|
Epervier de Madagascar ou Accipiter madagascariensis
|
Mâle
|
33
|
298
|
18,05
|
|
femelle
|
38,42
|
344
|
23,12
|
|
Faucon à ventre rayé ou Falco
zoniventris
|
Mâle
|
34
|
146
|
11,3
|
|
Femelle
|
36
|
178
|
12
|
Ce tableau nous montre qu'il y a une différence
morphologique entre les deux sexes au niveau de taille pour les deux
espèces. Les femelles semblent plus avantageuses que les males surtout
au niveau de poids.
Chez l'Epervier de Madagascar, la femelle tient la taille plus
grande que le mâle, le dimorphisme sexuel est bien évident au
niveau de la taille, poids, et de la longueur de la rémige à part
de la coloration.
Tandis que chez le faucon à ventre rayé, la
différence de longueur du corps entre les deux sexes semble moins
évidente. En effet, la détermination du dimorphisme sexuel peut
s'avancer à l'aide de la taille, particulièrement au niveau de
poids.
Vue les résultats sur ce tableau, l'Epervier de
Madagascar est une espèce de rapace de grande taille que le faucon
à ventre à rayé.
40
II.3. BIOLOGIE DE LA REPRODUCTION
II.3.1. Période de formation de couple, accouplement
et ponte
La formation du couple est l'ensemble des comportements ou des
activités de couple avant la période de la reproduction en vue de
l'accouplement. Tandis que ce dernier signifie la relation sexuelle directe
entre le couple. Les données de notre étude sur terrains ne
permettent pas de répondre cette partie à cause du retard pour la
descente sur terrain. En effet, notre effort lors de la recherche
bibliographique nous permettra d'avancer quelques informations dans la partie
des discussions.
II.3.2. Période de Ponte et taille des oeufs
La ponte se réalise après la construction du
nid. Comme le cas des activités ci-dessus, nous avons manqué
l'enregistrement de l'intervalle de ponte pour les couples
étudiés.
Pour l'Epervier de Madagascar, cette activité commence
à partir de début de mois d'octobre et le nombre des oeufs par
nid varie entre 3 à 4. La dernière ponte a été
trouvée pour le nid 2, le 14 novembre2019. Pour les deux nids
trouvés, ils contiennent au total 7 oeufs, chacun contient en moyenne
3,5 (n=3oeufs nid1 et n=4 oeufs nid 2). Ces oeufs sont colorés
uniformément blancs ou avec des marques plus sombres variables. La
largeur des oeufs varie entre 36,1 et 36,3 mm et entre 39 et 39,5 pour sa
longueur et ils pèsent entre 30 à 34g (n= 7 oeufs). Le premier
oeuf semble plus petit que les autres derniers.
Pour le faucon à ventre rayé, la période
et l'intervalle de ponte n'ont été pas enregistrée, tous
les nids ont été trouvés durant la période de
l'incubation. Mais ceci ne nous a empêchés pas d'identifier leurs
contenus. Le nid 3 qui se trouvant dans le site I (dans l'aire
protégée) contient 2 oeufs tandis que l'autre (nid 4)
situé dans le site 2 (aux alentours du parc) contient 3 oeufs. L'oeuf de
cette espèce pèse entre 23,80 à 25g (n= 5). Il mesure
entre 32 et 32,4 mm pour sa largeur et entre 38 à 38,4 pour sa
longueur.
II.3.3. Incubation et éclosion
Ce sous chapitre va nous exhiber les données concernant
la période de l'incubation et sa durée pendant la photophase,
ainsi que l'information sur le taux d'éclosion.
41
II.3.3.1. Incubation
Pour tous les nids étudiés pour les deux
espèces, les femelles seules occupent l'activité incubatrice
entre les couples, tandis que les males assurent le ravitaillement des proies.
La livraison des proies se réalisent en dehors de l'arbre de
nidification, distant environ 15 à 30 m par rapport à l'arbre du
nid.
Pour l'Epervier de Madagascar, le repérage de la
durée d'incubation a été faite sur le nid 2, cette
activité dure 35 jours. La fin de l'incubation est marquée par
l'éclosion. Pour cette couple, la femelle occupe l'incubation 83,7% pour
les 162 heures du temps d'observation pendant la phase diurne, tandis que le
reste du temps, les nids est abandonnées au moment que la femelle
cherche des proies ou réalisent d'autres activités. Pendant la
nuit, la femelle reste sur le nid, continue l'incubation tandis que le
mâle reste sur son dortoir tous près mais en dehors de l'arbre du
nid.
Tous les 2 nids du Faucon à ventre rayé ont
été trouvés en plein période d'incubation,
après 8 jours de la découverte du nid 3 dans le site 1, tous les
oeufs ont été éclos. Tandis que l'éclosion du
deuxième nid a été observée juste 4 jours
après sa découverte. Ceci nous rend difficile à
déterminer la durée de l'incubation. Mais notre approche
bibliographique avance les informations concernant cette activité dans
la partie des discussions.
II.3.3.2. Période d'incubation et taux
d'éclosion
Pour l'Epervier de Madagascar, le taux de réussite a
été 100% (n = 7 poussins pour les nids 1 et 2).
Les oeufs du nid 1 ont été tout éclos le 3 décembre
2019 tandis que ceux du nid 2 ont été éclos le 16
décembre 2019. Ceci nous permet donc d'estimer que l'incubation du nid 1
a été commencée le 30 octobre 2019. Le taux de
réussite a été 100% (n=7 poussins pour les nids 1et 2)
Pour les Faucon à ventre rayé, les oeufs du nid
3 ont été éclos le 13 Novembre 2019 tandis que les autres
oeufs se trouvant dans le nid 4 du site 2 ont été éclos le
22 novembre 2019. Ces deux nids ont un taux d'éclosion 100% qui donne au
total cinq (05) poussins issus des 5oeufs de deux nids (nid 3=2 poussins et nid
4= 3poussins).
Après l'éclosion, les poussins présentent
morphologiquement une couleur presque entièrement blanche qui sont les
duvets pour les deux espèces particulièrement la partie ventrale
et dorsale. Tandis que le bec, la zone perioculaire et les pattes sont nus avec
une couleur au
42
début jaune sombre s'évolue en gris
foncé. Ces colorations s'évoluent et varient suivant l'âge
des poussins
II.3.4. Elevage des poussins dans le nid
L'élevage des poussins est l'ensemble des
activités du couple envers leur génération à partir
de l'éclosion jusqu'à l'indépendance total des
poussins.
Pour l'Epervier de Madagascar, l'observation a
été faite sur le nid 1 à partir de l'éclosion 03
décembre 2019. Les poussins commencent à quitter leur nid
à partir de 35é jours pour le mâle et
38é jours pour les femelles après l'éclosion
mais ils retournent au nid au soir. Ces poussins deviennent indépendants
total aux 61 à 63é jours après
l'éclosion pour les mâles et 65 à 66é
jours pour les femelles. Mais entre cette période leur vie est à
la charge des parents comme la couvaison, le nourrissage, la défense,
etc. Pendant ces 66 jours, le temps d'observation a été 252
heures. La couvaison a été seulement effectuée par la
femelle tandis que le mâle assure la capture et le ravitaillement des
proies. Ces derniers ont été livrés à la femelle en
dehors de l'arbre du nid puis la femelle assure le nourrissage des poussins. La
femelle reste toujours sur le nid presque 92% du temps d'observation pendant la
phase claire pour la couvaison sauf en cas de livraison des proies. Pendant
cette étude, 72 proies ont été identifiées
livré par le mâle, 82% Oiseaux, 8% reptiles (lézard et
gecko...), 6 % insectes et 4% non identifié. Ensuite, la quantité
des proies données aux poussins diminue à partir de
38é jours de l'âge des poussins. Les poussins du nid 1
ont été tous disparus le 07 février 2020, tandis que ceux
du nid 2, le 21 février 2019.
Pour le faucon à ventre rayé, l'observation des
activités du couple pour l'élevage des poussins a
été faite au nid 3 à partir de l'éclosion qui a eu
lieu le 13 novembre 2019 jusqu'à l'indépendance total des
poussins. Celle-ci dure 62 heures au total pour le temps d'observation. La
femelle seule assure la plupart du temps environ 60% pour la couvaison et le
nourrissage des poussins tandis que le mâle assure principalement la
capture et le ravitaillement des proies mais ceci n'empêche pas la
femelle de se débrouiller. A partir de 10 jours de l'âge des
poussins, la femelle quitte le nid à la fois pour se nourrir et chercher
des proies pour leurs progénitures. A partir de 27 à
31éme jours de l'âge, les poussins apprennent à
voler et à capturer leur proie. A partir de ce moment la quantité
des proies livrés par les parents commence à diminuer mais cette
intervention continue jusqu'à ce qu'ils soient indépendants
totalement aux 53 à 55è jours
43
d'âge. Les poussins du nid 3 et du nid 4 ont disparus
respectivement le 07 janvier 2019 et le 15 janvier 2020.
Bref pour ces deux espèces des rapaces, les femelles
occupent principalement l'incubation des oeufs, la couvaison et le nourrissage
des poussins tandis que les mâles s'occupent les captures et les
ravitaillements des proies.
II.4. MENSURATION ET PESAGE DES POUSSINS
Les résultats de la mensuration servent à
connaitre le taux de croissance pondérale et morphométrique des
poussins. Alors les données du pesage sont présentées dans
le tableau ci-dessous. La mensuration a été faite tous les cinq
jours et a été commencée à partir de
cinquième jour jusqu'à vingt cinquième jour d'âge
des poussins car à part de cette période, les poussins commencent
à quitter leur nid et il est devient difficile de les capturer pour
poursuivre la mensuration.
Evolution pondérale des poussins en fonction
l'âge
|
Poids en g
|
180 160 140 120 100 80 60 40 20
|
|
|
0
|
5éj 10ej 15ej 20ej 25j
|
|
Jour de mensuration
|
|
Accipiter
madagascariensis
|
42
|
75
|
108
|
140
|
170
|
|
Falco zoniventris
|
27
|
49
|
82
|
111
|
129
|
Accipiter madagascariensis Falco
zoniventris
Figure n° 13 : Poids des poussins par
cinq jours
Le tableau et le figure ci-dessus affirme que le poids des
poussins s'évolue rapidement et presque constante. Pendant cette
période, les adultes se débrouillent beaucoup pour nourrir et
défendre leurs progénitures.
44
II.3.4. Productivité
Pour l'Epervier de Madagascar, la productivité est
égale à 100% car les sept poussins issus de sept oeufs de deux
nids donnent sept individus qui ont réussi envol au moment dispersion
des poussins. Aucune pression et menace n'a été observée
chez les poussins.
Pour le Faucon à ventre rayé, la
productivité est égale 80% car l'un des poussins issus des cinq
oeufs est mort. Il a été trouvé sous l'arbre du nid 4,
situé dans le site 2 lors de la pluie et du vent violent du 15 et 16
décembre 2019. Par contre, les quatre poussins restants ont
réussi à s'envoler jusqu'à l'indépendance total.
En général, le succès de la reproduction
de ces deux espèces est élevé et atteint la valeur moyenne
de 90%.
45
II.4. ECOLOGIE
II.4.1. Caractéristique des nids ? Emplacement des
nids
Pour l'Epervier de Madagascar, tous les nids ont
été trouvés dans le milieu forestier des zones
protégées. Le nid 1 s'est situé dans le Parc Mosa, est
placé à l'extrémité des deux petites branches en
forme d'une fourche d'un baobab fony (Adansonia rubrostipa),
14m de hauteur avec une DBH de 132cm. Cet arbre se situe dans le milieu
forestier mais tous près d'une piste du circuit de parc, avec une
distance de 14m. Ce nid se place à 12m de haut par rapport au sol. Ceci
se situe dans les coordonnés suivants : Latitude sud :
23°07'29» et Longitude Est 43°37'42»
Tandis que le nid 2, s'est trouvé à
l'intérieur de la réserve Reniala, s'est placé sur des
quatre branches en forme « V » de Boy Commifora lami. Cet
arbre possède une hauteur de 17m avec une DBH de 44cm. La distance du
nid par rapport au sol est à 14, 3m. Ceci se trouve dans le
coordonné suivant : Latitude sud : 23°07'13,1» et Longitude
Est 043°37'29,1»
Les nids des Epervier de Madagascar montrent une ouverture en
haut et se placent dans la partie Est de l'arbre de nidification afin de
bénéficier le rayon solaire pour le matin. Mais ces nids sont
généralement cachés sous des feuilles en raison
d'éviter la chaleur du rayonnement solaire et les prédateurs des
oeufs et des poussins. Les hauteurs de l'arbre du nid ont une moyenne de 15,5 m
(n=2 arbres du nid). Le choix de l'emplacement du nid est donc en raison de la
sécurité.
Pour les faucons à ventre rayé, le nid 3
localisé dans la réserve Réniala se situe dans les
coordonnés Latitude sud : 23°07'22,1» et Longitude Est
43°37'42,3», est placé sur un Farafatsy (Givotia
madagascariensis) de 13m de hauteur et avec une DBH de 98 cm. Le nid a
été placé de 8m par rapport au sol.
Tandis que l'autre, (nid 4) situé dans le milieu ouvert
en dehors du parc se situe sur les coordonnées Latitude sud :
23°07'21,9» et Longitude Est 043°37'14,3», se repose sur un
manguier (Mangifera indica) de 14m de haut avec une DBH de 143cm. Le
nid a été porté par des branches en forme d'une fourche de
cet arbre et situé une hauteur de 11m par rapport au sol. Ce nid se
situe au bord de l'agroforesterie de l'ONG Bel Avenir et distant de 102m d'une
habitation humaine. Le nid 5 non actif déjà abandonné
s'est trouvé sur un pied de Tamarinier
46
(Tamaridus indica), 15m de haut avec une DBH 173cm.
Cet arbre de nidification distant de 2,2km par rapport à l'arbre du Nid4
du site 2 et 540m par rapport au nid1 du site 1. Il se situe
précisément sur les coordonnées Latitude sud :
23°07'33,1» et Longitude Est 043°37'29,1».
Bref, l'hypothèse 3 est annulé car l'arbre de
nidification est différent pour les deux espèces voir même
pour les individus d'une même espèce.
? Dimension de nid
Pour déterminer les caractéristiques de nids,
nous avons faites une observation directe et une mensuration après la
dispersion totale des poussins. Tous les nids pour les deux espèces se
ressemblent au niveau des matériaux de construction. Ils sont
généralement construits à base des petites branches ou des
rameaux avec des diamètres varient entre 1 à 2,5 cm et de 14
à 48cm de longueur, des feuilles mortes et des herbes sèches dans
la partie internes.
Les paramètres considérés et les
données de la mensuration sont présentés sur le tableau
suivant :
Tableau n° 3 : Forme et dimension des
nids
|
Espèces
|
Accipiter madagascariensis
|
Falco zoniventris
|
Moyenne
|
|
Nids
Paramètres considérés
|
Nid 1
|
Nid2
|
Nid3
|
Nid4
|
N1& N2
|
N3&N4
|
|
Diamètre total du nid (cm)
|
38
|
36
|
35
|
36,5
|
37
|
35,75
|
|
Largeur total du nid (cm)
|
34
|
35
|
32
|
34
|
34,5
|
33
|
|
Diamètre interne
|
31,5
|
30
|
28
|
31
|
31,25
|
29,5
|
|
Hauteur (cm)
|
22
|
21
|
16
|
18
|
21,5
|
17
|
|
Profondeur intérieur (cm)
|
13
|
12
|
09
|
11
|
12,5
|
10
|
Rakotoson, 2020
Accipiter madagascariensis Nid 1 Accipiter madagascariensis
Nid2
Falco zoniventris Nid3 Falco zoniventris Nid4
40
30
20
10
0
Accipiter madagascariensis Nid 1
Accipiter madagascariensis Nid2
Falco zoniventris Nid3
Falco zoniventris Nid4
47
Figure n° 14 : Résultats de la
mensuration des nids
Le tableau et la figure ci-dessus montrent que les formes des
nids entre les couples des mêmes espèces sont presque semblables
avec une différence seulement entre de 1 à 2cm pour chaque
paramètre étudiée. Tandis que cette différence peut
atteindre de 5 cm entre les deux espèces. Les nids de l'Accipiter
madagascariensis sont un peu plus larges que ceux des Falco
zoniventris. Ces écarts sont quasiment constants pour chaque
paramètre considéré et pratiquement moins évidents
pour la même espèce.
II.4.2. Analyse de l'habitat de nidification
Ce sous chapitre donne la structure et la
caractéristique des territoires aux alentours de l'arbre du nid. Pour
cette étude, nous avons inventorié au total 342 pieds d'arbres.
Ces arbres se répartissent de façon inégale pour chaque
territoire d'un nid donné surtout entre les deux sites. (Tableau
n°4).
Tableau n° 4 : Résultats de la
mensuration des arbres aux alentours de nid
|
Type de
site
|
Habitat du
nid
|
Nombre de pied
d'arbres
enregistré
|
Nombre
d'espèces
d'arbre
|
Hauteur
moyenne
(en m)
|
DBH
moyenne
(en cm)
|
|
Site 1
|
Nid 1
|
123
|
25
|
13,5#177;4,2
|
33,12#177;8,3
|
|
Nid 2
|
111
|
22
|
14,8#177;2,1
|
41,22#177;1,1
|
|
Nid 3
|
97
|
18
|
15#177;2,2
|
38,21#177;2
|
|
Site 2
|
Nid 4
|
11
|
6
|
14,3#177;3,3
|
39, 6#177;12,1
|
Rakotoson, 2020
48
Ce tableau affirme que les territoires du nid du site 1
renferment beaucoup des pieds et d'espèces d'arbres que du site2. Ceci
est dû à la conservation de l'habitat entant qu'un habitat
protégé. Le territoire du nid 1 est plus riche en pied et en
espèce d'arbre tandis que le nid2 tient le record au niveau du DBH par
rapport au territoire des autres nids. Les hauteurs moyennes des arbres dans
tous les territoires des nids varient entre 13,5#177;4,2 à 15#177;2,2 m
avec une moyenne totale de 14,4 (n=4 territoires des nids). Ils se
présentent quasiment d'une même strate.
II.4.3. Territoire de chasse, perchoir et dortoir ?
Territoire de chasse
Pendant le suivi des couples, les Epervier de Madagascar
occupent un vaste et variable territoire pour la chasse, des milieux forestiers
des zones protégées et des milieux ouverts aux alentours du parc
voire dans les zones d'habitation humaine pour chasser les poussins domestiques
tandis que les milieux arborés ou forestiers sont le plus
utilisés pour chasser les petits reptiles et des oiseaux. Pour les deux
couples suivis, les proies ont été capturées
généralement entre 18 à 1600m par rapport à l'arbre
du nid. Mais cette espèce utilise fréquemment l'habitat forestier
pour cette activité, rarement dans le milieu ouvert.
Pour le Faucon à ventre rayé, les territoires de
chasse sont facultatifs, l'espèce peut chasser même sur l'arbre de
nidification, pour la capture des insectes mais les territoires utilisés
peuvent s'étendre dans une distance plus de 1230m par rapport à
l'arbre du nid. Le couple dans le site 1 chasse souvent dans le milieu
forestier de la zone protégée tandis que l'autre couple
fréquente le milieu ouvert aux alentours du nid mais le mâle peut
parfois effectuer la chasse dans le milieu forestier du site 1. En
général, l'endroit de chasse est totalement variable.
? Perchoir et dortoir
Pour l'Epervier de Madagascar, les individus
fréquentent de se jucher sur un arbre ayant une hauteur moyenne de 9,5m
(n=24 pieds d'arbres) de haut. L'arbre de perchoir distant de 100 à 250m
de cibles de chasses et l'arbre de dortoir distant entre 80 à 150m de
l'arbre du nid pour le mâle. Parmi ces 24 pieds d'arbre constitués
par cinq espèces, les Didiera trollii (pour le juchoir) et les
baobabs (pour le dortoir) sont les types d'arbres les plus utilisés par
les individus.
Pour les faucons à ventre rayé, le couple 1 du
site 1 utilise habituellement les pieds d'arbres ayant une hauteur moyenne de
8,2m de haut (n=13pieds d'arbres). Cette espèce utilise
fréquemment Didiera trollii surtout pendant la chasse car ces
espèces cherchent des insectes
49
comme des papillons et autres, cette plante attire beaucoup
d'insectes surtout pendant la floraison. Mais parfois ces faucons se perchent
sur un arbre comme Adansonia sp, Givotia sp, Alluaudia
sp et commiphora sp. Ces sont des pieds d'arbres ayant une
hauteur un peu élevé environ 11m de hauteur, utilisé
surtout pour le dortoir.
Tandis que le couple 2 dans le site 2 (site non
protégé), utilise les pieds d'arbres un peu plus
élevé que ceux utilisé dans le site 1 avec une moyenne de
14m de haut (n=9 pieds d'arbres). Les espèces d'arbres le plus
utilisées pour le juchoir et le dortoir sont : le Tamarinier
(Tamaridus indica), le Manguier, Eucaliptus et Accacia
sp.
II.4.3. Régime alimentaire
Cette sous chapitre exhibe les données concernant les
informations sur la quantité et la qualité des régimes
alimentaires des espèces étudiées. Dans cette
étude, nous avons seulement tenir compte les proies capturées et
portées par les adultes du couple. L'observation a été
faite à partir de la période de l'incubation jusqu'à
l'indépendance total des poussins. L'alimentation de chaque
espèce est connue grâce à l'identification des proies vues
ou apportées par des individus, ou encore par les restes de nourritures
récoltées sous l'arbre du nid. L'alimentation diffère
surtout du point de vue qualitative et quantitative selon la période
d'observation, elle est composée en grande partie d'une grande
variété d'insectes et de petits vertébrés comme les
oiseaux, les lézards, les geckos et les caméléons.
Pour les couples d'Epervier de Madagascar, 182 proies ont
été recensées (99 pour le couple 1 et 83 pour le couple
2). Parmi eux, 81,86% (n= 149 proies) ont été identifiés
et 18,14% (n= 33 proies) restent non identifiés lors de la taille
parfois plus petite ou problème du choix des points d'observation semble
un peu loin du nid. En général par rapport aux proies
identifiées, la ration alimentaire est constituée principalement
par des oiseaux forestiers des petites taille (n=72 proies) et des poussins
domestique (n=34 proies), au total 71,14% (n=106 proies) mais ces oiseaux
chassent aussi des reptiles 21,47% (n=32proies) comme les geckos (Phelsuma
sp) (n=5 proies), les lézards (Chalaradon sp) (n=8 proies)
et les caméléons (n=19 proies) et des insectes 7, 38%
(n=11proies) dans les ordres des orthoptères (criquets et des papillons)
et des coléoptères (fig n°8). Aucuns serpents et des
grenouilles n'ont été identifiés. Ces proies sont
généralement capturées sur le sol à l'aide de leurs
serres mais parfois capturés en vol particulièrement les
papillons. Ces résultats nous permettent de dire que les Eperviers de
50
Madagascar ne sont pas des espèces totalement
spécialistes mais seulement on peut dire que la ration est
essentiellement basée en oiseaux.
Pour les Faucons à ventre rayé, 88 proies ont
été chassés au total pour les deux couples
étudiés, 78, 40% (n=69 proies) parmi eux ont été
identifiés tandis que les restes 11,59% (n=19 proies) sont inconnue
comme de l'épervier de Madagascar. Par rapport aux proies
identifiées, la ration est constituée principalement par des
petits reptiles (geckos (n=5 proies), caméléons (n= 21 proies),
lézards (n=12 proies), au total 55,07% (n=38 proies) et des insectes
(Papillons (n=17 proies), Menthes religieuses (Mentis sp) (n=2),
criquets (Locusta migratoria et Nomadocris septemphasciata)
(n=7 proies) et des libellules (Libellula sp) (n=5)) soit 44,92% (n=
31proies) au total (fig n°15). Aucuns oiseaux et serpent n'ont
été capturés et livrés pendant cette étude
pour cette espèce. La plupart des proies sont capturés sur le sol
mais certains insectes sont chassés en vol ou sur d'autres substrats
comme les geckos. Les Faucons à ventre sont des espèces
polyphagiques et leur alimentation se base principalement sur les petits
reptiles et des insectes.
Pour les deux espèces étudiées, les
insectes sont généralement chassés pendant la
période d'élevage des poussins pour nourrir les poussins. Tandis
que les adultes des Eperviers de Madagascar préfèrent beaucoup
des oiseaux et des petits reptiles pour les Faucons à ventre
rayé.
|
160 140 120 100 80 60 40 20
|
|
|
0
|
Insectes
|
Reptiles
|
Oiseaux
|
Total
|
|
Accipiter madagascariensis
|
11
|
32
|
106
|
149
|
|
Falco zoniventris
|
31
|
38
|
0
|
69
|
Accipiter madagascariensis Falco zoniventris
Figure n° 15 : Les régimes
alimentaires des oiseaux étudiés
51
Cette figure montre que les deux espèces ont à
peu près la même proportion au niveau des reptiles, mais
l'alimentation est basée principalement en oiseaux pour les Epervier de
Madagascar. Tandis que pour les Faucons, leur alimentation se base en insectes
et reptiles.

Figure n° 16 : Faucons à Ventre
rayé sur le perchoir avec les proies capturées, Reniala 2020
II.4.4. Pressions et menaces
Les deux espèces étudiées ont les
degrés des menaces différentes mais en général, ils
sont liés à l'activité anthropique.
Pour les Eperviers de Madagascar, leur menace est la chasse
directe par la population locale, en raison de lutter contre l'ennemi des
poussins domestiques. Pour ce cas, la population piège l'espèce
à l'aide d'un filet ou d'un piège à bâton. Pendant
notre observation sur terrain, la plupart des individus capturés a
été le sexe masculin. Ces sont des mâles qui chassent afin
de nourrir leurs partenaires et leurs progénitures. Ensuite, la
deuxième menace possible mais n'a été pas
été vu pendant cette étude, c'est la chasse par les autres
espèces de rapaces (Milvus sp) envers leurs poussins et
également par le serpent particulièrement les Boa (Sanzinia
madagascariensis) envers leurs oeufs voire leur poussin aussi. Certes,
cette espèce fréquente les milieux forestiers de l'aire
protégée entant qu'une espèce forestière et capable
de défendre leur territoire de nidification, elle est alors
généralement sécurisée face à la pression
humaine et des autres prédateurs terrestres.
Pour les Faucons à ventre rayé, la population
riveraine ne chasse pas cette espèce car elle ne porte pas aucune menace
envers ses domestiques mais le problème c'est au niveau de l'arbre de
nidification et de perchoir dans le milieu ouvert. La population exploite les
grands arbres dans le milieu ouvert pour accomplir leur besoin quotidien
(Charbons, bois de construction et des matériels à la maison,
pirogues, etc). Ceci appauvrisse donc l'arbre
52
disponible pour la nidification or ces espèces choisit
fréquemment l'habitat ouvert pour la reproduction. En plus, la
période de novembre et décembre est une période des
récoltes des mangues et des tamarins, qui entraine donc la perturbation
des poussins sur leur nid. Les individus dans le milieu forestier
protégé n'ont aucune menace liée à l'action
humaine, mais la chasses des autres prédateurs comme le cas de
l'Epervier de Madagascar est considérables.
D'autre part, les enfants chasseur des oiseaux à l'aide
d'une flèche ne choisissent pas ces deux espèces comme cible de
chasse à cause du tabou et de la croyance.
53
II.5. ETHOLOGIE
Pendant cette étude, les comportements pendant la
période de reproduction à partir de l'incubation jusqu'à
l'indépendance des poussins ont été enregistré tels
que les comportements de chasse, la livraison des proies, l'élevage des
poussins, la défense territoriale et quelques autres comportements.
II.5.1. Comportement de chasse
Le comportement de chasse pour les deux espèces est
quasiment semblable. Les individus se cachent et restent en silence sous des
feuilles d'arbre de perchoir ou d'autres abris pour localiser les cibles avant
de se lancer, puis ils les attaquent rapidement par un vol rapide (vol
piqué tout en fermant les ailes) puis ils ouvrent leurs ailes et
immobilisent les proies avec les serres quand il s'agit des grosses proies. Ces
dernières sont généralement capturées à
l'aide de leurs serres soit sur la terre soit sur les feuilles soit sur les
branches soit en vol (pour certains insectes tels les papillons) puis
transporté vers les grands arbres de perchoir pour les tuer et les
consommées mais ils peuvent tuer les proies directement sur la terre.
Ces dernières seront mangées directement par les chasseurs ou
transportées vers les nids pour les cas de femelles afin de nourrir les
poussins ou livré aux partenaires pour le cas de mâles.
Pour les Eperviers de Madagascar, les proies comme les oiseaux
sauvages et les poussins domestiques sont capturées sur la terre ou sur
des branches de canopée à basse altitude. Avant l'attaque, ils se
cachent et restent en silence sur un abri d'un arbre généralement
avec une hauteur plus de 8,5m puis lance une attaque rapide par un vol
piqué jusqu'au cible pour la capture. Puis ils tuent directement les
proies sur le substrat soit ils les transportent d'abord sur le perchoir avant
le tuer. Cette espèce est généralement vue seul soit en
couple sur pendant la chasse. Il est souvent posé sur la canopée
de la forêt, mais il lui arrive de planer au-dessus des arbres.
Les Faucons à ventre rayé peuvent rester
immobiles sur leurs perchoirs pendant 3h15mn pour repérer les proies.
Dès qu'ils trouvent leurs cibles, ils font d'un vol rapide pour les
capturer comme l'Epervier de Madagascar aussi. Cette espèce
préfère beaucoup le milieu forestier ouvert ou
dégradé pour chasser surtout les reptiles (les lézards et
caméléons) tandis que les Epervier de Madagascar restent souvent
dans le milieu forestier protégé afin de se satisfaire aux petits
oiseaux forestiers.
54
II.5.2. Livraison des proies entre le couple et Elevage des
poussins ? Livraison des proies entre le couple en reproduction
La livraison des proies se fait en dehors de l'arbre de
nidification pour les deux espèces. Le males chasse et transfert la
proie au femelle sur un arbre de perchage. La livraison se fait du mâle
au femelle, on ne trouvait pas le cas contraire. En effet, pour informer son
partenaire, le mâle du couple crie dès qu'il soit là avec
les provisions et la femelle rejoint directement son partenaire, avec un cri
aussi comme réponse pour les femelles faucons à ventre
rayé. La livraison se fait soit laisser devant le femelle (du pied
à bec ou de pied à pied) soit du bec à bec. Cette
activité a été observé à partir de
l'incubation jusqu'à l'indépendance des poussins. En dehors de
cette période, le couple ne s'entre occupe plus.
? Elevage des poussins
L'élevage des poussins pour les deux espèces est
généralement assuré par la femelle tels la couvaison, le
nourrissage et le gardiennage des poussins tandis que le mâle assure la
recherche des proies et la défense du territoire. Mais, en cas de la
défaillance de chasse de son partenaire, la femelle remplit la lacune et
assure aussi la défense territoriale. Pendant, la livraison des proies
aux poussins, le transfert se fait du bec à bec jusqu'à la
période que les poussins soient capable de se débrouiller. Et
pour les nids 1 et nid 3 comme nos nids de référence pour les
deux espèces, une compétition des poussins pendant la prise de
nourriture a été observée. L'un des poussins qui semble
être plus vigoureux, accapare les proies en premier et les restes ne
mangent que ceux qui restent. Des agressions entre eux ne sont pas
observées à part de lancement de cris défensifs pour que
l'autre n'approche pas. Le transfert de proie pour la plupart du temps entre
les adultes et les juvéniles s'effectue dans le nid. Les
juvéniles émettent des cris répétitifs quand ils
ont faim. Lors de ses interventions, le parent nourrisseur donne des
becquées aux poussins l'un après l'autre. Et l'adulte charge le
partage assure le la dissection des proies avant le nourrissage. Pour
l'Epervier de Madagascar si la proie s'agit d'un oiseau, avant la dissection,
le parent charge d'abord la déplumâtes. La quantité des
proies données aux poussins diminue progressivement à partir de
30ème jours pour l'Epervier de Madagascar et de
25ème jours pour les Faucons à ventre rayé des
poussins jusqu'à l'indépendance total des poussins. A part de
cette période, les poussins commencent capable de quitter les nids pour
apprendre à voler et à chasser. La diminution de cette
quantité continue jusqu'à l'absence total du nourrissage aux 61
à 66 jours de l'âge des poussins pour l'Epervier
55
de Madagascar et aux 53 à 55 jours des poussins pour le
Faucons à ventre rayé. Ceci entraine la dispersion totale des
juvéniles
II.5.3. Relation interspécifique et défense
territoriale
La relation interspécifique dans le sens positif est
moins évidente dans cette étude. Le cas de Faucon avec les
Gymnogène de Madagascar Polybroides radiatus (Fihiaka) est l'un
exemple, l'adulte du Faucon fait le chevauchement de territoire de cette
dernière. Les faucons osent à chasser dans le territoire des
Gymnogènes. Ceci entraine cette dernière à lancer des
attaques jusqu'à ce que l'intrus ait quitte son territoire. Ce cas n'a
été trouvé pour le cas de l'Epervier de Madagascar.
Pour les deux espèces étudiées, le
comportement de la défense territoriale est presque semblable, s'est
manifesté d'une manière plus agressive et par trois gestes bien
distinctes. La première geste correspond à une émission de
cris d'alertes en cas de présence d'intrus juste pour le faire fuir, la
deuxième correspond l'intrus arrive tous près du nid dans un
rayon environ moyenne: 132 m pour l'Epervier de Madagascar et 118m du nid pour
Faucons à ventre rayé, l'un des adultes occupe le pourchasse hors
du territoire et la troisième dernière correspond l'intrus est
entièrement pénétrer dans le territoire de nidification,
l' adule lance l'attaque avec un cris aigus jusqu'à ce que l'intrus
quitte définitivement leur territoire. Accipiter francesii,
Milvus migrans, Couas (Coua cristata, Coua cursor, Coua
gigas, Coua sp), Falco Newtoni, Aviceda Madagascariensis, Buteo
Brachypyerus sont des espèces listées comme des intruses.
En général, la sympatrie pour les deux
espèces étudiées est liée aux conditions de
territoire de la reproduction et à la disponibilité des
régimes alimentaires.
II.5.4. Vol et cris
? Type des vols
Quatre types des vols ont été trouvés pour
les deux espèces étudiée : Vol plané :
Pratiqué pour la localisation des proies
Vol piqué : pratiqué pendant la chasse, avant la
capture, l'oiseau ferme les ailes afin d'avoir une vitesse plus
élevée.
Vol stationnaire pour les faucons : pratiqué pour
piéger les cibles, l'oiseau reste immobile à l'air libre avec
ailes et queue écarté.
56
Vol battu : pratiqué pendant l'attaque des intrus et
pour le déplacement à longue distance. Ceci est souvent
intercalé par un autre type de vol cité ci-dessus.
? Type de cris
L'Epervier de Madagascar émet habituellement des cris
« whik », « whisk » ou « eeesk » court, aigu et
haut-perché, répété et très
mécanique. Nous pouvons aussi entendre des notes rapides « kee
» pendant le transfert de nourriture du mâle à la femelle
près du nid. Les cris sont souvent entendus tôt le matin.
Pour les faucons à ventre rayé, l'espèce
a un cri perçant, staccato, bavardant et un appel aigu et criant mais
est habituellement silencieux surtout en dehors de la saison de reproduction.
Les individus adultes de cette espèce ont émis trois cris pour
cette étude. L'un est pratiqué
pendant la livraison des proies, la femelle émit un cri
« Peetss ! Peetss ! Peetss ! ». Ensuite
la
deuxième, en cas de présence d'ennemi, la femelle signale son
partenaire avec un cri « tsu ! tsu....tsuuu ! ». Pour la
défense du territoire, le couple prévient les intrus avec le cri
« ket !
ket ! ket ! » suivi « kiwi ! kiwi ! kiwi ! » s'il
fait un attaque direct.
PARTIE III : DISCUSSION
59
III.1. DISTRIBUTION ET TAILLE DES POPULATIONS
L'abondance et la distribution des animaux dans un habitat ne
sont pas constantes vue la situation présente dans son environnement
(Cody, 1995). Ce changement peut être mené dans le sens positif si
les facteurs bioécologiques sont favorables, dans ce cas la taille et la
distribution de la population augmente. Mais dans le cas contraire, la taille
de la population diminue c'est-à-dire le degré de menace devient
élevé voire l'extinction si aucune mesure n'est prise. Ces
facteurs peuvent être des conditions suivantes : disponibilité des
régimes alimentaires, site de reproduction, sécurité et
d'autres facteurs abiotiques (Rene de Roland, 2010).
? Epervier de Madagascar
L'épervier de Madagascar est une espèce
endémique de la grande île et classée parmi les
espèces quasi-menacées selon l'UICN en 2009. Cette espèce
est distribuée dans la forêt pluviale de l'Est ainsi que dans les
broussailles et la forêt épineuse sèche de l'Ouest et du
sud-ouest de l'île ; rarement vue dans la partie sud de l'île (Rene
de Roland, 2010). C'est une espèce forestière peu observée
dans le milieu dégradé ou dans les habitats secondaires. Mais
l'espèce est probablement plus commune que prévu car c'est un
rapace discret qui reste souvent posé en haut de la canopée des
arbres. La taille de la population est généralement encore faible
en comparant à l'effectif des autres espèces d'oiseaux
présents dans ces sites. Mais l'augmentation de la taille de la
population est considérable si aucune menace n'est survenue vue le
succès de la reproduction comme montré par cette présente
étude.
? Faucon à ventre rayé
Les faucons à ventre rayé sont la seule
espèce endémique des faucons à Madagascar, occupant
généralement une distribution un peu plus élevé,
particulièrement dans les côtes de l'ile. C'est une espèce
très discrète qui peut être observée à partir
du niveau de la mer jusqu'à une altitude de 2000 m (Langrand, 1990). Ils
se trouvent dans les forêts denses humides de l'Est et Nord Est, dans les
forêts sèche de l'Ouest et du Sud-Ouest de l'île, on peut
les trouver aussi dans les habitats ouverts et dégradés sur ces
écorégions, mais quasiment rare, voire totalement absente dans le
haut plateau et dans la partie septentrionale (Langrand, 1995 ;
Robenimangarison, 1999). Malgré, la taille de la population est
quasiment toujours faible (Ferguson-Lees & Christie, 2001).
60
Le faucon à ventre rayé est une espèce
très rare et difficile à localiser ou à observer car cette
espèce reste souvent silencieuse vu son comportement très
discret, même pendant la période de la reproduction. Cependant,
elle est très facile à identifier par rapport au faucon de Newton
par sa taille plus grande, son iris de couleur jaune, sa grande tête et
les stries au niveau de la partie ventrale. Raveloson L, 2017, pendant son
étude bioécologique sur cette espèce dans le complexe
Tsimembo Manambolomaty, centre ouest Madagascar, n'a trouvé qu'un seul
couple et Rene de Roland (comm. Pers), n'a rencontré que deux individus
adultes et deux juvéniles seulement entre Mangily-Ifaty et le Sept lacs
d'Ifanoto (in Raveloson, 2017) et trois couples ont été
étudiés pendant la période de reproduction de 1997 en 1999
(Rene de Roland et al., 2005).
Le résultat de cette présente étude en
comptant les juvéniles, donne des effectifs 18 pour l'Epervier de
Madagascar et 22 pour le Faucon à ventre rayé, ceci nous informe
que la taille de cette population dans les forêts sèches de
Mangily augmente et presque élevé vue le succès de la
reproduction et la faiblesse des pressions et des menaces dans leur habitat. Ce
résultat donc va changer le statut de cette espèce si les
conditions bioécologiques sont toujours favorables. Malgré, cet
effectif est toujours faible par rapport à la densité de la
plupart des autres rapaces dans ce site (Reniala, 2019 comm. pers).
III.2. RECHERCHE DE NIDS
Après avoir inventorié les individus, la
recherche de nids a été faite et ne semble pas être
difficile après l'identification des individus couple. La combinaison
des méthodes telles que la localisation des individus, l'écoute
des cris et le suivi des individus nous donnent la chance pour cette recherche.
Tous cela a été effectué dans deux sites :
protégé (=site 1) et non protégée =site2)
grâce à la collaboration avec les villageois, les autres
collègues et les guides locaux. Pour cette étude, nous avons
trouvé 2 nids pour l'Epervier de Madagascar, tout a été
trouvé dans le site 1(nid 1et nid2) tandis que pour le Faucon à
ventre rayé, nous en avons trouvé 03, un dans le site 1 et deux
dans le site 2. L'un nid de ce dernier a été déjà
abandonné.
Ceux-ci expliquent que l'Epervier de Madagascar se reproduit
seulement dans le milieu forestier mais non dans les habitats ouvert ou
dégradé. Ceci a été prouvé par John
(1899), lors de son étude bioécologique. Cette
espèce peut être considérée comme une espèce
bioindicatrice d'un habitat. Le nombre des nids trouvés semble moins
nombreux par rapport au nombre des individus trouvés et à la
surface de l'habitat. Rene de Roland (2000) n'a trouvé que 5 nids
lors
61
de son étude dans les presqu'îles de Masoala.
Alors, l'insuffisance des nids est l'une raison qui conditionne l'insuffisance
d'effectif de cette espèce à Madagascar.
Pour le Faucon à ventre rayé, le choix du site
de reproduction n'est pas obligatoire dans le milieu forestier
c'est-à-dire certains individus utilisent l'habitat ouvert et les autres
préfèrent l'habitat forestier. Vue l'effectif des nids
trouvé par site, cette espèce préfère l'habitat
ouvert ou dégradé pour la reproduction que le milieu forestier.
Mais le type d'arbre du nid est tellement considérable (Voir chap
écologie). Pour cette étude, le nombre des nids rencontré
parait faible. Un nid en 1997 et deux nids en 1998 ont été
trouvés par (Robenarimangason ,1999) et il a signalé que cette
espèce est difficile à localiser car elle ne crie pas beaucoup.
Cette particularité a été considérée comme
un problème pendant la localisation du nid (Raveloson, 2017).
René de Roland et al, (2000) n'a trouvé que deux nids
même cas au Raveloson, 2017.
Notre méthode d'enquête envers la population
locale nous a permis de signaler que les enquêtés n'ont pas bien
connus les espèces étudiées, ils confondent par exemple
les Faucons à ventre rayé avec les Faucons Concolore et Faucons
de Newton ; L'Epervier de Madagascar avec l'Epervier de France et l'Autour de
Henst. Raveloson 2017, a signalé que les villageois au Nord-ouest de
Madagascar confondent le Faucon à ventre rayé avec le Faucons de
Newton et avec l'Epervier de France. Cela signifie que cette méthode ne
nous a pas efficacement aidés pour cette recherche. Cette confusion de
ces rapaces a déjà été relatée comme un
sujet ; Langrand (1990) et Morris & Hawkins (1998). D'après ces
auteurs, la morphologie quasi-semblable entre les espèces
étudiées et ceux des autres rapaces reflètent cette
circonstance.
Les deux espèces étudiées sont
discrètes, quasiment rare et difficile à trouver Morris &
Hawkins (1998). Ces caractères sont donc considérables l'un
facteur de l'insuffisance des nids trouvés pour cette étude ou
certains couples pourraient se reproduisent dans un habitat plus loin de notre
zone d'étude. Tout cela signifie que la méthode adoptée
pour cette recherche parait moins efficace et beaucoup de raisons peuvent
être l'origine de l'insuffisance des nids trouvés.
III.3. METHODE DE CAPTURE
Pour cette présente étude, nous avons
adopté pendant la capture des espèces la méthode de
Bal-Chatri et le Piège à bâton, installé tous
près des nids pour cibler les couples étudiés (Strena
& Boileau., 2011). En effet, un couple par espèce
étudiée a été capturé. L'utilisation de ces
mêmes méthodes a permis Robenarimangason (1999) à capturer
trois individus des faucons à ventre rayé à Masoala ;
Andriamalala (2005) a capturé 2 couple des Milans au complexe
62
Tsimembo Manambolomaty ; Rafarantsoa F. (2008) a
capturé 45 individus des Milans à Beheloka (Berenty) entre 2005
à 2006 et Raveloson (2017) a capturé deux individus des faucons
à ventre rayé au complexe Tsimembo Manambolomaty. Tout
ceux-là signifie que cette technique de capture parait convenable
à la capture des rapaces. Mais Strena & Boileau., (2011) ont
suggéré que l'utilisation de l'épuisette et un filet
rabattant automatiquement soit également efficace pour la capture des
rapaces.
III.4. MORPHOLOGIE ET MORPHOMETRIE DES ADULTES
John. (1899), a trouvé que l'Epervier de
Madagascar a une longueur entre 29 à 42 cm, envergure : 50 à 69
cm et Poids : 285 à 348 g, Rene de Roland (2000), a trouvé que la
femelle de cette espèce possède une longueur 38,3 et un poids
348g. Le résultat de cette présente étude montre que le
mâle possède une longueur 33cm, pèse 298g tandis que la
femelle mesure 38,42 et pèse 344g. Ceux-ci expliquent que la taille des
individus est incluse dans l'intervalle de John. (1899), et le
dimorphisme sexuel entre eux semble évident, le mâle parait plus
petit que la femelle. La morphologie de la femelle ressemble avec le mâle
Accipiter henstii non seulement au niveau de taille mais aussi
à la couleur. Ceci rend la population locale d'avoir une
difficulté pour la distinction. Pourtant, la longueur du doigt central
(troisième doigt) de l'Epervier de Madagascar est très
remarquable par rapport aux autres doigts. Cette particularité joue un
rôle crucial sur le choix de sa proie. Sa queue brune est striée
de cinq à six bandes 12 claires. Les sous caudaux sont de couleur
claire. Cette caractéristique diffère de l'épervier de
Madagascar femelle et du mâle d'autour de Henst.
En général les résultats morphologique et
morphométrique de cette espèce n'ont aucune remarque signifiant
comme au niveau de la couleur et du poids en comparant aux informations
déjà publiées.
Pour le Faucon à ventre rayé, elle est une
espèce morphologiquement semblable pour les deux sexes surtout au niveau
de couleur et de la longueur selon Langrand, (1990), Morris & Hawkins
(1998). C'est pourquoi certains auteurs qualifient que cette espèce ne
présente pas un dimorphisme sexuel. Ceci est valable pour l'observation
à distance mais Raveloson, (2017) défend énergiquement que
les deux sexes diffèrent surtout au niveau de la taille et certaines
formes des plumes au niveau des ailes et de la queue. Cet auteur a bien
confirmé que la femelle est légèrement plus grande que son
conjoint, c'est qui conforme aux résultats de Robenarimangason (1999) et
Rene De Roland et al, 2005. Cette information contre la vision de
63
Olsen (1990) que la plupart des oiseaux de proie, les
mâles ont une taille plus grande que la femelle. Par rapport à la
taille, on peut dire que la taille de cette espèce diminue pour la
présente étude (146g mâle contre 178 g femelle) en
comparant aux informations publiées par Robenarimangason, 1999 (175g
mâle contre 222g femelle).
III.5. LA REPRODUCTION
III.5.1. Période de formation du couple,
nidification et ponte
La période de formation de couple chez les oiseaux de
proies est mal connue car il se passe rapidement (Village, 1990). Les
résultats de cette présente étude ne donnent pas
l'information concernant cette sous chapitre lors du retard de la
période d'étude sur terrain. Mais notre recherche bibliographique
nous permet de donner les informations suivantes.
La formation du couple de l'Epervier de Madagascar commence
vers la première semaine du mois de septembre et le repérage du
lieu de nidification s'effectue dès l'appariement jusqu'à la
deuxième semaine de mois d'octobre Rene de Roland (2000). John (1899) a
bien signalé que les deux adultes participent à la construction
du nid, mais le mâle est le plus actif. Par contre, Rene de Roland (2000)
contre cette affirmation, il a dit que la femelle ne participe pas à la
nidification, seulement le mâle qui charge cette activité. La
ponte a lieu dès la première semaine d'octobre. Le nombre d'oeufs
coloré uniformément blancs ou avec des marques plus sombres
variables varie d'un à quatre par couvée, Rene de Roland
(2010).
Pour les Faucons à ventre rayé, Rene de Roland
(2010), Raveloson, (2017) ont réussi pendant leur étude sur cette
espèce à Masoala et à Tsimemo Manambolomaty que la
formation du couple se situe entre les mois d'Août et de Septembre. Cette
précision a été faite grâce à l'écoute
de cris particulier Langrand (1999), Robenarimangason (1999) et Rene de Roland
(2010). La ponte commence à partir de mois d'octobre et se termine
à la deuxième semaine de mois de Novembre (Coolebrook-Robjent,
(1973), Robenarimangason, (1999). Cela signifie que la période de ponte
s'effectue entre la deuxième semaine de mois de septembre jusqu'à
la deuxième semaine de mois de Novembre. Par rapport aux grands rapaces,
le nombre d'oeufs par nid est un peu élevé (Hartley &
Hustler, 1993 ; Hartley, 1995), le nombre d'oeufs pondu par nid varie entre 2
à 4, (n=5 oeufs pour deux nids pour cette présente étude).
Ceci est proportionnel à l'information publiée par Rene de Roland
(2010). Selon ce dernier auteur, les oeufs ont une dimension de 38,5 X 30,5 mm
et le poids moyen est de 19,7g tandis que pour cette présente
étude, les oeufs coloré beige au fond blanc pèsent entre
23,80 à 25g. Il mesure entre 38 à 38,4mm
64
pour sa longueur contre 32 et 32,4 mm pour sa largeur. Tous
cela veut dire que les oeufs de cette espèce à Mangily sont un
peu plus grands que ceux de Masoala mais l'écart est entièrement
faible.
En conclusion, la période de reproduction de ces deux
espèces est quasiment pareille car cette période correspond au
bon climat (température), favorable à la reproduction et à
l'abondance de la ressource alimentaire. Ensuite, la différence de
nombre des oeufs par nids peut être déterminée par les
facteurs bioécologiques tels que la disponibilité des ressources
alimentaire et l'âge du couple.
III.5.2. Incubation, éclosion et couvaison
La plupart de rapaces, l'incubation débute avant que
leur ponte soit complète (Newton, 1979) mais ceci n'a pas
été vue pour le cas des espèces étudiées.
L'Epervier de Madagascar débute cette activité après la
dernière ponte. Tandis que pour le Faucon ventre rayé, cette
étude a raté l'identification le début de l'incubation
mais Thorstrom comm pers, Raveloson (2017) ont bien signalé que cette
activité débute juste avant que le pénultième oeuf
soit pondu c'est-à-dire au moment de l'avant dernier oeuf. Ces
mêmes auteurs ont bien précisé que cette activité
commence entre la deuxième semaine de mois d'octobre et se termine
à la première semaine de mois de décembre, qui dure
environ 30 à 32 jours qui correspond bien aux résultats de cette
présente étude.
L'activité incubatrice dure environ 35 jours, une
période identique à celle de l'espèce africaine
Accipiter rufiventris (Grobler, 1980) est assurée seulement par
la femelle pour l'Epervier de Madagascar même cas pour la couvaison. Ce
cas est trouvé pour les espèces de rapaces ayant un dimorphisme
sexuel marqué comme l'Autour de Henst Accipiter henstii, la
femelle avec une plus grande surface de plaque incubatrice assure la
majorité de l'incubation (79,5%) (Rene de Roland, 2000). La femelle ne
quitte pas leur nid qu'au moment où le mâle apporte la proie et le
transfert de proies se réalise plus souvent au niveau d'une branche
d'arbre autre que l'arbre de nidification. Mais à partir de la
deuxième semaine pour cette présente étude, nous avons
trouvé que la femelle assure aussi la sécurité
territoriale et parfois effectue la chasse pour se nourrir et nourrir ses
poussins surtout en cas de défaillance de chasse de son conjoint. Mais
en général, la femelle reste sur le nid pendant plus de 80% de
temps de notre observation tandis que le mâle assure la
sécurité du territoire et la recherche des proies. A partir de 36
à 38éme jours de l'âge, les poussins commence
à quitter leurs nids pour apprendre à voler et à
65
chasser mais encore ils retournent au nid chaque soir. A ce
moment, le couple continue à ravitailler ses poussins en matière
alimentaire mais la quantité livrée diminue puis voire absente
à partir de 60é jour. Cette dernière entraine
la dispersion des poussins à l'âge de 65 à 66
é jours. Pour cette présente étude, le taux de
réussite de la reproduction est 100% alors que Rene De Roland, (2000) a
trouvé aussi 99%. Ceci peut entrainer l'augmentation de taille de la
population si aucune menace n'est survenue.
Pour le Faucon à ventre rayé, la durée de
l'incubation pour cette présente étude est mal
précisée mais Rene de Roland L. A (2010), Raveloson (2017) ont
dit que la durée de cette activité varie entre 29 à
32jours. Cette durée est similaire à celle de la
crécerelle des Seychelles Falco araea (Watson, 1981) et de la
crécerelle d'Amérique Falco sparverius (Balgooyen, 1976)
qui ont toutes de tailles proches au faucon à ventre rayé. Ceci
conforme aux informations vécu pendant notre terrain vue la
période d'éclosion de poussins par rapport aux informations
concernant la période de ponte. La différence de période
d'incubation d'une espèce est en fonction de taille de l'oeuf et leur
masse, principalement en raison de perte d'eau (Newton, 1979 ; Gill, 1989 et
Burnham, 1997). Tout cela pourrait donc justifier cette légère
variation de période d'incubation sur cette espèce et même
cas pour l'espèce précédente. L'activité
incubatrice est assurée par la femelle tandis que le mâle assure
la recherche des proies même cas pour l'Epervier de Madagascar. La
femelle commence à quitter le nid à la deuxième semaine de
l'éclosion pour aider le mâle pour la chasse Rene de Roland
(2010). Pour cette présente étude, le taux de réussite
pour l'éclosion est 100% (n=5oeufs donne 5poussins). Malgré, l'un
de ces poussins a été tué lors de la pluie violente du
mois 16 décembre 2019 qui entraine le taux de réussite pour le
succès de la reproduction comme 80%. Raveloson, (2017) avait un taux de
réussite lors de son étude à Masoala 83,3 % pour
l'éclosion et 0% pour les poussins indépendants. Il a
signalé que cette défaillance est due un facteur climatique
défavorable et une perturbation de l'habitat de nidification par
l'action anthropique et d'autres rapaces. Robenarimangason (1999) a
mentionné que l'insuccès de cette espèce à Masoala
est dérivé de l'abandon du nid avant l'éclosion lors de la
perturbation du Rollier malgache Eurystomus glaucurus. Ceci pourrait
donc l'un facteur qui entraine certain couple des faucons à installer
leur nid dans un habitat ouvert pour fuir les prédateurs sympatriques
dans leur habitat.
66
III.6. ECOLOGIE
III.6.1. Habitats de nidification et caractéristique
de nids
Les nids de ces deux rapaces étudiés sont
localisés dans deux sites différents : trois nids ont
été trouvés à l'intérieur de l'aire
protégée c'est-à-dire dans un habitat forestier dont 2
nids pour l'Epervier de Madagascar et un nid pour les faucons à ventre
rayé et les restes localisés dans le site 2 ou l'habitat ouvert
sont seulement pour les faucons.
Les deux nids de l'Epervier de Madagascar sont trouvés
dans le site protégée cela signifie que cette espèce se
reproduisent exclusivement dans un habitat forestier. Ces nids ont
été placé sur une branche portant une hauteur par rapport
au sol entre de 13 m à 15m et sur un arbre ayant une Dhp 132cm (nid 1sur
un baobab fony) et 44cm (nid 2 sur un Commifora lamii). Alors que John
G. K. (1899) et Rene de Roland (2000) ont mentionné que les nids de
cette espèce sont placés sur un arbre possédant une
hauteur de 20 à 32m par rapport au sol lors de leur étude dans la
côte Nord-Est de Madagascar. Ceci nous permet de dire que le choix de
l'arbre de nidification dépend la structure présente dans un
habitat, dans notre zone d'étude aucun arbre ne dépasse pas de
23m de hauteur, pratiquement, il est tellement rare de trouver un pied d'arbre
ayant une hauteur plus de 18m. Les nids ayant une ouverture en haut sont
placé dans la partie orientale, est en raison pour permettre
l'accès au rayon solaire pour la matinée mais les feuilles vertes
de la plante qui englobent ces nids servent comme abri à éviter
la température élevée, la pluie et les prédateurs.
Ces nids ont un moyen diamètre interne 31,25 cm et externe 37cm, une
profondeur interne 12 et externe 21,5cm. La taille de ces nids est plus petite
que ceux des nids de l'Aigle pêcheur de Madagascar lors des
résultats d'étude de Rasolonjatovo, (2017), cette dernière
espèce ayant un nid mesure 70,75 x 53 cm, la moyenne de profondeur
externe et interne du nid mesure respectivement 55,5 cm et 13,75 cm.
Pour les Faucons à ventre rayé, un nid a
été trouvé dans la zone protégée et deux
nids dans la zone non protégée du site 2 et l'un de ces derniers
a été déjà abandonné. Meyburg & Langrand
(1985), Robenarimangason, (1999) ont trouvé des nichés de faucons
dans des habitats assez loin de la forêt intacte, la plupart d'entre eux
sont tous près d'un village ou d'une agriculture. Raveloson, (2017) a
trouvé deux nids de cette espèce lors de son étude
à Tsimembo Manambolomaty dont un dans la forêt intact et l'autre
dans la zone dégradé ou milieu ouvert.
67
Bref, les Faucons à ventre rayé semble comme
d'autre d'espèces de rapace tels les Faucons de Newton, les Faucons
pèlerin, les Faucons de Dickinson, l'habitat de nidification n'est pas
obligatoire milieu forestier, ils peuvent se reproduire dans l'habitat ouvert
ou dégradé.
Pour cette présente étude, les nids ont
été placés sur des pieds d'arbre d'espèces
différentes (Givotia madagascariensis, Mangifera indica, Tamaridus
indica) ayant une hauteur moyenne de 14m et sur des branches portant une
hauteur moyenne par rapport au sol de 9,5m. Ces nids portant une longueur 35,75
cm et une largeur 33cm, une profondeur interne et externe respectivement 10 et
17cm. René de Roland et al, (1999) ont trouvé que les nids de
cette espèce se sont trouvés sur des arbres ayant une hauteur de
25m de hauteur et les nids sont placés sur des branches en forme de
fourche portant une altitude par rapport au sol 19m. Robenarimangason (1999) a
mesuré deux nids de dimension moyenne 60cm x 68,5cm (largeur et
longueur) sur un arbre de 20,4 à 32 ,5 de haut. Alors que Raveloson,
(2017) a trouvé que les nids ont une dimension moyenne 32,5 cm de
largeur et 54 cm de longueur sur un arbre de 12 à 15m. En effet, la
hauteur des arbres de nidification et la caractéristique des nids
dépends de l'endroit choisi, leur dynamique et leur environnement.
III.6.2. Type des proies
La quantité et la qualité des proies semblent
être variables pendant la période de reproduction chez les rapaces
(Rene de Roland, 2010). Ces espèces s'adaptent facilement à leur
alimentation et prennent tous les animaux qu'ils peuvent tuer (Village, 1990).
John (1899) et Rene de Roland, (2010) ont mentionné que les Eperviers de
Madagascar se nourrissent principalement des petits oiseaux forestiers qui
peuplent la canopée, quasi 90% du régime mais ils chassent aussi
des reptiles tels les lézards et des insectes. Pour la présente
étude, nous avons trouvé différente type des proies, en
plus des oiseaux forestiers, cette espèce chasse des poussins domestique
aux alentours du parc, des caméléons, des lézards, des
Gecko et des insectes. Toutes ces proies sont capturées au sol ou sur
d'autres substrats à l'aide de leurs serres sauf les derniers, qui sont
capturés souvent en vol. Mais l'alimentation se base en oiseaux, plus de
80%.
Pour les Faucons à ventre rayé, leur
alimentation se base en deux types différents Reptiles et insectes (Rene
de Roland (1999), Robenarimangason (1999) et Raveloson, (2017). Cette
espèce se nourrisse des caméléons, des lézards, des
geckos pour les reptiles et les insectes tels que les papillons et des
criquets. Le type alimentaire chassé et livré pendant la
reproduction est différent, par exemple pendant l'incubation, le male
chasse des reptiles tandis que pendant
68
l'élevage de poussins, les adultes chassent des
insectes particulièrement jusqu'à la troisième semaine de
l'âge des poussins puis en dehors de cette période, les adultes
apportent des nourritures à grandes tailles, ces sont des reptiles
particulièrement des caméléons genre Furcifer.
Plusieurs auteurs ont aussi mentionné que la majeure partie des
proies chassées par le faucon à ventre rayé est
composée par des caméléons (Rand, 1936 ;
Colebrook-Robjent, 1973 ; Thorstrom, 1999). En général,
l'alimentation n'est pas considérée comme une menace pour les
espèces étudiées dans ces sites.
III.7. COMPORTEMENT
III.7.1. Comportement pendant la reproduction
Pour les rapaces, la plupart des femelles ont la taille plus
grande que les mâles occupent plusieurs activités pendant la
reproduction (Magali, 2009). Pour les deux espèces
étudiées, seules les femelles occupent l'activité
incubatrice et la couvaison. Celle-ci est une particularité des oiseaux
de proie ayant un dimorphisme sexuel important Simmons (1986), Robenarimangason
(1999). Selon Raveloson, (2017) lors de son étude au ComplexeTsimembo
Manambolomaty, le Faucon femelle assure la plupart de temps pour l'incubation
et l'élevage des poussins. Ce même auteur a bien mentionné
que le mâle assure la tâche pour la recherche des proies et aussi
la défense territoriale.
III.7.2. Relation interspécifique
La relation interspécifique a été
identifiée pour la compétition alimentaire et la défense
territoriale. Les Eperviers de Madagascar font une compétions
alimentaire avec les autres rapaces aviphages comme le Milan, l'Autour de
Hensti, etc. Dans ce cas, l'Epervier lance l'attaque envers les adversaires
s'ils dominent, par contre ils restent en silence.
L'agressivité de ces deux espèces envers les
individus de la même espèce s'observe seulement dans le territoire
de nidification. Plus un individu s'éloigne de son territoire, plus la
tolérance envers les autres s'établissent. En revanche, ces
espèces sont très agressives envers les Buses de Madagascar,
Milan Milvus sp, Buteo brachypterus et Polyboroides
radiatus. Ces espèces semblent être à la fois les
principales compétitrices de l'espèce étudiée pour
la chasse, car leurs nids se localisent à 100m de celui de M.
migrans. En plus, P. radiatus est le principal prédateur
des poussins des oiseaux. Rene de Roland (2000) avait confirmé que de
P. radiatus est un facteur limitant pour le développement des
oiseaux, car cette espèce attaque les
69
différentes proies en allant d'une espèce de
petite taille ou même les oeufs dans le nid. Mais les facteurs naturels
biotiques ne sont pas considérés comme des principales menaces
pour les espèces étudiées.
70
CONCLUSION
La présente étude bioécologique de
l'Epervier de Madagascar et du Faucon à ventre rayé a
été faite dans la forêt sèche de Mangily, plus
précisément dans les aires protégées :
réserve Reniale et Parc Mosa, dans la partie Sud-ouest de Madagascar.
Aucune étude bioécologique n'a été encore
effectuée sur ces espèces dans ces sites. Les résultats
obtenus de cette présente étude serviront comme outil et moyen
indispensables pour la protection et la conservation de la biodiversité
particulièrement ces rapaces dans la forêt sèche
caducifoliée, de l'écorégion du Sud-ouest malgache. En
effet, elle nous a aidés à compléter les informations sur
les rapaces à Madagascar et de nous fournir les informations
indispensables sur la conservation.
L'étude sur terrain a été
réalisée entre le début mois de novembre 2019 au
début de Mars 2020 soit 4 mois. Cette période correspond à
la période de reproduction des espèces. Comme
méthodologie, la documentation, l'enquête et l'observation directe
en utilisant des appareils visuels et des mesures ont été
adoptées pendant les collectes de données.
Deux couples de chaque espèce ont été
étudiés pour cette étude, les nids des couples de
l'Epervier de Madagascar ont été localisés dans le centre
forestier de l'aire protégée tandis que pour les faucons à
ventre rayé, le nid d'un couple a été trouvé dans
le milieu forestier de l'aire protégée et les deux autres nids
ont été trouvés dans le milieu dégradé,
constitué par un nid actif et un nid abandonné. Tous ces nids
sont placés sur les pieds d'arbres de différentes espèces,
avec une hauteur importante par rapport au sol.
Les résultats recueillis durant cette étude nous
permettent d'avancer que ces deux espèces ont une même
période de reproduction, qui se réalise pendant
l'été, la ponte et l'incubation se localisent durant le mois de
Novembre et de Décembre et l'élevage des poussins se termine en
fin de Février et au début de mois de Mars. Chaque nid contient 3
et 4 oeufs pour l'Epervier de Madagascar et 2 et 3 oeufs pour les Faucons
à ventre rayé, les oeufs de l'Epervier de Madagascar semblent
plus grands que ceux du Faucon à ventre rayé. Pour les deux
espèces étudiées, les femelles assurent l'incubation, la
couvaison et le nourrissage des poussins tandis que les mâles s'occupent
la recherche des proies pour nourrir leurs partenaires et aussi leurs poussins.
Le transfert des proies entre le couple se réalise en dehors de l'arbre
de nidification. Le type de proies chassés et livrés par
l'Epervier de Madagascar est basé principalement en petits oiseaux, mais
cette espèce chasse aussi de reptiles et des insectes tandis que les
Faucons à ventre rayé ne chassent seulement que les deux derniers
types.
71
L'habitat de ces espèces est favorable à leur
besoin, en général sécurisé face aux perturbations
anthropiques, d'où les taux de productivités pour les deux
espèces atteignent 90%. Toutefois, les aléas climatiques (pluie
et vent violent) entrainent la mort d'un poussin des Faucons à ventre
rayé dans le milieu dégradé. L'exploitation des grands
arbres de nidifications dans le milieu dégradé par la population
locale est l'une de menace considérable pour cette espèce tandis
que la chasse pour les Eperviers de Madagascar en raison d'éviter la
menace pour les poussins domestiques.
D'ailleurs, il s'avère important de souligner que cette
étude est loin d'être terminée. La recherche sur ces
espèces nécessite encore des efforts pour sa conservation et la
gestion durable dans ces sites.
72
RECOMMANDATIONS
Les recommandations suivantes demeurent nécessaires afin
d'avoir une bonne connaissance sur ces espèces et une meilleure
conservation :
? La descente sur terrain doit commencer quelques semaines
avant la formation du couple
? Utilisation des matériels satisfaisants comme une
caméra de surveillance, pour enregistrer les évènements
dans les nids et aux alentours, installation de puce électronique aux
espèces afin de faciliter le suivi écologique,
? Sensibiliser la population locale sur l'importance de la
biodiversité et de l'aire protégée
73
BIBLIOGRAPHIE
1. Andriamalala Tolojanahary R. A., 2005
Contributions à l'étude de la biologie de reproduction
du Milan Noir à bec jaune Milvus aegyptius (Hartlaub, 1877)
dans le complexe des trois lacs (Ankerika, Befotaka, Soamalipo) du District
d'Antsalova, Mémoire de D.E.A en Biologie et Ecologie Animale,
Université Antananarivo, 69 p.
2. ANDRIANJAFY H. P.N., 2017, Etude de
biologie et écologie de Falco concolor (Temminck, 1825) (Aves,
Falconidae) dans sa zone d'hivernage, Madagascar, Mémoire de D.E.A en
Biologie et Ecologie Animale, Université Antananarivo, 37p
3. Andriantsaralaza S. 2016 Rapport annuel
ONG RENIALA 24p
4. Bothel Daurelis, 2018, Inventaire et
étude de la nidification du Rollier Terrestre à longue queue
Uratelornis chimaerae (Rothchild 1895) dans la réserve de
Reniala, Mangily, Sud-Ouest de Madagascar, mémoire de DEA,
48p
5. Del Hoyo, J., A. Elliott and J. Sargatal.
1994. Handbook of the Birds of the World. Vol. 2: New World Vultures
to Guine a fowl. Barcelona: Lynx Edicions. 638 pp.
6. Fenn, M.D. 2003. The spiny forest
ecoregion; pp. 1525-1529, in: S.M. Goodman and J.P. Benstead (ed.). The Natural
History of Madagascar. Chicago: The University of Chicago Press.
7. Fomenjanahary F. R, 2014, Inventaire de
la population de Rollier terrestre à longue queue Uratelornis
chimaerae et Monias Benschi, dans la réserve de Reniala,
Mangily Ifaty, Mémoire de Licence en Biodiversité et
environnement, Université de Toliara, 40p
8. Gardner, C.J. and L. Jasper 2009.
Possible new Mirza taxon from the Fiherenana River valley,
Atsimo-Andrefana Region. Lemur News 14: 46-49.
9. Gardner, C.J., D. Kidney and H. Thomas.
2009. First comprehensive avifaunal survey of PK32-Ranobe, a new
protected area in southwestern Madagascar. Phelsuma 17(1): 20-39.
10. Gardner C.J., Accipiter henstii
(Schlegel, 1873) (Falconiformes: Accipitridae): New distribution record
from southwest Madagascar, 18p
11. Gardner, C.J., C. De Ridder, B. De Ridder and L.
Jasper 2012. Birds of Ambondrolava mangrove complex, southwest
Madagascar. Check List 8(1): 1-7.
74
12. Goodman, S.M., M.J. Raherilalao, D. Rakotomalala,
D. Rakotondravony, A.P. Raselimanana, H.V. Razakarivony and V. Soarimalala.
2002. Inventaire des vertèbres du Parc National de
Tsimanampetsotsa (Toliara). Akon'ny Ala 28(1):1-36.
13. IUCN. 2016. The IUCN red list of threatened
species. Version 2018-2. <
www.iucnredlist.org>.
Downloaded on 10 Janvier 2019
14. Karpanty, S.M. 2006. Direct and indirect
impact of raptor predation on lemurs in Southeastern Madagascar. International
Journal of Primatology 27(1): 239-261.
15. Langrand, O. 1990. Guide to the Birds of
Madagascar. New Haven and London: Yale University Press. 364 pp.
16. Langrand, O and B.-U. Meyburg. 1984.
Birds of prey and owls in Madagascar: their distribution, status and
conservation; pp. 3-13, in: J.-M. Mendelsohn and C.W. Saps ford (ed).
Proceedings of the Second Symposium on African Predatory Birds, 22-26 August
1983. Durban: Natal Bird Club.
17. Langrand, O. and S.M. Goodman. 1997. Les
oiseaux; pp. 131-143, in: O. Langrand and S.M. Goodman (ed). Inventaire
Biologique, Forêt de Vohibasia et d'Isoky-Vohimena. Recherches pour le
Développement, Série Sciences Biologiques 12.
18. Langrand O., 1995, Guide des Oiseaux de
Madagascar, Delachaux et Niestlé, Lausanne, Paris
19. Mahafety, SC, 2013, Rapport annuel,
suivi environnemental Reniala, 19p
20. Megali, T. 2009, Dossier
pédagogique « Poussin kunken » Museum d'Histoire Naturelle
Fribourg. 45p
21. Mittermeier, R.A,2014, Hot spot revised,
CEMEX, Mexico, 390p
22. Moat, J. and P. Smith 2007. Atlas of the
Vegetation of Madagascar. Kew Publishing, Royal Botanical Gardens, Kew. 124
pp.
23. Newton, I., 1979, Population ecology of
raptors, Buteo books, Vermillons SD USA. 399p
24. Olson, D. and E. Dinerstein 1998. The
Global 200: A representation approach to conserving the world's most
biologically valuable ecoregions. Conservation Biology 12(3): 502-515.
25. Olsen, P. 1990, Raptor biology: Social
behavior, in bird of prey Ian Newton(ed), fact on file , INC, USA, Pp
108-123
26.
75
Rabarijerivelo. V. G., 2013. Inventaire et
suivi écologique du Mésite Monias benschi dans la
réserve Reniala à Mangily-Ifaty. Mémoire pour obtention du
D.E.A. Université de Toliara
27. Rafarantsoa F., 2008, Contribution
à l'étude bio-éco-éthologique, comparative des
couples hétéromorphes et homomorphes des espèces de milan
dans la réserve privée de Berenty, partie sud de Madagascar,
Mémoire de D.E.A. en Biodiversité et Environnement,
université de Toliara, 52 p
28. Raherilalao, M.J. and Goodman S.M. 2005.
Modèles d'endémisme des oiseaux forestiers des hautes terres de
Madagascar. Revue d'Ecologie (La Terre et la Vie) 60(4): 355-368.
29. Raherilalao J. M ; & Goodman S M, 2011,
Les familles et les sous familles des oiseaux endémiques de
Madagascar. Association Vahatra / Guide sur la diversité biologique, 148
p.
30. Ramanitra N. A. 2006, Contribution aux
études écologique et biologique de trois espèces
sympatriques de cuculidae dans les forêts d'Andapa : Coua caerulea,
Coua serriana et Coua reynaudi, thèses de Doctorat 192
pages
31. Rand, A.L. 1936. Distribution and habits
of Madagascar birds. Summary of the field notes of the Mission
Franco-Anglo-Américaine à Madagascar. Bulletin of the American
Museum of Natural History 72: 143-499.
32. Rasolonjatovo Delphin, 2018, Etude de la
taille de population, de distribution et de l'écologie d'aigle
pêcheur Madagascar Haliaetus vociferoides, dans la partie
occidentale de Madagascar, mémoire de DEA, Université de Toliara,
79p
33. Raveloson Laurent, 2017, Etude
biologique et écologique du faucon à ventre rayé
Falcon zoniventris (Peter, 1854) dans le complexe Tsimembo
Manambolomaty, centre Ouest de Madagascar, mémoire de DEA,
Université de Toliara, 63p
34. Rejo-Fienena F. 1995, Etude
phytosociologique de la végétation de la région de Toliara
(Madagascar) et des gestions des ressources végétales par les
populations locales, Thèse de Doctorat du Museum National d'Histoire
Naturelle (paris), 182p
35. René de Roland, L.-A., R. Thorstrom and
R.T. Watson 1996. Breeding records and nestling predation of Henst
Goshawks on Masoala Peninsula, Madagascar. Ostrich 67: 168-170
36.
76
Rene de Roland L. A. 2000, Contribution
à l'étude biologique, écologique et éthologique de
trois espèces d'Accipiter dans la presqu'île de
Masoala: A francesii (Smith, 1834), A. madagascariensis
(Smith, 1834) et A. henstii (Shlegel, 1873),
thèse de doctorat de troisième cycle en Biologie, Ecologie et
Conservation animale, Faculté de science de l'Université
d'Antananarivo, 118p
37. René de Roland, L.-A. 2001,
Comparaison du régime alimentaire des trois espèces d'Accipiter
dans la forêt pluviale, au nord-est de Madagascar. Ostrich
supplément 15: 206-209
38. René de Roland, L. -A., Rabearivony, J.,
Razafimanjato, G., Robenarimangason, H. & Thorstrom, R. 2005.
Breeding biology and diet of Banded Kestrels Falco zoniventris on
Masoala Peninsula, Madagascar. Ostrich. 76 (1&2): 32-36
39. René de Roland, L.-A. 2013a.
Henst's goshawk, Accipiter henstii, Autour de Henst; pp. 291-293, in:
R. Safford and F. Hawkins (ed.). Birds of Africa, vol. VI: the Malagasy region.
London: Christopher Helm
40. René de Roland, L.-A. 2013b.
Madagascar sparrowhawk, Accipiter madagascariensis, Epérvier de
Madagascar ; pp. 289-291, in: R.
41. Robenarimangason, H. 1999, Contribution
à l'étude biologique, écologique et éthologique de
la famille de la Falconidé, Faucon de newton (Falcon newton) et
Faucon à ventre rayé Falco zoniventris dans la partie
occidentale de presqu'île de Masoala, Mémoire de DEA,
Université d'Antananrivo, S1p.
42. Safford, R. and Hawkins, A. F. A. (ed.)
2013. Birds of Africa, vol. VI: the Malagasy region. London:
Christopher Helm. 1024 pp
43. Simmons, R.E.1986, Food provisioning,
nestling growth and experimental manipulation of brood size in the African
Red-Breasted Accipiter rufiventris. Ornis scandinavica, 17:31-40
44. Strenna, L., & Boileau, N. 2011, La
capture de faucon crécerelle Falco tinnunculus au bal chatri in
Bourgogne (1992-2005), Aluadia, 79 : 43-52
45. Thiollay, J.M. & Meyburg, B.M. 1983,
Remarque sur l'organisation d'un peuplement insulaire de rapace : Madagascar ;
Aluaudia, 49(3) : 216-226
46. Thomasson, 1991, Essai sur la flore du
sud-ouest malgache, originalité, affinité et origine, Bulletin du
Museum National d'Histoire Naturelle, Paris 4ème, Adansonia
no 12, 13 : 71- 89
47.
77
Thorstrom, R. 1996, Methods of capturing
tropical forest bird of prey, wildlife society bulletin 3:516-520p
48. Thorstrom, R., Ramos, José, D. &
Castillo, J.M. 2000, Breeding biology and behavior of the collard
forest Falcon (Micrastur semitorquatus) in Guatemala, volume 11 Ornithologia
Neotropical. 12p
49. Villages, A., 1990, Raptor biology,
Reproduction in bird of prey Ian Newton (ed) fact on file, INC, USA, Pp
124-140
50. Waston, R. T; & Lewis R. 1992, Raptors
studies in Madagascar rain forest. In raptor conservation today
I
ANNEXES
Annexe 1 : Tableau ombrométrique de la
température moyenne et de la précipitation à Mangily,
2016
|
Mois
|
Précipitation (mm)
|
T moyennes
|
T min
(°C)
|
T max (°C)
|
|
Janvier
|
10,3
|
28,8
|
24,0
|
33,6
|
|
Février
|
96,4
|
28,4
|
24,1
|
32,7
|
|
Mars
|
116
|
28,6
|
24,6
|
32,6
|
|
Avril
|
0
|
25,6
|
19,8
|
31,4
|
|
Mai
|
1,1
|
23,0
|
16,5
|
29,4
|
|
Juin
|
7,9
|
21,9
|
15,6
|
28,3
|
|
Juillet
|
13,2
|
22,1
|
16,2
|
28,0
|
|
Aout
|
0
|
21,8
|
15,8
|
27,8
|
|
Septembre
|
0
|
23,4
|
16,9
|
29,9
|
|
Octobre
|
13,1
|
24,1
|
19,1
|
29,0
|
|
Novembre
|
0,5
|
26,4
|
21,9
|
30,8
|
|
Décembre
|
11,9
|
27,7
|
23,1
|
32,3
|
Source : Service météorologique de Toliara, 2016
II
Annexe 2 : Distribution mensuelle des
précipitations durant 30 ans (1981-2010) en mm.
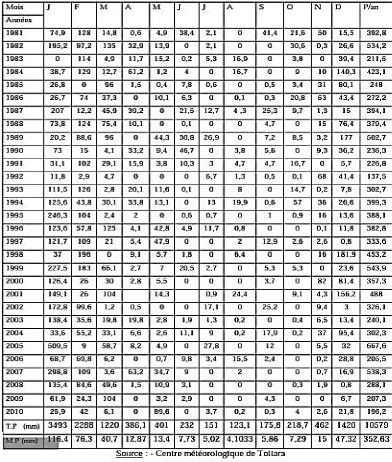
Annexe 3 : Poids des poussins par 5 cinq jours,
Rakotoson, 2020
|
Espèces
|
Jour de mensuration
|
|
|
5éj
|
10ej
|
15ej
|
20ej
|
25j
|
|
Accipiter madagascariensis
|
42
|
75
|
108
|
140
|
170
|
Poids en gramme (g)
|
|
Falco zoniventris
|
27
|
49
|
82
|
111
|
129
|
III
Annexe 4 : Régime alimentaire
|
Type de régime alimentaire
|
Accipiter madagascariensis
|
Falco zoniventris
|
|
Insectes
|
11
|
31
|
|
Reptiles
|
32
|
38
|
|
Oiseaux
|
106
|
0
|
|
Total
|
149
|
69
|
Annexe 5 : Photos Faucons à ventre
rayé (en haut) et Eperviers de Madagascar (en bas)


IV
Faucon à ventre rayé Falco zoniventris
(Cliché par Rene de Rolland, 2008)

Adulte femelle de l'Epervier de Madagascar Accipiter
madagascariensis (cliché par Rene de Roland en 2007)
V
Annexe 6 : Listes des oiseaux visibles au Reniala
|
Nom scientifique
|
Nom Français
|
L.C (en cm)
|
|
Leptopterus chabert
|
Artamie de Chabert
|
14
|
|
Leptopterus viridis
|
Artamie en tête blanche
|
20
|
|
Lonchura nana
|
Mannikin de Madagascar
|
9
|
|
Oena capensis
|
Tourterelle à masque de fer
|
28
|
|
Merops sperciliosus
|
Guêpier de Madagascar
|
27 à 33
|
|
Monias benschI
|
Monias de Bensch
|
32
|
|
Milvus migranus
|
Milan noir
|
55 à 60
|
|
Nectarinia notata
|
Souimanga angladian
|
14
|
|
Nectarinia souimanga
|
Souimanga malgache
|
10 à 11
|
|
Neomixis striatigula
|
Grande eroesse
|
12
|
|
Neomixis tenella
|
Petite eroesse
|
10
|
|
Nesillas typica
|
Fauvette de Madagascar
|
17 à 18
|
|
Newtonia archboldi
|
Newtonie d'Archobold
|
12
|
|
Newtonia brunneicauda
|
Newtonie commune
|
12
|
|
Numida meleagris
|
Pintade mitrée
|
58 à 65
|
|
Otus rotilus
|
Petit duc de Madagascar
|
22 à 24
|
|
Polyboroides radiatus
|
Polyboroide rayé
|
68
|
|
Streptolia picturata
|
Tourterelle peinte
|
28
|
|
Terpsiphone mutata
|
Gobe-mouche de paradis de
Madagascar
|
18 à 30
|
|
Thamnornis chloropetoides
|
Thamnornis
|
15
|
|
Turnix nigricollis
|
Turnix de Madagascar
|
14 à 16
|
|
Tyto alba
|
Chouette effraie
|
32 à 36 19
|
|
Upupa epops
|
Huppe fasciée
|
32
|
|
Uratelornis chimaera
|
Rollier terrestre a longue queue
|
47
|
|
Vanga Curvirostris
|
Vanga écorcheur
|
25 à 29
|
VI
Xenopirostris xenopirostris
|
Vanga de Lafresnaye
|
24
|
|
Zoovena grandidieri
|
Martinet de Grandidier
|
12
|
|
Falco concolor
|
Faucon concolore
|
31 à 33
|
|
Asio madagascariensis
|
Hibou de Madagascar
|
40 à 50
|
|
Ninox superciliaris
|
Ninox à sourcils
|
30
|
|
Accipiter francesI
|
Epervier de Frances
|
30 à 35
|
|
Accipiter madagascariensis
|
Epervier de Madagascar
|
34 à 40
|
|
Acridotheres tristis
|
Martin Triste
|
24
|
|
Agapornis cana
|
Inséparable à tête grise
|
14 à 16
|
|
Treron austra
|
Pigeon vert de Madagascar
|
32
|
|
Aviceda madagascariensis
|
Baza malgache
|
40 à 45
|
|
Buteo brachypyerus
|
Buse de Madagascar
|
48 à 51
|
|
Calicalicus madagascariensis
|
Vanga à queue rousse
|
13.5 à 14
|
|
Caprimulgus
madagascariensis
|
Engoulevent de Madagascar
|
21
|
|
Caprimulgus enarratus
|
Engoulevent à collier
|
24
|
|
Centropus toulou
|
Coucal malgache
|
45 à 50
|
|
Copsychus albospecularis
|
Dyal malgache
|
18
|
|
Coracina cineria
|
Echenilleur malgache
|
24
|
|
Coracopsis nigra
|
Petit perroquet noir
|
35
|
|
Coracopsis vasa
|
Grand perroquet vasa
|
50
|
|
Coua cristata
|
Coua huppée
|
40 à 44
|
|
Coua cursor
|
Coua coureur
|
40
|
|
Coua gigas
|
Coua géant
|
62
|
|
Coua R. olivaceiceps
|
Coua Olivier de Madagascar
|
43
|
|
Cuculus rochu
|
Coucou de Madagascar
|
28
|
|
Dicrurus forficatus
|
Drongo Malgache
|
26
|
|
Eurystomus glaucurus
|
Rollier Malgache
|
32
|
|
Falco newtoni
|
Faucon de Newton
|
30
|
VII
Falco zoniventris
|
Faucon à ventre rayé
|
35
|
|
Falculea palliata
|
Falculie mantelée
|
32
|
|
Foudia madagascariensis
|
Foudi de Madagascar
|
13.5 à 14
|
|
Foudia omissa
|
Foudi de forêt
|
14.5
|
|
Plocus sakalava
|
Tisserin Sakalava
|
15 à 15.5
|
|
Hypsipetes madagascariensis
|
Bulbul noir
|
24
|
Source: Reniala, 2018
Annexe 7 : Listes des plantes
répertoriées au Reniala (32 espèces)
|
Nom vernaculaire
|
Nom scientifique
|
Famille
|
|
Boy
|
Commiphora lami
|
Burseraceae
|
|
Dangoa
|
Strychnos decussak
|
Loganiaceae
|
|
Famata
|
Euphorbia stenoclada
|
Euphorbiaceae
|
|
Fandriandambo
|
Rhopalopilia sp
|
Opiliaceae
|
|
Fandrifotry
|
Euphorbia pervilleana
|
Euphorbiaceae
|
|
Farafatry
|
Givotia madagascariensis
|
Euphorbiaceae
|
|
Fengoke
|
Delonix decaryi
|
Fabaceae
|
|
Fony
|
Adansonia rubrostipa ou fony
|
Malavaceae
|
|
Hatratra
|
Jatropha mahafaliensis
|
Euphorbiaceae
|
|
Hazomafio
|
Zanha slaveolens
|
Sapindaceae
|
|
Hazomena
|
Securinega perrierI
|
Euphorbiaceae
|
|
Hazondragnaty
|
Sapium sp
|
Euphorbiaceae
|
|
Holaboay
|
Adenia olaboensis
|
Passifloraceae
|
|
Karabo
|
Cordyla madagascariensis
|
Fabaceae
|
|
Katrafay
|
Cedrelopsis grevei
|
Rutaceae
|
|
Kifafa
|
Henonia scoparia
|
Amaranthaceae
|
|
Laro
|
Euphorbia laro
|
Euphorbiaceae
|
|
Lovainafy
|
Lovanafia capuroniana
|
Fabaceae
|
|
Manary Foty
|
Dalbergia purpurescens
|
Fabaceae
|
VIII
Monongo
|
Zanthoxylum decaryi
|
Rutaceae
|
|
Nato
|
Capurodendron perrieri
|
Sapotaceae
|
|
Pisopiso zalazala
|
Croton sp
|
Euphorbiaceae
|
|
Ranga
|
Cynanchum perrierI
|
Apocynaceae
|
|
Romby
|
Commiphora mahafaliensis
|
Burseraceae
|
|
Sanganakoholahy
|
Chadsia grevei
|
Fabaceae
|
|
Sogno
|
Didierea madagascariensis
|
Didieraceae
|
|
Sonotsoy
|
Fernandoa madagascariensis
|
Bignoniceae
|
|
Vahontsoy
|
Aloe divaricata
|
Liliaceae
|
|
Vaovy
|
Tetrapterocarpon geayi
|
Fabaceae
|
|
Varo
|
Cordia madagascariensis
|
Boraginaceae
|
|
Velomihanto
|
Bakerella sp
|
Loranthaceae
|
|
Vontake
|
Pachypodium sp
|
Apocynaceae
|
Source: Reniala, 2018
IX
TABLE DES MATIERES
REMERCIEMENTS i
LISTE DES FIGURES vi
LISTE DES TABLEAUX vi
INTRODUCTION GENERALE 1
INTRODUCTION 2
PARTIE I : METHODOLOGIE 1
I.1. PRESENTATION DE LA ZONE D'ETUDE 6
I.1.1. Contexte historique et localisation géographique
de la zone d'étude 6
I.1.2. Milieu abiotique 9
I.1.2.1. Le climat 9
I.1.2.2. Géo-pédologie 12
I.1.3. Milieu Biologique 12
I.1.3.1. Flore et Végétation 12
I.1.3.2. Faune 13
I.1.4. Population et ses activités 14
I.2. PRESENTATION DES ESPECES ETUDIEES 16
I.2.1. Position Systématique 17
I.2.1.1. Epervier de Madagascar (Accipiter madagascariensis)
17
I.2.1.2. Faucon à ventre rayé (Falco
zoniventris) 18
I.2.2. Description morphologique 18
I.2.2.1. Epervier de Madagascar (Accipiter madagascariensis)
18
I.2.2.2. Faucon à ventre rayé (Falco
zoniventris) 19
I.2.3. Distribution et statut de l'espèce 20
I.2.3.1. Epervier de Madagascar (Accipiter madagascariensis)
20
I.2.3.2. Faucon à ventre rayé (Falco
zoniventris) 20
I.3. MATERIELS UTILISES SUR LE TERRAIN ET LEURS UTILISATIONS
22
I.4. METHODES DE COLLECTES DE DONNEES 22
I.4.1. Période d'étude sur le terrain 22
I.4.2. Technique de recensement pour l'inventaire des individus
et des nids 22
I.4.2.1. Enquête 23
I.4.2.2. Observation directe et suivi de l'individu 23
I.4.2.3. Méthode de point d'écoute et visite de
sentier 24
X
I.4.3. Observation du nid 24
I.4.4. Capture et relâchement 24
I.4.5. Mensuration et pesage des adultes et des poussins 26
I.4.6. Biologie de la reproduction des espèces 26
I.4.6.1 Formation du couple et Accouplement 27
I.4.6.2. Ponte 27
I.4.6.2. Incubation et éclosion 27
I.4.6.3. Elevage et croissance des poussins 28
I.4.6.4. Productivité 28
I.4.7. Etude écologique des espèces 28
I.4.7.1. Etude de nid 29
I.4.7.2. Régime alimentaire 31
I.4.7.3. Evaluation des menaces et des pressions dans l'ensemble
d'habitat 31
I.4.8. Comportements des adultes 31
I.5. ANALYSE DE DONNEES 33
I.5.1. Moyen et écart-type 33
I.5.2. Test de Chi-deux (÷2) 33
PARTIE II : RESULTATS ET INTERPRETATIONS 35
II.1. NOMBRE DES INDIVIDUS ET DES NIDS TROUVES 37
II.1.1. Nombre des individus 37
II.1.2. Nombre des nids 38
II.2. MORPHOMETRIE 39
II.3. BIOLOGIE DE LA REPRODUCTION 40
II.3.1. Période de formation de couple, accouplement et
ponte 40
II.3.2. Période de Ponte et taille des oeufs 40
II.3.3. Incubation et éclosion 40
II.3.3.1. Incubation 41
II.3.3.2. Période d'incubation et taux d'éclosion
41
II.3.4. Elevage des poussins dans le nid 42
II.4. MENSURATION ET PESAGE DES POUSSINS 43
II.3.4. Productivité 44
II.4. ECOLOGIE 45
II.4.1. Caractéristique des nids 45
XI
II.4.2. Analyse de l'habitat de nidification 47
II.4.3. Territoire de chasse, perchoir et dortoir 48
II.4.3. Régime alimentaire 49
II.4.4. Pressions et menaces 51
II.5. ETHOLOGIE 53
II.5.1. Comportement de chasse 53
II.5.2. Livraison des proies entre le couple et Elevage des
poussins 54
II.5.3. Relation interspécifique et défense
territoriale 55
II.5.4. Vol et cris 55
PARTIE III : DISCUSSION 37
III.1. DISTRIBUTION ET TAILLE DES POPULATIONS 59
III.2. RECHERCHE DE NIDS 60
III.3. METHODE DE CAPTURE 61
III.4. MORPHOLOGIE ET MORPHOMETRIE DES ADULTES 62
III.5. LA REPRODUCTION 63
III.5.1. Période de formation du couple, nidification et
ponte 63
III.5.2. Incubation, éclosion et couvaison 64
III.6. ECOLOGIE 66
III.6.1. Habitats de nidification et caractéristique de
nids 66
III.6.2. Type des proies 67
III.7. COMPORTEMENT 68
III.7.1. Comportement pendant la reproduction 68
III.7.2. Relation interspécifique 68
CONCLUSION 70
RECOMMANDATIONS 72
BIBLIOGRAPHIE 73
ANNEXES I
Keywords: biology, ecology, Accipiter
madagascariensis, Falco zoniventris, Reniala, Parc Mosa, Madagascar.
RAKOTOSON Mauhamed Galien, Contact: 0343867058 (
galienohh@gmail.com)
RESUME
Cette présente étude se focalise sur la biologie
et l'écologie des rapaces sympatriques Epervier de Madagascar
Accipiter madagascariensis et Faucon à ventre rayé
Falco zoniventris, dans les forêts sèches de Mangily,
à savoir la réserve privée Reniala et Parc Mosa
(forêt de baobab), Sud-ouest de Madagascar. L'étude sur terrain
dure quatre mois, début novembre 2019 au début de mars 2020.
Comme méthodologie, les méthodes d'enquête sous forme de
dialogue auprès des villageois et d'écoute ont été
utilisées pour la localisation des nids ; comptage direct en suivant le
transect pour connaitre l'effectif des individus, observation directe et
continue en utilisant des matériels des mesures tels que les
télescopes, le GPS, la balance, le chronomètre, etc. ont
été adoptés pendant les collectes de données
bioécologiques. Deux couples de chaque espèce ont
été étudiés durant cette étude.
D'après les résultats, 29 individus ont été
recensés dont 11 Accipiter madagascariensis et 18 Falco
zoniventris. Ces deux espèces fréquentent dans le milieu
forestier protégé. Les résultats ont aussi
révélé que la période de reproduction de ces
espèces se réalise pendant l'été. L'effectif des
oeufs par nids varie de 3 à 4 (n=3 nid1 et n=4 nid2) pour Accipiter
madagascariensis et 2 à 3(n=2 nid3 et n=3 nid4) pour Falco
zoniventris et la période d'éclosion se situe entre les mois
de novembre et de décembre pour les deux espèces. Tous les oeufs
colorés en blancs pèsent en moyenne 32g et mesure 39,25*36,2 mm
pour Accipiter madagascariensis et 24,4g et 38,2*32,2mm pour Falco
zoniventris, sont tous éclos en donnant 12 poussins,
malheureusement l'un des poussins de Faucon à ventre rayé n'a pas
pu atteindre le stade en vol lors de la pluie violente qui donne enfin les taux
de productivité 100% pour Accipiter madagascariensis et 80%
pour Falco zoniventris. Les activités telles que l'incubation,
la couvaison et le nourrissage des poussins sont assurées par les
femelles tandis que la défense du territoire et la chasse sont le
responsable des mâles, et la livraison de proies entre le couple se
réalise en dehors de l'arbre de nidification. Le type de proies
chassés et livrés par l'Epervier de Madagascar est basé
principalement en petits oiseaux, mais cette espèce chasse aussi les
reptiles et les insectes tandis que les Faucons à ventre rayé ne
chassent seulement que les deux derniers types. Les menaces de ces
espèces sont la chasse de la population locale en raison d'éviter
la lutte des ennemis des poussins domestiques pour Accipiter
madagascariensis et l'exploitation des arbres de nidification pour
Falco zoniventris. Les données obtenues sur cette recherche nous
aident à fournir des informations sur ces deux rapaces sympatriques afin
de permettre et d'évaluer la conservation.
Mots-clés : biologie, écologie,
Accipiter madagascariensis, Falco zoniventris, Reniala, Parc Mosa,
Madagascar.
ABSTRACT
This present study focuses on the biology and ecology of
sympatric raptors Madagascar's sparrowhawk Accipiter madagascariensis
and Madagascar's Banded kestrel Falco zoniventris, in the dry
forests of Mangily, namely the private reserve Reniala and Parc Mosa (baobab
forest), Southwest of Madagascar. The field study lasts four months, November
2019 to early March 2020. As a methodology, survey methods in the form of
dialogue with the villagers and listening were used for the location of the
nests; direct counting by following the transect to know the number of
individuals, direct and continuous observation using measuring equipment such
as telescopes, GPS, scales, chronometers, etc. were adopted during
bioecological data collections. Two pairs of each species were studied during
this study. According to the results, 29 individuals were identified including
11 Accipiter madagascariensis and 18 Falco zoniventris. These
two species frequent in the protected forest environment. The results also
revealed that the breeding season for these species occurs during the summer.
The number of eggs per nest varies from 3 to 4 (n = 3 nest1 and n = 4 nest2)
for Accipiter madagascariensis and 2 to 3 (n = 2 nest 3 and n = 3 nest
4) for Falco zoniventris and the hatching period is between the months
of November and December for both species. All the colored eggs white, weigh on
average, 32g and measure 39.25 * 36.2 mm for Accipiter madagascariensis
and 24.4g and 38.2 * 32.2mm for Falco zoniventris, are all
hatched giving 12 chicks, unfortunately the one of the Banded kestrel chicks
could not reach the mid-flight stage during the heavy rain, which finally gives
the productivity rates 100% for Accipiter madagascariensis and 80% for
Falco zoniventris. Activities such as incubation, brooding and feeding
of the chicks are carried out by the females while the defense of the territory
and hunting are responsible for the males, and the delivery of prey between the
pair takes place outside the tree nesting. The type of prey hunted and
delivered by the Madagascan Hawk is based primarily on small birds, but this
species also hunts reptiles and insects while the Madagascar's Banded kestrel
hunts only the latter two types. Threats of these species are hunting by local
people due to the avoidance of pest control of domestic chicks for
Accipiter madagascariensis and exploitation of nesting trees for
Falco zoniventris. The data obtained from this research helps us to
provide information on these two sympatric raptors to enable and assess
conservation.
| 

