|
Université de Paris-Sud XI
UFR STAPS
Laboratoire Contrôle Moteur et
Perception
UPRES EA 4042
Mémoire de recherche
pour
l'obtention du Master 2e année
en Sciences de la Motricité
Spécialité : Contrôle Moteur
Effet de la fatigue sur la variation des
paramètres
mécaniques et métaboliques au cours de
répétitions
de 100m sprint
Présenté par :
Mohamed-Amine
CHOUKOU
Sous la Codirection de :
Guillaume Laffaye et Serge
Le Bo2ec
2008-2009
Plan
1. Introduction
1
2. Partie
théorique
3
2.1. La performance en course de vitesse
2.1.1. La phase d'accélération initiale
2.1.2. La phase de course à vitesse maximale
2.1.3. La phase de décélération
2.2. La modélisation de la raideur
mécanique au cours de la course : rêve et
choix 6
2.2.1. Le modèle symétrique 2.2.2. Le
modèle asymétrique
2.3. Les méthodes de calcul de la raideur
mécanique 7
2.3.1. La méthode analytique
2.3.2. La méthode sinusoïdale
2.4. Les paramètres mécaniques du
SMM 9
2.4.1. La force musculaire
2.4.2. La vitesse de la course
2.4.3. La raideur mécanique
2.5. La fatigue
musculaire 11
2.5.1. Effet de la fatigue sur la
performance
2.5.2. Modèle de la fatigue
2.5.2.1. Modèle
énergétique/déplétion des réserves
énergétiques
2.5.2.2. Modèle neuromusculaire
2.5.2.3. Modèle biomécanique
2.5.2.4. Modèle complexe
Hypothèses
15
3. Méthodologie
........16
3.1. Population
3.2. Protocol expérimental
3.3. Modélisation de la course
3.4. Variables étudiées
3.5. Mesure des paramètres cinétiques
3.6. Observations de la cinématique de la course
3.7. Mesure de la lactatémie
3.8. Analyse statistique
4. Résultats
.....22
5. Discussion
......30
6. Conclusion
.....33
Bibliographie
........36
Annexes
39
1. Introduction
Le sprint regroupe les courses à pied dans lesquelles
le sprinter doit courir sur une faible distance à la vitesse la plus
rapide possible. Le 100 mètres (100m) est un sprint court parcouru en
ligne droite. Le record actuel de la « distance reine » est
détenu par le jamaïcain Usain Bolt 9.69s, selon le site officiel du
mouvement olympique (16/08/2008, Jeux Olympiques de Pékin).
C'est-à-dire que l'homme le plus rapide de la planète parcoure le
100m à une vitesse moyenne de 10.3 1 m.s-1.
Le 100m sprint est reparti en 3 phases ; successivement
l'accélération initiale, la course à vitesse maximale et
la décélération (voir figure n°1). Un tel effort
intense et de courte durée nécessite des sources
énergétiques immédiatement disponibles
(Phosphocréatine) permettant une accélération maximale
lors de la première phase de la course. L'effort est supramaximal au
cours de la deuxième phase de la course. Il permet d'atteindre et de
maintenir la vitesse maximale. Ce rendement mécanique ne peut pas
être enduré jusqu'à la fin du 100m, ce qui explique la
décélération.

Figure n°1 : Les trois phases du 100m
sprint
La diminution de la vitesse de course entraîne des
modifications des différents paramètres mécaniques et
énergétiques du 100m. Cette étude traitera les causes
possibles de la diminution de la performance avec la fatigue
induite par la répétition de sprint. Cette dernière est
habituellement utilisée dans la programmation des séances de
sprint. Quelles sont les causes de la fatigue ? Quelles sont les
conséquences sur la performance ? Comment l'athlète s'y
adaptera-t-il ?
2. Partie théorique
2.1. La performance en course de vitesse
Lors d'une compétition de sprint, l'athlète doit
courir sur une faible distance le plus rapidement possible. Le 100m sprint
requiert des ressources anaérobies pour produire l'énergie
nécessaire à la contraction musculaire dans un délai aussi
bref que le temps écroulé au 100m. Les réactions chimiques
qui se déroulent dans la cellule musculaire en manque d'oxygène,
produisent de l'adénosine triphosphate (ATP) et génère du
lactate (voir annexe 1). Ce rendement métabolique est converti en une
puissance mécanique (« mechanical output ») qui permet de
créer et d'entretenir la vitesse de course.
De nos jours, les écarts entre les performances chez
les athlètes de haut niveau se calculent aux centièmes de
secondes en dessous des 10s pour les hommes et en dessous de 11s pour les
femmes. Le 100m implique une contribution de 48% de phosphocréatine, de
48% de glycolyse anaérobie et de 4% de glycolyse aérobie dans la
production d'adénosine triphosphate (ATP) [25] (voir figure n°2).
Lors d'une étude plus récente, Bret et al. (2001) ont
constaté une part plus importante de l'utilisation des glycogènes
(76.2 %) qui s'accompagne de 20% de phosphagènes.
L'épreuve de 100m sprint commence par un départ
en starting-block. Ce dernier est donné par un coup de pistolet du
starter (stimulus sonore). Le temps de latence qui s'étale entre la
présentation de ce stimulus auditif et la réponse de
l'athlète est appelé communément « temps de
réaction ». Il est estimé à 1 20ms environ [20]. Mero
et al. (1992) ont démontré par des enregistrements
électromyographiques de l'activité musculaire lors du
départ, que la réponse au signal peut être
découpée en deux périodes : un temps «
pré-moteur » au cours duquel on n'enregistre qu'une activité
neuromusculaire (sans mouvement), et un temps « moteur » au cours
duquel on enregistre une activité musculaire, qui traduit la force
musculaire et qui est suffisante pour déclencher le mouvement de
départ. Le temps de réaction lors du départ est
très court chez les experts mais n'est pas corrélé avec le
niveau de performance [20].
Le 100m sprint se déroule en trois phases : «
accélération initiale », « course à vitesse
maximale » et « la décélération » [7, 9,
20, 21] (voir figure n°1). Dans les paragraphes suivants, nous allons
détailler les différents phénomènes physiologiques
et mécaniques spécifiques à chacune des phases.
2.1.1. La phase d'accélération
initiale
L'accélération initiale requiert principalement
la dégradation des phosphagènes qui sont disponibles
immédiatement mais en très faibles réserves, et
secondairement la phosphocréatine pour régénérer
rapidement de l'ATP. Les glycogènes sont aussi sollicités
dès les premières secondes du sprint [16, 28].
D'un point de vue technique, le sprinter
accélère après avoir quitté le bloc de
départ, en augmentant la longueur et la fréquence de ses
foulées. Le centre de gravité du corps de l'athlète se
situe en avant par rapport au point de contact avec le sol pendant les deux
premiers appuis, et passe derrière le point de contact à partir
du troisième contact au sol (bassin en rétroversion). Le sprinter
attaque, appuie et pousse sur l'avant- pied uniquement et ainsi, les pressions
se concentrent au niveau des articulations métatarsiennes. Au cours de
cette phase, la fréquence de la foulée (SF) et le temps d'envol
(tf) augmentent alors que le temps de contact (tc) diminue [23].
Par ailleurs, Mero et al. (1981) ont constaté une forte
corrélation entre la vitesse de course (v) et les forces propulsives (F)
précisément lors du premier contact au sol témoignant
ainsi du caractère propulsif du départ et de
l'accélération de début de course. Selon Chelly et al.
(2001), l'accélération initiale de début de course est
l'expression de la puissance mécanique et la production de forces
propulsives des membres inférieurs. La puissance mécanique
détermine l'accélération initiale au début du 100m
et permet aussi de soutenir la vitesse maximale au cours de la deuxième
phase du sprint [9, 10, 20].
2.1.2. La phase de course à vitesse maximale
Cette phase représente la période au cours de
laquelle l'athlète peut atteindre sa vitesse maximale de course. Cette
dernière correspond à un effort supramaximal qui
épuise la glycolyse anaérobie [3]. Cette
dernière se déroule en manque d'oxygène et extrait des
glycogènes les protons (H+) et leur électron. Au cours
d'un effort intense et court tel que le 100m, les protons s'accumulent dans la
cellule musculaire en entraînant une acidose (acidité du milieu).
La présence des lactates dans le sang ([La]) témoigne ainsi de
l'état d'oxydation du milieu lors de l'effort. En effet, Hirvonen et al.
(1987) ont constaté une augmentation de la concentration sanguine en
lactate (4.5 mmol.l-1) après 40m sprint chez des
athlètes de haut niveau. Les concentrations moyennes en lactates
enregistrées après le 100m chez 12 athlètes (1 0.7s), en
compétition nationale officielle, ont été proches de
8.51mmol.l-1 [15].
Au cours de cette phase, la raideur mécanique du
système musculosquelettique permet, selon une étude menée
par Chelly et al. (2001) chez une population de handballeurs confirmés,
de maintenir le plus haut niveau de vitesse par l'optimisation du rapport
amplitude-fréquence de la foulée. D'après Mero et al.
(1992), l'amplitude de la foulée stagne au cours de la deuxième
phase du sprint alors que l'athlète tente d'augmenter la
fréquence de sa foulée (estimée à 5 Hz à
vitesse maximale) afin de courir le plus vite possible.
Par ailleurs, Bret et al. (2002) ont démontré,
que la raideur des membres inférieurs (kleg) prédit
l'accélération lors de cette phase chez des sprinters de niveau
régional à national. McMahon et al. (1987) ont
précédemment constaté que kleg restait
constante quelque soit la vitesse de course. Cela n'est pas le cas pour Chelly
et al. (2001), qui considèrent qu'une raideur très importante des
membres inférieurs constitue un facteur limitant de la vitesse. Cela se
traduit par une diminution des oscillations verticales du centre de
gravité du corps à vitesse maximale quasi-constante. La raideur
mécanique pourrait ainsi être un facteur déterminant de la
performance dans cette zone du 100m sprint [7, 9, 20, 21]. C'est dans cette
optique que les membres inférieurs peuvent être
modélisés par un ressort (R) portant une charge (M)
représentant le poids et l'inertie du reste du corps (tronc + membres
supérieurs), constituant ainsi le modèle masse-ressort de la
course (ou « spring mass model », SMM) [17, 22, 23]. Ce modèle
sera discuté ultérieurement.
2.1.3. La phase de décélération
Cette phase correspond à la fin de la phase de maintien
de la vitesse maximale qui correspond à la déplétion des
réserves en glycogènes. L'acidose métabolique
altère le fonctionnement de la glycolyse anaérobie et provoque la
diminution de la puissance mécanique. Au cours de la
décélération, la fréquence de la foulée
diminue et l'amplitude augmente [20]. Selon une étude menée par
Morin et al. (2003) sur une population de sprinters « experts »,
aucun facteur mécanique mesuré n'a été lié
à la décélération à la fin du 100m. En
cherchant l'effet de la fatigue sur la variation des paramètres
mécaniques du SMM chez des « novices », ces mêmes
chercheurs ont démontré que la décélération
est due à la diminution de la raideur verticale [22].
Dans les paragraphes suivants, on exposera les différents
modèles et méthodes de calcul de la raideur mécanique au
cours de la course.
2.2. La modélisation de la raideur
mécanique au cours de la course
La modélisation du corps de l'athlète en
situation de course permet de mesurer, entre autres, la raideur
mécanique des membres inférieurs qui complète
l'observation cinétique et cinématique de sa performance. Les
modèles de la raideur mécanique permettent de comprendre les
transferts de l'énergie élastique au niveau des membres
inférieurs. On distingue 2 modèles disponibles à ce jour :
le modèle symétrique et le modèle asymétrique.
2.2.1. Le modèle symétrique
Au cours de l'appui, le pied est fixé au sol avec une
vitesse égale à zéro, alors que le centre de masse
continue son mouvement vers l'avant avec sa propre vitesse. Cela peut
être modélisé par un pendule inversé (ou «
spring loaded inverted pendulum » S.L.I.P) constitué par un ressort
linéaire fixé au sol, qui représente les membres
inférieurs, et qui supporte une masse (M) représentant la masse
du haut du corps. Le S.L.I.P illustre la succession de mouvements de
flexion-extension des membres inférieurs. Ceci correspond à un
cycle d'étirement-raccourcissement (« stretch shortening cycle
» SSC), durant lequel l'énergie élastique est stockée
et restituée (voir Figure n° 2) [18].

Figure n° 2: Modèle symétrique
(S.L.I.P). [McMahon et al. (1990)]
« è » : angle d'envol, «
AL » : déplacement du centre de masse, «
L0 »: longueur de la jambe-ressort,
«
Lmax »: longueur maximale de la jambe au cours de
l'appui, « CM » centre de masse de M.
2.2.2. Le modèle asymétrique
Le modèle asymétrique d'Alexander (1990)
considère le corps humain comme étant trois segments rigides,
i.e. un tronc et deux segments égaux représentant les membres
inférieurs (cuisse + jambe). Le centre de masse se trouve au niveau du
bas du tronc et représente le centre de masse de tout le modèle.
Le système est analysé en deux dimensions (2D) avec le pied comme
origine (voir figure n°3) [1].

Figure n°3: modèle
asymétrique. [Alexander (1990)]
(« a » : longueur des membres
inférieurs, « è »: angle d'incidence
au sol, « Ö » : angle du genou)
2.3. Les méthodes de calcul de kleg
L'acquisition et les calculs des différentes
données cinétiques et cinématiques dépendent de la
disponibilité des outils techniques technologiques et de la nature de
l'expérimentation (voir tableau n°1).
Tableau n°1 : méthodes de calcul de
kleg au cours du sprint
|
Méthode
|
Auteur
|
Formules
|
Avantages et inconvénients
|
|
Farley et al.
(1999)
|
kleg= Ó kjoint
kjoint = ÄMjoint
Äèjoint
|
- Nécessite un système de capture du mouvement
(caméras), et des plates formes de force.
- calculs des forces et moments de
force par la dynamique
inverse.
|
|
Morin et al.
(2005)
|
-1
kleg = F · ÄL
max
|
- Utilisable dans des conditions de terrain
- Calcul rapide de la raideur mécanique
- Nécessite un radar pour
enregistrer les vitesses instantanées et des semelles
équipées de
capteurs de pression (switches) pour enregistrer les temps
d'envol et de contact
|
|
Ä = L - L 2 - ( vt 2)
2 + Ä y
L c c
F ( t ) = F max × sin[( Ð
t c ) · t ]
- 1
k vert = F · Ä y c
max
F max =mg·(Ð
2)·[(t f t c )+1)]
2 2
t t
F max c c
Ä = - · + ·
|
|
y c2 g
m Ð 8
|
Abréviations : « m »: la
masse corporelle (m), « g » :
accélération de la pesanteur (m.s-2), « F
»: force (N), « k »: la raideur
(kN.m-1), « Ayc » : le
déplacement vertical du centre de masse, « AL
» : l'abaissement de la jambe ressort (m), « L
»: la longueur de la jambe (m), « M »:
le moment de force (N.m-1), « è »
: angle articulaire (° : degré), « tf
»: temps d'envol (s), « tc
»: temps de contact (s).
2.3.1. La méthode analytique
La méthode analytique suppose que la raideur totale des
membres inférieurs est égale à la somme des raideurs
articulaires des articulations (kjoint) impliquées dans le
mouvement, i.e. la hanche, le genou et la cheville (voir
figure n° 4). La raideur articulaire est égale au produit de la
variation du moment de force par la variation de l'angle articulaire (voir
tableau n°1).

Figure n° 4 : Modèle masse
ressort. [Farley et al. (1999)]
La raideur de la jambe est la somme des raideurs de chaque
articulation, « è »
représente l'articulation des
trois angles de la jambe.
2.3.2. La méthode sinusoïdale
La courbe de la force en fonction du temps (F(t))
de la jambe ressort est périodique. Cette oscillation revête une
forme sinusoïdale [2]. La courbe F(t) est définie par
l'équation suivante [22] (voir annexe 2):
F(t) = Fmax × sin ( Ð t)
CT
Cette méthode ne nécessite pas la mesure de la
force. Elle utilise un minimum de paramètres anthropométriques et
cinétiques pour calculer la raideur des membres inférieurs
(kleg) et la raideur verticale du système (kvert)
(voir annexe 2): la vitesse au moment du contact au sol (vc), la
masse corporelle (m), la longueur des membres inférieurs
(lleg), le temps de contact (tc), et le temps d'envol
(tf). Elle a été récemment utilisée par Morin et
al. (2005) pour évaluer la course sur tapis de course et sur piste.
2.4. Les paramètres mécaniques du SMM
2.4.1. La force musculaire
Mero et al. (1981) ont démontré l'effet du type
de fibre sur la vitesse de course. En effet, les vitesses de départ, de
la phase d'accélération et de la phase de course à vitesse
constante ont été corrélées avec la proportion de
fibre de type II dites rapides, ou «fast twitch » (FT) du vaste
externe (muscle de la cuisse dont la fonction principale est l'extension du
genou). Une corrélation positive a été mise en
évidence également entre la proportion de fibres FT du même
muscle et la résultante des forces propulsives au cours de la vitesse
maximale, ce qui témoigne de l'importance de la force musculaire dans la
production de mouvement rapide.
La course est une activité cyclique dont les
contractions musculaires sont plyométriques. Dans ce type de mouvements,
une force élevée est produite par la composante contractile des
muscles et/ou grâce à la capacité des structures
musculotendineuses à stocker et restituer l'énergie
élastique, et permet donc de développer une grande puissance, une
fois couplée avec des valeurs maximales de vitesse [10, 19, 20].
2.4.2. La vitesse de course
La vitesse de course est le produit de la fréquence de
la foulée (SF) et la longueur de la foulée. Elle a
été calculée, par exemple, aux alentours de 8.1
m.s-1 selon une étude récente de Morin et al (2005)
menée avec des étudiants en éducation physique (non
spécialistes en sprint). L'augmentation de la vitesse est une
élévation de la fréquence gestuelle tout en augmentant
l'amplitude de la foulée. Cette dernière, étant
mécaniquement et morphologiquement limitée, est optimisée
lors de la course à vitesse maximale, par une fréquence gestuelle
maximale, pour favoriser une meilleure production de vitesse [20, 23]. Mero et
al (1992) suggèrent que SF joue un rôle plus décisif sur la
vitesse de course que la longueur de la foulée.
On remarque que la vitesse de course est significativement
différente aussi bien entre 40 et 60 mètres qu'entre 60 et 80 m,
ce qui témoigne de découpage commun de la course de vitesse en
trois phases qui ont été déjà décrites
auparavant et schématisées sur la figure n°1.
2.4.3. La raideur mécanique prédit-elle
la vitesse de course ?
Morin et al. (2005) ont montré que la performance au
100m sprint était proportionnelle à la raideur verticale et que
la raideur des membres inférieurs restait inchangée
(kleg =19,6 kN.m-1, environ) quelque soit la valeur
maximale de la vitesse. Cette étude confirme les résultats
observés par Chelly et al. (2001) qui ont mis en évidence une
corrélation significative entre la raideur des membres inférieurs
au cours de sauts verticaux et la vitesse maximale atteinte au cours d'un 40m
sprint.
Par ailleurs, Bret et al. (2002) ont constaté que la
raideur des membres inférieurs prédit les changements de vitesse
entre les différentes phases du sprint chez les experts. Ainsi, la
valeur la plus élevée de la raideur détermine la
capacité du sprinter d'atteindre sa vitesse maximale au cours de la
deuxième phase du 100m. Par ailleurs, Chelly et al. (2001) ont
suggéré une limitation probable de kleg à une
valeur optimale même à des vitesses élevées,
étant donné que le cycle étirement-raccourcissement des
muscles des membres inférieurs nécessite un minimum de flexion
des genoux et des chevilles. La vitesse dépend, selon ces chercheurs, de
deux paramètres : la puissance musculaire, qui est nécessaire
pour l'accélération initiale au cours de la première phase
de la course, et la raideur des membres inférieurs, qui permet
d'atteindre la vitesse maximale au cours de la deuxième phase du sprint
[9].
Dans la littérature, la raideur mécanique semble
affecter directement la performance au 100m sprint et constitue ainsi un
facteur mécanique important pour la compréhension de la
régulation des facteurs biomécaniques au cours de la course ainsi
que la variation de l'effort au cours de chaque phase du 100m.
2.5. La fatigue musculaire 2.5.1. Modèles de la
fatigue
2.5.1.1. Modèle
énergétique/déplétion des réserves
énergétiques
Le corps a besoin de sources énergétiques
indispensables à la réalisation du mouvement. Le
métabolisme s'adapte à chaque type d'effort en approvisionnant
les muscles. Les réserves énergétiques sont susceptibles
d'être épuisées. La déplétion des
réserves provoque l'atténuation de l'activité musculaire.
Selon le modèle énergétique, la
fatigue est donc provoquée par deux mécanismes
principaux : un approvisionnement insuffisant des muscles actifs en ATP, et une
déplétion des substrats énergétiques.
La fatigue se manifeste par « la réduction de la
capacité de performance lors d'un travail intense ». « La
fatigue musculaire ou physique relève de modification dans le muscle
squelettique (chutes des réserves, accumulation d'acide lactique) »
[27]. D'après Cerretelli (2002), les mécanismes oxydatifs
impliqués dans une épreuve de courte durée ne sont pas en
mesure, une fois épuisées les réserves de
phosphocréatine, de fournir l'énergie nécessaire pour
soutenir (avec l'énergie venant de la glycolyse anaérobie) les
charges imposées.
La fatigue peut être expliquée par l'augmentation
de la concentration de sang en lactate ([La]). Il provient de la transformation
des pyruvates en lactates sous l'influence de la
lacticodéshydrogénase (LDH), en présence d'un co-facteur
le NAD+. Le lactate peut être formé à partir d'acides
aminés, en particulier dans les états de catabolisme
protéique intense (effort bref et/ou intense) [24].
2.5.1.2. Modèle neuromusculaire
Dans ce modèle, la diminution de la capacité de
production de force maximale n'est pas due à l'incapacité des
processus énergétiques de fournir l'énergie
nécessaire à l'effort au niveau de l'organe
périphérique, y compris les muscles squelettiques. Elle est
plutôt d'ordre neuromusculaire, c'est-à-dire au niveau du
recrutement des unités motrices par le système nerveux
central.
La fatigue est associée à des perturbations au
niveau de la commande nerveuse et aussi à des modifications au niveau
musculaire. Les principales causes de la fatigue peuvent être la
réduction de la conduction nerveuse ou la réduction de la
réponse musculaire au stimulus transmis par le système nerveux.
Le modèle neuromusculaire explique la fatigue, à un niveau
périphérique, par la détérioration de
l'efficacité des composantes intramusculaires [14].
2.5.1.3. Modèle biomécanique
Le sprint est l'expression d'une puissance de travail
importante qui permet de produire et maintenir un haut niveau de vitesse. Cette
puissance est le résultat d'une dépense
énergétique. L'efficacité technique dépend alors de
la disponibilité des réserves énergétiques.
La fatigue correspond à la diminution de
l'efficacité technique. Selon cette théorie, l'optimisation de
l'efficacité du mouvement résulte d'une déplétion
moins rapide des réserves énergétiques, et d'une
élévation de la température du corps
atténuée. Seulement 25 % de l'énergie produite par
l'organisme est utilisé pour la contraction musculaire et le reste
assure la régulation des échanges thermiques [3].
2.5.1.4. Modèle complexe
La fatigue est une expression physique d'une sensation. Lors
de la course, l'effort est régulé en boucle, à un niveau
central dans le système nerveux et dans les muscles à un niveau
périphérique. Gibson et al. (2004) suggère que la
réalisation d'un exercice physique est gouvernée par une commande
centrale qui intègre et calcule tous les signaux sensoriels afin de
maintenir une homéostasie et permettre une « sensation de fatigue
acceptable par l'athlète ». Ce processus est un exemple de
système dynamique non-linéaire dans lequel les processus
physiologiques interagissent comme une partie du système global en
régulant l'effort avant, pendant et après l'exercice.
La fatigue provoque un changement inconscient de la
foulée qui sera véhiculé vers le système nerveux
central (SNC) qui contrôle le recrutement des unités motrices
(afférences) [26]. Lors du sprint en particulier, l'effort est à
la fois court et intense. Cela ne laisse pas de délais aux commandes
centrales pour varier les degrés de sollicitation musculaire. La
manifestation de la fatigue est dans ce cas là inconsciente. Elle est en
effet la résultante d'un ensemble de feedbacks physiologiques,
biochimiques et sensoriels qui ne sont pas forcément associés
à la variation de la production de force musculaire et qui sont
ressentis lors de la récupération.
2.5.2. Effet de la fatigue sur la performance
La fatigue diminue la performance globale au 100m sprint [23].
Elle se manifeste par l'incapacité d'endurer la vitesse maximale
atteinte lors des premières secondes du sprint. La
décélération de fin de course est une conséquence
directe de la fatigue musculaire qui est expliquée par
l'épuisement des réserves de glycogènes vers la fin de la
deuxième phase du sprint [28]. En effet, Zouhal et al. (2002) ont
démontré que la glycolyse anaérobie jouait un rôle
non négligeable dans la production de l'énergie indispensable
à la réalisation de sprint de durée inférieure
à 6 secondes.
Morin et al. (2006) ont démontré que la
performance au 100m sprint diminue avec la fatigue induite par la
répétition d'un effort maximal. En effet, la vitesse moyenne du
quatrième 100m, mesurée chez des novices, correspond à
92.7 % de leur première répétition.
Nous nous intéressons à l'étude de la
fatigue sur la performance au 100m par la mise en relief de la relation entre
la déplétion des réserves énergétiques et la
variation des paramètres mécaniques, principalement la raideur
des membres inférieurs, au cours de la répétition de
sprint.
Hypothèses :
Fort de ces connaissances théoriques, nous émettons
les hypothèses suivantes :
(1) La raideur mécanique permet de conserver la vitesse
maximale de course au cours du 100m sprint.
(2) La raideur des membres inférieurs est
corrélée avec la diminution de la vitesse de course lors de la
répétition de 100m sprint.
(3) La diminution de la vitesse avec la fatigue est liée
à l'augmentation des taux de lactates sanguins.
*******
3. Méthodologie
Tous les sujets ont donné leur libre consentement pour
participer à cette expérience et nous ont autorisés
à utiliser leurs résultats à des fins scientifiques.


Figure n°5 : Schéma récapitulatif
du protocole expérimental
3.1. Population
Notre population se compose de huit sprinters (hommes) de
niveau régional (Ile de France) qui courent le 100m en 1 1,54s (#177;
0,37). Les sujets sont en moyenne âgés de 19 #177;2,06 ans,
mesurent 1,80 #177;0,07m et pèsent 72,88 #177;5,78 kg.
3.2. Protocole expérimental
Nous avons demandé aux sujets de courir 4 × 100m,
avec un départ debout, le plus rapidement possible, après avoir
effectué leur échauffement standard. Les sprinters étaient
équipés de chaussures d'athlétisme avec pointes.
Les 4 répétitions sont séparées de
3 minutes de récupération semi-active (marche). Nous
considérons ce type de répétition comme un protocole
induisant une fatigue musculaire. La récupération imposée
est insuffisante pour laisser apparaître des quantités maximales
de lactates sanguins après chaque 100 m. Dans la littérature, les
concentrations sanguines en lactate augmentent entre 5 et 9 minutes suite
à un sprint court et intense [11, 13].
3.3. Modélisation de la course
Nous avons choisi le modèle symétrique de
McMahon et al. (1990) car il considère les membres inférieurs
comme une seule composante du système sans tenir compte des proportions
segmentaires (voir figure n°3). Le bas du corps du sprinter est donc
modélisé par un ressort linéaire simple de raideur (k) et
d'une longueur égale à la distance entre le grand trochanter et
le sol.
Dans ce modèle, les membres supérieurs font
partie du haut du corps. Leurs mouvements ne seront pas étudiés
en tant que tel durant ce travail de recherche, bien que leur rôle soit
primordial dans la mobilisation du tronc par une flexion constante de l'angle
bras/avant-bras qui évite l'excès de mouvement naturel
d'antépulsion (dans le plan antéropostérieur) et de
rétropulsion (vers l'extérieur) dus au positionnement vers
l'avant, le haut, et l'extérieur de la cavité
glénoïde de l'articulation de l'épaule.
Nous avons opté pour la méthode sinusoïdale
proposée par Morin et al. (2005) pour évaluer les
paramètres mécaniques de la course (voir tableau n°1). Cette
méthode permet de calculer la raideur des membres inférieurs et
la raideur verticale avec un minimum de paramètres (tc,
tv, v, L, M) (voir annexe n°2) et dans des conditions de
terrain.
3.4. Variables étudiées
Les variables étudiées sont réparties en 3
groupes :
3.4.1. paramètres de la foulée : - temps
de contact (tc),
- temps d'envol (tf),
- L'abaissement du centre de masse (?L)
- l'amplitude et la fréquence (SF) de la foulée.
3.4.2. paramètres mécaniques de la
course: - la force (F),
- la vitesse (v),
- la raideur des membres inférieurs (kleg) - la raideur
verticale (kvert).
3.4.3. variable physiologique :
- concentration sanguine en lactate [La].
3.5. Mesure des paramètres cinétiques
Le temps de contact et le temps d'envol ont été
mesurés par un « Locotest » (Techno-Concept) (voir Image
n°1). Ce dernier consiste en un système d'acquisition sans fils du
mouvement qui se compose de deux semelles et d'une ceinture, contenant des
capteurs de pressions et des accéléromètres. Cet
appareillage est embarqué sur le sujet. Le Locotest est connecté
via des modules à un ordinateur par une liaison « Bluetooth »
qui facilite l'expérimentation sur terrain (voir figure n°5). Le
deuxième avantage de ce dispositif est la mesure des
accélérations sur 2 axes, haut-bas et avant- arrière.

Image n°1 : Locotest
(Techno-concept)
Les mesures des vitesses instantanées du coureur ont
été réalisées grâce à un radar «
Stalker ATS » (image n°2) échantillonné à 250 Hz
et placé à une hauteur de 1
mètre du sol ( hauteur de la taille du sprinter), et
à 3 mètres de la ligne de départ (voir figure
n°5).
Nous avons déterminé les vitesses
instantanées au 30, 60, 80 et 100m en correspondant le moment de passage
de l'athlète entre les lattes, obtenus sur la vidéo, et la
vitesse correspondante obtenues par le radar.

Image n°2: Radar (Stalker ATS)
3.6. Observation de la cinématique de la
course
La course a été filmée à l'aide
d'une caméra (25hz) placée à 7 mètres du couloir de
course et centrée au niveau du 50m de façon à filmer la
totalité du 100m. Ce dernier a été découpé
en 4 portions en plaçant des lattes, de part et d'autre du couloir, au
niveau du 30m, 60m, 80m et 100m, pour éviter la parallaxe du mouvement
et compter le nombre d'appuis à chaque phase. Les athlètes ont
été avisés de leur temps au 100m et la
récupération a été limitée à 3
minutes, en utilisant un chronomètre manuel. Ce découpage nous
permet d'observer la variation des paramètres de la foulée au
cours des différentes phases du 100m, i.e. l'amplitude et la
fréquence de la foulée. Pour ce faire, Nous compterons le nombre
d'appuis par portions. Ceci nous permettra de calculer l'amplitude moyenne de
la foulée, en divisant la distance par le nombre d'appui d'un seul pied,
et la fréquence moyenne de la foulée qui est le produit du nombre
d'appui d'un pied par le temps parcouru par phase.
3.7. Mesure de la lactatémie
La prise de sang lors de la récupération est la
seule méthode de mesure de la lactatémie sur terrain. Nous avons
prélevé du sang en appui sur l'index d'une main et en faisant une
piqure de l'autre. Le doigt est désinfecté à chaque
prélèvement avec du coton imbibé d'alcool. Une fois
l'index piqué, nous récupérons une goutte de sang par une
lamelle pour l'introduire dans le Lactate Pro (voir image n° 3). La
lecture du taux de lactate se fait directement sur l'écran de
l'appareil.

Image n° 3 : Lactate Pro
Les mesures ont été réalisées par un
appareil de mesure du lactate sanguin, le « lactate-pro », de la
façon suivante (voir tableau n°2):
· 1 premier prélèvement sanguin au repos
[L0],
· 4 prélèvements (après une minute de
repos) après chaque
100m, et à 3mn, 5mn et 7mn lors de la
récupération passive
jusqu'à enregistrer la concentration maximale des
lactates
dans le sang.
Tableau n° 2 : Organisation des
prélèvements sanguins
|
Au Repos
|
Après 100m-1
|
Après 100m-2
|
Après 100m-3
|
Après 100m-4
|
Après
3mn
|
Après 5mn
|
Après 7mn
|
|
[La] 0
|
[La] 1
|
[La] 2
|
[La] 3
|
[La] 4
|
[La] 5
|
[La] 6
|
[La] 7
|
La variation de la lactatémie (?[La]) est
calculée en déduisant la concentration de sang en lactate du
repos ([La0]) de la valeur de la lactatémie à un instant t de
l'expérience [Lax].
?[La] = [Lax] - [La0]
3.8. Analyse statistique
On a effectué une analyse de la variance « ANOVA
» avec mesures répétées et le test post-hoc LSD de
Fisher afin de tester l'effet de la fatigue sur la variation des
paramètres cinétiques et cinématiques de la course au
cours des 4 répétitions de sprint. Nous avons également
étudié les corrélations (r de Bravais-Pearson) entre les
concentrations sanguines de lactate, les paramètres biomécaniques
et les paramètres de performance des 4 répétitions de
100m. Le seuil de signification a été fixé à
p<0,05.

138
136
134
132
130
128
126
124
122
120
118
116
114
195
190
185
180
175
170
165
160
155
150
145
140
Figure n°6: Evolution du temps de contact au cours de la
répétition
(4 x 100m)
Figure n° 7: Evolution du temps d'envol au cours de la
répétition
(4 x 100m)
1 2 3 4
1 2 3 4
Séries de 100m
Séries de 100m
4. Résultats
4.1. Le temps de contact (tc) et le temps d'envol
(tf)
Le temps de contact moyen est égal à 125
#177;0.01ms. Le temps d'envol est égal à 170 #177;0.01ms. Ni l'un
ni l'autre ne varie avec la répétition de 100m sprint [F (3,21)
<1].
4.2. L'abaissement du centre de masse (AL)
L'abaissement du centre de masse est égal à 0.17
#177; 0.02m en moyenne. L'abaissement du centre de masse ne varie pas avec la
répétition [F (3,21) <1].

0,22
0,21
0,20
0,19
0,18
0,17
0,16
0,15
0,14
0,13
Figure n°8: Evolution de l'abaissement du centre de masse
au
cours de la répétition (4 x 100m)
1 2 3 4
Séries de 100m
4.3. La fréquence (SF) et l'amplitude de la
foulée
La fréquence de la foulée est calculée
à 3.9 #177;0.27Hz en moyenne avec des valeurs comprises entre 3.45 et
4.4 Hz. Elle diminue au cours des 4 x 100m de 3.99 #177;0.25Hz au cours du
premier sprint à 3.9 #177;0.26Hz au cours de la quatrième
répétition [F (3,21) = 3.13; p<0.05]. Le test post hoc LSD de
Fisher a montré une différence significative entre SF du
3ème et le 4ème sprint. La fréquence
de la foulée diminue avec la fatigue.
L'amplitude de la foulée est égale à 2.08
#177;0. 14m en moyenne avec des valeurs qui fluctuent entre 1.83 et 2.33m. Elle
diminue avec la répétition de 2.06 #177;0.14m lors de la
première répétition à 2.03 #177;0.12m au cours du
quatrième sprint [F (3,21) = 4.22; p<0.05]. Le test post-hoc LSD de
Fisher a montré une différence significative entre la
deuxième et la quatrième répétition.

La longueur de la foulée est corrélée avec
la vitesse au 100m [r = 0.67; p<0.05]. L'amplitude de la foulée
diminue avec la diminution de la vitesse de course.

2,4
2,3
2,2
2,1
2,0
1,9
1,8
7,0 7,2 7,4 7,6 7,8 8,0 8,2 8,4 8,6
Vitesse de course du premier 100m (m.s-1)
Figure
n°11: Relation entre la vitesse et l'amplitude de la foulée
95% de confiance

29
28
27
26
25
24
23
22
21
31
30
Figure n° 12: Evolution de la force maximale au cours de
la
répétition ( 4 x 100m)
1 2 3 4
Séries de 100m
4.4. La force (F)
La force des sujets qui constituent notre échantillon
est égale à 2.68 #177; 0.26 kN en moyenne. Elle ne varie pas
significativement avec la répétition [F (3,21) <1]. La
répétition n'affecte pas la force des membres
inférieurs.
4.5. La performance au 100m
Le temps au 100m (12.43 #177;0.55s) augmente significativement
de 12.24 à 12.71 m.s-1 ; [F (3,21) = 4.19; p<0.05]. La
vitesse de course est égale à 8.06 #177;0.34m.s-1.
Elle diminue significativement (8.36%): [F (3,21) = 4,03 ; p<0.05], de 8.18
#177;0.29m.s-1 au premier 100m à 7.89 #177;0.42
m.s-1 au quatrième sprint. Le test post-hoc LSD de Fisher
indique une différence significative entre les performances du premier
et du dernier sprint et aussi celles de la deuxième et de la
quatrième répétition. La performance au 100m diminue avec
la répétition.

4.6. La raideur des membres inférieurs
(kleg)
La raideur des membres inférieurs a été
évaluée à 15.86 #177;2.76 kN.m-1 en moyenne et
les valeurs sont comprises entre 11.06 et 19.84 kN.m-1. Elle
évolue de 16.05 #177;2.59 kN.m-1 lors du première 100m
à 17,16 #177;1.42 kN.m-1 lors du dernier sprint. Cependant,
la différence n'est pas significative entre les 4
répétitions [F (3,21) <1]. La raideur des membres
inférieurs n'est pas affectée par la fatigue au cours du 4×1
00m.

21
20
19
18
17
16
15
14
13
12
11
10
Figure n°14 : Evolution de la raideur des membres
inférieurs au cours de la
répétition (4 x 100m)
1 2 3 4
Séries de 100m
4.7. La raideur verticale (kvert)
La raideur verticale a été évaluée
à 69.53 #177;6.35kN.m-1 en moyenne avec des valeurs
rangées entre 57.21 et 77.95 kN.m-1. Elle évolue de
72.57 #177;5.65 kN.m-1 lors du premier 100m à 69.5 #177;2.85
kN.m-1 lors du dernier sprint, mais la différence n'est pas
significative entre les 4 répétitions [F (3,21) <1].
La raideur verticale varie dans le même sens que le
temps de contact [r = 0.77; p<0.05] et l'abaissement maximal des membres
inférieurs [r = 0.8; p<0.05], au cours du 100m. Elle est inversement
corrélée [r = -0.62; p<0.05] avec la variation de la force
maximale.

82
(r = -,76)
80
95% de confiance
78
76
74
72
70
68
66
64
62
60
58
56
112 114 116 118 120 122 124 126 128 130 132 134 136 138
Figure n°15: Relation de la raideur verticale avec le temps
de contact
Temps de contact (ms)
(56)
(58)
95% de confiance
(60)
(62)
(64)
(66)
(68)
(70)
(72)
(74)
(76)
(78)
(80)
0,13 0,14 0,15 0,16 0,17 0,18 0,19 0,20 0,21 0,22 0,23
Abaissement du centre de masse (m)
Figure n°16: Relation entre l'abaissement du centre de
masse et la raideur
verticale

(56)
(58)
(60)
(62)
(64)
(66)
(68)
(70)
(72)
(74)
(76)
(78)
(80)
22 23 24 25 26 27 28 29 30 31 32
Figure n°17: Relation entre la raideur verticale et la
force maximale
aucours d'un 100m sprint
Force maximale (kN)
95% de confiance
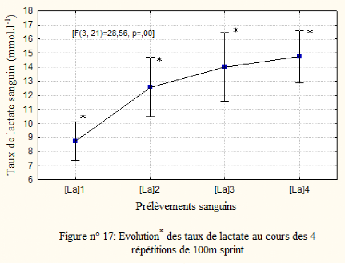
4.8. Les concentrations sanguines en lactate ([La])
Les concentrations de sang en lactates augmente
significativement de 8.71 #177; 1.56 au premier sprint à 14.76 #177;
2.08 mmol/l à la quatrième répétition: [F (3, 21) =
35.37; p<0.05] (Voir Figure n°7). Le test post-hoc LSD de Fisher montre
une différence significative entre les 4 répétitions de
100m. Les concentrations sanguines en lactate augmentent avec la
répétition de 100m à vitesse maximale.
5. Discussion
Les valeurs moyennes du temps de contact (125 ms), du temps
d'envol (170 ms), et de l'abaissement du centre de masse (0.17 m) sont
supérieures à celles de la littérature. Par exemple, Morin
et al. (2006) ont constaté, chez des étudiants en
éducation physique et sportive, un temps de contact qui varie avec la
répétition (120.5 ms) et un temps d'envol moyen égal
à 144.5 ms qui reste inchangé. Dans les 2 études,
l'abaissement du centre de masse ne fluctue pas significativement au cours de
la répétition. La fatigue n'affecte pas l'abaissement du centre
de masse.
La différence de durée des temps de contact et
d'envol peut s'expliquer par le port de chaussure d'athlétisme chez les
sujets de notre population ce qui n'est pas le cas chez les novices qui ont
participé à l'étude de Morin et al. (2006). Le temps de
contact est plus élevé chez les experts. Selon Bishop et al.
(2006), le port de chaussures à pointes augmente la flexion dorsale de
la cheville lors de l'atterrissage. Cela peut expliquer partiellement
l'élévation des valeurs du temps de contact au sol chez les
experts sans pour autant expliquer la différence des valeurs en fonction
de l'expertise.
L'amplitude de la foulée, qui est égale à
2.08 m en moyenne, semble normale par rapport aux résultats de la
littérature. Mero et al. (1992) l'ont moyenné entre 2 et 2.6m. En
comparant nos résultats à cette étude, nous remarquons que
la fréquence de la foulée (3.9 Hz) était inférieure
à 5Hz, sachant que cette valeur correspondait à une valeur
enregistrée chez des sprinters experts. Les fréquences de la
foulée de notre population ont diminué de 9 % entre la
première et la dernière répétition. Ces
résultats ressemblent à ceux de Morin et al. (2006) chez une
population de novices, i.e. une diminution de 8.03 % dans des conditions
identiques à celle de notre protocole expérimental.
L'entraînement ne permet pas de diminuer l'effet de la fatigue sur la
fréquence de la foulée.
Les valeurs de la force maximale du premier sprint (2.76 kN)
sont supérieures à celles de Mero et al. (1992) chez des experts
en condition de course à vitesse maximale (1.76 kN). Ces chercheurs ont
démontré que la force maximale augmentait dans le même sens
que la vitesse de course (9.96 m.s-1). La différence des
valeurs de la force
maximale peut donc être expliquée par les
différences de performance entre les deux populations.
Les valeurs moyennes de la force maximale (2.68 kN) sont
similaires aux résultats de Morin et al. (2006) chez des novices et dans
un protocole identique à celui de notre expérimentation. A notre
connaissance, aucune étude n'a démontré, à ce jour,
que la répétition de 100m à vitesse maximale affecte la
force des membres inférieurs.
Le temps au 100m a été calculé à
12.43 s, c'est à dire que nos sujets ont couru en moyenne à une
vitesse égale à 8.06 m.s-1 sur l'ensemble des 4
répétitions. La performance du premier 100m (12.43 s) est
nettement inférieure au record du monde du 100m qui est égale
à 9.69 s. La performance de notre population est éloignée
de celle énoncée au début (11.54 s). Cela peut être
expliqué par la différence de motivation entre une
compétition officielle et une expérimentation scientifique. Le
protocole (4 x 100m) est compris par les athlètes comme étant un
protocole de fatigue. Cela n'éloigne pas l'hypothèse que les
sprinters répartissent leurs énergies, et ne courent pas chaque
sprint avec la vitesse maximale possible, comme l'indique notre consigne
orale.
La vitesse de course a diminué de 8.18 m.s-1
au premier 100m à 7.89 m.s-1 au quatrième sprint,
c'est-à-dire de 8.36 %. Morin et al. (2006) ont constaté une
diminution de 11.6 % dans des conditions similaires chez des novices, avec une
vitesse moyenne qui diminue de 8.10 m.s-1 au premier sprint à
7.10 m.s-1 à la quatrième répétition. La
baisse de la performance avec la fatigue due à la
répétition est plus élevée chez les novices (3.5
%). L'expertise permet de diminuer les effets de la fatigue sur la performance
au 100m.
La raideur des membres inférieurs a
évolué de 16.05 #177;2.59 kN.m-1 lors de la
première répétition de 100m à 17,16 #177;1.42
kN.m-1 au cours de la dernière répétition. Elle
diminue de 19.5 kN.m-1 à 18.3 kN.m-1 chez des
novices dans des conditions similaires [23]. La fatigue n'affecte la raideur
des membres inférieurs, ni chez experts, ni chez les novices. Est-ce une
stratégie du système nerveux central pour contrôler les
différents paramètres mécaniques ? La raideur des membres
inférieurs est-elle une
variable macroscopique commandée par le SNC pour conserver
et optimiser les paramètres de la foulée au cours de la course
?
La raideur verticale diminue de 72.57 à 69.5
kN.m-1 entre le premier et le dernier sprint. Ces valeurs sont
supérieures à celles des novices [23] dont la raideur verticale
diminue significativement de 93.9 à 74.9 kN.m-1. Morin et al.
(2006) ont constaté que la diminution de la raideur verticale est la
conséquence de la diminution de la vitesse. Cela n'est pas le cas chez
les experts dont la raideur verticale ne varie pas avec la
répétition. La fatigue n'affecte pas la raideur verticale chez
les sprinters confirmés.
Au cours du 100m, la raideur verticale varie dans le
même sens que le temps de contact et l'abaissement maximal des membres
inférieurs. La raideur verticale permet de soutenir le temps de contact
et l'abaissement du centre de masse au cours d'un 100m sprint. Cela permet au
système musculotendineux de restituer de l'énergie
élastique au cours d'un temps court, ce qui explique
l'insensibilité de l'abaissement du centre de masse à la
décélération de fin de course. La raideur verticale permet
d'optimiser la foulée lors d'un 100m en réduisant le temps de
contact et la fluctuation verticale du centre de masse.
Les taux de lactate enregistrés sont similaires
à ceux de Bret et al. (2001) qui ont démontré une
corrélation entre la performance (11.7s) et les concentrations de
lactate sanguin (12.5 mmol.l-1) lors de compétition
officielle de 100m, ce qui n'est pas le cas pour Hautier et al. (1994) qui ont
constaté des taux de lactates inférieurs aux valeurs
enregistrées par Bret et al. (2001) (8.5 mmol.l-1) et
indépendants des performances au 100m (valeurs de lactates
enregistrées lors d'une compétition officielle). De même,
Nummela et al. (1992) ont démontré que les concentrations de sang
en lactates n'étaient corrélées qu'avec la performance
finale au 400m et qu'elles étaient indépendantes de celles des
distances inférieures, i.e. le 300, le 200 et le 100m. Les
filières énergétiques sollicitées lors d'une
série de 4 répétitions de 100m (anaérobie
alactique) ne sont pas les mêmes que celles sollicitées lors d'une
épreuve 400m (anaérobie lactique), donc, il n'est pas possible de
comparer nos résultats à ceux de Nummela et al. (1992).
6. Conclusion
Le but de cette étude était d'étudier la
variation des paramètres cinétiques et cinématiques au
cours de la répétition de sprint (4 × 100m) et l'effet de la
fatigue physiologique sur la performance au 100m.
Notre première hypothèse était l'effet de
la raideur mécanique sur la conservation de la vitesse maximale de
course. Nous avons prouvé que la raideur verticale affecte indirectement
la performance au cours du 100m, en faisant varier les paramètres de la
foulée. Notre hypothèse n'est pas confirmée car nous
n'avons démontré aucune relation directe entre la raideur des
membres inférieurs ou la raideur verticale et la vitesse de course. Au
cours du 100m sprint, l'abaissement des membres inférieurs et le temps
de contact varient dans le même sens que la raideur verticale. Cela nous
amène à conclure que des valeurs optimales de raideur verticale
sont nécessaires pour optimiser la restitution de l'énergie
élastique au cours du cycle étirement- raccourcissement, ce qui
permet le maintien de la vitesse maximale.
Nous avons émis l'hypothèse que la raideur des
membres inférieurs est corrélée avec la diminution de la
vitesse de course au cours de la répétition de 100m. Nous avons
démontré que la performance au 100m diminue avec la
répétition de 4 × 100m, et que l'expertise permet de
diminuer les effets de la fatigue. Cette dernière n'affecte ni la
raideur des membres inférieurs ni la raideur verticale.
Notre protocole amène à une fatigue
périphérique qui affecte les paramètres de la
foulée. Le temps de contact, le temps d'envol et l'abaissement du centre
de masse reste inchangés au cours de la répétition. La
fatigue induit une diminution de l'amplitude et de la fréquence de la
foulée à travers les répétitions de 100m. Cela
empêche les sprinters experts de garder leur haut niveau de vitesse et
à décélérer à la fin de la course. Chez les
novices [23], la fatigue induit une diminution de la fréquence de la
foulée qui s'accompagne d'une diminution du temps de contact. Le temps
d'envol reste inchangé. Cela n'est pas le cas pour les experts de notre
expérience car la fatigue affecte leurs fréquences et leurs
amplitudes de foulée indépendamment des paramètres
temporels.
L'insensibilité de la raideur des membres
inférieurs et de la raideur verticale des athlètes experts
à la répétition se présente comme un processus
d'adaptation à la fatigue. Selon Nummela et al. (1992), cette
dernière induit une augmentation de la durée de la contraction
musculaire responsable de la prolongation du temps de contact au sol. Le temps
de contact ne varie pas avec la fatigue due à la
répétition de sprint. Les fluctuations verticales du centre de
masse sont ainsi réduites par la conservation du temps de contact et de
l'abaissement des membres inférieurs. Chez les experts, la raideur des
membres inférieurs est restée inchangée avec la
répétition de 100m. La conservation d'une certaine valeur de la
raideur mécanique est-elle une stratégie du SNC pour gérer
les différents paramètres mécaniques ?
La diminution de la performance avec la
répétition peut être expliquée, au niveau
énergétique, par l'augmentation des concentrations de lactate.
Notre troisième hypothèse est confirmée. Le
métabolisme anaérobie est incapable d'endurer un effort
supramaximal sur une durée telle que celle du 100m. L'accumulation des
lactates sanguins avec la répétition de sprint est due à
l'enchaînement de répétition de course à vitesse
maximale avec des récupérations incomplètes, donc à
la sommation des taux de lactate de chaque 100m.
Notre travail de recherche a permis de mieux comprendre la
variation des paramètres biomécaniques avec la
répétition et l'effet de la fatigue physiologique sur la
performance au 100m sprint. Ceci présente un constat intéressant
qui permet d'appréhender les causes et les effets de la fatigue sur la
performance, sans pour autant expliquer les interactions entre les
différents systèmes du corps humain. En effet, le système
musculosquelettique est piloté par des commandes centrales,
c'est-à-dire par le SNC qui reçoit des afférences
relatives à la fatigue, i.e. des informations physiologiques,
biomécaniques, sensorielles et psychologiques. Dans le modèle
complexe [26], ces rétroactions sont envoyées par les
différents systèmes impliqués dans la réalisation
de l'exercice, sont centralisées et intégrées par le
système nerveux et enfin transformée de façon inconsciente
en sensation de fatigue.
Les résultats de notre étude sont à
compléter avant de pouvoir être publiés et
généralisés. Les données qu'y ont été
recueillies présentent un échantillon de 8
sprinters. Des études neuromusculaire et psychologique
s'avèrent nécessaires pour compléter notre
interprétation de la fatigue et répondre aux questionnements
suivants:
- Comment sont traitées les différentes
rétroactions dans le système nerveux ?
- Quels sont les modes et les délais d'ajustement de
l'effort ?
Pour répondre à cela, nous proposons d'utiliser
notre méthode d'expérimentation et de la compléter par des
enregistrements électromyographiques (« EMG ») de
l'activité musculaire des membres inférieurs et une étude
de la poulaine afin de comprendre l'adaptation des experts à l'effort et
à la fatigue.
Bibliographie
1. Alexander, R. McN. (1990). Optimum take-off techniques for
high and long jump. Philosophical Transactions of the Royal Society of
London, 329(B), 3-10.
2. Alexander, R. (1992). A model of bipedal locomotion on
compliant legs. Philosophical Transactions of the Royal Society of
London, 338(B), 189- 198.
3. Billat, V. (1996). Use of blood lactate measurements for
prediction of exercise performance and for control of training. Sports
Medicine, 22(3), 157-175.
4. Bishop, M., Fiolkowsski, P., Conrad, B., Brunt, D.,
Horodyski, M. (2006). Athletic footwear, leg stiffness, and running kinematics.
Journal of athletic training, 41(4), 387-392
5. Blickhan, R. (1989). The spring-mass model for running and
hopping. Journal of Biomechanics, 22, 1217-1227.
6. Bret, A., Rahmani, A., Messonnier, L., Bourdin, M., Bedu,
E., Lacour, J.R. (2001). Relation entre la concentration sanguine en lactate
mesurée en fin de compétition et la performance sur 100m.
Science & Motricité, 42, 24-28.
7. Bret, C., Rahmani, A., Dufour, A.B., Messonnier, L.,
Lacour, J.R. (2002). Leg strength and stiffness as ability factors in 100m
sprint running. Journal of Sports Medicine & Physical Fitness, 42,
274-81.
8. Cerretelli, P. (2002). Traité de physiologie de
l'exercice et du sport. Paris : Masson.
9. Chelly, S.M., Denis, C. (2001). Leg power and Hopping
Stiffness: relationship with sprint running performance. Medicine and
Sciences in Sports and Exercise, 33(2), 326-333.
10. Dalleau, G., Rahmani, A., Verkindt, C. (2007). Relation
entre la puissance et la raideur musculotendineuse chez les sportifs de haut
niveau. Science & Sports, 22, 110-116
11. Di Prampero, P.E. Energetics of muscular exercise. (1981).
Reviews of Physiology, Biochemistry & Pharmacology, 89,144-222.
12. Farley, C.T., Morgenroth, D.C. (1999). Leg stiffness depends
on ankle stiffness during human hopping. Journal of Biomechanics, 32,
267-273.
13. Fujitsuka, N., Yamamoto, T., Ohkuwa, T. (1982). Peak
blood lactate after short periods of maximal treadmill running. European
Journal of Applied Physiology, 48, 289-96.
14. Gibson, A.S.C., Noakes, TD. (2004). Evidence for complex
system integration and dynamic neural regulation of skeletal muscle recruitment
during exercise in humans. British journal of sports medicine, 38,797-
806.
15. Hautier, C.A., Wouassi, D., Arsac, L.M., Bitanga, E.,
Thiriet, P., Lacour, J.R. (1994). Relationships between postcompetition blood
lactate concentration and average running velocity over 100m and 200m races.
European Journal of Applied Physiology, 68, 508-513.
16. Hirvonen, J., Rehunen, S., Rusko, H., Harkonen, M.
(1987). Breakdown of high energy phosphate compounds and lactate accumulation
during short supramaximal exercise. European journal of Applied
Physiology, 56, 253-259.
17. McMahon, T.A., Valiant, G., Frederick, E.C. (1987). Groucho
running. Journal of Applied Physiology, 62(6), 2326-2337.
18. McMahon, T.A., Cheng, G.C., (1990). The mechanics of
running: How does stiffness couple with speed? Journal of
Biomechanics, 23(1), 65-78.
19. Mero, A., Luthtanen, P., Viitasalo, J.T., Komi, P.V.
(1981). Relationships between the maximal running velocity, muscle fiber
characteristics, force production and force relaxation of sprinters.
Scandinavian Journal of Sports Sciences, 3(1), 16-22.
20. Mero, A., Komi, P.V., Gregor, R.J. (1992). Biomechanics of
running. Sports Medicine, 13(6), 376-92.
21. Morin, J.B., Belli, A. (2003) .Mechanical factors of 100m
sprint performances in trained athletes. Science & Sports, 18:
161-163.
22. Morin, J.B., Dalleau, G., Kyrolainen, H., Jeannin, T.,
Belli, A. (2005). A simple method for measuring stiffness during running.
Journal of Applied Biomechanics, 21, 167-180.
23. Morin, J.B., Jeannin, T., Chevalier, B., Belli, A.
(2006). Spring-Mass Model characteristics during sprint running: correlation
with performance and fatigue induced changes. International Journal of
Sports Medicine, 27, 158-165.
24. Nummela, A., Vuorimaa, T., Rusko, H. (1992). Changes in
force production, blood lactate and EMG activity in the 400-m sprint.
Journal of Sports Sciences, 10, 2 17-228.
25. Newsholme, E.A., Blomstrand, E., Ekblom, B. (1992).
Physical and mental fatigue: metabolic mechanisms and importance of plasma
amino acids. British Medical Bulletin, 48(3), 477-95.
26. Noakes, T.D., St Clair Gibson, A., Lambert, E.V. (2005).
From catastrophe to complexity: a novel model of integrative central neural
regulation of effort and fatigue during exercise in humans: summary and
conclusions. British Journal of Sports Medicine, 39, 120-124.
27. Schmidt, R.F. (1999). Physiologie. De Boeck
& Larcier: Paris, Bruxelles.
28. Zouhal, H., Moussa, E., Forichier, J.M.,
Gratas-Delamarche, A., Delamarche, P. (2002). Évolution de la
lactatémie lors d'une séance de vitesse chez des sprinters de
niveau national. Science & Sports, 17, 31-4.
Annexe 1
Schéma du métabolisme anaérobie du
glucose [Cerretelli (2002)]

ATP : adénosine-triphosphate, (-1 ATP) = 1 molécule
d'ATP utilisée, (+2) =2 molécule d'ATP produite.
Annexe 2
Calcul de la raideur mécanique par la
méthode sinusoïdale [Morin et al. (2005)]
- Liste des paramètres utilisés pour le
calcul de kleg et kvert :
|
Abréviation
|
Paramètres utilisés
|
Unité de mesure
|
|
Fmax
|
la force de réaction au sol
|
N (Newton)
|
|
G
|
accélération de la Pesanteur = 9.81 à
Paris
|
m.s-2
|
|
L
|
distance grand trochanter-sol (dans la position
debout)
|
m (mètre)
|
|
ÄL-1
|
la compression maximale du ressort (membre
inférieur)
|
M
|
|
M
|
la masse corporelle
|
kg (kilogramme)
|
|
V
|
la vitesse de course
|
m.s-1
|
|
tc
|
temps contact
|
ms (Millisecondes)
|
|
tf
|
temps d'envol
|
ms (Millisecondes)
|
|
Äyc
|
déplacement maximal du centre de masse pendant la
phase de contact (déterminé par double intégration de
l'accélération verticale par rapport au temps)
|
M
|
· Equation de calcul de la raideur verticale
(kvert) en kN.m-1 :
- 1
kvert = F ·Äy c
max
Avec ; F max =mg·(Ð
2)·[(t f t c )+1)]


2
Avec ; ÄL = L -
L2 - (vt c 2)2 + Äy
c


2
F t
Et, Ä = - ·
y + ·
max c c
t
g
c Ð 2
m 8
·
ÄL- 1
Equation de calcul de la raideur du membre
inférieur (kleg) en kN.m-1 :
max
kleg=F ·
| 

