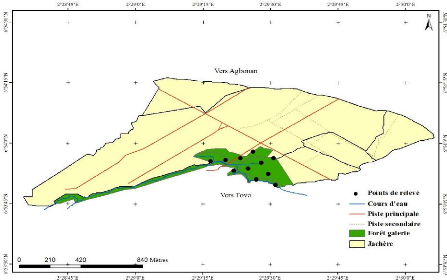
Caractéristiques structurales et écologiques des forêts de Bonou et d'Itchèdè au sud- est du Bénin( Télécharger le fichier original )par Sunday Berlioz KAKPO Université d'Abomey- Calalvi, faculté des sciences agronomiques - Diplome d'ingénieur agronome spécialiste en aménagement et gestion des ressources naturelles 2011 |

Où x = diamètre des arbres; a =10 cm pour les structures en diamètre; a=5 cm pour les structures en hauteur; b = paramètre d'échelle lié à la valeur centrale des diamètres ou hauteurs; c= paramètre de forme lié à la structure en diamètre ou hauteur considérée. Pour chaque peuplement et chaque population concernée, les données de diamètre ou de hauteur des arbres ont servi à l'estimation des paramètres a, b et c grâce à la méthode du maximum de vraisemblance disponible dans le langage MatLab (versionR2006a). La distribution de Weibull peut prendre plusieurs formes selon la valeur du paramètre de forme c. Tableau 1 : Interprétations des paramètres de forme « c »
Source : Rondeux, 1999
14 L'analyse log-linéaire (ou analyse de variance basée sur le maximum de variance) est réalisée pour comparer dans chaque cas, la distribution observée à la distribution théorique de Weibull avec estimation des paramètres. 3.2.4. Répartition spatiale des espèces de valeurPour étudier la répartition spatiale des espèces de valeur, nous avons compté par espèce et pour chaque individu de diamètre = 1 cm, le nombre d'individus de l'espèce se trouvant dans un rayon de 10 m, puis sur une bande de 10 à 20 m et de 20 à 30 m autour de chacun des individus de l'espèce considérée (Condit et al., 2000) (Figure 5). Cette méthode permet le calcul de la densité relative de voisinage Ù de chaque espèce étudiée. Selon Condit et al. (2000), la densité relative de voisinage est un indice qui n'exprime pas seulement la structure des peuplements ou des populations d'espèces en termes de valeurs moyennes ou de distribution, mais elle décrit la structure spatiale de manière continue. Cet indice peut être comparé à la fonction K de Ripley. Il permet aussi d'avoir des connaissances plus explicites sur la spatialisation et sur des interactions entre les arbres sur des séquences de distances. Figure 5 : Dispositif d'échantillonnage des espèces de valeur
Repère de comptage 10m 30m 20m ??? = ???? ???? (1) Avec Dx la densité spécifique et Ds la densité relative ???? = ???? ???? et ???? = ???? ???? Ax la surface inventoriée par rapport à l'anneau considéré ; Nx le nombre d'individu de l'espèce dans l'anneau considéré ; Ni le nombre d'individu de l'espèce de valeur dans le placeau ; Sp la surface du placeau. L'interprétation se fait par rapport aux différentes valeurs obtenues. En effet, si ?x=1, la distribution est dite aléatoire ?x >1, la distribution est dite agrégative ?x < 1 ; on parle d' hyperdispersion. 3.2.5. Traitement des données? La densité du peuplement (N, en arbres/ha) : il s'agit du nombre moyen d'arbres de dbh = 10cm sur pied par hectare (1) étant le nombre total d'arbres par placette et s la surface en ha de la placette ? La densité de régénération Nr= i ???? ?? ??=i (2) ?? yi étant la densité de régénération dans l'unité d'échantillonnage j tandis que n est le nombre total d'unités d'échantillonnage. g h i i ? Le diamètre de l'arbre de surface terrière moyenne (g, en cm) Dg ? 1 n ? 2 , (3) n
15 n est le nombre d'arbre de dbh = 10 cm à l'intérieur du placeau et diamètre en cm de l'arbre j. ? La hauteur moyenne de Lorey (H, en mètre) est la hauteur moyenne de tous les arbres inventoriés à l'intérieur d'un placeau, pondéré par leur surface terrière: n ? g i ? 1 2 H ? avec g ? d
(4) i h i ? n i i ? gi 4
i ? , et dj respectivement la surface terrière et la hauteur totale et le diamètre de l'arbre i.
16 ? La surface terrière (, en m2/ha) est la somme, ramenée à l'hectare des sections transversales à hauteur d'homme (1,3 m) de tous les arbres de dbh = 10 cm, à l'intérieur du placeau ; (5) étant le diamètre en centimètre (cm) de l'arbre i du placeau. ? La richesse spécifique La richesse spécifique est apparemment un indice de diversité extrêmement simple. Notée S, elle est le nombre (ou une fonction croissante du nombre) de catégories différentes présentes dans le système étudié, par exemple le nombre d'espèces d'arbres dans une forêt (Marcon, 2011). ? L'indice de diversité de Shannon (H, en bits) L'indice de Shannon (Shannon, 1948 ; Shannon et Weaver, 1963), aussi appelé indice de Shannon-Weaver ou Shannon-Wiener, est dérivé de la théorie de l'information. ?? = - ???? ?? ??=?? ???????? ????? ? (6) ?? ni = nombre d'individus de l'espèce i ; n = nombre total d'individus inventoriés dans les placeaux. Cet indice est maximal quand tous les individus sont répartis de façon équitable sur toutes les espèces. Il rend compte de l'organisation des espèces et des individus au sein de la communauté végétale. Très souvent la valeur de H est comprise entre 0 et 5 (normalement la limite supérieure est log2S). Si H est proche de 0, la communauté est peu diversifiée et si H est compris entre 3 et 4,5 bits, la communauté est relativement diversifiée (Marcon, 2011). ? L'équitabilité de Pielou La régularité de la distribution des espèces (équitabilité en Français, evenness en Anglais) est un élément important de la diversité. Une espèce représentée abondamment ou par un individu n'apporte pas la même contribution à l'écosystème (Marcon, 2011). ???? = ?? ?? ???????? ???? ???? = ?????????? (7) ?????? Hmax indice de diversité maximale théorique de Shannon lié au peuplement. L'indice Eq varie de 0 à 1; il est maximal quand les espèces ont des abondances identiques dans le peuplement et minimal quand un petit groupe d'espèces domine tout le peuplement. Lorsque « Eq » est compris entre 0,7 et 0,9, toutes les espèces sont bien représentées au sein des individus
17 (Marcon, 2011). Lorsque Eq < 0,6, il y a une espèce dominante et les autres le sont moins ou il s'agit d'une communauté peu diversifiée. S le nombre total d'espèces. A partir des données d'inventaire des forêts de Bonou et d'Itchèdè la structure en hauteur et en diamètre du peuplement ont été établies ainsi que leur ajustement à la distribution de Weibull qui caractérise mieux la variabilité des formes des structures de peuplements forestiers. Pour s'assurer du bon ajustement de la structure observée à la distribution théorique, il est nécessaire de réaliser un test statistique d'ajustement. La méthode statistique souvent utilisée à cet effet est le test Chi-carré d'ajustement de Pearson ou l'une de ses variantes (Dagnelie, 1998). Néanmoins, il est à reconnaître que ces tests sont imprécis dans des situations de faibles effectifs de classes ou de faible nombre de classes. Des procédures Minitab et SAS relatives à la construction de ces structures, à leur ajustement à la distribution théorique de Weibull par l'analyse log linéaire ont été conçues.
18 4. RéSULTATS4.1. Caractéristiques structurales et écologiques des forêts de Bonou et d'Itchèdè4.1.1. Paramètres dendrométriquesLes paramètres dendrométriques des forêts de Bonou et d'Itchèdè sont présentés au tableau 2 et les indices relatifs à leur dispersion sont présentés au tableau 3. On note que la densité moyenne des arbres sur pied (N) et le diamètre moyen des arbres sont significativement différents (Prob ? 0,05). Par contre la surface terrière (G) et la hauteur moyenne de Lorey (HL) présentent des différences non significatives entre les deux forêts (Prob > 0,05). Tableau 2 : Moyenne des paramètres dendrométriques des forêts de Bonou et d'Itchèdè
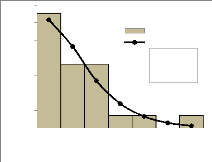
Moy : moyenne ; sur une même ligne, les chiffres affectés des mêmes lettres ne présentent pas de différence significative au seuil de 5%. 4.1.2. Structure en diamètre des forêtsUne différence très hautement significative entre les classes de diamètre du point de vue de leurs fréquences est notée au niveau de la structure en diamètre des deux forêts. Ceci est assez évident puisque dans une telle structure, les individus jeunes sont nettement plus fréquents que les individus adultes. On remarque, en analysant la structure des deux forêts, que la différence entre la distribution observée et la distribution théorique de Weibull n'est pas significative (Prob = 0.62 à Bonou et 0.73 à Itchèdè) et indique globalement une concordance entre la structure observée et celle déduite de la distribution de Weibull. On note des figures 6 et 7 que la structure en diamètre des peuplements présente une forme en « J renversé » avec le paramètre de forme, c prenant une valeur inférieure à 1. Cette structure est donc régressive. Il est rare d'observer plus de 10 arbres à l'hectare pour des diamètres allant au delà de 60 cm. Que ce soit dans la forêt de Bonou ou d'Itchèdè, les individus de diamètre compris entre 10 cm et 20 cm sont les plus représentés à l'hectare. Les individus de diamètre supérieur à 110 cm sont rares au sein des deux forêts. 10_20 20_30 30_40 40_50 50_60 60_70 70_80 80_90 90_100 100_110 110_120 120_130 130_140 140_150 150_160 160_170 170> Classe de diamètre (cm)
Densité (arbre/ha) 200 Bonou a = 10 b =13,44 c = 0,83 Densité observée Weibull 0 150 100 50 10--20 20--30 30--40 40--50 50--60 60--70 70--80 80--90 90--100 100--110 110--120 120--130 130--140 140--150 150--160 160--170 170--180 >180 Classe de diamètre (cm)
Densité (arbre/ha) 40 80 70 60 50 30 20 10 0 Itchèdè a = 10 b = 20,38 c = 0,9349 Densité observée Weibull
19 Figure 6 : Structure en diamètre de la forêt de Bonou Figure 7 : Structure en diamètre de la forêt d'Itchèdè 4.1.3. Structure en hauteur des forêtsOn remarque en analysant la structure en hauteur des deux forêts que le test lié à la différence entre la distribution observée et la distribution théorique de Weibull n'est pas significative (Prob = 0,88 à Bonou et 0,74 à Itchèdè) et indique globalement une adéquation entre la structure observée et celle attendue sur la base de la distribution de Weibull. La distribution en hauteur dans les forêts de Bonou et d'Itchèdè est de forme exponentielle négative (figures 8 et 9). Le paramètre de forme est égal à 1,31 dans la forêt de Bonou et 1,18 dans la forêt d'Itchèdè. On note une prédominance d'individus de petites hauteurs. Les sujets dont la hauteur est comprise entre 4 m et 12 m sont les plus nombreux à l'hectare. Les sujets de hauteur supérieure à 36 m sont faiblement représentés dans les différentes forêts.
Densité (arbre/ha 180 160 140 120 100 40 80 60 20 0 4_12 12_20 20_28 28_36 36_44 > 44 Classe de hauteur (m) Bonou Densité observée Weibull a = 4 b = 10,97 c = 1,318
100 Itchèdè Densité observée Weibull a = 4 b = 11,35 c = 1,181 0 80 60 40 20 Densité (arbre/ha) 4_12 12_20 20_28 28_36 36_44 >44
20 Figure 8 : Structure en hauteur de la forêt de Bonou Figure 9 : Structure en hauteur de la forêt d'Itchèdè 4.1.4. Densité de régénérationLa densité moyenne globale de régénération (toutes espèces confondues) est estimée à 2006 tiges/ha dans la forêt de Bonou alors qu'elle est de 1987 tiges/ha dans la forêt d'Itchèdè (tableau 3). Il est à constater que la densité de régénération va décroissant suivant les stades croissants de développement quelle que soit la forêt. Tableau 3 : Densité moyenne de régénération des forêts de Bonou et d'Itchèdè
4.1.5. Diversité floristique des forêtsDe façon globale, la richesse en espèces des individus arborescents (dbh = 10cm) dans la forêt de Bonou est de 38 espèces avec un indice de diversité de Shannon égal à 4,6 bits qui traduit une inégale répartition des individus des espèces présentes et aussi une forte diversité spécifique, en comparaison avec l'indice de diversité maximal théorique de Shannon lié au peuplement étudiée (H max = Log S = 5,2 bits). L'indice d'équitabilité de Piélou est égal à 0,9. On note dans l'ordre de rang décroissant d'abondance certaines espèces : Dyctiandra arborescens, Triplochiton scleroxylon, Aidia genipiflora, Tabernaemontana pachysiphon, Anthonotha macrophylla, Terminalia superba et Cola millenii. Dans la forêt d'Itchèdè, la richesse en espèces des individus arborescents (dbh = 10cm) est de 53 espèces avec un indice de diversité de Shannon égal à 5,03 bits qui traduit une forte diversité spécifique, en comparaison de l'indice de diversité maximal théorique de Shannon (H
21 max = Log S = 5,73 bits). L'indice d'équitabilité de Piélou est égal à 0,88. On note dans l'ordre de rang décroissant d'abondance certaines espèces : Dyctiandra arborescens, Cola millenii, Triplochiton scleroxylon, Ceiba pentandra, Celtis zenkeri, Olax subscorpinoides, Antiaris toxicaria. 4.2. Caractéristiques structurales et écologiques de la population de Cola millenii4.2.1. Paramètres dendrométriquesLes paramètres dendrométriques des populations de Cola millenii dans les différentes formations sont présentés au tableau 4. On note que la densité moyenne des arbres sur pied (N), le diamètre de l'arbre de surface terrière moyenne (Dg), la surface terrière (G), la hauteur moyenne et la hauteur moyenne de Lorey (HL) présentent tous des différences non significatives entre les deux forêts (Prob > 0,05). Tableau 4 : Paramètres dendrométriques de Cola millenü au sein des forêts de Bonou et d'Itchèdè
Moy : moyenne 4.2.2. Structure en diamètreEn analysant la structure eu diamètre de Cola millenii dans les des deux forêts, on remarque que la différence entre la distribution observée et la distribution théorique de Weibull n'est pas significative (Prob = 0,77 à Bonou et 0,43 à Itchèdè) et indique globalement une concordance entre la structure observée et celle déduite de la distribution de Weibull. On note des figures 10 et 11 que la structure en diamètre de la population de Cola millenii dans les deux forêts présente une distribution asymétrique droite ou asymétrique positive. Le paramètre de forme est égal à 1,228 dans la forêt de Bonou et 1,327 dans la forêt d'Itchèdè; tous compris entre 1 et 3,6. On note une prédominance d'individus de Cola millenii de petits diamètres. Les sujets dont le diamètre est compris entre 10 cm et 18 cm sont les plus nombreux à l'hectare. Les classes modales dans les forêts de Bonou et d'Itchèdè sont respectivement 10 à 14cm et 14 à 18cm. Les sujets de diamètre supérieur à 26 cm sont faiblement représentés dans les différentes forêts. Nous notons cependant que les fréquences de distribution sont irrégulières dans les classes de diamètre supérieures (dg>20cm) Classe de diamètre (Cm)
Densité (arbre/ha) 10 12 12 14 14 16 16 18 18 20 20 22 22 24 24 26 26 28 >28 4,5 3,5 2,5 0,5 1,5 4 5 3 2 0 1 Bonou Densité observée Weibull a = 10 b = 5,327 c = 1,228
Densité (arbre/ha) 2,5 0,5 1,5 3 2 0 1 10 12 12 14 14 16 16 18 18 20 20 22 22 24 24 26 26 28 >28 Classe de diamètre (Cm) Itchèdè Densité observée Weibull a =10 b = 6,92 c = 1,327
22 Figure 10 : Structure en diamètre du C. millenii dans la forêt de Bonou Figure 11 : Structure en diamètre du C. millenii dans la forêt d'Itchèdè 4.2.3. Structure en hauteurOn remarque en analysant la structure en hauteur de Cola millenii dans les deux forêts que la différence entre la distribution observée et la distribution théorique de Weibull n'est pas significative (Prob = 0,74 à Bonou et 0,91 à Itchèdè) et indique globalement une adéquation entre la structure observée et celle attendue sur la base de la distribution de Weibull. Les distributions en hauteur de la population de Cola millenii dans les différentes formations végétales sont présentées aux figures 12 et 13. Le paramètre de forme (c) est égal à 1,747 dans la forêt de Bonou et 1,416 dans la forêt d'Itchèdè. Le paramètre c est compris entre 1 et 3,6 quelle que soit la forêt. Il est donc évident que la distribution en hauteur de la population de Cola millenii dans les forêts de Bonou et d'Itchèdè est asymétrique droite ou asymétrique positive. Les sujets dont la hauteur est comprise entre 4 m et 12 m sont les plus nombreux à l'hectare. Les sujets de hauteur supérieure à 16 m sont faiblement représentés dans les différentes forêts. 8
4_8 8_12 12_16 16_20 >20 7 6 5 4 3 2 Densité (arbre/ha) 1 0 Bonou
Densité observée Weibull a = 4 b = 6,101 c = 1,747 Classe de hauteur (m)
Itchèdè 6 Densité observée Weibull a = 4 b = 6,095 c = 1,416 0 4 2 1 Densité (arbre/ha) 4_8 8_12 12_16 16_20 >20 Classe de hauteur (m)
23 Figure 12 : Structure en hauteur du C. millenii dans la forêt de Bonou Figure 13 : Structure en hauteur du C. millenii dans la forêt d'Itchèdè 4.2.4. Répartition spatialeD'après la méthode de densité relative de voisinage que nous avons utilisée, on déduit trois classes de densité relative de voisinage dans chaque anneau pour lesquelles ? prend les valeurs suivantes : ? >1 ; ? =1 et ? <1. La proportion de chaque classe par rapport au nombre total de placeaux inventoriés et suivant les catégories d'anneau est consignée dans le tableau ci-après. Tableau 5 : Densité relative de voisinage par catégorie d'anneau de Cola millenii dans les forêts de Bonou et d'Itchèdè
Le tableau 5 montre que sur un rayon de 10 m autour d'un pied de Cola millenii, la densité relative de voisinage de ce dernier est supérieure à 1 dans l'ensemble des placeaux inventoriés à Bonou. A Itchèdè, dans 95% des cas, la densité relative de voisinage de Cola millenii est supérieure à 1 (soit 19 sur 20 observations). Nous pouvons conclure que sur un rayon de 10 m la distribution de Cola millenii est agrégative (?>1). Au-delà d'un rayon de 10 m autour d'un pied de Cola millenii plus précisément sur les anneaux de 10 à 20 m et de 20 à 30 m la totalité des placeaux présente une répartition spatiale non agrégative et donc relativement dispersée de Cola millenii (Ù < 1), quelle que soit la forêt.
24 4.2.5. Densité de régénérationLa densité moyenne globale de régénération (tableau 6) est estimée à 254,54 tiges/ha pour Cola millenii dans la forêt de Bonou alors qu'elle est de 153,33 tiges/ha dans la forêt d'Itchèdè. Notons que la densité de régénération va décroissant suivant les stades de développement. Tableau 6 : Densité moyenne de régénération de Cola millenii
4.3. Caractéristiques structurales et écologiques des populations de Dialium guineense4.3.1. Paramètres dendrométriquesLes paramètres dendrométriques des populations de Dialium guineense dans les différentes formations sont présentés au tableau 11. On note que la densité moyenne des arbres sur pied (N), le diamètre de l'arbre de surface terrière moyenne (Dg), la surface terrière (G) et la hauteur moyenne de Lorey (HL) présentent tous des différences non significatives entre les deux forêts (Prob > 0,05). Tableau 7 : Paramètres dendrométriques de Dialium guineense au sein des forêts de Bonou et d'Itchèdè
Moy : moyenne 4.3.2. Structure en diamètreLes résultats d'analyse log-linéaire effectuée sur ces structures indiquent un bon ajustement des données à la distribution de Weibull (Prob = 0.81 à Bonou et 0.87 à Itchèdè) au seuil de 5%. On note des figures 14 et 15 que la structure en diamètre de la population de Dialium guineense dans les deux forêts présente une distribution asymétrique droite ou asymétrique positive. Le paramètre de forme est égal à 1,253 dans la forêt de Bonou et 1,709 dans la forêt d'Itchèdè; tous compris entre 1 et 3,6. On note une prédominance d'individus de Dialium guineense de petits diamètres. Les sujets dont le diamètre est compris entre 10 cm et 20 cm sont les plus nombreux à l'hectare. Les sujets de diamètre supérieur à 60 cm sont faiblement représentés dans les différentes forêts.
10--20 20--30 30--40 40--50 50--60 60--70 >70 Classe de diamètre (cm) 3,5 Bonou 3 2,5 2 1,5 1 Densité (arbre/ha) 0,5 0 Densité observée Weibull a = 10 b = 18,86 c = 1,253
Densité (arbre/ha) 0,5 1,5 2 0 1 10--20 20--30 30--40 40--50 50--60 >60 Classe de diamètre (cm) Itchèdè Densité observée Weibull a = 4 b = 11,97 c = 1,709
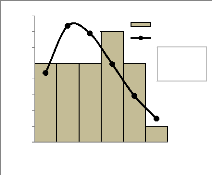
25 Figure 14 : Structure en diamètre de D. guineense dans la forêt de Bonou Figure 15 : Structure en diamètre de D. guineense dans la forêt d'Itchèdè 4.3.3. Structure en hauteurOn remarque en analysant la structure en hauteur des deux forêts que la différence entre les deux distributions n'est pas significative (Prob = 0,06 à Bonou et 0,98 à Itchèdè) et indique globalement une adéquation entre la structure observée et celle attendue sur la base de la distribution de Weibull. Le paramètre de forme (c) est égal à 1,759 dans la forêt de Bonou et 1,709 dans la forêt d'Itchèdè. Le paramètre c est compris entre 1 et 3,6 quelle que soit la forêt. Il est donc évident que la distribution en hauteur de la population de Dialium guineense dans les forêts de Bonou et d'Itchèdè est asymétrique droite ou asymétrique positive. Les classes modales dans les forêts de Bonou et d'Itchèdè sont respectivement 8 à 14 m et 16 à 20 m. Les sujets de hauteur supérieur à 20 m sont faiblement représentés dans la forêt de Bonou et ceux supérieur à 24 m dans la forêt d'Itchèdè.
Densité (arbre/ha) 3,5 2,5 0,5 1,5 3 2 0 1 Classe de hauteur (m) Bonou Densité observée Weibull a = 1,759 b = 13,26 c = a
Densité (arbre/ha) 0,8 0,6 0,4 0,2 1,6 1,4 1,2 0 1 4--8 8--12 12--16 16--20 20--24 >24 Classe de hauteur (m) Itchèdè Densité observée Weibull a = 4 b = 11,97 c = 1,709
26 Figure 16 : Structure en hauteur de D. guineense dans la forêt de Bonou Figure 17 : Structure en hauteur de D. guineense dans la forêt d'Itchèdè 4.3.4. Répartition spatialeD'après la méthode de densité relative de voisinage, on déduit trois classes la densité relative de voisinage dans chaque anneau pour lesquelles ? prend les valeurs suivantes : ?>1 ; ?=1 et ? <1. La proportion de chaque classe par rapport au nombre total de placeaux inventoriés et suivant les catégories d'anneau est consignée dans le tableau 8 Tableau 8 : Densité relative de voisinage par catégorie d'anneau de Dialium guineense dans les forêts de Bonou et d'Itchèdè
Le tableau 8 montre que sur un rayon de 10m autour d'un pied de Dialium guineense, la densité relative de voisinage de ce dernier est supérieure à 1 dans 81,81% (soit 9 sur 11 observations) des placeaux inventoriés à Bonou. A Itchèdè, dans 70% des cas, la densité relative de voisinage de Dialium guineense est supérieure à 1 (soit 14 sur 20 observations). Nous pouvons conclure que sur un rayon de 10 m, la distribution de Dialium guineense est agrégative (?>1). Sur les anneaux de 10 à 20 m, 81,81% (9 sur 11 observations) des placeaux à Bonou et 90% (18 sur 20 observations) à Itchèdè ont une densité relative de voisinage de Dialium guineense inférieure à 1. La répartition spatiale de Dialium guineense sur les anneaux de 10 à 20
27 m est relativement dispersée. Sur un anneau de 20 à 30 m dans l'ensemble des placeaux, quelque soit la forêt, la densité relative de voisinage de Dialium guineense est inférieur à 1. La population de Dialium guineense est donc relativement au-delà d'un rayon de 20 m jusqu'à 30 m. 4.3.5. Densité de régénérationLa densité moyenne globale de régénération (tableau 9) est estimée à 57,57 tiges/ha pour Dialium guineense dans la forêt de Bonou alors qu'elle est de 58,33 tiges/ha dans la forêt d'Itchèdè. Comme dans le cas de Cola millenii, la densité de régénération va décroissant suivant la classe d'âge. Dans la forêt de Bonou la densité de régénération est de 42,42 tiges/ha au niveau des plantules (dbh<10 mm), 12,12 tiges/ha au niveau des juvéniles (10 mm = dbh<50 mm) et de 3,03 tiges/ha au niveau des petites perches (50mm = dbh<100 mm). Dans la forêt d'Itchèdè, la densité de régénération est de 35 tiges/ha au niveau des plantules, 20 tiges/ha au niveau des juvéniles et de 3,33 tiges/ha au niveau des petites perches. Tableau 9 : Densité moyenne de régénération de Dialium guineense dans les forêts de Bonou et d'Itchèdè
4.4. Caractéristiques structurales et écologiques des populations de Afzelia africana4.4.1. Paramètres dendrométriquesLes paramètres dendrométriques des populations de Afzelia africana dans les différentes formations sont présentés au tableau 10. On note que la densité moyenne des arbres sur pied (N), le diamètre de l'arbre de surface terrière moyenne (Dg), la surface terrière (G) et la hauteur moyenne de Lorey (HL) présentent tous des différences non significatives entre les deux forêts (Prob > 0,05).
28 Tableau 10 : Paramètres dendrométriques de Afzelia africana au sein des forêts de Bonou et d'Itchèdè
Moy : moyenne 4.4.2. Structure en diamètreLes résultats d'analyse log-linéaire effectuée sur la structure en diamètre de Afzelia africana indiquent un bon ajustement des données à la distribution de Weibull (Prob = 0,86 à Bonou et 0,79 à Itchèdè) au seuil de 5%. On note des figures 18 et 19 que la structure en diamètre de la population de Afzelia africana dans les deux forêts présente une distribution irrégulière en fonction des classes de diamètre. On note une prédominance d'individus de Afzelia africana de petits diamètres ; ceci est beaucoup plus visible dans la forêt de Bonou. Les sujets dont le diamètre est compris entre 10 cm et 20 cm sont les plus nombreux (à l'hectare) dans la forêt de Bonou. Les sujets de diamètre supérieur à 60 cm sont quasi absents dans les différentes forêts.
Itchèdè 10--20 20--30 30--40 40--50 50--60 60--70 70--80 80--90 >90 Densité (arbre/ha) 0,45 0,35 0,25 0,15 0,05 0,4 0,3 0,2 0,1 0 Densité observée Weibull a = 10 b = 53,71 c = 2,274 Classe de diamètre (Cm) Bonou 2 Densité observée Weibull a = 10 b = 18,84 c = 1,126 0 1,5 1 Densité (arbre/ha) 0,5 10--20 20--30 30--40 40--50 >50 Classe de diamètre (Cm) Figure 18 : Structure en diamètre de A. africana dans la forêt de Bonou Figure 19 : Structure en diamètre de A. africana dans la forêt d'Itchèdè 4.4.3. Structure en hauteur
29 On remarque en analysant la structure en hauteur des deux forêts que la différence entre les deux distributions n'est pas significative (Prob = 0,92 à Bonou et 0,99 à Itchèdè) et indique globalement une adéquation entre la structure observée et celle attendue sur la base de la distribution de Weibull. L'analyse des figures 20 et 21 indique de façon générale une allure en cloche pour la structure en diamètre de la population de Afzelia africana dans la forêt d'Itchèdè. Le paramètre de forme (c) est égal à 1,147 dans la forêt de Bonou et 2,117 dans la forêt d'Itchèdè. Le paramètre c est compris entre 1 et 3,6 quelque soit la forêt. Il est donc évident que la distribution en hauteur de la population de Afzelia africana dans les forêts de Bonou et d'Itchèdè est asymétrique droite ou asymétrique positive. Les classes modales dans les forêts de Bonou et d'Itchèdè sont de 24 m à 28 m.
0,8 0,7 0,6 0,5 0,4 0,3 0,2 0,1 0 Densité (arbre/ha) Densité observée Weibull a = 4 b = 14,03 c = 1,147 Bonou 4 8 8 12 12 16 16 20 20 24 24-28 >28 Classe de hauteur (m) Itchèdè 4--12 12--20 20--28 28--36 >36 Densité (arbre/ha) 0,7 0,6 0,5 0,4 0,3 0,2 0,1 0 Densité observée Weibull a = 4 b = 23,96 c =2,117 Classe de hauteur (m) Figure 20 : Structure en hauteur de A. africana dans la forêt de Bonou Figure 21 : Structure en hauteur de A. africana dans la forêt d'Itchèdè 4.4.4. Répartition spatialeD'après la méthode de densité relative de voisinage que nous avons utilisée, on déduit trois classes de densité relative de voisinage dans chaque anneau pour lesquelles ? prend les valeurs suivantes : ? >1 ; ? =1 et ? <1. La proportion de chaque classe par rapport au nombre totale de placeau inventorié et suivant les catégories d'anneau est consignée dans le tableau 11.
30 Tableau 11 : Densité relative de voisinage par catégorie d'anneau de Dialium guineense dans les forêts de Bonou et d'Itchèdè
Le tableau 11 montre que sur un rayon de 10m autour d'un pied de Afzelia africana, la densité relative de voisinage de ce dernier est supérieure à 1 dans 54,54% (soit 6 sur 11 observations) des placeaux inventoriés à Bonou. A Itchèdè, dans 30% des cas, la densité relative de voisinage de Afzelia africana est supérieure à 1 (soit 6 sur 20 observations) contre 70% des cas où la densité relative de voisinage est inférieure à 1. Nous pouvons conclure qu'à Bonou, sur un rayon de 10 m, la distribution de Afzelia africana est agrégative (?>1) mais tend vers l'hyperdispersion puisque le nombre de cas où la densité relative de voisinage de Afzelia africana n'est pas négligeable. Par contre à Itchèdè Afzelia africana, est relativement dispersée. Au-delà d'un rayon de 10 m autour d'un pied de Afzelia africana plus précisément sur les anneaux de 10 à 20 m et de 20 à 30 m la totalité des placeaux présente une densité relative de voisinage relatif à Afzelia africana inférieur à 1, quelle que soit la forêt. Sur les bandes de 10 à 20 m et de 20 à 30 m Afzelia africana est relativement dispersée. 4.4.5. Densité de régénérationLa densité moyenne globale de régénération (tableau 12) est estimée à 57,57 tiges/ha pour Afzelia africana dans la forêt de Bonou alors qu'elle est de 16,66 tiges/ha dans la forêt d'Itchèdè. Comme dans le cas de Cola millenii et de Dialium guineense, la densité de régénération va décroissant suivant la classe d'âge. Dans la forêt de Bonou, la densité de régénération est de 51,51 tiges/ha au niveau des plantules (dbh<10 mm), 6,06 tiges/ha au niveau des juvéniles (10 mm = dbh<50 mm) et nulle au niveau des petites perches (50mm = dbh<100 mm). Dans la forêt d'Itchèdè, la densité de régénération est de 10 tiges/ha au niveau des plantules, 6,67 tiges/ha au niveau des juvéniles et nulle au niveau des petites perches. Tableau 12 : Densité moyenne de régénération de A. africana dans les forêts de Bonou et d'Itchèdè
31
32 5. Discussion5.1. Caractéristiques dendrométriques des forêts étudiéesLa densité moyenne globale présente une différence significative entre les forêts de Bonou et d'Itchèdè. La densité moyenne globale de la forêt classée de Bonou (316 tiges/ha) est relativement plus élevée que celle de la forêt classée d'Itchèdè (171 tiges/ha). Comita et al. (2007) trouvent que plus le nombre d'individus reproducteurs est élevé, plus la densité globale est élevé. La valeur de densité relativement faible observée dans la forêt classée d'itchèdè pourrait s'expliquer par une faible présence d'individus reproducteurs due aux exploitations illicites organisées par les populations riveraines de cette forêt. La densité moyenne des populations de Cola millenii, de Dialium guineense et de Afzelia africana, présente une différence non significative entre les forêts classées de Bonou et d'Itchèdè. La densité moyenne de Cola millenii dans la forêt classée de Bonou (4,18 tiges/ha) est très faible par rapport celle dans la forêt d'Itchèdè (11 tiges/ha). Etant donné que la densité globale de régénération de la forêt de Bonou (254 tiges/ha) est relativement plus élevée que celle de la forêt d'Itchèdè (153 tiges/ha), la différence de densité ainsi constatée entre les deux forêts serait liée à l'insuffisance de la lumière qui entraverait la survie de l'espèce dans le sous-bois (Gourlet-Fleury, 1998). Contrairement au Cola millenii, la densité moyenne de Dialium guineense dans la forêt de Bonou (8 tiges/ha) est relativement supérieure à celle dans la forêt d'Itchèdè (6 tiges/ha). Ceci est d'autant plus normal que la forêt de Bonou est moins dégradée que la forêt d'Itchèdè. La densité moyenne de Afzelia africana est relativement plus élevée dans la forêt classée de Bonou (2,54 tiges/ha) que dans celle d'Itchèdè (1,8 tiges/ha), ce qui est comparable à la densité moyenne (2,8 tiges/ha) trouvée par Bonou (2007) dans la forêt de Lama. La différence de densité entre les deux forêts est due au faite que la forêt de Bonou est moins dégradée que la forêt d'Itchèdè. Mais, il est à remarquer que la densité moyenne de Afzelia africana est relativement faible dans les deux forêts. Cela se justifie par le fait que le recrutement des populations de l'espèce est difficile (Sokpon et Biaou, 2002). De plus, l'espèce a un faible pouvoir de rejet, de sorte que la destruction de la partie aérienne (par les feux ou par les animaux) entraîne la mort chez la plupart des individus de l'espèce (Bationo et al., 2001). Selon Hubbell et Foster (1986) cités par Herrero-Jáuregui (2012), une espèce peut être considérée comme rare en forêt lorsque sa densité est inférieure à un individu par ha. Plusieurs espèces commerciales répondent à ce critère (Schulze et al., 2008, cité par Herrero-Jáuregui, 2012). D'après nos résultats, aucune des espèces de valeur étudiées ne peut être considérée comme rare dans notre zone d'étude.
33 La structure d'un peuplement est le reflet d'un dépassement en hauteur, d'une suppression et d'un encombrement des individus les plus faibles par les individus les plus vigoureux (Rondeux, 1999). Etant donné que le couvert est beaucoup plus dense dans la forêt de Bonou, les individus de Cola millenii ne possédant pas une grande capacité de dominance se trouvent ralentis dans leur croissance (Brower et al., 1990). Ce qui fait qu'on note un diamètre moyen et une hauteur moyenne de Lorey relativement plus faible dans la forêt de Bonou que dans la forêt d'Itchèdè. Aussi, cette relative densité de la forêt de Bonou limite la croissance en diamètre et en hauteur de Afzelia africana. Salomon (1998) décrit ce phénomène dans son étude sur la structure spatiale d'une population de Spirotropis longifolia en forêt guyanaise et conclut que ces espèces ont des diamètres quadratiques significativement petits, tandis que les autres espèces voisines ont des diamètres importants. Ces derniers semblent se maintenir en canopée. Ils sont alors autant de « voisins gênants » compétitifs, en termes d'accès à la lumière (Gourlet-Fleury 1998). 5.2. Structure en diamètre et en hauteurLa forme classique de la structure d'un peuplement inéquienne ou multispécifique est caractérisée par une fréquence élevée de jeunes individus dans les petites classes de diamètre et une diminution progressive des individus au fur et à mesure que le diamètre devient grand (Glèlè Kakaï, 2010). L'analyse de la structure en diamètre du peuplement arborescent des forêts classées de Bonou et d'Itchèdè révèle une distribution exponentielle négative caractérisée par plus d'individus de petits diamètres et moins d'individus de gros diamètre. Cette structure est régressive. Cette forme de distribution de diamètre rejoint celle des forêts denses tropicales humides (Rollet, 1974). Dans les peuplements inéquiennes, les arbres présentent souvent des canopées différentes constituant des strates (Olivier & Larson, 1996). La distribution en hauteur devrait être similaire à la distribution en diamètre dans ce type de peuplement, mais moins variable que cette dernière. Les forêts classées de Bonou et d'Itchèdè présentent une distribution en hauteur exponentielle négative, avec plus d'arbre de petites tailles que de grandes tailles. Mais, la distribution en hauteur des arbres d'une formation naturelle présente globalement une forme gaussienne qui peut être asymétrique selon les conditions de vie du peuplement (Glele Kakaï, 2010), ce qui n'est pas le cas dans les forêts classées de Bonou et d'Itchèdè. Les forêts de Bonou et d'Itchèdè possèdent donc une bonne capacité de régénération. Cola millenii dans les forêts de Bonou et d'Itchèdè présente une distribution en diamètre comme en hauteur de forme exponentielle décroissante ou en diminution régulière des effectifs avec les classes de diamètre et de hauteur. Selon Pascal (2003), ce type de distributions
34 est caractéristique des espèces sciaphiles, tolérant l'ombre. On la retrouve aussi bien chez les espèces qui passent tout leur cycle en sous-bois (les diamètres maximaux sont alors peu importants) que chez celles qui commencent leur développement en sous-bois et le terminent au niveau de la canopée, atteignant ainsi de forts diamètres (Pascal, 2003). Dialium guineense a également une distribution en diamètre de forme exponentielle décroissante dans les deux forêts. Cela se justifie par le fait que l'espèce tolère l'ombrage. Par contre Dialium guineense dans les deux forêts étudiées présente une distribution en hauteur en forme de cloche et on note une décroissance des effectifs quand la hauteur augmente même si cette décroissance est faible dans la forêt d'Itchèdè. La structure en forme de « J renversé » est la juxtaposition d'un certain nombre de courbes en cloche formée à partir des événements de régénération individuels et réguliers pour les espèces adaptées à leurs conditions stationnelles. En forêts naturelles, la structure en forme de cloche typique est due à une régénération sporadique, du fait de la non-adaptation des espèces aux conditions stationnelles (Geldenhuys, 2010). La structure en diamètre et en hauteur de Afzelia africana dans la forêt de Bonou présente une structure en « J renversé », caractérisée par un plus grand nombre de petits arbres que de grands arbres, et une réduction progressive du nombre d'arbres d'une classe à la suivante. En s'intéressant à la structure en diamètre et en hauteur de Afzelia africana dans la forêt d'Itchèdè, on remarque qu'elle est caractérisée par une distribution en cloche et par un effectif plus important des individus de classes de diamètres intermédiaires. Ces différences entre classes de diamètre pourraient être dues à des irrégularités de fructification de l'espèce. Ceci peut aussi s'expliquer par la croissance différentielle de certains pieds d'arbres et à la concurrence dans les groupes du fait des mauvaises conditions du milieu pour la régénération (Geldenhuys, 2010). Cela entraîne donc, une irrégularité de la succession des vagues de régénération (Geldenhuys, 2010). Ce qui peut s'expliquer par une interruption temporaire de la régénération (conditions du milieu devenant défavorables) du fait d'une récolte excessive, de dommages directs aux semis ou de l'absence d'agents de pollinisation ou de dissémination (Peters, 1994). De telles distributions ont été observées par Sokpon & Biaou (2002) dans la forêt classée de Bassila, Sinsin et al. (2004) dans les différentes zones climatiques du Bénin, notamment chez Afzelia africana. Les espèces fidèles à ce type de distribution sont dites déstructurantes car menacées de disparition dans les peuplements (Sokpon & Biaou, 2002). Geldenhuys (2010) fait des observations similaires en parlant de croissance différentielle et de mortalité des jeunes tiges due à la concurrence entre les individus et aux mauvaises conditions du milieu. Ces constats sont identiques à ceux effectuées par Sinasson (2010) dans
35 les forêts naturelles d'Itchèdè-Toffo (Sud-Bénin) et Hounkpèvi (2010) dans les groupements végétaux des secteurs forestiers de Massi et Koto de la forêt classée de la Lama (Sud et centre-Bénin). 5.3. Répartition spatialeL'indice de Blackman et de Green présentent l'inconvénient de décrire la structure spatiale à l'échelle du plus proche voisin, les interactions au-delà du plus proche voisin sont ignorées (Stoyan & Penttinen, 2000). Pour pallier à cette difficulté, nous avons utilisé pour étudier la répartition spatiale des espèces de valeur, la densité relative de voisinage ? (Condit et al, 2000) qui parait être une meilleure méthode. La densité relative de voisinage est un indice qui n'exprime pas la structure des peuplements seulement en termes de valeurs moyennes ou de distribution, mais qui décrit la structure spatiale de manière continue. Cet indice peut être comparé à la fonction K de Ripley mais n'est pas une fonction cumulative de la distance d'observation comme celle de Ripley (Condit et al., 2000). La densité relative de voisinage permet des investigations spatialement explicites des interactions entre les arbres sur de grandes échelles de distance. A cet effet, la densité relative de voisinage indique que dans un rayon de 10 m la distribution de Cola millenii et de Dialium guineense est agrégative dans les deux forêts étudiées. Au-delà d'un rayon de 10m plus précisément entre 10 m et 30 m, les individus de ces deux espèces sont relativement dispersés, quelle que soit la forêt. La population de Afzelia africana dans la forêt de Bonou a une distribution agrégative mais tendant vers une dispersion. Par contre à Itchèdè, Afzelia africana dans un rayon de 10 m est relativement dispersés. Au-delà d'un rayon de 10 m plus précisément entre 10 m et 30 m les individus de Afzelia africana sont relativement dispersés, quelle que soit la forêt considérée. Ces résultats sont différents de ceux de Bonou et al. (2009) sur la caractérisation structurale des formations végétales hébergeant Afzelia africana dans la forêt classée de la Lama au Sud du Bénin. Bonou et al. (2009) trouvent que les valeurs des indices de Blackman et de Green indiquent un regroupement très faible des individus de l'espèce. Cependant, il note que quelques regroupements des individus sont parfois observés mais sur de faibles étendues généralement de moins d'un quart d'hectare. Nos résultats se rapprochent de ceux trouvés par Fandohan (2006) dans la forêt classée de Wari-Maro qui a noté une répartition aléatoire avec une tendance à l'agrégation pour de faibles rayons (30 m) autour de tout point arbitrairement fixé au sein de la population. Le développement des plantes dans l'espace est à la base des phénomènes fondamentaux que sont la régénération forestière, et l'extension des écosystèmes forestiers.
36 Mais les modalités de la dissémination des graines sont certainement parmi les facteurs décisifs qui règlent le comportement d'une espèce au sein d'un groupement végétal (Comita et al., 2007 ; Nishimura et al., 2008 ; Mussenac, 2009). Les stratégies de dispersion des graines adoptée par une plante peuvent être liées à l'agent de dispersion. Ainsi, Cola millenii et Dialium guineense sont des espèces sarcochore (Diaspores totalement ou partiellement charnues). Ils ont généralement besoin des animaux en particulier les frugivores pour se disséminer (zoochorie), ce qui explique la raison pour laquelle on retrouve des individus de ces espèce au-delà de 10 m. La structure spatiale agrégative observée chez ces deux espèces peut être liée à une faible activité des agents de dispersion. Afzelia africana par contre est une espèce barochore (Diaspore non charnue, lourde). Ses graines ne se disséminent pas par nature sur de longue distance. A cause de sa faible distance de dispersion, les graines se retrouvent en agrégats autour de l'arbre « mère » (Menaut et al., 1990; Collinet, 1997; Barot et al., 1999). Nishimura (2008) en étudiant la répartition spatiale et écologique des Fagaceae dans la forêt de Sumatra (Indonésie), remarque qu'une espèce peut avoir sur une distance donnée une première distribution qui est agrégative et sur une distance plus longue grâce aux agents de dispersion une deuxième distribution qui est aléatoire. La dissémination des graines par les animaux permet d'augmenter la probabilité que les espèces colonisent des milieux plus favorables (Thompson et Willson (1978) cité par Nishimura et al. (2008)). Si la distribution agrégative est donc observée dans la forêt de Bonou et pas dans celle d'Itchèdè, c'est parce que les conditions stationnelles ne s'y prêtent pas. La forêt d'Itchèdè dans un état de dégradation poussée n'offre plus l'écosystème favorable à Afzelia africana. Mais pour que Afzelia africana se retrouve sur une distance supérieure à 10 m, il a fallu l'action disséminateur des animaux, rongeurs et herbivores en particulier. 5.4. Contraintes sylvicoles et recommandations pour une gestion durable des forêtsLes différentes interprétations de la structure spatiale et écologique des forêts de Bonou et d'Itchèdè et des espèces de valeur (Cola millenii, Dialium guineense et Afzelia africana), nous permettent de faire des diagnostics et de proposer des mesures sylvicoles à prendre pour une gestion durable des différentes formations étudiées. En effet, la densité moyenne relativement faible des espèces de valeur étudiées ne garantit pas un recrutement continu dans les forêts de Bonou et d'Itchèdè. Spécifiquement Afzelia africana est faiblement représenté dans les deux forêts et est menacée de disparition dans la forêt d'Itchèdè. Pour restaurer ces espèces de valeur, la régénération naturelle doit être assistée
37 à travers le repérage des semis et le dégagement périodique des adventices tout autour. Le peuplement des forêts doit être ouvert en desserrant les arbres (éclaircies sélectives) pour permettre à la lumière de pénétrer dans le sous-bois afin de favoriser la germination et le développement des semis (Geldenhys, 2010). Pour cela, on pourra supprimer aux endroits denses des forêts, les arbres ayant atteint le diamètre moyen d'exploitabilité tout en préservant un minimum de densité pour servir de semenciers. Il faut faire également des enrichissements des forêts surtout en Afzelia africana avec des plants produits en pépinière. Dans ce cas, les endroits convenables sont les troués, les zones de reconstitution lente de la forêt afin de permettre la croissance adéquate de la régénération. Il faut envisager la protection des graines de Afzelia africana contre les déprédateurs et une assistance qui consisterait au moins à protéger les jeunes plants durant les premières années de leur croissance en faisant de la régénération assistée. Il s'agira par exemple d'identifier les plantules et de les entourer d'enclos inaccessibles aux prédateurs.
38 CoNcLusIoNLes formations naturelles comme les forêts Classées de Bonou et d'Itchèdè qui constituent encore des reliques forestières dans la région Sud du Bénin, méritent d'être suivies en permanence afin de maintenir leur structure spatiale et temporelle en un état comparable aux forêts denses semi-décidues. L'étude de la structure des populations de Cola millenii, de Dialium guineense et de Afzelia africana et de leur écologie contribue à une meilleure connaissance de ces espèces dans les formations naturelles du Sud-Bénin. Une telle étude constitue un préalable fondamental à l'élaboration de stratégies de conservation et de gestion durable des forêts à Cola millenii, Dialium guineense et Afzelia africana. Il ressort de cette étude que la densité moyenne globale est significativement différente au seuil de probabilité de 5% entre les deux forêts (316 pieds/ha dans la forêt de Bonou et 171 pieds/ha dans la forêt d'Itchèdè). Par contre, on ne note pas une différence entre la structure des deux forêts. Les structures en hauteur et en diamètre dans les deux forêts sont exponentielles négatives caractérisées par plus d'individus de petits diamètres et de petites tailles et moins d'individus de gros diamètre et de grande taille. Ceci montre globalement une bonne capacité de régénération de ces forêts. Quant aux espèces de valeur (Cola millenii, Dialium guineense et Afzelia africana) leurs paramètres dendrométriques ne présentent aucune différence significative entre les forêts de Bonou et d'Itchèdè. Les densités des individus (dbh?10cm) de ces espèces de valeur sont relativement faibles quelle que soit la forêt considérée, ce qui est indicateur d'un risque de disparition de ces espèces. Aussi la structure spatiale de ces espèces de valeur interpelle sur leur viabilité dans les deux forêts étudiées. Cola millenii dans les forêts de Bonou et d'Itchèdè présente une distribution de forme exponentielle décroissante caractérisée par une dominance des individus jeunes. Cela laisse suggérer que Cola millenii présente une bonne capacité de régénération contrairement au Dialium guineense qui a une distribution en hauteur en forme de cloche avec une décroissance des effectifs quand le diamètre augmente. Dialium guineense est donc confronté à des problèmes de régénération probablement dus aux conditions stationnelles qui ne lui sont pas favorables. Afzelia africana par contre à une distribution déstructurante et menacée de disparition dans la forêt d'Itchèdè, car cette espèce présente une structure en cloche avec un effectif plus important des individus de classes de diamètres intermédiaires. Ce qui n'est pas le cas à Bonou où Afzelia africana présente une structure en « J renversé », caractérisée par un plus grand nombre de petits arbres que de grands arbres. Il faut retenir également que les populations de Cola millenii et de Dialium guineense dans un rayon de 10m ont une répartition agrégative. Au-delà des 10m plus précisément entre 10
39 m et 30 m, cette répartition devient progressivement dispersée. Cela est constaté dans les deux forêts. Afzelia africana par contre observé dans la forêt d'Itchèdè a une distribution relativement dispersée sur un rayon de 30 m or dans la forêt de Bonou la distribution ne devient relativement dispersée qu'au-delà de 10m, si non avant cette distance elle est relativement agrégative. Cette différence entre les deux forêts dans un rayon de 10m autour d'un pied de Afzelia africana est due aux actions anthropiques beaucoup plus accentuées dans la forêt d'Itchèdè que celle de Bonou. D'une façon générale, les résultats obtenus suggèrent que les populations des espèces cibles présentent dans l'ensemble une structure peu stable et présentent donc à terme une propension à la régression si rien n'est fait pour assurer leur renouvellement dans les deux forêts. Il est donc important de mettre en oeuvre une régénération naturelle assistée et un enrichissement en espèces de valeur afin d'assurer la pérennité des services ecosystémiques forestiers. Par ailleurs, des soins culturaux tels que l'ouverture de la canopée, le délianage... sont indispensables pour garantir la survie des plantules dans les stades de développement de la forêt.
40 RÉFÉRENCES BIBLIOGRAPHIQUES Adjanohoun E. et al, 1989. Contribution aux études ethnobotaniques et floristiques en République du Bénin. Médecine traditionnelle et pharmacopée : 895 p. ACCT. Paris. France. Adomou C. A. 2005. Vegetation Patterns and Environmental gradients in Benin: implications for biogeography and conservation. PhD Thesis. Wageningen University, Pays-Bas. 133p. Ahouangonou, S., Bris, B., 1997. Afzelia africana. Le Flamboyant 42, 7-10. Akouègninou A. et al, 2006. Flore analytique du Bénin. 1063p Andriamalala F. et al, 2011. Outils pour la caractérisation des milieux forestiers et de leur diversité végétale Atindogbé G., 2004. Analyse de la structure spatiale des arbres : Application à l'enrichissement des savanes boisées de la zone soudanienne au Bénin. Thèse d'Ingénieur agronome, FSA/UAC. 97p Aubreville A., 1937. Les forêts du Dahomey et du Togo. Bulletin du comité d'études historique et scientifiques de l'Afrique occidentale française. Tome XX (1-2) : 1-112 Aussenac G., 1966. Observations sur la dissémination des graines chez abies grandis. Awokou K. S., Ganglo C. J., Azontondé H. A., Adjakidjè V. & De Foucault B., 2009 « Caractéristiques structurales et écologiques des phytocénoses forestières de la forêt classée d'Itchèdè (Département du Plateau, Sud-est Bénin) », Sciences & Nature Vol.6 N°2 : 125 - 138 Barot S., Gignoux J., Menaut J.C., 1999. Demography of a savanna palm tree: predictions from comprehensive spatial pattern analyses. Ecology 80 (6): 1987-2005. Bayer, W., Waters-Bayer, A., 1999. La gestion des fourrages. CTA, Wageningen. Bonou W., Glèlè Kakaï R., Assogbadjo A. E., Fonton H. N., Sinsin B., 2009. Characterisation of Afzelia africana Sm. habitat in the Lama Forest reserve of Benin. Forest Ecology and Management 258.1084-1092p. Brower et al, 1990. Methods for General Ecology. Third ed., Wm. C. Brown Publishers, Dubuque.
41 C/BADI., 2001. Atlas monographique des Communes du Bénin. Service Allemand de Développement. Chakirou, M. R., 1991. Les crues du fleuve Ouémé et leurs incidences biogéographiques dans la basse vallée entre les latitudes de Bonou et d'Adjohoun FLASH / UAC. Collinet F., 1997. Essais de regroupements des principales espèces structurantes d'une forêt dense humide d'après l'analyse de leur répartition spatiale (Forêt de Paracou-Guyane). Comita L. S., Condit R. Hubbell S. P., 2007. Developmental changes in habitat associations of tropical trees. Journal of Ecology 95, 482-492 Condit R., Ashton P. S., Baker P., Bunyavejchewin S., Gunatilleke S., Gunatilleke N., Hubbell S. P., Foster R. B., Itoh A., LaFrankie J. V., Lee H. S., Losos E., Manokaran N., Sukumar R., Yamakura T., 2000. Spatial Patterns in the Distribution of Tropical Tree Species, Sciences Vol.288: 1414-1417p Dadjo C. P. A. F., 2010. Caractérisation structurale et ethnobotanique de Vitex doniana sweet dans la forêt classée de la lama. Thèse d'ingénieur agronome, FSA/UAC. Dagnelie P., 1998. Statistique théorique et appliquée volume 2. Paris et Bruxelles, De Boeck et Larcier, 508-659 p. Dupuy B., 1998. Bases pour une sylviculture en forêt dense tropicale humide africaine. 328p. Fandohan B., 2006. Abondance et structure des peuplements de sept espèces ligneuses médicales dans la forêt classée de Wari-Maro (région des Monts Kouffé). Thèse d'ingénieur agronome, 109p. Gbaguidi, 1998. Forêts sacrées et conservation de la biodiversité dans le département de l'Ouémé au sud Est du Bénin. Thèse d'Ingénieur Agronome 161p FSA/ UAC. Bénin Geldenhuys CJ., 2010. Managing forest complexity through application of disturbance-recorvery knowledge in development of silvicultural systems and ecological rehabilitation in natural forest systems in Africa. Journal Forest Research, 15 : 3-13. Glele Kakaï R., Bonou W., Pelz D., 2010. Modélisation et interprétation des structures en diamètre et en hauteur des peuplements forestiers. 15p
42 Gnonlonfoun I., 2011. Effet combine de la spatialisation et des caractéristiques des placettes d'inventaire sur la précision de caractérisation structurale de la régénération en forêt dense: cas de la forêt classée de Lama au Sud- bénin. Thèse d'ingénieur agronome. 68p Goji T. C. & Ayodele A. E., 2006. Foliar epidermal and Pollen Characters in the genus Cola Schott. & Endl. in Nigeria. acta SATECH 2(2): 57-63 (2005) Goreaud F. et Pélissier R., 2000. Analyse de la structure spatiale de semis de points hétérogènes : exemples d'application à des peuplements forestiers. Gourlet-Fleury S., 1998. In JATBA (Journal d'Agronomie Tropicale et de Botanique Appliquée), revue d'ethnobiologie, vol. 40 (1-2), spécial Guyane : Conserver, gérer la biodiversité : quelle stratégie pour la Guyane ? p.p. 79-102. Herrero-Jáuregui C, García-Fernández C, Sist PL, Casado MA. 2012. Recruitment dynamics of two low-density neotropical multiple-use tree species. Plant Ecology, 212 (9): 1501-1512. Hitimana J, Kiyiapi JL, Njunge TJ. 2004. Forest structure characteristics in disturbed and undisturbed sites of Mt. Elgon Moist Lower Montane Forest, western Kenya. Department of Forestry, Moi University. Eldoret, Kenya. Houessou A., 1974. Etudes des formations détritiques de l'Ouest du bassin sédimentaire côtier du Dahomey. Houndonougbo J. S. H., 2011. Caractéristiques structurales et écologiques des populations de Triplochiton scleroxylon et Terminalia superba dans les forêts reliques du Sud-Benin. Thèse d'ingénieur agronome, FSA/UAC. 53p. Hounkpevi A. et al, 2010. Contribution à la gestion durable des forêts: caractéristiques écologiques et structurales de la forêt naturelle de Massi (Lama) au Sud du Bénin. Thèse d'ingénieur agronome, FSA/UAC. 137p. INSAE. 2003. Troisième recensement général de la population et de l'habitation (rgph-3). Résultats définitifs: caractéristiques générales de la population, Bénin. 404p. Kerharo & Adam g. J. (1974), La pharmacopée sénégalaise traditionnelle. Plantes médicinales et toxiques. Vigot Frères, Paris, 488.
43 Kiki J. M., 2005. Analyse de la structure spatiale des arbres des peuplements a dominance Isoberlinia spp du centre-bénin. Thèse d'ingénieur agronome, FSA/UAC. Lanly, J.-P., 1969. Forêts tropicales et inventaires. Revue Forestière Française [ISSN 00352829], 1969, Vol. 21, N° 6; p. 602-602. Mahugnon J., 2005. Analyse de la structure spatiale des arbres des peuplements à dominance Isoberlinia spp. Craib & stapf du centre-bénin. Thèse d'ingénieur agronome. 101p Marcon E., 2011. Mesures de la biodiversité. 37 p. McNeely J. A., 1996. Conservation and future: Trends and options toward the year 2025. A discussion paper. IUCN, Gland Switzerland.66p Menaut J.C., Gignoux J., Prado C., Clobert J., 1990. Tree community dynamics in a humid savanna of the Côte d'Ivoire: modelling the effects of fire and competition with grass and neighbours. Journal of Biogeography, 17:471-481 Morakinyo J. A., 1995. Mitosis in Cola lepidota, Cola millenii and Sterculia tragacantha. Bioscience Research communications Vol. 7, N°2. N'dri, M.T.K.; Gnahoua, M.G.; Konan, E.K.; Traoré, D., 2008. Plantes alimentaires spontanées de la région du Fromager (Centre Ouest de la Côte d'Ivoire) flore, habitats et organes consommés. Sci. & Nat. 5, 61-70. Nishimura S, Yoneda T, Fujii S, Mukhtar E, Kanzaki M. 2008. Spatial patterns and habitat associations of Fagaceae in a hill dipterocarp forest in Ulu Gadut, West Sumatra. Journal of Tropical Ecology. 24: 535-550. Oliver C.D. and Larson B.C., 1996. Forest stand dynamics. John Wiley and Sons, Inc., 520 p. Onana J. 1998. Sur l'utilisation et la régénération naturelle de Afzelia africana Smith ex Person. Le Flamboyant n° 46 Juin 1998 :33. Pascal JP., 2003. Dynamique spatiale de vouacapoua americana (aublet), arbre de foret tropicale humide a répartition agrégée. Thèse de doctorat. 230p Pascal JP., 2003. Notions sur les structures et dynamiques des forêts tropicales humides. Rev. For. Fr. LV - numéro spécial 2003
44 Peters, C.M. 1994. Exploitation soutenue des Produits Forestiers Autres que le Bois d'oeuvre en forêt tropicale humide : Manuel d'initiation écologique, série générale du programme d'appui à la biodiversité, Washington. No2, 49p. Philip M.S., 2002. Measuring Trees and Forests, 2nd ed. CABI, London. Projet Bois de Feu, 1999. Plan d'aménagement de la forêt classée d'Itchèdè-Tofo. 51 p. Rollet B., 1974, L'architecture des forêts denses humides sempervirentes de plaines. C.T.F.T., Paris, France. 298 p. Rondeux J., 1999. La mesure des peuplements forestiers. Presses agronomiques de Gembloux. 522p. Sagbo P. A., 2000. Etude des caractéristiques dendrométriques des peuplements naturels à dominance Isoberlinia spp : Cas de la Forêt Classée de l'Ouémé Supérieur au Nord du Bénin. Thèse d'ingénieur agronome, FSA/UAC. 93 p. Salomon L., 1998. Structuration spatiale d'une population de Spirotropis longifolia (DC.) Baill. (Leguminosae-Papilionoideae): Cas de monodominance en forêt guyanaise. 15p. Sinasson S. K., 2010. Dynamique des forêts naturelles de Bonou et d'Itchèdè-Toffo, Sud-Bénin. Thèse de Diplôme d'Etude Approfondie (DEA), FSA/UAC. 143p. Sinsin B., 2003. Dynamique des plantations d'enrichissement des galeries forestières de la région de Bassila au Bénin. P 13-26. In Sokpon, N., Sinsin, B. et Eyog Matig, O. (Eds). Acte du IIè séminaire international sur l'aménagement intégré des forêts naturelles des zones tropicales sèches en Afrique de l'Ouest, Parakou, Bénin, 25-29juin. 410p. Sinsin, B., 1993. Phytosociologie, écologie valeur pastorale, productivité et capacité de charge des pâturages naturels des périmètres Nikki-kalalé au Nord-Bénin. Thèse de Doctorat. Université Libre de Bruxelles. Belgique 390p. Sinsin, B. et al, 1997. Faune sauvage des forêts classées de Goungoun, de la Sota et des Environs. Directions des forêts et des Ressources Naturelles, Projet UNSO/BEN/93/G31. Cotonou.
45 Sinsin, B., Eyog Matig, O., Assogbadjo, A.E., Gaoué, O.G., Sinadouwirou, T., 2004. Dendrometric characteristics as indicators of pressure of Afzelia africana Sm. trees dynamics in different climatic zones of Benin. Biodiversity and Conservation 13(8), 1555-1570. Sinsin, B., Eyog Matig, O., Sinadouwirou, T., Assogbadjo, A.E., 2002. Caractérisation écologique des essences fourragères Khaya senegalensis Desr. et Afzelia africana Sm. Suivant les gradients de latitude et de station au Bénin, In: Eyog Matig, O., Gaoue, O. G., Obel-Lawson (Eds.), Development of appropriate conservation strategies for African forest trees identified as priority species by SAFORGEN member countries, SAFORGEN, 15-50. Sokpon N., 2003. Aménagement des forêts denses sèches du Nord Bénin: Bases techniques pour une gestion durable. Pp 140-158. In Sokpon, N., Sinsin, B. et Eyog Matig, O. (Eds). Acte du IIè séminaire international sur l'aménagement intégré des forêts naturelles des zones tropicales sèches en Afrique de l'Ouest, Parakou, Bénin, 25-29juin. 410p. Sokpon N., et BIAOU H. S., 2002. The use of diameter distribution in sustained-use management of remnant forests in Benin: case of Bassila forest reserve in North Benin. Forest Ecology and Management, 161 : 13-25. Stoyan, D. & Penttinen, A. 2000. Recent applications of point process methods in forestry statistics. Forest Science 15(1): 61-78. Tohngodo B., 2004. Phytosociologie appliquée à l'aménagement de la forêt classée de Bonou. Thèse d'ingénieur agronome. Faculté des Sciences Agronomiques. Université d'Abomey-Calavi, 209p. UICN. 2008. The UICN Red list of Threatened Species. www.iucnredlist.org update of 2008. Volkoff B. & Willalme P., 1976. Carte pédologique de reconnaissance de la République Populaire du Bénin au 1/200000. Feuille Porto-Novo (1). Notice Explicative n° 66 - ORSTOM - Paris, 39p. Multigr. Yehouenou Tessi D. R..,2011. Caractéristiques structurales et écologiques des populations d'Antiaris toxicaria (Pers) Lesh et Ceiba pentandra (L.) Gaertn dans les forêts reliques du Sud-Bénin. Thèse d'ingénieur agronome, FSA/UAC.
46 ANNEXESAnnexe 1 : Fiche d'inventaire (Mesure dendrométrique)Nom de la forêt Date Code placeau Pente Latitude Longitude .
Annexe 2 : Fiche d'inventaire (dénombrement des individus).
C = Circonférence Annexe 3 : Fiche d'inventaire des espèces de valeurNom de la forêt Date Code placeau Pente Latitude Longitude
Annexe 4 : Résultats d'analyse log-linéaire d'ajustement à la distribution de Weibull : Annexe 4a: Distribution en diamètre des forêts de Bonou et d'Itchèdè Forêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Annexe 4b : Distribution en hauteur des forêts de Bonou et d'ItchèdèForêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Annexe 4c : Distribution en diamètre de Cola millenii dans les forêts de Bonou et d'Itchèdè Forêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Annexe 4d : Distribution en hauteur de Cola millenii dans les forêts de Bonou et d'ItchèdèForêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Annexe 4e : Distribution en diamètre de Dialium guineense dans les forêts de Bonou et d'Itchèdè Forêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Forêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Annexe 4f : Distribution en hauteur de Dialium guineense dans les forêts de Bonou et d'Itchèdè Annexe 4g : Distribution en diamètre de Afzelia africana dans les forêts de Bonou et d'Itchèdè Forêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Forêt de Bonou Maximum Likelihood Analysis of Variance
Forêt d'Itchèdè Maximum Likelihood Analysis of Variance
Annexe 4h : Distribution en hauteur de Afzelia africana dans les forêts de Bonou et d'Itchèdè Annexe 5 : Résultats des tests de comparaison des paramètres dendrométriquesAnnexe 5a : Résultats des tests de comparaison des paramètres dendrométriques des forêts de Bonou et d'Itchèdè
Annexe 5b : Résultats des tests de comparaison des paramètres dendrométriques des populations de Cola millenii dans les forêts de Bonou et d'Itchèdè
Annexe 5c : Résultats des tests de comparaison des paramètres dendrométriques des populations de Dialium guineense dans les forêts de Bonou et d'Itchèdè
Annexe 5d : Résultats des tests de comparaison des paramètres dendrométriques des populations de Afzelia africana dans les forêts de Bonou et d'Itchèdè
Annexe 6 : Liste des espèces inventoriées dans les forêts de Bonou et d'Itchèdè
| 
Changeons ce systeme injuste, Soyez votre propre syndic 
"Là où il n'y a pas d'espoir, nous devons l'inventer" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||