
FOSAS PROJECT
UNIVERSITE DE DSCHANG
IRAD



IRAD
FACULTE D'AGRONOMIE ET DES SCIENCES AGRICOLES
(FASA)
FACULTY OF AGRONOMY AND AGRICULTURAL SCIENCES
(FASA)
DEPARTEMENT DE FORESTERIE
DEPARTMENT OF FORESTRY
INVENTAIRE DE LA FAUNE SAUVAGE SUR LES
TRANSECTS
PERMANENTS EN PERIPHERIE NORD-EST ET DANS LE
PARC
NATIONAL DE BOUMBA-BEK, SUD-EST CAMEROUN
Mémoire présenté en
vue de l'obtention du diplôme d'Ingénieur des Eaux, Forêts
et Chasses
Par
DJEKDA Daniel
Matricule:
CM04-08ASA0110
Septembre 2014
UNIVERSITE DE DSCHANG

FOSAS PROJECT
UNIVERSITE DE DSCHANG



IRAD
FACULTE D'AGRONOMIE ET DES SCIENCES AGRICOLES
(FASA)
FACULTY OF AGRONOMY AND AGRICULTURAL SCIENCES
(FASA)
DEPARTEMENT DE FORESTERIE
DEPARTMENT OF FORESTRY
INVENTAIRE DE LA FAUNE SAUVAGE SUR LES
TRANSECTS
PERMANENTS EN PERIPHERIE NORD-EST ET DANS LE
PARC
NATIONAL DE BOUMBA-BEK, SUD-EST CAMEROUN
Mémoire présenté en
vue de l'obtention du diplôme d'Ingénieur des Eaux, Forêts
et Chasses
Par
DJEKDA Daniel
Matricule : CM04-08ASA0110
16e Promotion FASA
Encadreur
Mr KAMGAING William
Ingénieur des Eaux, Forêts et
Chasses
Direction de la Faune et des Aires
Protégées, Ministère des Forêts et de la
Faune
Superviseur
Dr BOBO Kadiri Serge
Directeur de l'Ecole de Faune de Garoua, Chargé de
cours au Département de foresterie, FASA
Université de Dschang
Septembre 2014
FICHE DE CERTIFICATION DE L'ORIGINALITE DU
TRAVAIL
Je soussigné, DJEKDA Daniel, atteste
que le présent mémoire est le fruit de mes propres travaux
effectués en périphérie Nord-est et dans le Parc National
de Boumba-Bek, dans le cadre de l'initiative «Projet JICA/FOSAS:
IRAD de Yaoundé ``Forest Savana Sustainability'', Projet
Cameroun ». Ce travail a été effectué
sous la supervision scientifique de Dr BOBO Kadiri Serge, enseignant au
Département de Foresterie, Faculté d'Agronomie et des Sciences
Agricoles de l'Université de Dschang, Directeur de l'Ecole de Faune de
Garoua et, l'encadrement technique de M. KAMGAING William, en service au
Ministère des Forêts et de la Faune.
Ce mémoire est authentique et n'a été
l'objet d'aucune présentation antérieure pour l'acquisition de
quelque grade universitaire que ce soit.
Visa de l'auteur
DJEKDA Daniel
Date :
Visa du Superviseur Visa du Chef de Département de
Foresterie
Dr Bobo Kadiri Serge
Date : Date
FICHE DE CERTIFICATION DES CORRECTIONS APRES
SOUTENANCE
Le présent mémoire a été
revu et corrigé conformément aux observations du jury.
Visa du Superviseur :
Dr Bobo Kadiri Serge
Date :
Visa du Président du Jury:
Date :
Visa du Chef de département de Foresterie:
Date :
DEDICACE
Je dédie ce mémoire de Master Professionnel en
Faune Aquatique et Terrestre à mon Dieu Le Tout Puissant et à
toutes personnes passionnées par la faune sauvage de part le monde
entier.
REMERCIEMENTS
La réalisation de ce mémoire a été
rendue possible grâce au concours de plusieurs personnes à qui
j'exprime ici ma profonde gratitude.
Je pense de prime abord au Dr Bobo Kadiri Serge, Directeur de
l'Ecole de Faune de Garoua et Chargé de Cours au département de
Foresterie, à la Faculté d'Agronomie et des Sciences Agricoles de
l'Université de Dschang pour la supervision scientifique de ce document,
malgré ses multiples occupations.
Je remercie aussi toute l'équipe du Projet JICA/FOSAS:
IRAD de Yaoundé qui a bien accepté et financé ces travaux
de recherche, malgré une démarche administrative très
longue, je tiens encore à vous adresser mes sincères
remerciements.
Ma reconnaissance va également:
ü A mon encadreur KAMGAING William, qui a voulu bien
consacré son temps pour ses multiples orientations vis-à-vis de
l'analyse des données et les commentaires associés lors de la
relecture dudit document;
ü Au Pr TCHAMBA Martin, enseignant à la
Faculté d'Agronomie et des Sciences Agricoles de l'Université de
Dschang et Chef de Département de Foresterie, pour ses multiples
conseils;
ü Au Pr TSI Evaristus, et Dr Tumenta tous enseignants
à la Faculté d'Agronomie et des Sciences Agricoles de
l'Université de Dschang, pour leurs multiples conseils ;
ü Au Doyen de la Faculté d'Agronomie et des
Sciences Agricoles, le Professeur MANJELI Yacouba et son staff administratif,
ainsi qu'à tous les enseignants, particulièrement ceux du
Département de Foresterie ;
ü Au Pr ICHIKAWA fondateur du Projet FOSAS, Pr ARAKI,
tous pour leurs encouragements sans cesse ;
ü Au Dr HAYASHI Koji, anthropologue et chercheur
Japonais, qui, durant ces travaux de recherche sur le terrain m'a beaucoup
encouragé, avec qui, nous avions effectué des multiples descentes
de terrain à Gribé pour d'éventuel travaux des
caméras infra-rouges ;
ü Au Dr MASSAI Hirai, chercheur Japonais, pour sa bonne
collaboration ;
ü Au Dr FOAHAM enseignant à l'Université de
Yaoundé 1, team leader dans le Projet JICA/FOSAS, pour son encouragement
sans cesse;
ü A Mme MARUTA EIKO et Mr SHINNA, tous deux responsables
administratifs dans le projet FOSAS pour leurs encouragements et les conseils
respectifs qu'ils m'ont toujours prodigué durant ces travaux de
recherche ;
ü A tous mes camarades stagiaires NDZEFACK Brice, BOBO
René, BALANGOUNDE, NGONGANG pour leurs encouragements tout au long du
stage ;
ü A Mr Bonito CHIA, Paulin KENGNE, qui n'ont
orienté dans l'analyse de mes données et dans l'apprentissage des
logiciels éventuels ;
ü A tous mes camarades de la 16e Promotion avec lesquels
nous avons partagé des moments simplement inoubliables. Je prie que
l'Eternel se souvienne de chacun d'eux
ü A toute la population de zone d'étude
Gribé et Gouonepoun pour leur accueil chaleureux et pour avoir
accepté notre présence dans leur milieu, je pense
particulièrement à mes redoutables guides de terrain entre autres
MBIAP Mermoze, MEDJINANDJO Bertrand, LENGO Jean Paul, KANI Remi, KITA
Christaller, pour le site de Gribé et MEDOUMETO Hilaire, SERBOT Simon,
BONOULOU Mathias, pour le site de Gouonepoun, tous pour leur courage et leur
solidarité et aussi sans qui toutes les données d'inventaire
faunique n'auraient pu être collectées, mes chers commandos je
tiens sincèrement ici à vous tirer un coup de chapeau, encore
merci ;
ü A ma dulcinée ZADANA SALI Sandrine, pour tout
son amour, son soutien et sa dévotion à mon égard;
Je souhaite enfin exprimer tout mon amour et ma reconnaissance
envers ma famille qui m'a soutenu, m'a encouragé tout au long de mes
études et m'a permis de trouver la motivation nécessaire pour
avancer sur le plan professionnel et personnel.
SOMMAIRE
REMERCIEMENTS
I
DEDICACE
I
SOMMAIRE
IV
LISTE DES TABLEAUX
VIII
LISTE DES FIGURES
XI
LISTE DES ANNEXES
XIV
LISTE DES ACRONYMES ET ABREVIATIONS
XV
RESUME
XVI
ABSTRACT
XVII
CHAPITRE 1:
INTRODUCTION
1
1.1 CONTEXTE
1
1.2 PROBLÉMATIQUE
1
1.3 OBJECTIFS
2
1.3.1 Objectif Global
2
1.3.2 Objectifs spécifiques
2
1.4 IMPORTANCE DE L'ÉTUDE
3
1.5 LIMITES DE L'ÉTUDE
3
CHAPITRE 2: REVUE
DE LA LITTERATURE
4
2.1 CLARIFICATION DES CONCEPTS
4
2.1.1 Aire protégée
4
2.1.2 Diversité biologique
4
2.1.3 Espèces rares
5
2.1.4 Grands et moyens mammifères
5
2.1.5 Parc National
5
2.1.6 Transect
6
2.1.7 Zone périphérique
6
2.2 GÉNÉRALITÉS SUR QUELQUES
TECHNIQUES D'INVENTAIRE DE LA FAUNE EN ZONE DE FORET
6
2.2.1 Généralités
6
2.2.2 Quelques méthodes d'inventaire de
la faune en zone de forêt.
6
2.2.2.1 Cris d'appel des chasseurs, attractifs et
postes d'observation
6
2.2.2.2 Recensement au moyen des attractifs
7
2.2.2.3 Postes d'observation
7
2.2.2.4 Transects en forêt
7
2.2.2.5 Pièges photographiques
7
2.3 THÉORIE DES TRANSECTS
LINÉAIRES
8
2.4 PRINCIPE DES TRANSECTS LINÉAIRES
9
2.5 DÉTERMINATION DES DENSITÉS DES
GRANDS ET MOYENS MAMMIFÈRES À PARTIR DES CROTTES
10
2.6 VITESSE DE DÉGRADATION OU DISPARITION
DES CROTTES DES GRANDS ET MOYENS MAMMIFÈRES.
10
2.7 SYNTHÈSE DE QUELQUES ÉTUDES SUR
LE POTENTIEL FAUNIQUE EN PÉRIPHÉRIE NORD DU PNBB
11
2.8 RICHESSE SPECIFIQUE DANS LES ZICGC 13 ET 14 AU
NORD DU PARC NATIONAL DE BOUMBA - BEK
11
CHAPITRE 3:
METHODOLOGIE
12
3.1 DESCRIPTION DE LA ZONE D'ÉTUDE
12
3.1.1 Localisation
12
3.1.2 Caractéristiques biophysiques et
écologiques
13
3.1.2.1 Climat
13
3.1.2.2 Relief et topographie
13
3.1.2.3 Pédologie et géologie
13
3.1.2.4 Hydrologie
13
3.1.2.5 La végétation
13
3.1.2.6 La faune
14
3.1.3 Relations Faune - Flore
16
3.1.4 Caractéristiques socioculturelles
et démographiques
16
3.1.4.1 Populations natives
16
3.1.4.2 Populations migrantes
17
3.1.4.3 Répartition spatiale
18
3.1.4.4 Infrastructures sociales
19
3.1.5 Caractéristiques
socioéconomiques
20
3.1.5.1 Agriculture
20
3.1.5.2 Chasse
20
3.1.5.3 Pêche
21
3.1.5.4 Cueillette
22
3.1.5.5 Exploitation forestière
22
3.1.5.6 Chasse sportive
22
3.1.5.7 Capture des oiseaux
22
3.1.5.8 Exploitation minière artisanale
23
3.2 COLLECTE DES DONNÉES
23
3.2.1 Le choix de site
23
3.2.2 Données de sources secondaires
23
3.2.3 Données de sources primaires
23
3.2.3.1 Technique d'inventaire diurne:
méthode directe
24
3.2.3.2 Technique d'inventaire diurne :
méthode indirecte
25
3.2.3.3 Technique d'inventaire nocturne :
méthode directe
25
3.2.3.4 Utilisation des caméras
infra-rouges: méthode directe
26
3.2.3.5 Suivi de dégradation des crottes des
grands et moyens mammifères
28
3.2.3.6 Caractérisation de la
végétation de la faune dans la zone d'étude
29
3.3 ANALYSE DES DONNÉES
29
3.3.1 Inventaires diurnes et nocturnes
29
3.3.1.1 Estimation des densités et de
l'abondance relative
29
3.3.1.2 Comparaison des densités et de
l'abondance relative (Inventaires diurnes et nocturnes)
30
3.3.2 Caméras infra-rouges
31
3.3.2.1 Estimation des densités et de
l'abondance relative
31
3.3.2.2 Comparaison des densités et de
l'abondance relative (WPI)
31
3.3.3 Evaluation de la performance de
différentes méthodes
31
3.3.4 Analyse de la vitesse de
décomposition ou disparition de crottes
32
3.3.5 Caractérisation de la
végétation de la faune dans la zone d'étude
32
CHAPITRE 4:
RESULTATS ET DISCUSSION
33
4.1 COMPARAISON DES DENSITÉS ET DES INDICES
KILOMÉTRIQUES D'ABONDANCE (IKA) DES GRANDS ET MOYENS MAMMIFÈRES
SUIVANTS QUATRE MÉTHODES D'INVENTAIRE DANS LES TROIS SITES
33
4.1.1 Inventaires diurnes indirects
33
4.1.1.1 Richesse spécifique
33
4.1.1.2 Estimation des densités des grands
et moyens mammifères dans les trois sites (Inventaires diurnes
indirects)
34
4.1.1.3 Comparaison des densités du
céphalophe bleu entre les trois sites (Inventaires diurnes
indirects)
37
4.1.1.4 Estimation des densités de
Céphalophe roux entre les trois sites (Inventaires diurnes
indirects)
38
4.1.1.5 Comparaison des densités de
Céphalophe roux entre les trois sites (Inventaires diurnes
indirects)
42
4.1.1.6 Estimation de l'abondance relative des
espèces entre les trois sites (Inventaires diurnes indirects)
42
4.1.1.7 Comparaison des IKA du Céphalophe
bleu entre les trois sites (Inventaires diurnes indirects)
44
4.1.1.8 Comparaison des IKA du Céphalophe
roux entre les trois sites (Inventaires diurnes indirects)
46
4.1.1.9 Comparaison des IKA du Céphalophe
à dos jaune entre les trois sites (Inventaires diurnes indirects)
47
4.1.1.10 Comparaison des IKA Bongo entre les trois
sites (Inventaires diurnes indirects)
48
4.1.1.11 Comparaison des IKA Eléphant entre
les trois sites (Inventaires diurnes indirects)
53
4.1.1.12 Comparaison des IKA Gorilles entre les
trois sites (Inventaires diurnes indirects)
54
4.1.1.13 Comparaison des IKA Pangolin à
longue queue entre les trois sites (Inventaires diurnes indirects)
56
4.1.1.14 Comparaison des IKA Pangolin géant
entre les trois sites (Inventaires diurnes indirects)
57
4.1.1.15 Comparaison des IKA Athérure
africain entre les trois sites (Inventaires diurnes indirects)
59
4.1.1.16 Comparaison des IKA Potamochère
entre les trois sites (Inventaires diurnes indirects)
61
4.1.1.17 Comparaison des IKA des grands et moyens
mammifères entre les trois sites (Inventaires diurnes indirects)
62
4.1.1.18 Activités Humaines
63
4.1.1.19 Comparaison des IKA des activités
humaines entre les trois sites (Inventaires diurnes indirects)
63
4.1.2 Inventaires diurnes directs
64
4.1.2.1 Estimation des densités de groupes
des petits singes entre les trois sites (Inventaires diurnes directs)
64
4.1.2.2 Comparaison des densités de groupes
de petits singes entre les trois sites (Inventaires diurnes directs)
68
4.1.2.3 Estimation de l'abondance relative des
espèces entre les trois sites (Inventaires diurnes directs).
69
4.1.2.4 Comparaison des IKA de petits singes entre
les trois (Inventaires diurnes directs)
69
4.1.2.5 Comparaison IKA global de grands et moyens
mammifères entre les trois sites (inventaires diurnes directs)
70
4.1.3 Inventaires nocturnes
71
4.1.3.1 Estimation de l'abondance relative des
espèces entre les trois sites (Inventaires nocturnes)
71
4.1.3.2 Comparaison de l'abondance relative des
espèces entre les trois sites (Inventaires nocturnes)
72
4.1.3.3 Comparaison IKA de tous les grands et
moyens mammifères entre les trois sites (Inventaires nocturnes)
73
4.1.3.4 Estimation des densités des
espèces recensées entre les trois sites (Inventaires
nocturnes)
73
4.1.3.5 Comparaison des densités de
Céphalophes bleus entre les trois sites (Inventaires nocturnes)
77
4.1.4 Caméras infra-rouges
78
4.1.4.1 Estimation des densités au moyen des
Caméras infra-rouges entre les trois sites
78
4.1.4.2 Comparaison des densités des
espèces au moyen des Caméras infra-rouges entre les trois
79
4.1.4.3 Estimation de Wildlife Picture Index (WPI)
dans les trois sites
80
4.1.4.4 Comparaison WPI des différentes
espèces entre les trois sites (Caméras infra-rouges)
82
4.1.4.5 Comparaison de WPI de tous les
mammifères entre les trois sites (Caméra infra-rouge)
83
4.1.5 Comparaison des résultats avec
ceux obtenus dans la région du Sud-est, Cameroun
84
4.1.6 Quelques vitesses de dégradation
des crottes des espèces identifiées dans la zone
d'étude
87
4.1.7 Stade de décomposition des
crottes
87
4.1.8 Caractérisation de la
végétation le long des transects dans la zone d'étude
89
4.1.8.1 Inventaires des ligneux le long des
transects dans la zone d'étude
89
4.1.8.2 Inventaires des herbacées le long
des transects dans la zone d'étude
90
4.2 EVALUATION DE LA PERFORMANCE DES
MÉTHODES D'INVENTAIRES DE LA FAUNE TESTÉES
90
4.3 DÉTECTION DES ESPÈCES RARES
92
4.4 FORMULATION DE QUELQUES SUGGESTIONS AFIN
D'ORIENTER LE MODÈLE DE GESTION DE LA RESSOURCE FAUNE SAUVAGE DANS LA
ZONE D'ÉTUDE
93
4.4.1 Information, éducation et
Communication efficace au profit de la communauté locale
93
4.4.2 Lutte anti-braconnage
93
4.4.3 Suivi permanent de la faune
95
CHAPITRE 5:
CONCLUSION ET RECOMMANDATIONS
96
5.1 CONCLUSION
96
5.2 RECOMMANDATIONS
97
BIBLIOGRAPHIE
99
ANNEXES
105
LISTE
DES TABLEAUX
Tableau 1: Caractéristiques des
caméras Bushnell Trophy Cam
8
Tableau 2: Richesse spécifique des grands et
moyens mammifères dans quelques sites du Sud-est, Cameroun
11
Tableau 3: Définition d'âges des
indices de présence des espèces rencontrées
29
Tableau 4: Statistique d'estimation de la
densité de Céphalophe bleu dans le site de Gribé
(Inventaires diurnes indirects)
34
Tableau 5: Statistique d'estimation de la
densité de Céphalophe bleu dans le site de Gouonepoun
(Inventaires diurnes indirects)
35
Tableau 6: Statistique d'estimation de la
densité de Céphalophe bleu dans le PNBB (Inventaires diurnes
indirects)
36
Tableau 7: Statistique d'estimation de la
densité de Céphalophe roux à Gribé (Inventaires
diurnes indirects)
38
Tableau 8: Statistique d'estimation de la
densité de Céphalophe roux à Gouonepoun (Inventaires
diurnes indirects)
39
Tableau 9: Statistique d'estimation de la
densité de Céphalophe roux dans au PNBB (Inventaires diurnes
indirects)
41
Tableau 10: IKA moyen des espèces
recensées par site dans la zone d'étude (inventaires diurnes
indirects)
43
Tableau 11: IKA Céphalophe Bleu par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
44
Tableau 12: IKA Céphalophe roux par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
45
Tableau 13: IKA Céphalophe à dos
jaune par signe d'activité et par âge dans les trois sites
d'étude (Inventaires diurnes indirects)
46
Tableau 14: IKA Bongo par signe d'activité
et par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
48
Tableau 15: IKA Buffle par signe d'activité
et par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
49
Tableau 16: IKA petits singes par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
50
Tableau 17: IKA Chat sauvage par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
50
Tableau 18: IKA Chevrotain aquatique par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
51
Tableau 19: IKA Civette par signe d'activité
et par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
51
Tableau 20: IKA Chimpanzé par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
52
Tableau 21: IKA Eléphant par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
52
Tableau 22: IKA Antilope de Bates par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
53
Tableau 23: IKA Gorille par signe d'activité
et par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
54
Tableau 24: IKA Oryctérope par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
55
Tableau 25: IKA Pangolin à longue queue par
signe d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
56
Tableau 26: IKA Pangolin géant par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
57
Tableau 27: IKA Léopard par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
58
Tableau 28: IKA Athérure africain par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
59
Tableau 29: IKA Potamochère par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
60
Tableau 30: IKA Sitatunga par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
62
Tableau 31: IKA Activités humaines par signe
d'activité et par âge dans les sites d'étude (Inventaires
diurnes indirects)
63
Tableau 32: Individus vus physiquement et par site
(inventaires diurnes directs)
64
Tableau 33: Statistique d'estimation de la
densité de groupes de petits singes à Gribé (Inventaires
diurnes directs)
65
Tableau 34: Statistique d'estimation de la
densité de groupes de petits singes à Gouonepoun (Inventaires
diurnes directs)
66
Tableau 35: Statistique d'estimation de la
densité de groupes de petits singes dans le PNBB (Inventaires diurnes
directs)
67
Tableau 36: IKA des espèces dans l'ensemble
de la zone d'étude (Inventaires diurnes directs)
69
Tableau 37: IKA des observations directes des
petits singes entre les trois sites (Inventaires diurnes directs)
70
Tableau 38: IKA des espèces dans l'ensemble
de la zone d'étude (Inventaires nocturnes)
71
Tableau 39: Espèces vues physiquement et par
site (Inventaires nocturnes)
74
Tableau 40: Statistique d'estimation de la
densité de Céphalophe bleu à Gribé (Inventaires
nocturnes)
74
Tableau 41: Statistique d'estimation de la
densité de Céphalophe bleu à Gouonepoun (Inventaires
nocturnes)
75
Tableau 42: Statistique d'estimation de la
densité de Céphalophe bleu dans le PNBB (Inventaires
nocturnes)
76
Tableau 43: Densité des espèces
capturées au moyen des Caméras infra-rouges dans les trois
sites
78
Tableau 44: Indice photographique de la faune (WPI)
des espèces dans les trois sites
81
Tableau 45: Richesse spécifique des grands
et moyens mammifères dans quelques sites du Sud-est Cameroun
84
Tableau 46: Abondance relative (IKA) des grands et
moyens mammifères dans quelques sites du Sud-est Cameroun
85
Tableau 47: Densité des grands et moyens
mammifères dans quelques sites du Sud-est Cameroun
86
Tableau 48: WPI des grands et moyens
mammifères dans quelques sites du Sud-est Cameroun
86
Tableau 49: Vitesse moyenne de décomposition
des crottes de quelques espèces dans la zone d'étude
87
Tableau 50: Quelques ligneux recensés le
long des transects à Gouonepoun et à Gribé
89
Tableau 51: IKA et WPI des espèces
recensées par type de méthode testées dans la zone
91
Tableau 52: Coefficient de variation des
méthodes d'évaluation de l'abondance de la faune
91
Tableau 53: Liste des espèces rares
détectées au moyen des caméras infra-rouges
92
LISTE DES FIGURES
Figure 1: Localisation de la zone d'étude
dans la région et du PNBB
12
Figure 2 : Visée d'angle pour
ouverture des transects
24
Figure 3 : Ouverture des transects
24
Figure 4: Site proposé pour le suivi de la
faune
24
Figure 5: Inventaires nocturnes
26
Figure 6: Programmation des caméras
27
Figure 7: Hauteur de fixation de la
caméra
27
Figure 8: Distribution des caméras
infra-rouges sur les transects dans la zone d'étude
27
Figure 9: BongoTragelaphus
euryceros
28
Figure 10: Potamochère Potamochoerus
porcus
28
Figure 11: IKA des espèces recensées
dans toute la zone d'étude (Inventaires diurnes indirects)
33
Figure 12: Courbe de probabilité de
détection des crottes de Céphalophe bleu dans le site de
Gribé en fonction de la distance perpendiculaire au transect
(Inventaires diurnes indirects)
34
Figure 13: Courbe de probabilité de
détection des crottes de Céphalophe bleu dans le site de
Gouonepoun en fonction de la distance perpendiculaire au transect (Inventaires
diurnes indirects)
36
Figure 14: Courbe de probabilité de
détection des crottes de céphalophe bleu dans le PNBB en
fonction de la distance perpendiculaire au transect (Inventaires diurnes
indirects)
37
Figure 15: Comparaison des densités des
Céphalophe bleus entre les trois sites (Inventaires diurnes
indirects)
37
Figure 16: Courbe de probabilité de
détection des crottes de Céphalophe roux à Gribé en
fonction de la distance perpendiculaire au transect (Inventaires diurnes
indirects)
39
Figure 17: Courbe de probabilité de
détection des crottes de Céphalophe roux à Gouonepoun en
fonction de la distance perpendiculaire au transect (Inventaires diurnes
indirects)
40
Figure 18: Courbe de probabilité de
détection des crottes de Céphalophe roux dans le PNBB en fonction
de la distance perpendiculaire au transect (Inventaires diurnes indirects)
41
Figure 19: Comparaison des densités de
Céphalophe roux entre les trois sites (Inventaires diurnes
indirects)
42
Figure 20: Comparaison des IKA du Céphalophe
bleu entre les trois sites (Inventaires diurnes indirects)
45
Figure 21: Comparaison des IKA du Céphalophe
roux entre les trois sites (Inventaires diurnes indirects)
46
Figure 22: Comparaison des IKA du Céphalophe
à dos jaune entre les trois sites (Inventaires diurnes indirects)
47
Figure 23: Comparaison des IKA de Bongo entre les
trois sites (Inventaires diurnes indirects)
48
Figure 24: Comparaison IKA éléphants
entre les trois sites (Inventaires diurnes indirects)
53
Figure 25: Comparaison des IKA Gorilles entre les
trois sites (Inventaires diurnes indirects)
55
Figure 26: Comparaison des IKA de pangolin à
longue queue entre les trois sites (Inventaires diurnes indirects)
56
Figure 27: Comparaison des IKA de pangolin
géant entre les trois sites (Inventaires diurnes indirects)
58
Figure 28: Comparaison des IKA de l'Athérure
africain entre les trois sites (Inventaires diurnes indirects)
60
Figure 29: Comparaison des IKA de
potamochère entre les trois sites (Inventaires diurnes indirects)
61
Figure 30: Comparaison des IKA des grands et moyens
mammifères entre les trois sites (Inventaires diurnes indirects)
62
Figure 31: Comparaison des IKA Activités
humaines entre les trois sites (Inventaires diurnes indirects)
64
Figure 32: Courbe de probabilité de
détection des petits singes à Gribé en fonction de la
distance perpendiculaire au transect (Inventaires diurnes directs)
65
Figure 33: Courbe de probabilité de
détection des petits singes à Gouonepoun en fonction de la
distance perpendiculaire au transect (Inventaires diurnes directs)
67
Figure 34: Courbe de probabilité de
détection des petits singes dans le PNBB en fonction de la distance
perpendiculaire au transect (Inventaires diurnes directs)
68
Figure 35: Comparaison des densités des
groupes de petits singes entre les trois sites (Inventaires diurnes
directs)
68
Figure 36: Comparaison des IKA de Petits singes
entre les trois sites (inventaires diurnes directs)
70
Figure 37: Comparaison IKA global des grands et
moyens mammifères entre les trois sites (Inventaires diurnes
directs)
71
Figure 38: Comparaison des IKA des
céphalophes bleus entre les trois sites (Inventaires nocturnes)
72
Figure 39: Comparaison des IKA de daman des arbres
entre les trois sites (Inventaires nocturnes)
73
Figure 40: Comparaison IKA global des grands et
moyens mammifères entre les trois sites (Inventaires nocturnes)
73
Figure 41: Courbe de probabilité de
détection de Céphalophe bleu à Gribé en fonction de
la distance perpendiculaire au transect (Inventaires nocturnes)
75
Figure 42: Courbe de probabilité de
détection de Céphalophe bleu à Gouonepoun en fonction de
la distance perpendiculaire au transect (inventaires nocturnes)
76
Figure 43: Courbe de probabilité de
détection de Céphalophe bleu dans le PNBB en fonction de la
distance perpendiculaire au transect (inventaires nocturnes)
77
Figure 44: Comparaison des densités de
Céphalophes bleus entre les trois sites (inventaires nocturnes)
77
Figure 45: Comparaison des densités de
Céphalophe bleu entre les trois sites (Caméra infra-rouge)
79
Figure 46: Comparaison des densités de
Céphalophe roux entre les trois sites (Caméra infra-rouge)
79
Figure 47: Comparaison des densités de
Petits singes entre les trois sites (Caméra infra-rouge)
80
Figure 48: WPI moyen des espèces
capturées dans les trois sites (Caméra infra-rouge)
81
Figure 49: Comparaison des WPI de Céphalophe
bleu entre les trois sites (Caméra infra-rouge)
82
Figure 50: Comparaison des WPI de Céphalophe
roux entre les trois sites (Caméra infra-rouge)
83
Figure 51: Comparaison des WPI de Petits singes
entre les trois sites (Caméra infra-rouge)
83
Figure 52: Comparaison de WPI de tous les
mammifères entre les trois sites (Caméra infra-rouge)
84
Figure 53: Crotte fraîche (0 jours)
88
Figure 54: Crotte récente (5ème
jour)
88
Figure 55: Crotte vieille (10ème
jour)
88
Figure 56: Crotte très vieille
(15ème jour)
88
Figure 57: Crotte disparue
88
LISTE DES ANNEXES
Annexe 1: Fiche de collecte des données
d'inventaire de la faune
105
Annexe 2: Fiche d'identification des crottes des
animaux le long des transects
106
Annexe 3: Fiche d'inventaire des caméras
infra-rouges le long des transects
107
Annexe 4: fiche de suivi des différents
stades de décomposition des crottes des céphalophes
108
Annexe 5: Fiche de caractérisation de la
végétation de la faune dans la zone d'étude
109
Annexe 6: Liste des espèces de grands et
moyens mammifères recensées dans la zone
110
Annexe 7: Quelques espèces rares
photographiées par les Caméras infra-rouges dans la zone
d'étude
111
LISTE DES ACRONYMES ET
ABREVIATIONS
|
AAPPEC
|
Association pour l'Auto-Promotion des Populations de l'Est
Cameroun
|
|
CITES
|
Convention on International Trade on Endangered Species of
Flora
and Fauna
|
|
DEPFOR
|
Département de Foresterie
|
|
DSCN
|
Direction de la Statistique et de la Comptabilité
Nationale
|
|
FASA
|
Faculté d'Agronomie et des Sciences Agricoles
|
|
GIC
|
Groupe d'Initiative Commune
|
|
GPS
|
Global Positioning System.
|
|
GTZ
|
Gesellschaft für Technische Zusammenrbeit
|
|
FOSAS
|
Forest Savana Sustainability
|
|
IKA
|
Indice Kilométrique d`Abondance
|
|
IUCN
|
Union Mondiale pour la Conservation de la Nature
|
|
JICA
|
Agence Internationale de Coopération Japonaise
|
|
MINEF
|
Ministère de l'Environnement et des Forêts
|
|
MINFOF
|
Ministère des Forêts et de la Faune
|
|
MINMEE
|
Ministère des Mines, de l'Eau et de l'Énergie
|
|
PNBB
|
Parc National de Boumba-Bek
|
|
Pro/PSFE/GTZ
|
Programme d'Appui au Programme Sectoriel Foret Environnement
à travers la Deutsche Gesellschaft fur Technische Zusammenarbeit
|
|
SIBAF
|
Société Industrielle des Bois Africains
|
|
SIM
|
Société Industrielle de Mbang
|
|
STBK
|
Société de Transformation des Bois de la
Kadei
|
|
UFA
|
Unité Forestière d'Aménagement
|
|
UDS
|
Université de Dschang
|
|
UTO
|
Unité Technique Opérationnel
|
|
WCS
|
Wildlife Conservation Society
|
|
WWF
|
World Wide Funds for Nature
|
|
ZIC
|
Zone d'Intérêt Cynégétique
|
|
ZICGC
|
Zone d'Intérêt Cynégétique à
Gestion Communautaire
|
|
|
RESUME
L'inventaire de la faune sauvage sur les transects permanents
en périphérie Nord-est et dans le Parc National de Boumba-Bek
(PNBB) s'est déroulé du 22 juillet au 21 janvier 2014. Il
s'agissait de comparer les densités et/ou l'indice de présence
des espèces de grands et moyens mammifères à Gribé,
à Gouonepoun et dans le Parc, d'évaluer la performance des
méthodes d'inventaires testées, de détecter les
espèces rares et de formuler quelques suggestions afin d'orienter le
modèle de gestion de la ressource faune. Pour estimer les
densités et l'abondance relative de chaque espèce, les
inventaires diurnes directs, indirects, nocturnes et les caméras
infra-rouges ont été réalisés.
Vingt-huit espèces de grands et moyens
mammifères ont été identifiées sur un ensemble de
18 transects pour une distance totale de 108 km dans la zone d'étude.
Les inventaires diurnes indirects ont donné un total de 4529 signes
d'activité soit un Indice Kilométrique d'Abondance (IKA) de
41,935, avec une différence significative entre les sites pour le
céphalophe à dos jaune, le buffle, le pangolin à longue
queue et le pangolin géant. Seules les densités des
céphalophes bleus et céphalophes roux ont pu être
estimées par le Programme DISTANCE 6.0, à Gribé (2,915
céphalophes bleus/km2 et 1,364 céphalophes
roux/km²), à Gouonepoun (1,678 céphalophes
bleus/km2 et 0,745 céphalophes roux/km²) et dans le PNBB
(0,634 céphalophes/km2 et 0,837 céphalophes
roux/km²). Aucune différence significative n'a été
observée entre les trois sites. Pour ce qui était des inventaires
diurnes directs, seules les densités des petits singes ont pu être
calculées par le Programme DISTANCE 6.0, à Gribé (6,027
groupes de petits singes /km²), à Gouonepoun (3,848 groupes de
petits singes/km²) et dans le PNBB (8,500 groupes de petits
singes/km²). Un total de 337 signes (vu/entendu) a été
enregistré soit un IKA de 3,5 avec aucune différence
significative observée entre les sites.
Un total de 1105 signes (vu/entendu) ont été
enregistrés lors des inventaires nocturnes (IKA = 10,32). Ainsi, seules
les densités de céphalophes bleus ont pu être
estimées par le Programme DISTANCE 6.0, notamment à Gribé
(34,799 céphalophes bleus/km²), à Gouonepoun (91,328
céphalophes bleus /km²) et dans le PNBB (78,423 céphalophes
bleus/km²). De la comparaison des moyennes des densités et des IKA,
aucune différence significative n'a été observée
entre les trois sites. Douze caméras ont été fixées
dans chaque site, soit un total de 578 captures de toutes les espèces de
faune dans la zone d'étude, dont 156 captures à Gribé
(Densité= 19,713#177;35,653 individus/km²), de 231 captures
à Gouonepoun (Densité = 27,017#177;34,528 individus/km²) et
de 191 captures dans le PNBB (Densité = 20,813#177;31,976
individus/km²). L'effort total de capture était de 1080
caméra-jours. Le Wildlife Picture Index (WPI) a été
estimée pour toutes les espèces de la faune. Le calcul des
densités et de WPI des différentes espèces n'ont
présenté aucune différence significative entre les sites.
La vitesse de décomposition de crottes des cinq espèces
rencontrées a été suivie jusqu'à disparition
complète et quelques ligneux ont été inventoriés le
long des transects à Gribé et à Gouonepoun.
L'évaluation des méthodes d'inventaires testés ont
révélé que, la meilleure méthode d'inventaire sur
transect linéaire dans la zone d'étude est celle des inventaires
diurnes indirects. Les espèces rares ou en voie de disparition
capturée par les caméras infra-rouges appartenaient à
quatre familles avec huit espèces. Trois axes ont été
développés afin d'orienter le modèle de gestion de la
ressource faune sauvage.
Mots clés: Caméra infra-rouge;
Inventaire de la faune; Parc National de Bounba-Bek; Performance des
méthodes; Transects permanents; Wildlife Picture Index.
ABSTRACT
The inventory of wildlife on permanent transect in the North
east Periphery Zone of the Boumba-Bek National Park (BBNP) was carried out on
July 22, 2013 to January 21, 2014 in the park and the surrounding villages of
Gribé and Gouonepoun. The study had the following objectives that is, to
compare the density and the abundance of large and medium-sized mammals within
these sites, to evaluate the performance of the survey methods used, to detect
the rare species and to provide some suggestions to guide the management model
of fauna. To estimate the densities and the abundance of each species, direct
diurnal, indirect diurnal, nocturnal surveys, and camera trapping were used.
Twenty eight large and medium-sized mammal species were
identified on 18 permanent transects. A distance of 108 kilometers was
effectively covered. The indirect diurnal surveys gave 4492 signs of presence
with Mean Encounter Rates (MER) of 41,935, with a significant difference among
the sites for the yellow back duiker, the buffalo, long tail pangolin and the
giant pangolin. Only the densities of the blue duikers and russet-red duikers
were estimated by the DISTANCE Program 6.0, in Gribé (2,915 blue
duikers/km² and 1,364 russet-red duikers/km²), in Gouonepoun (1,678
duikers/km² and 0,745 russet-red duikers/km²) and in the BBNP (0,634
duikers/km² and 0,837 russet-red duikers/km²). There is no
significant difference observed among the three sites. Only the densities of
the small monkeys for the direct diurnal survey were estimated by the DISTANCE
Program 6.0, in Gribé (6,027 groups of small monkeys/km²), in
Gouonepoun (3,848 groups of small monkeys/km²) and in the BBNP (8,500
groups of small monkeys/km²). 337 signs were recorded, MER of 3,5 per
transect, there is no significant difference observed among the sites.
A total of 1105 signs were recorded during the nocturnal
survey (MER = 10,32), for this survey, only blues duikers densities was
estimated by DISTANCE program 6.0, in Gribé (34,799 blues
duikers/km²), in Gouonepoun (91,328 blues duikers /km²) and in the
BBNP (78,423 blues duikers/km²). There is no significant difference among
the three sites. Twelve cameras trappings were fixed per site, where 578
captures were realized, with 156 captures in Gribé (Densities =
19,713#177;35,653 individual's/km²), 231 captures in Gouonepoun (Densities
= 27,017#177;34,528 individual's/km²) and 191 captures in BBNP (Densities
= 20,813#177;31,976 individual's/km²). The total effort of capture was
1080 camera-days. The Wildlife Picture Index (WPI) was estimated for all
species. There is no significant difference in densities and WPI estimated. The
dung decay's rate is estimated for five species. The habitat of fauna on
transects in Gribé and Gouonepoun was characterized. The evaluation of
the surveys methods tested, revealed that, the best method of linear transect
in the study zone was the indirect diurnal surveys. The rare species that were
captured by the cameras trapping belong to 4 families with 8 species. Three
axes were developed in order to guide management model of the wildlife
resources in the study area.
Keywords: Boumba-Bek National Park; Camera
trapping; Performance of surveys methods; Permanent transects; Wildlife Picture
Index; Wildlife survey.
CHAPITRE 1: INTRODUCTION
1.1
Contexte
Le suivi est la clé à la conservation. Il est
nécessaire de disposer des techniques les plus appropriées pour
mieux connaître les richesses faunistiques d'un site, leur distribution
et leur évolution dans le temps. Ainsi, le dénombrement de la
faune contribue activement au maintien de l'intégrité des
écosystèmes et de la biodiversité (Poilecot, 2009).
Cependant, la nécessité dans le cadre d'une
gestion rationnelle de bien connaître la situation actuelle et les
tendances évolutives de la ressource animale est donc importante dans le
développement des stratégies de conservation de la
biodiversité (Hacker et al., 1998; Halford, 2003).
Historiquement, les biologistes de la conservation
s'intéressaient majoritairement à des espèces que l'on
pourrait qualifié d'emblématiques. Ces espèces
étaient majoritairement des oiseaux ou de grands mammifères dont
l'écologie est bien connue mais aussi aisés à suivre
techniquement (forte détectabilité, capture et marquage
relativement simple, ...).
Au vue de l'importance de cette diversité biologique,
le Cameroun accorde une place importante à la gestion durable des
écosystèmes forestiers et de toutes les composantes qui y vivent.
Ceci s'observe notamment à travers l'arrêté N°
0222/MINFOF du 02 mai 2006 fixant les normes d'inventaire des espèces
fauniques en zone de forêt et l'arrêté N° 0244/MINFOF
du 02 mai 2006 fixant les normes d'inventaire des espèces fauniques en
zone de savane. Ces textes préconisent entre autres l'élaboration
des plans d'aménagement des aires protégées et mettent un
accent particulier sur le suivi de la faune qui passe par l'évolution
des densités des populations animales suivant différentes
méthodes. Le Cameroun possède aujourd'hui des aires
protégées qui comptent parmi les plus vastes d'Afrique. Ces aires
protégées abritent des communautés biotiques, animales et
végétales, ayant un endémisme assez important (UICN,
1992).
1.2
Problématique
Le suivi de la faune sauvage est fondamental pour la
conservation de la biodiversité. Il peut être utilisé pour
estimer la durabilité de la chasse et l'efficacité de la
conservation (Waltert et al., 2006). Les mêmes auteurs affirment
que, pour la gestion adaptative de la faune sauvage et la mesure de
l'efficacité de la conservation, le suivi a besoin d'être à
la fois fiable et efficace. Ce suivi implique une disponibilité de
moyens financiers, humains et matériels, donc des arbitrages pouvant
conduire à concentrer les efforts sur les habitats et les espèces
les plus pertinentes (Besnard, 2010).
Pour la réalisation des travaux d'inventaires
fauniques, plusieurs méthodes sont utilisées notamment, les
inventaires diurnes et nocturnes basées sur la méthodologie des
transects linéaires (White et Edward, 2000), les captures au filet telle
que décrite par Wilson (2005) et l'utilisation des caméras
infra-rouges (Rowcliffe et al., 2011; Nkwetaketu, 2011) La
méthode de recensement la plus efficace pour les grandes zones de
forêts est celle de transects linéaires (Barnes et Jensen,
1987 ; Buckland et al., 1993 ; Tutin et Fernandez,
1984 ; White et Edward, 2000), car les analyses par transect
linéaire utilisent la distribution des distances perpendiculaires le
long des transects pour calculer une probabilité de détection
(Buckland et al.,1993). Une évaluation de l'efficacité
de différentes méthodes d'inventaire faunique basée sur
l'analyse de l'estimation du coefficient de variation avait été
réalisée par (Viquerat et al., 2012) dans le Parc
National de Korup car le test statistique Z des paramètres de
populations recommandé par Buckland et al.,. (2001) est
insensible au changement de l'estimation associée au coefficient de
variation (Plumptre, 2000). Selon Viquerat et al., (2012), sur les
trois méthodes testées sur les transects linéaires
à savoir les inventaires directs diurnes, nocturnes et la méthode
indirecte basée sur les crottes, la meilleure des méthodes est
celle des inventaires diurnes directs. La méthode indirecte sur
transects linéaires reste cependant très utilisée en zone
de forêt (Burnham et al., 1980; Buckland et al., 1993;
2001; White et Edwards 2000). Par ailleurs, des inventaires nocturnes peuvent
s'avérer nécessaires pour estimer plus précisément
l'abondance de certaines espèces dont les céphalophes et la
plupart des petits carnivores. (Larrubia et Arnhem 2009).
La méthode d'inventaire basée sur les
caméras infra-rouges est très peu utilisée en zone
tropicale (ex : Nkwetaketu, 2011; Silveira et al., 2003). Il en
est de même de la méthode d'inventaire au moyen des captures au
filet (ex : Kostner et Hart 1988; Hart, 2000; Djetkeu, 2010). La question
mise en relief dans la présente étude est celle de savoir la
méthode adéquate d'estimation de l'abondance et/ou de la richesse
spécifique de la faune sauvage autour du Parc National de Boumba-Bek
(PNBB), notamment dans les ZICGC 13 et 14.
1.3
Objectifs
1.3.1
Objectif Global
L'objectif global de la présente étude est de
contribuer à la meilleure connaissance du potentiel faunique par la mise
à disposition des gestionnaires une méthode adéquate
d'estimation de l'abondance et/ou de la richesse spécifique de la faune
sauvage au Nord du PNBB.
1.3.2
Objectifs spécifiques
Plus spécifiquement, l'étude se propose de :
- Comparer les densités et/ou les indices de
présence des espèces de grands et moyens mammifères
rencontrées suivant quatre méthodes d'inventaire faunique dans
trois sites ;
- Evaluer la performance des quatre méthodes
d'inventaire faunique testées;
- Détecter les espèces rares;
- Formuler des suggestions afin d'orienter le modèle de
gestion durable de la ressource faune sauvage en cours de construction dans la
zone d'étude.
1.4
Importance de l'étude
La présente étude va contribuer à mieux
connaître les contours de la gestion des ressources fauniques tant sur le
plan théorique que pratique.
ü Sur le plan théorique, les résultats de
cette étude constitueront une importante base de données pour
initier d'autres travaux de recherche par les scientifiques
(universités, Instituts de recherche etc.) et certains organismes de
conservation.
ü Sur le plan pratique, la connaissance des
densités ou de l'abondance relative et la méthode adéquate
sont des éléments pertinents de gestion. Les résultats de
cette étude vont efficacement contribuer à l'élaboration
des plans de gestion spécifiques, notamment le modèle de gestion
de la ressource faune. Les services de conservation et les guides de chasse
pourront utiliser ces informations dans leur planification stratégique.
1.5
Limites de l'étude
La principale limite de l'étude est la réticence
des pisteurs vis-à-vis des inventaires nocturnes, occasionnant parfois
des interruptions des travaux.
CHAPITRE 2: REVUE DE LA LITTERATURE
2.1
Clarification des concepts
La gestion durable des ressources fauniques en
général et le suivi de la faune sauvage en particulier
comprennent de nombreux concepts dont certains sont importants à
clarifier dans le cadre de la présente étude pour mieux
appréhender le sujet traité.
2.1.1
Aire protégée
Selon l'Union Mondiale pour la Nature, une aire
protégée est une portion de terre et/ou de mer vouée
spécialement à la protection et au maintien de la
diversité biologique, ainsi que des ressources naturelles et culturelles
associées, et gérée par des moyens efficaces, juridiques
ou autres (UICN, 1994). D'après la législation camerounaise il
s'agit d'une zone géographique délimitée et
gérée en vue d'atteindre des objectifs spécifiques de
conservation et de développement durable d'une ou plusieurs ressources
données (Décret n 95-466-PM-du 20 juillet 1995). D'après
les dispositions légales les aires protégées au Cameroun
sont regroupées en Unités techniques opérationnelles (UTO)
et reparties suivant trois catégories en fonction de la superficie.
- catégorie I : superficie supérieure à
100 000 ha
- catégorie II : superficie comprise entre 50 000 ha et
100 000 ha
- catégorie III : superficie inférieure à
50 000 ha
L'UTO peut dans une certaine mesure contenir plusieurs aires
protégées selon les cas. Cependant la loi N° 94/01 du 20
janvier 1994 définit comme étant des aires
protégées de faune:
- les parcs nationaux
- les réserves de faune
- les zones d'intérêt
cynégétique
- les jardins zoologiques
- les sanctuaires de faune
- les zones tampons
2.1.2
Diversité biologique
Au sommet de Rio de Janeiro en 1992, la diversité
biologique a été définie comme étant la
variabilité des organismes vivants de toute origine y compris les
écosystèmes terrestres, marins et les écosystèmes
aquatiques et les complexes écologiques dont ils font partie. La notion
de diversité est liée à ce que les naturalistes appellent
subjectivement la richesse d'un peuplement (Ramade, 1993). La diversité
peut être divisée en trois catégories hiérarchiques : gène, espèce et
écosystème
2.1.3 Espèces rares
Ce sont des espèces avec une faible population. La
rareté peut être naturelle ou résultant de la pression
humaine (Simberloff, 1998).
2.1.4
Grands et moyens mammifères
Les mammifères regroupent la classe des
vertébrés la plus évoluée. Les glandes mammaires
qui caractérisent leurs femelles secrètent du lait pour nourrir
les jeunes. Leur coeur est divisé en quatre cavités et
possèdent un encéphale volumineux, leur corps est le plus souvent
couvert de poils avec des organes de sens sont très
développés. Les grands et moyens mammifères, sont les
espèces de mammifères qui vont de la taille de l'Athérure
à celle de l'Eléphant. (White et Edwards, 2000). Les
ongulés des forêts (mammifères pourvus de sabots) se
divisent en deux catégories: les espèces de petite taille (de 5
à 70 kg) notamment les céphalophes, le chevrotain et les guibs
harnachés qui vivent dans des aires stables et bien définies et
les espèces de grande taille (plus de 100 kg), telles que
potamochères, hylochères, bongos, okapis, buffles,
rhinocéros et éléphants qui sont des espèces
souvent très mobiles, voire migratrices. Chez les espèces de
grande taille les densités de population sont moins
élevées que chez les espèces de grandes tailles par
exemple chez les céphalophes. Les grands mammifères sont ceux qui
se reproduisent le plus lentement (White et Edwards, 2000).
2.1.5
Parc National
Le décret 95/466 du 20 juillet 1995 en son article 2,
alinéa 8 définie un parc national comme étant : un
périmètre d'un seul tenant, dont la conservation de la faune, de
la flore, du sol, du sous-sol, de l'atmosphère, des eaux, et en
général, du milieu naturel, présente un
intérêt spécial qu'il importe de préserver contre
tout effort de dégradation naturelle, et de soustraire à toute
intervention susceptible d'en altérer l'aspect, la composition et
l'évolution.
Sont prises en considération à ce titre :
- la préservation d'espèces animales ou
végétales et d'habitats en voie de disparition sur tout ou partie
du territoire national;
- la préservation ou la constitution d'étapes
sur les grandes voies de migrations de la faune sauvage;
- les études scientifiques ou techniques indispensables
au développement des connaissances humaines.
Les activités suivantes sont interdites :
- la chasse et la pêche, sauf dans le cadre d'un
aménagement;
- les activités industrielles;
- l'extraction des matériaux;
- les pollutions de toute nature;
- les activités agricoles, pastorales et
forestières;
- la divagation des animaux domestiques ;
- le survol par aéronefs à une altitude
inférieure à 200 m;
- l'introduction d'espèces zoologiques ou botaniques
indigènes ou importées, sauf dans un but scientifique ou dans le
cadre d'opérations d'aménagement autorisées par le
ministre chargé de la Faune.
2.1.6
Transect
C'est une piste ouverte en forêt couvrant plusieurs
kilomètres de long et de largeur variable à partir de laquelle
les animaux ou les indices de leur présence sont vus et comptés
(White et Edwards, 2000). Les transects permanents sont des lignes bien
entretenues pendant une période de temps bien déterminée,
sur lesquels les recensements de la faune sont effectués de façon
régulière (Whitesides, 1988). Ainsi la méthode la plus
efficace pour compter les grands et moyens mammifères en zone de
forêt est celle des transects linéaires (Tutin et Fernandez,
1984)
2.1.7
Zone périphérique
Zone créée autour de l'aire
protégée (3 km sur la périphérie) dont l'objectif
est d'apporter un appui à la conservation et au développement,
face aux villages qui dépendent du parc pour satisfaire leurs besoins
quotidiens (MINEF, 2002).
2.2
Généralités sur quelques techniques d'inventaire de la
faune en zone de foret
2.2.1
Généralités
Les recensements de mammifères fournissent aux
gestionnaires des données de trois types.
- Au niveau le plus élémentaire, ils permettent
de déterminer la présence ou l'absence des espèces dans
différents sites en vue de l'élaboration des cartes de leur aire
de répartition.
- À un niveau plus détaillé, un simple
échantillonnage permet de déterminer l'abondance relative d'une
espèce dans différents sites ou dans un site donné sur une
période prolongée.
- Au troisième et dernier niveau de complexité,
des travaux d'échantillonnage bien plus rigoureux, une collecte plus
systématique de données et de robustes analyses statistiques
permettent parfois d'élaborer des estimations quantitatives de la
densité des populations (Davies et al., 2001).
2.2.2
Quelques méthodes d'inventaire de la faune en zone de forêt.
2.2.2.1 Cris d'appel des chasseurs, attractifs et postes
d'observation
Les chasseurs ont toute une gamme de grognements, de cris et
de sifflements pour attirer différentes espèces. Le meilleur
exemple en est le bêlement nasillard qui permet d'attirer les
céphalophes. (par exemple Wilson, 1990). Les appels doivent être
répétés à des distances d'au moins 250 m. Les
chercheurs doivent se dissimuler soigneusement, par exemple entre les
contreforts d'un arbre, et rester silencieux et immobiles pendant que les
chasseurs appellent. Cette méthode livre de précieux
enseignements sur la présence des espèces dans un habitat
donné.
2.2.2.2 Recensement au moyen des attractifs
On peut aussi attirer les animaux par
d'autres méthodes: des pierres à sel, naturelles ou
artificielles, dans le cas des herbivores, et de la viande ou des marques
odorantes pour les carnivores. Il faut parfois plusieurs semaines avant que les
animaux ne découvrent un attractif, et cette méthode ne se
prête donc pas aux enquêtes ponctuelles de courte durée.
Mais une des particularités de cette méthode est d'étudier
le comportement des animaux.
2.2.2.3 Postes d'observation
Les points d'observation stratégiques comprennent les
affleurements salins, les trous d'eau, les bourbiers, les arbres chargés
de fruits, les trouées dues à la chute des arbres et
occupées par des pousses fraîches, les clairières, les
routes d'exploitation forestière et les pistes
régulièrement empruntées par les animaux. Une cache simple
peut être aménagée en suspendant du filet de camouflage
entre les arbres ou leurs contreforts, ou en coupant des palmes pour former un
écran derrière lequel on peut se dissimuler tout en observant
l'extérieur (Wilson, 1990). Cette méthode permet de
déterminer le potentiel faunique dans une zone afin de mettre en place
un tourisme d'observation.
2.2.2.4 Transects en forêt
Le transect est par définition une piste ouverte
couvrant plusieurs kilomètres de long à partir de laquelle les
animaux ou leurs indices de présence sont vus et comptés (White
et Edwards, 2000). La méthode de recensement la plus efficace pour les
grandes zones de forêt pluviale est celle des transects linéaires
(Tutin et Fernandez, 1984).
2.2.2.5 Pièges photographiques
La caméra infra-rouge est un appareil qui est
déclenché suite à un mouvement focalisé dans le
champ de vision de la caméra (Chapman, 1927, cité par Rowcliffe
et Carbone, 2008), cités par Nkwetaketu (2011). La photo est
extrêmement précieuse pour enregistrer les espèces
difficiles à dépister. La pose d'appareils à
déclenchement automatique permet de surveiller avec peu de personnel les
sites fréquemment visités par les animaux (par exemple, les
affleurements de sel), les sites appâtés et leurs lieux de passage
habituels. Les pièges photo permettent aussi de déterminer les
schémas d'activité (diurne, nocturne ou crépusculaire),
les réactions aux perturbations (par exemple Griffiths, 1994), les
déplacements, les comportements de reproduction saisonniers et les
structures sociales. Si l'on a assez d'appareils, on peut aussi réunir
des informations sur l'abondance des espèces.
2.2.2.5.1 Quelques
caractéristiques techniques de Bushnell Trophy Cam
Plusieurs marques de caméras infra-rouges sont
utilisées pour la détection des animaux de la faune. L'emploi
d'une caméra dépend des objectifs visés et des moyens
financiers pour une acquisition d'une telle. On rencontre entre autres, Reconyx
HC500, Scoutguard SG550, Moultrie I65 et bien d'autres. Le tableau 1
présente les caractéristiques de Caméra Bushnell Trophy
Cam.
Tableau 1: Caractéristiques des caméras
Bushnell Trophy Cam
|
Eléments photographiques
|
Caractéristiques
|
|
Capteur d'images
|
5 Mégapixels couleur CMOS
|
|
Résolution Maximale
|
3264x2448 (8 MP)
|
|
Objectif
|
Focale = 3,1, Champ de Vision = 50°
|
|
Portée maximale du flash infrarouge
|
15 mètres
|
|
Taille des photos
|
8 MP = 3264x2448; 5 MP = 2560x1920; 3 MP =
2048x1536
|
|
Taille des vidéos
|
720x480/30 fps, 640x480/30 fps, 320x240/30 fps
|
|
Sensibilité de
l'infrarouge passif
|
Capteur Infrarouge Passif (PIR) à 3 niveaux de
sensibilité : Élevée / Normale / Faible
|
|
Fonctionnement
|
Jour/Nuit
|
|
Temps de réponse
|
1s
|
|
Intervalle de déclenchement
|
Programmable 1s - 60 mn
|
|
Nombres de prises
de vue
|
Programmable 1- 3
|
|
Alimentation
|
8xAA recommandées, 4xAA alimentation d'urgence
|
|
Interface
|
Sortie TV (NTSC/PAL) ; USB ; emplacement pour carte
mémoire ; prise pour alimentation externe 6V
|
|
Température de
fonctionnement
|
De -20°C à +60°C
(température de stockage : de -30°C à
+70°C)
|
Source: Meek et al. (2012).
2.3
Théorie des transects linéaires
Les inventaires de faune dans les zones forestières se
font à travers la méthode des transects linéaires à
largeur variable. Cette méthode est la meilleure pour l'estimation des
potentialités fauniques dans les zones de forêts tropicales
humides (Buckland et al., 1993). Elle permet d'identifier les
principales tendances concernant l'abondance, la distribution et les mouvements
saisonniers des espèces clés en relation avec la présence
et les implantations humaines, d'évaluer l'efficacité de la
protection, d'identifier les zones concernées par les problèmes
de protection et de fournir la base de suivi des changements écologiques
et des activités humaines (White et Edwards, 2000). Relativement facile
à mettre en place, cette méthode fournit des résultats
fiables et représentatifs de la réalité (Tutin et
Fernandez, 1984). Cette méthode est largement utilisée en Afrique
Centrale, et plus particulièrement au Cameroun où elle a
d'ailleurs fait l'objet d'un arrêté ministériel fixant les
normes d'inventaires des espèces fauniques en zone forestière du
pays, connu sous l'arrêté 0244/MINFOF du 23 mai 2006. Elle a
été ainsi utilisée par exemple au Parc National de Korup
(Ekobo, 2006; Djetkeu, 2010) et à Ebo (Ekobo, 2006), dans les Parc
Nationaux de Lobéké, Boumba-Bek et Nki (Ekobo, 1998 ;
Nzooh-Dongmo, 2005, Nzooh-Dongmo, 2006 cités par Adama, 2010) et
même dans certaines UFA du Cameroun (Etoga, 2002 ; Nzooh-Dongmo et
al., 2004 cités par Adama, 2010.; Bobo et al, 2006a; Bobo,
2009a). Le comptage par transects linéaires est fondé sur
l'hypothèse selon laquelle la probabilité de détecter un
animal, un nid ou une crotte décroît au fur et à mesure que
de l'axe de déplacement. Plus simplement, vous avez plus de chance de
voir un tas de crottins d'éléphant à vos pieds qu'un autre
situé à 12 m de votre ligne de transects, caché dans une
végétation luxuriante. La probabilité de voir des objets
à des distances différentes dépend de la densité de
la végétation. Sur une piste d'exploitation dépourvue de
végétation, vous pouvez voir un éléphant à
300 m, tandis que dans une forêt à Marantacées dense, vous
pouvez passer à 5 m d'un éléphant au repos sans le voir
(White et Edwards, 2000). C'est dire que l'inventaire faunique en milieu
fermé, comme les forêts, oblige une attention toute
particulière à cause du champ visuel très limité
par la densité de la végétation, soit au sol pour les
mammifères terrestres, soit en hauteur pour les espèces
arboricoles.
2.4
Principe des transects linéaires
La pratique consiste, pour un observateur, à
évoluer sur une ligne de longueur déterminée (li) de
manière aléatoire et noter les contacts avec les animaux en
mesurant (ou estimant), avec le plus de précision possible (Burnham
et al., 1980 ; Buckland et al., 1993).
- la distance (ri) qui le sépare de l'animal,
- l'angle (ai) formé entre la ligne de marche et la
ligne imaginaire reliant l'observateur à l'animal,
- la distance orthogonale (xi) de l'animal à l'axe du
transect qui est mesurée, puisqu'il existe une relation
trigonométrique entre les trois mesures : xi = ri x sin (ai).
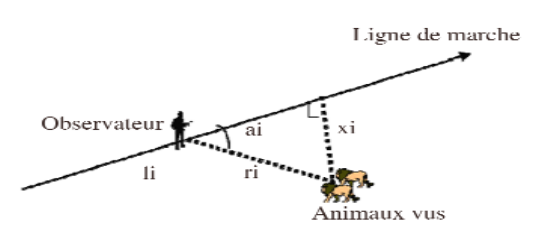
Légende :
xi = distance perpendiculaire animal - ligne de
transect
ri = distance observateur - animal
ai = angle d'observation de l'animal à
partir de la ligne de marche
li = distance parcourue par l'observateur sur
le transect
L'estimation de la densité se résume ainsi
à :
D = n/A = n/2La
D = densité
n = nombre d'objets (animaux) observés
A = surface échantillonnée
L = longueur du transect
a = largeur de bande d'observation d'un côté du
transect
2.5
Détermination des densités des grands et moyens mammifères
à partir des crottes
D'après White et Edwards (2000), pour estimer la
densité animale grâce aux crottes, il faut connaître :
- La densité des crottes (nombre de
tas/km2);
- Le taux de défécation ou taux de
création (nombre de tas de crottes /jour)
- La vitesse de dégradation ou disparition des crottes
(nombre de jours/tas)
Pour déterminer la densité d'animaux
(animaux/km²), on divise la densité de crottes (nombre de tas
/km²) par le produit de la durée moyenne de disparition (nombre
moyen de jours nécessaires pour que la crotte disparaisse après
sa fabrication) et par taux de création (nombre de crottes produites par
un animal en un jour).
Densité d'animaux = Densité de
crottes / (durée moyenne de disparition des crottes x taux de
défécation).
2.6
Vitesse de dégradation ou disparition des crottes des grands et moyens
mammifères.
Lorsque l'on utilise la méthode des transects
linéaires pour estimer les densités, une des hypothèses
cruciales est que tous les objets situés sur la ligne médiane
doivent être détectés avec certitude. Ainsi, lors de
recensement des crottes, la considération de la vitesse de
dégradation s'avère nécessaire pour calculer la
densité animale. (White et Edwards, 2000). La dégradation des
crottes est un processus complexe. Elle est fonction de la diminution de la
disponibilité en fruits, de l'activité des insectes, de
l'humidité, de la pluviométrie et de la saison sèche (Van
Vliet et Nasi, 2008, cités par Djetkeu, 2010).
La disponibilité en fruit affecte la consistance de la
crotte et par conséquent, la vitesse de décomposition de la
crotte. De nombreux insectes (bousiers et termites) sont des
décomposeurs importants des crottes, mais ont des rythmes
d'activité saisonniers. Les crottes déposées sur les
berges des rivières ou dans les ruisseaux peuvent être
emportées par la pluie, mais celles qui sont sur un sol
marécageux peuvent sembler fraîches pendant de longues
périodes. Celles qui sont exposées au soleil peuvent
sécher plus rapidement et garder leur forme pendant longtemps.
Un autre facteur peut jouer sur la longévité des
crottes, il s'agit du régime alimentaire, car celui-ci joue sur la
consistance de la crotte (Koster et Hart, 1988; Wiles, 1980). Ainsi, dans tout
échantillon de crottes suivi pour déterminer la vitesse de
disparition, il y aura un certain degré de variabilité. Certains
tas de crottes disparaîtront rapidement, d'autres dureront des mois.
Selon Barnes et al. (1994),pour un bon suivi de la
dégradation des crottes, tout chercheur doit éviter les sentiers
et pistes, mais plutôt favoriser le suivi de crottes trouvées dans
la végétation représentative de la zone recensée.
Cela est facile sur des transects linéaires permanents pouvant
être parcourus plusieurs fois à une ou deux semaines d'intervalle
(Tchamba, 1992).
2.7
Synthèse de quelques études sur le potentiel faunique en
périphérie Nord du PNBB
Pour améliorer la connaissance sur la faune sauvage, il
est indispensable d'estimer la richesse spécifique des grands et moyens
mammifères. C'est ainsi, plusieurs études ont été
faites au Sud -est du Cameroun et dont les résultats sont
consignés dans le tableau 2.
Tableau 2: Richesse spécifique des grands et
moyens mammifères dans quelques sites du Sud-est, Cameroun
|
Site
|
Richesse spécifique
|
Auteurs
|
|
Parc National de Lobéké
|
32-45
|
WCS (1996); Stromayer et Ekobo (1991); Elkan (1994); Ekobo
(1995); Nzooh (2003)
|
|
Parc National de BoumbaBek et Nki
|
34
|
Ekobo (1998)
|
|
Forêt de Ngoila-Mintom
|
37
|
Nzooh (2003)
|
|
ZICGC N°1, 2, 3, 8 et 9
|
26-33
|
Nzooh et al. (2002)
|
|
UFA 10-018
|
27
|
Nzooh et al. (2003a)
|
|
UFA 10-022
|
38
|
Nzooh et al. (2004a)
|
|
UFA 10-013
|
28
|
Nzooh et al. (2004b)
|
|
UFA 10-051
|
22
|
Bobo et al. (2006a)
|
|
UF1 10-053
|
23
|
Bobo et al. (2006b)
|
|
UFA 10-013
|
22
|
Bobo (2009a)
|
|
UFA 10-025
|
20
|
Bobo (2009b)
|
|
UFA 10-063
|
24
|
Bobo (2009c)
|
|
ZIGC N°13 et 14
|
26
|
Djafsia et Kegne (2012)
|
Source : Djafsia et Kegne (2012).
On y retrouve des espèces qui sont très
sensibles aux pressions anthropiques ou aux perturbations de leurs habitats :
c'est le cas du bongo, du buffle, du céphalophe à dos jaune, du
chevrotain aquatique, de l'éléphant, du chimpanzé, du
pangolin géant et du sitatunga
2.8
Richesse specifique dans les ZICGC 13 et 14 au Nord du Parc National de Boumba
- Bek
Les signes d'activités de 26 espèces de grands
et moyens mammifères appartenant à six familles (bovidés,
primates, proboscidés, rongeurs, tragulidés, pholidotes) ont
été recensés dans la zone d'étude pour un total de
9921 observations avec le céphalophe roux, le céphalophe bleu, le
porc-épic, et l'éléphant qui sont les plus
représentés avec respectivement un IKA de 8,74 ; 6,11 ; 4,42 ; et
1,40 alors que le daman des arbres, le léopard et le buffle sont presque
inexistants dans la zone avec respectivement un IKA de 05 ; 0,015 ; 0,015
(Djafsia et Kegne, 2012).
CHAPITRE 3: METHODOLOGIE
3.1
Description de la zone d'étude
3.1.1
Localisation
Le PNBB est situé entre les latitudes Nord de
2°09' à 2°20' et les longitudes Est de 15°35' à
15°50'. Il couvre une superficie d'environ 278.252 ha. Du point de vue de
l'organisation administrative, il est situé dans la région de
l'Est, à cheval entre les arrondissements de Moloundou et de Yokadouma
dans le département de la Boumba-et-Ngoko (Figure 1). Sa zone tampon se
partage entre plusieurs zones de chasse comprenant : à l'Est, les
ZICGC n° 7 et 8 assises sur la zone Agroforestière qui
s'étend de part et d'autre de la route Yokadouma - Moloundou; au Nord,
les ZICGC n° 13 et 14 est au Sud, la ZIC n° 38 et les ZICGC
n° 9 et 10. Plusieurs UFA sont définies dans cette zone tampon, en
superposition aux ZIC et des ZICGC, notamment les UFA n° 10-018 au Nord et
10-015 au Sud. L'Ouest de cette zone tampon est constitué du PNN. Il
faut aussi ajouter la présence de la Forêt Communale de Moloundou
au Sud-Est du PNBB.
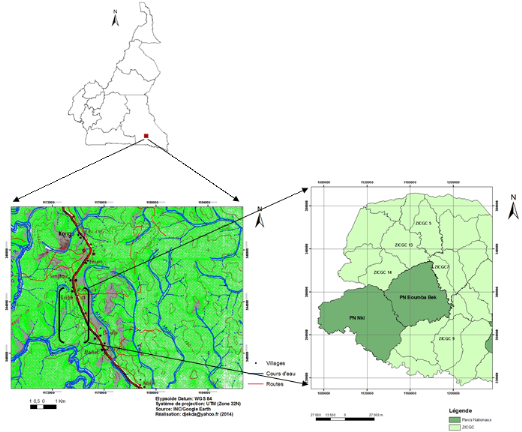

Figure 1: Localisation de
la zone d'étude dans la région et du PNBB
Source: Atlas du Cameroun (2011) adapté et
actualisé par l'auteur (2014).
3.1.2
Caractéristiques biophysiques et écologiques
3.1.2.1 Climat
La région du PNBB est caractérisée par un
climat de type équatorial à quatre saisons. La hauteur des
précipitations est d'environ 1.500 mm/an. Les précipitations
s'étalent tout au long de l'année avec deux pics en avril et en
octobre (Harrison et Agland, 1987 ; Ekobo, 1995). La grande saison des
pluies s'étend de septembre en novembre alors que la petite va de mars
en juin. La grande saison sèche dure de décembre en
février et la petite de juillet en août. Les moyennes mensuelles
de températures varient de 23,1°C à 25°C. La
température annuelle moyenne est de 24°C. L'humidité
relative de l'air varie de 60 à 90 %. Des taux de saturation peuvent
être enregistrés pendant les nuits de saison sèche.
3.1.2.2 Relief et topographie
La région de Boumba-Bek, située sur le versant
nord-ouest de la cuvette congolaise, est caractérisée par des
formations métamorphiques typiques d'âge Précambrien
à Cambrien appartenant à la série du Dja inférieur.
Ces formations métamorphiques sont composées essentiellement de
tillites, quartzites, schistes, calcaires et dolérites. Le socle
Boumba-Bek est d'origine précambrienne, constitué en une base
cristalline de granites et de roches métamorphiques recouvertes de
schistes et de pierres à chaux et quartzite de grès appelé
série basse du Dja (Laclavère, 1979 ; Ekobo, 1998). Son
relief est accidenté par endroits. On y retrouve de nombreux
affleurements rocheux comparables à ceux observés dans le Dja
ainsi qu'au niveau du corridor entre le Dja et le futur PNN.
3.1.2.3 Pédologie et géologie
Les sols de la région de Boumba-Bek appartiennent au
sous-ordre des sols ferrallitiques typiques définis par une
séparation du fer et de l'oxyde d'alumine. Ils sont argileux et la fine
couche superficielle d'humus contient peu de matière organique. Ils sont
acides et pauvres en azote et bases échangeables. Ces sols peuvent
être divisés en deux groupes qui sont les sols rouges
dérivant de l'ancienne roche mère métamorphique et
éruptive et les rouges dérivant des anciennes roches basaltiques
(Laclavère, 1979).
3.1.2.4 Hydrologie
Le système hydrographique de Boumba-Bek coule vers le
Sud jusqu'aux rivières Dja puis Ngoko, deux affluents du fleuve Congo.
Il est formé des rivières Boumba à l'Est, Bek à
l'Ouest et au Sud, Apom et Gbwogbwo au Nord. Les sources de ces
différentes rivières entretiennent un complexe de
clairières marécageuses communément appelées
baï en référence à leur nom dans la langue Baka
(Harrison et Agland, 1987).
3.1.2.5 La
végétation
Sur le plan phytogéographique, la région de
Boumba-Bek est située sur la limite Est du système du Dja. Elle
est sous l'influence de la forêt du Dja dans sa partie Ouest et de la
forêt semi-décidue dans la région de Boumba-Bek. Cette
diversité d'influences phytogéographiques y entretient une
variété d'habitats naturels.
Le PNBB est caractérisé par un mélange de
forêt semi-décidue (98%) et de forêt marécageuse
à Raphia (2%) (Letouzey, 1985). Ce massif forestier n'a jamais subi
l'exploitation forestière.
La flore de Boumba-Bek est très diversifiée. Les
résultats des inventaires botaniques ont permis d'identifier près
de 831 espèces avec un DHP > 10 cm. Ces espèces sont
réparties en 111 familles différentes (Ekobo, 1998). La
région de Boumba-Bek se situe dans la zone d'inversion de la
phénologie des plantes ; le mouvement des espèces animales dans
cette région apparaît lié au rythme de nombreux
végétaux. Près de 44 espèces
végétales sont des essences d'une très grande valeur
commerciale. Les écorces, les graines et les fruits secs de beaucoup
d'espèces végétales de la région sont
exploités et commercialisés par la population locale. On peut
citer Irvingia gabonensis, Ricinodendron
heudelotii, Tetrapleura tetrapleura, Gnetum
africanum, Afromomum dalzeillii, Cola spp., Baillonella
toxisperma (Ekobo, 1998).
Les résultats des études de la
végétation ont permis de constater que la forêt de
Boumba-Bek est une mosaïque de végétation variant entre la
forêt semi-décidue à l'extrême Est
(Moloundou-Mimbo-Mimbo) et la forêt sempervirente à
l'extrême Ouest. La végétation ici est
caractérisée par une canopée fragmentée entretenue
par l'activité des éléphants. Par endroits, cette
forêt comporte de grandes superficies de forêt
monospécifique à Gilbertiodendron dewevrei
(Letouzey, 1985). Une nouvelle variété de Lophira
alata (Ochnaceae) a été trouvée dans la
région de Boumba-Bek et de Ndongo-Adjala. Deux espèces
endémiques de lianes (Milletia duchesnei et M.
sp.) y ont été identifiées (Ekobo, 1998). Un total de
près de 14 types de formations végétales y a
été trouvé parmi lesquelles de nombreuses
clairières marécageuses et de clairières sur schiste. La
quasi-dominance des Gramineae dans les clairières sur schiste constitue
un facteur majeur d'attrait des herbivores dans ces sites, notamment les
buffles, dont les jeunes repousses sont très prisées par ces
derniers. Environ 41 sur 131 espèces végétales ligneuses
identifiées dans la région font partie de la pharmacopée
Baka (Kenfack et Fimbel, 1995). A cette diversité d'habitats naturels,
est associée une importante biodiversité animale que
révèlent les résultats de l'inventaire de quelques groupes
zoologiques (mammifères, poissons) (Nzooh Dongmo et Bene Bene, 2005).
3.1.2.6 La faune
3.1.2.6.1
Mammifères
Les résultats des inventaires de la faune mammalienne
confirment la présence de 34 espèces de grands mammifères
communs aux sites de Boumba-Bek et de Nki dont 11 espèces de primates,
12 espèces d'ongulés et quatre espèces de carnivores. Les
densités d'éléphants et de gorilles sont respectivement
0,3 et 0,9 individus au km2 (Ekobo, 1998). Il faut noter que dans
cette région, compte tenu de la diversité des habitats et du
caractère intact de la forêt primaire, le nombre d'espèces
probables de mammifères dont la présence reste encore à
confirmer se situe autour de 180. Dans cette forêt, cohabitent deux
sous-espèces de colobe : Colobus polykomos
guereza et C. p. satanas.
La faune des insectivores et des rongeurs, qui regroupe le
grand nombre d'espèces de mammifères, reste encore inconnue dans
différents habitats naturels de la région. A titre d'exemple,
l'écureuil pygmée Myosciurus pumilio avait
récemment été découvert au Sud du PNBB,
précisément au Nord de l'UFA 10 015 (coordonnées
d'observation 2°11'62''N ; 14°51'35'E) (Bobo, 2002). Ce minuscule
écureuil, ayant un statut de Vulnérable (UICN, 2000), a une
distribution très restreinte (du sud-est Nigeria au Gabon) et est
essentiellement confiné à la forêt atlantique.
L'espèce atteint ici la limite orientale de sa répartition et il
s'agit apparemment de la donnée la plus à l'Est pour
l'espèce (Bobo, 2002).
3.1.2.6.2 La faune aviaire
Les connaissances de l'avifaune de la région de
Boumba-Bek restent parcellaires. Dowsett-Lemaire et Dowsett (1999) y ont
effectué un inventaire préliminaire. Un autre inventaire partiel
y avait aussi été effectué, précisément dans
les UFA 10 015 et 10-018 situées respectivement au Sud et au Nord du
PNBB où 194 espèces respectivement 121 espèces avaient
été identifiées (Bobo, 2002). Un complément
d'inventaire s'impose notamment dans la partie Ouest du PNBB sous l'influence
du Dja. Mais, l'avifaune de la région de Boumba-Bek peut en partie
(surtout pour sa région Est) être assimilée à celle
de la région voisine de Lobéké où 305
espèces ont déjà été identifiées
(Dowsett-Lemaire et Dowsett, 1997 ; 1999 ; 2000) et dont :
- trois espèces, notamment Ageslaster niger,
Apaloderma equatoriale et Criniger olivaceus, ont une
répartition très localisée et figurent sur la liste rouge
de l'UICN (UICN 2000, Dowsett-Lemaire et Dowsett, 1997 ; Collart et Stuart
,1985) ; Apaloderma equatoriale a déjà été
identifiée au sud du PNBB (Bobo, 2002) et beaucoup d'autres
espèces de ce statut y existent sûrement ;
- trois autres espèces, notamment Glaucidium
capense, Phylloscopus budongoensis et Ortygospiza locustella,
n'avaient été observées nulle part ailleurs dans le pays
sauf dans la région de Lobéké (Dowsett-Lemaire et Dowsett,
1997), sûrement existent aussi à Boumba-Bek car Phylloscopus
budongoensis a déjà aussi été observée
dans l'UFA 10 015 au Sud du PNBB (Bobo 2002) ;
- la plus grande colonie de fauvette du Dja Bradypterus
grandis observée (20 couples) se trouve dans les marais à
Rhynchospora de la forêt de Lobéké (Dowsett-Lemaire et
Dowsett, 1999) ; Cette espèce a aussi été observée
dans le PNBB, précisément dans le complexe de baïs de
Kopandako au Nord du village Ndongo dans l'UFA 10 015 (au Sud du PNBB) (Bobo,
2002);
- le site de Lobéké abrite plusieurs autres
espèces forestières rares comme l'Ibis olivâtre
Bostrychia olivacea, l'indicateur de Zenker Melignomon
zenkeri, le Gobemouche de Tessmann Muscicapa tessmanni et le
tisserin à cape jaune Ploceus dorsomaculatus qui existent aussi
sûrement à Boumba-Bek.
Dans le PNBB, il existe d'importantes colonies de perroquets
à queue rouge (Psittacus erithacus) qui sont
très recherchés pour le marché international.
3.1.2.6.3 La faune
ichtyologique
Environ près de 121 espèces de poissons y ont
déjà été identifiées, dont deux sont
nouvelles pour la science à savoir Aphyosemion sp. et
Phenacogrammus sp. Sur le plan écologique, ces
espèces de poissons appartiennent au groupe de « big brother fish
» et de « rapid fish » (Makazi et al., 1998). Cette
région n'est pas marquée par un haut degré
d'endémisme. Un inventaire complémentaire est
nécessaire.
3.1.2.6.4
Lépidoptères
Les collectes effectuées dans UFA 10-015 et 10-018,
respectivement situées au Sud et au Nord du PNBB (Bobo, 2002) ont permis
d'identifier 96 espèces respectivement 20 espèces de papillons
appartenant à 17 sous-familles respectivement à neuf
sous-familles (Bobo, 2002). Cette liste préliminaire et
incomplète en de la période d'échantillonnage très
courte, mais démontre cependant la grande richesse spécifique de
la faune des Lépidoptères de la région de Boumba-Bek, qui
s'apparenterait bien à celle observée à
Lobéké où près de 215 espèces y ont
été répertoriées dont 80 % d'espèces
forestières et huit espèces endémiques au Cameroun
notamment Liptena sauberii, Liptena yakadumae, Aslauga modesta, Cymothoe
crocea, Cymothoe radialis, Berbearia jolyana, Euphaedra margaritifera et
Euphaedra stellata (Davenport, 1998). Un inventaire
complémentaire est indispensable.
3.1.3
Relations Faune - Flore
Dans le PNBB, il existe une spécialisation relative du
milieu pour l'habitat des espèces animales, ainsi que des liaisons
spécifiques entre espèces végétales et
espèces animales. Dans cet ordre :
Les clairières marécageuses ou baïs
constituent des pôles d'attraction et des refuges pour plusieurs
espèces de mammifères (éléphant de forêt,
gorille de plaine, buffle, bongo, sitatunga, hylochère,
potamochère, léopard, etc.) et d'oiseaux (perroquets, pigeons
verts, fauvette du Dja, etc.). Ces clairières, en plus des salines
naturelles, regorgent plusieurs espèces végétales dont les
organes sont très appréciés par les animaux. Il s'agit des
bulbes de Rhyncospra corymbosa et Kyllinga polyphylla pour les
gorilles, des feuilles d'herbacées telles Brillantesia sp.,
Indigofera hendendeca-phylla, Heteranthera callifolia,
Heteranthoecia guineensis, Ludwigia erecta, etc... pour les
Artiodactyles et des fruits de Cyperaceae pour les perroquets et pigeons verts
(Nzooh Dongmo, 2005).
Les forêts secondaires très anciennes en
périphérie du PNBB, notamment au Nord de l'UFA 10 015,
constituent en plus des clairières sur sols marécageux, des
pôles d'attraction des éléphants et buffles dont des
couloirs de migration avaient été mis en évidence entre
les baïs de cette UFA et la région Ouest du PNBB (Bobo, 2002) est
formellement établi par le suivi des éléphants du groupe
« Djako » par émetteur radio VHS/UHF (Nzooh Dongmo, 2005).
Dans ces formations végétales, les tiges de Marantaceae telles
Hypselodelphis scandens, Haumania danckelmaniana, Magaphrynium spp.
Sarcophrynium spp. et de Zingiberaceae telles Aframomum spp. et
Renealmia spp. sont, non seulement consommées par les gorilles,
mais aussi utilisées pour la fabrication de leurs nids, tandis que les
feuilles de la plupart des arbustes et lianes sont consommées par les
éléphants.
Les fruits de plusieurs espèces végétales
sont abondamment consommés par les grands mammifères. Il s'agit
de Autranella congolensis, Omphalocarpum spp.,
Gambeya spp., Landolphia spp., Strychnos spp., Irvingia spp., Klianedoxa
gabonensis, etc. La distribution et le rythme de fructification de ces
espèces influencent directement les mouvements de ces animaux.
3.1.4
Caractéristiques socioculturelles et démographiques
Bien que la densité des populations humaines vivant
dans la région de Boumba-Bek soit parmi les plus faibles du Cameroun, le
peuplement y est ancien et relativement hétérogène. On
distingue globalement deux groupes de populations : les populations natives (ou
autochtones) qui vivent dans la région depuis plusieurs
générations et les populations immigrantes (ou allogènes)
arrivées plus récemment au cours des deux dernières
décennies à la suite de l'ouverture du massif forestier par
l'exploitation forestière.
3.1.4.1 Populations natives
Suivant un modèle courant dans l'Est Cameroun, les
populations natives de la région de Boumba-Bek forment
traditionnellement un complexe pluriethnique composé d'un
côté de la population semi-nomade Pygmées dites Baka qu'on
retrouve un peu partout, et de l'autre des populations traditionnels dites
villageoises ou d'agriculteurs Bantou, composées de Boman, Bangando,
Bakwélé et Sangha-Sangha au Sud et de Bidjuki, Mbimo, Mvongmvong
et Konabembe au Nord (Ekobo, 1998).
Les Baka, et leurs voisins/associés Bantou,
résident en règle générale au sein de villages
communs situés au bord des routes, où ils sont cependant
établis dans des hameaux distincts. Certains hameaux Baka sont
implantés en retrait dans la forêt. Des relations de
pseudo-parenté ou d'amitié rituelle, perpétuées de
génération en génération, déterminent une
communauté de vie entre les Baka et leurs voisins. Il en résulte
une co-exploitation des mêmes espaces forestiers par des membres des
lignages Baka et Bantou associés.
Les communautés natives de la région de
Boumba-Bek sont héritières du régime politique
acéphale des peuples forestiers. Cette forme d'organisation de la vie
politique est caractérisée, aujourd'hui, d'une part par une
résistance à toute forme de hiérarchie rigide. D'autre
part, elle met en scène une multiplicité de forme
d'autorité politique qui s'exerce surtout sur le mode de l'influence, de
la persuasion et non de la décision. Il existe souvent une
disparité entre les représentants officiels (chefs traditionnels)
et d'autres formes d'autorité découlant tantôt de
l'application de règles coutumières et tantôt de la
disposition d'un capital de richesses et d'influences pouvant être
investies dans le milieu local.
L'organisation politique est aussi partiellement fondée
sur une différenciation socioculturelle qui tient en minorité les
femmes, les jeunes et les Pygmées Baka. Les femmes et les jeunes Bantou
peuvent, à l'occasion des initiatives locales de développement
montées par leurs propres soins, jouir d'un certain pouvoir
économique et partant d'une certaine influence politique. Le projet
AAPPEC (Association pour l'Auto-Promotion des Populations de l'Est Cameroun),
oeuvre depuis une trentaine d'années en faveur des Baka. Cependant, les
clivages socio-politiques demeurent importants et se révèlent
difficiles à concilier avec les exigences du développement
communautaire, inhérentes aux politiques actuelles en faveur du monde
rural.
3.1.4.2 Populations migrantes
Aux populations natives, s'ajoutent progressivement de
nombreux immigrants aux origines et aux modes d'établissement
très divers. On peut en distinguer plusieurs grandes catégories
:
La population allogène, travaillant pour les
sociétés d'exploitation forestière. Elle vit dans les
agglomérations de type urbain (villes-usines) situés à
proximité des usines de transformation du bois (Lokomo, Masséa,
Ngolla 35 et Sengbot). Les régions d'émigration sont très
diverses. Le cercle des cadres des différentes sociétés
forestières est en majorité constitué par des
expatriés (européens et asiatiques) et des Camerounais
recrutés parmi la population des grandes villes. En raison de la
sous-scolarisation de la région, les responsables des
sociétés forestières constituent également leurs
équipes d'ouvriers spécialisés en déplaçant
des individus d'autres régions du Cameroun.
Les marchands musulmans appelés couramment «
Haoussas » ou « Bamouns ». Ils sont originaires du Grand-Nord,
de la province de l'Ouest, des pays voisins et même de la
sous-région d'Afrique de l'Ouest. Installés comme distributeurs
des produits manufacturés et de la viande de boeuf en milieu urbain,
certains n'hésitent pas à s'investir dans le négoce plus
lucratif du trafic des matières précieuses (ivoire, or, diamant,
etc.). Ils sont présents dans une diversité de lieux incluant les
centres urbains (chef-lieux d'unités administratives et
villes-usines).
Les pêcheurs artisanaux, installés sur les bords
des grandes rivières (Boumba, Bek, Ngoko) et le long du fleuve Sangha,
souvent originaires de la sous-région d'Afrique de l'Ouest
(Sénégal, Mali).
Les exploitants illégaux des produits fauniques
(gibier, trophées, perroquets vivants, etc.). Les populations
autochtones les désignent comme étant les « vrais
braconniers », certainement en raison de leur établissement en
forêt profonde qui tranche avec leur origine urbaine. Leurs
activités profitent des pistes et des chantiers forestiers ouverts par
les sociétés forestières.
Les exploitants miniers artisanaux (or, diamant, etc.). Ils
viennent essentiellement des pays voisins (RCA, Congo). Quelques-uns sont
associés à des Camerounais qui disposent de permis d'exploitation
en règle, établis par les services locaux du Ministère des
Mines, de l'Eau et de l'Énergie (MINMEE). Cependant la plupart des
artisans miniers mènent leurs activités illégalement.
3.1.4.3 Répartition spatiale
Les villages riverains du PNBB s'égrènent le
long des routes Yokadouma-Moloundou-Ndongo sur un tronçon d'environ 180
km compris entre les lieux dits Carrefour-Sengbot au Nord et entre l'axe
Moloundou-Ndongo plus au Sud d'une part et entre les lieux dits
Carrefour-Sengbot au Nord et le village Maléa ancien au Sud-Ouest sur
environ 50 km. Ces localités comptent l'essentiel des populations
natives de Boumba-Bek. La ville de Moloundou et les sites forestiers
industriels de Sengbot, Masséa, Ngolla 35 et Lokomo rassemblent le gros
des populations allogènes.
En raison du relatif éloignement (40 km en moyenne) des
principaux centres urbains de la région (Yokadouma, Moloundou, Libongo,
Kika) du site de Boumba-Bek, on peut considérer que les populations
riveraines sont presque entièrement rurales. Le niveau
d'hétérogénéité y est très faible.
Les seuls habitants non autochtones des villages sont quelques fonctionnaires,
des commerçants et des « vrais braconniers ». Sans qu'il ne
soit possible d'indiquer avec précision les effectifs de ces
populations, on peut, sur la base des données du recensement
préliminaire effectué par le PROFORNAT/GTZ (Zouya-Mimbang, 1998)
et les estimations des autorités locales, estimer que les populations
Baka représentent environ 50 à 55% de la population autochtone.
Cependant, une étude faite par Madzou (2003) sur l'état des
tendances démographiques de tous les 14 villages et deux campements
forestiers périphériques au Nord du PNBB indique une population
estimée à 7239 habitants dont 50,15% d'hommes contre 49,84% de
femmes. Cette population du Nord du PNBB est globalement jeune, moins
scolarisée et quasi-analphabète.
Les densités de population calculées pour les
arrondissements de Yokadouma et de Moloundou dans le département de la
Boumba-et-Ngoko, à partir des données fournies par les services
de la Direction de la Statistique et de la Comptabilité Nationale
(DSCN), s'élevaient en 1997 à 6,4 hab./km2 pour l'arrondissement
de Yokadouma contre 2,1 hab./km2 pour l'arrondissement de Moloundou. Ces
densités sont à prendre avec beaucoup de précautions pour
deux raisons : Premièrement, si on fait foi aux analyses de
AIDEnvironment (1998), les effectifs totaux de la population seraient
sous-estimés à cause de la méthode utilisée par la
DSCN pour évaluer annuellement la croissance naturelle de la population
et des migrations ; Deuxièmement, si on considère que la
population urbaine du département de la Boumba-et-Ngoko était
estimée à environ 40% de l'effectif total en 1998
(AIDEnvironment, 1998), ces densités seraient surévaluées
pour la périphérie immédiate de Boumba-Bek où les
populations sont essentiellement rurales.
3.1.4.4 Infrastructures sociales
3.1.4.4.1 Santé
La région de Boumba-Bek relève à la fois
des districts de santé de Yokadouma et de Moloundou qui couvre la
totalité du département de la Boumba-et-Ngoko. L'administration
publique y dispose de deux hôpitaux de district (Yokadouma et Moloundou)
et d'un centre de santé intégré (Salapoumbé). Les
sociétés forestières opérant dans la région
ont mis en place des infirmeries d'entreprises (Lokomo, Sengbot, Ngolla 35 et
Massea) pour leurs ouvriers et les familles de ces derniers. La mission
catholique dispose d'un hôpital à Salapoumbé. Les
hôpitaux de Salapoumbé et de Moloundou sont les seules formations
sanitaires équipées pour des examens sommaires. En dehors des
pharmacies d'entreprises, le système de distribution des
médicaments est mal organisé. Les besoins sanitaires des
populations natives ne sont que très insuffisamment couverts en raison
du coût inaccessible des soins de santé qui a favorisé
l'émergence des soigneurs et pharmacies ambulantes. La pauvreté
et certaines traditions culturelles poussent les populations locales à
se tourner en priorité vers la médecine traditionnelle. Cependant
les solutions thérapeutiques de la pharmacopée sont très
limitées face au surgissement et à la propagation rapide de
certaines affections dues principalement à l'exploitation
forestière (insalubrité et promiscuité dans les
villes-chantiers et les villages relais). Il s'agit notamment du paludisme, des
infections respiratoires, des infestations parasitaires, des infections
cutanées (AIDEnvironment, 1998).
3.1.4.4.2 Education
Les établissements scolaires existant dans la
région du PNBB comprennent 16 écoles primaires, sept centres
préscolaires pour les Baka, un collège d'enseignement secondaire
(Salapoumbé) et un lycée d'enseignement général
(Moloundou). Ces dernières années le domaine de
l'éducation a bénéficié de quelques projets de
viabilisation ou de construction des infrastructures grâce aux fonds
provenant des redevances forestières. Cependant, faute de personnels et
d'allocation budgétaire en rapport avec les besoins, les
établissements scolaires dans l'ensemble fonctionnent difficilement. De
plus, la très grande dispersion des populations empêche le
remplissage optimal des écoles qui aurait permis un déploiement
optimal des ressources humaines. Les communes et certaines entreprises
forestières prennent en charge des maîtres bénévoles
pour pallier le manque criard de personnel (AIDEnvironment, 1998).
3.1.4.4.3 Réseau de
communication
Le PNBB est partiellement entouré par un réseau
routier composé de deux axes principaux, à savoir la route
Yokadouma-Moloundou-Ndongo et l'axe Ngatto nouveau-Maléa ancien, qui
desservent les sites industriels de Sengbot, Masséa, Lokomo, Ngola 35 et
Kika.
Le fleuve Sangha et la rivière Dja au Sud du PNBB,
servant de frontière entre le Cameroun, la RCA et le Congo, sont
également des voies de communication assez fréquentées.
Elles desservent les sites de Lidjombo/Bayanga (RCA), Libongo/Bela (Cameroun),
Bomassa/Kabo (Congo).
La région du PNBB n'est pas couverte par un
réseau de télécommunication. Dans toute la région
du Sud-Est, seule la ville de Yokadouma à plus de 200 km était
par intermittences connectée au réseau de téléphone
par câble, actuellement non fonctionnelle. Cependant la communication
téléphonique entre Yokadouma et les grands centres urbains se
fait par téléphone satellite du genre Turaya. Depuis le
début du mois de juin 2005, Yokadouma est dotée d'un
système de communication téléphonique mobile (Orange et
sûrement MTN un peu plutard). La commune de Moloundou dispose depuis 2002
d'une station de radio diffusion rurale, dont la zone de réception ne
dépasse cependant pas les limites de la ville. Les communications entre
les différents sites du Sud-Est Cameroun et même avec les pays
voisins (Bayanga en RCA, Bomassa au Congo) se font surtout par
radio/fréquence (AIDEnvironment, 1998).
3.1.5
Caractéristiques socioéconomiques
3.1.5.1 Agriculture
Chaque ménage entretient plusieurs parcelles de
cultures vivrières et de cultures de rente. L'agriculture
vivrière est surtout destinée à l'autoconsommation. Tout
au long de l'année, la banane plantain constitue la base de
l'alimentation des populations natives de la région de Boumba-Bek. Avec
une demande importante dans les villes et les sites forestiers industriels, un
surplus est vendu régulièrement tout au long de l'année.
La production de maïs, relativement importante, est essentiellement
destinée à la fabrication d'un vin local très
alcoolisé (arki ou ngolos). Ce produit dont la commercialisation
représente, surtout pour les femmes, une importante source de revenus
cause cependant des problèmes d'alcoolisme parmi les populations
locales.
Le cacao est la principale culture de rente. Comme dans
l'ensemble du département de Boumba-et-Ngoko, de nombreuses plantations
sont temporairement à l'abandon ou ne sont pas suffisamment entretenues
à cause des mauvais prix actuels sur le marché mondial et du
coût élevé des traitements phytosanitaires. Sur 29.007 ha
de cacaoyères plantées dans la région du Sud-Est,
seulement 26.030 ha sont entretenus (5 ha / producteur pour une récolte
de 250-350 kg/ha en moyenne) (Hecketsweiler et al., 2001).
Cependant la culture de cacao demeure la principale source de revenus agricoles
des villageois Bantou.
Malgré l'ancienneté de l'établissement
des villages de long des axes routiers Yokadouma-Moloundou-Ndogo et Ngatto
nouveau-Maléa ancien (depuis environ 70 ans), les systèmes
agro-forestiers demeurent peu développés, dépassant
rarement le stade primaire de jardins de case. Le faible peuplement, la
prépondérance d'une agriculture itinérante sur
brûlis, les meilleurs rendements réalisés avec la chasse,
une commercialisation agricole fluctuant selon l'humeur des transporteurs et
l'état des routes, une forte proportion de Pygmées et de jeune
fuyant tout « asservissement » agricole peu lucratif sont autant
d'éléments qui peuvent expliquer cette situation (Hecketsweiler
et al., 2001).
3.1.5.2 Chasse
On peut distinguer globalement deux modes de chasse par les
populations riveraines de Boumba-Bek : la chasse de subsistance et la chasse
commerciale. La chasse de subsistance peut être définie comme
celle dont le produit est essentiellement destiné à
l'autoconsommation bien qu'un faible surplus soit régulièrement
dégagé et vendu dans les villages. Quant à la chasse
commerciale, elle peut être considérée comme celle qui est
pratiquée essentiellement dans un but de commercialisation du gibier et
des trophées. Elle s'apparente au braconnage car s'effectue dans la
plupart des cas dans les zones protégées.
Les chasseurs originaires de la région pratiquent la
chasse de subsistance toute l'année avec cependant une baisse
considérable de l'activité pendant la grande saison sèche
(Jell, 1998). Les ongulés constituent l'essentiel des prises
(Zouya-Mimbang, 1998 ; Ngandjui, 1998). Chaque ménage collecte en
moyenne six gibiers par mois. Le céphalophe bleu Cephalophus
monticola (Ngandjui, 1998) constitue plus de la moitié des
prises. Le piège à collet d'acier, procure jusqu'à 84 %
des prises (WCS, 1995). Cette technique de piégeage dont l'usage est
très répandu est cependant illégale.
La majorité des acteurs de la chasse commerciale se
recrutent parmi les immigrants. La saisonnalité ici est identique
à celle de la chasse traditionnelle. Seulement 10 % du gibier
chassé est destiné à l'autoconsommation. Les produits de
la chasse sont écoulés sur les marchés situés
à Kika, Moloundou, Yokadouma et Bertoua au Cameroun, à Ouesso,
Kabo et Pokola au Congo. Les fusils (16 %) et les câbles d'acier (84 %)
sont utilisés. Le Céphalophe de Peters Cephalophus
callipygus constitue l'essentiel des captures (86 %). Chaque chasseur
commercial collecte en moyenne 29 ongulés par mois, ce qui correspond
à une valeur monétaire moyenne de 87.000 FCFA / mois.
La raréfaction de la faune dans les zones proches des
villages oblige les chasseurs locaux à se rendre de plus en plus loin
dans la forêt. Pendant les pics de cette activité en saison des
pluies les chasseurs peuvent parcourir plusieurs dizaines de kilomètres
et établir des campements temporaires de chasse à
l'intérieur de la forêt. Ils franchissent surtout les limites Est
et Nord du Parc pour se retrouver autour des baïs et le long des
principales rivières (Boumba, Bek, Apom). L'approvisionnement des grands
centres urbains et des sites industriels en gibier est facilité par les
transporteurs locaux.
3.1.5.3 Pêche
En zone rurale, la pêche est une activité
très marginale mais se pratique régulièrement le long des
cours d'eau (Boumba, Bek, Apom et Gbwogbwo). Les populations natives pratiquent
régulièrement la pêche en amont des rivières qui
inondent périodiquement les clairières marécageuses
(baïs) à l'intérieur et hors (UFA 10 015) du PNBB pendant la
première moitié de la grande saison sèche
(décembre-janvier). En période de décrue et
d'étiage, la pêche à barrages (à l'écope et
à la nasse), surtout pratiquée par les femmes et les enfants est
une activité traditionnelle très appréciée. La
production (poissons, crevettes, moules, huîtres, crabes, tortues etc.)
est en quasi-totalité autoconsommée. Elle représente un
complément alimentaire non négligeable. Certaines espèces
(Dalophys sp., Malapterus electricus, Pollimyrus kingsleyae, Nannochromis
candifacsiatus) sont utilisées en médecine
traditionnelle.
Les ethnies natives de la région du Sud-Est Cameroun
n'ont pas dans l'ensemble développé une tradition de pêche
dans les grandes rivières pourtant abondantes et poissonneuses (Sangha,
Ngoko, Boumba, Bek etc.). Il existe cependant quelques exceptions : les
minorités ethniques Sangha-Sangha et Bakwélé (Essel)
vivant le long de la Ngoko pratiquent la pêche en pleine eau tandis que
le long de la Sangha, les Kaka pratiquent la pêche à la nasse en
barrant les embouchures des marigots et des petits confluents. Toutefois ce
sont surtout les immigrants Ouest-africains et les ethnies Centrafricaines qui
exploitent les ressources halieutiques de ce secteur fluvial frontalier. A
proximité des sites industriels forestiers, quelques immigrants
pratiquent la pêche commerciale mais leur faible nombre ne permet pas de
garantir un approvisionnement suffisant et régulier (Hecketsweiler
et al., 2001.).
3.1.5.4 Cueillette
Sur 131 espèces végétales ligneuses
identifiées dans la région de Lobéké qui est
voisine à celle de Boumba-Bek, 41 font partie de la pharmacopée
Baka (Kenfack et Fimbel, 1995). Une enquête ethnobotanique
effectuée auprès des populations des villages de l'axe
Kouméla-Nguilili (Gwet, 2003) fait état de l'utilisation de 584
espèces végétales (arbres, herbes et lianes) ; dont 28,8 %
pour l'alimentation, 17,8 % pour l'équipement, 15,7 % pour la
construction et 37,7 % pour la médecine traditionnelle. Il
apparaît dans l'ensemble que 22,6 % de ces espèces sont uniquement
récoltés dans la zone agroforestière située entre
le PNBB et le PNL. Les ignames sauvages (7 espèces) y sont aussi
récoltées. Les écorces, les graines et les fruits secs de
beaucoup d'espèces végétales de la région sont
exploités et commercialisés par la population locale. On peut
citer Irvingia gabonensis, Ricinodendron heudelotii, Tetrapleura
tetrapleura, Gnetum africanum, Afromomum dalzeillii, Cola spp., Baillonella
toxisperma (Ekobo, 1998).
3.1.5.5 Exploitation forestière
L'exploitation forestière dans la zone de Boumba-Bek a
commencé vers début des années 1970. Mais, l'exploitation
n'a jamais concerné l'intérieur du Parc. Quatre UFA (10 018 et 10
015) contiguës au PNBB sont aujourd'hui en exploitation active. Elles sont
attribuées à quatre compagnies forestières dont la STBK
(qui remplace aujourd'hui la SIBAF) et la CIBC, la 10 022 attribuée
à la SIM, la 10 023 attribuée à la TTS. Elles
génèrent annuellement (redevances forestières et taxes
d'abattage confondues) pour l'État, les communes riveraines (Moloundou,
Salapoumbé et Yokadouma) et les communautés villageoises des
recettes. Dans le souci d'atténuer l'impact de l'exploitation
forestière autour du PNBB, et dans l'intérêt d'une gestion
intégrée de l'ensemble de l'UTO Sud-Est, des études
d'impact environnementales ont été faites dans ces UFA (Nzooh
Dongmo, 2005).
3.1.5.6 Chasse sportive
Les entreprises de safari qui pour la plupart
exerçaient déjà en savane dans la province du Nord ont
commencé à s'installer, dans la région du Sud-Est et
particulièrement autour du PNL, vers les années 1980. Ce n'est
que cers le milieu des années 1990 qu'un guide de chasse professionnel
s'est installé dans la zone de Ndongo au Sud du PNBB (ZIC 38). La saison
cynégétique va de décembre en juillet. Le bongo est la
principale espèce recherchée en zone forestière. D'autres
espèces telles que l'éléphant, le buffle, le sitatunga,
l'hylochère ou le céphalophe à dos jaune complètent
régulièrement les tableaux de chasse (Nzooh Dongmo, 2005).
Faute d'une délimitation précise et d'un
affermage en règle des zones de chasse pendant les premières
années, l'activité était régulièrement
perturbée par de nombreux conflits entre les l'entreprise de safari et
les communautés villageoises riveraines. A la faveur du processus de
zonage et de classement des forêts, conduit sous la houlette de l'UTO
Sud-Est entre 1999 et 2001, les concessions de chasse safari (ZIC) et des zones
de chasse communautaire (ZIC à Gestion Communautaire - ZICGC) ont
été délimitées en conciliant les
intérêts des différents acteurs (Nzooh Dongmo, 2005).
3.1.5.7 Capture des oiseaux
La capture du perroquet gris à queue rouge
(Psittacus erithacus) et du pigeon vert (Treron
australis) dans la région du Sud-Est Cameroun remonte à
plusieurs dizaines d'années. Les baïs constituent les principaux
sites de capture même si ils sont situés dans les Parcs Nationaux
(donc dans l'illégalité). Les perroquets prélevés
illégalement sont intégrés dans des filières
régulières d'exportation, faute d'organisation adéquate de
l'activité (Nzooh Dongmo, 2005).
La capture des perroquets est exercée par des
opérateurs économiques en provenance des grands centres urbains
(Yaoundé et Douala). La filière est presque entièrement
aux mains d'acteurs immigrants, les populations natives n'y jouant que des
rôles secondaires de manoeuvres et de porteurs. Les pigeons sont revendus
localement comme gibier tandis que les perroquets sont impliqués dans un
trafic international florissant (Nzooh Dongmo, 2005).
3.1.5.8 Exploitation minière artisanale
Le diamant et l'or ne sont pas encore exploités dans le
PNBB. Mais, ils le sont déjà dans sa zone
périphérique, notamment la partie qui jonche avec le PNL
où cette activité est florissante.
3.2
Collecte des données
3.2.1
Le choix de site
L'emplacement exact de la zone d'étude à
l'intérieur des territoires a été
sélectionné parce qu'il abrite le Parc National de Boumba-Bek, la
zone est dotée d'une grande richesse floristique et est convoitée
par de nombreuses sociétés d'exploitation forestière qui
exploitent de nombreuses UFA, deux groupes éthiques peuplent la zone
notamment les Baka et les Konabembé dont les sources alternatives de
revenus sont rares et la faune constitue pour eux la principale source de
protéine.
3.2.2
Données de sources secondaires
Les données secondaires ont été
tirées sur internet, des rapports d'activités JICA-FOSAS ainsi
que des documents de la bibliothèque du Département de Foresterie
(DEPFOR) de l'Université de Dschang (UDs) traitant ou se rapprochant de
notre étude dans d'autres sites.
3.2.3
Données de sources primaires
Des études
précédentes réalisées par Kengne (2012) et Djafsia
(2012), trois sites ont été identifiés donc l'un
situé en forêt primaire, l'autre en forêt secondaire et le
dernier dans le PNBB (Figure 2). Dans chaque site, six transects de 2 km de
longueur et espacés de 2 km les uns des autres, reliés par des
recces de 2 km ont été installés et suivis de façon
permanente. L'ouverture des transects (Figures 3 et 4) sur le terrain a
consisté à dégager légèrement la
végétation à l'aide des machettes sans pour autant
perturber l'habitat de la faune. La carte a été utilisée
pour localiser le point de départ par rapport à la ligne de base
(135°). Le long de chaque transect, des marques étaient
portées tous les 25 m. Les rubans utilisés sur les points de
départ et fin des transects portaient les indications suivantes: N°
du transect, la longueur du transect; l'orientation et une mention,
début recce sur les fins transects.

Figure 2: Site
proposé pour le suivi de la faune
Source : Kengne (2012).
Figure 3: Visée
d'angle pour ouverture des transects
Figure 4: Ouverture des transects


Le potentiel faunique a été évalué
par quatre différentes méthodes dont les inventaires diurnes sur
transects linéaires basés sur les observations directes
(spécimens vus ou cris entendus), les inventaires diurnes sur transects
linéaires basées sur les observations indirectes (empreintes,
crottes, nids, épines, pistes...), les inventaires nocturnes sur
transects linéaires basés sur des observations directes (White et
Edward, 2000) et l'utilisation des caméras infra-rouges.
3.2.3.1 Technique d'inventaire diurne: méthode
directe
Pendant les inventaires diurnes basée sur la
méthode directe, de 6h à 15h, dépendamment des conditions
météorologiques, les différentes espèces fauniques
ont été observées directement (spécimens vus ou
cris entendus), sur les transects permanents prédéfinis comme
décrite par (Bobo, 2009 ; Viquerat et al., 2012; Djetkeu,
2010; Whitesides et al., 1988). La
1ère collecte des données ont été faites
deux semaines après installation des transects permanents. Le long des
transects, des fiches de collecte de données pré-établies
(Annexe 1) ont été utilisées pour collecter des
informations sur des observations directes (spécimens vus ou cris
entendus). Les coordonnées des points où les animaux ont
été observés et celles des activités anthropiques
étaient enregistrées dans le GPS. Il a été
relevé en plus des données précédentes, les
distances le long du transect (du point de rencontre), le type de formation
végétale et/ou habitat, les distances perpendiculaires à
la direction du transect pour des observations directes d'individus, le nom de
l'espèce identifié etc.
3.2.3.2 Technique d'inventaire diurne : méthode
indirecte
Pour cette méthode indirecte, les inventaires
commençaient à 6h et s'achevaient à 15h. Pendant ces
inventaires, les différentes espèces fauniques ont
été observées indirectement (empreintes, crottes, nids,
épines, pistes ...) et les signes des activités humaines (chasse,
pêche, cueillette, culture, feux de brousse...) sur les transects
permanents prédéfinis comme décrite par (Bobo, 2009;
Viquerat et al., 2012; Djetkeu, 2010; Whitesides et al.,
1988). Tous ces indices de présences ont
été notés sur la fiche de données
pré-établies (voir annexe 1). Il a été
enregistré en plus des données précédentes, les
distances le long du transect (du point de rencontre), le type de formation
végétale et/ou habitat, les distances perpendiculaires à
la direction du transect pour des rencontres de crottes de mammifères ou
de nids de primates, l'âge des indices de présence des
espèces rencontrées, le nom de l'espèce identifié ont été
relevés.
3.2.3.3 Technique d'inventaire nocturne : méthode
directe
A cause de l'éloignement des transects par rapport au
campement, les inventaires nocturnes ont été combinés aux
inventaires diurnes afin de réduire non seulement le nombre
d'entrée en forêt, mais aussi les dépenses
financières. Chaque entrée en forêt était
consécutive à un entretien avec deux chasseurs qui devaient
servir de guide tout au long des inventaires. Ces derniers étaient
informés sur le type d'information à collecter, et enfin sur le
comportement qu'ils doivent afficher pendant l'inventaire.
Pendant les inventaires nocturnes, de 20h à 5h,
l'énergie lumineuse des torches frontales nous permettait de distinguer
les différentes espèces fauniques à partir de la couleur
provenant soit du reflet de leurs yeux ou de leur pelage, le long des transects
permanents. Chaque individu ou chaque groupe d'individus vu ou entendu a
été compté comme décrite par (Waltert et
al., 2006 ; Djetkeu, 2010 ; Viquerat et al.,
2012). Les informations à relever sur des fiches
d'inventaires ont concerné l'espèce, le nombre, la distance par
rapport au transect, la position sur le transect, le nom de l'espèce et
enfin l'heure de la collecte de l'information (voir annexe 1). Les
coordonnées des points où les animaux ont été
observés et celles des activités anthropiques étaient
enregistrées dans le GPS. La figure 5 illustre quelques les images lors
des inventaires nocturnes.




Figure 2: Inventaires
nocturnes
3.2.3.4 Utilisation des caméras infra-rouges:
méthode directe
Elle a consisté à l'installation de deux
caméras infra-rouges sur des arbres à une hauteur de 45 cm du sol
ou au niveau du genou le long de chaque transect de 2 km; la 1er
à 500 m du début du transect et la seconde à 1000 m du
1er. Une capture a été constituée d'images
filmées par les caméras pendant 30 jours. Une capture a
été réalisée pendant cette étude afin de
photographier tous les animaux qui passeront sur lesdits transects, notamment
des animaux « disparus » et de découvrir des animaux inconnus,
dont l'activité est presque totalement nocturne, dont on ignorait la
présence ou dont on ne possédait que des indices de
présence indirects (traces, mues, observations furtives, etc.) ou les
espèces rares difficiles à observer. Le détecteur
enregistre, en même temps le mouvement de l'animal, la date, l'heure et
l'année de cet événement, sur une distance effective de 15
à 30 m comme décrite par (Silveira et al., 2003 ;
Collen et al., 2008; Nkwetaketu, 2011). Les fiches de collecte ont
été conceptualisées pour des informations sur la date
d'installation, les coordonnées et autres (Annexes 2 et 3). Ces
caméras ont été programmées pour ménager un
intervalle entre deux photos successives, de façon à
éviter de multiplier inutilement le portrait d'animaux restant longtemps
sur ces points, tout en ayant une bonne chance d'intercepter les visiteurs
« de passage ». Les piles utilisées, étaient de type
Panasonic de 1,5V. Deux paires de piles ont été installées
dans chaque caméra et ont été rechangées tous les
15 jours. Les SD carte (carte mémoire) étaient de type Transcend
d'une capacité de 2G, ont été installées dans
chaque caméra et rechangée après 15 jours. Une collecte
éventuelle des données (une capture) de sa mémoire vers un
ordinateur a été faite pour un traitement ultérieur de la
fréquentation au point surveillé comme décrite par (Manet
et Herman, 2003). Un silicate de sodium a été installé
dans chaque caméra pour absorber l'humidité contenu dans la
caméra, chaque silicate était renouvelé tous les 15 jours.
Une tôle plastique superficielle a été installée en
dessus de la caméra pour une protection éventuelle contre la
pluie. Les rubans adhésifs ont servi de sceller les bordures de chaque
caméra une fois que ces dernières étaient
paramétrées ceci dans le but d'éviter toutes pluies
tombant sur les flancs de la caméra. Les figures 6 et 7
représentent respectivement la programmation des caméras et la
hauteur à laquelle la caméra était fixée. En effet,
toutes les caméras sont programmées et contrôlées
à la base (Gribé) avant toute opération de fixation sur
les transects permanents.
Figure 3: Programmation des
caméras


Figure 4: Hauteur de
fixation de la caméra
Pour chaque type d'inventaire, 3 passages par transect ont
été réalisés, correspondant à une distance
totale parcourue de 108 km dans la zone d'étude. La figure 8
présente la distribution des caméras sur les 18 transects
parcourus pendant les inventaires.

Figure 5: Distribution des
caméras infra-rouges sur les transects dans la zone
d'étude
Source: Atlas du Cameroun (2011) adaptée et
réactualisé par l'auteur (2014).
Toutes les caméras ont été
distribuées de manières systématiques dans la zone
d'étude. La caméra infra-rouge est dotée d'un
détecteur qui s'adapte en fonction de la lumière émise par
le rayonnement solaire. En effet lorsqu'il fait jour et que le degré
d'ensoleillement semble important, les images résultants sont clairs et
nets (Figure 9). Par contre, dans la nuit les images enregistrées par
les détecteurs sont de couleur noire sur blanc (Figure 10).
Figure 6:
BongoTragelaphus euryceros




Figure 7: Potamochère
Potamochoerus porcus
3.2.3.5 Suivi de dégradation des crottes des grands et
moyens mammifères
Lorsqu'une crotte très fraîche a
été rencontrée (âge 0 c.-à-d. quelques heures
après défécation) pour la première fois, une balise
était posée tout près de celle-ci pour un suivi
éventuel c'est-à-dire, à l'aide d'un marqueur
indélébile le numéro de code et la date de
découverte sur un ruban surplombant a été attribuée
à chaque crotte. Les informations comme, le temps de rencontre, les
données habituelles sur l'état du transect notamment sa
localisation le long du transect à l'aide d'un GPS et la distance
perpendiculaire au transect ont été enregistrés. Pour
chaque parcours du transect, une fiche de suivi a été
préparée (Annexes 2 et 4). Cette dernière a
été visitée tous les 5 jours du à
l'éloignement des sites, jusqu'à la disparition complète
(pourriture). Lors de chaque visite les informations sur l'état de
décomposition de crottes ont été consignées dans la
fiche de collecte pré établie (Annexe 4). Pour une bonne
estimation de densité des céphalophes, il faut suivre un
échantillon de 20-30 crottes pour chaque zone d'étude et chaque
saison comme décrite par (White, 1994).
Tableau 3: Définition d'âges des indices
de présence des espèces rencontrées
|
Nature
|
Age des indices
|
Nature
|
Age des indices
|
Nature
|
Age des indices
|
|
Trace / crotte d'espèces de
mammifères
|
Très frais = quelques
heures
Frais = 1 à 2 jours
Récent = 3 à 5 jours
Vieille = 6 à 14 jours
Très vieille = plus de 14 jours.
|
Nids de gorille et chimpanzé
|
Frais = Feuilles encore vertes - 1 à 5
jours;
Récent = Feuilles mortes, mais nids
toujours intacts - 6 à 14 jours;
Vieux = Feuilles complètement mortes
et sèches mais toujours attachés aux branches - 2 à 4
semaines);
Pourri = Feuilles tombées des
branches, et parfois repousse de la végétation pour les nids au
sol (plus de 4 semaines)
|
Crottes
(Eléphants)
|
A =Crottes intactes, très
fraîches, humides, avec odeur;
B = Crottes intactes, fraîches mais
sèches, sans odeur ;
C = Quelques crottes sont
désintégrées, et d'autres toujours reconnaissable ;
D = Toutes les crottes sont
désintégrées, le tas d'excrément devint une masse
plate;
E = Crottes sur lesquelles on observe des
plantules issues de la germination des graines de fruits consommés.
|
N.B. : Traces = Empreintes, Signes
d'alimentation, Poils, Epines, Plumes
Source : Bobo (2009a).
3.2.3.6 Caractérisation de la végétation
de la faune dans la zone d'étude
Les inventaires des ligneux ont été
réalisés à Gribé et à Gouonepoun le long des
transects permanents. Deux placettes de 25 m x 25 m ont été
installées le long de chaque transect de 2 km pour le comptage des
ligneux. Au total 24 placettes ont été installées sur les
12 transects. La première placette est installée à 500 m
de début de transect et la seconde placée à 1000 m de
cette dernière. A l'intérieur de chaque, placettes toutes les
espèces herbacées et ligneuses ont été
comptées et le degré de recouvrement a été
estimé pour chaque site. Une fiche de comptage des ligneux et des
herbacées a été préparée (Annexe 5).
3.3 Analyse des données
3.3.1
Inventaires diurnes et nocturnes
3.3.1.1 Estimation des densités et de l'abondance
relative
Les données ont été introduites, puis
encodées dans Microsoft office Excel 2010. Les estimations des
densités des crottes ont été obtenues à l'aide du
programme DISTANCE 6.0 (Half-normal Cosine adjustment model) comme
décrite par (Laake et al., 1994). Les densités de
crottes ont été converties en individu/km², en utilisant le
taux de défécation et de taux de dégradation de crottes
estimé par Koster et Hart (1988) dans la forêt dense en
République Démocratique du Congo. Ce programme exige une grande
précision dans les mesures de distances perpendiculaires et permet une
analyse des données pour les espèces suffisamment
observées (au moins 30 observations). Néanmoins, pour les
densités obtenues ayant une faible observation (< 30), le coefficient
de variation a été estimé avec précision. Toutes
les densités de crottes estimées, dont le coefficient de
variation est inférieur à 25% ont été
considéré comme acceptable (comme dans Nzooh-Dongmo, 2003; Bobo,
2006 a). L'intervalle de confiance a été pris à 95% pour
toutes les densités estimées. La fonction de probabilité
de détection des données le long des transects a
été calculée, car elle n'est pas connue d'avance, et varie
avec les facteurs tels que l'environnement et l'expérience de
l'observateur à détecter les animaux. La formule pour
l'estimation des densités est la suivante :
[Densité: E = (Y× R)/ D ou E = Y/ (D×
R')].
Avec: E: Estimation du
nombre d'animaux d'une espèce donnée dans une unité de
surface;
Y: Densité de crottes ou de
nids (n/km2);
R: Vitesse de dégradation de
crottes ou de nids;
R'= 1/R: Période de
dégradation (en jour);
D: Taux de défécation
ou de déposition de nids.
Les observations ont été regroupées en
espèces de mammifères et par type d'activités
anthropiques. Les taux de rencontre au kilomètre ou Indices
Kilométriques d'Abondance (IKA) par transect de différents
groupes d'information ont été calculés. L'IKA est le
rapport entre le nombre total d'observations de chaque signe d'activités
recensées (N) le long des transects et la distance parcourue (D en km).
Il se calcule pour une espèce ou pour l'ensemble des espèces dans
une zone. Effectué régulièrement dans la même zone
et dans les mêmes conditions, il est un bon indicateur de la tendance des
populations animales, car il permet de savoir si la population augmente,
diminue ou stagne. Il se calcule par la formule suivante:
[Abondance relative: IKA = N/D]
Toutes les cartes ont été produites à l'aide
du logiciel ArcGIS 10.0. Le Programme Distance 6.0 nous a permis d'estimer
toutes les densités de crottes et la production éventuelle des
courbes de probabilité de détection des crottes en fonction de la
distance perpendiculaire au transect.
3.3.1.2 Comparaison des densités et de l'abondance
relative (Inventaires diurnes et nocturnes)
Ø Densités de grands et moyens
mammifères
La comparaison des densités des grands et moyens
mammifères pour les inventaires diurnes indirects s'est basée sur
les densités des crottes obtenues au travers de programme DISTANCE 6.0.
En utilisant les taux de défécation et le taux de pourriture des
crottes déterminé par Koster et Hart (1988) en forêt dense
de la République Démocratique du Congo, les densités en
crottes ont été converties en individu/km². Ensuite, les
tests d'analyse de variance (ANOVA) et de Tukey pour la séparation des
moyennes de Tukey ont été utilisés pour voir s'il y a une
différence significative entre les moyennes de densités des trois
sites.
La comparaison des densités des grands et moyens
mammifères pour les inventaires diurnes directs et nocturnes des
individus vus physiquement s'est basée sur les densités obtenues
au travers de programme DISTANCE 6.0. Ensuite, le test de séparation des
moyennes de Tukey a été utilisé pour voir s'il y a une
différence significative entre les moyennes de densités dans les
trois sites.
Ø IKA des grands et moyens
mammifères
La comparaison des moyennes des IKA nécessite une
grande précision (au moins 30 observations) pour qu'une
différence significative soit observée entre les sites.
Néanmoins, toutes les données (= 20 observations) pour une
espèce donnée et dans un site donné ont été
comparées. La comparaison n'est possible pour une espèce dont
l'IKA est égale à zéro dans un site donné. Le test
de séparation des moyennes des IKA de Tukey a été
utilisé pour voir s'il y a une différence significative entre les
moyennes les IKA dans les trois sites.
3.3.2
Caméras infra-rouges
3.3.2.1 Estimation des densités et de l'abondance
relative
L'estimation des densités au moyen des caméras
infra-rouges s'est faite sur la base des enregistrements photographiques des
caméras Bushnell Trophy Cam. Les différentes densités
relatives ont été calculées suivant la formule
développée par Rowcliffe et al. (2011).
[D = Pi/r(2+ ?)]
Où D = densité
(ind/km²)
Pi = passage d'une espèce
i
? = champ de vision en (o)
r = portée maximale du flash
infra-rouge en m
Le champ de vision et la portée maximale du flash sont
respectivement de l'ordre 50o et de 15 mètre. Dans chaque
site, ont été installées 12 caméras infra-rouges.
Les images photographiques résultantes ont été
identifiées et triées grâce à l'aide de deux
chasseurs Baka, ensuite ces images ont été filtrées par le
Logiciel Excel (Tableau Croisé Dynamique). Le WPI a été
calculé pour chaque espèce identifiée. Cet indice
représente le nombre des mouvements des espèces obtenues
automatiquement par les caméras infra-rouges (Nkwetaketu, 2011). Cet
indice a été calculé par la formule:
[WPIspx =
(Nsp/T)x100]
Où: WPI = Indice Photographique de la
Faune (Nombre des passages des espèces
photographiées)
Nsp : Nombre de passage pour
l'espèce x identifiée
T: Effort de capture (caméra-jours)
3.3.2.2 Comparaison des densités et de l'abondance
relative (WPI)
Ø Seules les densités ayant une grandes
précisons des données (= 20 observations) par espèces ont
été comparées par le test de Tukey de séparation
des moyennes, afin de déceler une différence significative entre
les trois sites Il en est de même pour les WPI.
3.3.3
Evaluation de la performance de différentes méthodes
L'évaluation de la performance des différentes
méthodes d'inventaire des grands et moyens mammifères entre les
trois sites, dont le premier est situé en forêt primaire
(Gouonepoun), le second en forêt secondaire (Gribé) et le
troisième dans le PNBB s'est basée sur l'analyse de l'estimation
du coefficient de variation des IKA des espèces recensées, les
IKA des activités humaines, les différentes densités et/ou
les indices de présence des espèces recensées, la vitesse
de décomposition des crottes comme décrite par (Viquerat et
al., 2012). Cependant, la meilleure méthode était celle
ayant le plus petit coefficient de variation. La formule de coefficient de
variation est la suivante :
[CV
 x100]
x100]
Où : CV= Coefficient de
Variation
SD = Standard Deviation (Ecart Type)
u = Moyenne
3.3.4
Analyse de la vitesse de décomposition ou disparition de crottes
Toutes les informations résultantes de la
décomposition des crottes, notamment la date de détection, la
durée de décomposition effective, la position de la crotte par
rapport au transect, ont été encodées dans Excel. Ensuite
la durée moyenne a été calculée en divisant la
durée totale de toutes les crottes par le nombre total de crottes
suivies effectivement, puis à cette durée a été
associée la valeur de son écart type.
3.3.5
Caractérisation de la végétation de la faune dans la zone
d'étude
Les données obtenues ont été saisies et
analysées à partir du logiciel Microsoft EXCEL 2010. Tous les
noms des ligneux commerciaux, obtenus en Baka, ont été
identifiés à l'aide du délégué du
Comité de Valorisation des Ressources Forestières (COVAREF). Pour
chaque ligneux identifié en langue française, leurs noms
scientifiques ont été associés. La proportion de
l'espèce la plus abondante et le degré de recouvrement de chaque
ont été calculées.
CHAPITRE 4: RESULTATS ET
DISCUSSION
4.1
Comparaison des densités et des Indices Kilométriques d'Abondance
(IKA) des grands et moyens mammifères suivants quatre méthodes
d'inventaire dans les trois sites
4.1.1
Inventaires diurnes indirects
4.1.1.1 Richesse spécifique
Les signes d'activités de 26 espèces de grands
et moyens mammifères appartenant à six familles ont
été recensés dans la zone d'étude lors des
inventaires diurnes indirects (Annexe 6). Un total de 4529 observations ont
été relevées pour une distance totale de 108 km (IKA =
41,935). Sur la base des IKA, le céphalophe roux et le céphalophe
bleu sont les plus abondants avec respectivement 11,324 et 9,519 indices de
présence au kilomètre. La panthère, la civette et
l'oryctérope, sont presque absents avec respectivement 0,019; 0,046 et
0,074 signes de rencontre au km (Figure 11).

Figure 8: IKA des
espèces recensées dans toute la zone d'étude (Inventaires
diurnes indirects)
4.1.1.2 Estimation des densités des grands et moyens
mammifères dans les trois sites (Inventaires diurnes indirects)
4.1.1.2.1 Céphalophe
bleu Cephalophus monticola à Gribé
L'estimation de la densité de Céphalophe bleu
s'est faite sur la base des crottes. Au total, 53 crottes ont été
relevées sur les six transects parcourus. La distance perpendiculaire
maximale au transect est de 620 cm. Le tableau 4 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 12 illustre bien l'allure de la courbe
de probabilité de détection des crottes en fonction de la
distance perpendiculaire au transect.
Tableau 4: Statistique d'estimation de la
densité de Céphalophe bleu dans le site de Gribé
(Inventaires diurnes indirects)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
|
|
|
|
|
AIC
|
651,61
|
|
|
|
|
f(0)
|
0,34929E-02
|
0,36705E-03
|
10,51
|
0,28305E-02
|
0,43104E-02
|
|
p
|
0,46177
|
0,48525E-01
|
10,51
|
0,37419
|
0,56984
|
|
ESW
|
286,30
|
30,085
|
10,51
|
232,00
|
353,30
|
|
n/L
|
1,4722
|
|
31,84
|
0,76427
|
2,8360
|
|
D
|
257,12
|
86,208
|
33,53
|
130,37
|
507,08
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des crottes
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen de crottes par unité de distance du
transect en km,
D = densité des crottes (nombre/km2)
AIC = Akaike Information Criterion

Figure 9: Courbe de
probabilité de détection des crottes de Céphalophe bleu
dans le site de Gribé en fonction de la distance perpendiculaire au
transect (Inventaires diurnes indirects)
Au travers de DISTANCE, nous obtenons une densité
moyenne de 257,12 crottes/km2, avec un coefficient de variation de
33,53%: ce qui est acceptable mais, légèrement supérieur
à 25%.
En utilisant les valeurs du taux de défécation
(4,9 crottes/jour) et de dégradation des crottes (18 jours)
déterminées par Koster et Hart (1988) en forêt dense de la
République Démocratique du Congo, nous obtenons une
densité moyenne de 2,915 individus/km2, variant entre les
valeurs 1,478 et 5,749 individus/km2.
4.1.1.2.2 Céphalophe
bleu Cephalophus monticola à Gouonepoun
L'estimation de la densité de Céphalophe bleu
s'est faite sur la base des crottes. Au total, 18 crottes ont été
relevées sur les six transects parcourus. La distance perpendiculaire
maximale au transect est de 410 cm. Le tableau 5 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 13 illustre bien l'allure de la courbe
de probabilité de détection des crottes en fonction de la
distance perpendiculaire au transect.
Tableau 5: Statistique
d'estimation de la densité de Céphalophe bleu dans le site de
Gouonepoun (Inventaires diurnes indirects)
|
Paramètre
|
Valeur Estimée
|
Erreur
standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
211,46
|
|
|
|
|
f(0)
|
0,59190E-02
|
0,14146E-02
|
23,90
|
0,35913E-02
|
0,97552E-02
|
|
p
|
0,41207
|
0,98485E-01
|
23,90
|
0,25002
|
278,45
|
|
ESW
|
168,95
|
40,379
|
23,90
|
102,51
|
278,45
|
|
n/L
|
0,50000
|
|
26,81
|
0,28677
|
0,87177
|
|
D
|
147,97
|
53,151
|
35,92
|
72,830
|
300,65
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des crottes
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen de crottes par unité de distance du
transect en km,
D = densité des crottes (nombre/km2);
AIC = Akaike Information Criterion.
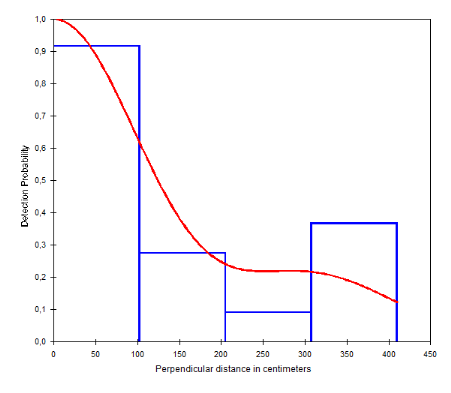
Figure 10: Courbe de
probabilité de détection des crottes de Céphalophe bleu
dans le site de Gouonepoun en fonction de la distance perpendiculaire au
transect (Inventaires diurnes indirects)
Au travers de DISTANCE, nous obtenons une densité
moyenne de 147,97 crottes/km2, avec un coefficient de variation de
35,92%: ce qui est acceptable, mais légèrement supérieur
à 25%.
En utilisant les valeurs du taux de défécation
(4,9/jour) et de dégradation des crottes (18 jours)
déterminées par Koster et Hart (1988) en forêt dense de la
République Démocratique du Congo, nous obtenons une
densité moyenne de 1,678 individus/km2, variant entre les
valeurs 0,826 et 3,409 individus/km2.
4.1.1.2.3
Céphalophe bleu Cephalophus monticola dans le PNBB
L'estimation de la densité de Céphalophe bleu
s'est faite sur la base des crottes. Au total, 14 crottes ont été
relevées sur les six transects parcourus. La distance perpendiculaire
maximale au transect est de 930 cm. Le tableau 6 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 14 illustre bien l'allure de la courbe
de probabilité de détection des crottes en fonction de la
distance perpendiculaire au transect.
Tableau 6: Statistique d'estimation de la
densité de Céphalophe bleu dans le PNBB (Inventaires diurnes
indirects)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
188,07
|
|
|
|
|
f(0)
|
0,28768E-02
|
0,72007E-03
|
25,03
|
0,16813E-02
|
0,49223E-02
|
|
p
|
0,37378
|
0,93559E-01
|
25,03
|
0,21845
|
0,63955
|
|
ESW
|
347,61
|
87,010
|
25,03
|
203,16
|
594,78
|
|
n/L
|
0,38889
|
|
33,77
|
0,19440
|
0,77794
|
|
D
|
55,937
|
23,513
|
42,04
|
24,501
|
127,71
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des crottes
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen de crottes par unité de distance du
transect en km,
D = densité des crottes (nombre/km2)
AIC = Akaike Information Criterion
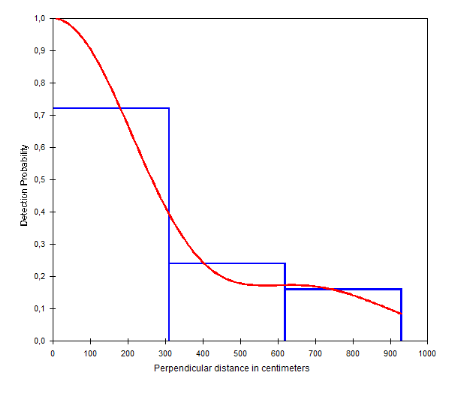
Figure 11: Courbe de
probabilité de détection des crottes de céphalophe bleu
dans le PNBB en fonction de la distance perpendiculaire au transect
(Inventaires diurnes indirects)
Au travers de DISTANCE 6.0, nous obtenons une densité
moyenne de 55,937 crottes/km2, avec un coefficient de variation de
42,04%: ce qui est acceptable, mais légèrement supérieur
à 25%.
En utilisant les valeurs du taux de défécation
(4,9/jour) et de dégradation des crottes (18 jours)
déterminées par Koster et Hart (1988) en forêt dense de la
République Démocratique du Congo, nous obtenons une
densité moyenne de 0,634 individus/km2, variant entre les
valeurs 0,278 et 1,448 individus/km2.
4.1.1.3 Comparaison des densités du céphalophe
bleu entre les trois sites (Inventaires diurnes indirects)
La comparaison des densités moyennes entre les trois
sites a été réalisée sur la base des
densités obtenues au travers de DISTANCE 6.0. De l'analyse des variances
des densités de Céphalophe bleu, il en découle qu'il n'y a
pas de différence significative entre les trois sites (one way
ANOVA : F2;15= 2,2271; p = 0,18905) (Figure
15).

Figure 12: Comparaison des
densités des Céphalophe bleus entre les trois sites (Inventaires
diurnes indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.4 Estimation des densités de Céphalophe
roux entre les trois sites (Inventaires diurnes indirects)
4.1.1.4.1
Céphalophe roux à Gribé
Les céphalophes roux rencontrés dans la
région du Sud-est Cameroun regroupent trois espèces dont
Cephalophus callipygus, C. dorsalis et C. nigrifons
dont leurs signes d'activités sont difficilement différentiables
(Bobo, 2009).
L'estimation de la densité de Céphalophe roux
s'est faite sur la base des crottes. Au total, 22 crottes ont été
relevées sur les six transects parcourus. La distance perpendiculaire
maximale au transect est de 1030 cm. Le tableau 7 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 16 illustre bien l'allure de la courbe
de probabilité de détection des crottes en fonction de la
distance perpendiculaire au transect.
Tableau 7: Statistique d'estimation de la
densité de Céphalophe roux à Gribé (Inventaires
diurnes indirects)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
188,07
|
|
|
|
|
f(0)
|
0,41237E-02
|
0,96837E-03
|
23,48
|
0,25433E-02
|
0,66861E-02
|
|
p
|
0,23544
|
0,55288E-01
|
23,48
|
0,14521
|
0,38174
|
|
ESW
|
242,50
|
56,946
|
23,48
|
149,56
|
393,19
|
|
n/L
|
0,61111
|
|
31,41
|
0,31990
|
1,1674
|
|
D
|
126,00
|
49,420
|
39,22
|
58,345
|
272,11
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des crottes
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen de crottes par unité de distance du
transect en km,
D = densité des crottes (nombre/km2)
AIC = Akaike Information Criterion

Figure 13: Courbe de
probabilité de détection des crottes de Céphalophe roux
à Gribé en fonction de la distance perpendiculaire au transect
(Inventaires diurnes indirects)
Au travers de DISTANCE, nous obtenons une densité
moyenne de 126,00 crottes/km2, avec un coefficient de variation de
39,22%: ce qui est acceptable, mais légèrement supérieur
à 25%.
En utilisant les valeurs du taux de défécation
(4,4/jour) et de dégradation des crottes (21 jours)
déterminées par Koster et Hart (1988) en forêt dense de la
République Démocratique du Congo, nous obtenons une
densité moyenne de 1,364 individu/km², variant entre les valeurs
0,632 et 2,945 individu/km².
4.1.1.4.2
Céphalophe roux à Gouonepoun
L'estimation de la densité de Céphalophe roux
s'est faite sur la base des crottes. Au total, neuf crottes ont
été relevées sur les six transects parcourus. La distance
perpendiculaire maximale au transect est de 500 cm. Le tableau 8
récapitule les résultats statistiques de l'estimation de la
densité obtenus au travers du programme DISTANCE. La figure 17 illustre
bien l'allure de la courbe de probabilité de détection des
crottes en fonction de la distance perpendiculaire au transect.
Tableau 8: Statistique d'estimation de la
densité de Céphalophe roux à Gouonepoun (Inventaires
diurnes indirects)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
110,97
|
|
|
|
|
f(0)
|
0,55046E-02
|
0,18061E-02
|
32,81
|
0,25844E-02
|
0,11725E-01
|
|
p
|
0,36333
|
0,11921
|
32,81
|
0,17058
|
0,77388
|
|
ESW
|
181,66
|
59,606
|
32,81
|
85,290
|
386,94
|
|
n/L
|
0,25000
|
|
46,44
|
0,98401E-01
|
0,63515
|
|
D
|
68,808
|
39,126
|
56,86
|
23,067
|
205,2
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des crottes
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen de crottes par unité de distance du
transect en km,
D = densité des crottes (nombre/km2)
AIC = Akaike Information Criterion


Figure 14: Courbe de
probabilité de détection des crottes de Céphalophe roux
à Gouonepoun en fonction de la distance perpendiculaire au transect
(Inventaires diurnes indirects)
Au travers de DISTANCE, nous obtenons une densité
moyenne de 68.808 crottes/km2, avec un coefficient de variation de
56.86%: ce qui est acceptable, mais légèrement supérieur
à 25%.
En utilisant les valeurs du taux de défécation
(4,4/jour) et de dégradation des crottes (21 jours)
déterminées par Koster et Hart (1988) en forêt dense de la
République Démocratique du Congo, nous obtenons une
densité moyenne de 0,745 individu/km², variant entre les valeurs
0,250 et 2,221 individu/km².
4.1.1.4.3
Céphalophe roux dans le PNBB
L'estimation de la densité de Céphalophe roux
s'est faite sur la base des crottes. Au total, 22 crottes ont été
relevées sur les six transects parcourus. La distance perpendiculaire
maximale au transect est de 1200 cm. Le tableau 9 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 18 illustre bien l'allure de la courbe
de probabilité de détection des crottes en fonction de la
distance perpendiculaire au transect.
Tableau 9: Statistique d'estimation de la
densité de Céphalophe roux dans au PNBB (Inventaires diurnes
indirects)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
298,77
|
|
|
|
|
f(0)
|
0,25303E-02
|
0,46008E-03
|
18,18
|
0,17369E-02
|
0,36860E-02
|
|
p
|
0,32934
|
0,59885E-01
|
18,18
|
0,22608
|
0,47977
|
|
ESW
|
395,21
|
71,862
|
18,18
|
271,30
|
575,73
|
|
n/L
|
0,61111
|
|
23,44
|
0,37517
|
0,99544
|
|
D
|
77,314
|
22,935
|
29,66
|
42,831
|
139,56
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des crottes
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen de crottes par unité de distance du
transect en km,
D = densité des crottes (nombre/km2)
AIC = Akaike Information Criterion

Figure 15: Courbe de
probabilité de détection des crottes de Céphalophe roux
dans le PNBB en fonction de la distance perpendiculaire au transect
(Inventaires diurnes indirects)
Au travers de DISTANCE, nous obtenons une densité
moyenne de 77,314 crottes/km2, avec un coefficient de variation de
29,66%: ce qui est acceptable, mais légèrement supérieur
à 25%.
En utilisant les valeurs du taux de défécation
(4,4/jour) et de dégradation des crottes (21 jours)
déterminées par Koster et Hart (1988) en forêt dense de la
République Démocratique du Congo, nous obtenons une
densité moyenne de 0,837 individu/km², variant entre les valeurs
0,464 et 1,510 individu/km².
4.1.1.5 Comparaison des densités de Céphalophe
roux entre les trois sites (Inventaires diurnes indirects)
La comparaison des densités moyennes des individus
entre les trois sites a été réalisée sur la base
des densités obtenues au travers de DISTANCE 6.0. De l'analyse des
variances des densités moyennes de Céphalophe roux, il en
découle qu'il n'y a pas de différence significative entre les
trois sites (one way ANOVA: F2;15 = 0,46865; p =
0,64696) (Figure 19).

Figure 16: Comparaison des
densités de Céphalophe roux entre les trois sites (Inventaires
diurnes indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.6 Estimation de l'abondance relative des espèces
entre les trois sites (Inventaires diurnes indirects)
A partir des données recensées dans les trois
sites, nous avons dressé le tableau 10 qui récapitule les IKA
moyens des espèces recensées par site dans la zone
d'étude.
Tableau 10: IKA moyen des espèces
recensées par site dans la zone d'étude (inventaires diurnes
indirects)
|
Espèce
|
Gribé
|
Gouonepoun
|
PNBB
|
|
Total
|
|
n
|
IKA moyen
|
n
|
IKA moyen
|
n
|
IKA moyen
|
n
|
IKA total
|
|
Bongo
|
15
|
0,417#177;0,705
|
29
|
0,806#177;0,826
|
28
|
0,778#177;0,292
|
72
|
2,001#177;1,823
|
|
Buffle
|
34
|
0,944#177;0,917
|
0
|
0,0
|
13
|
0,361#177;0,068
|
47
|
1,305#177;0,985
|
|
Céphalophe à dos jaune
|
59
|
1,639#177;1,092
|
37
|
1,028#177;0,627
|
87
|
2,417#177;0,376
|
183
|
5,084#177;2,095
|
|
Céphalophe bleu
|
373
|
10,361#177;2,489
|
357
|
9,917#177;1,797
|
298
|
8,278#177;1,566
|
1028
|
28,556#177;5,852
|
|
Céphalophe roux
|
376
|
10,444#177;2,736
|
457
|
12,694#177;2,388
|
390
|
10,833#177;2,532
|
1223
|
33,971#177;7,656
|
|
Chat sauvage
|
5
|
0,139#177;0,068
|
2
|
0,056#177;0,086
|
1
|
0,028#177;0,068
|
8
|
0,223#177;0,222
|
|
Chevrotain aquatique.
|
3
|
0,083#177;0,139
|
1
|
0,028#177;0,068
|
4
|
0,111#177;0,136
|
8
|
0,222#177;0,343
|
|
Chimpanzé
|
15
|
0,417#177;0,376
|
5
|
0,139#177;0,125
|
23
|
0,639#177;0,645
|
43
|
1,195#177;1,146
|
|
Civette
|
5
|
0,139#177;0,195
|
0
|
0,0
|
0
|
0,0
|
5
|
0,139#177;0,195
|
|
Eléphant
|
94
|
2,611#177;3,117
|
15
|
0,417#177;0,274
|
87
|
2,417#177;1,177
|
196
|
5,445#177;4,568
|
|
Espèce de petits singes
|
16
|
0,444#177;0,344
|
5
|
0,306#177;0,195
|
5
|
0,167#177;0,149
|
26
|
0,917#177;0,688
|
|
Antilope de Bates
|
17
|
0,472#177;0,464
|
3
|
0,083#177;0,091
|
16
|
0,444#177;0,228
|
36
|
0,999#177;0,783
|
|
Gorille
|
55
|
1,528#177;1,062
|
43
|
1,194#177;0,627
|
70
|
1,944#177;1,104
|
168
|
4,666#177;2,793
|
|
Oryctérope
|
0
|
0,0
|
0
|
0,0
|
8
|
0,222#177;0,344
|
8
|
0,222#177;0,344
|
|
Pangolin à longue queue
|
152
|
4,222#177;1,219
|
53
|
1,472#177;1,199
|
110
|
3,056#177;1,608
|
315
|
8,75#177;4,026
|
|
Pangolin géant
|
16
|
0,444#177;0,344
|
16
|
0,444#177;0,502
|
68
|
1,889#177;1,015
|
100
|
2,777#177;1,861
|
|
Léopard
|
2
|
0,056#177;0,086
|
0
|
0,0
|
0
|
0,0
|
2
|
0,056#177;0,086
|
|
Porc épic
|
236
|
6,556#177;3,489
|
161
|
4,472#177;1,035
|
229
|
6,361#177;2,101
|
626
|
17,389#177;6,625
|
|
Potamochère
|
108
|
3#177;2,060
|
111
|
3,083#177;1,718
|
181
|
5,028#177;2,758
|
400
|
11,111#177;6,536
|
|
Sitatunga
|
18
|
0,5#177;0,548
|
5
|
0,139#177;0,164
|
13
|
0,361#177;0,245
|
36
|
1#177;0,957
|
|
TOTAL
|
1599
|
44,022#177;21,451
|
1300
|
36,008#177;11,725
|
1631
|
46,078#177;16,412
|
4529
|
126,108#177;49,584
|
n= nombre total d'indice observé
A partir du tableau 10 nous remarquons que le Parc National de
Boumba-Bek présente le plus grand nombre d'indice soit un total de 1630
signes d'activités des grands et moyens mammifères (IKA =
46,078#177;16,412) suivis, par le site de Gribé (IKA =
44,022#177;21,451). Tandis que le site de Gouonepoun situé en
forêt primaire dont les activités sont absentes, présente
les signes d'activités les plus faibles (IKA = 36,008#177;11,725). Ce
dernier s'expliquerait notamment par la forte pression anthropique
exercée sur la faune notamment le braconnage, qui aurait sans doute
entrainée la migration des animaux vers les endroits de la forêt
où les activités humaines n'ont pas encore un impact très poussé comme le Parc.
4.1.1.6.1
Céphalophe bleu Cephalophus monticola
Les signes d'activité de Céphalophe bleu
ont été observés dans toute la zone d'étude.
Le tableau 11 récapitule les IKA moyens du céphalophe bleu par
signe d'activité et par âge dans les trois sites.
Tableau 11: IKA Céphalophe Bleu par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
3
|
0,083
|
|
crottes
|
11
|
0,306
|
4
|
0,111
|
18
|
0,500
|
13
|
0,361
|
7
|
0,194
|
53
|
1,472
|
|
empreinte
|
45
|
1,250
|
94
|
2,611
|
47
|
1,306
|
5
|
0,139
|
0
|
0
|
191
|
5,306
|
|
piste
|
0
|
0
|
3
|
0,083
|
117
|
3,250
|
0
|
0
|
6
|
0,167
|
126
|
3,500
|
|
Global Gribé
|
56
|
1,556
|
102
|
2,833
|
183
|
5,084
|
18
|
0,500
|
14
|
0,389
|
373
|
10,361
|
|
couchage
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
Gouonepoun
|
crottes
|
6
|
0,167
|
4
|
0,111
|
5
|
0,139
|
3
|
0,083
|
0
|
0
|
18
|
0,500
|
|
empreinte
|
27
|
0,750
|
41
|
1,139
|
9
|
0,250
|
0
|
0
|
0
|
0
|
77
|
2,139
|
|
piste
|
1
|
0,028
|
0
|
0,028
|
257
|
7,139
|
0
|
0
|
1
|
0,028
|
260
|
7,223
|
|
Global Gouonepoun
|
35
|
0,972
|
46
|
1,278
|
272
|
7,556
|
3
|
0,083
|
1
|
0,028
|
357
|
9,917
|
|
couchage
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
crottes
|
1
|
0,028
|
4
|
0,111
|
6
|
0,167
|
2
|
0,056
|
1
|
0,028
|
14
|
0,389
|
|
PNBB
|
empreinte
|
19
|
0,528
|
45
|
1,250
|
75
|
2,083
|
16
|
0,444
|
5
|
0,139
|
160
|
4,444
|
|
piste
|
0
|
0
|
1
|
0,028
|
96
|
2,667
|
0
|
0
|
26
|
0,722
|
123
|
3,417
|
|
Global PNBB
|
20
|
0,556
|
50
|
1,389
|
178
|
4,945
|
18
|
0,500
|
32
|
0,889
|
298
|
8,278
|
Il en
ressort que la majorité des observations est constituée de pistes
où l'animal passe régulièrement et des empreintes frais et
récents. Ainsi, le site de Gribé est le plus abondant en indice
de présence de céphalophe bleu (IKA = 10,361), ceci
s'expliquerait par le fait le site a été repeuplé
immédiatement suite à une exploitation forestière quelques
années bien avant où le calme est de retour.
4.1.1.7 Comparaison des IKA du Céphalophe bleu entre
les trois sites (Inventaires diurnes indirects)
Sur l'ensemble des trois sites, ont été
enregistrés 1028 signes de présence de céphalophe bleu
dont 373 à Gribé (10,361#177;2,489), 357 à Gouonepoun (IKA
de 9,917#177;1,797) et 298 dans le PNBB (IKA de 8,278#177;1,566) (Tableau 11).
De l'analyse des variances des IKA des céphalophes bleus, il en
découle qu'il n'y a pas de différence significative entre les
trois sites (one way ANOVA: F2;15 = 1,8246; p =
0,19531) (Figure 20).

Figure 17: Comparaison des
IKA du Céphalophe bleu entre les trois sites (Inventaires diurnes
indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.7.1
Céphalophe roux
Le tableau 12 récapitule les IKA moyen de
Céphalophe roux par signe d'activité et par âge dans les
trois sites d'étude afin de voir si ceux-ci sont significativement
différents ou non.
Tableau 12: IKA Céphalophe roux par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
couchage
|
0
|
0
|
2
|
0,056
|
2
|
0,056
|
0
|
0
|
1
|
0,028
|
5
|
0,139
|
|
crottes
|
3
|
0,083
|
2
|
0,056
|
7
|
0,194
|
6
|
0,167
|
4
|
0,111
|
22
|
0,611
|
|
empreinte
|
37
|
1,028
|
98
|
2,722
|
68
|
1,889
|
11
|
0,306
|
9
|
0,250
|
223
|
6,194
|
|
piste
|
0
|
0
|
0
|
0
|
107
|
2,973
|
2
|
0,056
|
17
|
0,472
|
126
|
3,500
|
|
Global Gribé
|
40
|
1,111
|
102
|
2,833
|
184
|
5,111
|
19
|
0,528
|
31
|
0,861
|
376
|
10,444
|
|
Gouonepoun
|
couchage
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
crottes
|
0
|
0
|
1
|
0,028
|
2
|
0,056
|
3
|
0,083
|
3
|
0,083
|
9
|
0,250
|
|
empreinte
|
31
|
0,861
|
73
|
2,028
|
31
|
0,861
|
0
|
0
|
0
|
0
|
135
|
3,750
|
|
piste
|
0
|
0
|
2
|
0,056
|
305
|
8,472
|
0
|
0
|
4
|
0,111
|
311
|
8,639
|
|
Global Gouonepoun
|
31
|
0,861
|
77
|
2,139
|
339
|
9,417
|
3
|
0,083
|
7
|
0,194
|
457
|
12,694
|
|
PNBB
|
couchage
|
2
|
0,056
|
2
|
0,056
|
1
|
0,028
|
0
|
0
|
0
|
0
|
5
|
0,139
|
|
crottes
|
5
|
0,139
|
8
|
0,222
|
4
|
0,111
|
3
|
0,083
|
2
|
0,056
|
22
|
0,611
|
|
empreinte
|
15
|
0,417
|
63
|
1,750
|
66
|
1,833
|
22
|
0,611
|
11
|
0,306
|
177
|
4,917
|
|
piste
|
0
|
0
|
0
|
0
|
130
|
3,611
|
0
|
0
|
56
|
1,556
|
186
|
5,167
|
|
Global PNBB
|
22
|
0,612
|
73
|
2,028
|
201
|
5,583
|
25
|
0,694
|
69
|
1,917
|
390
|
10,833
|
Il en
ressort que la majorité des observations est constituée de pistes
où l'animal passe régulièrement et des empreintes frais et
récents. Ainsi, le site de Gouonepoun est le plus abondant en indice de
présence de céphalophe roux (IKA = 12,694), ceci s'expliquerait
par le fait le site de Gouonepoun est situé en forêt primaire
où les activités humaines ont un impact relativement faible sur
la survie des animaux.
4.1.1.8 Comparaison des IKA du Céphalophe roux entre
les trois sites (Inventaires diurnes indirects)
Sur l'ensemble des trois sites, ont été
enregistrés 1613 signes de présence de céphalophe roux
dont 376 signes à Gribé (IKA de 10,444#177;2,736), de 457 signes
à Gouonepoun (IKA de 12,694#177;2,388) et de 390 signes dans le PNBB
(IKA de 10,833#177;2,532) (Tableau 12). De l'analyse des variances des IKA des
céphalophes roux, il en découle qu'il n'y a pas de
différence significative entre les trois sites (one way ANOVA:
F2;15 = 1,3283; p = 0,29436) (Figure 21).
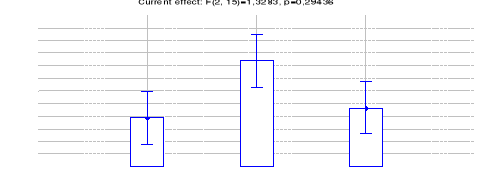
Figure 18: Comparaison des
IKA du Céphalophe roux entre les trois sites (Inventaires diurnes
indirects)
4.1.1.8.1
Céphalophe à dos jaune Cephalophus silvicultor
Très peu de crottes ont été
relevées sur les 18 transects permanents parcourus dans la zone
d'étude, avec seulement trois à Gribé, deux à
Gouonepoun et six dans le PNBB, rendant impossible l'estimation de la
densité pour cette espèce. Le tableau 13 récapitule les
IKA de Céphalophe à dos jaune par signe d'activité et par
âge dans les trois sites d'étude.
Tableau 13: IKA Céphalophe à dos jaune
par signe d'activité et par âge dans les trois sites
d'étude (Inventaires diurnes indirects)
|
Sites
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
couchage
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
crottes
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
3
|
0,083
|
|
empreinte
|
5
|
0,139
|
16
|
0,444
|
22
|
0,611
|
2
|
0,056
|
6
|
0,167
|
51
|
1,417
|
|
piste
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
3
|
0,083
|
4
|
0,111
|
|
Global Gribé
|
6
|
0,167
|
17
|
0,472
|
23
|
0,639
|
3
|
0,083
|
10
|
0,278
|
59
|
1,639
|
|
Gouonepoun
|
crottes
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
2
|
0,056
|
|
empreinte
|
3
|
0,083
|
13
|
0,361
|
16
|
0,444
|
1
|
0,028
|
1
|
0,028
|
34
|
0,944
|
|
traces
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gouonepoun
|
3
|
0,083
|
14
|
0,389
|
16
|
0,444
|
2
|
0,056
|
2
|
0,056
|
37
|
1,028
|
|
PNBB
|
alimentation
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
crottes
|
0
|
0
|
1
|
0,028
|
2
|
0,056
|
0
|
0
|
3
|
0,083
|
6
|
0,167
|
|
empreinte
|
1
|
0,028
|
15
|
0,417
|
20
|
0,556
|
17
|
0,472
|
19
|
0,528
|
72
|
2,000
|
|
piste
|
0
|
0
|
0
|
0
|
4
|
0,111
|
0
|
0
|
4
|
0,111
|
8
|
0,222
|
|
Global PNBB
|
2
|
0,056
|
16
|
0,444
|
26
|
0,722
|
17
|
0,472
|
26
|
0,722
|
87
|
2,417
|
Il en ressort que la majorité des observations est
constituée des empreintes récents. Cependant, le site de PNBB est
le plus abondant en indice de présence de céphalophe à dos
jaune, (IKA = 2,417), car pour la plupart, ces animaux ont migré dans le
Parc où toutes activités humaines (chasse, pêche et autres)
sont strictement interdite selon la loi camerounaise.
4.1.1.9 Comparaison des IKA du Céphalophe à dos
jaune entre les trois sites (Inventaires diurnes indirects)
Sur l'ensemble des trois sites, ont été
enregistrés 183 signes de présence de céphalophe à
dos jaune dont 59 signes à Gribé (IKA de 1,639#177;1,092), 37
signes à Gouonepoun (IKA de 1,028#177;0,627) et 87 signes dans le PNBB
(IKA de 2,417#177;0,376) (Tableau 13). De l'analyse des variances des IKA de
céphalophe à dos jaune, il en découle qu'il y a une
différence significative entre les trois sites (one way ANOVA:
F2;15= 5,0429; p = 0,02113) (Figure 22).
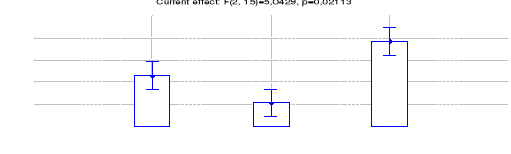
Figure 19: Comparaison des
IKA du Céphalophe à dos jaune entre les trois sites (Inventaires
diurnes indirects)
Notes : a, b: expression de la
différence significative au seuil de probabilité de 5%
Du test de Tukey de la séparation des moyennes, la
moyenne des IKA à Gribé est significativement différente
de celle de Gouonepoun (p = 0,016603) et de celle dans le PNBB
(p = 0,016603). La moyenne des IKA à Gouonepoun (p =
0,368663) n'est pas significativement différente de la moyenne des IKA
dans le PNBB (p = 0,211876).
4.1.1.9.1 Bongo
Tragelaphus euryceros
Le tableau 14 récapitule les IKA de Bongo par signe
d'activité et par âge dans les trois sites d'étude.
Tableau 14: IKA Bongo par signe d'activité et
par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
|
sites
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
crottes
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
empreinte
|
0
|
0
|
1
|
0,028
|
5
|
0,139
|
2
|
0,056
|
2
|
0,056
|
11
|
0,278
|
|
piste
|
0
|
0
|
0
|
0
|
4
|
0,112
|
0
|
0
|
0
|
0
|
4
|
0,112
|
|
Total Gribé
|
0
|
0
|
2
|
0,056
|
9
|
0,250
|
2
|
0,056
|
2
|
0,056
|
15
|
0,417
|
|
Gouone-
poun
|
crottes
|
2
|
0,056
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
empreinte
|
0
|
0
|
3
|
0,083
|
12
|
0,333
|
5
|
0,139
|
1
|
0,028
|
21
|
0,583
|
|
piste
|
0
|
0
|
0
|
0
|
5
|
0,139
|
0
|
0
|
0
|
0
|
5
|
0,139
|
|
Total Gouonepoun
|
2
|
0,056
|
3
|
0,083
|
1
|
0,500
|
5
|
0,139
|
1
|
0,028
|
29
|
0,806
|
|
PNBB
|
couchage
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
crottes
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
empreinte
|
0
|
0
|
4
|
0,111
|
8
|
0,222
|
1
|
0,028
|
9
|
0,250
|
22
|
0,611
|
|
piste
|
0
|
0
|
0
|
0
|
2
|
0,056
|
0
|
0
|
2
|
0,056
|
4
|
0,111
|
|
Total PNBB
|
0
|
0
|
5
|
0,139
|
10
|
0,278
|
1
|
0,028
|
12
|
0,333
|
28
|
0,778
|
Il en
ressort que la majorité des observations est constituée des
empreintes récents. Cependant, le site de Gouonepoun est le plus
abondant en indice de présence de bongo (IKA = 0,806).
4.1.1.10 Comparaison des IKA Bongo entre les trois sites
(Inventaires diurnes indirects)
Sur l'ensemble des trois sites, ont été
enregistrés 72 signes de présence de Bongo dont 15 signes
à Gribé (IKA de 0,417#177;0,705), 29 signes à Gouonepoun
(IKA de 0,806#177;0,826) et 28 signes dans le PNBB (IKA de 0,778#177;0,292)
(Tableau 14). De l'analyse des variances, il en découle qu'il n'y a pas
de différence significative entre les trois sites (one way ANOVA:
F2;15 = 0,66997; p = 0,52639) (Figure 23).
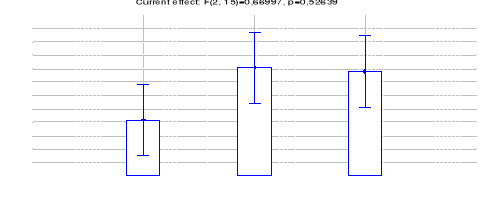
Figure 20: Comparaison des
IKA de Bongo entre les trois sites (Inventaires diurnes indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.10.1 Buffle Syncerus
cafer nanus
Le tableau 15 récapitule les IKA de Céphalophe
bleu par signe d'activité et par âge dans les trois sites
d'étude.
Tableau 15: IKA Buffle par signe d'activité et
par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très
vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
empreinte
|
1
|
0,028
|
6
|
0,167
|
9
|
0,250
|
5
|
0,139
|
6
|
0,167
|
27
|
0,750
|
|
piste
|
0
|
0
|
0
|
0
|
4
|
0,111
|
0
|
0
|
0
|
0
|
4
|
0,111
|
|
traces
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
0
|
0
|
3
|
0,083
|
|
Global Gribé
|
2
|
0,056
|
7
|
0,194
|
13
|
0,361
|
6
|
0,167
|
6
|
0,167
|
34
|
0,944
|
|
Gouonepoun
|
/
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Global Gouonepoun
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
PNBB
|
couchage
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
crottes
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
|
empreinte
|
0
|
0
|
0
|
0
|
5
|
0,139
|
1
|
0,028
|
2
|
0,056
|
8
|
0,222
|
|
piste
|
0
|
0
|
0
|
0
|
2
|
0,056
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
Global PNBB
|
0
|
0
|
0
|
0
|
9
|
0,250
|
2
|
0,056
|
2
|
0,056
|
13
|
0,361
|
Il ressort du tableau 15, que aucun signe de buffle n'a
été observé sur les transects à Gouonepoun rendant
impossible la comparaison des moyennes des IKA entre les trois sites.
4.1.1.10.2 Espèce
de Petits singes
Les petits singes relevés regroupent six espèces
notamment, le Hocheur Cercopithecus nictitans ; le
Cercocèbe agile Cercocebus agilis, le Cercopithèque
pogonias, Cercopithecus pogonias, le Colobe Gureza Colobus
guereza le Cercocèbe à joues grises Cercocebus albigena,
le Moustac Cercopithecus cephus. Le tableau 16 récapitule
les IKA des petits singes par signe d'activité et par âge dans
les trois sites d'étude
Tableau 16: IKA petits singes par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Sites
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
2
|
0,056
|
6
|
0,167
|
2
|
0,056
|
|
0
|
1
|
0,028
|
11
|
0,306
|
|
crottes
|
0
|
0
|
3
|
0,083
|
1
|
0,028
|
|
0
|
1
|
0,028
|
5
|
0,139
|
|
Total Gribé
|
2
|
0,056
|
9
|
0,250
|
3
|
0,083
|
0
|
0
|
2
|
0,056
|
16
|
0,444
|
|
Gouonepoun
|
alimentation
|
4
|
0,111
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
0
|
0
|
6
|
0,167
|
|
crâne
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
|
crottes
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
empreinte
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Total Gouonepoun
|
6
|
0,167
|
1
|
0,028
|
2
|
0,056
|
2
|
0,056
|
0
|
0
|
11
|
0,306
|
|
PNBB
|
alimentation
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
crottes
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
2
|
0,056
|
|
piste
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
2
|
0,028
|
|
Total PNBB
|
2
|
0,056
|
1
|
0,028
|
1
|
0,028
|
1
|
0,028
|
1
|
0,028
|
6
|
0,167
|
Il
ressort du tableau 16 que très peu de signes d'observation des petits
singes ont été enregistrés pour cette espèce,
rendant impossible la comparaison des IKA entre les trois sites.
4.1.1.10.3 Chat sauvage
Nandinia binotata
Les signes d'activité de Chat sauvage ont
été enregistrés dans tous les sites d'étude. Le
tableau 17 récapitule les IKA de chat sauvage par signe
d'activité et par âge dans les trois sites d'étude.
Tableau 17: IKA Chat sauvage par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
crottes
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
empreinte
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
piste
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
|
terrier
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
Global Gribé
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
1
|
0,028
|
1
|
0,028
|
5
|
0,139
|
|
Gouonepoun
|
crottes
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
empreinte
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gouonepoun
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
PNBB
|
crottes
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
|
Global PNBB
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
Il
ressort du tableau 17 que très peu des signes d'observation de chat
sauvage ont été observés pour cette espèce, rendant
impossible la comparaison des IKA entre les trois sites
4.1.1.10.4 Chevrotain
aquatique Hyemoschus aquaticus
Le tableau 18 récapitule les IKA de Chevrotin aquatique
par signe d'activité et par âge dans les trois sites
d'étude
Tableau 18: IKA Chevrotain aquatique par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
piste
|
0
|
0
|
0
|
0
|
3
|
0,083
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
Global Gribé
|
0
|
0
|
0
|
0
|
3
|
0,083
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
Gouonepoun
|
empreinte
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gouonepoun
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
PNBB
|
empreinte
|
0
|
0
|
0
|
0
|
3
|
0,083
|
1
|
0,028
|
0
|
0
|
4
|
0,111
|
|
Global PNBB
|
0
|
0
|
0
|
0
|
3
|
0,083
|
1
|
0,028
|
0
|
0
|
4
|
0,111
|
Il ressort du tableau 18 que très peu de signes
d'observation de chevrotain aquatique ont été enregistrés
pour cette espèce, rendant impossible la comparaison des IKA entre les
trois sites.
4.1.1.10.5 Civette
Civettictis civetta
Le tableau 19 récapitule les IKA des Civettes par signe
d'activité et par âge dans les trois sites d'étude.
Tableau 19: IKA Civette par signe d'activité et
par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
crottes
|
1
|
0,028
|
1
|
0,028
|
2
|
0,056
|
|
empreinte
|
0
|
0
|
2
|
0,056
|
2
|
0,056
|
|
Global Gribé
|
1
|
0,028
|
4
|
0,111
|
5
|
0,139
|
|
Gouonepoun
|
/
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Global Gouonepoun
|
0
|
0
|
0
|
0
|
0
|
0
|
|
PNBB
|
/
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Global PNBB
|
0
|
0
|
0
|
0
|
0
|
0
|
Il ressort du tableau 19 que seule la civette a
été observée à Gribé, rendant impossible la
comparaison des IKA de cette espèce entre les trois sites.
4.1.1.10.6
Chimpanzé Pan troglodites
Le tableau 20 récapitule les IKA de Chimpanzé
par signe d'activité et par âge dans les trois sites
d'étude.
Tableau 20: IKA Chimpanzé par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
repos
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
3
|
0,084
|
|
crottes
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
empreinte
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
3
|
0,083
|
|
nid
|
0
|
0
|
1
|
0,028
|
6
|
0,167
|
0
|
0
|
0
|
0
|
7
|
0,195
|
|
piste
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gribé
|
1
|
0,028
|
2
|
0,056
|
9
|
0,250
|
0
|
0
|
3
|
0,083
|
15
|
0,417
|
|
Gouonepoun
|
empreinte
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
nid
|
0
|
0
|
0
|
0
|
0
|
0
|
3
|
0,084
|
0
|
0
|
3
|
0,084
|
|
Global Gouonepoun
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
3
|
0,083
|
0
|
0
|
5
|
0,139
|
|
PNBB
|
alimentation
|
0
|
0
|
5
|
0,139
|
2
|
0,056
|
3
|
0,083
|
2
|
0,056
|
12
|
0,333
|
|
crottes
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
3
|
0,083
|
|
empreinte
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
|
nid
|
0
|
0
|
1
|
0,028
|
3
|
0,083
|
0
|
0
|
0
|
0
|
4
|
0,111
|
|
traces
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
3
|
0,084
|
|
Global PNBB
|
1
|
0,028
|
8
|
0,222
|
5
|
0,139
|
5
|
0,139
|
4
|
0,111
|
23
|
0,639
|
Il
ressort du tableau 20 que très peu de signes d'observation de
chimpanzé ont été enregistrés rendant impossible la
comparaison des IKA entre les trois sites.
4.1.1.10.7 Eléphant
Loxodonta africana cyclotis
Le tableau 21 récapitule les IKA de
l'éléphant par signe d'activité et par âge dans les
trois sites d'étude.
Tableau 21: IKA Eléphant par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
2
|
0,056
|
|
crottes
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
4
|
0,111
|
10
|
0,278
|
16
|
0,444
|
|
piste
|
1
|
0,028
|
3
|
0,083
|
19
|
0,528
|
11
|
0,306
|
42
|
1,166
|
76
|
2,111
|
|
Global Gribé
|
1
|
0,028
|
4
|
0,111
|
20
|
0,556
|
15
|
0,417
|
54
|
1,500
|
94
|
2,611
|
|
Gouonepoun
|
empreinte
|
0
|
0
|
0
|
0
|
1
|
0,028
|
2
|
0,056
|
8
|
0,222
|
11
|
0,306
|
|
piste
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
4
|
0111
|
|
Global Gouonepoun
|
0
|
0
|
1
|
0,028
|
2
|
0,056
|
2
|
0,056
|
10
|
0,278
|
15
|
0,417
|
|
PNBB
|
alimentation
|
0
|
0
|
0
|
0
|
4
|
0,111
|
0
|
0
|
5
|
0,139
|
9
|
0,250
|
|
crottes
|
0
|
0
|
0
|
0
|
1
|
0,028
|
2
|
0,056
|
5
|
0,139
|
8
|
0,222
|
|
empreinte
|
1
|
0,028
|
2
|
0,056
|
4
|
0,111
|
1
|
0,028
|
26
|
0,722
|
34
|
0,944
|
|
nettoyage
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
2
|
0,056
|
|
piste
|
0
|
0
|
1
|
0,028
|
8
|
0,222
|
0
|
0
|
25
|
0,694
|
34
|
0,945
|
|
Global PNBB
|
1
|
0,028
|
4
|
0,111
|
17
|
0,472
|
3
|
0,083
|
62
|
1,722
|
87
|
2,417
|
Il en ressort que la majorité des observations
est constituée des piste utilisées fréquemment par des
éléphants. Ces pistes sont pour la plupart constituées des
indices vieux. Le site de Gribé est le plus abondant en indice soit un
IKA de 2,611. Ceci s'expliquerait par le fait que l'on rencontre abondamment
des parasoleiliers qui sont consommés par ces derniers.
4.1.1.11 Comparaison des IKA Eléphant entre les trois
sites (Inventaires diurnes indirects)
Dans l'ensemble des trois sites, ont été
relevés 196 signes de présence des éléphants dont
94 signes à Gribé (IKA de 2,611#177;3,117), 15 signes à
Gouonepoun (IKA de 0,417#177;0,274) et 87 signes dans le PNBB (IKA de
2,417#177;1,177) (Tableau 21). De l'analyse des variances des IKA de
l'éléphant, il en découle qu'il n'y a pas de
différence significative entre les trois sites (one way ANOVA:
F2;15 = 2,3755; p = 0,12698) (Figure 24).

Figure 21: Comparaison IKA
éléphants entre les trois sites (Inventaires diurnes
indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.11.1 Antilope de
Bates Neotragus batesi
Le tableau 22 récapitule les IKA Antilope de Bates par
signe d'activité et par âge dans les trois sites d'étude.
Tableau 22: IKA Antilope de Bates par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
village
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
empreinte
|
3
|
0,083
|
9
|
0,250
|
2
|
0,056
|
0
|
0
|
14
|
0,389
|
|
piste
|
0
|
0
|
0
|
0
|
2
|
0,056
|
0
|
0
|
2
|
0,056
|
|
Global Gribé
|
3
|
0,083
|
10
|
0,278
|
4
|
0,111
|
0
|
0
|
17
|
0,472
|
|
Gouonepoun
|
empreinte
|
2
|
0,056
|
1
|
0,028
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
Global Gouonepoun
|
2
|
0,056
|
1
|
0,028
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
PNBB
|
empreinte
|
2
|
0,056
|
8
|
0,222
|
2
|
0,056
|
4
|
0,111
|
16
|
0,444
|
|
Global PNBB
|
2
|
0,056
|
8
|
0,222
|
2
|
0,056
|
4
|
0,111
|
16
|
0,444
|
Il ressort du tableau 12 que très peu de signes
d'observation ont été enregistrés pour l'antilope de Bates
rendant impossible la comparaison des IKA de cette espèce.
4.1.1.11.2 Gorille Gorilla
g, gorilla
Le tableau 23 récapitule les IKA des Gorilles signe
d'activité et par âge d'indice dans les trois sites
d'étude.
Tableau 23: IKA Gorille par signe d'activité et
par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Pourri
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
5
|
0,139
|
5
|
0,139
|
5
|
0,139
|
0
|
0
|
6
|
0,167
|
0
|
0
|
21
|
0,583
|
|
crottes
|
0
|
0
|
0
|
0
|
2
|
0,056
|
0
|
0
|
7
|
0,194
|
0
|
0
|
9
|
0,250
|
|
nid au sol
|
0
|
0
|
0
|
0
|
2
|
0,056
|
8
|
0,222
|
0
|
0
|
0
|
0
|
10
|
0,278
|
|
Piste
|
1
|
0,028
|
8
|
0,222
|
3
|
0,084
|
0
|
0
|
3
|
0,084
|
0
|
0
|
15
|
0417
|
|
Global Gribé
|
6
|
0,167
|
13
|
0,361
|
12
|
0,333
|
8
|
0,222
|
16
|
0,444
|
0
|
0
|
55
|
1,528
|
|
Gouonepoun
|
alimentation
|
0
|
0
|
8
|
0,222
|
17
|
0,472
|
0
|
0
|
5
|
0,139
|
0
|
0
|
30
|
0,833
|
|
crottes
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
|
nid au sol
|
0
|
0
|
0
|
0
|
0
|
0
|
3
|
0,083
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
Piste
|
1
|
0,028
|
1
|
0,028
|
6
|
0,116
|
0
|
0
|
0
|
0
|
0
|
0
|
8
|
0,222
|
|
Global Gouonepoun
|
1
|
0,028
|
9
|
0,250
|
24
|
0,667
|
3
|
0,083
|
6
|
0,167
|
0
|
0
|
43
|
1,194
|
|
PNBB
|
alimentation
|
3
|
0,083
|
6
|
0,167
|
13
|
0,361
|
6
|
0,167
|
15
|
0,417
|
0
|
0
|
43
|
1,194
|
|
crottes
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
0
|
0
|
4
|
0,111
|
|
nid au sol
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
8
|
0,222
|
0
|
0
|
6
|
0,167
|
16
|
0,444
|
|
Piste
|
0
|
0
|
2
|
0,056
|
2
|
0,056
|
1
|
0,028
|
2
|
0,056
|
0
|
0
|
7
|
0,194
|
|
Global PNBB
|
3
|
0,083
|
10
|
0,278
|
17
|
0,472
|
15
|
0,417
|
19
|
0,528
|
6
|
0,167
|
70
|
1,944
|
Il ressort du tableau 23 que 168 indices de
présence ont été relevés le long de 18 transects
parcourus. La plupart des observations est constituée des
récentes traces d'alimentation de l'espèce.
4.1.1.12 Comparaison des IKA Gorilles entre les trois sites
(Inventaires diurnes indirects)
Dans l'ensemble des trois sites, ont été
enregistrés 168 signes de présence de gorilles dont 55 signes
à Gribé (IKA de 1,528#177;1,062), 43 signes à Gouonepoun
(IKA de 1,194#177;0,627) et 70 signes dans le PNBB (IKA de 1,944#177;1,104)
(Tableau 23). De l'analyse des variances des IKA, il en découle qu'il
n'y a pas de différence significative entre les trois sites (one way
ANOVA: F2;15 = 0,92794; p = 0,41692) (Figure
25).

Figure 22: Comparaison des
IKA Gorilles entre les trois sites (Inventaires diurnes indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.12.1
Oryctérope Orycteropus afer
Le tableau 24 récapitule les IKA de l'oryctérope
par signe d'activité et par âge dans les trois sites
d'étude.
Tableau 24: IKA Oryctérope par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Récent
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
/
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Global Gribé
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Gouonepoun
|
/
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Global Gouonepoun
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
PNBB
|
alimentation
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
empreinte
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
|
terrier
|
3
|
0,083
|
1
|
0,028
|
2
|
0,056
|
6
|
0,167
|
|
Global PNBB
|
3
|
0,083
|
2
|
0,056
|
3
|
0,083
|
8
|
0,222
|
Il ressort du tableau 24 que seul l'oryctérope a
été enregistré dans le PNBB, rendant impossible la
comparaison des IKA entre les trois sites.
4.1.1.12.2 Pangolin
à longue queue Manis tetradactyla
Le tableau 25 récapitule les IKA de pangolin à
longue queue par signe d'activité et par âge dans les trois sites
d'étude.
Tableau 25: IKA Pangolin à longue queue par
signe d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
38
|
1,056
|
48
|
1,333
|
34
|
0,944
|
10
|
0,278
|
2
|
0,056
|
132
|
3,667
|
|
piste
|
0
|
0
|
2
|
0,056
|
17
|
0,473
|
0
|
0
|
0
|
0
|
19
|
0,529
|
|
terrier
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gribé
|
39
|
1,083
|
50
|
1,389
|
51
|
1,417
|
10
|
0,278
|
2
|
0,056
|
152
|
4,222
|
|
Gouonepoun
|
alimentation
|
8
|
0,222
|
21
|
0,583
|
17
|
0,472
|
1
|
0,028
|
3
|
0,083
|
50
|
1,389
|
|
piste
|
0
|
0
|
1
|
0,028
|
2
|
0,056
|
0
|
0
|
0
|
0
|
3
|
0,084
|
|
Global Gouonepoun
|
8
|
0,222
|
22
|
0,611
|
19
|
0,528
|
1
|
0,028
|
3
|
0,083
|
53
|
1,472
|
|
PNBB
|
alimentation
|
4
|
0,111
|
29
|
0,806
|
45
|
1,250
|
19
|
0,528
|
5
|
0,139
|
102
|
2,833
|
|
crottes
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
piste
|
0
|
0
|
0
|
0
|
4
|
0,111
|
0
|
0
|
1
|
0,028
|
5
|
0,139
|
|
terrier
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
2
|
0,056
|
|
Global PNBB
|
5
|
0,139
|
29
|
0,806
|
49
|
1,361
|
19
|
0,528
|
8
|
0,222
|
110
|
3,056
|
Il ressort du tableau 25 que 315 indices de
présences ont été relevés le long de 18 transects
parcourus. La plupart des observations est constituée des fraiches et
récentes traces d'alimentation de l'espèce.
4.1.1.13 Comparaison des IKA Pangolin à longue queue
entre les trois sites (Inventaires diurnes indirects)
Il en ressort que dans l'ensemble des trois sites, a
été enregistré un total de 315 signes de présence
des Pangolin à longue queue dont 152 signes à Gribé (IKA
de 4,222#177;1,219), 53 signes à Gouonepoun (IKA de 1,472#177;1,199) et
110 signes dans le PNBB (IKA de 3,056#177;1,608) (Tableau 25). De l'analyse des
variances il en découle qu'il y a une différence significatives
entre les trois sites (one way ANOVA: F2;15= 6,2274 ;
p = 0,01074) (Figure 26).

Figure 23: Comparaison des
IKA de pangolin à longue queue entre les trois sites (Inventaires
diurnes indirects)
Notes : a, b: expression de la
différence significative au seuil de probabilité de 5%
Il ressort de la figure 26 que les moyennes des IKA du
Pangolin à longue queue sont significativement différentes entre
les différents sites (one way ANOVA: F2;15= 6,2274;
p = 0,01074). Il ressort du test de Tukey de la séparation des
moyennes, que la moyenne des IKA à Gribé (p = 08361) est
significativement différente à celle de Gouonepoun (p =
0,140722), mais n'est pas significativement différent de celle du PNBB.
De même la moyenne des IKA à Gouonepoun (p = 08361) est
significativement différente de la moyenne des IKA dans le PNBB
(p = 0,140722).
4.1.1.13.1 Pangolin
géant Smutsia gigantea
Les signes d'activité de Pangolin géant ont
été observés dans toute la zone d'étude. Le tableau
26 récapitule les IKA de pangolin géant par signe
d'activité et par âge dans les trois sites d'étude.
Tableau 26: IKA Pangolin géant par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
village
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
1
|
0,028
|
3
|
0,083
|
0
|
0
|
4
|
0,111
|
8
|
0,222
|
16
|
0,444
|
|
Global Gribé
|
1
|
0,028
|
3
|
0,083
|
0
|
0
|
4
|
0,111
|
8
|
0,222
|
16
|
0,444
|
|
Gouonepoun
|
alimentation
|
0
|
0
|
4
|
0,111
|
4
|
0,111
|
3
|
0,083
|
5
|
0,139
|
16
|
0,444
|
|
Global Gouonepoun
|
0
|
0
|
4
|
0,111
|
4
|
0,111
|
3
|
0,083
|
5
|
0,139
|
16
|
0,444
|
|
PNBB
|
alimentation
|
0
|
0
|
9
|
0,250
|
16
|
0,444
|
16
|
0,444
|
22
|
0,611
|
63
|
1,750
|
|
empreinte
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
piste
|
0
|
0
|
0
|
0
|
3
|
0,083
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
Global PNBB
|
0
|
0
|
10
|
0,278
|
20
|
0,556
|
16
|
0,444
|
22
|
0,611
|
68
|
1,889
|
Il ressort du tableau 26 que 100 indices de
présence ont été relevés le long de 18 transects
parcourus. La plupart des observations est constituée des vieilles
traces d'alimentation de l'espèce.
4.1.1.14 Comparaison des IKA Pangolin géant entre les
trois sites (Inventaires diurnes indirects)
Dans l'ensemble des trois sites, ont été
enregistrés 100 signes de présence des Pangolin géant,
dont 16 signes à Gribé (IKA de 0,444#177;0,344), 16 signes
à Gouonepoun (IKA de 0,444#177;0,502) et 68 signes dans le PNBB (IKA de
1,889#177;1,015) (Tableau 26). De l'analyse des variances il en découle
qu'il y a de différences significatives entre les trois sites (one way
ANOVA: F2;15 = 8,9357; p = 0278)
(Figure 27).

Figure 24: Comparaison des
IKA de pangolin géant entre les trois sites (Inventaires diurnes
indirects)
Notes : a, b:
expression de la différence significative au seuil de probabilité
de 5%
Du test de Tukey de séparation des moyennes,
l'abondance relative moyenne du Pangolin géant dans le PNBB est
significativement supérieure à celle de Gribé (p
= 06288) et à celle de Gouonepoun (p = 06283). Mais, la
moyenne des IKA à Gribé (p = 06283) n'est pas
significativement différente de celle de Gouonepoun (p =
06283)
4.1.1.14.1 Léopard
Panthera pardus
Les signes d'activité de Léopard ont
été observés exclusivement à Gribé et nul
par ailleurs, rendant impossible la comparaison des IKA de cette
espèces. Le tableau 27 récapitule les IKA de léopard par
signe d'activité et par âge dans les trois sites
d'étude.
Tableau 27: IKA Léopard par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
sites
|
Signe d'activité
|
Très frais
|
Récent
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
empreinte
|
1
|
0,028
|
1
|
0,028
|
2
|
0,056
|
|
Global Gribé
|
1
|
0,028
|
1
|
0,028
|
2
|
0,056
|
|
Gouonepoun
|
/
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Global Gouonepoun
|
0
|
0
|
0
|
0
|
0
|
0
|
|
PNBB
|
/
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Global PNBB
|
0
|
0
|
0
|
0
|
0
|
0
|
Il ressort du tableau 27 que seul
le léopard a été enregistré à Gribé,
rendant impossible la comparaison des IKA entre les trois sites.
4.1.1.14.2 Athérure africain Atherurus
africanus
Le tableau 28 récapitule les IKA de Athérure
africain par signe d'activité et par âge dans les trois sites
d'étude.
Tableau 28: IKA Athérure africain par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
crottes
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
2
|
0,056
|
|
empreinte
|
2
|
0,056
|
7
|
0,194
|
4
|
0,111
|
0
|
0
|
0
|
0
|
13
|
0,361
|
|
piste
|
0
|
0
|
0
|
0
|
167
|
4,639
|
0
|
0
|
30
|
0,833
|
197
|
5,472
|
|
terrier
|
4
|
0,111
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
4
|
0,111
|
|
traces
|
1
|
0,028
|
3
|
0,083
|
10
|
0,278
|
3
|
0,083
|
1
|
0,028
|
18
|
0,500
|
|
Global Gribé
|
8
|
0,222
|
11
|
0,306
|
181
|
5,028
|
4
|
0,111
|
32
|
0,889
|
236
|
6,556
|
|
Gouonepoun
|
alimentation
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
|
crottes
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
empreinte
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
piste
|
0
|
0
|
0
|
0
|
143
|
3,972
|
0
|
0
|
0
|
0
|
143
|
3,972
|
|
terrier
|
6
|
0,167
|
0
|
0
|
0
|
0
|
0
|
0
|
5
|
0,139
|
11
|
0,306
|
|
traces
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
3
|
0,083
|
|
Global Gouonepoun
|
7
|
0,194
|
2
|
0,056
|
145
|
4,028
|
2
|
0,056
|
5
|
0,139
|
161
|
4,472
|
|
PNBB
|
alimentation
|
1
|
0,028
|
3
|
0,083
|
8
|
0,222
|
2
|
0,056
|
3
|
0,083
|
17
|
0,472
|
|
crottes
|
0
|
0
|
0
|
0
|
3
|
0,083
|
0
|
0
|
0
|
0
|
3
|
0,083
|
|
empreinte
|
0
|
0
|
3
|
0,083
|
2
|
0,056
|
0
|
0
|
0
|
0
|
5
|
0,139
|
|
piste
|
0
|
0
|
0
|
0
|
158
|
4,389
|
0
|
0
|
40
|
1,111
|
198
|
5,500
|
|
terrier
|
3
|
0,083
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
5
|
0,139
|
|
traces
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global PNBB
|
5
|
0,139
|
6
|
0,167
|
171
|
4,750
|
2
|
0,056
|
45
|
1,250
|
229
|
6,361
|
Il ressort du tableau 28 que 626 indices de
présence ont été relevés le long de 18 transects
parcourus. La plupart des observations est constituée de pistes
où l'animal passe régulièrement.
4.1.1.15 Comparaison des IKA Athérure africain entre
les trois sites (Inventaires diurnes indirects)
Il en ressort que dans l'ensemble des trois sites, ont
été enregistrés 626 signes de présence de
l'athérure africain dont 236 signes à Gribé (IKA de
6,556#177;3,489), 161 signes à Gouonepoun (IKA de 4,472#177;1,035) et
229 signes dans le PNBB (IKA de 6,361#177;2,101) (Tableau 28). De l'analyse des
variances des IKA de l'Athérure africain, il en découle qu'il n'y
a pas de différence significative entre les trois sites (one way ANOVA :
F2;15 = 1,3495; p = 0,28911) (Figure 28).
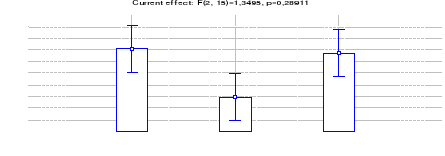
Figure 25: Comparaison des
IKA de l'Athérure africain entre les trois sites (Inventaires diurnes
indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.15.1
Potamochère Potamochoerus porcus
Le tableau 29 récapitule les IKA de potamochère
par signe d'activité et par âge dans les trois sites
d'étude.
Tableau 29: IKA Potamochère par signe
d'activité et par âge dans les trois sites d'étude
(Inventaires diurnes indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
alimentation
|
2
|
0,056
|
10
|
0,278
|
5
|
0,139
|
6
|
0,167
|
19
|
0,528
|
42
|
1,167
|
|
couchage
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
0
|
0
|
2
|
0,056
|
|
crottes
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
empreinte
|
3
|
0,083
|
14
|
0,389
|
19
|
0,528
|
5
|
0,139
|
2
|
0,056
|
43
|
1,194
|
|
piste
|
0
|
0
|
0
|
0
|
17
|
0,472
|
0
|
0
|
2
|
0,056
|
19
|
0,528
|
|
traces
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gribé
|
5
|
0,139
|
24
|
0,667
|
42
|
1,167
|
13
|
0,361
|
24
|
0,667
|
108
|
3,000
|
|
Gouonepoun
|
alimentation
|
0
|
0
|
14
|
0,389
|
26
|
0,722
|
10
|
0,278
|
11
|
0,306
|
61
|
1,694
|
|
bain de boue
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
0,056
|
|
crottes
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
empreinte
|
4
|
0,111
|
12
|
0,333
|
10
|
0,278
|
4
|
0,111
|
2
|
0,056
|
32
|
0,889
|
|
piste
|
0
|
0
|
0
|
0
|
13
|
0,361
|
0
|
0
|
1
|
0,028
|
14
|
0,389
|
|
traces
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gouonepoun
|
5
|
0,139
|
29
|
0,806
|
49
|
1,361
|
14
|
0,389
|
14
|
0,389
|
111
|
3,083
|
|
PNBB
|
alimentation
|
0
|
0
|
28
|
0,778
|
29
|
0,806
|
36
|
1,000
|
34
|
0,944
|
127
|
3,528
|
|
crottes
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
empreinte
|
1
|
0,028
|
9
|
0,250
|
13
|
0,361
|
3
|
0,083
|
7
|
0,194
|
33
|
0,917
|
|
nettoyage
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
piste
|
0
|
0
|
0
|
0
|
3
|
0,083
|
0
|
0
|
15
|
0,417
|
18
|
0,500
|
|
traces
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global PNBB
|
1
|
0,028
|
37
|
1,028
|
47
|
1,306
|
39
|
1,083
|
57
|
1,583
|
181
|
5,028
|
Il ressort du tableau 29 que 400 indices de
présence ont été relevés le long de 18 transects
parcourus. La plupart des observations est constituée des alimentations
et des empreintes de l'espèce.
4.1.1.16 Comparaison des IKA Potamochère entre les
trois sites (Inventaires diurnes indirects)
Il en ressort que dans l'ensemble des trois sites, ont
été enregistrés 400 signes de présence de
potamochère dont 108 signes à Gribé (IKA de
3,000#177;2,060), 111 signes à Gouonepoun (IKA de 3,083#177;1,718), et
181 signes dans le PNBB (IKA de 5,028#177;2,758) (Tableau 29). De l'analyse des
variances des IKA de potamochère, il en découle qu'il n'y a pas
de différence significative entre les trois sites (one way ANOVA :
F2;15= 1,6010; p = 0,23432) (Figure 29).
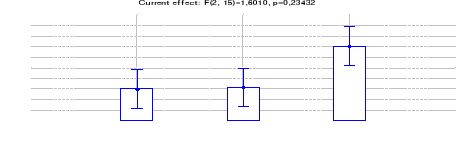
Figure 26: Comparaison des IKA de potamochère
entre les trois sites (Inventaires diurnes indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.16.1 Sitatunga Tragelaphus spekei
Le tableau 30 récapitule les IKA de Sitatunga par signe
d'activité et par âge d'indice dans les trois sites
d'étude.
Tableau 30: IKA Sitatunga par signe d'activité
et par âge dans les trois sites d'étude (Inventaires diurnes
indirects)
|
Site
|
Signe d'activité
|
Très frais
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
empreinte
|
2
|
0,056
|
4
|
0,111
|
8
|
0,222
|
0
|
0
|
2
|
0,056
|
16
|
0,444
|
|
piste
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
traces
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gribé
|
2
|
0,056
|
4
|
0,111
|
10
|
0,278
|
0
|
0
|
2
|
0,056
|
18
|
0,500
|
|
Gouonepoun
|
empreinte
|
2
|
0,056
|
1
|
0,028
|
1
|
0,028
|
0
|
0
|
0
|
0
|
4
|
0,111
|
|
piste
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gouonepoun
|
2
|
0,056
|
1
|
0,028
|
2
|
0,056
|
0
|
0
|
0
|
0
|
5
|
0,139
|
|
PNBB
|
empreinte
|
1
|
0,028
|
2
|
0,056
|
8
|
0,222
|
2
|
0,056
|
0
|
0
|
13
|
0,361
|
|
Global PNBB
|
1
|
0,028
|
2
|
0,056
|
8
|
0,222
|
2
|
0,056
|
0
|
0
|
13
|
0,361
|
Il ressort du tableau 30 que très peu d'observation a
été enregistrée pour le Sitatunga, rendant impossible la
comparaison de l'IKA de cette espèce entre les trois sites.
4.1.1.17 Comparaison des IKA des
grands et moyens mammifères entre les trois sites (Inventaires diurnes
indirects)
Il en ressort que dans l'ensemble des trois sites, ont
été enregistrés 4492 de signes d'observations de tous les
mammifères dont 1562 signes à Gribé (IKA de
44,222#177;21,451), 1300 signes à Gouonepoun (IKA de 36,278#177;11,725)
et 1630 signes dans le PNBB (IKA de 47,278#177;16,412) (Tableau 30). De
l'analyse des variances des IKA, il en découle qu'il n'y a pas de
différence entre les trois sites (one way ANOVA: F2;15
= 3,3932; p = 0,06085) (Figure 30).

Figure 27: Comparaison des
IKA des grands et moyens mammifères entre les trois sites (Inventaires
diurnes indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.1.18 Activités Humaines
Les activités humaines ont été
présentes le long de 18 transects dans la zone d'étude. Le
tableau 31 récapitule les IKA des Activités humaines par signe
d'activité et par âge dans les trois sites d'étude.
Tableau 31: IKA Activités humaines par signe
d'activité et par âge dans les sites d'étude (Inventaires
diurnes indirects)
|
Site
|
Signe d'activité
|
Frais
|
Récent
|
Vieux
|
Très vieux
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Gribé
|
campement
|
0
|
0
|
1
|
0,028
|
0
|
0
|
2
|
0,056
|
3
|
0,083
|
|
chasse
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
cueillette
|
1
|
0,028
|
2
|
0,056
|
0
|
0
|
1
|
0,028
|
4
|
0,111
|
|
douille
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
|
piège
|
0
|
0
|
0
|
0
|
0
|
0
|
3
|
0,083
|
3
|
0,083
|
|
piste de chasse
|
0
|
0
|
5
|
0,139
|
0
|
0
|
1
|
0,028
|
6
|
0,167
|
|
sachet whisky
|
1
|
0,028
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
|
Global Gribé
|
2
|
0,056
|
8
|
0,222
|
1
|
0,028
|
8
|
0,222
|
19
|
0,528
|
|
Gouonepoun
|
campement
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
cueillette
|
0
|
0
|
0
|
0
|
0
|
0
|
4
|
0,111
|
4
|
0,111
|
|
douille
|
0
|
0
|
0
|
0
|
1
|
0,028
|
0
|
0
|
1
|
0,028
|
|
piège
|
0
|
0
|
2
|
0,056
|
0
|
0
|
1
|
0,028
|
3
|
0,083
|
|
piste de chasse
|
0
|
0
|
15
|
0,417
|
0
|
0
|
1
|
0,028
|
16
|
0,444
|
|
Global Gouonepoun
|
0
|
0
|
17
|
0,472
|
1
|
0,028
|
7
|
0,194
|
25
|
0,694
|
|
PNBB
|
cueillette
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
0,028
|
1
|
0,028
|
|
piège
|
0
|
0
|
5
|
0,139
|
0
|
0
|
2
|
0,056
|
7
|
0,194
|
|
piste de chasse
|
0
|
0
|
3
|
0,083
|
0
|
0
|
4
|
0,111
|
7
|
0,194
|
|
sachet whisky
|
0
|
0
|
0
|
0
|
3
|
0,083
|
0
|
0
|
3
|
0,083
|
|
Global PNBB
|
0
|
0
|
8
|
0,222
|
3
|
0,083
|
7
|
0,194
|
18
|
0,500
|
Il en ressort que la majorité des observations
est constituée des pistes de chasses régulièrement
fréquentées par des chasseurs. La chasse est beaucoup plus
accentuée à Gouonepoun soit un IKA de 0,694. La majorité
des chasseurs viennent des villages environnants comme Song ancien, et
Maléa ancien. Le produit de la chasse est orienté vers le centre
urbain.
4.1.1.19 Comparaison des IKA des activités humaines
entre les trois sites (Inventaires diurnes indirects)
Dans l'ensemble des trois sites, ont été
enregistrés 62 observations des activités humaines dont 19 signes
d'activité humaine à Gribé (IKA de 0,528#177;0,306), 25
signes à Gouonepoun (IKA de 0,696#177;0,400) et 18 signes dans le PNBB
(IKA de 0,500#177;0,471) (Tableau 31). De l'analyse des variances des IKA, il
en découle qu'il n'y a pas de différence significative entre les
trois sites (one way ANOVA: F2;15= 0,41774; p =
0,66596) (Figure 31).

Figure 28: Comparaison des
IKA Activités humaines entre les trois sites (Inventaires diurnes
indirects)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.2
Inventaires diurnes directs
4.1.2.1 Estimation des densités de groupes des petits
singes entre les trois sites (Inventaires diurnes directs)
Très peu d'individus ont été vus
physiquement sur les 18 transects permanents parcourus dans la zone
d'étude, rendant impossible l'estimation de la densité pour
plusieurs espèces. Seule la densité des espèces de petits
singes a été calculée par le programme DISTANCE. Le
tableau 32 présente le nombre des individus vus pendant les inventaires
diurnes directs sur les transects à Gribé, Gouonepoun et PNBB.
Tableau 32: Individus vus physiquement et par site
(inventaires diurnes directs)
|
Gribé
|
Gouonepoun
|
PNBB
|
|
Espèces
|
Nombre vu
|
Nombre
vu
|
Nombre vu
|
|
Bongo
|
6
|
0
|
0
|
|
Céphalophe bleu
|
2
|
3
|
1
|
|
Céphalophe roux
|
3
|
4
|
1
|
|
Pangolin à longue queue
|
3
|
0
|
0
|
|
Petits singes
|
45
|
32
|
35
|
|
Porc épic
|
0
|
1
|
0
|
|
Potamochère
|
0
|
2
|
0
|
|
Totale
|
59
|
42
|
37
|
Il ressort du tableau 32 que les espèces de petits
singes sont les plus abondants dans toute la zone d'étude, soit un total
de 112 individus vus physiquement dont 12 groupes (45 individus) de petits
singes à Gribé, neuf groupes (32 individus) à Gouonepoun
et 13 groupes (35 individus) dans le PNBB.
4.1.2.1.1 Densité
de groupes de petits singes à Gribé (Inventaires diurnes
directs)
L'estimation de la densité des petits singes à
Gribé s'est faite sur la base des groupes d'individus vus physiquement.
Au total, 45 individus ont été relevés sur les six
transects parcourus. Parmi les 45 individus rencontrés sur les
transects, 22 individus étaient constitués de Hocheur
Cercopithecus nictitans, 12 de Mécanicien Cercocebus
albigena, neuf individus de Cercopithèque de brazza
Cercopithecus negletus et deux individus de Magistrat Colobus
guereza. La distance perpendiculaire maximale au transect est de 4200 cm.
Le tableau 33 récapitule les résultats statistiques de
l'estimation de la densité obtenus au travers du programme DISTANCE. La
figure 32 illustre l'allure de la courbe de probabilité de
détection des groupes de petits singes à Gribé en fonction
de la distance perpendiculaire au transect.
Tableau 33: Statistique d'estimation de la
densité de groupes de petits singes à Gribé (Inventaires
diurnes directs)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
200,266
|
|
|
|
|
f(0)
|
0,36163E-03
|
0,82944E-04
|
22,94
|
0,21970E-03
|
0,59526E-03
|
|
p
|
0,65839
|
0,15101
|
22,94
|
0,39999
|
1,0000
|
|
ESW
|
2765,2
|
634,23
|
22,94
|
1679,9
|
4551,6
|
|
n/L
|
0,33333
|
|
40,22
|
0,14726
|
0,75454
|
|
D
|
6,0272
|
2,7906
|
46,30
|
2,4347
|
14,921
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des
individus
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen des individus par unité de distance
du transect en km,
D = densité des individus (nombre/km2)
AIC = Akaike Information Criterion

Figure 29: Courbe de
probabilité de détection des petits singes à Gribé
en fonction de la distance perpendiculaire au transect (Inventaires diurnes
directs)
Au travers de DISTANCE, il en résulte une
densité moyenne de 6,027 groupes d'individus/km² variant entre les
valeurs 2,435 et 14,921 avec un coefficient de variation de 46,30: ce qui est
supérieur à 25%.
4.1.2.1.2 Densité
de groupes de petits singes à Gouonepoun (Inventaires diurnes
directs)
L'estimation de la densité des petits singes à
Gouonepoun s'est faite sur la base des groupes d'individus vus physiquement. Au
total, neuf groupes (32 individus) ont été relevés sur les
six transects parcourus. Parmi les 32 individus rencontrés sur les
transects, 16 individus étaient constitués de Hocheur
Cercopithecus nictitans, quatre individus de Mécanicien
Cercocebus albigena, 10 individus de Cercopithèque de brazza
Cercopithecus negletus et deux individus de Cercocèbe agile
Cercocebus agilis. La distance perpendiculaire maximale au transect
est de 4000 cm. Le tableau 34 récapitule les résultats
statistiques de l'estimation de la densité obtenus au travers du
programme DISTANCE. La figure 33 illustre l'allure de la courbe de
probabilité de détection des groupes de petits singes en fonction
de la distance perpendiculaire au transect.
Tableau 34: Statistique d'estimation de la
densité de groupes de petits singes à Gouonepoun (Inventaires
diurnes directs)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
150,95
|
|
|
|
|
f(0)
|
0,30780E-03
|
0,95877E-04
|
31,15
|
0,15259E-03
|
0,62091E-03
|
|
p
|
0,81221
|
0,25299
|
31,15
|
0,40263
|
1,0000
|
|
ESW
|
3248,8
|
1012,0
|
31,15
|
1610,5
|
6553,6
|
|
n/L
|
0,25000
|
|
29,15
|
0,13686
|
0,45667
|
|
D
|
3,8476
|
1,6414
|
42,66
|
1,6426
|
9,0123
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des individus
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen des individus par unité de distance
du transect en km,
D = densité des individus (nombre/km2)
AIC = Akaike Information Criterion

Figure 30: Courbe de
probabilité de détection des petits singes à Gouonepoun en
fonction de la distance perpendiculaire au transect (Inventaires diurnes
directs)
Au travers de DISTANCE, il en résulte une
densité moyenne de 3,848 groupes d'individus/km² variant entre les
valeurs 1,643 et 9,012 avec un coefficient de variation de 42,66 : ce qui
est légèrement supérieur à 25%, mais acceptable.
4.1.2.1.3 Densité
de groupes de petits singes dans le PNBB (Inventaires diurnes directs)
L'estimation de la densité des petits singes dans le
PNBB s'est faite sur la base de groupes d'individus vus physiquement. Au total,
13 groupes (35 individus) ont été relevées sur les six
transects parcourus, 13 individus étaient constitués de Hocheur
Cercopithecus nictitans, 10 individus de Mécanicien
Cercocebus albigena, 7 individus constitués de
Cercopithèque de brazza Cercopithecus negletus, deux individus
de singes Magistrat Colobus guereza et trois individus de
Cercocèbe agile Cercocebus agilis La distance perpendiculaire
maximale au transect est de 7000 cm. Le tableau 35 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 34 illustre l'allure de la courbe de
probabilité de détection des groupes de petits singes en fonction
de la distance perpendiculaire au transect.
Tableau 35: Statistique d'estimation de la
densité de groupes de petits singes dans le PNBB (Inventaires diurnes
directs)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
221,37
|
|
|
|
|
f(0)
|
0,47079E-03
|
0,12972E-03
|
27,55
|
0,25957E-03
|
0,85388E-03
|
|
p
|
0,30344
|
0,83607E-01
|
27,55
|
0,16730
|
0,55036
|
|
ESW
|
2124,1
|
585,25
|
27,55
|
1171,1
|
3852,5
|
|
n/L
|
0,36111
|
|
29,20
|
0,19748
|
0,66033
|
|
D
|
8,5004
|
3,4128
|
40,15
|
3,8471
|
18,782
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des
individus
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen des individus par unité de distance
du transect en km,
D = densité des individus (nombre/km2)
AIC = Akaike Information Criterion

Figure 31: Courbe de
probabilité de détection des petits singes dans le PNBB en
fonction de la distance perpendiculaire au transect (Inventaires diurnes
directs)
Au travers de DISTANCE, il en résulte une
densité moyenne de 8,500 groupes d'individus/km² variant entre les
valeurs 3,847et 18,782 avec un coefficient de variation de 40,15: ce qui est
légèrement supérieur à 25%, mais acceptable.
4.1.2.2 Comparaison des densités de groupes de petits
singes entre les trois sites (Inventaires diurnes directs)
La comparaison des densités moyennes des individus
entre les trois sites a été réalisée sur la base
des densités obtenues au travers de DISTANCE 6.0. De l'analyse des
variances des densités moyennes de groupe de petits singes, il en
découle qu'il n'y a pas de différence significative entre les
trois sites (one way ANOVA : F2;6= 0,60699; p
= 0,57535) (Figure 35).

Figure 32: Comparaison des
densités des groupes de petits singes entre les trois sites (Inventaires
diurnes directs)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.2.3 Estimation de l'abondance relative des espèces
entre les trois sites (Inventaires diurnes directs).
L'estimation de l'abondance relative s'est faite sur la base
des individus vus physiquement et les cris entendus. Le tableau 36
présente l'abondance relative individuelle des différentes
espèces identifiées sur les transects à Gribé,
à Gouonepoun et dans le PNBB.
Tableau 36: IKA des espèces dans l'ensemble de
la zone d'étude (Inventaires diurnes directs)
|
Espèces
|
Gribé
|
Gouonepoun
|
PNBB
|
Total
|
|
vu/
entendu
|
IKA
|
vu/
entendu
|
IKA
|
vu/
entendu
|
IKA
|
vu/
entendu
|
IKA
|
|
Bongo
|
6
|
0,167#177;0,408
|
0
|
0,0
|
0
|
0,0
|
6
|
0,167#177;0,408
|
|
Céphalophe bleu
|
2
|
0,056#177;0,086
|
3
|
0,083#177;0,091
|
1
|
0,028#177;0,068
|
6
|
0,167#177;0,245
|
|
Céphalophe roux
|
3
|
0,083#177;0,139
|
4
|
0,111#177;0,086
|
1
|
0,028#177;0,068
|
8
|
0,222#177;0,294
|
|
Chat sauvage
|
0
|
0,0
|
0
|
0,0
|
1
|
0,028#177;0,068
|
1
|
0,028#177;0,068
|
|
Chimpanzé
|
2
|
0,056#177;0,136
|
3
|
0,083#177;0,091
|
7
|
0,194#177;0,195
|
12
|
0,333#177;0,422
|
|
Eléphant
|
0
|
0,0
|
1
|
0,028#177;0,068
|
1
|
0,028#177;0,068
|
2
|
0,056#177;0,136
|
|
Gorille
|
0
|
0,0
|
0
|
0,0
|
2
|
0,056#177;0,086
|
2
|
0,056#177;0,086
|
|
Pangolin à longue queue
|
3
|
0,083#177;0,139
|
0
|
0,0
|
0
|
0,0
|
3
|
0,083#177;0,139
|
|
Petits singes
|
108
|
3#177;2,188
|
109
|
3,028#177;1,019
|
114
|
3,167#177;1,703
|
331
|
9,194#177;4,910
|
|
Athérure africain
|
0
|
0,0
|
1
|
0,028#177;0,068
|
0
|
0,0
|
1
|
0,028#177;0,068
|
|
Potamochère
|
0
|
0,0
|
3
|
0,083#177;0,139
|
2
|
0,056#177;0,136
|
5
|
0,139#177;0,276
|
|
Global
|
124
|
3,444#177;3,098
|
124
|
3,444#177;1,563
|
129
|
3,583#177;2,392
|
377
|
10,472#177;5,518
|
Il en ressort que dans l'ensemble des trois sites, ont
été enregistrés 377 signes de présence (vu/entendu)
des animaux lors des inventaires diurnes directs soit un IKA de
10,472#177;5,518, répartit comme suit : 124 signes à
Gribé (IKA de 3,444#177;3,089), 124 signes à Gouonepoun (IKA de
3,444#177;1,563) et 129 signes dans le PNBB (IKA de 3,583#177;2,392) (Tableau
36).
4.1.2.4 Comparaison des IKA de petits singes entre les trois
(Inventaires diurnes directs)
Fondamentalement, nous ne pouvons pas utiliser de test
statistique (Tukey test) lorsqu'on a zéro observation dans un
échantillon donné et aussi lorsque le nombre d'observation
apparait très peu (< 20). Seuls les IKA des petits singes entre les
trois sites ont été comparés.
4.1.2.4.1 Petits singes
Les petits singes relevés regroupent sept
espèces (Tableau 37). Dans l'ensemble, 331 groupes de petits singes ont
été observés, dont 108 groupes à Gribé (IKA
de 3,000#177;1,019), 109 à Gouonepoun (IKA de 3,028#177;2,188) et 114
dans le PNBB (IKA de 3,167#177;1,703). Le hocheur est l'espèce la plus
rencontrée dans les trois sites soit un groupe de 192 (IKA de 5,333),
tandis que le houstac est assez rare avec seulement deux rencontres sur 108 km
parcourus. De l'analyse des variances des IKA de Petits singes, il en
découle qu'il n'y a pas de différence significative entre les
trois sites (one way ANOVA: F2;15 = 0,01642;
p = 0,98373) (Figure 36).
Tableau 37: IKA des observations directes des petits
singes entre les trois sites (Inventaires diurnes directs)
|
Espèces
|
Nom scientifique
|
Gouonepoun
|
Gribé
|
PNBB
|
Total
|
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
n
|
IKA
|
|
Cercocèbe agile
|
Cercocebus agilis
|
6
|
0,167
|
0
|
0
|
13
|
0,361
|
19
|
0,528
|
|
Cercopithèque de brazza
|
Cercopithecus negletus
|
17
|
0,472
|
16
|
0,444
|
10
|
0,278
|
43
|
1,194
|
|
Cercopithèque poganias
|
Cercopithecus pogonias
|
0
|
0
|
1
|
0,028
|
5
|
0,139
|
6
|
0,167
|
|
Hocheur
|
Cercopithecus nictitans
|
72
|
2,000
|
58
|
1,611
|
62
|
1,722
|
192
|
5,333
|
|
Magistrat
|
Colobus guereza
|
2
|
0,056
|
5
|
0,139
|
7
|
0,194
|
14
|
0,389
|
|
Cercocèbe à joue grise
|
Cercocebus albigena
|
12
|
0,333
|
21
|
0,583
|
15
|
0,417
|
48
|
1,333
|
|
Moustac
|
Cercopithecus cephus
|
0
|
0
|
7
|
0,194
|
2
|
0,056
|
9
|
0,250
|
|
Global
|
|
109
|
3,028
|
108
|
3,000
|
114
|
3,167
|
331
|
9,194
|

Figure 33: Comparaison des
IKA de Petits singes entre les trois sites (inventaires diurnes
directs)
Notes: La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.2.5 Comparaison IKA global de grands et moyens
mammifères entre les trois sites (inventaires diurnes directs)
Il ressort du tableau 36 que sur l'ensemble des trois sites, a
été enregistré un total de 377 signes d'animaux de toutes
les espèces confondues (soit un IKA de 10,472#177;5,518) le long de 18
transects, sur une distance parcourue de 108 km. De l'analyse des variances de
IKA global de toutes les espèces recensées, il en découle
qu'il n'y a pas de différence significative entre les sites (one way
ANOVA : F2;15 = 0,01307; p = 0,98702) (Figure
37).

Figure 34: Comparaison IKA
global des grands et moyens mammifères entre les trois sites
(Inventaires diurnes directs)
NB : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.3
Inventaires nocturnes
4.1.3.1 Estimation de l'abondance relative des espèces
entre les trois sites (Inventaires nocturnes)
L'estimation de l'abondance relative s'est faite sur la base
des individus vus physiquement et les cris entendus. Le tableau 38
présente l'abondance relative individuelle des différentes
espèces identifiées lors des inventaires nocturnes directes sur
les transects à Gribé, à Gouonepoun et dans le PNBB.
Tableau 38: IKA des espèces dans l'ensemble de
la zone d'étude (Inventaires nocturnes)
|
Espèces
|
Gribé
|
Gouonepoun
|
PNBB
|
Total
|
|
vu/
entendu
|
IKA
|
vu/
entendu
|
IKA
|
vu/
entendu
|
IKA
|
vu/
entendu
|
IKA
|
|
Céphalophe bleu
|
28
|
0,778#177;0,612
|
52
|
1,444#177;0,765
|
53
|
1,472#177;0,414
|
133
|
3,694#177;1,790
|
|
Céphalophe roux
|
8
|
0,222#177;0,251
|
8
|
0,222#177;0,172
|
4
|
0,111#177;0,136
|
20
|
0,556#177;0,559
|
|
Chat sauvage
|
0
|
0
|
1
|
0,028#177;0,068
|
0
|
0
|
1
|
0,028#177;0,068
|
|
Chimpanzé
|
5
|
0,139#177;0,125
|
2
|
0,056#177;0,086
|
5
|
0,139#177;0,195
|
12
|
0,333#177;0,406
|
|
Civette
|
18
|
0,500#177;0,683
|
14
|
0,389#177;0,202
|
10
|
0,278#177;0,202
|
42
|
1,167#177;1,087
|
|
Daman des arbres
|
256
|
7,111#177;2,444
|
276
|
7,667#177;1,434
|
273
|
7,583#177;0,815
|
805
|
22,361#177;4,693
|
|
Eléphant
|
4
|
0,111#177;0,172
|
0
|
0
|
1
|
0,028#177;0,068
|
5
|
0,139#177;0,240
|
|
Gorille
|
5
|
0,139#177;0,125
|
3
|
0,083#177;0,139
|
7
|
0,194#177;0,245
|
15
|
0,417#177;0,510
|
|
Pangolin à longue queue
|
3
|
0,083#177;0,139
|
4
|
0,111#177;0,136
|
11
|
0,306#177;0,125
|
18
|
0,500#177;0,401
|
|
Petits singes
|
14
|
0,389#177;0,202
|
9
|
0,250#177;0,230
|
6
|
0,167#177;0,105
|
29
|
0,806#177;0,537
|
|
Porc épic
|
2
|
0,056#177;0,086
|
4
|
0,111#177;0,086
|
4
|
0,111#177;0,136
|
10
|
0,278#177;0,308
|
|
Potamochère
|
5
|
0,139#177;0,267
|
4
|
0,111#177;0,136
|
1
|
0,028#177;0,068
|
10
|
0,278#177;0,471
|
|
Potto de Bosman
|
2
|
0,056#177;0,086
|
1
|
0,028#177;0,068
|
2
|
0,056#177;0,086
|
5
|
0,139#177;0,240
|
|
Total global
|
350
|
9,722#177;5,193
|
378
|
10,500#177;3,523
|
377
|
10,472#177;2,596
|
1105
|
30,694#177;11,311
|
Il en
ressort que dans l'ensemble des trois sites, ont été
enregistrés 1105 signes de présence (vu/entendu) des animaux lors
des inventaires nocturnes directs soit un IKA moyen global de
30,694#177;11,311, répartit comme suit : 350 signes à
Gribé (IKA de 9,722#177;5,193), 378 signes à Gouonepoun (IKA de
10,5#177;3,523) et 377 signes dans le PNBB (IKA de 10,472#177;2,596) (Tableau
38).
4.1.3.2 Comparaison de l'abondance relative des
espèces entre les trois sites (Inventaires nocturnes)
Fondamentalement, nous ne pouvons pas utiliser de test
statistique (Tukey test) lorsqu'on a zéro observation dans un
échantillon donné et aussi lorsque le nombre d'observation
apparait très peu (< 20). Seuls les IKA de céphalophes bleus
et des damans des arbres ont été comparés. Entre les trois
sites.
4.1.3.2.1
Céphalophe bleu Cephalophus monticola
Il ressort du tableau 38 que sur l'ensemble des trois sites un
total de 133 céphalophes bleus a été enregistré
pendant les inventaires nocturnes, dont 28 à Gribé (IKA de
0,778#177;0,612), 52 à Gouonepoun (IKA de 1,444#177;0,765) et 53 dans le
PNBB (IKA de 1,472#177;0,414). De la comparaison des moyennes des IKA de
céphalophe bleu, il en découle qu'il n'y a pas de
différence significative entre les trois sites (One-way ANOVA,
F2;15 = 2,4609, p = 0,11905) (Figure
38).

Figure 35: Comparaison des
IKA des céphalophes bleus entre les trois sites (Inventaires
nocturnes)
Notes: La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.3.2.2 Daman des arbres
Dendrohyrax arboreus
Il ressort du tableau 38 que sur l'ensemble des trois sites un
total de 805 cris de daman des arbres a été enregistré
pendant les inventaires nocturnes directs, dont 256 à Gribé (IKA
de 7,111#177;2,444), 276 à Gouonepoun (IKA de 7,667#177;1,434) et 273
dans le PNBB (IKA de 7,583#177;0,815). De la comparaison des moyennes des IKA
de daman des arbres, il en découle qu'il n'y a pas de différence
significative entre les trois sites (One-way ANOVA,
F2;15 = 0,18594, p = 0,83221) (Figure
39).
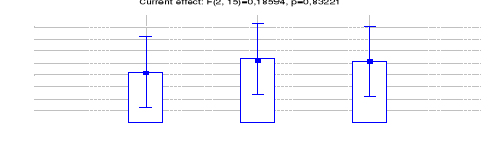
Figure 36: Comparaison des
IKA de daman des arbres entre les trois sites (Inventaires
nocturnes)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.3.3 Comparaison IKA de tous les grands et moyens
mammifères entre les trois sites (Inventaires nocturnes)
Il ressort du tableau 38 que sur l'ensemble des trois sites un
total 1105 animaux a été enregistré pendant les
inventaires nocturnes directs, dont 350 à Gribé (IKA de
9,722#177;5,193), 378 à Gouonepoun (IKA de 10,500#177;3523) et 377 dans
le PNBB (IKA de 10,472#177;2,596). De la comparaison des moyennes de
l'abondance relative de tous les animaux recensés, il en découle
qu'il n'y a pas de différence significative entre les trois sites
(One-way ANOVA, F2;15 = 0,028052, p = 0,75927)
(Figure 40).
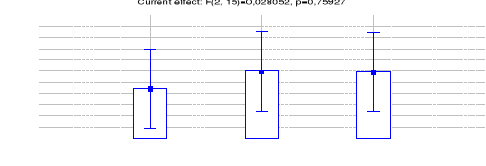
Figure 37: Comparaison IKA
global des grands et moyens mammifères entre les trois sites
(Inventaires nocturnes)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.3.4 Estimation des densités des espèces
recensées entre les trois sites (Inventaires nocturnes)
L'estimation des densités s'est faite sur la base des
animaux vus physiquement. Le tableau 39 présente le nombre des
espèces par observation directe pendant les inventaires nocturnes sur
les transects à Gribé, Gouonepoun et PNBB.
Tableau 39: Espèces vues physiquement et par
site (Inventaires nocturnes)
|
Gribé
|
Gouonepoun
|
PNBB
|
|
Espèce
|
Nombre vu
|
Nombre vu
|
Nombre vu
|
|
Céphalophe bleu
|
28
|
52
|
53
|
|
Céphalophe roux
|
8
|
8
|
4
|
|
Civette
|
18
|
14
|
10
|
|
Eléphant
|
2
|
0
|
0
|
|
Pangolin à longue queue
|
3
|
4
|
9
|
|
Athérure africain
|
2
|
4
|
4
|
|
Potamochère
|
4
|
2
|
0
|
|
Potto de Bosman
|
2
|
1
|
2
|
|
Totale
|
67
|
85
|
82
|
Il ressort du tableau 38 que les Céphalophes bleus sont
les plus abondants dans toute la zone d'étude lors des inventaires
nocturnes directs, soit un total de 133 individus vus physiquement dont 28
à Gribé, 52 à Gouonepoun et 53 dans le PNBB.
4.1.3.4.1 Densité
de Céphalophe bleu à Gribé
L'estimation de la densité de Céphalophe bleu
Cephalophus monticola s'est faite sur la base des individus vus
physiquement. Au total, 28 individus ont été observés sur
les six transects parcourus à Gribé. La distance perpendiculaire
maximale au transect est de 1560 cm. Le tableau 40 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 41 illustre bien l'allure de la courbe
de probabilité de détection des individus en fonction de la
distance perpendiculaire au transect.
Tableau 40: Statistique d'estimation de la
densité de Céphalophe bleu à Gribé (Inventaires
nocturnes)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
410,90
|
|
|
|
|
f(0)
|
0,89482E-03
|
0,17348E-03
|
19,39
|
0,60336E-03
|
0,13271E-02
|
|
p
|
0,71637
|
0,13888
|
19,39
|
0,48303
|
1,0000
|
|
ESW
|
1117,5
|
216,65
|
19,39
|
753,53
|
1657,4
|
|
n/L
|
0,77778
|
|
20,28
|
0,50925
|
1,1879
|
|
D
|
34,799
|
9,7623
|
28,05
|
19,958
|
60,674
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des
individus
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen des individus par unité de distance
du transect en km,
D = densité des individus (nombre/km2)
AIC = Akaike Information Criterion

Figure 38: Courbe de
probabilité de détection de Céphalophe bleu à
Gribé en fonction de la distance perpendiculaire au transect
(Inventaires nocturnes)
Au travers de DISTANCE, il en résulte une
densité moyenne de 34,799 individus/km² variant entre les valeurs
19,958 et 60,674 avec un coefficient de variation de 28,05: ce qui est
légèrement supérieur à 25%, mais acceptable.
4.1.3.4.2 Densité
de Céphalophe bleu à Gouonepoun
L'estimation de la densité de Céphalophe bleu
Cephalophus monticola s'est faite sur la base des individus vus
physiquement. Au total, 52 individus ont été observés sur
les six transects parcourus à Gouonepoun. La distance perpendiculaire
maximale au transect est de 2100 cm. Le tableau 41 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 42 illustre bien l'allure de la courbe
de probabilité de détection des individus en fonction de la
distance perpendiculaire au transect.
Tableau 41: Statistique d'estimation de la
densité de Céphalophe bleu à Gouonepoun (Inventaires
nocturnes)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
761,77
|
|
|
|
|
f(0)
|
0,10859E-02
|
0,11009E-03
|
10,14
|
0,88635E-03
|
0,13303E-02
|
|
p
|
0,43854
|
0,44459E-01
|
10,14
|
0,35797
|
0,53725
|
|
ESW
|
920,93
|
93,365
|
10,14
|
751,73
|
1128,2
|
|
n/L
|
1,4444
|
|
23,56
|
0,88454
|
2,3588
|
|
D
|
78,423
|
20,116
|
25,65
|
46,557
|
132,10
|
Avec:
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des
individus
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen des individus par unité de distance
du transect en km,
D = densité des individus (nombre/km2)
AIC = Akaike Information Criterion
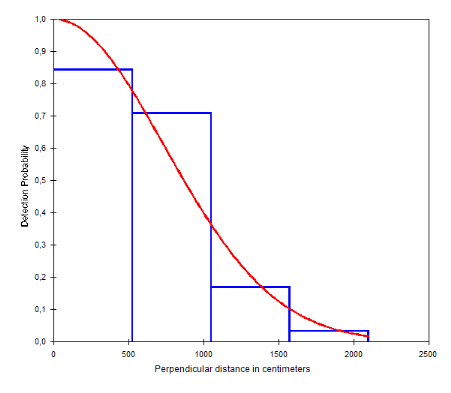
Figure 39: Courbe de
probabilité de détection de Céphalophe bleu à
Gouonepoun en fonction de la distance perpendiculaire au transect (inventaires
nocturnes)
Au travers de DISTANCE, il en résulte une
densité moyenne de 91,328 individus/km² variant entre les valeurs
63,903 et 130,520 avec un coefficient de variation de 17,750: ce qui est bon
car inférieur à 25%.
4.1.3.4.3 Densité
de Céphalophe bleu dans le PNBB
L'estimation de la densité de Céphalophe bleu
Cephalophus monticola s'est faite sur la base des individus vus
physiquement. Au total, 53 individus ont été observés sur
les six transects parcourus dans le PNBB. La distance perpendiculaire maximale
au transect est de 1700 cm. Le tableau 42 récapitule les
résultats statistiques de l'estimation de la densité obtenus au
travers du programme DISTANCE. La figure 43 illustre bien l'allure de la courbe
de probabilité de détection des individus en fonction de la
distance perpendiculaire au transect.
Tableau 42: Statistique d'estimation de la
densité de Céphalophe bleu dans le PNBB (Inventaires
nocturnes)
|
Paramètre
|
Valeur Estimée
|
Erreur standard
|
Coefficient de variation (%)
|
Intervalle de confiance à 95%
|
|
AIC
|
760,76
|
|
|
|
|
f(0)
|
0,12407E-02
|
0,12884E-03
|
10,38
|
0,10079E-02
|
0,15273E-02
|
|
p
|
0,47412
|
0,49234E-01
|
10,38
|
0,38516
|
0,58364
|
|
ESW
|
806,01
|
83,698
|
10,38
|
654,77
|
992,19
|
|
n/L
|
1,4722
|
|
17,00
|
1,0884
|
1,9914
|
|
D
|
91,328
|
16,208
|
17,75
|
63,903
|
130,52
|
Avec:
Modèle: Half-normal, Cosine adjustments
f(0) = fonction de la probabilité de densité de
la distance perpendiculaire à la ligne évaluer à 0
p = probabilité de détection des
individus
ESW = largeur effective du transect (en centimètre)
n/L = nombre moyen des individus par unité de distance
du transect en km,
D = densité des individus (nombre/km2)
AIC = Akaike Information Criterion
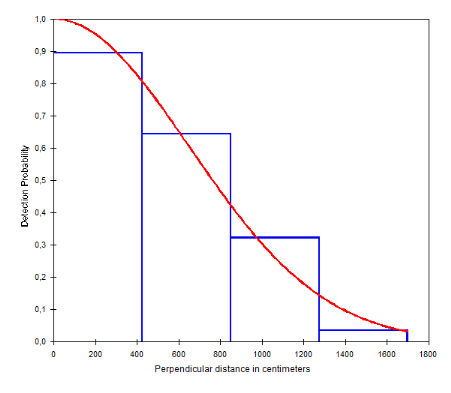
Figure 40: Courbe de
probabilité de détection de Céphalophe bleu dans le PNBB
en fonction de la distance perpendiculaire au transect (inventaires
nocturnes)
Au travers de DISTANCE, il en résulte une
densité moyenne de 78,423 individus/km² variant entre les valeurs
46,557 et 132,10 avec un coefficient de variation de 25,65: ce qui est
légèrement supérieur à 25%, mais acceptable.
4.1.3.5 Comparaison des densités de Céphalophes
bleus entre les trois sites (Inventaires nocturnes)
La comparaison des densités moyennes des individus
entre les trois sites a été réalisée sur la base
des densités obtenues au travers de DISTANCE 6.0. De l'analyse des
variances des densités moyennes de Céphalophes bleus, il en
découle qu'il n'y a pas de différence significative entre les
trois sites (one way ANOVA : F2;6= 2,4354; p
= 0,16814) (Figure 44).
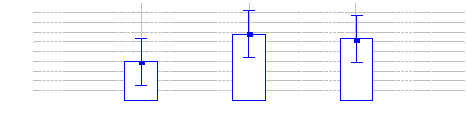
Figure 41: Comparaison des
densités de Céphalophes bleus entre les trois sites (inventaires
nocturnes)
Notes: La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.4
Caméras infra-rouges
4.1.4.1 Estimation des densités au moyen des
Caméras infra-rouges entre les trois sites
L'estimation des densités étaient fonction du
nombre de pièges photographiques (après filtrage des images
capturés), de la portée maximale de flash infra rouge, et du
champ de vision de la caméra en degré donnée par la
formule de (Rowcliffe et al., 2011). Le tableau 43
présente les densités des différentes espèces
animales au moyen des inventaires par les Caméras infra-rouges sur les
transects à Gribé, Gouonepoun et PNBB.
Tableau 43: Densité des espèces
capturées au moyen des Caméras infra-rouges dans les trois
sites
|
|
Gribé
|
Gouonepoun
|
PNBB
|
|
Espèces
|
Nombre de
capture
|
Ind/km²
|
Caméra nuit
|
Nombre de
capture
|
Ind/km²
|
caméra nuit
|
Nombre de
capture
|
caméra nuit
|
Ind/km²
|
|
Antilope de Bates
|
0
|
0
|
0
|
1
|
0,100#177;0,347
|
12
|
0
|
0
|
0
|
|
Bongo
|
1
|
0,100#177;0,332
|
12
|
5
|
0,700#177;2,426
|
24
|
0
|
0
|
0
|
|
Buffle
|
1
|
0,100#177;0,332
|
12
|
1
|
0,100#177;0,347
|
12
|
0
|
0
|
0
|
|
Céphalophe à dos jaune
|
4
|
0,400#177;0,895
|
36
|
14
|
1,501#177;1,629
|
156
|
14
|
156
|
1,401#177;1,761
|
|
Céphalophe bleu
|
69
|
8,405#177;13,317
|
240
|
110
|
12,408#177;11,989
|
360
|
100
|
336
|
11,507#177;17,123
|
|
Céphalophe roux
|
33
|
3,402#177;6,217
|
168
|
46
|
5,203#177;5,868
|
360
|
30
|
216
|
3,002#177;3,218
|
|
Chat sauvage
|
1
|
0,100#177;0,332
|
12
|
0
|
0
|
0
|
2
|
24
|
0,200#177;0,467
|
|
Chevrotain aquatique
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
24
|
0,200#177;0,467
|
|
Chimpanzé
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
12
|
0,100#177;0,347
|
|
Civette
|
1
|
0,100#177;0,332
|
12
|
4
|
0,400#177;1,066
|
24
|
2
|
24
|
0,200#177;0,693
|
|
Daman des arbres
|
0
|
0
|
0
|
0
|
0
|
0
|
1
|
12
|
0,100#177;0,347
|
|
Eléphant
|
5
|
0,801#177;1,576
|
60
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Gorille
|
0
|
0
|
0
|
3
|
0,600#177;1,402
|
36
|
2
|
24
|
0,200#177;0,467
|
|
Oryctérope
|
0
|
0
|
0
|
3
|
0,400#177;0,782
|
48
|
2
|
24
|
0,300#177;0,746
|
|
Pangolin à longue queue
|
0
|
0
|
0
|
0
|
0
|
0
|
7
|
36
|
0,700#177;1,489
|
|
Pangolin géant
|
0
|
0
|
0
|
2
|
0,200#177;0,467
|
24
|
1
|
12
|
0,100#177;0,347
|
|
Petits singes
|
32
|
5,103#177;8,961
|
72
|
23
|
3,102#177;4,418
|
192
|
9
|
84
|
1,001#177;1,126
|
|
Porc épic
|
3
|
0,300#177;0,715
|
36
|
11
|
1,201#177;1,774
|
132
|
18
|
168
|
1,801#177;3,377
|
|
Potamochère
|
6
|
0,901#177;2,646
|
48
|
8
|
1,101#177;2,013
|
72
|
0
|
0
|
0
|
|
Total général
|
156
|
19,713#177;35,653
|
708
|
231
|
27,017#177;34,528
|
1452
|
191
|
1152
|
20,813#177;31,976
|
Ind = individu; P = Piège photographique
Il ressort du tableau 43 que 19 espèces ont
été capturées par les caméras le long des 18
transects dans la zone d'étude. Les Céphalophes bleus
étaient les plus abondants dans toute la zone d'étude, soit un
total de 279 pièges photographiques capturés après
filtrage des images dont 69 à Gribé (densité de
8,405#177;13,317), 110 à Gouonepoun (densité 12,408#177;11,989)
et de 100 dans le PNBB (densité de 11,507#177;17,123). Par ailleurs, le
Chimpanzé et le daman des arbres étaient les espèces les
moins capturées par les caméras de surveillance.
4.1.4.2 Comparaison des densités des espèces au
moyen des Caméras infra-rouges entre les trois
4.1.4.2.1
Céphalophe bleu Cephalophus monticola
Lors des inventaires au moyen de Caméra infra-rouge, un
total de 279 pièges photographiques a été
enregistré dont 69 à Gribé (densité moyenne =
8,405#177;13,317), de 110 à Gouonepoun (densité moyenne =
12,408#177;11,989) et de 100 dans le PNBB (densité moyenne =
11,507#177;17,123) De la comparaison des densités moyennes des
céphalophes bleus, il en découle qu'il n'y a pas de
différence significative entre les différents sites (One-way
ANOVA, F2, 33 = 0,25176; p = 0,77891) (Figure
45).
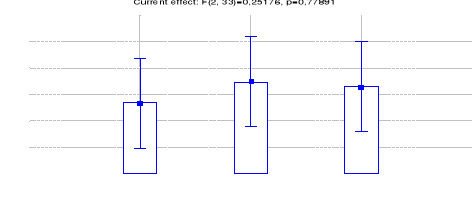
Figure 42: Comparaison des
densités de Céphalophe bleu entre les trois sites (Caméra
infra-rouge)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.4.2.2
Céphalophe roux
Lors des inventaires au moyen de Caméra infra-rouge, un
total de 109 pièges photographiques a été
enregistré dont 33 à Gribé (densité moyenne =
3,402#177;6,217), de 46 à Gouonepoun (densité moyenne =
5,203#177;5,868) et de 30 dans le PNBB (densité moyenne =
3,002#177;3,218) De la comparaison des densités moyennes de
céphalophe roux, il en découle qu'il n'y a pas de
différence significative entre les différents sites (One-way
ANOVA, F2, 33 = 0,56921; p = 0,57143) (Figure
46).
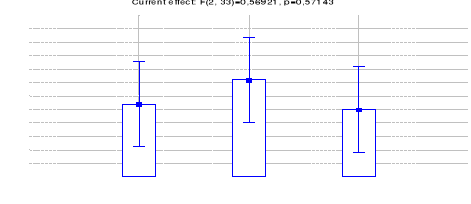
Figure 43: Comparaison des
densités de Céphalophe roux entre les trois sites (Caméra
infra-rouge)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.4.2.3 Petits singes
Lors des inventaires au moyen des Caméras infra-rouges,
les petits singes regroupent cinq espèces dont, le Hocheur
Cercopithecus nictitans, le Cercocèbe agile Cercocebus
agilis, le Magistrat Colobus guereza, le Mangabé à
joues grises ou mécanicien Cercocebus albigena, la Queue rouge
Cercopithecus cephus. Les densités moyennes de Petits singes
étaient de 5,103#177;8,961 à Gribé, de 3,102#177;4,418
à Gouonepoun et de 1,001#177;1,126 au PNBB (Tableau 43). De la
comparaison des densités moyennes des petits singes, il en
découle qu'il n'y a pas de différence significative entre les
différents sites (One-way ANOVA, F2, 15 = 1,3980;
p = 0,26134) (Figure 47).

Figure 44: Comparaison des
densités de Petits singes entre les trois sites (Caméra
infra-rouge)
Notes: La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.4.3 Estimation de Wildlife Picture Index (WPI) dans les
trois sites
L'estimation de WPI s'est basée sur les observations
faites à l'aide des caméras Bushnell Trophy Cam. 12
caméras ont été installées dans chaque site pour
une durée de 30 jours par site, soit un effort de capture de 360
(nuits-pièges) par site, sur les 18 transects permanents dans la zone
d'étude. Le tableau 44 récapitule l'indice photographique de la
faune (WPI) des différentes espèces prises au moyen des
caméras infra-rouges sur les transects à Gribé, Gouonepoun
et PNBB.
Tableau 44: Indice photographique de la faune (WPI)
des espèces dans les trois sites
|
Espèces
|
Gribé
|
Gouonepoun
|
PNBB
|
|
Nsp
|
WPI
|
Nsp
|
WPI
|
Nsp
|
WPI
|
|
Bongo
|
1
|
0,278#177;0,680
|
7
|
1,944#177;4,763
|
0
|
0
|
|
Buffle
|
1
|
0,278#177;0,680
|
1
|
0,278#177;0,680
|
0
|
0
|
|
Céphalophe à dos jaune
|
4
|
1,111#177;1,721
|
15
|
4,167#177;2,739
|
14
|
3,889#177;2,277
|
|
Céphalophe bleu
|
84
|
23,333#177;26,916
|
124
|
34,444#177;23,038
|
100
|
31,944#177;32,325
|
|
Céphalophe roux
|
34
|
9,444#177;12,590
|
52
|
14,444#177;11,816
|
30
|
8,333#177;4,714
|
|
Chat sauvage
|
1
|
0,278#177;0,680
|
0
|
0
|
2
|
0,556#177;0,861
|
|
Chevrotain aquatique
|
0
|
0
|
0
|
0
|
2
|
0,556#177;1,361
|
|
Chimpanzé
|
0
|
0
|
0
|
0
|
1
|
0,278#177;0,680
|
|
Civette
|
1
|
0,278#177;0,680
|
4
|
1,111#177;1,361
|
2
|
0,556#177;1,361
|
|
Daman des arbres
|
0
|
0
|
0
|
0
|
1
|
0,278#177;0,680
|
|
Eléphant
|
8
|
2,222#177;3,443
|
0
|
0
|
0
|
0
|
|
Gorille
|
0
|
0
|
6
|
1,667#177;2,582
|
2
|
0,556#177;0,861
|
|
Oryctérope
|
0
|
0
|
4
|
1,111#177;1,361
|
2
|
0,833#177;1,394
|
|
Pangolin à longue queue
|
0
|
0
|
0
|
0
|
7
|
1,944#177;2,670
|
|
Pangolin géant
|
0
|
0
|
2
|
0,556#177;0,861
|
1
|
0,278#177;0,680
|
|
Petits singes
|
51
|
14,167#177;15,977
|
31
|
8,611#177;7,025
|
9
|
2,778#177;2,018
|
|
Athérure africain
|
3
|
0,833#177;1,394
|
12
|
3,333#177;3,162
|
18
|
5,000#177;5,774
|
|
Potamochère
|
9
|
2,500#177;5,349
|
12
|
3,056#177;3,402
|
0
|
0
|
|
Total général
|
197
|
54,722#177;70,112
|
269
|
74,722#177;62,790
|
191
|
57,778#177;57,657
|
Nsp= nombre total d'une espèce donnée
Il ressort du tableau 44 que les céphalophes bleus
étaient les plus abondants dans toute la zone d'étude, soit un
total de 308 individus enregistrés par les caméras de
surveillance dont 84 à Gribé (WPI de 23,333#177;26,916), 124
à Gouonepoun (WPI de 34,444#177;23,038) et 100 dans le PNBB (WPI de
31,944#177;32,325). Par ailleurs, le chimpanzé et le daman des arbres
étaient les espèces les moins détectées par les
caméras de surveillance (Figure 48).

Figure 45: WPI moyen des
espèces capturées dans les trois sites (Caméra
infra-rouge)
Il ressort de la figure 48 que les céphalophes bleus
sont les plus abondants dans la zone d'étude. Ceci s'explique par le
fait que les céphalophes sont très prolifiques. Par contre, les
damans et les chimpanzés sont les moins abondants dans la zone
d'étude.
4.1.4.4 Comparaison WPI des différentes espèces
entre les trois sites (Caméras infra-rouges)
Fondamentalement, nous ne pouvons pas utiliser de test
statistique (Tukey test) lorsqu'on a zéro observation dans un
échantillon donné et aussi lorsque le nombre d'observation
apparait très peu (< 20) (Tableau 44). Seuls les WPI de
céphalophes bleus, céphalophes roux et des petits singes ont
été comparés entre les trois sites.
4.1.4.4.1
Céphalophe bleu Cephalophus monticola
Lors des inventaires au moyen de Caméra infra-rouge,
les WPI des céphalophes bleus étaient de 23,333#177;26,916
à Gribé, de 34,444#177;23,038 à Gouonepoun et de
31,944#177;32,325 au PNBB (Tableau 44). De la comparaison des moyennes de WPI
de céphalophe bleu, il en découle qu'il n'y a pas de
différence significative entre les différents sites (One-way
ANOVA, F2, 15 = 0,26588; p = 0,77007) (Figure
49).
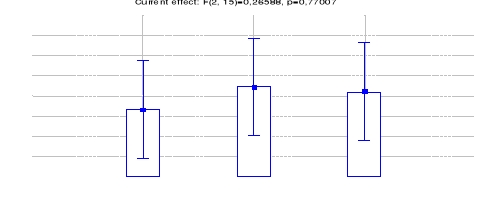
Figure 46: Comparaison des
WPI de Céphalophe bleu entre les trois sites (Caméra
infra-rouge)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.4.4.2
Céphalophe roux
Lors des inventaires au moyen de Caméra infra-rouge,
les WPI des céphalophes roux étaient de 9,444#177;12,590 à
Gribé, de 14,444#177;11,816 à Gouonepoun et de 8,333#177;4,714 au
PNBB (Tableau 44). De la comparaison des moyennes de WPI de céphalophe
roux, il en découle qu'il n'y a pas de différence significative
entre les différents sites (One-way ANOVA, F2, 15 =
0,59532; p = 0,56390) (Figure 50).

Figure 47: Comparaison des
WPI de Céphalophe roux entre les trois sites (Caméra
infra-rouge)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.4.4.3 Petits singes
Lors des inventaires au moyen de Caméra infra-rouge,
les WPI de Petits singes étaient de 14,167#177;15,977 à
Gribé, de 8,611#177;7,025 à Gouonepoun et de 2,778#177;2,018 au
PNBB (Tableau 44). Les espèces de petits singes regroupent ici cinq
espèces entre autres, le cercocèbe agile, le hocheur, le
magistrat, cercocèbe à joue grise et le moustac. De la
comparaison des moyennes de WPI des espèces de petits singes, il en
découle qu'il n'y a pas de différence significative entre les
différents sites (One-way ANOVA, F2, 15 = 1,8912;
p = 0,18516) (Figure 51).
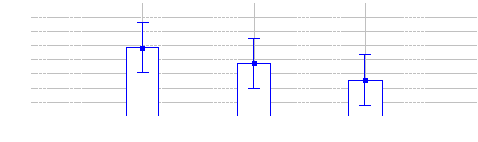
Figure 48: Comparaison des
WPI de Petits singes entre les trois sites (Caméra
infra-rouge)
Notes : La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.4.5 Comparaison de WPI de tous les mammifères
entre les trois sites (Caméra infra-rouge)
De la comparaison des WPI global de toutes les espèces
capturées, il en découle qu'il n'y a pas une différence
significative entre les différents sites (One-way ANOVA, F2, 15
= 0,35992; p = 0,70359) (Figure 52).

Figure 49: Comparaison de
WPI de tous les mammifères entre les trois sites (Caméra
infra-rouge)
Notes: La lettre a indique qu'il n'y a pas de
différence significative au seuil de 5 %.
4.1.5
Comparaison des résultats avec ceux obtenus dans la région du
Sud-est, Cameroun
Les inventaires
réalisés dans la zone d'étude (PNBB, ZICGC 13 et 14) font
état de la présence de 28 espèces de grands et moyens
mammifères. Il faut signaler que dans la ZICGC N°3 superposée en
très grande partie à l'UFA 10-063, 28 espèces avaient
été recensées en 2002 (Nzooh et al., 2002).
Tableau 45: Richesse spécifique des grands et
moyens mammifères dans quelques sites du Sud-est Cameroun
|
Site
|
Richesse spécifique
|
Auteurs
|
|
Parc National de Lobéké
|
32-45
|
WCS (1996); Stromayer et Ekobo (1991); Elkan (1994); Ekobo
(1995); Nzooh (2003)
|
|
Parc National de Boumba Bek et Nki
|
34
|
Ekobo (1998)
|
|
Forêt de Ngoila-Mintom
|
37
|
Nzooh (2003)
|
|
ZICGC N°1, 2, 3, 8 et 9
|
26-33
|
Nzooh et al. (2002)
|
|
UFA 10-018
|
27
|
Nzooh et al. (2003)
|
|
UFA 10-022
|
38
|
Nzooh et al. (2004a)
|
|
UFA 10-013
|
28
|
Nzooh et al. (2004b)
|
|
UF1 10-053
|
23
|
Bobo (2006b)
|
|
UFA 10-051
|
22
|
Bobo (2006a)
|
|
UFA 10-013
|
22
|
Bobo (2009a)
|
|
UFA 10-025
|
20
|
Bobo (2009b)
|
|
UFA 10-063
|
24
|
Bobo (2009c)
|
|
PNBB, ZICGC 13 et 14
|
28
|
Présente étude
|
Il ressort du tableau 45 que 28
espèces de grands et moyens mammifères ont été
recensées dans la zone d'étude. Cette valeur se rapproche de
celle obtenue par Nzooh et al. (2003) qui avait trouvé une
richesse spécifique estimée à 27 espèces. Cette
stabilité s'expliquerait par les efforts consentis par les organismes de
conservation en charge de la faune.
Le taux de rencontre au km des signes d'activité des
différentes espèces pour les inventaires diurnes indirects est de
41,935. Ce taux a subi une hausse considérable comparée aux
données de 2002 sur le même site qui indiquaient un taux de
rencontre de 28,02 (Nzooh et al., 2002), sûrement en raison de
l'effort de conservation dans la zone d'étude malgré une forte
pression de braconnage. Ce taux est relativement important si l'on compare aux
autres sites semblables de la région du Sud-est Cameroun. Sur le
même site en 2002, des IKA plus élevés avaient
été relevés pour les espèces comme le gorille, le
chimpanzé, le buffle, les petits singes. Ceux de
l'éléphant, des Suidées et du bongo sont en augmentation
(Nzooh et al., 2002) (Tableau 46).
Tableau 46: Abondance relative (IKA) des grands et
moyens mammifères dans quelques sites du Sud-est Cameroun
|
Site
|
Eléphant
|
Gorille
|
Chimpanzé
|
Buffle
|
Bongo
|
Petits
singes
|
Suidée
|
Toute la faune
|
Auteurs
|
|
Dja
|
0,93
|
0,22
|
0,27
|
0,06
|
0,01
|
|
|
2,03
|
Nzooh (2001)
|
|
Forêt de Ngoila-Mintom
|
3,6
|
1,6
|
0,27
|
0,05
|
0,025
|
0,792
|
1,49
|
20,37
|
Nzooh (2003)
|
|
ZICGC N°1, 2, 3, 8 et 9
|
0,5-2,18
|
1,4-4,95
|
0,03-0,24
|
0,01-0,30
|
0-1,29
|
0,55-0,806
|
0,48-1,35
|
10,98-45,71
|
Nzooh et al. (2002)
|
|
PN Lobéké
|
18,38
|
3,23
|
0,33
|
0,406
|
0,945
|
0,682
|
2,072
|
47,037
|
Nzooh (2003)
|
|
UFA 10-018
|
0,37
|
0,73
|
0,15
|
0,07
|
0,08
|
0,35
|
1,28
|
9,84
|
Nzooh et al. (2003)
|
|
UFA 10-052
|
0,616
|
0,632
|
0,022
|
|
|
0,271
|
0,674
|
14,346
|
Nzooh et Bassama (2003)
|
|
UFA 10-022
|
4,292
|
3,047
|
0,171
|
0,07
|
0,053
|
0,167
|
3,38
|
28,93
|
Nzooh et al. (2004a)
|
|
UFA 10-013
|
3,917
|
3,373
|
0,21
|
0,713
|
0,535
|
0,559
|
0,567
|
19,95
|
Nzooh et al. (2004b)
|
|
UFA 10-051
|
0,05
|
0,01
|
0,05
|
-
|
0,41
|
0,11
|
0,44
|
15,85
|
Bobo et al. (2006a)
|
|
UFA 10-053
|
-
|
0,51
|
0,02
|
-
|
-
|
0,39
|
1,76
|
21,21
|
Bobo et al. (2006b)
|
|
UFA 10-013
|
1,37
|
2,27
|
0,11
|
0,34
|
0,6
|
0,5
|
0,97
|
21,26
|
Bobo (2009a)
|
|
UFA 10-025
|
0,13
|
0,19
|
0,02
|
-
|
-
|
0,05
|
0,3
|
19,93
|
Bobo (2009b)
|
|
UFA 10-063
|
2,71
|
1,70
|
0,079
|
0,25
|
0,97
|
0,28
|
1,48
|
20,65
|
Bobo (2009c)
|
|
PNBB, ZICGC 13 et 14
|
1,815
|
1,556
|
0,398
|
0,435
|
0,667
|
0,240
|
-
|
41,935
|
Présente étude
|
Il ressort du tableau 46 que
l'abondance relative totale de la faune pour la présente étude
d'étude (IKA = 41,935) se rapproche à celle obtenue par Nzooh
et al. (2002) dans la ZICGC 9 (IKA = 45,71) et à celle de Nzooh
(2003) dans le PN Lobéké (IKA = 47,037). Cette valeur
s'avère plus petite que les deux autres pour des raisons sûrement
liées aux braconnages qui ont pris de l'ampleur ces dernières
années, malgré l'effort de conservation consenti par les
organismes en charge de la gestion de la faune. Comparées à
d'autres sites, les densités obtenues pour les céphalophes roux
et céphalophes bleus sont plus importantes (Tableau 47).
Tableau 47: Densité des grands et moyens
mammifères dans quelques sites du Sud-est Cameroun
|
Site
|
Eléphant
|
Gorille
|
Céphalophe à dos Jaune
|
Céphalophe roux
|
Céphalophe bleu
|
Auteurs
|
|
Parc National de Lobéké
|
0,96
[0,81-1,14]
|
1,59
[1,13-2,24]
|
1,19
[0,85-1,66]
|
11,04
[9,28-13,15]
|
3,12
[2,54-3,84]
|
Nzooh (2003)
|
|
Parc National de Boumba Bek et Nki
|
0,3
[0,23-0,37]
|
0,9
[0,7-1,1]
|
0,85-1,7
|
5,3-11
|
0,4-3,7
|
Ekobo (1998)
|
|
UFA 10-052
|
-
|
-
|
-
|
1,36
[0,80-2,33]
|
0,91
[0,54-1,54]
|
Nzooh et Bassama (2003)
|
|
UFA 10-022
|
0,092
[0,036-0,232]
|
5,81
[2,59-13,01]
|
10,90
[7,53-15,79]
|
4,58
[3,03-6,93]
|
7,66
[5,72-10,25]
|
Nzooh et al. (2004a)
|
|
UFA 10-013
|
0,356
[0,212-0,6]
|
4,72
[2,79-7,98]
|
-
|
15,47
[8,18-29,24]
|
2,92
[1,62-5,27]
|
Nzooh et al. (2004b)
|
|
UFA 10-051
|
-
|
-
|
-
|
1,73
[130-2,29]
|
1,91
[1,37-2,64]
|
Bobo (2006a)
|
|
UFA 10-053
|
-
|
1,44
[0,67-3,06]
|
2,6
[1,55-4,34]
|
2,17
[1,49-3,09]
|
1,39
[0,98-1,97]
|
Bobo (2006b)
|
|
UFA 10-013
|
0,077
[0,044-0,134]
|
2,82
[1,91-4,18]
|
-
|
2,51
[1,83-3,45]
|
1,38
[0,88-2,19]
|
Bobo (2009a)
|
|
UF1 10-025
|
-
|
-
|
-
|
0,74
[0,49-1,11]
|
0,58
[0,35-0,96]
|
Bobo (2009b)
|
|
UF1 10-063
|
0,28
[0,19-0,43]
|
0,82
[0,54-1,24]
|
1,31
[0,81-2,12]
|
4,9
[3,58-6,73]
|
3,53
[2,28-5,47]
|
Bobo (2009c)
|
|
PNBB, ZICGC 13 et 14
|
-
|
-
|
-
|
2,946
[1,346-6,676]
|
5,227
[2,582-10,606]
|
Présente étude
|
Comparées à d'autres sites, les WPI obtenus par
images photographiques des certaines espèces sont plus importantes
(Tableau 48).
Tableau 48: WPI des grands et moyens mammifères
dans quelques sites du Sud-est Cameroun
|
Sites
|
Céphalophe à dos jaune
|
Céphalophe bleu
|
Céphalophe roux
|
Chimpanzé
|
Eléphant
|
Gorille
|
Potamochère
|
Auteurs
|
|
UFA 10 038
|
0,4
|
3,6
|
0,6
|
0,1
|
-
|
0,7
|
0,1
|
Nkwetaketu (2011)
|
|
PNBB, ZICGC 13 et 14
|
3,06
|
28,52
|
10,74
|
0,1
|
0,74
|
0,74
|
1,94
|
Présente étude
|
Il ressort du tableau 48 que les valeurs de WPI de
chimpanzé et de gorille de la présente étude corroborent
avec celles obtenues par Nkwetaketu (2011), pour des raisons liées
à la reproduction et au prélèvement illégal. En
effet, la taille de population chez les primates est très faible et le
rythme de reproduction est lent avec une mise bat par an et une longue
période de gestation qui dure environ neuf mois. Par contre chez les
céphalophes, la vitesse de reproduction est importante malgré le
braconnage.
4.1.6
Quelques vitesses de dégradation des crottes des espèces
identifiées dans la zone d'étude
Lors des inventaires de la faune dans la zone d'étude,
toutes les crottes très fraîches (âge 0) des espèces
ont été identifiées et suivies jusqu'à
décomposition complète. Le tableau 49 présente la moyenne
de vitesse de décomposition des crottes de quelques espèces de la
faune rencontrées dans la zone d'étude.
Tableau 49: Vitesse moyenne de décomposition
des crottes de quelques espèces dans la zone d'étude
|
Nom Français
|
Nombre de crotte
|
Vitesse de dégradation (j)
|
|
Céphalophe bleu Cephalophus monticola
|
25
|
16,04#177;2,79
|
|
Céphalophe roux*
|
05
|
21,20#177;1,79
|
|
Chat sauvage Nandinia binotata
|
02
|
26,50#177;16,26
|
|
Pangolin à longue queue
|
02
|
8#177;4,24
|
|
Athérure africain Atherurus africanus
|
01
|
5#177;0,0
|
* ici regroupe deux espèces le Céphalophe bai
Cephalophus dorsalis et le Céphalophe de Peter's
Cephalophus callipygus
Il ressort du tableau 49 que 25 crottes de Céphalophes
bleus ont été identifiées dans toute la zone
d'étude, soit une vitesse moyenne de dégradation estimée
à 16,04#177;5,64 jours (environ 16 jours). Ces résultats
corroborent avec ceux déterminés par Koster et Hart (1988) en
forêt dense dans la République Démocratique du Congo avec
une vitesse de décomposition de 18 jours. Cinq crottes de
Céphalophes roux ont été suivies jusqu'à
disparition complète avec une moyenne de dégradation
estimée à 21,20#177;1,79 jours. Ces résultats corroborent
avec ceux déterminés par Koster et Hart (1988) en forêt
dense dans la République Démocratique du Congo avec une vitesse
de décomposition de 21 jours. Par ailleurs le nombre de crottes de chat
sauvage, de Pangolin à longue queue et de l'athérure africain
était très peu représenté dans la zone
d'étude avec respectivement, 2, 2 et une crotte par espèce ce qui
nous a permis de calculer malgré un nombre insuffisant leur vitesse de
dégradation comme consigné dans le tableau 46.
4.1.7
Stade de décomposition des crottes
La vitesse de décomposition des crottes
s'avèrent indispensables pour le calcul des densités des animaux.
Le rythme de pourriture des crottes varie largement en fonction des zones et
des saisons. Les figures 53, 54, 55, 56 et 57 illustrent le stade de
décomposition des crottes de céphalophe bleu à
Gouonepoun.
A l'âge 0 (0 jour), toutes les crottes ont une couleur
gris luisante, elles sont tendres, humides et très odorantes. Elles sont
décrites comme étant très fraîches (Figure 53).
Arrivé au 5ème jour, les crottes prennent une couleur
gris terne, leur odeur devient de moins en moins perceptible et la crotte
commence à se décomposer sous l'action des termites et fourmis
(dégradation de 20-25% de la crotte) et la crotte est décrite
comme étant récente (Figure 54). Au fur et à mesure que le
temps passe (10ème jour), la crotte perd complètement
son odeur et la décomposition s'accélère davantage sous
l'action des champignons et des larves d'insectes (crottes
dégradées de 75-90%) et la crotte est décrite comme
étant Vieille (Figure 55). Au 15ème jour, la
décomposition s'achève peu à peu (crotte
dégradée à 95%) et la crotte est décrite comme
étant très vieille (Figure 56). Ensuite la décomposition
est totale (crotte dégradée à 100%) lorsque la motte de
terre a réduit considérablement, la crotte est décrite
comme disparue (Figure 57).
Figure 50: Crotte
fraîche (0 jours)
IRAD


Figure 51: Crotte vieille
(10ème jour)
Figure 52: Crotte
très vieille (15ème jour)


Figure 53: Crotte
disparue

4.1.8
Caractérisation de la végétation le long des transects
dans la zone d'étude
4.1.8.1 Inventaires des ligneux le long des transects dans la
zone d'étude
Les résultats des inventaires des ligneux le long des
transects à Gribé (forêt secondaire) et à Gouonepoun
(forêt primaire), ont permis d'identifier 30 espèces
végétales appartenant à 20 familles avec un
Diamètre à Hauteur de Poitrine (DHP) supérieur à 10
cm. Le tableau 50 présente les espèces végétales
recensées le long des transects à Gribé et à
Gouonepoun.
Tableau 50: Quelques ligneux recensés le long
des transects à Gouonepoun et à Gribé
|
Famille
|
Nom usuel
|
Nom scientifique
|
|
Gouonepoun
|
Bombacaceae
|
Baobab
|
Adansonia digitata
|
|
Mimosaceae
|
Dabéma
|
Piptadeniastrum africanum
|
|
Apocynaceae
|
Emien
|
Alstonia boonei
|
|
Fabaceae
|
Padouk rouge
|
Pterocarpus soyauxii
|
|
Combretaceae
|
Fraké
|
Terminalia superba
|
|
Annonaceae
|
Ebon
|
Anonidium mannii
|
|
Burséraceae
|
Aiélé
|
Canarium schweinflurtili
|
|
Dilleniaceae
|
Liane à eau
|
Tetracera alnifolia
|
|
Euphorbiaceae
|
Njansang
|
Ricinodendron heudelotii
|
|
Meliaceae
|
Sapelli
|
Entandrophragma cylindricum
|
|
Meliaceae
|
Tiama
|
Entandrophragma angolense
|
|
Annonaceae
|
Moambé jaune
|
Enantia chlorantha
|
|
Caesalpiniaceae
|
Abem nlong
|
Gilbertiodendron grandiflorum
|
|
Sterculiaceae
|
Ayous
|
Triplochyton scleroxylon
|
|
Ochnaceae
|
Azobé
|
Lophira alata
|
|
Fabaceae
|
Padouk blanc
|
Pterocarpus mildbraedii
|
|
Rutaceae
|
Bongo Yokadouma
|
Fagara welwitschii
|
|
Sterculiaceae
|
Eyong
|
Eribroma oblongum
|
|
Gribé
|
Urticaceae
|
Parasolier
|
Musanga cecropioides
|
|
Ulmaceae.
|
Kekelé
|
Holoptelea grandis
|
|
Caesalpiniaceae
|
Nganga
|
Cynometra hankei
|
|
Annonaceae
|
Moambé blanc
|
/
|
|
Malvaceae
|
Kola
|
Cola acuminata
|
|
Rhizophoraceae
|
Palétuvier
|
Rhizophora racemosa
|
|
Sapotaceae
|
Mukulungu
|
Autranella congolensis
|
|
lrvingiaceae
|
Andok
|
Irvingia gabonensis
|
|
Fabaceae
|
Assamela
|
Pericopsis elata
|
|
Mimosaceae
|
Okan*
|
Cylicodiscus gabonensis
|
|
Rubiaceae
|
Johimbe*
|
Pausinystalia johimbe
|
|
Fabaceae
|
Pao rosa*
|
Swartzia fistuloides
|
*= espèce rencontrée à la fois à
Gouonepoun et à Gribé
Sur les 12 placettes installées à Gouonepoun, la
végétation est caractérisée par une canopée
fermée soit un degré de recouvrement moyen de (85#177;8,8)% par
transect avec un sous-bois clair. Les espèces les plus
rencontrées sur les transects étaient constituées pour la
plupart des Fraké Terminalia superba soit une proportion de
33,33%. Par contre à Gribé (UFA 10 018), la canopée est
ouverte, avec un degré de recouvrement moyen par transect de
(25#177;12,9)% La plupart des ligneux étaient constitués par les
tiges d'avenir de faible diamètre et de palétuviers soit une
proportion de 66% sur les transects. Les graines et les fruits secs de beaucoup
d'espèces végétales de la zone sont exploités et
commercialisés par la population locale. On peut citer Irvingia
gabonensis, Ricinodendron heudelotii, Gnetum africanum, Afromomum dalzeillii,
Cola acuminata. Le Johimbé Pausinystalia johimbe est
utilisé dans la pharmacopée, jouant le rôle de
d'aphrodisiaque dans la zone.
4.1.8.2 Inventaires des herbacées le long des
transects dans la zone d'étude
A l'intérieur des placettes, quelques espèces
herbacées ont été identifiées dans la zone
d'étude. Parmi les herbacées les plus représentées,
on distingue les feuilles de marantacées, les fougères, certains
herbes de la famille des Euphorbiaceae comme Alchornea
cordilolia, de la famille des Asteraceae comme Chromolaena
odorata et de la famille de Poaceae comme Setaria
megaphylla.
4.2
Evaluation de la performance des méthodes d'inventaires de la faune
testées
Les signes d'activités des espèces de grands et
moyens mammifères lors des inventaires sur les transects permanents
étaient de 26 espèces pour les inventaires diurnes indirects
(Tableau 10), de 18 espèces pour les inventaires diurnes directs
(Tableau 37), de 17 espèces pour les inventaires nocturnes directs
(Tableau 38) et de 24 espèces capturées par les caméras
infra-rouges (Tableau 45). Les densités des différentes
espèces ont été calculées par le Programme DISTANCE
6.0, pour ce qui était des inventaires diurnes indirects et des
inventaires nocturnes directs. En ce qui concerne les inventaires diurnes
directs, seules les abondances relatives ont été estimées
car, le nombre des observations (Tableau 32) était très petit
pour utiliser le Programme DISTANCE 6.0. Les densités des espèces
capturées au moyen des caméras infra-rouges ont été
calculées (Tableau 43), ainsi que le l'indice photographique de la faune
(WPI) (Tableau 44). La vitesse de marche pour les inventaires diurnes
était environ de 2,00#177;1,50 km/h, par contre dans la nuit, la vitesse
de marche était estimée à 2,35#177;1,23 km/h. La
différence entre ces deux valeurs s'expliquerait par le fait que, dans
la nuit, tous les 50m le long des transects, on s'arrêtait pour suivre
les cris des différents animaux et pouvoir les identifiés. En
outre, seules trois méthodes dont les densités de quelques
espèces ont été estimées entre autre, la
méthode indirecte, la méthode nocturne et la méthode au
moyen de caméra infra-rouge, rendant impossible l'estimation de la
performance des méthodes par les densités. Cependant,
l'estimation de performance des méthodes testées a
été déterminée par le calcul de coefficient de
variation de l'abondance relative (IKA et WPI) des toutes les espèces
recensées dans les trois sites. Le tableau 51 présente les IKA et
WPI des espèces recensées par type de méthode.
Tableau 51: IKA et WPI des espèces
recensées par type de méthode testées dans la zone
|
Espèce
|
Inventaire diurne
indirect
|
Inventaire diurne direct
|
Inventaire nocturne
|
Camera infra-rouges
|
|
ni
|
IKA
|
ni
|
IKA
|
ni
|
IKA
|
Pi
|
WPI
|
|
Bongo
|
72
|
0,667
|
6
|
0,056
|
0
|
0
|
8
|
0,741
|
|
Buffle
|
47
|
0,435
|
0
|
0
|
0
|
0
|
2
|
0,185
|
|
Céphalophe à dos jaune
|
183
|
1,694
|
0
|
0
|
0
|
0
|
33
|
3,056
|
|
Céphalophe bleu
|
1028
|
9,519
|
6
|
0,056
|
133
|
1,231
|
308
|
28,519
|
|
Céphalophe roux
|
1223
|
11,324
|
8
|
0,074
|
20
|
0,185
|
116
|
10,741
|
|
Chat sauvage
|
8
|
0,074
|
1
|
09
|
1
|
09
|
3
|
0,278
|
|
Chevrotain aquatique
|
8
|
0,074
|
0
|
0
|
0
|
0
|
2
|
0,185
|
|
Chimpanzé
|
43
|
0,398
|
12
|
0,111
|
12
|
0,111
|
1
|
0,093
|
|
Civette
|
5
|
0,046
|
0
|
0
|
42
|
0,389
|
7
|
0,648
|
|
Daman des arbres
|
0
|
0
|
0
|
0
|
805
|
7,454
|
1
|
0,093
|
|
Eléphant
|
196
|
1,815
|
2
|
0,019
|
5
|
0,046
|
8
|
0,741
|
|
Petits singes
|
26
|
0,241
|
331
|
3,065
|
29
|
0,269
|
91
|
8,426
|
|
Antilope de Bates
|
36
|
0,333
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Gorille
|
168
|
1,556
|
2
|
0,019
|
15
|
0,139
|
8
|
0,741
|
|
Oryctérope
|
8
|
0,074
|
0
|
0
|
0
|
0
|
6
|
0,556
|
|
Pangolin à longue queue
|
315
|
2,917
|
3
|
0,028
|
18
|
0,167
|
7
|
0,648
|
|
Pangolin géant
|
100
|
0,926
|
0
|
0
|
0
|
0
|
3
|
0,278
|
|
Léopard
|
2
|
0,019
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Porc épic
|
626
|
5,796
|
1
|
09
|
10
|
0,093
|
33
|
3,056
|
|
Potamochère
|
400
|
3,704
|
5
|
0,046
|
10
|
0,093
|
21
|
1,944
|
|
Potto Bosman
|
0
|
0
|
0
|
0
|
5
|
0,046
|
0
|
0
|
|
Sitatunga
|
36
|
0,333
|
0
|
0
|
0
|
0
|
0
|
0
|
|
TOTAL
|
4529
|
41,935
|
377
|
3,491
|
1105
|
10,231
|
685
|
63,426
|
ni = nombre total d'indice de presences d'une
espèce i sur les 18 transects
Pi = nombre total de photographie prise pour une
espèce i par les cameras de surveillance
Il ressort du tableau 51 que la méthode d'inventaire
diurne indirecte présente un grand nombre d'indice de présence
soit n = 4529 (IKA = 41,935 indice au km). Cette valeur est plus importante,
pour des raisons liées à la présence incontestable des
pisteurs Baka et Konabembé expérimentés, qui tout au long
de cette étude nous ont permis de pister les animaux dont leurs signes
étaient bien visibles sur le sol. Le tableau 52 présente les
coefficeints de variation des différentes méthodes.
Tableau 52: Coefficient de
variation des méthodes d'évaluation de l'abondance de la
faune
|
IKA/WPI
|
|
Méthodes d'inventaire
|
SD
|
u
|
CV (%)
|
|
Méthode diurne directe*
|
3,492
|
0,650
|
18,609
|
|
Méthode diurne indirecte*
|
3,126
|
41,945
|
7,453
|
|
Méthode nocturne*
|
1,584
|
10,232
|
15,476
|
|
Caméra infra-rouge directe**
|
6,386
|
60,929
|
10,481
|
* = méthode utilisant l'IKA ; **= méthode
utilisant le WPI
Il ressort du tableau 52 que, la méthode diurne directe
(CV = 18,609) basée sur des observations directes s'avère moins
performante dans la zone d'étude, car il est très difficile
d'observer les animaux, parce que, ces derniers sont très sensibles aux
bruits et aux odeurs corporelles provoqués par la présence des
humains. Cette méthode oblige une attention toute particulière
à cause du champ visuel très limité par la densité
de la végétation. Les inventaires nocturnes (CV = 15, 476)
présentent un avantage de détecter les animaux dont leurs
activités sont beaucoup plus difficiles à observer en
journée comme la civette, le daman des arbres, le Potto de Bosman. Les
camera infra-rouges (CV = 10,481) sont des engins de captures passives, seules
les cibles (animaux) se déplaçant dans le champ de vision de
l'objectif (caméra) sont capturées, ce qui traduit une bonne
certitude de détection. Parmi les avantages qu'offrent les pièges
photographiques, la détection des espèces rares demeurent la plus
importante. Ainsi, la méthode qui a été retenue pour
l'estimation des abondances relatives des grands et moyens mammifères
est celle des inventaires diurnes indirects (CV = 7,481%), car
cette dernière présente la plus petite valeur de coefficient de
variation. Ces résultats corroborent avec ceux déterminés
par (Burnham et al., 1980; Buckland et al., 1993; 2001; White
et Edwards, 2000) qui stipule que la méthode d'inventaire diurne
indirecte sur transects linéaires reste cependant très
utilisée en zone de forêt. Par contre, Viquerat et
al. (2012), dans le Parc National de Korup révélait que
sur trois méthodes testées à savoir, la méthode
diurne directe, diurne indirecte basée sur l'analyse des crottes et
nocturne, la méthode la plus performante était celle des
inventaires diurnes directs.
4.3
Détection des espèces rares
L'observation des animaux en forêt s'avère
difficile, car la végétation est très dense, les bruits et
les odeurs dégagées par les hommes lors de leur
déplacement provoquent la fuite des animaux. Cependant, les
caméras infra-rouges dotées d'un système de capture
relativement passive et sans aucun contact avec l'animal capturé, permet
de détecter avec précision un animal donné et dans un
habitat donné. Le tableau 53 présente les espèces rares
détectées par les camera infra-rouges
Tableau 53: Liste des espèces rares
détectées au moyen des caméras infra-rouges
|
|
Nom Français
|
Nom anglais
|
Nom scientifique
|
Statut1
|
|
Primates
|
|
Chimpanzé
|
West Afican Chimpanzee
|
Pan troglodytes
|
A
|
|
|
Gorille
|
Gorilla
|
Gorilla g, gorilla
|
A
|
|
|
Magistrat
|
Black and white Colobus
|
Colobus guereza
|
A
|
|
|
Cercocèbe agile
|
Agile Mangabey
|
Cercocebus agilis
|
A
|
|
Proboscidea
|
|
Eléphant de forêt
|
Forest elephant
|
Loxodonta a. cyclotis
|
A/B
|
|
Artiodactiles
|
|
Oryctérope
|
Aardvark
|
Orycteropus afer
|
A
|
|
Tragulidae
|
|
Chevrotain aquatique
|
Water Chevrotain
|
Hyemoschus aquaticus
|
A
|
|
Bovidae
|
|
Céphalophe à dos jaune
|
Yellow-backed Duiker
|
Cephalophus silvicultor
|
A
|
1Au Cameroun, la classe A regroupe les
espèces rares ou en voie de disparition qui bénéficient
d'une protection intégrale, la classe B regroupe les espèces
à protection partielle et la classe C les espèces qui ne
bénéficient d'aucune protection.
Il ressort du tableau 53 que les espèces rares ou en
voie de disparition capturée par les caméras infra-rouges
appartenaient à quatre familles avec huit espèces (Annexe 7).
4.4
Formulation de quelques suggestions afin d'orienter le modèle de gestion
de la ressource faune sauvage dans la zone d'étude
Compte tenu de l'importance de la zone aux plans
écologiques et économiques, et des menaces auxquelles elle doit
faire face, trois axes suivants ont être développés afin
d'orienter le modèle de gestion de la ressource faune sauvage.
4.4.1
Information, éducation et Communication efficace au profit de la
communauté locale
La communauté locale n'est véritablement
sensibiliser sur la question de conservation des ressources naturelles. Les
menaces qui pèsent sur la faune s'expliquent par un manque
d'information sur l'éducation environnementale, la méconnaissance
du cadre réglementaire en matière de conservation de la
biodiversité par les populations riveraines leur permettant de jouir
efficacement de leur droit d'usage. Pour une gestion durable des ressources
naturelles dans la zone, il est important de :
- Organiser des campagnes d'information et de sensibilisation
sur la protection des espèces menacées de disparition via les
dépliants et les posters ;
- Elaborer et mettre en oeuvre une stratégie sur
l'éducation environnementale des jeunes et des femmes;
- Enseigner dans les écoles villageoises le bienfait de
la faune et les conséquences sur l'extinction des animaux;
- Désigner et nommés les chefs traditionnels de
la zone comme éco gardes locaux et les impliquer tous dans le processus
de conservation et la protection des aires protégées, car toutes
les décisions venant de ce dernier devrait être
considéré par les villageois;
- Organiser des concours inter village et motiver les villages
qui protégeraient mieux ou ayant réussi à sensibiliser les
populations dans la protection des ressources naturelles, après une
analyse des données collectées sur le terrain;
- Organiser les discussions avec la communauté locale
de la zone sur la question des espèces chassées et les techniques
de la chasse villageoise.
4.4.2
Lutte anti-braconnage
Malgré les efforts consentis par les organismes en
charge de la faune dans la région comme le la GTZ, le WWF et d'autres
ONGs nationales, depuis plus d'une décennie n'ont pas pu renverser les
tendances, mais plutôt ces dernières années, on assiste
à une recrudescence du grand braconnage et du sciage sauvage.
Cependant pour préserver la faune et son habitat, la
lutte anti-braconnage/anti-sciage sauvage est une nécessité. Ceci
passe d'abord par:
- Fermeture de toutes les routes forestières
après exploitation notamment à Gribé par exemple où
les routes sont à la merci des braconniers, facilitant rapidement
l'accès dans les aires protégées;
- Renforcement du contrôle à l'accès aux
armes et munitions dans la zone. Il s'agit de constituer un agent de
renseignement secret par village afin de démenteler tout le
réseau de traficants des armes dans la zone, ainsi que les villageois
qui hebergéraient les braconniers allogènes;
- Création dans chaque village un poste de
contrôle fixe et opérationnel opérant de 5h à 20h et
impliquer le chef traditionnel dans le processus de contrôle des produits
issus de la chasse villageoise et leurs dérivés. Il en est de
même avec les barrières fixes où les heures d'ouverture et
de fermeture doivent être strictement respectées, notamment la
barrière de Ngato;
- création des emplois dans le cadre de la gestion des
activités des Parcs et de ZIC/ZICGC. Il s'agit d'organiser la
communauté locale en GIC, puis signer les contrats entre cette
dernière et l'administration forestière en vue de
développer des micro projets autour des aires protégées
concernées, ainsi le partage des revenus sont équitables entre
les deux parties ;
- Constituer des brigades mobiles d'intervention
multisectorielle, opérant avec une unité mixte entre autres les
forestiers, les forces de l'ordre nationale (gendarmes et les millitaires);
- Faciliter le processus de la pratique de la chasse
traditionnelle ou droit d'usage qui permet aux populations de prélever
le petit gibier autour des villages pour leur alimentation;
- Une concertation entre toutes les parties prenantes
(Concessionnaires de toutes les UFA voisines Alpicam-Grumam, STBK, ...), Chefs
de postes forestiers et de chasses, Conservateur du PNBB, ONG locales et
internationales (WWF-E, GTZ), Guides de chasse de toutes les ZIC et ZICGC
voisines, Populations riveraines, Confessions religieuses, Autorités
administratives, Forces de l'ordre, ...) par exemple lors d'un atelier à
l'issue duquel des engagements formels et concrets doivent être pris afin
de juguler ce fléau, du moins au sein des sociétés
forestières;
- Promotion de l'élevage non conventionnel pour
minimiser le braconnage. Par exemple l'élevage de lapin très
prolifique permettra à la population de subvenir au besoin en proteine,
il faut promouvoir aussi les élevages des poissons car la zone dispose
des cours d'eau potentielle par exemple la Boumba et le Bek très riches
en poissons;
- Des patrouilles mixtes de reconnaissance doivent être
régulièrement effectuées dans la zone ;
- Les braconniers et leurs managers (fournisseurs d'armes et
des munitions) doivent être sévèrement sanctionnés
et condamnés par le tribunal et les peines réellement
appliquées.
4.4.3
Suivi permanent de la faune
Toutes ces actions devront être préparées,
exécutées et suivies sur le terrain. Un bilan annuel devra
être établi et les priorités devront être
redéfinies et réorientées. Plus spécifiquement il
s'agira de:
- Collecter une fois tous les trois mois les données
relatives au suivi de la faune sauvage (inventaires, suivi de
fréquentation des sites particuliers, taux de pourriture, taux de
defécation...) dans la zone, par la méthode habituelle distance,
sur les transects permanents prédefinis ;
- Etablir une base de données et un système SIG
afin de pouvoir suivre l'évolution des populations animales et des
activités de braconnage dans la zone ;
- Etablir une base de données afin d'exploiter les
informations sur le braconnage à collecter au niveau des
barrières d'entrée et/ou de sortie de L'UFA;
- Si le parcours ci-dessus proposé est
réalisé, il faudrait ensuite faire un inventaire complet de tous
les trois sites par la méthode distance une fois tous les cinq ans afin
d'apprécier, d'évaluer et de ré-orienter les efforts de
conservation entrepris, l'inventaire de 2002 (Nzooh et al.,
2002) servant de référence ;
- Si le parcours ci-dessus proposé n'est pas
réalisé, nous recommandons de faire un inventaire complet par la
méthode distance une fois tous les deux ans pour les mêmes
fins.
CHAPITRE 5: CONCLUSION ET RECOMMANDATIONS
5.1
Conclusion
La présente étude fait ressortir la
variation des densités et des abondances relatives des espèces de
grands et moyens mammifères dans le PNBB et sa périphérie
Nord-est représentée par les villages Gribé et Gouonepoun,
situé dans la région de l'Est Cameroun. L'inventaire faunique
réalisé sur les transects linéaires des trois sites (PNBB,
Gribé et Gouonepoun) suivant quatre méthodes (méthode
diurne indirecte, méthode diurne directe, méthode nocturne
directe et les caméras infra-rouges) a permis de recenser une richesse
spécifique estimée à 28 espèces de grands et moyens
mammifères sur un ensemble de 18 transects pour une distance totale de
108 km. Les inventaires diurnes indirects ont présenté, une
différence significative entre les sites pour le céphalophe
à dos jaune, le buffle, le pangolin à longue queue et le pangolin
géant. Seules les densités des céphalophes bleus et
céphalophes roux ont été estimées par le Programme
DISTANCE 6.0, entre les trois sites, mais aucune différence
significative n'a été observée dans les trois sites. Pour
ce qui était des inventaires diurnes directs, seules les densités
des petits singes ont été calculées par le Programme
DISTANCE 6.0, dans les trois sites. Seules les densités des
céphalophes bleus lors des inventaires nocturnes ont été
estimées par le Programme DISTANCE 6.0, aucune différence
significative observée dans les trois sites. Douze caméras ont
été installées dans chaque site, soit un total de 578
captures de toutes les espèces de faune dans la zone d'étude. Le
WPI a été estimée pour toutes les espèces de la
faune. Le calcul des densités et de WPI des différentes
espèces n'ont présenté aucune différence
significative entre les sites. Cinq espèces de faune dont les crottes
ont été suivies jusqu'à disparition complète,
à savoir céphalophes bleus, céphalophe roux, chat sauvage,
pangolin à longue et l'athérure africain. Trente espèces
végétales appartenant à 20 familles avec un
Diamètre à Hauteur de Poitrine (DHP) supérieur à 10
cm ont été comptées le long des transects permanents en
forêt secondaire et en forêt primaire. A Gouonepoun, les ligneux
les plus rencontrées sur les transects étaient constituées
pour la plupart des Fraké Terminalia superba et des Mukulungu
Autranella congolensis. Par contre à Gribé (UFA 10 018),
les ligneux étaient constitués par les tiges d'avenir de faible
diamètre et de palétuviers. Parmi les herbacés les plus
représentés, on distingue les feuilles de marantacées, et
quelques fougères
L'évaluation des méthodes d'inventaires
testés ont révélé que, la meilleure méthode
d'inventaire sur transect linéaire dans la zone d'étude est celle
des inventaires diurnes indirects basés sur les indices de
présence des grands et moyens mammifères.
Les espèces rares ou en voie de disparition
capturée par les caméras infra-rouges appartenaient à 4
familles avec huit espèces, dont le Gorille Gorilla g, gorilla,
le Chimpanzé Pan troglodytes, le Magistrat Colobus
guereza, le Cercocèbe agile, Cercocebus agilis,
l'Eléphant, Loxodonta africana, cyclotis, l'Oryctérope,
Orycteropus afer, le Chevrotain aquatique
Hyemoschus aquaticus et le Céphalophe à dos jaune
Cephalophus silvicultor
Compte tenu de l'importance de la zone, aux plans
écologiques et économiques, et des menaces auxquelles elle doit
faire face, trois axes ont été développés afin
d'orienter le modèle de gestion de la ressource faune sauvage, parmi
lesquels il faut:
- Privilégier l'information, l'éducation et la
communication au profit de la communauté locale;
- Promouvoir la lutte anti-braconnage;
- Envisager un suivi permanent de la faune.
5.2 Recommandations
Au terme de cette étude, nous recommandons:
AU MINFOF dans le but de réduire la
chasse illégale et le prélèvement des espèces
menacées constaté dans le PNBB et dans sa zone
périphérique :
Ø d'approfondir les connaissances et la
compréhension de toutes les interactions existantes entre la zone
périphérique et les populations riveraines afin de réduire
l'incidence des activités anthropiques sur les populations
animales ;
Ø procéder à une application effective
des sanctions en vigueur contre toute personne pris en flagrant délit et
contre les personnels de l'administration soupçonné de
corruption ;
Ø renforcer le dispositif de surveillance en mettant en
place des barrières de contrôles forestiers fixes et mobiles, en
permanence le long des axes : Ngato-Gribé-Song ancien et
Gouonepoun-Zoulabot ;
Ø mettre en place une unité commando
opérationnelle constituée par des forestiers et des militaires
afin d'assurer la surveillance au moins une fois tous les mois dans le Nord du
PNBB ;
Ø déployer les éco gardes afin de
détruire les ponceaux sur le cours d'eau POKPOK construit par les
braconniers au Sud de la ZICGC 13 qui relie la ZICGC 13 au PNBB.
A LA RECHERCHE dans le but d'améliorer
la connaissance sur le potentiel faunique :
Ø utiliser d'autres techniques d'inventaires dans la
zone d'étude, comme la capture au filet, le marché de la viande
de brousse afin de voir leur efficacité et de les comparer aux
méthodes déjà testées dans la zone
d'étude ;
Ø A cause du faible nombre des crottes suivies, la
vitesse de décomposition des crottes de certaines espèces n'a pu
être estimée à une valeur fiable comme pour le chat sauvage
(26,50#177;16,26), le pangolin à longue queue (8#177;4,24) et
l'athérure africain (5#177;0,0). Néanmoins, il faudra que la
recherche envisage de poursuivre l'étude sur la vitesse de
décomposition sur au moins 20 crottes, et le taux de
défécation des crottes des espèces citées. En
effet, si ces études fournissent des valeurs fiables, elles pourront
permettre un calcul raisonné des densités sur la base des crottes
dans la zone du PNBB et ses environs ;
Ø développer des programmes d'éducation
environnementale pour les jeunes afin de les sensibiliser dès le bas
âge sur la nécessité de la conservation de la
biodiversité ;
Ø installer les caméras infra-rouges le long des
pistes de débardages et sur les rives de cours d'eau à
Gribé, afin d'identifier avec précision les espèces rares.
A Gribé, le long des pistes de débardage, il faut mener les
études afin de savoir pourquoi tous ces animaux aiment marcher sur le
débardage.
AUX ONGs ET AUX EXPLOITANTS FORESTIERS pour
réduire la pression sur la faune
Ø développer les alternatives au braconnage et
mettre en place des dispositifs pour s'assurer de l'effectivité de ces
alternatives ;
Ø promouvoir l'élevage local des espèces
couramment consommées, comme l'extension de poulet de village et des
petits ruminants ;
Ø fermer toutes les routes forestières
après exploitation.
A JICA/FOSAS ET IRAD DE YAOUNDE d'approfondir
les recherches dans la section faune, en mettant sur pied une unité de
suivi permanent constituée d'un Ingénieur et des trois
Techniciens Supérieurs des Eaux, Forêts et Chasses. Cette
équipe sera, capable de :
Ø accroître de quatre le nombre des transects,
par site afin d'augmenter la probabilité de détection des indices
de faune et les crottes pour d'éventuelle suivie
(décomposition) ;
Ø continuer le suivi de décomposition des
crottes des éléphants à Gribé, les nids frais de
gorille et de chimpanzé identifiés dans le PNBB, afin d'estimer
les densités de ces derniers par leur valeur propre ;
Ø continuer la collecte des données sur le suivi
de la faune une fois tous les trois mois sur les transects permanents dans la
zone d'étude sur une durée d'au moins deux ans, afin d'avoir les
données beaucoup plus fiables.
AUX POPULATIONS LOCALES pour améliorer
leur perception, leur participation active à la protection des
ressources fauniques et leur condition de vie :
Ø s'impliquer activement dans les opérations de
protection et de conservation de la faune sauvage, ici les actions à
entreprendre sont par exemples, de limiter l'ouverture des pistes des chasses
et la pose des pièges, de limiter voir interdire l'utilisation de
matériels de chasse destructifs, de pratiquer une chasse
sélective et de réglementer la chasse villageoise surtout en
période de reproduction ;
Ø s'intéresser activement à l'agriculture
non seulement à celle de subsistance, mais aussi celle de
commercialisation comme par exemple le cacao, le café, le plantain qui
sont très productifs dans la zone d'étude, et s'avère de
plus en plus cher dans les centres urbains.
BIBLIOGRAPHIE
Adama, M. 2010. Etat des lieux et
perspectives de gestion durable de la chasse villageoise en
périphérie Nord-est du parc national de Korup,-Région du
Sud-ouest Cameroun Mémoire de fin d'études. FASA.
Université de Dschang. Cameroun. 73p.
AIDEnvironment. 1998. Initiative Jengi-
Esprit de la forêt. Analyse économique de la zone
forestière du Sud-Est du Cameroun. Rapport final provisoire.
AIDEnvironment, Amsterdam, 120P +annexes.
Barnes, R.F.W., Jensen, K.L. 1987. How to
count Elephant in forest IUNC, African Elephant and Rhino.
Specialist group Technical Bulletin 1: 1-6.
Barnes, R.F.W., Asamoah-Boeteng, B, Naada Majam, J;
Tchamba, M.N., Agyei-Ohemeng, J, Ekobo A, Nchanji, A. 1994.
Improving the accurency of forest elephants census methode: studies of dung
decay rates in Ghana and Cameroon., European Comission DG VIII: Ecology in
Developing Countries Programme.
Barnes, R.F.W., Beardsley, K., Michelmore, F., Alers,
M.P.T. Blom, A. 1997. Estimating forest elephant numbers with dung
counts and a geographic information system. Journal of Wildlife
Management 61: 1384-1393.
Besnard, A. 2010. Suivi scientifique
d'espèces animales. Aspects méthodologiques
essentiels pour l'élaboration de protocoles de suivis.
Laboratoire biologie et écologie des vertébrés, à
Montpellier.
Bobo, K.S. 2009a Statut des grands et moyens
mammifères, et des activités anthropiques dans l'UFA 10-013.
ALPICAM/GRUCM - JMN Consultant. 59p.
Bobo, K.S. 2009b Statut des grands et moyens
mammifères, et des activités anthropiques dans l'UFA 10-025.
SFIL/DECOLVENAERE - JMN Consultant. 56p.
Bobo, K.S. 2009c. Statut des grands et moyens
mammifères, et des activités anthropiques dans l'UFA 10-063.
ALPICAM/GRUCM - JMN Consultant. 61p.
Bobo, K.S., Tougoulou, J., Njie, M.F. 2006a.
Statut des grands et moyens mammifères, des activités
anthropiques et des oiseaux dans l'UFA 10-051. Cabinet JMN consultant. 51p.
Bobo, K.S., Tougoulou, J., Njie, M.F. 2006b.
Statut des grands et moyens mammifères, des activités
anthropiques et des oiseaux dans l'UFA 10-053. Cabinet JMN consultant. 54p.
Bobo, K.S. 2002. Bilan diagnostic de
l'état de la `'biodiversité'' dans quelques UFA du Sud Cameroun
(10-018, 10-015, 10-063, 10-011, 10-012, 10-047 et 10-037). Rapport de
consultation pour le WWF/SFM-C.
Buckland, S.T., Anderson, D.R., Burnham, K.P., Laake,
J.L. 1993. Distance sampling: estimating abundance of biological
populations. Champan and Hill, London.
Buckland, S.T., Anderson, D.R., Burnham, K.P., Laake,
J.L., Borchers, D.L., Thomas L. 2001. Introduction to distance
sampling: estimating abundance of biological populations. Oxford University
Press.
Burnham, K.P., Anderson, D.R., Laake, J.L. 1980
Estimation of density from line
transect sampling of biological populations. Wildlife
Monographs 72: 1-202.
Coopération Internationale en Recherche
Agronomique pour le Développement (CIRAD). 2006. Module 7
prendre en compte la faune sauvage dans l'aménagement. Formation de
forestier aménagiste et de gestionnaire forestier. Ecole Nationale des
Eaux et Forêts du Cap Estérias. Libreville,
Gabon.
Collart, N.J., Stuart, S.N. 1985. Threatened
birds of Africa and related islands. The ICPB / IUCN red data book. Part I.
ICPB, Cambridge, UK, and IUCN, Gland Switzeland.
Collen, B., Konie, J., Rist, J., Howard, R.
2008. Constructing the evidence for pygmy hippo conservation
in Sapo National Park, Liberia. Project progress report. Zoological
Society of London, Forestry Development Authority, Fauna and Flora
International. 19p.
Davenport, T. 1998. The Butterflies of
Lobéké reserve. Report WWF-CPO.
Davies, G., Heydon, M., Leader-Williams, N.,
Mackinnon, J., Newing, H. 2001. The effects of logging on tropical
forests ungulates. In The Cutting Edge: conserving wildlife in logged
tropical forests pp93-124. Fimbel, R.A., Grajal, A. & Robinson, J.G.
(eds) Columbia Press, New York.
Djafsia, K. 2012. Statuts des grands et
moyens mammifères dans les ZICGC 13 et 14 à la
périphérie Nord des parcs nationaux de Boumba-bek et Nki.
Rapport de stage Pré-insertion professionnel. FASA,
Université de Dschang. 61p.
Djetkeu, T.S. 2010. Evaluation du potentiel en cephalophes
en périphérie nord-est du parc national de korup, sud-ouest
Cameroun. Mémoire. FASA, Université de Dschang.51p
Doumenge, C., Garcia, J.J.E., Gartlan, S., Landgrand,
O., Ndinga, A. 2001. Conservation de la biodiversité
forestière en Afrique Centrale Atlantique : le réseau
d'aires protégées est-il adéquat. Bois et Forêts
des Tropicales 268: 5-27.
Dowsett-Lemaire, F., Dowsett, R.J. 2000.
Birds of the Lobéké Faunal Reserve, Cameroon, and its regional
importance for conservation. Bird Conservation International 10:
67-87.
Dowsett-Lemaire, F., Dowsett, R.J. 1999.
Zoological survey (birds, amphibians) of Lobéké Faunal Reserve in
April 1999, with special reference to Dja River Warbler Bradyptera grandis.
Final report by Tauraco a.s.b.l. for WWF-Cameroon. 25p.
Dowsett-Lemaire, F., Dowsett, R.J. 1997.
Enquête préliminaire sur la faune (en particulier l'avifaune) de
la réserve de Lobéké, Cameroun. Report WWF-CPO / Tauraco
a.s.b.l. 24p.
Dubost, G. 1983. Le comportement de
Cephalophus monticola Thunberg et C dorsalis Gray,et la place
des céphalophes au sein des ruminants. Mammalia 47: 141 -
177.
Eboko, A. 2006. Large mammals surveys in the
proposed Ebo protected Area.
Ekobo, A. 1998. Large mammals and
vegetation surveys in Boumba-Bek and Nki project area. WWF Cameroon
Internal Report. 63p. + annexes.
Ekobo, A. 1995. Conservation of the African
forest elephant (Loxodonta africana cyclotis) in Lobéké,
Southeast Cameroon. Ph. D. thesis, University of Kent 151p.
Elkan, P.W. 1994. A preliminary survey of
Bongo antelope assessment of Safari hunting in the Lobéké region
of south eastern Cameroon. Report, WCS/Cameroon
Griffiths, M. 1994. Population density of
Sumatran tigers in Gunung Leuser National Park. In:Sumatran Tiger
population and Habitat Viability Analysis Report. (Eds. R. Tilson, K.
Soemarna, W.Ramono, S. Lusli, K. Traylor-Holzer et U. Seal). Indonesian
Directorate of Forest Protection and Nature Conservation and IUCN/SSC
Conservation Breeding Specialist Group. Apple Valley, Minnesota, USA.
Gwet, J.-P. 2003. Enquête
socioéconomique et ethnobotanique sur l'état des lieux des
pro-duits forestiers non ligneux autour et dans la zone communautaire du Parc
National de Lobéké. Rapport, GTZ-PROFORNAT, Yokadouma 35p.
Harrisson, M., Agland, P. 1987. A draft
proposal for the designation of three new national forest parks. Dja River
Films, LTD.
Hart, J. A. 2000. Impact and sustainability
of indigeous hunting in the Ituri forest, Congo - Zaire: A comparison of
unhunted and hunted duiker populations. P 106 - 153 in J.G. Robinson and E.L
Bennet, editors. Hunting for sustainability in tropic forests.
Columbia University press, New York, USA.
Hecketsweiler, P.H., Boukong, M.B., Nsengue, B.E.,
François, J-R., Perthuisot, N. 2001. Étude
environnementale stratégique pour la planification et
l'aménagement de l'exploitation forestière dans l'UTO Sud-Est.
WWF/GTZ 160p.
Jell, B. 1998. Utilisation des produits
secondaires par les Baka et les Bangando dans la région de
Lobéké au Sud-Est Cameroun : Étude de cas. Rapport,
GTZ-PROFORNAT, Yokadouma, 40p.
Kenfack, D., Fimbel, R. 1995. Contribution
à l'étude des plantes médicinales de la réserve de
Lobéké : point sur la médecine traditionnelle
camerounaise. Rapport WCS / Cameroun.
Kengne, P. 2012. Statut des grands et
moyens mammifères à la périphérie nord du parc
national de Boumba-Bek, région du sud-est Cameroun, cas des ZICGC 13 et
14. Rapport de stage Pré-insertion
professionnel. FASA, Université de Dschang. 60p.
Koster, S.H., Hart, J.A. 1988. Methods of
estimating ungulate populations in tropical forests. African Journal of
Ecology 26: 117-126.
Laclavère, G. 1979. Atlas de la
république unie du Cameroun. Ed. Jeune Afrique, Paris. 72p.
Larrubia, J C., Arnhem. E. (2009)
Dénombrer la grande et moyenne faune mammalienne en milieu forestier.
Dans Triplet, P. (ed). Manuel de gestion des aires
protégées d'Afrique francophone Awely, Paris. 1234p.
Letouzey, R. 1985. Notice de la carte
phytogéographique du Cameroun au 1:500.000. Institut de la cartographie
internationale de la végétation, Toulouse, France.
Madzou, Y.C. 2003. Etat actuel des tendances
démographiques dans la region Nord de Boumba-Bek (Sud-Est Cameroun).
Rapport de recherche (phase 1 - Volume 2). Université Bordeaux 3.
48p.
Manet, B., Herman, R. 2003. Photographie
automatique et animaux à activité nocturne. Centre de
Recherche de la Nature, des Forêts et du Bois, Laboratoire de la Faune
sauvage et de Cynégétique, avenue Maréchal Juin, 23 B-5030
Gembloux.
Makazi, L.C., Usongo, L., Davenport, T. 1998.
Indigenous aquatic resource use in the proposed protected area of
Lobéké, south east Cameroon. Report WWF-CPO. 27p.
Meek, P., Ballard, G., Fleming, P. 2012. An
introduction to camera trapping for wildlife surveys in Australia. 94p.
Ministère de l'Environnement et des
Forêts (MINEF). 1995. Décrets n° 95/466/PM du 20
juillet 1995 et n° 95/531/PM du 23 août 1995 fixant les
modalités d'application du régime des Forêts et de la
Faune.
Ministère de l'Environnement et des
Forêts (MINEF). 2002. A Management Plan for
Korup National Park and its Peripheral Zone 2002-2007. WWF/EU/GTZ.
Ngandjui, G. 1998. Étude de la chasse
villageoise en vue de sa gestion durable : cas du site Sud-Est Cameroun.
Rapport, GTZ-PROFORNAT, Yokadouma 47p.
Nkwetaketu, N.I. 2011. A comparative
evaluation of camera traps and line transects as methods of survey for large
and medium sized fauna in the FMU 10 038 in East Cameroon.
Dissertation Submitted in partial fulfillment of requirements for the award of
a professional Master's (DESS) in Forest Science. University of Yaounde.
188p.
Nzooh-Dongmo, Z-L. 2001. Dynamique de la
faune sauvage et des activités anthropiques dans la Réserve de
Biosphère du Dja. Rapport ECOFAC. 75p.
Nzooh-Dongmo, Z-L., Koulbout, D., Bene Bene, L.,
Konje, P. 2002. Evaluation des potentialités fauniques des
zones d'intérêt cynégétique a gestion communautaire
(ZICGC) n° 1, 2, 3, 8 et 9 au Sud-est Cameroun. Rapport WWF CPO. 99p +
annexes.
Nzooh-Dongmo, Z-L. 2003. Suivi
écologique dans le Parc National de Lobéké et sa zone
périphérique : données de base sur la dynamique des
populations de grands et moyens mammifères et des activités
anthropiques. Rapport WWF/Jengi. 203p.
Nzooh-Dongmo, Z-L., Ngniado Wouala, A., Mahop, J.P.
2003. Statut des grands et moyens mammifères et des
activités humaines dans l'UFA 10-018, Rapport WWF CPO-SIBAF. 48p+
annexes.
Nzooh-Dongmo, Z-L., Bassama, C. 2003.
Evaluation préliminaire de l'environnement
bioécologique : Statut des grands et moyens mammifères dans
l'UFA 10-052. Rapport WWF CPO-Decolvenance. 48p + annexes.
Nzooh-Dongmo, Z-L., Mahop, J.P., Fouda, E. 2004a.
Statut des grands et moyens mammifères et des activités
humaines dans l'UFA 10-022. Rapport WWF CARPO-SCIFO. 50p.
Nzooh-Dongmo, Z-L., Bassama, C., Konje, P.
2004b. Statut des Grands et Moyens Mammifères et des
Activités Humaines dans l'UFA 10-013. Rapport WWF CCPO/JSEFP/CFE.
54p.
Nzooh-Dongmo, Z-L., Bené, B-L. 2005.
Suivi écologique dans le PNBB et sa zone
périphérique: données de base sur la dynamique des
populations des grands et moyens mammifères et des activités
anthropiques. 103p.
Poilecot, P. 2009 Dénombrer la grande
faune en milieu de savane: dans Triplet. P (ed), Manuel de
gestion des aires protégées d'Afrique francophone Awely, Paris,
1234p
Plumptre, A.J. 2000. Monitoring mammal
populations with line transect techniques in African forests. J. Appl.
Ecol. 37: 356-368.
Ramade, F. 1993. Dictionnaire
encyclopédique de l'écologie et des sciences de l'environnement.
EDISCIENCE International, Paris, 822p.
Rowcliffe, J.M., Carbone, C., Jansen, P.A., Kays, R.,
Kranstauber, B. 2011. Quantifying
the sensitivity of camera traps: an adapted distance sampling
approach. Methods in Ecology and Evolution 2, 464-476.
Simberloff, D. 1998 - Flagships, umbrellas, and
keystones: is single-species management in the landscape era? Elsevier,
247-257.
Silveira, L., Jacomo, A.T.A., Diniz, J.A.F.
2003. Camera trap, line transect census and track surveys: a
comparative evaluation. Biol. Conserv. 114:351-355.
Stromayer, K.A.K., Ekobo, A. 1991. Biological
survey of southeastern Cameroon. Report, WCI/IAR, MHESR. 40p.
Tchamba, N. M. 1992. Defecation by the
African forest elephant Loxodonta africana cyclotis
in the Santchou Reserve, Cameroon. Mammalia 56: 155-158.
Tutin, C.E.G., Fernandez, M. 1984. Nationwide
census of Gorilla (Gorilla g. gorilla) and Chimpanzee (Pan. t troglodytes).
Population in Gabon. American journal of primatology 6: 313-336
Union Internationale pour la Conservation de la Nature
(UICN). 1992. Protected areas of the world: A review of national
systems. IUCN, Gland Switzerland.
Union Internationale pour la Conservation de la Nature
(UICN). 1994. Lignes directrices pour les catégories de gestion
des aires protégées. Commission des parcs nationaux et des aires
protégées de l'Union Mondial.
Union Internationale pour la Conservation de la Nature
(UICN). 2000. IUCN Red List of Threatened Species. IUCN
Conservation Monitoring Center, Cambridge, UK.
Viquerat, S. M. A, Bobo, K. S., Mareike, M., Kiffner,
C., Waltert, M. 2012. Estimating Forest duiker (Cephalophinae) density
in Korup National Park: a case study on the performance of three line transect
methods. South African Journal of Wildlife Research 42:
1-10.
Waltert, M., Heber, S., Riedelbauch, S., Lien, J.L.,
Mühlenberg, M. 2006. Estimates of blue duiker densities from
diurnal and nocturnal line transects in the Korup region, SW Cameroon.
African Journal of Ecology 44: 290-292.
Wildlife Conservation Society (WCS). 1996.
The Lobeke forest, South-East Cameroon. Summary of activities period 1986-1995.
Report WCS/Yaounde. 217p.
Wildlife Conservation Society (WCS). 1995.
Integrated monitoring program for trans-boundary forest conservation and
management in Congo, Cameroon and the Central African Republic. Proceedings of
the Bomassa Workshop, September 1994.
White, L. 1994. Biomasse des
mammifères de foret pluviale dans la réserve de Lopé,
Gabon. Journal of Animal Ecology 63: 499 - 512.
White, L., Edwards A. (Eds). 2000. Conservation
en forêt pluviale africaine: Méthode de recherche. Wildlife
Conservation Society, New York. 456p.
Whitesides, G.H., Oates, J.F., Green, S.M., Kluberdanz,
R.P. 1988. Estimating primates densities from transect in a west
African rain forest: A campararison of technique. Journal of Animal
Ecology 57: 345-367.
Wiles, G.J. 1980. Feces deterioration rate of
four wild ungulates in Thailand. Natural History Bulletin of the Siam
Society 28: 121-134.
Wilson, V. 2005. Duikers of Africa.
Masters of the African forest Floor. Zimbi books. Pretoria. South Africa.
795p.
Zouya-Mimbang, L. 1998. Les circuits de
commercialisation des produits de chasse dans le Sud-Est Cameroun. Rapport,
GTZ-PROFORNAT, Yokadouma. 65p.
ANNEXES
Annexe 1: Fiche de
collecte des données d'inventaire de la faune
|
Date :
|
Météo :
|
Village :
|
Longueur totale transect (m):
|
Coordonnées début transect
Long. (m) :
Lat. (m) :
|
Coordonnées milieu transect
Long. (m) :
Lat. (m) :
|
Coordonnées fin transect
Long. (m) :
Lat. (m) :
|
|
Heure début :
|
Heure fin :
|
|
Transect N°:
|
Orientation transect :
|
|
Noms des collecteurs
|
|
Espèce/ indicateur
|
Nom Baka
|
Type de rencontre(2)
|
Nombre d'observation
|
Age indices (3)
|
Heure
|
Micro-habitat(1)
|
Distance sur le transect(m)
|
Distance perpendiculaire(m)
|
Observations
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
NB: (1): Colline, rocher, cours d'eau,
forêt galerie, marécage, peuplement mono-spécifique,
ouvert, fermé...
(2): Nids sur arbre, nids au sol, traces, crottes,
individus, excavations, terrier, cartouche, pièges, chasseurs, champs,
...
(3):Nombre de jours depuis le dépôt de
l'indice (crottes, nids, os)/décomposition
Annexe 2: Fiche
d'identification des crottes des animaux le long des transects
|
Village :
|
Espèce :
|
Noms des collecteurs :
|
|
|
Date
|
Code crotte
|
L'heure identification
|
No transect
|
Orientation
transect
|
Age de crotte
|
Distance perpendiculaire (m)
|
Distance le long du transect (m)
|
Coordonnées GPS :
|
Observations
|
|
long(m)
|
Lat. (m)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Annexe 3: Fiche
d'inventaire des caméras infra-rouges le long des transects
|
Village :
|
Noms des collecteurs :
|
|
|
Date début
|
Heure début
|
no caméra
|
no T
|
météo
|
Micro habitat (1)
|
Pente (2)
|
Hauteur fixation au-dessus du sol
|
Coordonnées GPS :
|
Date fin
|
Heure fin
|
Observation
|
|
long(m)
|
Lat. (m)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
NB : (1) : Colline, rocher, cours d'eau, forêt
galerie, marécage, peuplement mono-spécifique, ouvert,
fermé
(2) : légère, raide...
Annexe 4: fiche de suivi
des différents stades de décomposition des crottes des
céphalophes
|
Village :
|
Code de crotte :
|
Date de détection :
|
|
Transect no :
|
Noms collecteurs :
|
|
Nom de l'espèce :
|
|
Date de visite
|
Heure de visite
|
Description de stades de décomposition des crottes
|
Remarques
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Annexe 5: Fiche de
caractérisation de la végétation de la faune dans la zone
d'étude
|
Site :
|
Date :
|
Noms de collecteurs :
|
|
no T
|
no placette
|
Coordonnées
|
Nom commercial
|
Nom en Baka
|
DHP (cm)
|
Haut (m)
|
Eff
(1)
|
Dr
(2)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(1) : Eff = effectif des espèces identifiées
(2) : Dr = Degré de recouvrement
Annexe 6: Liste des espèces de grands et moyens
mammifères recensées dans la zone
|
Nom Français
|
Nom anglais
|
Nom scientifique
|
Faux nom
|
|
Primates
|
|
|
|
|
|
Chimpanzé
|
West Afican Chimpanzee
|
Pan troglodytes
|
|
|
Gorille
|
Gorilla
|
Gorilla g, gorilla
|
|
|
Magistrat
|
Black and white Colobus
|
Colobus guereza
|
|
|
Cercocèbe agile
|
Agile Mangabey
|
Cercocebus agilis
|
Agile
|
|
Cercopithèque pogonias
|
Crowned Monkey
|
Cercopithecus pogonias
|
|
|
Hocheur
|
White-nosed Monkey
|
Cercopithecus nictitans
|
Nez blanc
|
|
Moustac
|
Moustached Monkey
|
Cercopithecus cephus
|
Queue rouge
|
|
Mangabé à joues grises
|
Grey-checked Mangabe
|
Cercocebus albigena
|
Mécanicien
|
|
Rongeurs
|
|
|
|
|
|
Athérure africain
|
Brush-tailed Porcupine
|
Atherurus africanus
|
Porc-épic
|
|
Pholidotes
|
|
|
|
|
|
Pangolin à longue queue
|
Long-tailed Pangolin
|
|
Petit Pangolin
|
|
Pangolin géant
|
Giant Pangolin
|
Smutsia gigantea
|
Grand Pangolin
|
|
Proboscidea
|
|
|
|
|
|
Eléphant de forêt
|
Forest elephant
|
Loxodonta africana cyclotis
|
|
|
Artiodactiles
|
|
|
|
|
|
Potamochère
|
Red river Hog
|
Potamochoerus porcus
|
Sanglier
|
|
Oryctéropidea
|
|
|
|
|
|
Oryctérope
|
Aardvark
|
Orycteropus afer
|
|
|
Tragulidae
|
|
|
|
|
|
Chevrotain aquatique
|
Water Chevrotain
|
Hyemoschus aquaticus
|
|
|
Bovidae
|
|
|
|
|
|
Buffle
|
Buffalo
|
Syncerus cafer nanus
|
|
|
Bongo
|
Bongo
|
Tragelaphus euryceros
|
Zèbre
|
|
Sitatunga
|
Sitatunga
|
Tragelaphus spekei
|
Gazelle
|
|
Cephalophe bleu
|
Blue Duiker
|
Cephalophus monticola
|
Lièvre
|
|
Cephalophe de Peter's
|
Peter's Duiker
|
Cephalophus callipygus
|
|
|
Cephalophe à dos jaune
|
Yellow-backed Duiker
|
Cephalophus silvicultor
|
|
|
Cephalophe bai
|
Bay Duiker
|
Cephalophus dorsalis
|
Biche
|
|
Antilope de Bate's
|
Dwarf Antelope
|
Neotragus batesi
|
|
|
Carnivore
|
|
|
|
|
|
Civette
|
African civet
|
Civettictis civetta
|
|
|
Léopard
|
Leopard
|
Panthera pardus
|
|
|
Chat sauvage
|
Wild cat
|
Nandinia binotata
|
|
|
Hyracoides
|
|
|
|
|
|
Daman des arbres
|
Tree Dassie
|
Dendrohyrax arboreus
|
|
|
|
|
|
|
|
Potto de Bosman
|
|
Perodicticus potto
|
Potto
|
Annexe 7: Quelques
espèces rares photographiées par les Caméras infra-rouges
dans la zone d'étude







