Etude de l'effet de lisière sur la distribution spatio-temporelle des rongeurs dans un écosystème anthropisé en région tropicale: cas de la Réserve forestière de Masako à Kisangani en RDC( Télécharger le fichier original )par Jean Pierre Pitchou MENIKO TO HULU Université de Kisangani - Diplôme d'études supérieures en aménagement durable des forêts tropicales 2011 |

Fig 1.4 : Représentation schématique des trois mécanismes par lesquels les corridors facilitent la continuité entre population dans les habitats en tâches 9 Fig 1.5 : Représentation de la formation de lisière associée avec la fragmentation 10 Fig 3.1: Localisation de la Réserve Forestière de Masako 15 Fig 3.2 : Description du dispositif expérimental ..19 Fig 4.1 : Corrélation sur base de la vitesse du vent ..22 Fig 4.2 : Corrélation sur base de la température de l'air ..23 Fig 4.3 : Corrélation sur base de l'humidité relative de l'air 23 Fig 4.4 : Corrélation sur base des points de rosée 24 Fig 4.5 : Similarité des habitats en fonction de la composition floristique .26 Fig 4.6 : Comparaison des effectifs des rongeurs dans les trois habitats 31 Fig 4.7 : Comparaison des moyennes des Hybomys lunaris capturés dans les trois habitats .31 Fig 4.8 : Comparaison des moyennes des Hylomyscus spp capturés dans les trois habitats...32 Fig 4.9 : Comparaison des moyennes des Lophuromys dudui capturés dans les trois habitats 32 Fig 4.10 : Comparaison des moyennes des Praomys spp capturés dans les trois habitats 33 Fig 4.11 : Comparaison des moyennes des Stochomys longicaudatus capturés dans les trois habitats .33 Fig 4.12 : Corrélation des espèces avec les différents habitats 34 Liste des tableaux.
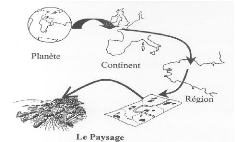
RésuméLa présente étude se propose d'analyser l'impact de l'anthropisation sur la population des Rongeurs en région tropicale. La relation faune - habitat par évaluation de la sensibilité des espèces aux effets de lisière a été établit. La démarche adoptée a conduit à la mise en place d'un dispositif de trois grilles de piégeages placées le long d'un transect de 500m traversant trois types d'occupations du sol (Jachère, Lisière et Forêt secondaire). Dans chaque grille, cinq layons étaient ouverts parallèlement à une équidistance de 10m pour le prélèvement des paramètres environnementaux et la capture des Rongeurs. La récolte des données s'était effectuée à Masako de Juin à Août 2010. Ces données ont été complétées par celles D'avril et Mai disponibilisées par le Doctorant Léon Iyongo encadreur de ce mémoire. L'étude a conduit à la capture de 348 Rongeurs repartis dans trois familles et douze espèces différentes. Les pièges Clapettes de type « Lucifer rat traps » et Sherman, ont servi pour cette fin. L'identification des espèces était faite sur base des seuls caractères morphologiques externes. Les abondances relatives des espèces des Rongeurs dans la zone d'étude et par habitat, les similarités entre les habitats sur base des données des présences et absences, la réponse des espèces à l'hétérogénéité d'habitats ainsi que les corrélations espèces - habitats ont été déterminées et analysées. Cinq espèces ont répondu à l'hétérogénéité d'habitats dont trois se sont montrées réellement sensibles aux effets de lisière. L'impact de l'anthropisation a été ressenti par la présence de deux individus de l'espèce Mastomys natalensis en jachère. Deux espèces de Rongeurs sont corrélées à la jachère, une à la lisière et enfin deux à la forêt secondaire. Deux espèces évitent la jachère et une évite la lisière. Le prélèvement des paramètres environnementaux (vitesse du vent, température, humidité relative, fraicheur de l'air, indice de chaleur et point de rosée) a été rendu possible grâce à l'anémomètre Kestrel 3000 de marque américaine. La description de la végétation dans chaque type d'occupation du sol à partir des données d'inventaires a été faite et les caractéristiques de chaque habitat mises en exergue. L'analyse des données a conduit à l'acceptation des hypothèses 1, 2, 3 et au rejet de l'hypothèse 4. Mots clés : Lisière, Rongeurs, Spatio- temporelle, Ecosystème, Anthropisé, Masako, Occupation du sol, Habitat, Sherman, Lucifer rat-traps, Typologie, Similarité. AbstractThis study aims to analyze the impact of urbanization on the population of Rodents in tropical regions. The relationship - wildlife habitat assessment of species sensitivity to edge effects has been established. The approach led to the establishment of a system of three trapping grids placed along a 500m transect through three types of land uses (Fallow, Forest Edge and secondary). In each grid, five transects were open along with a contour interval of 10m for the collection of environmental parameters and capture of rodents Data collection was conducted at Masako from June to August 2010. These data were supplemented by those of April and May availability by Leon Iyongo PhD supervisor of this thesis. The study led to the capture of 348 rodents in three families and twelve different species. Traps type valve "Lucifer Rat Trap" and Sherman, were used for this purpose. Species identification was made based only on external morphological characters. The relative abundances of species of rodents in the study area and habitat similarities between habitats based on attendance and absence data, species response to the heterogeneity of habitats and species correlations - habitats have been determined and analyzed. Five species have responded to heterogeneity of habitats, three of which are designed really sensitive to edge effects. The impact of human impact has been felt by the presence of two individuals of the species Mastomys natalensis in fallow. Two species of rodents are correlated with fallow, on the edge and finally two in secondary forest. Both species avoid the fallow and avoids the edge. The removal of environmental parameters (wind speed, temperature, relative humidity, fresh air, heat index and dew point) was made possible by the anemometer Kestrel 3000 American brand. The description of vegetation in each type of land from the inventory data was made and the characteristics of each habitat highlighted. Data analysis led to the acceptance of hypotheses 1, 2, 3 and the rejection of Hypothesis 4. Keywords: Edge, Rodents, Space-temporal ecosystem, human impact, Masako, Land Use, Housing, Sherman, Lucifer rat-traps, Typology, Similarity. APERÇU, CONTEXTE ET PROBLEMATIQUELa connaissance de la diversité biologique d'un territoire est la première étape à franchir dans toute tentative de gestion responsable et durable de ses ressources biologiques. En ce début du 21ème siècle, l'attention du monde quant à son avenir est entre autres focalisée sur la dégradation accélérée de l'environnement et donc de la biodiversité qui semble hâter le processus de modification du climat et des habitats et amplifier l'indisponibilité des ressources biologiques (Katuala, 2009). Richard Fays (2009) considère que les forêts tropicales, et plus particulièrement les forêts denses humides, abritent plus de 50% voire 80% de la diversité terrestre. Cette richesse est aujourd'hui menacée et selon la FAO (2005), le quart de la diversité biologique de la planète risque de disparaître d'ici à 2020. La dégradation et la destruction des espaces forestiers additionnées à l'effet des pollutions de l'air et de l'eau et aux effets négatifs d'une chasse non contrôlée en constituent les principales causes. La pratique de la culture itinérante sur brûlis, encore fréquente en Afrique équatoriale, est une des causes importantes de la déforestation dans les régions où une pression démographique excessive a fait accélérer le rythme des rotations agricoles. Alors qu'il faut une vingtaine d'années pour la restauration de la forêt secondaire, bien souvent la mise en jachère aujourd'hui est inférieure à sept ans (Ramade, 1984). La principale cause de déforestation mais aussi la principale menace, en République Démocratique du Congo (RDC) est donc l'agriculture vivrière. Le développement envisagé des plantations industrielles (palmiers à huile notamment) fait planer une menace supplémentaire sur le massif forestier de la RDC (Duveiller et al, 2008). Les écosystèmes forêts tropicales malgré leur grande complexité, leur grande diversité et leur grande richesse de formes biologiques (Wilson, 1988) n'échappent pas à la déforestation. Ils sont aussi très menacés de dégradation suite aux divers facteurs, les uns intrinsèques tels que le niveau de réchauffement général de la planète (Leroux, 2004), et les autres autochtones, comme les types d'activités anthropiques qui y sont pratiquées : activités agricoles, notamment l'agriculture itinérante sur brûlis (Mate, 2001), exploitation du bois sous diverses formes (bois de chauffe, bois d'oeuvre et d'industrie), cueillette et chasse, urbanisation, installation des infrastructures de développement, exploitation minière, etc (Juo & Wilding, 1996 ; Katuala, 2005). Le climat est un facteur majeur qui contrôle la structure globale de la végétation et du sol, leur productivité et la composition spécifique de la faune et de la flore (Gitay et al, 2002). En outre, il est avéré que l'évolution du monde animal a de tout temps été influencée par l'environnement abiotique et le climat y a joué un rôle fondamental (Gautier- Hion et al, 1999). Tout changement important de climat affecte directement les fonctions des organismes individuels (croissance), modifie les caractéristiques des populations (importance numérique) et change la structure, la fonction des écosystèmes (composition des espèces et leurs interactions) et leur distribution dans le paysage. Cela aboutit à la sélection génétique, à des adaptations spécifiques, à la spéciation et à l'innovation (De Menocal, 2004). Dans les écosystèmes forestiers tropicaux d'Afrique, en raison du changement climatique mondial et de la déforestation croissante liée aux activités anthropiques (Juo & Wilding, 1996 ; Querouil, 2001 ; Katuala, 2005), un intérêt croissant s'observe face au problème de la perte (modification) de la biodiversité. C'est ainsi que des études directement ou indirectement liées à ce phénomène sont de plus en plus menées en vue de connaître la biodiversité des habitats et son évolution dans l'espace et dans le temps à la lumière de la dégradation et de la fragmentation accélérée des écosystèmes. Pourtant il est reconnu dans le bassin du Congo, que la vaste majorité de la population rurale dépend des forêts pour sa vie quotidienne. Ces gens puisent dans la forêt l'essentiel de leurs protéines, médicaments, énergie, matériaux et revenu (Cifor, 2007). La Réserve de Masako qui fait l'objet de notre étude n'est pas épargnée de cette pression anthropique. Elle subit une pression de plus en plus croissante de la part de la population environnante, toujours en quête de nouvelles terres plus fertiles. L'agriculture itinérante sur brûlis y est pratiquée abondamment. Cette pratique culturale favorise la fragmentation des habitats forestiers et la multiplication des habitats de lisière (Iyongo, 2008). Elle peut aussi reproduire les effets des catastrophes naturelles et contribuer à créer et agrandir les vides (Ngo Bieng, 2004). Dans le cadre du Projet Interuniversitaire Ciblé : Aménagement des Forêts Congolaises « AFORCO » financé par la Coopération Universitaire pour le Développement (CUD), l'écologie du paysage y est un des domaines clés où les recherches se focalisent sur les effets de lisière et la distribution spatiale des espèces car la fragmentation des habitats est reconnu comme étant un des phénomènes majeurs ayant marqué la plupart des paysages au cours du 20ème siècle (Paillat et Butet, 1994). Ce morcellement des habitats en taches dans l'espace conditionne l'organisation spatiale des populations, affectant par conséquent les processus écologiques. Cette étude qui cadre dans ce domaine de l'écologie du paysage voudrait se consacrer à l'analyse des effets de lisière sur la distribution spatio - temporelle des Rongeurs pour une meilleure connaissance de l'impact de la fragmentation et de l'anthropisation à Masako PARTIE I : INTRODUCTION GENERALE1.1. ECOLOGIE DU PAYSAGE.Toute discipline scientifique nait et se développe dans la continuité des théories et des méthodes antérieures, en s'y appuyant pour les réfuter ou pour les dépasser. L'écologie du paysage ne fait pas exception à cette continuité de l'histoire des sciences. 1.1.1. Définition et ImportanceL'écologie du paysage est une science qui décrit la structure, le fonctionnement, les qualités, les fonctions et la dynamique spatio-temporelle des paysages. Elle s'intéresse à des systèmes posés d'emblée comme spatiaux et hétérogènes (Baudry, 1988) et peut apporter énormément de la compréhension des causes et conséquences de la transformation des paysages (Decamps, 2004). Elle est souvent définie comme la réunion de la géographie et de l'écologie et en tant que science, elle décrit et cherche à expliquer (Baudry, 1998 ; Gecopa, 2004) : - La structure des paysages (leur compositions et configuration) ; - Le fonctionnement des paysages (processus écologiques qui s'y déroulent) ; - Les qualités écologiques, économiques et sociales des paysages ; - La dynamique spatio-temporelle des paysages. Pour ces raisons, l'écologie du paysage fait appel à plusieurs disciplines telles que l'architecture du paysage, les sciences naturelles, la géographie, et les sciences sociales. Elle s'interroge sur la façon de cerner l'étude des systèmes complexes. D'où la notion de la complexité y est indissociable de celle de la perception (Cirad, 2001). 1.1.2. Historique de la discipline et son évolutionLe terme écologie, proposé en 1866 par Haeckel, biologiste allemand, désigne la science qui étudie les relations des êtres vivants avec leur milieu. Etymologiquement, il associe les mots grecs oikos et logos et signifie science de l'habitat. Pour pouvoir répondre aux questions des associations de protection de la nature concernant les effets de la fragmentation forestière sur le déclin des populations animales, et pour apporter des éléments de réponse relatifs aux conséquences écologiques des transformations de l'espace agricole, la communauté scientifique a dû évoluer et changer d'objet d'étude. L'étude des écosystèmes a fait place à l'étude des systèmes plus complexes pour laquelle la pluridisciplinarité s'est avérée indispensable. Les écologues se sont alors rapprochés de la communauté des biogéographes issus de la pensée de Troll qui créa le premier le terme Ecologie du paysage : << Landschaftökologie >> en 1939 (Bogaert et Mahamane, 2005) et il y a eu une volonté-nécessité-de partager les concepts, des outils et de méthodes. De ces échanges est née l'écologie du paysage telle qu'elle se développe actuellement rassemblant une gamme étendue des vues, des théories et des méthodologies (Bastian, 2001) et c'est cela qui fait sa force (Wiens, 1999). 1.2. LE PAYSAGE1.2.1. Origine, définitions, typologie et échelle.L'origine du terme << landscape >> ou << paysage >> serait la juxtaposition de deux mots : land qui est une portion délimitée de territoire et Scape qui signifie un assemblage d'objets similaires (Burel & Baudry, 2000). La définition du paysage que nous retiendrons par la suite de ce mémoire est la synthèse des définitions de Bertrand (1975) et de Forman & Godron(1986) qui considèrent que le paysage est un niveau d'organisation des systèmes écologiques, supérieur à l'écosystème ; il se caractérise pour partie par les activités humaines. Des différentes approches qui ont été développées en écologie du paysage, on peut en dégager trois principales qui ne s'excluent pas réciproquement, mais sont plutôt complémentaires (Iorgulescu & Schlaepfer, 2002) : - Une première approche selon laquelle le paysage est déterminé par les facteurs environnementaux (climat, topographie, etc.) ou par les facteurs dépendant de l'interaction de ces facteurs environnementaux (végétation, hydrologie, etc.) ; à ces facteurs on peut également ajouter l'impact anthropique ; - Une approche basée sur la perception de l'environnement par un organisme vivant quelconque ; - Une approche centrée sur la perception de l'environnement par l'homme ; De la définition du paysage telle que ci-haut adoptée et qui met en avant plan l'hétérogénéité et la dynamique des systèmes, peut s'appliquer une très grande gamme d'échelles, du continent au microsite (figure1).
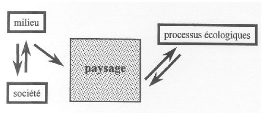
Fig 1.1 : Le paysage, niveau d'organisation des systèmes écologiques situé au dessus de l'écosystème, mais en dessous de la région et du continent (d'après Forman, 1995 dans Burel & Baudry, 2000). 1.2.2. Processus écologique se déroulant au sein de paysageLes sciences humaines : agronomie, géographie, histoire définissent la structure du paysage et sa dynamique en fonction de l'histoire passée et récente des sociétés (Burel & Baudry, 2000). Ainsi, la dynamique des paysages dépend des relations entre les sociétés et leur environnement, le paysage devient donc la résultante d'une dynamique entre le milieu physique et l'activité des sociétés humaines qui s'y développent. Par conséquent, l'organisation et la dynamique du paysage resteront en interaction constante avec les processus écologiques (figure2).
Fig 1.2 : Démarche scientifique adoptée en écologie du paysage : le paysage est la résultante de la dynamique du milieu et de la société qui s'y développe (Burel & Baudry, 2000) 1.3. PATTERN/PROCESS PARADIGM1.3.1. Fonctionnement d'un écosystèmeAu sein d'un écosystème, la dynamique de chaque individu est fortement influencée par les interactions entre cet individu et les autres éléments de l'écosystème (Begon et al, 1990). Ainsi pour un peuplement forestier, la structure spatiale joue un rôle clé dans sa dynamique. Elle décrit les relations de voisinages entre les individus et prend en compte autant les dimensions des individus que les relations spatiales entre les individus (Bouchon, 1979). Elle détermine en particulier l'environnement local autour de chaque arbre et donc ses conditions de croissance (Goreaud, 2000). Cet environnement local modifie l'expression des processus naturels comme la croissance, la mortalité et la régénération du peuplement. Inversement, ces processus naturels modifient à leur tour la structure spatiale, qui peut aussi être influencée par des actions anthropiques (Barot et al, 1999 dans Ngo Bieng, 2004). 1.3.2. Processus et structure spatiale d'un paysageL'importance de la structure spatiale des écosystèmes paysagers pour éclairer les processus écologiques est reconnue par la communauté écologique (Fortin, 2002 in Bogaert & Mahamane, 2005). Chaque système écologique est
caractérisé par une interdépendance de trois
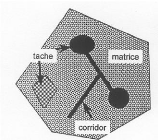
éléments clés : sa car les zones initialement couvertes par la forêt seront remplacées par une autre classe d'occupation du sol (Bogaert & Mahamane, 2005). En analysant les structures du paysage et leur dynamique, des déductions utiles au sujet des processus (écologiques) fondamentaux peuvent être faites, et vice versa (Bogaert et al, 2004). Cela est connu sous le terme « pattern/process paradigm >> et est une hypothèse centrale de l'écologie du paysage, aussi souvent définie pour cette raison comme « une branche de la science développée pour étudier les processus écologique dans leur contexte spatial >>. (Antrop, 2001 ; Stine & Hunsaker, 2001 in Bogaert & Mahamane, 2005). 1.4. CATEGORIES D'ELEMENTS DU PAYSAGE1.4.1. Tache, corridor et matriceDans un des textes fondateurs de l'écologie du paysage, Forman & Godron (1981) ont proposé une distinction entre les différents éléments que l'on peut distinguer dans un paysage (figure 3). La matrice est l'élément dominant, il est le type ou classe (ensemble des tache ayant des caractéristiques similaires pour le processus considéré, le plus répandu et le moins fragmenté (Iorgulescu & Schalaepfer, 2002) englobant en son sein des taches (bosquets, habitation) et des corridors, éléments linéaires résultant généralement des activités humaines. L'ensemble des taches constitue une mosaïque et l'ensemble de corridors, un réseau. Au sein des taches (et des corridors), on peut distinguer une lisière qui a de très fortes interactions avec la matrice ou les taches voisines, et un milieu intérieur dans lequel les interactions sont très faibles ou nulles. Plus les taches sont allongées, plus le ratio lisière/intérieur est élevé.
Fig 1.3 : Les catégories d'éléments du paysage. La structure du paysage peut être représentée comme un ensemble de tâches, éventuellement liées par des corridors. La structure englobant ces deux catégories est appelée la matrice, qui constitue « l'arrière-plan » du paysage. D'après Forman & godron (1986). Les habitats corridors peuvent dans leur forme, leur couvert végétal, leur disposition dans l'espace, produire des conditions du milieu hétérogène. Leur utilisation par les petits mammifères dépendra donc des modalités et des potentialités de réponses des différentes espèces à cette hétérogénéité. En facilitant les mouvements entre taches et en fournissant des habitats supplémentaires, ils seraient donc des éléments essentiels pour le maintien des populations à l'échelle des paysages (Paillat & Butet, 1994). Bennett (1990) distingue pour les petits mammifères trois types de mouvements à travers les corridors (figure 4).
Fig 1.4 : Représentation schématique des trois mécanismes par lesquels les corridors facilitent la continuité entre population dans les habitats en tâches : a- mouvement direct par un individu, b- mouvement d'un individu, ponctué par des pousses, c- flux de gènes à travers une population résidence reproductrice dans le corridor.D'après (Bennet, 1990). 1.4.2. Concepts d'écotone et typesEtymologiquement, le mot écotone est grec, il est composé d'oikos, maison et tonos, tension. Un écotone est une zone de transition écologique entre deux écosystèmes. Par exemple, le passage de la savane à la forêt. Selon les auteurs (Clements, 1905 ; Odum, 1971 ; Shelford dans Di Castri, 1981 ; Wiens et al, 1985 ; Risser, 1989, Baudiere et Gauquelin, 1990 ; Kolasa & Zalewski, 1995) on distingue trois principales approches de l'écotone : - L'approche descriptive de l'écotone ; - L'approche fonctionnelle de l'écotone ; et - L'approche dynamique de l'écotone. Se référant à Naiman & Decamps (1991) dans Iyongo (2008), trois principaux types d'écotones se distinguent : > Les lisières : ce sont des limites entre les formations forestières et les formations herbacées ou les landes. Cas de notre étude dans
la Reserve Forestière de Masako ; cède la place à une végétation uniquement herbacée ; > Les ripisylves : elles se définissent comme étant des écotones terre-milieux aquatiques. 1.5. LISIERE ET INTERIEURLes lisières jouent un rôle dans le mouvement des individus disperseurs : elles sont plus ou moins perméables en fonction de leur environnement immédiat, de leur structure et de l'espèce considérée. Elles peuvent être perçues comme des discontinuités ou comme des gradients marqués. Une lisière large peut être perçue comme une zone de transition par un organisme très mobile qui la traverse en peu de temps, alors qu'un autre organisme peu mobile, peut percevoir cette lisière comme une tache avec des limites étroites. Les organismes relativement sédentaires, parce qu'ils rencontrent peu de lisières dans leurs mouvements, peuvent percevoir des différences entre les taches comme des lisères plus marquées que les organismes plus mobiles.
Fig 1.5 : Représentation de la formation de lisière associée avec la fragmentation. Quand l'habitat original (a) est fragmenté, quatre fragments sont observés (b), chacun caractérisé par un intérieur et un habitat de lisière. La surface cumulative de lisière de (b) peut dépasser la surface de lisière de (a), pendant que la surface totale de l'intérieur est plus petite que dans (a). Une perturbation centripète avec profondeur de pénétration constante perpendiculaire à tous les côtés de la tâche est supposée. (Bogaert, 2000). 1.6. LES RONGEURS DANS LA RESERVE FORESTIERE DE MASAKO1.6.1. Pourquoi étudier les Rongeurs ?Les Rongeurs constituent une des composantes importante de la faune des écosystèmes terrestres totalisant près de 42% des espèces de Mammifères dans le monde (Dieterlen in Katuala, 2007 ; Happold ; 1996). Ils jouent un rôle important dans le fonctionnement des écosystèmes en général, et celui des forêts en particulier et forment l'un des ordres les plus diversifiés de la classe des Mammifères de la RD Congo (Dudu & Gevaerts, 1987). Ils sont un maillon important dans les réseaux trophiques (Carey & Johnson dans Katuala, 2009). Ils constituent parfois une ressource alimentaire importante notamment dans plusieurs contrées africaines (Wetsi et al ; 1988). Enfin, à l'instar d'autres petits mammifères, les Rongeurs conviennent pour évaluer le rôle relatif du changement des conditions écologiques et de la fragmentation des habitats, dans la mesure où des évidences paléontologiques et écologiques indiquent que beaucoup d'espèces de petits Rongeurs ont une forte association avec des habitats particuliers, de sorte que des changements dans la structure des habitats conduisent à des modifications spécifiques des Rongeurs (Nicolas et al dans Katuala, 2009). 1.6.2. Richesse spécifique de MasakoLa richesse spécifique des Rongeurs à Masako s'élève à 28 espèces. Les Muridés constituent la plus riche famille du peuplement avec 17 espèces. Parmi les Rongeurs de Masako, une à deux espèces dominantes se présentent, Praomys jacksoni et Hybomys lunaris ; celles-ci sont accompagnées de trois espèces très abondantes, Deomys ferrugineus, Hylomyscus stella et Lophuromys duduii. La composition numérique des espèces de Rongeurs à Masako suit la structure générale de la composition d'abondances dans des peuplements de vertébrés en forêts tropicales en général et en forêts équatoriales africaines en particulier, où quelques espèces très nombreuses cohabitent avec un grand nombre d'espèces moins abondantes (Dudu, 1991). Partie II : HYPOTHESES, INTERETS ET OBJECTIFS DE L'ETUDEII.1. HYPOTHESES DE L'ETUDENotre compréhension des processus d'anthropisation/et ou de fragmentation actuellement à l'oeuvre dans la Réserve Forestière de Masako et qui conduit à la multiplication des zones lisières, nous amène à émettre quelques hypothèses sur la distribution spatio-temporelle des Rongeurs, que nous pouvons ensuite tester sur notre dispositif. Ces hypothèses sont : - La distribution spatio-temporelle des Rongeurs serait conditionnée par les facteurs environnementaux (température, humidité relative etc.) caractérisant différents habitats considérés ; - La lisière se caractériserait par une abondance et une composition de Rongeurs différentes des zones intérieures ; - Certaines espèces de Rongeurs forestiers de Masako seraient sensibles aux effets de lisière (espèces indicatrices) et d'autres non ; - Il existerait des espèces de Rongeurs inféodées à chaque type d'habitat (Jachère, lisière, forêt secondaire). II.2. OBJECTIFS DE L'ETUDELa présente étude qui cadre avec les relations « faune-habitats » en milieu forestier tropical vise, en s'appuyant sur les facteurs environnementaux, à évaluer le degré de sensibilité des Rongeurs forestiers face aux effets de lisière. L'objectif global étant d'analyser la composition et la distribution des populations des Rongeurs en relations avec l'hétérogénéité d'habitats dans un écosystème anthropisé de la Réserve Forestière de Masako. Cela aboutira à une meilleure connaissance du fonctionnement de l'écosystème et de sa réponse aux processus de fragmentation sur la biodiversité des Rongeurs à l'échelle de la réserve. De cet objectif général, découlent les objectifs spécifiques ci-après :
II.3. INTERETLa connaissance des modifications des facteurs environnementaux et de l'impact de l'anthropisation associé aux effets de lisière sur la composition et la distribution spatiotemporelle des petits mammifères, notamment les Rongeurs de la Réserve Forestière de Masako, constitue tout simplement les buts social, économique et écologique de cette étude. Partie III : MILIEU D'ETUDE, MATERIELS ET METHODESIII. MILIEU D'ETUDEIII.1.1. Choix et localisation du siteLe choix de la Réserve Forestière de Masako comme cadre pour la réalisation de cette étude a été motivé par les raisons suivantes : - La Réserve Forestière de Masako compte tenu de son statut d'air protégée jouit du privilège de constituer une étendue proche de la ville de Kisangani susceptible d'offrir, pour des recherches écologiques d'une longue durée, une forêt plus ou moins intacte (Dudu, 1991 ; Upoki, 1997 ; Upoki, 2001) ; - La présence d'une mosaïque d'habitats probablement en interaction ; - La présence d'une gamme des données sur sa faune et sa végétation issues des recherches antérieures devant faciliter une bonne comparaison des résultats ; - La présence d'un bon écosystème pour une étude des rongeurs en milieu fragmenté ; - L'accessibilité de la réserve à partir de la ville de Kisangani facilitant le ravitaillement en denrées pendant la période de récolte des données ; - La présence d'un gite aux conditions internationales offrant des meilleures possibilités de logement et de travail (Mensuration, Prélèvement d'organes en biopsie, formalisation, etc.). La Réserve Forestière de Masako est située à 14 Km au Nord-Est de la Ville de Kisangani, sur l'ancienne route Buta. Elle a une superficie de 2.105 hectares et est entièrement comprises dans une grande boucle de la rivière Tshopo (Juakaly, 2002 ; Mukinzi et al 2005 ; Iyongo, 2008). Elle est une propriété du Ministère de l'Environnement, Conservation de la Nature et Tourisme, créée par l'ordonnance-loi n°052/378 du 12 novembre 1953.
Fig 3.1: Localisation de la Réserve Forestière de Masako. La carte montre l'emplacement de la Réserve Forestière de Masako située à proximité de la Ville de Kisangani, dans la Province Orientale, en République Démocratique du Congo. Elle est délimitée dans sa grande partie par la grande boucle de la rivière Thopo. III.1.2. HydrographieLe réseau hydrographique de la Réserve de Masako est dominé par une seule grande rivière, la Tshopo, et la présence de 13 ruisseaux qui s'y déversent tous. Parmi eux, nous pouvons citer Amakasampoko, Masanga-Mabe, Magima, Amandje, Mayi ya chumvi, Masanga - mabe et Masako le principal ruisseau ayant donné son nom à la Réserve (Iyongo, 1996 ; Juakaly, 2007). III.1.3. Situation climatiqueLe climat de la région étudiée est globalement celui de Kisangani, sauf quelques petites modifications dues à la présence du couvert végétal (Iyongo, 2008). Selon Goffaux (1990), cette région est caractérisée par un climat du type << Afi >> de la classification de Köpen. Dans cette classification, << A >> désigne un climat chaud avec les moyennes mensuelles supérieurs à 18°C ; << f >> le climat humide dont la pluviosité est répartie sur toute l'année ; c'est-à-dire sans saison sèche absolue et dont la hauteur mensuelle des pluies du mois le plus sec est supérieur à 60 mm et << i >> signale une très faible amplitude thermique (Nyakabwa, 1982 ; Upoki, 2001, Juakaly, 2002 et Juakaly, 2007). Les tendances saisonnières de la région de Kisangani sont consignées dans le tableau 3.1 reprenant les données climatiques disponibles de vingt quatre années récentes (Soki et Upoki, dans Juakaly, 2007). Tableau 3.1 : Moyennes mensuelles de température (°C), précipitation (mm) et de l'humiditérelative (%) de la région de Kisangani
Saison S1 S2 S3 S4 Légende : Tm : Température moyenne mensuelle HRm : Humidité relative moyenne mensuelle PPm : Précipitation moyennes mensuelles S1; S2 ; S3 ; S4 : Saisons. Le tableau ci-haut révèle que : v' La température varie entre 25,3°C en Mars et 23,5°C en Août avec une moyenne annuelle de 24,4°C ; v' L'humidité de l'air varie entre 79,1% en Février et 87,3% en Juillet avec une moyenne annuelle de 84,0% ; v' Les précipitations sont abondantes toute l'année, avec une hauteur moyenne annuelle de 1782,7 mm. On observe un déficit pluviométrique en Janvier (69,5 mm) et Juillet (95,9 mm), périodes qui correspondent aux saisons subsèches de notre région. Les maxima sont constatés en Mai (178,7 mm) et en Octobre (237,4 mm), mois qui correspondent aux périodes des grandes pluies à Kisangani. Ces précipitations montrent une certaine tendance qui renseigne sur les périodes suivantes : - Décembre, Janvier, Février : première période subsèche, S1 = 275,6 mm ; - Mars, Avril, Mai : Première période des pluies ; S2 = 494,3 mm ; - Juin, Juillet, Août : Deuxième période subsèche, S3 = 355,2 mm ; - Septembre, Octobre, Novembre : Deuxième période des pluies, S4 = 675,6 mm. III.1.4. VégétationFox & Fox dans Iyongo (2008), signalent que les plantes nourrissent la plupart des Rongeurs et entretiennent des microclimats propices à la vie de certains micromammifères. Dudu & Gevaerts (1987) ajoutent que la structure de la végétation influence fortement le choix des micromammifères, de leur habitat et de celui de leur nid ou gite. La végétation de la Réserve Forestière de Masako est constituée principalement d'une forêt primaire à Gilbertiodendron dewevrei (Caesalpiniaceae), des forêts secondaires, des forêts marécageuses et des jachères. La forêt primaire est caractérisée par 4 strates : la strate arborescente supérieure, la strate arborescente inférieure, la strate arbustive et la strate herbacée. On y rencontre des espèces telles que : Geophila renaris, Polyalthia suaveolen, Staudtia gabonensis, Scaphopetalum thonneri etc. La forêt secondaire est subdivisée en forêt secondaire vieille et en forêt secondaire jeune. La forêt secondaire veille est une forêt transitive pré climacique, suite à la densité de ses éléments caractéristiques. Elle se caractérise par 3 strates : La strate arborescente, la strate arbustive et la strate herbacée. Quelques espèces caractéristiques sont : Cynometra hankei, Petersianthus macrocarpus, Trichilia welwitschii, Barteria nigritiana, Palisota schweinfurthii, Haumannia leonardiana etc (Makana 1986 ; Kahindo, 1988; Mambangula, 1988 et Mabay, 1994). Les jachères sont constituées selon Dudu (1991) de deux principales associations suivantes : - Association à Aframomum laurentii et Costus lucanusianus avec Costus lucanusianus, Funtumia elastica, Dichapetalum mombuttense, etc. - Association à Triumfetta cordifolia et Selaginella myosurus avec Triumfetta cordifolia, Selaginella myosurus, Trema orientalis etc. III.1.5. Activités humainesD'une manière générale, l'activité principale du paysan de Masako est l'agriculture itinérante sur brûlis. La mise en jachère dépasse rarement sept ans, si bien que le sol subit une forte pression dont l'issu est encore inconnu à ce jour (Juakaly, 2002). En plus de l'agriculture, les paysans s'adonnent aussi à la fabrication des braises, une autre activité destructrice des forêts. Il trouve dans la forêt le nécessaire pour sa survie, notamment : bois de chauffe, bois de construction. Suivant les saisons, la population s'occupe de la cueillette des champignons et des végétaux comestibles, du ramassage des chenilles et des escargots, de la chasse, de la pêche et de la fabrication du vin de palme à partir d'Elaeis guineensis ou de Raphia gilletii. Souvent, les arbres à chenilles et ces palmiers sont simplement abattus pour faciliter les opérations de ramassage des chenilles et la fabrication du vin. III.2. MATERIEL BIOLOGIQUE ET EQUIPEMENTS TECHNIQUESIII.2.1. Matériel biologiqueLe matériel biologique de cette étude était principalement constitué des espèces végétales récoltées ainsi que des Rongeurs capturés au sein des différents habitats. III.2.2. Equipements techniquesLes équipements ci-après nous ont été d'un grand secours pour la réalisation des travaux de terrain (récolte des données). Il s'agit de : - Un GPS OREGON 300 pour la prise des coordonnés géographiques du site d'étude ; - Un pentadécamètre pour la mesure de la longueur du transect ; - Trois machettes pour le nettoyage du transect et des layons de piégeages ; - Un anémomètre de marque Kestrel 3000 pour la prise des paramètres environnementaux ; - Une montre de marque SEICKO pour noter l'heure de prise des paramètres environnementaux ; - Les pièges « clapettes » de marque Lucifer et les pièges Sherman pour la capture des Rongeurs ; - Les sachets rayés bleu-blancs pour la signalisation des stations de piégeages le long des layons ; - Les noix de palme comme appât pour attirer les Rongeurs vers les pièges. III.3. METHODESIII.3.1. Descriptions du dispositifTrois grilles de piégeages installées respectivement dans 3 types d'occupation du sol (Jachère, Lisière et Forêt secondaire) par le doctorant Léon IYONGO, ont été utilisées. A l'intérieur de ces grilles de 1ha chacune étaient ouverts cinq layons ou transects secondaires de 200 m de long et équidistant de 10m, qui contenaient 100 pièges à raison de 50 pour chaque type (Clapettes de marque Lucifer et Sherman). Un transect principal de 500m de long allant de la jachère à la forêt secondaire en passant par la zone de lisière a été tracé en vue d'installer les grilles de piégeages et de prélever les différentes variables environnementales. La figure 3.1 donne la cartographie du site. L5 L4 Grille Forêt secondaire L3 Grille Lisière L3 50m L2 L1
200m
L2 L4 L1 200m L5
10m Fig 3.2 : Description du dispositif expérimental. On observe trois grilles de piégeages de 50 m chacune placées dans chaque type d'occupation du sol et équidistants de 200 m les unes des autres. Chaque grille contenaient cinq layons de 200 m de long et équidistants de 10 m le long desquels étaient piégés les Rongeurs. III.3.2. Technique de piégeageAu total 150 pièges de marque Lucifer rat trap et 150 pièges Sherman ont été utilisés soit 300 pièges de 2 types à raison de 100 pièges par grille et par type d'occupation du sol (50 Clapettes et 50 Sherman). Ces pièges ont été placés le long de 5 layons de 200m situés dans chaque grille, à une équidistance de 10 m. Un signalement de la station était à chaque fois noté et attaché à un arbuste comme repère pour faciliter les opérations de piégeages, de relevé et la détection des pièges. III.3.3. Prélèvement des paramètres environnementauxLes paramètres environnementaux tels que la vitesse du vent, la température, l'humidité relative, la fraicheur de l'air, l'indice de chaleur et le point de rosée ont été prélevés à l'aide d'un anémomètre de marque Kestrel 3000. Ces relevés s'effectuaient chaque jour entre 8h et 11h00 de l'avant midi et entre 14h et 17h de l'après midi. III.3.4. Analyses statistiques.Nous avons commencé par tester la distribution de nos données afin d'être fixé sur quel type de tests appliquer. Ce test a laissé voir une distribution gaussienne de nos données (annexe 2.a). Le test de corrélation a été effectué sur base des variables environnementales afin de mesurer la partie de la variance qui est partagée entre ces variables dans les habitats pris deux à deux. Une forte corrélation traduirait que les habitats étaient fortement liés et que ces variables variaient de la même façon à l'intérieur de ces habitats. A l'inverse, les variables diffèrent d'un habitat à l'autre, ce qui traduit qu'on est face à des milieux différents. Une Analyse de Variance à un critère a été effectuée et les p-valeurs on été calculés afin de déterminer si les effectifs étaient significativement différents entre les trois habitats ou pas. Ensuite, le test de Kolmogorov Smirnov a permis de comparer ces effectifs entre les habitats pris deux à deux afin de confirmer le résultat obtenu à l'aide de l'ANOVA. Enfin, le test t de Student a été réalisé pour tester les réponses des espèces à l'hétérogénéité d'habitats par comparaison de leurs effectifs dans les habitats pris deux à deux. Ces différents calculs ont été effectués à l'aide du logiciel STATISTICA. Le Test de Kolmogorov Smirnov est basé sur la comparaison de la fonction cumulative de fréquence N(x) pour l'échantillon (c'est-à-dire le nombre d'observations inférieures à x) avec la fonction de répartition F(x)pour la population (c'est-à-dire la probabilité qu'une observation soit inférieure à x). De façon plus précise on déterminera l'écart maximum en valeur absolue entre ces deux fonctions et on compare cet écart à des valeurs critiques tabulées. Il est d'une utilisation plus large que le test du Chi carré, car il reste applicable pour des échantillons d'effectif réduit. Le test t tient compte de l'importance de la différence, de la taille et de la déviation standard des échantillons. Le calcul de P valeur permet de répondre pour notre cas à la question : « A quel point sommes nous sûrs qu'il existe effectivement une différence entre les habitats ? » (Motulsky, 1995). III.3.5. Les coefficients ou indices de similaritéLe calcul des coefficients de similitude permet de quantifier le niveau de similitude entre deux sites. De nombreuses mesures des coefficients de similarité existent dans la littérature (voir Legendre & Legendre, 1998 et Magurran, 2004). On ne retiendra que deux parmi les plus fréquemment utilisés. 'V Le coefficient de Jaccard Cj : Il représente le nombre de cas de présence simultanée de deux espèces considérées, divisé par le nombre de cas où au moins l'une de deux est présente. Cj= a/a+b+c a = nombre d'espèces présentes dans les deux habitats ; b et c = nombres d'espèces absentes d'un des deux habitats ; 'V Le coefficient de Sorensen Cs : Il est similaire à celui de Jaccard, cependant il pondère par deux, le terme de co-occurrence. Cs=2a/ (2a+b+c) III.3.6. Typologie de réponse des espèces à l'hétérogénéité des habitats.Pour mieux interpréter les résultats des tests statistiques appliqués à nos données, nous nous sommes référés à la typologie des espèces face à l'hétérogénéité d'habitats (tableau 3.2) selon Iyongo et al (2009). Tableau 3.2 : Typologie de réponse des espèces à l'hétérogénéité d'habitats Types Habitat 1 Habitat 2 Habitat 3 Interprétations I a a a pas d'effet de lisière, espèce est généraliste ; pas de différence d'abondance entre les habitats. II a b a effet de lisière: si b > a, espèce préfère la lisière et si b < a, espèce évite la lisière. effet de lisière: espèce s'adapte différemment à chacun des III a b c habitats ; si b > a et b>c, espèce préfère la lisière ; si b < a et b< c, espèce évite la lisière. IV a ab b effet de lisière: l'abondance de l'espèce dans la lisière est intermédiaire de celles des habitats adjacents. V a a b pas d'effet de lisière : si a > b, espèce évite habitat 3 et si b > a, espèce cherche habitat 3 VI ab a b pas de détection d'effet de lisière. Partie IV : RESULTATSIV.1. Corrélation des habitats sur base des variables environnementalesSur un total de six variables étudiées, quatre ont montré un effet réel de l'hétérogénéité d'habitats liée aux actions anthropiques. Il s'agit de : la vitesse du vent, la température, l'humidité relative de l'air et le point de rosée. IV.1.1. Vitesse moyenne du ventL'analyse de la corrélation entre les 3 milieux sur base de la vitesse du vent est donnée dans la figure 4.1.suivante :
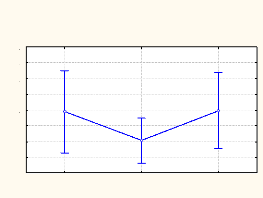
Vitesse moyenne du vent 0,07 0,06 0,05 0,04 0,03 0,02 0,01 0 a b c Jachère Lisière Forêt Grilles Fig. 4.1. Corrélation sur base de la vitesse du vent Cette figure laisse voir qu'il n'y a aucune corrélation significative entre les 3 milieux explorés du point de vue de leurs vitesses du vent (r2 = 0,1712 et p = 0,4148 entre la jachère et la lisière ; r2 = 0,4899 et p = 0,1215 entre la lisière et la forêt ; r2 = 0,0381 et p = 0,7110 entre la jachère et la forêt). Les vitesses varient d'un milieu à un autre avec la plus grande valeur dans la jachère, la valeur intermédiaire dans la lisière et la plus petite vitesse en forêt. IV.1.2. Température de l'airLa figure 4.2 donne la corrélation entre les trois habitats obtenue sur base de la température de l'air. Jachère Lisière Forêt secondaire Grilles
Temperatures ( °C ) 28,8 28,6 28,4 28,2 27,8 27,6 27,4 29 28 b a b Fig. 4.2. Corrélation sur base de la température de l'air On note de la figure 3.2 que la jachère est un milieu qui diffère de la lisière (r2 = 0,5204 ; p = 0,1056) et de la forêt secondaire (r2 = 0,2725 ; p = 0,2881) du point de vue de leurs températures. Par contre, la lisière et la forêt secondaire sont des habitats significativement corrélés (r2 = 0,8301 ; p = 0,0115). La température la plus élevée est enregistrée en forêt tandis que la plus faible est trouvée dans la jachère. IV.1.3. Humidité relative de l'airLes différentes valeurs du coefficient obtenues en mesurant l'humidité de l'air dans les trois milieux laissent voir une corrélation entre la lisière et les habitats adjacents. La jachère et la forêt secondaire n'étant pas significativement corrélées (Figure 4.3.).
a a b Jachère Lisière Forêt secondaire Grilles
94,5 94 93,5 93 92,5 92 91,5 91 Humidite relative de Pair (%) 90,5 90 89,5 Fig. 4.3. Corrélation sur base de l'humidité relative de l'air Il ressort de la figure 4.3.que l'humidité de l'air dans la jachère est différente de celle de la forêt secondaire (r2 = 0,4303 ; p = 0,1572). L'air paraît plus humide en forêt qu'en jachère. La zone de lisière est caractérisée par une humidité différente de celle de la forêt (r2 = 0,5044 ; p = 0,1138) et qui corrèle significativement avec celle de la jachère (r2 = 0,8711 et p = 0,0065 : très forte corrélation). IV.1.4. Point de roséeLa corrélation entre les habitats mesurée sur base de leurs points de rosée laisse voir que ces trois milieux ne sont pas corrélés (Figure 4.4.).
28,4 28,2 c b a 27,4 27,2 27 26,8 28 27,8 27,6 26,6 26,4 Jachère Lisière Forêt secondaire Habitats Point de rosee ( °C ) Fig. 4.4. Corrélation sur base des points de rosée On note de cette figure que les habitats exploités ne sont pas significativement corrélés (milieux différents) sur le plan de leurs points de rosée (r2 = 0,0754 et p = 0,5984 entre la jachère et la lisière ; r2 = 0,0935 et p = 0,5555 entre la lisière et la forêt ; r2 = 0,3297 et p = 0,2333 entre la jachère et la forêt). Les points de rosée varient d'un habitat à l'autre avec la plus grande valeur en forêt et la plus petite en jachère. La lisière compte une valeur située entre les deux extrêmes. IV.2. Description de la végétation de la zone d'étude.Une étude systématique de la végétation du milieu d'étude a été faite. Ainsi une tendance générale de la diversité du site en terme d'habitat est donnée des les lignes qui suivent.
Dans la jachère 189 individus on été identifiés dans 22 familles avec 31 espèces. Les dix familles les plus abondantes sont : Flacourtiaceae avec 23 individus, Arecaceae avec 22 individus, Malvaceae avec 17 individus, Euphorbiaceae avec 12 individus, Poaceae avec 12 individus, Apocynaceae avec 10 individus, Costaceae avec 8 individus, Commelinaceae avec 7 individus, Brommelinaceae avec 6 individus et Cannabaceae avec 5 individus. On y rencontre les espèces telles que : Rauvolfia vomitoria, Elaeis guineensis, Pteridium aquilinum, Alchornea cordifolia, Manihot esculenta, Palisota ambigua, Costus lucanusianus, Caloncoba welwitschii, Triumfetta cordifolia, Macaranga monandra... IV.3. Similarité entre habitats sur base de la végétationLa similarité entre habitats sur base de la composition floristique a été rendu possible par l'analyse factorielle des correspondances dont les résultats sont illustrés dans la figure 4.5 cidessous. PCA case scores
29.5
23.6
11.8 Axis 2
5.9
LF IF
-29.5 -23.6 -17.7 -11.8 -5.9 5.9 11.8 17.7 23.6 29.5
-5.9 -11.8 -17.7 -23.6 IJ
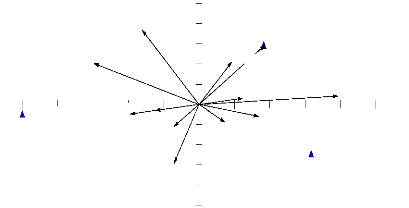
-29.5 Axis 1 Fig. 4.5. Similarité des habitats en fonction de la composition floristique Le résumé de la figure 4.5 indique que de manière générale les zones intérieures sont différentes du point de vue de la composition floristique. Les lisières (lisière jachère et lisière forêt) ont une composition floristique intermédiaire aux deux habitats mais beaucoup plus proche de celle de la forêt secondaire. IV.4. Situation des Rongeurs capturés.Le tableau 4.1 résume la composition des Rongeurs dans la zone d'étude Tableau 4.1: Inventaires des individus capturés
La lecture du tableau ci-dessus indique que 348 individus au total on été capturés pendant la période de récolte des données et dans les trois habitats concernés. L'espèce la plus abondante en termes de capture est le Praomys spp avec 145 représentants. Les espèces Graphiurus lorraineus et Oenomys hypoxanthus sont les moins représentées. La richesse spécifique de la zone d'étude est de 12 espèces des Rongeurs dont 10 de la famille des Muridae, 1 de la famille de Crecitidae et 1 de la famille de Gliridae. IV.5. Richesses spécifiques et abondance relative dans les habitats.IV.5.1. Fréquence relative des espèces par habitatLe tableau 4.2 donne les fréquences de capture des espèces de Rongeurs dans la jachère, lisière et forêt secondaire. Tableau 4.2: Fréquences des captures des espèces par habitat
Le tableau 4.2 fait remarquer que dix espèces se sont montrées présentes dans la jachère à l'exception de Malacomys longipes et Graphiurus lorraineus, huit dans la lisière exceptée Nannomys minutoides, Oenomys hypoxanthus, Mastomys natalensis et Graphiurus lorraineus et enfin neuf dans la forêt secondaire à l'exception de Lemniscomys striatus, Oenomys hypoxanthus et Mastomys natalensis. La forêt secondaire a regorgé le plus grand nombre d'individus, suivi de la jachère et enfin la lisière. IV.5.2. Abondance relative des espèces dans les trois habitats.Le tableau 4.3 donne l'abondance relative de chaque espèce pour chaque habitat.
Le tableau ci-dessus renseigne que les espèces Deomys ferrugineus, Hybomys lunaris et Praomys spp ont été les plus abondamment représentées dans la forêt secondaire ; Hylomyscus spp et Malacomys longipes dans la lisière ; Lophuromys dudui, Nannomys minutoides et Mastomys natalensis dans la jachère. Certaines espèces telles que Lemniscomys striatus et Stochomys longicaudatus sont bien représentées à chaque fois dans deux habitats. IV.5.3. Abondance relative des espèces dans la zone d'étude.La richesse spécifique par espèce et par habitat dans la zone d'étude est résumée dans le tableau 4.4. Tableau 4.4: Abondance relative des captures dans la zone d'étude
L'examen du présent tableau révèle que l'espèce Praomys spp est la plus abondante de toutes les espèces capturées dans la zone d'étude avec à elle seule 41,67% des individus du peuplement. Elle est suivie de Hylomyscus spp qui compte 13,79% des individus. Les espèces Graphiurus lorraineus et Oenomys hypoxanthus sont les moins abondantes voire rares de la zone d'étude. IV. 5.4. Richesse spécifique des habitats.IV.5.4.1. Forêt secondaire.La richesse spécifique dans cet habitat est de neuf espèces : Deomys ferrugineus, Hybomys lunaris, Hylomyscus spp, Lophuromys dudui, Nannomys minutoides, Praomys spp, Stochomys longicaudatus, Malacomys longipes et Graphiurus lorraineus. IV.5.4.2. Lisière.Dans la zone de lisière la richesse spécifique est de huit espèces : Deomys ferrugineus, Hybomys lunaris, Hylomyscus spp, Lemniscomys striatus, Lophuromys dudui, Praomys spp, Stochomys longicaudatus, Malacomys longipes. IV.5.4.3. Jachère.Dans la jachère enfin, la richesse en terme d'espèces est de dix espèces : Deomys ferrugineus, Hybomys lunaris, Hylomyscus spp, Lemniscomys striatus, Lophuromys dudui, Nannomys minutoides, Oenomys hypoxanthus, Praomys spp, Stochomys longicaudatus, Mastomys natalensis. IV.6. Similarité des habitats en fonction de la faune.Ces similarités entre habitats sont calculées sur base des données de présences et absences reprises dans le tableau 4.5. Tableau 4.5: Présences (+) et Absences (-) des espèces dans les habitats
Les tableaux 4.6 et 4.7 illustrent les degrés de similarité entre habitats, calculés respectivement avec le coefficient de Sørenson et celui de Jaccard. Tableau 4.6: Indice de Sørenson Habitats Jachère Lisière Forêt secondaire Jachère 1 0,77 0,73 Lisère 1 0,82 Forêt secondaire 1 Tableau 4.7: Indice de Jaccard Habitats Jachère Lisière Forêt secondaire Jachère 1 0,63 0,58 Lisère 1 0,70 Forêt secondaire 1 Les valeurs obtenues après calculs de ces deux coefficients révèlent que la lisière est beaucoup plus similaire à la forêt secondaire qu'à la jachère (0,82 pour Sorensen et 0,70 pour Jaccard) comme ce fût le cas pour la composition en espèces végétales. La jachère et la forêt secondaire présentent une similarité de 0,73 soit 73% pour Sørensen et de 0,58 soit 58% pour Jaccard. IV.7. Comparaison des effectifs des Rongeurs de la zone d'étude.Le résultat de l'analyse de variance effectuée est donné dans le graphique ci-après :
Effeclifs 4,5 4,0 2,5 2,0 3,5 3,0 0,5 1,5 1,0 Habitats; Moy. Pondérées Jachère Lisi ère Forêt Habitats Fig 4.6 : Comparaison des effectifs des Rongeurs dans les trois habitats. On note de ce graphique qu'il y a une différence significative entre les effectifs capturés dans les trois habitats (p = 0,0000). Ce résultat concorde avec ceux du test de Kolmogorov Smirnov (annexe 2.b.1) comparant les habitats deux à deux. Ce dernier test montre un réel effet de lisière traduit par des différences significatives entre la lisière et les intérieurs (p < 0,05). Par contre, aucune différence significative n'a été détectée en comparant les effectifs de la jachère avec ceux de la forêt secondaire (p > 0,10). IV.8. Réponses des espèces à l'hétérogénéité d'habitats.IV.8.1. Hybomys lunarisHybomys univittatus 5 b
Jachère Lisière Forêt secondaire Grille par habitat . Fig 4.7 : Comparaison des moyennes des Hybomys lunaris capturés dans les trois habitats. Ce graphique montre que Hybomys lunaris manifeste une adaptation du type V préférant la forêt secondaire par rapport aux autres habitats (annexe 2.b.2). IV.8.2. Hylomyscus sppHylomyscus spp
Moyennes par habitat 4 6 5 3 2 0 1 a b a Jachère Lisière Forêt secondaire Habitats Fig 4.8 : Comparaison des moyennes des Hylomyscus spp capturés dans les trois habitats. L'espèce Hylomyscus spp présente à la lumière de ce graphique une adaptation du type II ; l'espèce préfère la zone de lisière (annexe 2.b.2). IV.8.3. Lophuromys duduiLophuromys dudui
a ab b 0,5 0 2,5 2 1,5 1 Moyennes 3,5 3 Jachère Lisière Forêt secondaire Habitats . Fig 4.9 : Comparaison des moyennes des Lophuromys dudui capturés dans les trois habitats. Le graphique sous étude fait remarquer que l'espèce Lophuromys dudui présente une adaptation du type IV du tableau 3.2. Il ya donc effet de lisière et l'abondance de l'espèce est intermédiaire de celles des habitats adjacents (annexe 2.b.2). IV.8.4. Praomys sppPraomys sp
Jachère Lisière Forêt secondaire Habitats Fig 4.10 : Comparaison des moyennes des Praomys spp capturés dans les trois habitats. L'adaptation de Praomys spp est du type II. Il y a effet de lisière ; l'espèce évite la zone de lisière (annexe 2.b.2). IV.8.5. Stochomys longicaudatusStochomys longicaudatus
b b a 0,5 0 2,5 2 1,5 1 Moyennes 3,5 3 Jachère Lisière Forêt Habitats Fig 4.11 : Comparaison des moyennes des Stochomys longicaudatus capturés dans les trois habitats. Stochomys longicaudatus présente une adaptation du type V. Il n y a pas effet de lisière ; l'espèce évite la jachère (annexe 2.b.2). IV.9. Corrélation Espèces - Habitats.L'analyse en composantes principales corrélant les espèces aux habitats, donne les résultats illustrés par la figure 4.12 ci-dessous. PCA case scores
Praomys spp Nannomys munitoides 0.7 0.6 Forêt secondaire 0.3 Hybomys lunaris tomys natalensis Oenomys hypoxanthus -0.4 -0.3 0.1 Stochomys Malacomys longipe -0.1 01 Graphiurus lorraineus 0.3 0.4 0.6 0.7 Hylomyscus spp Mas Lophuromys dudui -0.1 Lisière -0.3 Lemniscomys striatus -0.4 -0.6 -0.7 Axis 2 Jachère -0.7 Axis 1 Vector scaling: 0,96 Fig 4.12 : Corrélation des espèces des Rongeurs avec différents habitats. Les espèces Lophuromys dudui et Mastomys natalensis sont corrélées à la jachère, l'espèce Hylomyscus spp préfère la lisière et les espèces Deomys ferrugineus, Hybomys lunaris sont corrélées à la forêt secondaire. Les espèces Stochomys longicaudatus et Malacomys longipes évitent la jachère tandis que l'espèce Praomys spp évite la lisière. Partie V : DISCUSSIONV.1. Aspects méthodologiquesV.1.1. Usage des grilles de piégeagesCe travail s'était réalisé sur un dispositif de 15 layons intérieures ou transects secondaires de 200m chacun et équidistant de 10m tracés dans trois grilles de piégeages à raison de 5 transects dans chaque grille de 1 ha installés dans chaque habitat. Un transect principal de 500m traversant les habitats était également tracé pour installer ces grilles de piégeages. Plusieurs auteurs ; pour des études des petits mammifères et de caractérisation des facteurs abiotiques du milieu, ont utilisé des transects principaux. A titre d'exemple, Malcolm et Ray (2000) en ont tracé 24 de 280m ; Alongo (2007) a tracé 2 de 200m et Iyongo (2008) 16 de 350m. Anderson et al (2003) ont observé la même équidistance que celle retenus pour ce travail alors que Boowers et Dooley (1993) ont adopté 12,5m entre les transects. White (1994) a utilisé la méthode des transects pour la capture des rongeurs à la Lopée au Gabon de même que Dudu (1991) dans la Réserve Forestière de Masako, Boowers & Dooley (1993) ; Boowers et Dooley (1996) aux Etats Unis ; Dunstan et Barry (1996) en Australie, Manson et Stiles (1998) aux Etats Unis ; Malcolm & Ray (2000) en République Centrafricaine ; Manson et al (2001); Anderson et al (2003); Wolf & Batzil (2004) aux Etats Unis ; Iyongo (2008) à Masako. D'autres auteurs tels que Chiarello (1999) et Barbosa & Marquet (2002) ont utilisé les transects pour l'étude des grands mammifères au Brésil et les coléoptères en Chili. Dudu (1991) ; Boowers et Dooley (1996) ; White et Edwards (2000) considèrent la méthode des transects comme celle de recensement la plus efficace pour les grandes zones de forêt pluviale africaine ; Wolf & Batzil (2004) affirment que c'est une méthode simple et pratique pour la recherche des espèces dans un biotope et permet une bonne estimation de leur abondance relative. Dunstan et Barry (1996) ; Chiarello (1999) ; Manson et al (2001); Anderson et al (2003) abordent dans le même sens mais précisent que cette méthode permet une récolte aisée aussi bien des données que des informations visuelles concernant la topographie et la structure de la végétation. Cependant, elle présente l'inconvénient de ne pas utiliser toutes les observations du milieu en ne se limitant qu'à celles le long des transects. V.1.2. Détection de la zone de lisièreTurner et al (1993) ; Walker et al (2003) se sont basés sur la végétation pour déterminer les écotones. La même méthodologie fut adoptée par Manson et al (2001); Anderson et al (2003) pour déterminer la zone de lisière entre la forêt et les vieux champs d'une part et entre la lisière et l'intérieur de la forêt d'autre part afin d'étudier la densité et la distribution des Rongeurs. Ainsi, la zone de lisière de 35m à 42m de long faisant objet de cette recherche a été délimitée à partir de l'étude de la végétation en allant de la jachère à la forêt secondaire. D'autres auteurs comme Alongo (2007) et Lokonda (2007) se sont servis des paramètres pédologiques pour déterminer la zone de lisière. V.1.3. Paramètres environnementauxPlusieurs auteurs antérieurs ont étudié l'évolution des facteurs environnementaux pour détecter les zones de lisière. On peut citer Ranney (1978) qui s'est intéressé à la température, à l'humidité, au vent et au degré de lumière au niveau du sol ; Alongo (2007) qui a observé l'humidité équivalente et la température du sol et Lokonda (2007) qui s'est intéressé à l'évolution du pH du sol. V.1.4. Capture des RongeursComme Dudu (1991) deux marques des pièges appâtés aux noix de palme ont été utilisées pour cette investigation. Les clapettes tuant les bêtes et les Sherman capturant vifs les animaux. Certains chercheurs ont utilisé exclusivement les pièges capturant vif les bêtes tels que Dunstan & Fox (1996) avec << Elliot live-traps >> ; Manson & Stiles (1998) ; Malcolm & Ray (2000) et Anderson et al (2003) avec << Sherman live-traps >> ; Mukinzi (2007) avec << Pit - falls >>... V.1.5. Indices ou coefficients de SimilaritéLes deux indices de similarité (Sørenson et Jaccard) montrent tous que la lisière est beaucoup plus similaire à la forêt secondaire qu'à la jachère en termes de la composition en espèces des Rongeurs. La même tendance a été observée par Iyongo (2008). L'indice de Sørenson a donné des valeurs supérieures à celui de Jaccard compte tenu du fait qu'il donne plus d'importance à la présence que Jaccard. Ces deux indices ont été utilisés par Nshimba (2008) dans son étude floristique, écologique et phytosociologique de l'île Mbiye. V.1.6. Analyses statistiquesLa corrélation linéaire a été calculée pour voir les affinités entre habitats sur base des variables environnementales (annxe1). L'analyse de la variance (ANOVA) à une voie a permis de comparer en une fois les effectifs de captures de trois habitats. Le test de Kolmogorov Smirnorv a permis la comparaison des effectifs des espèces des habitats pris deux à deux afin de vérifier l'ANOVA (annexe 2.b.1) et le test t a été calculé pour comparer les effectifs des espèces dans les habitats pris deux à deux afin d'apprécier la réponse des espèces à l'hétérogénéité d'habitats (annexe 2.b.2). Boowers & Dooley (1993) ont éffectué l'ANOVA à deux voie ; Boowers et Dooley (1996) ont comparés les densités de rongeurs entre taille de taches en utilisant l'analyse des mesures répétées de variance (RMA) ; Anderson et al (2003) ont testé les différences de densité de Peromyscus lecopus entre les lisières et intérieurs de petites et larges taches forestières avec l'ANOVA à deux voie ; Iyongo (2008) a utilisé les tests de Kruskall Wallis qui est une analyse de la variance non paramétrique et le U de Mann Whitney, l'équivalent non paramétrique du test t à cause de la distribution non normale de ses données. V.2. Résultats obtenusV.2.1. Paramètres environnementauxL'analyse des paramètres environnementaux ayant répondu à l'hétérogénéité d'habitats renseigne d'une part que (selon la température et l'humidité relative de l'air), la lisière présente des conditions similaires à l'un des deux habitats intérieurs qu'elle sépare et d'autre part que (selon la vitesse moyenne du vent et le point de rosée), la lisière présente ses conditions propres avec des différences significatives entre les trois habitats. Ces derniers résultats corroborent ceux de Schultz et al (1955) cité par Alongo (2007) qui attestent que de façon similaire, aux frontières entre prairie et forêt, il existe des changements nets de température, d'humidité et d'intensité lumineuse. Ranney (1978) a montré que près de lisière le sol de la forêt reçoit plus de lumière mais aussi expérimente des vents violents et des grandes variations de température. Alongo (2007) a observé de son coté que l'humidité équivalente et la température du sol entre la lisière et les zones de l'intérieur sont différentes et évoluent selon les habitats. V.2.2. Végétation de la zone d'étudeDudu & Gevaerts (1987) et Dudu (1991) signalent que la structure de la végétation influence fortement le choix des micromammifères, de leur habitat et de celui de leur nid ou gîte. Ce dernier affirme que la végétation joue un rôle important dans les communautés d'Insectivores et des Rongeurs. Anderson et al (2003) ont décrit la végétation de leur zone d'étude pour apprécier le lien entre la densité et la distribution de la souris à pattes blanches Peromyscus leucopus. Malcolm & Ray ont également décrit la végétation de la forêt à Gilbertiodendron dewevrei ainsi que les espèces accompagnatrices dans leur étude sur les Rongeurs en République Centre Africaine. Cette étude faisant suite à celles-ci haut citées, s'est proposée de caractériser les différents habitats de la zone d'étude par l'identification systématique des espèces. Cette étude a donné des résultats qui corroborent ceux de Makana (1986); Kahindo (1988); Mambangula (1988) ; Dudu (1991) et Mabay (1994) et Iyongo (2008) montrant que la jachère est constituée de deux strates (arbustive et herbacée) principalement et la forêt secondaire de trois (herbacée, arbustive et arborescente) comme l'indique la partie résultat. De même que Odum (1971), Forman et Godron (1995), Bogaert et al (2001) et Iyongo (2008) on peut noter que la lisière s'est montrée la plus riche en espèces par rapport aux autres habitats (jachère et forêt secondaire). V.2.3. Impacts des lisières sur les RongeursV.2.3.1. Présence des espèces dans différents habitats - TypologieL'analyse des données recueillies montre que la
jachère connait une richesse spécifique de dix secondaire avec au total 148 rongeurs, suivi de la jachère avec 116 rongeurs et enfin de la lisière avec 84 rongeurs. Ces résultats confirment l'hypothèse 2 de cette étude. La jachère et la lisière comptent en commun sept espèces. Trois des espèces présentes dans la jachère (Nannomys minutoides, Oenomys hypoxanthus et Mastomys natalensis) n'ont pas été signalées en lisière et une espèce en lisière (Malacomys longipes) n'a pas marqué sa présence en jachère. L'absence de trois espèces de la jachère en lisière et de celle de la lisière en forêt secondaire prouvent l'existence de la zone de lisière et donc des effets de lisière dans la zone d'étude. Ces résultats qui suivent la même tendance que ceux de Iyongo (2008) confirment l'hypothèse 3 de cette recherche. La jachère et la forêt secondaire ont également eu sept espèces en commun. Trois espèces de la jachère (Lemniscomys striatus, Oenomys hypoxanthus et Mastomys natalensis) ont été absentes de la forêt secondaire comme aussi deux espèces de la forêt secondaire (Nannomys minutoides et Graphiurus lorraineus) le sont de la jachère. La lisière et la forêt secondaire ont à leur tour une richesse commune de sept espèces, dont une de la lisière (Lemniscomys striatus) est absente de la forêt secondaire et deux espèces de la forêt secondaire (Nannomys minutoides et Graphiurus lorraineus) sont absentes de la lisière. C'est une deuxième confirmation de l'hypothèse 3 de l'étude. En sommes, toutes les espèces présentes dans la lisière ont aussi marqué leur présence dans les zones de l'intérieur (rejet de l'hypothèse 4). Cela pourrait s'expliquer dans un premier temps par le fait que ces espèces seraient mieux intégrées dans la jachère et la forêt secondaire de Masako (Dudu.1991) et seraient mobiles entre ces deux principaux habitats tout en exploitant la lisière qui présente les conditions de ces deux habitats réunis (Baudry & Merriam, 1988 dans Burel & Baudry, 2003 ; Kolasa & Zalewski, 1995 ; Iyongo, 2008). Une deuxième explication pourrait être la confirmation de l'hypothèse 1 de ce travail compte tenu du fait que l'examen avec minutie des différents paramètres environnementaux observés montre que la lisière présente des conditions environnementales instables par rapport aux habitats de l'intérieur. Par ailleurs, notons que l'espèce Graphiurus lorraineus considéré autre fois par Iyongo (2008) comme espèce spécifique à la lisière s'est plutôt révélé attachée à la forêt secondaire et l'espèce Oenomys hypoxanthus considéré par le même auteur comme généraliste s'est montrée spécifique à la jachère. Ces observations nous permettent de concéder avec Nicolas et al cités par Katuala (2009) qu'il se passe des changements dans la structure des habitats conduisant à des modifications spécifiques des Rongeurs. V.2.3.2. Abondance relative des espèces dans les habitatsLa répartition des espèces entre habitat a révélé que les abondances élevées ont été enregistrées en forêt secondaire et en jachère (les deux principaux types d'occupation du sol) et que de faibles valeurs ont généralement été observées dans la lisière. Ces résultats concordent avec ceux de Dudu (1991) qui signale que les jachères et les forêts secondaires de Masako sont les milieux les plus appropriés pour ces mammifères ayant des richesses et des abondances quasi égales, ne se différenciant uniquement que par l'inégale et l'irrégulière distribution des espèces rares ou celles inféodées à l'un des deux biotopes. Il précise que les abondances élevées seraient liées probablement aux conditions écologiques favorables de ces habitats, au faible taux de prédation et à une bonne intégration des espèces qui y ont installé leurs nids et s'y reproduisent. Dangerfield et al (2003) sur les invertébrés en Australie ; Iyongo (2008) sur les rongeurs à Masako, ont obtenu les mêmes résultats. Les faibles abondances en lisière seraient par contre liées à plusieurs facteurs parmi lesquels nous retenons que : Les lisières sont considérées comme des zones de tension où les principales espèces des communautés adjacentes atteignent leurs limites (Clements, 1905). Les lisières sont des << puits écologiques » dans le cadre du système dit << source - puits » où les espèces des milieux adjacents y disparaissent en plus grand nombre car y subissant une prédation accrue (Vanpeenne, 1998). Ce taux élevé de prédation serait dû aux faibles superficies qui caractérisent les lisières comparativement aux habitats qu'elles séparent et qui facilitent le repérage des espèces proies par leurs prédateurs (Henshaw, 2006). V.2.3.3. Sensibilité des espèces à l'hétérogénéité d'habitats - TypologieLe test de Kolmogorov Smirnov comparant en une fois les
effectifs des Rongeurs de trois l'intérieur. Les tests t testant la réponse des espèces à l'hétérogénéité des habitats par comparaison des habitats deux à deux ont montré que 5 espèces sont sensibles à l'hétérogénéité des habitats. On note que l'espèce Hybomys lunaris est mieux intégrée en forêt secondaire et présente une adaptation du type V ; Hylomyscus spp présente une adaptation du type II et préfère la lisière ; Lophuromys dudui présente une adaptation du type IV ; il ya effet de lisière car son abondance en lisière est intermédiaire aux deux autres habitats. ; Praomys spp présente une adaptation du type II, il évite la lisière et préfère les deux habitats de l'intérieur et enfin Stochomys longicaudatus qui évite la jachère et présente une adaptation du type V, il n'est pas sensible à l'effet de lisière. La réponse des espèces Hybomys lunaris et Praomys spp ont montré des préférences qui concordent avec les résultats de Dudu (1991) qui affirme qu'à Masako, les jachères et les forêts secondaires se caractérisent par la dominance partout et à chaque année de Praomys jacksoni, l'existence de deux espèces très abondantes Hybomys lunaris et Deomys ferrugineus L'espèce Lophuromys dudui présente une adaptation du type IV qui confirme les résultats de Iyongo (2008). Elle trouve dans la lisière les conditions réunies de deux habitats adjacents ; elle exploite aussi bien la lisière que les zones de l'intérieur. Hylomyscus spp dans sa sensibilité à l'hétérogénéité d'habitats a renvoyé à une adaptation du type II comme ce fût le cas pour l'espèce Hylomyscus stella dans la recherche de Iyongo en 2008. Cependant Hylomyscus spp a préféré la lisière alors que Hylomyscus stella a évité la lisière préférant la jachère. Ces différences pourraient s'expliquer par le fait qu'Iyongo a travaillé au niveau de l'espèce et non du genre comme c'est le cas ici et que les périodes de récoltes diffèrent également. Enfin, Stochomys longicaudatus présente une adaptation du type V, elle évite la jachère mais préfère la forêt secondaire tout en exploitant correctement la lisière. V.2.4. Corrélation espèces - habitatsL'analyse en composantes principales révèle que les espèces Lophuromys dudui et Mastomys natalensis sont corrélées à la jachère ; l'espèce Hylomyscus spp à la lisière et les espèces Deomys ferrugineus, Hybomys lunaris à la forêt secondaire. Stochomys longicaudatus et Malacomys longipes évitent la jachère alors que Praomys spp évite la lisière. CONCLUSION ET PERSPECTIVESLe présent travail avait pour objectif principal d'analyser la composition et la distribution des populations des Rongeurs en relation avec l'hétérogénéité des habitats dans un écosystème anthropisé de la Réserve Forestière de Masako. Il cadre avec les relations « faune-habitats » en milieu forestier tropical et visait ; en s'appuyant sur les facteurs environnementaux, à évaluer le degré de sensibilité des Rongeurs forestiers face aux effets de lisière. Les données ont été récoltées à Masako de juin à août 2010 puis complétées par celles d'avril et mai disponibilisées par le doctorant Leon Iyongo encadreur de ce mémoire. La méthode des grilles de piégeages placées à gauche d'un transect de 500m et traversant chaque type d'occupation du sol (jachère, Lisière et forêt secondaire) a été utilisée. Chaque grille contenait 100 pièges appâtés aux noix de palme dont 50 Clapettes et 50 Sherman placés sur cinq layons de 200m de long tracés à 10m d'équidistance. Les paramètres environnementaux étaient prélevés à l'aide de l'anémomètre de marque américaine Kestrel 3000. Au total 10 jours de captures et de prises des variables environnementales étaient organisés par campagne (mois). La végétation a également été décrite par identification des espèces dans les placeaux installés dans chaque habitat et l'identification des espèces de Rongeurs a été faite sur base des caractères morphologiques externes. La corrélation linéaire, l'analyse de la variance à une voie, le test de Kolmogorov Smirnov et le test t ont été appliqués selon les résultats recherchés. Cette étude a conduit à la récolte de 348 Rongeurs répartis en 3 familles et 12 espèces différentes avec une abondance différente en fonction d'habitat certainement due aux conditions environnementales et aux préférences des espèces. La sensibilité des espèces aux effets de lisière a été prouvée, laissant voir des adaptations différentes selon les espèces. La corrélation des espèces des Rongeurs à chaque type d'habitat a été réalisée grâce à l'analyse en composantes principales. Les hypothèses 1, 2 et 3 ont été affirmées et l'hypothèse 4 infirmée. La présence de l'espèce Mastomys natalensis en jachère est une preuve de l'anthropisation actuelle de la Réserve Forestière de Masako. La période de récolte étant courte et les effectifs des individus capturés petits, nous suggérons que les mêmes investigations soient réalisées sur un temps long (une année par exemple) afin de tirer au mieux les conclusions générales de l'impact réel des paramètres environnementaux sur la distribution des Rongeurs et du degré de sensibilité des ceux-ci aux effets de lisières. REFERENCES BIBLIOGRAPHIQUESAlongo, L., 2007 - Etude de l'effet de lisières sur l'humidité équivalente et la température du sol d'un écosystème forestier de la cuvette centrale congolaise. Cas de la Réserve forestière « jardin systématique »de l'INERA à Yangambi. Dissertation, DEA, Faculté des Sciences, Unikis:64p. Anderson, C.S., Cady, A.B.& Meikle, D., 2003. - Effects of vegetation structure and edge habitat on the density and distribution of white-footed mice (Peromyscus leucopus) in small and large forest patches. Canadian Journal of Zoology, 81: 897-904. Barbosa, O. & Marquet, P.A., 2002. - Effects of forest fragmentation on the beetle assemblage at the relict forest of Fray Jorge, Chile. Oecologia, 123: 296-306. Bastian, O., 2001 - Landscape ecology: towards a unified discipline? Landscape Ecol.16:757- 766. Baudry, J., 1988: Structure et fonctionnement écologique des paysages: Cas des bocages. Bulletin d'Ecologie, 19 [523-530] Baudière, A. & Gauquelin, T., 1990. - Successions, substitutions, écotones et systèmes phytomorphogénétiques pyrénées d'altitude. Botanica pirenaico-cantabrica Jaca y Huesca : 353-366. Begon, M ; Harper, J.L ; Townsend, CR., 1990 - «Ecology: Individuals, Populations and Communities» Blackwell Scientific Publications. 945p. Bennet, AF., 1990 - Habitats corridors: their role in wildlife management and conservation. Arthur Rylah Inst. Environ. Res, Depart. Conservation & Environment, Melbourne, 36p. Bertrand, G., 1975 - Pour une histoire écologique de la France rurale. In : Histoire de la France Rurale. Le seuil. Paris, 1 : 34-113. Bogaert, J., 2000. - Quantifying habitat fragmentation as a spatial process in a patch-corridormatrix landscape model. Thèse de doctorat, Université d'Anvers, 209 p. Bogaert, J., Van Eysenrode, D.S., Van Hecke, P., 2001. - The Interior-to-Edge Breakpoint Distance as a Guideline for Nature Conservation Policy. Environnemental Management, 27: 493-500. Bogaert, J., Ceulemans, R. & Van Eysenrode, S.D., 2004. - A decision tree algorithm for detection of spatial processes in landscape transformation. Environnemental Management, 33: 62-73. Bogaert, J. & Mahamane, A., 2005. - Ecologie du paysage: cibler la configuration et l'échelle spatiale. Annales des Sciences Agronomiques du Bénin 7 : 39 - 68 Bouchon, J., 1979 - Structure des peuplements forestiers. Annales des Sciences Forestières 36(3) : 175-209. Bowers, M.A. & Dooley, J.L., 1993. - Predation hazard and seed removal by small mammals: microhabitat versus patch scale effects. Oecologia 94: 247-254. Bowers, M.A. & Dooley, J.L., 1996. - Influences of patch size and microhabitat on the demography of two old-field rodents. Oikos 75: 453-462. Burel, F. & Baudry, J., 2000. - Ecologie du paysage. Concepts, méthodes et applications. Tec & Doc : Paris, France, 359 p. Chiarello, A.G., 1999. - Effects of fragmentation of the atlantic forest on mammal communities in south-eastern Brazil. Biological Conservation 89: 71-82 Cirad., 2001 - Approches écologiques. Ecologie et dynamique des resources naturelles. Centre de coopération international en recherché agronomique pour le développement. Site : cormas.cirad.fr/fr/demarch/ecologi.htm. Cifor et Cirad., 2007 - La forêt en République Démocratique du Congo post - conflit. Analyse d'un agenda prioritaire. www.cifor.cgiar.org/publication, 82p Clements, F.E., 1905. - Research methods in ecology. University publishing Company, Lincoln: 33-76 Dangerfield, J.M., Pik, A.J., Britton, D., Holmes, A., Gillings, M., Olivier, I., Briscoe, D. & Beattie, A.J., 2003. - Patterns of invertebrate biodiversity across a natural edge. Austral Ecology 28: 227-236. Decamps, H., 2004 - L'écologie du paysage ou l'ambition paysagère de l'écologie. Lettre de l'Institut Français de la Biodiversité, n°5, 2pp. De Menocal., PB 2004 - African climate change and farmal evolution during the Pliocene - Pliocene. Earth and planetary Sciences Letters 220: 3-24. Di Castri, F., 1981. - L'écologie : naissance d'une science de l'homme et de la nature. Le courrier de l'UNESCO, 34 : 6-11. Djibu, K., 2007. - Evaluation de l'état de la fragmentation des forêts au Katanga. Mémoire DEA, ULB, 85 p. Dudu, A.M. & Gevaerts, H., 1987. - Examen de la distribution des Muridés (Rodentia, Mammalia) dans quelques associations végétales des environs de Kisangani (Zaïre). Annales de la Faculté des Sciences, Kisangani, 4 : 103-110. Dudu, A.M., 1991. - Etude du peuplement d'insectivores et de rongeurs de la forêt ombrophile de basse altitude du Zaïre (Kisangani, Masako). Thèse de doctorat, Antwerpen, 171 p. Duveiller, G ; Defourny, P ; Desclee, B et mayaux, P ; 2008 - Déforestation in Central Africa : Estimates at regional, national and landscape levels by advanced processing of systematically distributed Landsat extracts. Remote Sensing of Environnment, 112 (5), pp 1969 - 1981. FAO, 2005. - Global forest resources assessment of 2005. Progress towards sustainable forestmanagement. FAO Forestry paper 147. FAO, Rome Fays., R.; 2009 - Des forêts... Des bois...Ambahibe, madagascar, Impribeau - sainte - ode - Belgique. Forman, R.T.T. & Godron, M., 1981. - Patches and structural components for a landscape ecology. Bioscience, 31: 733 - 740. Forman, R.T.T. & Godron, M., 1986. - Landscape ecology. John Wiley & sons, New York, USA, 640 p. Forman, R.T.T., 1995. - Land mosaics. The ecology of landscapes and regions. Cambridge University Press, Cambridge: 415-430. Fox, B.J. & Fox, M.D., 1981. A comparison of vegetation classifications as descriptors of small mammal habitat prefenrece. Vegetation classification in Australia. Australian University Press, Australia: 66-80. Gauthier-Hion, AColyn, M & Gauthier, JP., 1999 - Histoire naturelle des primates d'Afrique Centrale. Libreville : ECOFAC, 170p. Gecopa., 2004 - Site du GECOPA- Groupe de gestion écosystémiques et écologie du paysage. Site : http:// gecos.epfl.ch/gecopa/defpaysage.shtml. Gitay, H ; Suarez, A ; Watson, KT & Dokker, DJ.(Eds)., 2002 - Climate and Biodiversity. IPCC Technical Paper V.pp.86 Goreaud, F., 2000 - Analyse de la structure spatiale en forêt tempérée à l'étude et à la modélisation des peuplements complexes. Thèse, ENGREF. 524p. Happold, DCD., 1996 - Mammals of the Guinea - Congo rain forest. Proceeding of the Royal Society of Edinburgh. 104B: 243-284. Henshaw, B., 2006. - Oiseaux forestiers dans les zones urbaines : besoins des espèces sensibles à la superficie de l'habitat. ( http://www.on.ec.gc.ca/wildlife/factsheets/fs_forestbirds-f.html) Iorgulescu, I. & Schaepfer, R., 2002. - Paysage en tant qu'écocomplexe : définition, types, caractéristiques, fonctionnement et fonctions. Fiche d'enseignement 4.1., Laboratoire de Gestion des Ecosystèmes, Ecole Polytechnique de Lausanne, Suisse, 24 p. Iyongo, W.M., 1996. - Etude de la structure des populations de quelques Muridae et Cricetidae (Rodentia, Mammalia) : essai d'utilisation du poids de cristallin chez les espèces forestières. Mémoire, Faculté des Sciences, Unikis, 31p. Iyongo, W.M., 2008. - Etude des effets de lisière sur les populations de rongeurs dans la Réserve Forestière de Masako (Kisangani, R.D.Congo). DEA, Faculté des Sciences, ULB, 112p Iyongo W.M, Visser M, Verheyen E, Leirs H, Dudu A, Ulyel A.5 et Bogaert J., 2009 - Etude préliminaire des effets de lisière sur quatre espèces de rongeurs dans la Reserve Forestière de Masako (Kisangani, R.D. Congo), Annales I.S.E.A Vol (4) : 156-167. Juakaly, M., 2002. - Macrofaune et Mésofaune du sol dans un système de cultures sur brûlis en zone équatoriale (Masako, Kisangani, R.D. Congo). Dissertation DES, Faculté des Sciences, Unikis, 85 p. Juakaly, M., 2007 - Resilience et écologie des araignés du sol d'une forêt équatoriale de basse altitude. Reserve Forestière de Masako, Kisangani, R.D.Congo. Vol.1 :texte. Thèse de doctorat inédite, Faculté des Sciences, Unikis, p5-30. Juo, ASR & Wilding., 1996 - soils of the lowland forests of West and Central Africa. Proc. R. Soc. Edinburgh. 104B, p15-29. Kahindo, M., 1988. - Contribution à l'étude floristique et phytosociologique de la Réserve Forestière de Masako (Haut-Zaïre). Mémoire, Faculté des Sciences, Unikis: p5-30 Katuala, G.B., Hart, J.A., Hutterer, R., Leirs, H. & Dudu, A., 2005. - Biodiversity and ecology of small mammals (Rodents and Shrews) of the «Réserve de Faune à Okapi», Democratic Republic of the Congo. Belgian Journal of Zoology, 135: 191-196. Katuala, G.B., 2009. - Biodiversité et biogeography des rongeurs Myomorphes et Sciuromorphes (Rodentia: Mammalia) de quelques blocs forestiers de la region de Kisangani (R D Congo). Thèse de doctorat inédite, Faculté des Sciences, Unikis, Kolasa, J. & Zalewski, M., 1995. - Notes on ecotone attributes and functions. Hydrobiologia 303: 1-7. Legendre, L. & Legendre, L., 1998. - Numerical Ecology. Developments in Environmental Modelling. Elsevier Science B.V., Amsterdam, 853 p. Leroux, M., 2004 - Le réchauffement climatique est un mythe ! Agriculture et Environnement : 188, 6p Lokonda, O.W., 2007. - Etude de l'effet de lisière sur la réaction ou pH du sol dans un paysage fragmenté de la cuvette centrale congolaise : cas de la Réserve Forestière de Masako (Province Orientale, R.D. Congo). Mémoire DEA, Faculté des Sciences, Unikis, 80 p. Mabay, K., 1994. - Contribution à l'étude structurale, des forêts primaire et secondaire de la Réserve de Masako. Mémoire, Faculté des Sciences, Unikis, 73 p. Makana, M., 1986. Contribution à l'étude floristique et écologique de la forêt à Gilbertiodendron dewevrei (De Wild) J. Léonard de Masako (Kisangani). Mémoire, Faculté des Sciences, Kisangani, 62 p. Magurran, AE., 2004 - Measuring biological diversity. Blackwelle Publishing company, United Kingdom, 256p. Malcolm, J.R. & Ray, J.C., 2000. - Influence of timber extraction routes on central african small-mammal communities, forest structure and tree diversity. Conservation Biology 14 (6): 1623-1638. Mambangula, LN., 1988 - Etude floristique et biologique des lianes et herbes grimpantes des forêts secondaires et primaire de Masako (Haut Zaire). Mémoire inédit, Faculté des Sciences, Kisangani, 62 p. Manson, R.H. & Stiles, E.W., 1998. - Links between microhabitat preferences and seed predation by small mammals in old fields. Oikos 82(1): 37-50. Manson, R.H., Ostfeld, R.S. & Canham, C.D., 2001. - Long-term effects of rodent herbivores on tree invasion dynamics along forest-field edges. Ecology 82 (12): 3320-3329 Mate, M., 2001 - Croissance, phytomasse et minéralomasse des haies des légumineuses améliorantes en cultures en allées à Kisangani (R D Congo). Thèse de doctorat inédite, Faculté des Sciences, , ULB, 235 p. Motulsky, H.J., 1995. - Biostatistique: une approche intuitive. Oxford University press, Inc 484 p. Mukinzi, I., Katuala, G.B., Kennis, J., Gambalemoke, M., Kadange, N., Dudu, A.M., Colyn, M. & Hutterer, R., 2005. - Preliminary data on the biodiversity of rodents and insectivores (Mammalia) in the periphery of Kisangani (D.R. Congo). Belgian Journal of Zoology,135: 133-140. Mukinzi, I., 2007 - Contribution et structure du peuplement des soricidae (Soricomorpha, Mammalia) de la Réserve Forestière de la Yoko et de ses environs. (Kisangani, R.D.Congo). Dissertation, DES, Faculté des Sciences, Unikis Ngo Bieng, MA., 2004 - Analyse de la structure spatiale des peuplements forestiers mélangés Chêne sessile (Querus petraea) - Pin sylvestre (Pinus sylvestris) de la région Centre. DEA d'Ecologie, Université de Paris XI, Ecole Normale Supérieure I.N.A.P.G 30p Nshimba, S.W.M., 2008. - Etude floristique, écologique et phytosociologique des forêts de l'île Mbiye à Kisangani, R.D. Congo. Thèse de doctorat, ULB, 271 p. Nyakabwa, M., 1982. - Phytocénose de l'écosystème urbain de Kisangani. Thèse de doctorat, Faculté des Sciences, Kisangani, 998 p. Odum, E.P., 1971. - Fundamentals of ecology. W.B. Saunders Co. Philadelphia, 16: 262-270 Paillat, G et Butet, A., 1994 - Fragmentation et connectivité dans les paysages : Importance des habitats corridors pour les petits mammifères.URA 1853 « Ecobio », Laboratoire d'Evolution des Systèmes Naturels et Modifiés. Querouil, S., 2001 - Intérêts et limites de l'approche moléculaire pour aborder la biographie et la speciation: l'exemple de quelques mammifères d'Afrique tropicale. Thèse de doctorat, Université de Rennes 1310p Ranney, J., 1978. The importance of edge in the structure and dynamic of forest islands. Pages 67-95 in R.L. Burguess and M.D Sharpe editors. Forest island dynamics in man dominated landscape. Springer-Verlag, New York, USA Ramade., F., 1984 - Eléments d'écologie: Ecologie fondamentale. Mc graw - Hill, Paris IX - 397 p. Risser, P., 1989. - Landscape pattern and its effect on energy and nutrient distribution. Springer Verlag. New York, 45-56 Soki, K., 1994 - Biologie et écologie des termites (Isoptera) des forêts ombrophiles du Nord - Est du Zaire (Kisangani). Thèse de doctorat inédite, ULB, 329p. Turner, G.T., Hansen, R.M., Reid, V.H., Teitjen, H.P. & Ward, A.L., 1993. Landcape ecology: the effect of pattern on process. Ann. Rev. Ecol. Syst. 20 : 171-197. Upoki, A., 1997. - Aperçu systématique et écologie des espèces aviennes de la Réserve Forestière de Masako (Kisangani, Haut-Zaïre). Dissertation DES, Faculté des Sciences, Unikis, 77 p. Upoki, A., 2001.- Etude du peuplement en Bulbuls (Pycnonotidae, Passeriformes) dans la réserve forestière de Masako à Kisangani (R.D. Congo). Thèse de doctorat, Faculté des Sciences, Unikis, 160 p. Vanpeene, B.S., 1998. - Transformations des paysages et dynamique de la biodiversité végétale. Les écotones, un concept clé pour l'étude des végétations post-culturales. L'exemple de la commune d'Aussois (Savoie). Thèse de doctorat, Sciences de l'Environnement, Cemagref, Grenoble. Walker, S., Barstow, W.J., Steel, J.B., Rapson, G.L., Smith, B., King, W.M. & Cottam, Y.H., 2003. - Properties of ecotones: evidence from five ecotones objectively determined from a coastal vegetation gradient. Journal of Vegetation Science 14: 579-590. White, L.J.T., 1994. - The effects of commercial mechanized selective logging on a transect in lowland rainforest in the Lope Reserve, Gabon. Journal of Tropical Ecology 10: 313-322. White, L.J.T et Edwards, A., 2000 - Conservation en forêt pluvial africaine: méthodes de recherche. The Wildlife Conservation Society, New York USA, nombreuses illustrations, 456p Wiens, J.A., Crawford, C.S., Gosz, J.R., 1985. - Boundary dynamics: a conceptual framework for studying landscape ecosystems. Oikos 45: 421-427. Wiens, J.A., 1999. - Landscape ecology: the science and the action. Landscape Ecol.14: 88 Wolf, M. & Batzli, G., 2004. - Forest edge, high or low quality habitat for white-footed mice (Peromyscus leucopus)? Oecology 85 (3): 756-769. TABLE DES MATIERESAPERÇU, CONTEXTE ET PROBLEMATIQUE 9 PARTIE I : INTRODUCTION GENERALE 12 1.1. ECOLOGIE DU PAYSAGE. 12 1.1.1. Définition et Importance 12 1.1.2. Historique de la discipline et son évolution 13 1.2. LE PAYSAGE 13 1.2.1. Origine, définitions, typologie et échelle 13 1.2.2. Processus écologique se déroulant au sein de paysage 14 1.3. PATTERN/PROCESS PARADIGM 15 1.3.1. Fonctionnement d'un écosystème 15 1.3.2. Processus et structure spatiale d'un paysage 15 1.4. CATEGORIES D'ELEMENTS DU PAYSAGE 16 1.4.1. Tache, corridor et matrice 16 1.4.2. Concepts d'écotone et types 17 1.5. LISIERE ET INTERIEUR 18 1.6. LES RONGEURS DANS LA RESERVE FORESTIERE DE MASAKO 18 1.6.1. Pourquoi étudier les Rongeurs ? 18 1.6.2. Richesse spécifique de Masako 19 Partie II : HYPOTHESES, INTERETS ET OBJECTIFS DE L'ETUDE 20
Partie III : MILIEU D'ETUDE, MATERIELS ET METHODES 22 III. MILIEU D'ETUDE 22 III.1.1. Choix et localisation du site 22 III.1.2. Hydrographie 23 III.1.3. Situation climatique 23 III.1.4. Végétation 25 III.1.5. Activités humaines 26 III.2. MATERIEL BIOLOGIQUE ET EQUIPEMENTS TECHNIQUES 26 III.2.1. Matériel biologique 26 III.2.2. Equipements techniques 26 III.3. METHODES 27 III.3.1. Descriptions du dispositif 27 III.3.2. Technique de piégeage 27 III.3.3. Prélèvement des paramètres environnementaux 28 III.3.4. Analyses statistiques. 28 III.3.5. Les coefficients ou indices de similarité 29 III.3.6. Typologie de réponse des espèces à l'hétérogénéité des habitats. 29 Partie IV : RESULTATS 30
IV.5.1. Fréquence relative des espèces par habitat 35 IV.5.2. Abondance relative des espèces dans les trois habitats. 36 IV.5.3. Abondance relative des espèces dans la zone d'étude. 36 IV. 5.4. Richesse spécifique des habitats. 37 IV.5.4.1. Forêt secondaire. 37 IV.5.4.2. Lisière. 37 IV.5.4.3. Jachère. 37
Partie V : DISCUSSION 43
V.2.1. Paramètres environnementaux 45 V.2.2. Végétation de la zone d'étude 46 V.2.3. Impacts des lisières sur les Rongeurs 46 V.2.3.1. Présence des espèces dans différents habitats - Typologie 46 V.2.3.2. Abondance relative des espèces dans les habitats 48 V.2.3.3. Sensibilité des espèces à l'hétérogénéité d'habitats - Typologie 48 V.2.4. Corrélation espèces - habitats 49 CONCLUSION ET PERSPECTIVES 50 REFERENCES BIBLIOGRAPHIQUES 51 TABLE DES MATIERES 57 ANNEXES
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Tests Univariés de Significativité pour Effectifs (Données ULB rongeurs Meniko) |
|||||
|
Paramétrisation sigma-restreinte |
|||||
|
Décomposition efficace de l'hypothèse |
|||||
|
SC |
Degr. de |
MC |
F |
p |
|
|
ord. origine |
772,320 |
1 |
772,3198 |
45,30829 |
0,000000 |
|
Habitats |
32,427 |
2 |
16,2135 |
0,95117 |
0,388411 |
|
Erreur |
2795,525 |
164 |
17,0459 |
||
2. b.1. Test de Kolmogorov Smirnov
Effet
Test de Kolmogorov-Smirnov (Jachère - Lisière)
|
Tests Univariés de Significativité pour Effectifs (Données rongeurs Meniko) Paramétrisation sigma-restreinte Décomposition efficace de l'hypothèse |
|||||||||
|
SC |
Degr. de |
MC |
F |
p |
|||||

ord. origine
Habitats
Erreur
772,320 1 772,3198 45,30829 0,000000
|
32,427 |
2 |
16,2135 |
0,95117 0,388411 |
|
2795,525 |
164 |
17,0459 |
Test de Kolmogorov-Smirnov (Lisière-Forêt)
|
variable |
Test de Kolmogorov-Smirnov (Données rongeurs Meniko) Par var. Habitats Tests significatifs marqués à p <,05000 |
||||||
|
Max Nég |
Max Pos |
niv. p |
Moyenne |
Moyenne |
Ec-Type Jachère |
Ec-Type Lisière |
|
|
Effectifs |
0,00 0,112795 p < .05 2,444444 1,145455 4,753020 1,799364 |
||||||
Test de Kolmogorov-Smirnov (Jachère - Forêt)
|
variable |
Test de Kolmogorov-Smirnov (Données rongeurs Meniko) Par var. Habitats Tests significatifs marqués à p <,05000 |
|||||
|
Max Nég Différnc |
Max Pos |
niv. p |
Moyenne Moyenne Jachère Forêt |
Ec-Type Jachère |
Ec-Type |
|
|
Effectifs |
-0,040868 0,038314 p > .10 2,444444 2,482759 4,753020 4,608312 |
|||||
Test de distribution
|
variable |
Test de Kolmogorov-Smirnov (Données rongeurs Meniko) Par var. Habitats Tests significatifs marqués à p <,05000 |
|||||
|
Max Nég Différnc |
Max Pos |
niv. p |
Moyenne Moyenne Jachère Forêt |
Ec-Type Jachère |
Ec-Type |
|
|
Effectifs |
-0,040868 0,038314 p > .10 2,444444 2,482759 4,753020 4,608312 |
|||||
2.b.2. Test t de Student Test sur Hybomys univittatus
|
Effet |
Tests Univariés de Significativité pour Effectifs Paramétrisation sigma-restreinte Décomposition efficace de l'hypothèse |
||||||||||||||||||
|
SC |
Degr. de |
MC |
F |
p |
|||||||||||||||
|
ord. origine |
69,06154 |
1 |
69,06154 |
15,37808 |
0,002387 |
||||||||||||||
|
Habitats |
13,45714 |
2 |
69,07778 |
15,39889 |
0,002658 |
||||||||||||||
|
Erreur |
49,40000 |
11 |
4,49091 |
||||||||||||||||
|
Test t pour des Echantillons Indépendants (Don |
|||||||||||||||||||
|
Note : Variables traitées comme des échantillon |
|||||||||||||||||||
|
Moyenne |
Moyenne valeur t dl |
p |
|||||||||||||||||
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
|||||||||||||||||
|
Jachère vs. Lisière |
1,200000 1,600000 -0,447214 8 0,666581 |
||||||||||||||||||
|
Test t pour des Echantillons Indépendants |
|||||||||||||||||||
|
Note : Variables traitées comme des échantillo |
|||||||||||||||||||
|
Moyenne |
Moyenne |
valeur t dl |
p |
||||||||||||||||
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
|||||||||||||||||
|
Jachère vs. Forêt |
1,200000 3,600000 -1,54919 8 0,031599 |
||||||||||||||||||
|
Test t pour des Echantillons Indépendants |
|||||||||||||||||||
|
Note : Variables traitées comme des échantillon |
|||||||||||||||||||
|
Moyenne |
Moyenne |
valeur t |
dl |
p |
N Ac |
||||||||||||||
|
Groupe1 vs. Group |
Groupe 1 |
Groupe 2 |
Group |
||||||||||||||||
|
Lisière vs. Forêt |
1,60000 3,60000 -1,4433 8 0,03777 |
||||||||||||||||||
Test sur Hylomyscus spp Test sur Lophuromys duduiANOVA
|
Effet |
Tests Univariés de Significativité pour Effectifs Paramétrisation sigma-restreinte Décomposition efficace de l'hypothèse |
||||
|
SC |
Degr. de |
MC |
F |
p |
|
|
ord. origine |
166,6667 |
1 |
166,6667 |
7,342144 |
0,018968 |
|
Habitats |
54,9333 |
2 |
27,4667 |
1,209985 |
0,332125 |
|
Erreur |
272,4000 |
12 |
22,7000 |
||
Test t jachère - Lisière (p = 0,515466 ; différence non significative)
|
Test t pour des Echantillons Indépendants |
||||
|
Note : Variables traitées comme des échantillo |
||||
|
Moyenne |
Moyenne |
valeur t dl |
p |
|
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
||
|
Jachère vs. Lisière |
2,666667 2,000000 0,674200 10 0,515466 |
|||
Test t jachère - Forêt ( p = 0,032504 ; différence significative
|
Test t pour des Echantillons Indépendants |
||||
|
Note : Variables traitées comme des échantillo |
||||
|
Moyenne |
Moyenne |
valeur t dl |
p |
|
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
||
|
Jachère vs. Forêt |
2,666667 1,333333 2,480695 10 0,032504 |
|||
Test t Lisière - Forêt (p = 0,500827 ; non significatif)
|
Test t pour des Echantillons Indépendants |
||||
|
Note : Variables traitées comme des échantillo |
||||
|
Moyenne |
Moyenne |
valeur t dl |
p |
|
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
||
|
Lisière vs. Forêt |
2,000000 1,333333 0,698430 10 0,500827 |
|||
Test sur Praomys spp ANOVA
|
Effet |
Tests Univariés de Significativité pour Effectifs Paramétrisation sigma-restreinte Décomposition efficace de l'hypothèse |
||||
|
SC |
Degr. de |
MC |
F |
p |
|
|
ord. origine |
1401,667 |
1 |
1401,667 |
35,15886 |
0,000069 |
|
Habitats |
380,933 |
2 |
190,467 |
4,77759 |
0,029770 |
|
Erreur |
478,400 |
12 |
39,867 |
||
Test t (jachère - Lisière)
Test t pour des Echantillons Indépendants
Note : Variables traitées comme des échantillon
|
Moyenne |
Moyenne |
valeur t dl |
p |
|
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
||
|
Jachère vs. Lisière |
12,40000 2,600000 3,050607 8 0,015806 |
|||
Test t (jachère - Forêt)
|
Test t pour des Echantillons Indépendants |
||||
|
Note : Variables traitées comme des échantillons |
||||
|
Moyenne |
Moyenne |
valeur t dl |
p |
|
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
||
|
Jachère vs. Forêt |
12,40000 14,00000 -0,336861 8 0,744889 |
|||
Test t (Lisière - Forêt)
|
Test t pour des Echantillons Indépendants |
||||
|
Note : Variables traitées comme des échantillon |
||||
|
Moyenne |
Moyenne |
valeur t dl |
p |
|
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
||
|
Lisière vs. Forêt |
2,600000 14,00000 -2,94740 8 0,018499 |
|||
Test sur Stochomys longicaudatus ANOVA (p = 0,008566)
|
Effet |
Tests Univariés de Significativité pour Effectifs Paramétrisation sigma-restreinte Décomposition efficace de l'hypothèse |
||||
|
SC |
Degr. de |
MC |
F |
p |
|
|
ord. origine |
68,26667 |
1 |
68,26667 |
9,846154 |
0,008566 |
|
Habitats |
6,53333 |
2 |
3,26667 |
0,471154 |
0,635356 |
|
Erreur |
83,20000 |
12 |
6,93333 |
||
Test t (jachère - Lisière : p= 0,035119) Test t (Jachère - forêt : p = 0,035119) Test t (Lisière - forêt : p=1,000000)
|
Test t pour des Echantillons Indépendants |
||||
|
Note : Variables traitées comme des échantillo |
||||
|
Moyenne |
Moyenne |
valeur t dl |
p |
|
|
Groupe1 vs. Groupe2 |
Groupe 1 |
Groupe 2 |
||
|
Lisière vs. Forêt |
2,600000 2,600000 0,00 8 1,000000 |
|||
|
Date |
Numero |
Espèces |
Station |
Sexe |
LP |
LC |
LQ |
|||||||||||||||||||||||
|
20/06/2010 |
LM1 |
Hylomyscus spp |
FL2S1 |
F |
60,9 |
83,3 |
35,6 |
|||||||||||||||||||||||
|
LM2 |
Stochomys longicaudatus |
FL4S1 |
M |
62,1 |
102 |
199,1 |
||||||||||||||||||||||||
|
LM3 |
Hybomys lunaris |
JL4S1 |
M |
45,8 |
76,1 |
89,9 |
||||||||||||||||||||||||
|
LM4 |
Praomys spp |
FL3S5 |
F |
47,7 |
70,6 |
136,2 |
||||||||||||||||||||||||
|
LM5 |
Lophuromys dudui |
LL1S3 |
F |
45,7 |
78,4 |
63,2 |
||||||||||||||||||||||||
|
LM6 |
Hybomys lunaris |
FL1S1 |
F |
41,2 |
60,9 |
78,9 |
||||||||||||||||||||||||
|
LM7 |
Praomys spp |
JL4S10 |
M |
23,4 |
41,9 |
94,4 |
||||||||||||||||||||||||
|
LM8 |
Hylomyscus spp |
FL2S2 |
F |
60 |
82,3 |
35,5 |
||||||||||||||||||||||||
|
LM9 |
Stochomys longicaudatus |
FL4S1 |
M |
63,2 |
101,5 |
198,2 |
||||||||||||||||||||||||
|
LM10 |
Hybomys lunaris |
JL3S1 |
M |
44,8 |
75 |
90 |
||||||||||||||||||||||||
|
LM11 |
Praomys spp |
FL3S5 |
F |
46,6 |
71,7 |
135,3 |
||||||||||||||||||||||||
|
LM12 |
Lophuromys dudui |
LL1S3 |
F |
46,8 |
77,4 |
61,3 |
||||||||||||||||||||||||
|
LM13 |
Hybomys lunaris |
FL2S1 |
F |
41,2 |
60,3 |
77,8 |
||||||||||||||||||||||||
|
LM14 |
Praomys spp |
JL4S10 |
M |
24,5 |
43 |
95 |
||||||||||||||||||||||||
|
21/06/2010 |
LM15 |
Stochomys longicaudatus |
FL2S10 |
M |
57,6 |
99,4 |
211,6 |
|||||||||||||||||||||||
|
LM16 |
Deomys ferrugineus |
FL1S10 |
M |
54,1 |
75,4 |
189 |
||||||||||||||||||||||||
|
LM17 |
Praomys spp |
JL3S5 |
M |
44,4 |
62,8 |
117,3 |
||||||||||||||||||||||||
|
LM18 |
Deomys ferrugineus |
FL1S14 |
F |
53 |
64 |
142,2 |
||||||||||||||||||||||||
|
LM19 |
Praomys spp |
LL1S5 |
M |
37,1 |
61,4 |
117,7 |
||||||||||||||||||||||||
|
LM20 |
Praomys spp |
JL2S3 |
M |
35,8 |
50 |
79,5 |
||||||||||||||||||||||||
|
LM21 |
Praomys spp |
FL3S10 |
M |
42,4 |
70,7 |
127,3 |
||||||||||||||||||||||||
|
LM22 |
Praomys spp |
LL5S1 |
M |
42,4 |
74 |
130,2 |
||||||||||||||||||||||||
|
LM23 |
Praomys spp |
FL2S8 |
M |
35,5 |
63,7 |
117,4 |
||||||||||||||||||||||||
|
LM24 |
Praomys spp |
LL1S7 |
M |
42 |
56,8 |
122,2 |
||||||||||||||||||||||||
|
LM25 |
Praomys spp |
LL2S7 |
M |
33,4 |
58,6 |
121,2 |
||||||||||||||||||||||||
|
LM26 |
Nannomys minutoides |
JL3S1 |
M |
26,6 |
41,7 |
49,6 |
||||||||||||||||||||||||
|
LM27 |
Stochomys longicaudatus |
LL2S7 |
M |
58,1 |
99,5 |
210,2 |
||||||||||||||||||||||||
|
LM28 |
Deomys ferrugineus |
FL1S7 |
M |
53,1 |
74,4 |
189 |
||||||||||||||||||||||||
|
LM29 |
Praomys spp |
JL3S5 |
M |
45,5 |
62,9 |
118,6 |
||||||||||||||||||||||||
|
LM30 |
Deomys ferrugineus |
LL1S3 |
F |
53 |
63 |
143,2 |
||||||||||||||||||||||||
|
LM31 |
Praomys spp |
FL1S1 |
M |
37,5 |
62,4 |
117,1 |
||||||||||||||||||||||||
|
LM32 |
Praomys spp |
JL2S4 |
M |
36,4 |
51 |
78,8 |
||||||||||||||||||||||||
|
LM33 |
Praomys spp |
FL3S10 |
M |
41,9 |
70,7 |
126,6 |
||||||||||||||||||||||||
|
LM34 |
Praomys spp |
LL5S2 |
M |
42 |
71 |
129,2 |
||||||||||||||||||||||||
|
LM35 |
Praomys spp |
FL2S8 |
M |
35,9 |
62,5 |
117,3 |
||||||||||||||||||||||||
|
LM36 |
Praomys spp |
LL1S8 |
M |
41,5 |
55,2 |
121,3 |
||||||||||||||||||||||||
|
22/06/2010 |
LM37 LM38 LM39 LM40 LM41 LM42 LM43 |
Praomys spp Nannomys minutoides Malacomys longipes Praomys spp Hylomyscus spp Praomys spp |
FL2S1 JL3S1 LL2S6 FL2S6 FL2S2 FL2S10 JL1S9 |
M M M M M F F |
34,4 26,5 70,9 31,5 39 |
57,4 41 46,5 70,5 |
120,2 48,6 162,7 100,6 113 135,1 153,2 |
|||||||||||||||||||||||
|
LM44 |
Hybomys lunaris |
FL1S6 |
F |
54,5 |
82,4 |
111,6 |
||||||||||||||||||||||||
|
LM45 |
Malacomys longipes |
FL1S1 |
M |
71 |
81,1 |
163 |
||||||||||||||||||||||||
|
LM46 |
Praomys spp |
FL2S9 |
M |
31,3 |
40 |
101,6 |
||||||||||||||||||||||||
|
LM47 |
Hylomyscus spp |
FL3S1 |
M |
40 |
45,5 |
112 |
||||||||||||||||||||||||
|
LM48 |
Praomys spp |
FL2S10 |
F |
42,5 |
57,5 |
134,2 |
||||||||||||||||||||||||
|
LM49 |
Praomys spp |
JL1S10 |
F |
50,8 |
70,7 |
150,1 |
||||||||||||||||||||||||
|
LM50 |
Hybomys lunaris |
FL1S6 |
F |
54,7 |
82,1 |
110,6 |
||||||||||||||||||||||||
|
23/06/2010 |
LM51 |
Praomys spp |
FL2S7 |
M |
45,5 |
72,9 |
146,7 |
|||||||||||||||||||||||
|
LM52 |
Malacomys longipes |
LL5S9 |
M |
86 |
108,2 |
194,8 |
||||||||||||||||||||||||
|
LM53 |
Hybomys lunaris |
FL5S7 |
M |
57,6 |
83,2 |
97,6 |
||||||||||||||||||||||||
|
LM54 |
Praomys spp |
FL2S9 |
M |
44,4 |
71,1 |
147,7 |
||||||||||||||||||||||||
|
LM55 |
Malacomys longipes |
LL5S8 |
M |
85,6 |
109,1 |
193,7 |
||||||||||||||||||||||||
|
LM56 |
Hybomys lunaris |
FL5S7 |
M |
57 |
82,1 |
98,8 |
||||||||||||||||||||||||
|
24/06/2010 |
LM57 |
Praomys spp |
JL2S8 |
M |
47,7 |
71,3 |
146,5 |
|||||||||||||||||||||||
|
LM58 |
Praomys spp |
JL1S6 |
M |
42,6 |
67 |
149,6 |
||||||||||||||||||||||||
|
LM59 |
Praomys spp |
JL4S3 |
F |
36,8 |
46 |
98,4 |
||||||||||||||||||||||||
|
LM60 |
Praomys spp |
LL1S4 |
F |
43,7 |
73,2 |
104,9 |
||||||||||||||||||||||||
|
LM61 |
Praomys spp |
JL1S10 |
M |
40,1 |
64 |
128,1 |
||||||||||||||||||||||||
|
LM62 |
Praomys spp |
JL1S8 |
M |
48,8 |
73,2 |
145,8 |
||||||||||||||||||||||||
|
LM63 |
Praomys spp |
JL1S6 |
M |
42,2 |
66 |
150,6 |
||||||||||||||||||||||||
|
LM64 |
Praomys spp |
JL4S2 |
F |
35,9 |
45,8 |
98,8 |
||||||||||||||||||||||||
|
LM65 |
Praomys spp |
LL1S4 |
F |
42,9 |
71,7 |
105,3 |
||||||||||||||||||||||||
|
LM66 |
Praomys spp |
JL2S10 |
M |
39,9 |
65 |
127,5 |
||||||||||||||||||||||||
|
25/06/2010 |
LM67 |
Praomys spp |
FL4S6 |
M |
38 |
52,8 |
123,2 |
|||||||||||||||||||||||
|
LM68 |
Graphiurus lorraineus |
FL2S2 |
F |
35,9 |
55 |
47,3 |
||||||||||||||||||||||||
|
LM69 |
Hybomys lunaris |
FL1S3 |
F |
38 |
80,2 |
97 |
||||||||||||||||||||||||
|
LM70 |
Praomys spp |
FL4S6 |
M |
48 |
53 |
122,1 |
||||||||||||||||||||||||
|
LM71 |
Hybomys lunaris |
FL3S3 |
F |
40 |
81 |
98 |
||||||||||||||||||||||||
|
26/06/2010 |
LM72 |
Lophuromys dudui |
LL3S9 |
F |
29,5 |
79,9 |
41,4 |
|||||||||||||||||||||||
|
LM73 |
Praomys spp |
LL5S2 |
M |
51,3 |
90,4 |
127,8 |
||||||||||||||||||||||||
|
LM74 |
Praomys spp |
LL4S8 |
F |
52,1 |
60,2 |
140,7 |
||||||||||||||||||||||||
|
LM75 |
Malacomys longipes |
FL3S10 |
M |
77,7 |
102,4 |
173,9 |
||||||||||||||||||||||||
|
LM76 |
Stochomys longicaudatus |
LL2S4 |
M |
52,6 |
81,5 |
209,3 |
||||||||||||||||||||||||
|
LM77 |
Lophuromys dudui |
LL3S8 |
F |
30,5 |
78,8 |
40,5 |
||||||||||||||||||||||||
|
LM78 |
Praomys spp |
LL5S2 |
M |
50,1 |
90 |
126,3 |
||||||||||||||||||||||||
|
LM79 |
Praomys spp |
LL4S7 |
F |
51 |
63,2 |
140,2 |
||||||||||||||||||||||||
|
LM80 |
Malacomys longipes |
FL3S10 |
M |
78,1 |
100,8 |
171,9 |
||||||||||||||||||||||||
|
LM81 |
Stochomys longicaudatus |
LL2S4 |
M |
53,5 |
80,4 |
207,9 |
||||||||||||||||||||||||
|
27/06/2010 |
LM82 |
Deomys ferrugineus |
JL3S6 |
M |
63,4 |
83,1 |
172,3 |
|||||||||||||||||||||||
|
Pas de |
||||||||||||||||||||||||||||||
|
LM83 |
Lophuromys dudui |
JL4S12 |
F |
28,1 |
76 |
queue |
||||||||||||||||||||||||
|
LM84 |
Hybomys lunaris |
LL1S4 |
M |
45,6 |
60,2 |
99,2 |
||||||||||||||||||||||||
|
LM85 |
Praomys spp |
FL1S7 |
M |
40,1 |
68,7 |
139 |
||||||||||||||||||||||||
|
LM86 |
Deomys ferrugineus |
JL3S5 |
M |
64,1 |
82 |
170,7 |
||||||||||||||||||||||||
|
Pas de |
||||||||||||||||||||||||||||||
|
LM87 |
Lophuromys dudui |
JL4S12 |
F |
29,1 |
76 |
queue |
||||||||||||||||||||||||
|
LM88 |
Hybomys lunaris |
LL1S6 |
M |
44,6 |
61,9 |
97,8 |
||||||||||||||||||||||||
|
LM89 |
Praomys spp |
FL1S1 |
M |
41,1 |
67,2 |
140 |
||||||||||||||||||||||||
|
28/06/2010 |
LM90 |
Hylomyscus spp |
LL3S8 |
M |
30,2 |
59,1 |
115,2 |
|||||||||||||||||||||||
|
LM91 |
Deomys ferrugineus |
LL3S10 |
F |
19,2 |
43,3 |
43,9 |
||||||||||||||||||||||||
|
LM92 |
Praomys spp |
FL1S8 |
M |
47,9 |
71 |
144,8 |
||||||||||||||||||||||||
|
LM93 |
Hylomyscus spp |
LL3S9 |
M |
30 |
57,1 |
114 |
||||||||||||||||||||||||
|
LM94 |
Deomys ferrugineus |
LL3S10 |
F |
19,8 |
41,1 |
41,9 |
||||||||||||||||||||||||
|
LM95 |
Praomys spp |
FL3S8 |
M |
48 |
72 |
145 |
||||||||||||||||||||||||
|
29/06/2010 |
LM96 |
Praomys spp |
FL4S2 |
M |
41,9 |
70,3 |
125,1 |
|||||||||||||||||||||||
|
LM97 |
Praomys spp |
LL4S6 |
M |
38,8 |
42,6 |
74,2 |
||||||||||||||||||||||||
|
LM98 |
Praomys spp |
JL1S6 |
F |
26,9 |
32,7 |
52,9 |
||||||||||||||||||||||||
|
LM99 |
Praomys spp |
FL4S3 |
M |
42 |
71,2 |
124,9 |
||||||||||||||||||||||||
|
LM100 |
Praomys spp |
LL4S6 |
M |
39,9 |
41,5 |
74,2 |
||||||||||||||||||||||||
|
LM101 |
Praomys spp |
JL2S7 |
F |
26,7 |
30,8 |
53 |
||||||||||||||||||||||||
|
23/07/2010 |
LM102 |
Lemniscomys striatus |
LL1S1 |
M |
56,4 |
87,7 |
124,1 |
|||||||||||||||||||||||
|
LM103 |
Stochomys longicaudatus |
JL1S5 |
M |
58,1 |
107 |
196,4 |
||||||||||||||||||||||||
|
LM104 |
Lophuromys dudui |
LL1S3 |
F |
39,2 |
78,5 |
57,5 |
||||||||||||||||||||||||
|
LM105 |
Hybomys lunaris |
LL4S4 |
F |
52,6 |
71,3 |
96 |
||||||||||||||||||||||||
|
LM106 |
Praomys spp |
FL1S8 |
M |
45,4 |
71,1 |
139,2 |
||||||||||||||||||||||||
|
LM107 |
Praomys spp |
JL4S6 |
F |
34,2 |
45,9 |
96,8 |
||||||||||||||||||||||||
|
LM108 |
Praomys spp |
FL4S9 |
F |
30,6 |
43 |
88,5 |
||||||||||||||||||||||||
|
LM109 |
Nannomys minutoides |
JL2S9 |
M |
27,1 |
40,2 |
46,7 |
||||||||||||||||||||||||
|
LM110 |
Praomys spp |
JL3S2 |
M |
42,3 |
71,6 |
121,5 |
||||||||||||||||||||||||
|
LM111 |
Hylomyscus spp |
FL1S2 |
M |
35,4 |
50,8 |
121,2 |
||||||||||||||||||||||||
|
LM112 |
Lemniscomys striatus |
LL1S2 |
M |
55,7 |
87,9 |
123,1 |
||||||||||||||||||||||||
|
LM113 |
Stochomys longicaudatus |
JL1S5 |
M |
59,3 |
106,6 |
196,8 |
||||||||||||||||||||||||
|
LM114 |
Lophuromys dudui |
LL1S1 |
F |
38,9 |
77,4 |
56,6 |
||||||||||||||||||||||||
|
LM115 |
Hybomys lunaris |
LL4S4 |
F |
51,7 |
73,3 |
96 |
||||||||||||||||||||||||
|
LM116 |
Praomys spp |
FL1S8 |
M |
45,4 |
70,7 |
138,3 |
||||||||||||||||||||||||
|
LM117 |
Praomys spp |
JL4S6 |
F |
34 |
46 |
95,5 |
||||||||||||||||||||||||
|
LM118 |
Praomys spp |
FL4S9 |
F |
31 |
44 |
87,5 |
||||||||||||||||||||||||
|
LM119 |
Nannomys minutoides |
JL2S8 |
M |
28 |
41,1 |
47,4 |
||||||||||||||||||||||||
|
LM120 |
Praomys spp |
JL3S2 |
M |
42,5 |
70,6 |
121,8 |
||||||||||||||||||||||||
|
LM121 |
Hylomyscus spp |
FL2S1 |
M |
34,4 |
59 |
120 |
||||||||||||||||||||||||
|
24/07/2010 |
LM122 |
Hybomys lunaris |
FL3S2 |
F |
47,2 |
68,7 |
51,3 |
|||||||||||||||||||||||
|
LM123 |
Praomys spp |
LL1S7 |
M |
44,9 |
58,4 |
116,5 |
||||||||||||||||||||||||
|
LM124 |
Praomys spp |
FL5S1 |
F |
28 |
46 |
100,5 |
||||||||||||||||||||||||
|
LM125 |
Stochomys longicaudatus |
LL1S5 |
F |
49 |
84,3 |
99,3 |
||||||||||||||||||||||||
|
LM126 |
Nannomys minutoides |
JL1S2 |
F |
19,5 |
38,1 |
49,8 |
||||||||||||||||||||||||
|
LM127 |
Hybomys lunaris |
FL3S1 |
F |
48,1 |
67,7 |
50,6 |
||||||||||||||||||||||||
|
LM128 |
Praomys spp |
LL1S7 |
M |
45 |
57,7 |
117 |
||||||||||||||||||||||||
|
LM129 |
Praomys spp |
FL5S1 |
F |
27,5 |
46 |
100 |
||||||||||||||||||||||||
|
LM130 |
Stochomys longicaudatus |
LL1S3 |
F |
48 |
84 |
97,8 |
||||||||||||||||||||||||
|
LM131 |
Nannomys minutoides |
JL1S2 |
F |
19,7 |
37,6 |
48,8 |
||||||||||||||||||||||||
|
25/07/2010 |
LM132 |
Praomys spp |
JL1S5 |
M |
42,9 |
61,3 |
126,5 |
|||||||||||||||||||||||
|
LM133 |
Praomys spp |
FL1S7 |
M |
42,9 |
74,8 |
139,9 |
||||||||||||||||||||||||
|
LM134 |
Lophuromys dudui |
JL3S6 |
F |
33,4 |
73 |
50,2 |
||||||||||||||||||||||||
|
LM135 |
Hybomys lunaris |
JL2S4 |
F |
36,1 |
45 |
64,6 |
||||||||||||||||||||||||
|
LM136 |
Praomys spp |
FL2S6 |
F |
27,8 |
57,3 |
117 |
||||||||||||||||||||||||
|
LM137 |
Praomys spp |
JL1S4 |
M |
43 |
60,6 |
125,5 |
||||||||||||||||||||||||
|
LM138 |
Praomys spp |
FL1S7 |
M |
41 |
75 |
140 |
||||||||||||||||||||||||
|
LM139 |
Lophuromys dudui |
JL3S4 |
F |
32,6 |
73,3 |
51 |
||||||||||||||||||||||||
|
LM140 |
Hybomys lunaris |
JL2S4 |
F |
35,3 |
44,9 |
64,5 |
||||||||||||||||||||||||
|
LM141 |
Praomys spp |
FL1S6 |
F |
27 |
56,7 |
117,5 |
||||||||||||||||||||||||
|
26/07/2010 |
LM142 |
Praomys spp |
FLIS4 |
F |
24 |
43 |
113,8 |
|||||||||||||||||||||||
|
LM143 |
Stochomys longicaudatus |
FL1S8 |
M |
44,5 |
73,7 |
73,4 |
||||||||||||||||||||||||
|
LM144 |
Hybomys lunaris |
FL4S1 |
M |
52,6 |
82,9 |
116 |
||||||||||||||||||||||||
|
LM145 |
Praomys spp |
FLIS1 |
F |
25 |
42,8 |
112,8 |
||||||||||||||||||||||||
|
LM146 |
Stochomys longicaudatus |
FL2S2 |
M |
45 |
72,6 |
73,3 |
||||||||||||||||||||||||
|
LM147 |
Hybomys lunaris |
FL4S1 |
M |
53 |
83 |
115,8 |
||||||||||||||||||||||||
|
27/07/2010 |
LM148 |
Stochomys longicaudatus |
LL3S6 |
M |
44,4 |
76,5 |
93,2 |
|||||||||||||||||||||||
|
LM149 |
Praomys spp |
LL1S6 |
F |
33,9 |
54,6 |
101 |
||||||||||||||||||||||||
|
LM150 |
Stochomys longicaudatus |
LL4S6 |
M |
45,3 |
75,5 |
92,2 |
||||||||||||||||||||||||
|
LM151 |
Praomys spp |
LL1S6 |
F |
34 |
54 |
102 |
||||||||||||||||||||||||
|
28/07/2010 |
LM152 |
Praomys spp |
JL3S4 |
M |
38,5 |
61,3 |
126,2 |
|||||||||||||||||||||||
|
LM153 |
Oenomys hypoxanthus |
JL1S8 |
F |
34,4 |
55,2 |
103,3 |
||||||||||||||||||||||||
|
LM154 |
Praomys spp |
FL1S10 |
M |
43,9 |
74 |
134,8 |
||||||||||||||||||||||||
|
LM155 |
Praomys spp |
JL3S4 |
M |
39 |
60 |
125,3 |
||||||||||||||||||||||||
|
LM156 |
Praomys spp |
FL5S10 |
M |
44,9 |
74,5 |
133,7 |
||||||||||||||||||||||||
|
29/07/2010 |
LM157 |
Stochomys longicaudatus |
JL4S4 |
F |
46,2 |
59 |
99,2 |
|||||||||||||||||||||||
|
LM158 |
Deomys ferrugineus |
FL2S9 |
F |
61,1 |
96,1 |
110,4 |
||||||||||||||||||||||||
|
LM159 |
Lophuromys dudui |
LL2S1 |
F |
25,7 |
65,6 |
45,2 |
||||||||||||||||||||||||
|
LM160 |
Hybomys lunaris |
JL4S7 |
F |
47,2 |
75,9 |
101,9 |
||||||||||||||||||||||||
|
LM161 |
Praomys spp |
FL4S1 |
F |
29 |
42 |
94,2 |
||||||||||||||||||||||||
|
LM162 |
Stochomys longicaudatus |
JL4S4 |
F |
45,2 |
58,7 |
98,8 |
||||||||||||||||||||||||
|
LM163 |
Deomys ferrugineus |
FL2S8 |
F |
60 |
96,6 |
111 |
||||||||||||||||||||||||
|
LM164 |
Lophuromys dudui |
LL1S1 |
F |
26,1 |
66,6 |
44 |
||||||||||||||||||||||||
|
LM165 |
Hybomys lunaris |
JL4S7 |
F |
48 |
76 |
101,2 |
||||||||||||||||||||||||
|
LM166 |
Praomys spp |
FL4S2 |
F |
28,9 |
41,3 |
93,1 |
||||||||||||||||||||||||
|
30/07/2010 |
LM167 |
Nannomys minutoides |
FL5S3 |
F |
20,3 |
38,2 |
51,2 |
|||||||||||||||||||||||
|
LM168 |
Nannomys minutoides |
JL1S11 |
M |
25,5 |
40 |
48,3 |
||||||||||||||||||||||||
|
LM169 |
Nannomys minutoides |
FL5S5 |
F |
21 |
38 |
50,5 |
||||||||||||||||||||||||
|
31/07/2010 |
LM170 |
Praomys spp |
JL3S2 |
F |
33,5 |
60,6 |
119,1 |
|||||||||||||||||||||||
|
LM171 |
Praomys spp |
LL5S10 |
M |
30,6 |
44,9 |
86,4 |
||||||||||||||||||||||||
|
LM172 |
Hylomyscus spp |
LL1S2 |
F |
28,9 |
50,2 |
103,9 |
||||||||||||||||||||||||
|
LM173 |
Praomys spp |
JL2S2 |
F |
34,5 |
61,1 |
119,6 |
||||||||||||||||||||||||
|
LM174 |
Praomys spp |
LL5S10 |
M |
31 |
45 |
87 |
||||||||||||||||||||||||
|
LM175 |
Hylomyscus spp |
LL1S3 |
F |
29 |
51,3 |
102,9 |
||||||||||||||||||||||||
|
01/08/2010 |
LM176 |
Praomys spp |
LL4S9 |
F |
31,1 |
46,5 |
112,1 |
|||||||||||||||||||||||
|
LM177 |
Praomys spp |
LL3S6 |
M |
30,8 |
59,9 |
126,2 |
||||||||||||||||||||||||
|
LM178 |
Praomys spp |
LL5S5 |
F |
44,2 |
75,4 |
137,7 |
||||||||||||||||||||||||
|
LM179 |
Praomys spp |
LL4S10 |
F |
30 |
47,1 |
112 |
||||||||||||||||||||||||
|
LM180 |
Praomys spp |
LL3S6 |
M |
31 |
58,8 |
127,4 |
||||||||||||||||||||||||
|
LM181 |
Praomys spp |
LL5S5 |
F |
45,3 |
74,7 |
135,5 |
||||||||||||||||||||||||
|
17/08/2010 |
LM182 |
Praomys spp |
JL3S7 |
F |
39,3 |
62,9 |
123,6 |
|||||||||||||||||||||||
|
LM183 |
Deomys ferrugineus |
FL2S1 |
M |
58,5 |
102,7 |
189,5 |
||||||||||||||||||||||||
|
LM184 |
Malacomys longipes |
FL5S1 |
M |
49 |
80,2 |
125 |
||||||||||||||||||||||||
|
LM185 |
Praomys spp |
FL1S7 |
F |
40,1 |
65,6 |
109,4 |
||||||||||||||||||||||||
|
LM186 |
Hylomyscus spp |
JL1S1 |
M |
46,2 |
78,7 |
133 |
||||||||||||||||||||||||
|
LM187 |
Praomys spp |
JL1S7 |
F |
37,9 |
59 |
105,2 |
||||||||||||||||||||||||
|
LM188 |
Praomys spp |
LL5S8 |
F |
42,1 |
80 |
101,9 |
||||||||||||||||||||||||
|
LM189 |
Praomys spp |
JL3S13 |
F |
32,4 |
45,5 |
79,8 |
||||||||||||||||||||||||
|
LM190 |
Praomys spp |
JL4S12 |
M |
37,4 |
47,8 |
80,6 |
||||||||||||||||||||||||
|
LM191 |
Praomys spp |
FL3S6 |
M |
40,2 |
72,8 |
121,5 |
||||||||||||||||||||||||
|
LM192 |
Praomys spp |
JL3S7 |
F |
38,4 |
63,3 |
123,6 |
||||||||||||||||||||||||
|
LM193 |
Deomys ferrugineus |
FL2S1 |
M |
57 |
101,5 |
189,5 |
||||||||||||||||||||||||
|
LM194 |
Malacomys longipes |
FL5S7 |
M |
48,8 |
80,6 |
125 |
||||||||||||||||||||||||
|
LM195 |
Praomys spp |
FL1S8 |
F |
42,1 |
66 |
109,4 |
||||||||||||||||||||||||
|
LM196 |
Hylomyscus spp |
JL1S1 |
M |
46,6 |
77,8 |
130,1 |
||||||||||||||||||||||||
|
LM197 |
Praomys spp |
JL1S7 |
F |
38 |
58,8 |
103,2 |
||||||||||||||||||||||||
|
LM198 |
Praomys spp |
LL5S8 |
F |
41,1 |
81,8 |
102 |
||||||||||||||||||||||||
|
LM199 |
Praomys spp |
JL3S13 |
F |
33,3 |
45 |
78,7 |
||||||||||||||||||||||||
|
LM200 |
Praomys spp |
JL4S12 |
M |
38,2 |
47,7 |
81,6 |
||||||||||||||||||||||||
|
LM201 |
Praomys spp |
FL3S6 |
M |
41 |
73 |
120,6 |
||||||||||||||||||||||||
|
18/08/2010 |
LM202 |
Praomys spp |
FL5S9 |
F |
38,6 |
45,4 |
94,4 |
|||||||||||||||||||||||
|
LM203 |
Praomys spp |
JL1S1 |
M |
50,3 |
64,9 |
110,3 |
||||||||||||||||||||||||
|
LM204 |
Praomys spp |
JL3S13 |
M |
42,4 |
74 |
71,9 |
||||||||||||||||||||||||
|
LM205 |
Praomys spp |
FL5S5 |
F |
39 |
46,6 |
93,3 |
||||||||||||||||||||||||
|
LM206 |
Praomys spp |
JL1S1 |
M |
51,2 |
65,4 |
111,3 |
||||||||||||||||||||||||
|
LM207 |
Praomys spp |
JL3S13 |
M |
42 |
73,6 |
72 |
||||||||||||||||||||||||
|
19/08/2010 |
LM208 |
Hylomyscus spp |
JL2S2 |
F |
48 |
61,8 |
109,5 |
|||||||||||||||||||||||
|
LM209 |
Hylomyscus spp |
JL1S1 |
F |
47,9 |
60 |
105,4 |
||||||||||||||||||||||||
|
20/08/2010 |
LM210 |
Hybomys lunaris |
LL5S1 |
F |
50,6 |
71,1 |
83,4 |
|||||||||||||||||||||||
|
LM211 |
Hybomys lunaris |
LL5S5 |
F |
52 |
70,8 |
81,6 |
||||||||||||||||||||||||
|
21/08/2010 |
LM212 |
Hybomys lunaris |
FL5S6 |
M |
68,3 |
81,4 |
71,1 |
|||||||||||||||||||||||
|
LM213 |
Hylomyscus spp |
LL4S8 |
M |
45,1 |
57,1 |
125,2 |
||||||||||||||||||||||||
|
LM214 |
Praomys spp |
LL3S5 |
F |
42,2 |
42,9 |
75,8 |
||||||||||||||||||||||||
|
LM215 |
Hybomys lunaris |
FL5S6 |
M |
68,5 |
82,4 |
70,9 |
||||||||||||||||||||||||
|
LM216 LM217 |
Hylomyscus spp Praomys spp |
LL4S8 |
M |
44,4 |
58,1 |
126 76 |
||||||||||||||||||||||||
|
22/08/2010 |
LM218 |
Lemniscomys striatus |
JL2S8 |
F |
63,1 |
90 |
115,6 |
|||||||||||||||||||||||
|
LM219 |
Praomys spp |
FL4S1 |
F |
43,4 |
52,8 |
76,7 |
||||||||||||||||||||||||
|
LM220 |
Praomys spp |
FL2S1 |
F |
46,1 |
52,6 |
101,6 |
||||||||||||||||||||||||
|
LM221 |
Praomys spp |
FL5S10 |
F |
46,5 |
54,5 |
135 |
||||||||||||||||||||||||
|
LM222 |
Praomys spp |
LL2S5 |
M |
37,7 |
42,5 |
97,8 |
||||||||||||||||||||||||
|
LM223 |
Lemniscomys striatus |
JL3S8 |
F |
63 |
91,3 |
114,5 |
||||||||||||||||||||||||
|
LM224 |
Praomys spp |
FL4S1 |
F |
44,5 |
53 |
77 |
||||||||||||||||||||||||
|
LM225 |
Praomys spp |
FL1S1 |
F |
45,3 |
51,7 |
102,3 |
||||||||||||||||||||||||
|
LM226 |
Praomys spp |
FL5S10 |
F |
47 |
53,5 |
136 |
||||||||||||||||||||||||
|
23/08/2010 |
LM227 |
Praomys spp |
FL5S2 |
F |
53,2 |
80 |
138,5 |
|||||||||||||||||||||||
|
LM228 |
Praomys spp |
FL2S2 |
F |
53,4 |
79,5 |
139,6 |
||||||||||||||||||||||||
|
24/08/2010 |
LM229 |
Hylomyscus spp |
FL5S4 |
F |
31,3 |
55,3 |
99,1 |
|||||||||||||||||||||||
|
LM230 |
Praomys spp |
JL2S4 |
M |
37,2 |
60,2 |
112,1 |
||||||||||||||||||||||||
|
LM231 |
Hylomyscus spp |
FL5S4 |
F |
30,5 |
55 |
98,4 |
||||||||||||||||||||||||
|
LM232 |
Praomys spp |
JL2S4 |
M |
38,3 |
61 |
111,1 |
||||||||||||||||||||||||
|
25/08/2010 |
LM233 |
Lophuromys dudui |
JL4S4 |
M |
32,8 |
80,9 |
49 |
|||||||||||||||||||||||
|
LM234 |
Praomys spp |
FL3S6 |
M |
30,7 |
47,4 |
76,3 |
||||||||||||||||||||||||
|
LM235 |
Lophuromys dudui |
JL2S4 |
M |
33 |
81 |
50 |
||||||||||||||||||||||||
|
LM236 |
Praomys spp |
FL1S5 |
M |
31,5 |
47,5 |
77,7 |
||||||||||||||||||||||||