La mécanique statistique des membranes biologiques confinées( Télécharger le fichier original )par Khalid EL HASNAOUI Faculté des sciences Ben M'Sik Casablanca - Thèse de doctorat 2011 |

D/2f dzz2 (z) . (5.0) -D/2 D/2 f -D/2 dz (z) The restricted partition function is ? { j -H [h] (z) = Dhä [z - h (x0, y0)] exp . (5.0) kBT Here, H [h] is the original Hamiltonian defined in Eq. (3). Of course, this definition is independent on the chosen point (x0, y0), because of the translation symmetry along the parallel directions to plates. Notice that the above function is not singular, whatever the value of the perpendicular distance. Since we are interested in the confinement-regime, that is when the separation D is much smaller than (z h << L0 the membrane mean-roughness L0 ), we can replace the function par its value at z = 0, 1 1 denoted 0. In this limit, Eq. (A.2) gives the desired result. This ends the proof of the expected formula. ACKNOWLEDGMENTS We are much indebted to Professors T. Bickel, J.-F. Joanny and C. Marques for helpful discussions, during the »First International Workshop On Soft-Condensed Matter Physics and Biological Systems», 14-17 November 2006, Marrakech, Morocco. One of us (M.B.) would like to thank the Professor C. Misbah for fruitful correspondences, and the Laboratoire de Spectroscopie Physique (Joseph Fourier University of Grenoble) for their kinds of hospitalities during his regular visits. K. El Hasnaoui et al. African Journal Of Mathematical Physics Volume 8(2010)101-114 114 REFERENCES 1 M.S. Bretscher and S. Munro, Science 261, 12801281 (1993). 2 J. Dai and M.P. Sheetz, Meth. Cell Biol. 55, 157171 (1998). 3 M. Edidin, Curr. Opin. Struc. Biol. 7, 528532 (1997). 4 C.R. Hackenbrock, Trends Biochem. Sci. 6, 151154 (1981). 5 C. Tanford, The Hydrophobic Effect, 2d ed., Wiley, 1980. 6 D.E. Vance and J. Vance, eds., Biochemistry of Lipids, Lipoproteins, and Membranes, Elsevier, 1996. 7 F. Zhang, G.M. Lee, and K. Jacobson, BioEssays 15, 579588 (1993). 8 S. Safran, Statistical Thermodynamics of Surfaces, Interfaces and Membranes, Addison-Wesley, Reading, MA, 1994. 9 W. Helfrich, Z. Naturforsch. 28c, 693 (1973). 10 For a recent review, see U. Seifert, Advances in Physics 46, 13 (1997). 11 H. Ringsdorf and B. Schmidt, How to Bridge the Gap Between Membrane, Biology and Polymers Science, P.M. Bungay et al., eds, Synthetic Membranes: Science, Engineering and Applications, p. 701, D. Reiidel Pulishing Compagny, 1986. 12 D.D. Lasic, American Scientist 80, 250 (1992). 13 V.P. Torchilin, Effect of Polymers Attached to the Lipid Head Groups on Properties of Liposomes, D.D. Lasic and Y. Barenholz, eds, Handbook of Nonmedical Applications of Liposomes, Volume 1, p. 263, RCC Press, Boca Raton, 1996. 14 R. Joannic, L. Auvray, and D.D. Lasic, Phys. Rev. Lett. 78, 3402 (1997). 15 P.-G. de Gennes, Scaling Concept in Polymer Physics, Cornell University Press, 1979. 16 H.B.G. Casimir, Proc. Kon. Ned. Akad. Wetenschap B 51, 793 (1948). 17 S.K. Lamoreaux, Phys. Rev. Lett. 78, 5 (1997). 18 U. Mohideen and A. Roy, Phys. Rev. Lett. 81, 4549 (1998). 19 M.E. Fisher and P.-G. de Gennes, C. R. Acad. Sci. (Paris) Sér. B 287, 207 (1978); P.-G. de Gennes, C. R. Acad. Sci. (Paris) II 292, 701 (1981). 20 M. Krech, The Casimir Effect in Critical Systems, World Scientific, Singapore, 1994. 21 More recent references can be found in : F. Schlesener, A. Hanke, and S. Dietrich, J. Stat. Phys. 110, 981 (2003); M. Benhamou, M. El Yaznasni, H. Ridouane, and E.-K. Hachem, Braz. J. Phys. 36, 1 (2006). 22 R. Lipowsky, Handbook of Biological Physics, R. Lipowsky and E. Sackmann, eds, Volume 1, p. 521, Elsevier, 1995. 23 P.B. Canham, J. Theoret. Biol. 26, 61 (1970). 24 O. Farago, Phys. Rev. E 78, 051919 (2008). 25 We recover the power law II - D-3 that is known in literature (see, for instance, Ref. [8]), but the corresponding amplitude depends on the used model. 26 G. Gompper and D.M. Kroll, Europhys. Lett. 9, 59 (1989). 27 U. Seifert and R. Lipowsky, in Structure and Dynamics of Membranes, Handbook of Biological Physics, R. Lipowsky and E. Sackmann, eds, Elsevier, North-Holland, 1995. 28 J. Zinn-Justin, Quantum Field Theory and Critical Phenomena, Clarendon Press, Oxford, 1989. 29 H.K. Jansen, Z. Phys. B 23, 377 (1976). 30 R. Bausch, H.K. Jansen, and H. Wagner, Z. Phys. B 24, 113 (1976). 31 F. Langouche, D. Roekaerts, and E. Tirapegui, Physica A 95, 252 (1979). 32 R. Lipowsky, in Random Fluctuations and Growth, H.E. Stanley and N. Ostrowsky, eds, p. 227-245, Kluwer Academic Publishers, Dordrecht 1988. 33 R. Lipowsky, J. Phys. A 18, L-585 (1985). 34 R. Lipowsky, Physica Scripta T 29, 259 (1989). 35 F. Brochard and J.F. Lennon, J. Phys. (Paris) 36, 1035 (1975).
Article N°3 Mécanique Statistique des membranes confinées dans un liquide trouble
Dans cette troisième contribution originale, nous étudions les effets d'impuretés sur les propriétés statistiques des membranes fluides. Celles peuvent être attractives ou répulsives. En premier lieu, nous déterminons la rugosité moyenne de la membrane, en combinant la technique des répliques avec la méthode variationnelle. Le résultat s'exprime en fonction de la concentration des impuretés et l'amplitude de leur interaction avec la membrane. En second lieu, nous déterminons la taille d'une vésicule isolée, en fonction de ces mêmes paramètres. Enfin, l'étude est étendue à l'adhésion membranaire.
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their
version of the http://www.elsevier.com/copyright Author's personal copy Physica A 389 (2010) 3465-3475
Statistical mechanics of bilayer membranes in troubled aqueous media M. Benhamou*, K. Elhasnaoui, H. Kaidi, M. Chahid Laboratoire de Physique des Polymères et Phénomènes Critiques, Faculté des Sciences Ben M'sik, P.O. 7955, Casablanca, Morocco
Article history: Received 30 March 2010 Available online 14 April 2010 Keywords: Bilayer membranes Vesicles Impurities Equilibrium Statistical mechanics 1. Introduction We consider a bilayer membrane surrounded by small impurities, assumed to be attractive or repulsive. The purpose is a quantitative study of the effects of these impurities on the statistical properties of the supported membrane. Using the replica trick combined with a variational method, we compute the membrane mean-roughness and the height correlation function for almost-flat membranes, as functions of the primitive elastic constants of the membrane and some parameter that is proportional to the volume fraction of impurities and their interaction strength. As results, the attractive impurities increase the shape fluctuations due to the membrane undulations, while repulsive ones suppress these fluctuations. Second, we compute the equilibrium diameter of (spherical) vesicles surrounded by small random particles starting from the curvature equation. Third, the study is extended to a lamellar phase composed of two parallel fluid membranes, which are separated by a finite distance. This lamellar phase undergoes an unbinding transition. We demonstrate that the attractive impurities increase the unbinding critical temperature, while repulsive ones decrease this temperature. Finally, we say that the presence of small impurities in an aqueous medium may be a mechanism to suppress or to produce an unbinding transition, even the temperature and polarizability of the aqueous medium are fixed, in lamellar phases formed by parallel lipid bilayers. (c) 2010 Elsevier B.V. All rights reserved. Usually, the aqueous media supporting biological membranes are assumed to be homogeneous. Actually, any real system inevitably contains impurities. Under well-controlled conditions, the particles can be removed from the surrounding medium. But, if these entities are present, it is also interesting to study their effect on the statistical properties of the biomembranes, such as fluctuations' spectrum and dynamical behavior. In general, random inhomogeneities tend to disorder the system. It is important to make a distinction between annealed and quenched disorders. The former is used when impurities and host constituents (phospholipids) are in equilibrium [1]. This means that their respective mobilities are comparable. If it is not the case, that is host constituents and impurities are out of equilibrium, the disorder is rather quenched [1]. When the Statistical Mechanics is used, the latter consists to trace over all membrane undulations, before performing the summation over the impurities' disorder. Although the quenched disorder is harder to analyze, it remains more realistic than its annealed counterpart. Indeed, the thermal and the noise averaging have very different roles. In this paper, the physical system we consider is a fluid membrane (flat or closed) trapped in a troubled aqueous medium. The latter is impregnated by a weak amount of impurities that may be attractive or repulsive regarding the membrane. The aim is to show how these entities can modify the statistical properties of the fluid membrane. These properties will be studied through the fluctuations' amplitude. To model the system, we suppose that the impurities act as a random external potential with a Gaussian distribution (uncorrelated disorder). In addition, we suppose that the disorder is quenched. To * Corresponding author. E-mail address: benhamou.mabrouk@gmail.com (M. Benhamou). 0378-4371/$ - see front matter (c) 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.physa.2010.03.049 Author's personal copy 3466 M. Benhamou et al. / Physica A 389 (2010) 3465-3475 do calculations, the replica theory [2,3] that is based on a mathematical analytical continuation, usually encountered in Quantum Mechanics [4] and Critical Phenomena will be made use of [5,6]. Our finding are as follows. First, using the above evoked theory, we compute the mean-roughness of an almost-flat membrane, as a function of the primitive parameters of the pure membrane (free from impurities) and a certain parameter depending on the volume fraction of impurities and their interaction strength. The main conclusion is that, attractive impurities increase the shape fluctuations due to the thermal undulations, while repulsive ones tend to suppress these fluctuations and then lead to a strong membrane confinement. Second, we analyze the impurities effects on the equilibrium shape of closed vesicles solving the curvature equation (for spherical vesicles). We show that the vesicle is more stable in the presence of repulsive impurities, in comparison with attractive ones. Thereafter, the study is extended to lamellar phases formed by two parallel fluid membranes. We demonstrate that the presence of random impurities drastically affects the physical properties of the lamellar phase, in particular, the unbinding transition driving the system from a bind state to a state where the two membranes are completely separated. This paper is organized as follows. In Section 2, we describe the fundamentals of the used model. Section 3 deals with the computation of the fluctuations amplitude of a single almost-flat fluid membrane, surrounded by attractive or repulsive impurities. We compute, in Section 4, the equilibrium diameter of a closed vesicle in the presence of impurities. Extension of study to lamellar phases is the aim of Section 5. Finally, some concluding remarks are drawn in the last section. 2. Effective field theory Consider a fluctuating fluid membrane embedded in a three-dimensional liquid surrounded by very small impurities. For the sake of simplicity, we suppose that the impurities are point like. Within the framework of the Monge representation, a point on the membrane can be described by the position-vector (r, z = h (r)), where r = (x, y) ? R2 is the transverse vector and h (r) is the height function. The Statistical Mechanics of fluid membranes free from impurities is based on the Canham-Helfrich Hamiltonian [7] f [ê 2 h2] R0 [h] = d2r 2 (Äh)2 + u , (1) with ê the membrane bending modulus. The confinement energy (per unit area) uh2/2 (u > 0) is responsible for the localization of the membrane in some region of the Euclidean space, where it fluctuates around an equilibrium plane located at h = 0. Therefore, the height h takes either positive and negative values. For simplicity, the membrane is assumed to be tensionless. In fact, this assumption does not change conclusions made below. To model the impurities effects on the statistical properties of the system, we suppose that these tend to reinforce the membrane confinement, if they are repulsive, or to render this membrane more free, if the particles are rather attractive. These tendencies can be explained assuming that the confinement parameter is local in space and making the substitution u ? u + V (r) , (2) in the above Hamiltonian, where the variation V (r) can be regarded as a random external potential. To simplify, the corresponding probability distribution is supposed to be Gaussian (uncorrelated disorder), that is V (r) = 0, V (r) V (r0) = -vä2 (r - r0) . (3) Here, -v is a positive constant proportional to both concentration of impurities and strength of their interaction potential, and ä2 (r) denotes the two-dimensional Dirac distribution. Therefore, the new effective Canham-Helfrich Hamiltonian reads f Lê ] R [h] = d2r 2 (Äh)2 + 2 1 (u + V (r)) h2 , (4) for attractive impurities. For repulsive ones, V (r) must be replaced by iV (r), with i2 = -1. Since the disorder distribution is Gaussian, all its odd moments vanish, but the even ones do not. This implies that all physical quantities, calculated with the pure imaginary potential iV, are entire numbers. Since the impurities and membranes are not in equilibrium, the disorder is rather quenched, that is we have to average not the partition function, Z, but its logarithm, ln Z. The latter defines the free energy. For a given (quenched) configuration of impurities, the partition function is fZ = Dhe-A[h], (5) with the action
Author's personal copy M. Benhamou et al. / Physica A 389 (2010) 3465-3475 3467 We have used the notations ê u bê = bV = bu = kBT , kBT , V . (7) kBT In term of the reduced impurities potential bV, the second disorder law in Eq. (3) becomes: bV (r)bV (r') = -bvä (r - r'), with bv = v/ (kBT)2. A simple dimensional analysis shows that: [bê] = L0, [bu] = L-4 and [bv] = L-6, where L is some length that may be the membrane thickness. The main quantity to consider is the average of the logarithm of the partition function, ln Z, over disorder. To compute such a quantity, we use the replica trick [2]. This method consists to formally write Zn - 1 . (8) n ln Z = lim n?0 After performing the average over disorder, we get ZZn = Dh1 ...
Dhne-A[h1, ,hn], (9) Z " ! nX !# Xn Xn bê A [h1, ... , hn] = d2r (Ähá)2 + bu á + bv Xn h2 h2 h2 , (10) á â 2 2 8 á=1 á=1 á=1 â=1 where Greek indices denote replicas. The last term introduces an additional coupling constant bv < 0, which is directly responsible for the effective interaction between replicas due to the presence of impurities. The above action describes attractive impurities. For repulsive ones, the coupling bv must be replaced by -bv. The additive quartic term in the above action means that the presence of impurities is accompanied by an increase of entropy, when these are attractive, and by an entropy loss, if they are rather repulsive. The following paragraph is devoted to a quantitative determination of the fluctuations spectrum in the presence of impurities. 3. Single almost-flat membrane We start by considering the attractive impurities problem. Of course, the functional integral (9) cannot be exactly computed. One way is the use of a variational method. To this end, we consider a bilayer membrane in the presence of the following bare action Z " # Xn (Ähá)2 + bç Xn bê Aç [h1, ... , hn] = d2r h2 , (11) á 2 2 á=1 á=1 with the new confinement couplingbç > 0. With this action, the partition function, Zç, is exact. We have Zç = (Z0)n , (12) with Z ~ Z ~bê ~~ Z0 = Dh exp - d2r
2 (Äh)2 + bç 2 h2 .
(13) Introduce the mean-value of a functional X [h1, ... , hn], calculated with the bare action Aç [h1, ... , hn], (X)0 = Z Dh1 ... DhnX [h1, ... , hn] exp {-A,~ [h1, ... , hn]} . Zç (14) In term of this mean-value, the averaged partition function may write ~~~ Zn = Zç ~exp ~-~A - Aç 0 . (15) Using the standard inequality (eX)0 > e(X)0, (16) we get Zn > Zç exp~-~A - Aç ~ ~. (17) 0 Author's personal copy 3468 M. Benhamou et al. / Physica A 389 (2010) 3465-3475 This implies that
On the other hand, we have Z " # A - Aç~ Xn ~ Xn Xn h2 bu -bç h2 0 = d2r 0 + bv ~ áh2 . (19) á â 2 8 0 á=1 á=1 â=1 It is easy to see that
with the squared membrane roughness
With these considerations, bu -bç 2 ó2 + bv ~ó2~2~
(The subscript MF is for mean-field theory). The variational parameterbç is such that ~ bu -bç ~ó2~2~~ ? 2 ó2 + bv - ln Z0 + Ó = 0. (23) ?bç 4 Combining the relationship
and Eq. (20) yields the minimum value ofbç, which satisfies the following implicit relation bç = bu +bvó2. (25) ? ? ? 1 ó2 = The squared membrane roughness, ó2, is then given by the parametric equations 1
p , bç = 8 bê bç bu +bvó2. Eliminatingbç between these equations gives the implicit relation q8 bê Cbu +bvó2). 1
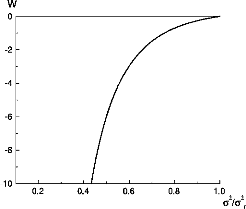
1 ó2 = p Introduce the usual squared roughness of a membrane free from impurities, ó2 0 = 1/8 bê bu. Then, we have 1 ó2 = ó2 . 0 (28) p1 +bvó2/bu 3 > 0, as The latter may be rewritten, in terms of dimensionless variables ó2/ó0 2 ,bê, and w = -64bê bv ~óÔ ó2 = ó2 0 1 q1 - wó2/óÔ , (attractive impurities) . (29) Author's personal copy M. Benhamou et al. / Physica A 389 (2010) 3465-3475 3469 Table 1 Some values of the coupling w and their corresponding ratios ó2/óô , for attractive impurities.
Notice that the coupling constant w > 0 is directly proportional to the volume fraction of impurities and their interaction strength. Let us comment about the obtained result. Firstly, this makes sense only when ó2/ó0 2 < 1/w. This condition is fulfilled for very weak disorders (w is small enough). Secondly, since w is positive definite, we have: ó2 > ó02. A comparison between this inequality and that just evoked above implies that 0 < w < 1. Thirdly, if we set x = ó2/ó02, the above relation can be rewritten as x2 - 1 w = . (30) x3 Therefore, w is a direct function of x (Fig. 1). This formula may be experimentally used to estimate the impurities coupling w, knowing the experimental value of the ratio ó2/ó20 . If w is fixed to some value, the x-variable then solves the following third-degree algebraic equation wx3 - x2 + 1 = 0. (31) The existence of its roots then depends on the value of the renormalized coupling w. We show that there exists a typical v value w* = 2/3 3 of the coupling w, such that:
- 4wx6 + 3x4 - 1 > 0. (32) In this case, the ratio ó2/ó02 can be obtained by numerically solving Eq. (31). We report in Table 1 some values of the impurities coupling w and the corresponding dimensionless squared membrane roughness ó2/ó02. It is easy to see that, the ratio ó2/ó02 increases with increasing coupling w, provided that w is in the interval w < w*. ó2 = ó2 0 1 , (repulsive impurities) , (33) Now, for repulsive impurities, the fundamental relationship (29) is replaced by q1- wó2/ó02 or equivalently - wx3 - x2 + 1 = 0,
(34) 4wx6 + 3x4 - 1 > 0.
(35) x2 - 1 w =3 (36) x with x < 1 and w < 0. As it should be, the membrane roughness is reduced by the presence of repulsive impurities, that is ó2 < óô, and in addition, ó2 decreases with increasing impurities coupling -w. As a matter of fact, the effect of these particles is to reinforce Author's personal copy
3470 M. Benhamou et al. / Physica A 389 (2010) 3465-3475
Fig. 1. Impurities' coupling w versus the ratio ó2/ó0 2 , for attractive impurities. Fig. 2. Impurities' coupling w versus the ratio ó2/ó2 0 , for repulsive impurities. Table 2 Some values of the coupling w and their corresponding ratios ó2/ó0 2 , for attractive impurities.
the confinement of the considered fluid membrane. Therefore, a bilayer membrane collapses, when it is trapped in a troubled aqueous media with repulsive impurities. In this case, it is easy to see that the algebraic equation has only one positive root. In Table 2, we give some values of the coupling w and the corresponding ratio ó2/ó2 0 . Another interesting physical quantity is the (connected) height correlation function: G (r - r') = (h (r) h (r')) - (h (r)) (h (r')). The latter measures the fluctuations of the height function h around its mean-value (h). Here, (.) denotes the thermal expectation mean-value, which must not be confused with average over disorder. Author's personal copy M. Benhamou et al. / Physica A 389 (2010) 3465-3475 3471 To compute the correlation functions, we start from the generating functional Z ~ Z ~ Z [J] = Loh exp -A [h] + d2r J (r) h (r) , (37) with the action A [h] = X' [h] /kBT, where X' [h] is the effective Canham-Helfrich Hamiltonian defined in Eq. (4). Here, J is an auxiliary source coupled to the field h. Functional derivatives of Z [J] with respect to source J give all height correlation functions, in particular, the propagator ~~ ä2ln Z [J] (38) ) J=0. G~r - r0) = ä ä 0 J (r) J (r This definition indicates that the main object to compute is the averaged connected generating functional ln Z [J]. To compute the functional ln Z [J], as before, we use the replica method based on the limit
Zn [J] = Z Loh1 ... Lohn exp -A [h1, ... , hn] + XZd2r J (r) hi (r) , (40) i=1 where A [h1, ... , hn] is the action, relation (10). Using the standard cumulant method [5,6] based on the approximate formula hexp {X}i0 = exp {hXi0 + (1/2!) ((X2)0 - hXi2 ~ + · · ·~ , (41) 0 we find that Z Zn [J] ' Zç exp~-~A - Aç ~ 0 + ···~exp~n Z d2r0J (r)G0~r - r0~J ~r0~~ d2r , (42) 2 with the usual bare propagator eiq.(r-r0) G0(r - r0)= Z d2q (43) (2ð)2 bêq4 +bç . In Eq. (42), we have ignored high-order terms in J that do not contribute to the propagator. After performing the limit n = 0, we find that the averaged connected generating functional reads bu -bç Z Z 2 ó2 + bv ~ó2~2~ + 1 ~r - r0 J ~r0~ . ln Z [J] ' - ln Z0 + Ó d2r d2r0J (r) G0 (44) 4 2 By simple functional derivation, we obtain the expected propagator eiq.(r-r0) d2q G{r - r0) = G0{r - r0) =Z (45) (2ð)2bêq4 +bç . Then, the expected propagator identifies with the bare one. Here, the variational parameterbç satisfies the implicit equation (25). We then recover the relationship: ó2 = G
(0), where ó2 is the squared membrane roughness computed
above. 4. Single vesicle We start by recalling some basic backgrounds dealt with the equilibrium shape of (spherical) vesicles, which may studied using Differential Geometry techniques. The vesicle is essentially formed by two adjacent monolayers (inner and outer) that are formed by amphiphile lipid molecules. These permanently diffuse with the molecules of the surrounded aqueous medium. Such a diffusion then provokes thermal fluctuations (undulations) of the membrane. This means that the latter experiences fluctuations around an equilibrium plane. Consider a biomembrane of arbitrary topology. A point of this membrane (surface) can be described by two local coordinates (u1, u2). At each point of the surface, there exists two particular curvatures (minimal and maximal), called Author's personal copy 3472 M. Benhamou et al. / Physica A 389 (2010) 3465-3475 principal curvatures, denoted C1 = 1/R1 and C2 = 1/R2. The quantities R1 and R2 are the principal curvature radii. With the help of the principal curvatures, one constructs two invariants that are the mean-curvature 1 C = 2 (C1 + C2) , (46) and the Gauss curvature R R0 = 1 #177; 8ky2 ö + O (ö2) , (56) K = C1C2. (47) The principal curvatures C1 and C2 are nothing else but the eigenvalues of the curvature tensor [8]. To comprehend the geometrical and physical properties of the biomembranes, one needs a good model. The widely accepted one is the fluid mosaic model proposed by Singer and Nicolson in 1972 [9]. This model consists to regard the cell membrane as a lipid bilayer, where the lipid molecules can move freely in the membrane surface like a fluid, while the proteins and other amphiphile molecules (cholesterol, sugar molecules, ...) are simply embedded in the lipid bilayer. We note that the elasticity of cell membranes crucially depends on the bilayers in this model. The elastic properties of bilayer biomembranes were first studied, in 1973, by Helfrich [7]. The author recognized that the lipid bilayer could be regarded as smectic-A liquid crystals at room temperature, and proposed the following curvature free energy (without impurities) F = 2 f (2C)2 dA + êG f KdA + f ãdA + p f dV. (48) where dA denotes the area element, and V is the volume enclosed within the lipid bilayer. In the above definition, ê accounts for the bending rigidity constant, êG for the Gaussian curvature, ã for the surface tension, and p for the pressure difference between the outer and inner sides of the vesicle. The spontaneous curvature is ignored. The first-order variation gives the shape equation of lipid vesicles [10] p - 2ãC + 4êC (C2 - K) + ê?2 (2C) = 0, (49) with the surface Laplace-Bertolami operator
where gij is the metric tensor on the surface and g = det (gij). For open or tension-line vesicles, local differential equation (49) must be supplemented by additional boundary conditions we do not write [11]. The above equation have known three v analytic solutions corresponding to sphere [10], 2-torus [12-15] and biconcave disk [16]. For spherical vesicles, the solution to the above curvature equation is exact, and we find that the equilibrium radius is 2ã R0 = . (51) Now, assume that the vesicle is trapped in a troubled aqueous medium. Usually, to take into account the presence of impurities, a low-order coupling between their volume fraction, ö, and the mean-curvature C is added to the above free energy F, that is fF ? F #177; kBT öCdA. (52) The positive sign is for attractive impurities and the negative sign for repulsive ones. The new free energy is then F = 2 f(2C #177; 2C0)2 dA + êGfKdA +fãIdA + pfdV (53)
Then, the impurities generate an extra spontaneous curvature. This means that these give arise to an asymmetry of the vesicle when its membrane is crossed. In addition, these impurities additively renormalize the interfacial tension coefficient. Minimizing this new curvature free energy yields p - 2ÿC + ê (2C #177; C0) (2C2 F C0C - 2K) + ê?2 (2C) = 0. (55) For a spherical vesicle, the mean curvature is a constant, and we have K = C2. In this case, the mean-curvature C is a root of a polynomial of degree 2. This means that we have an exact solution we do not write. In particular, for very small volume fractions (ö « 1), we find that the vesicle equilibrium radius writes Author's personal copy M. Benhamou et al. / Physica A 389 (2010) 3465-3475 3473 or ~~ ~~ R - R0 ~ ~ ~ R0 pkBT ^-' 8ã 2 ö. (57) The signs (+) and (-) describe attractive and repulsive impurities, respectively. Here, R0 = 2ã/p denotes the equilibrium radius for a spherical vesicle free from impurities. The above results call the following remarks. Firstly, as it should be, the equilibrium radius of the vesicle increases with increasing volume fraction of impurities, when these are attractive. This means that the vesicle is swollen, but it radius remains close to the unperturbed one R0, as long as the volume fraction of impurities is very small. For repulsive impurities, however, the vesicle is collapsed. Secondly, formula (57) makes sense only when the volume fraction ö is below some threshold ö* = 8ã2/kBTp. Since the latter must be small in comparison with unity, the vesicle bears interfacial tensions of coefficients that do not exceed some typical value ã* = .ABTp/8. Finally, at fixed volume fraction ö < ö*, this formula may be used to estimate the experimental value of the ratio p/ã2, by a simple measurement of the equilibrium radii R (with impurities) and R0 (without impurities). The following paragraph will be devoted to the study of lamellar phases made of two parallel lipid bilayers. 5. Lamellar phases The natural question is the extension of the study to the case when we have more than one membrane immersed in a aqueous media, which is impregnated by small random impurities. In our analysis, we start from a lamellar phase composed of two parallel (neutral) fluid membranes free from impurities. The effects of these entities on physics will be discussed below. The cohesion between these bilayer membranes is ensured by long-ranged attractive van der Waals forces [17], which are balanced, at short membrane separation, by strong repulsion coming from hydration forces [18] and by steric shape fluctuations ones resulting from the membrane undulations [7]. For two parallel bilayer membranes separated by a finite distance l, the total interaction energy per unit area is V (l) = VH (l) + VW
(l) + VS (l) . (58) VH (l) =
AHe-l/ëH
(59) ~ 1 ~ W VW (l) = - (60) l2 - 2 (l + ä)2 + 1 12ð (l + 2ä)2 is the attractive van der Waals potential (per unit area) that originates from polarizabilities of lipid molecules and water molecules. Here, W accounts for the Hamaker constant that is in the range W ^-' 10-22 - 10-21 J, and ä for the bilayer thickness. The latter is of the order of ä ^-' 4 mm. The last part is the steric shape fluctuations potential (per unit area) [7] Vs (l) = cH (kBT)2 , (61) We note that the lamellar phase remains stable at the minimum of the potential, provided that the potential depth is comparable to the thermal energy kBT. This depends, in particular, on the value of amplitude W of the direct van der Waals energy. The Hamaker constant W may be varied changing the polarizability of the aqueous medium. In a pioneered theoretical paper, Lipowsky and Leibler [19] have shown that there exists a certain threshold Wc beyond which the van der Waals attractive interactions are sufficient to bind the membranes together, while below this characteristic amplitude, the membrane undulations dominate the attractive forces, and then, the membranes separate completely. According to the authors, Wc is in the interval Wc ^-' (6.3 - 0.61) x 10-21 J when the bending rigidity constant is in the range ê ^-' (1 - 20) x 10-19 J. We note that the typical value Wc corresponds to some temperature, Tc, called unbinding critical temperature [19,20]. In particular, it was found [19] that, when the critical amplitude is approached from above, the mean-separation between the two membranes, (l)0, diverges according to (l)0=î0?~
(Tc -T)-ø,
T?Tc-, (62) Author's personal copy 3474 M. Benhamou et al. / Physica A 389 (2010) 3465-3475 Now, assume that the lamellar phase is trapped in a troubled aqueous medium. As first implication, the impurities drastically modify the unbinding transition phenomenon, in particular, the critical temperature. For simplicity, suppose that the two adjacent fluid membranes are physically identical. Then, we have to consider two distinct physical situations: The impurities attract or push the two membranes. As we have shown above, for attractive impurities, the membrane roughness î? is important, and then, the steric shape fluctuation energy dominates. Therefore, we expect that the unbinding transition occurs at a low critical temperature Tc* we will determine below. For repulsive impurities, however, we have an opposite tendency, that is the unbinding transition takes place at a high temperature greater than Tc (absence of impurities). This can be explained by the fact that the membrane roughness is less important, and then, the direct van der Waals energy dominates. In any case, the mean-separation hli scales as hli = î? -- (Tc* - T)-ø , T ? Tc- . (63) The new unbinding critical temperature Tc* can be estimated as follows. Using formulae (29) and (33), we show that, at first order in the impurities coupling w,
Combing Eqs. (62) to (65), we find that the difference between the two critical temperatures Tc and Tc* (without and with impurities) is as follows Tc - Tc* -- w + (9 (w2) , (attractive impurities) , (66) Tc - Tc* -- -w + (9 (w2) , (repulsive impurities) . (67) The prefactors in the above behaviors remain unknown. The temperature shift is then proportional to the volume fraction of impurities and their interaction strength through the coupling w. Then, we are in a situation similar to finite size scaling. For charged membranes forming the lamellar phase, it was found [19] that the mean-separation between two adjacent bilayers scales as [19] hli0 = î0? -- (÷ - ÷c)-ø , (68) in the vicinity of ÷c, with ÷ the ionic concentration of the aqueous medium and ÷c its critical value. For instance, for DPPC in CaCl2 solutions, ÷c is in the interval [21]: ÷c ' 84-10 mM. In the presence of impurities, we state that the mean-separation behaves as hli = î? -- (÷ - ÷* )-ø , (69) c with the same exponent ø. In this case, we have ÷*c - ÷c -- w + (9 (w2) , (attractive impurities) , (70) ÷*c - ÷c -- -w + (9 (w2) , (repulsive impurities) . (71) We note that the same comments may by done in this case. 6. Conclusions In this paper, we presented a large scope about the effects of impurities on the statistical properties of fluid membranes. In fact, these drastically affect the living systems behavior. As an example, we can quote a very recent experimental study [22], where the authors have undertaken a series of comparative experiments, in order to explore the effect of impurities in the form of proteins and lipids on the crystallization of membrane proteins in vapor diffusion. For the present study, the impurities were assumed to be attractive or repulsive. Using the replica trick combined with a variational method, we have computed the membrane mean-roughness, as a function of the parameters associated with the pure membrane and some parameter that is proportional to the volume fraction of impurities and their interaction strength. The main conclusion is that, attractive impurities increase the shape fluctuations due to the membrane undulations, but repulsive ones tend to suppress these fluctuations. Also, we have computed the equilibrium diameter of (spherical) vesicles surrounded by small random particles solving the curvature equation. Thereafter, we extended discussion to lamellar phase formed by two parallel fluid membranes that are a finite distance apart. This lamellar phase may undergo an unbinding transition. We have shown that, attractive impurities increase the unbinding critical temperature, while repulsive ones tend to decrease this temperature. Author's personal copy M. Benhamou et al. / Physica A 389 (2010) 3465-3475 3475 We point out that, the incorporation of a small amount of impurities in an aqueous medium may be a mechanism to suppress or to produce an unbinding transition (even the temperature and polarizability of the aqueous medium are fixed) within lamellar phases composed of fluid membranes. Finally, the present study may be extended to bilayer membranes of arbitrary topology. The essential conclusion is that, attractive impurities tend to swell the membrane, while in the presence of repulsive ones, this membrane is collapsed. Also, the used method can be applied to multilayers constituted by several parallel lamellar phases. Acknowledgements We are much indebted to Professors T. Bickel, J.-F. Joanny and C. Marques for helpful discussions, during the «First International Workshop On Soft-Condensed Matter Physics and Biological Systems», 14-17 November 2006, Marrakech, Morocco. One of us (M.B.) would like to thank Professor C. Misbah for fruitful correspondences, and the Laboratoire de Spectroscopie Physique (Joseph Fourier University of Grenoble) for their kind hospitality during his regular visits. References 111 J. Cardy, Scaling and Renormalization in Statistical Physics, Cambridge University Press, 1996. 121 V.J. Emery, Phys. Rev. B 11 (1975) 239. 131 S.F. Edvards, P.W. Anderson, J. Phys. (France) 5 (1975) 965. 141 L.D. Landau, E.M. Lifshitz, Quantum Mechanics, 3rd edition, Pergamin Press, 1991. 151 J. Zinn-Justin, Quantum Field Theory and Critical Phenomena, Clarendon Press, Oxford, 1989. 161 C. Itzykson, J.-M. Drouffe, Statistical Field Theory: 1 and 2, Cambridge University Press, 1989. 171 W. Helfrich, Z. Natureforsch 28c (1973) 693. 181 K. Wolfgang, Differential Geometry: Curves - Surfaces - Manifolds, American Mathematical Society, 2005. 191 S.J. Singer, G.L. Nicolson, Science 175 (1972) 720. 1101 O.-Y. Zhong-Can, W. Helfrich, Phys. Rev. Lett. 59 (1987) 2486. 1111 Z.C. Tu, Z.C. Ou-Yang, Phys. Rev. E 68 (2003) 061915. 1121 O.-Y. Zhong-Can, Phys. Rev. A 41 (1990) 4517. 1131 Z. Lin, R.M. Hill, H.T. Davis, L.E. Scriven, Y. Talmon, Langmuir 10 (1994) 1008. 1141 M. Mutz, D. Bensimon, Phys. Rev. A 43 (1991) 4525. 1151 A.S. Rudolph, B.R. Ratna, B. Kahn, Nature 352 (1991) 52. 1161 H. Naito, M. Okuda, O.Y. Zhong-Can, Phys. Rev. E 48 (1993) 2304. 1171 J.N. Israelachvili, Intermolecular and Surface Forces, 2nd edition, Academic Press, London, 1991. 1181 R.P. Rand, V.A. Parsegian, Biochim. Biophys. Acta 988 (1989) 351. 1191 R. Lipowsky, S. Leibler, Phys. Rev. Lett. 56 (1986) 2541. 1201 R. Lipowsky, E. Sackmann (Eds.), An Extensive List of References on the Subject can be Found in: Structure and Dynamics of Membranes: Generic and Specific Interactions, vol. 1B, Elsevier, 1995. 1211 L.J. Lis, W.T. Lis, V.A. Parsegian, R.P. Rand, Biochemistry 20 (1981) 1771. 1221 A. Christopher, et al., Acta Cryst. D 65 (2009) 1062-1073. Article N°4 Conformation d'un polymère confiné dans des domaines délimités par des
biomembranes Dans cette quatrième contribution originale, nous étudions la conformation d'un polymère isolé, qui est confiné entre deux membranes lipidiques parallèles ou dans une vésicule tubulaire. Pour rester plus général, nous supposons que le polymère est de topologie arbitraire, qu'on appelle D-manifold, où D est la dimension spectrale (par exemple, D =1, pour les polymères linéaires, et D =4?3, pour les polymères branchés). En fait, D est le nombre de coordonnées locales permettant de caractériser géométriquement le polymère 55
Statistics of a single D-manifold restricted to two
parallel M. Benhamou* , K. Elhasnaoui, H. Kaidi, M. Chahid Laboratoire de Physique des Polymères et
Phénomènes Critiques abstract The purpose is an extensive conformational study of a single polymer immersed in an aqueous medium (good solvent) delimitated by bilayer membranes. To be more general, we assume that the polymer is of arbitrary topology we term D-polymeric fractal or D-manifold, where D is the spectral dimension (for instance, D = 1, for linear polymers, and 4/3, for branched ones). The main quantity to consider is the parallel extension of the confined polymer. To make explicit calculations, we suppose that the polymer is restricted to a tubular vesicle or two parallel biomembranes. We first show that, for the first geometry, the polymer is confined only when the tubular vesicle is in equilibrium state. For the second geometry, the confinement is possible if only if the two parallel membranes are in their binding state, that is below the unbinding or adhesion temperature. In any case, the parallel gyration radius of the confined polymer is computed using an extended Flory-de Gennes theory. As result, this radius strongly depends on the polymer topology (through the spectral dimension D) and on the membranes sizes, which are the equilibrium diameter (function of bending modulus, pressure difference between inner and outer sides of the membrane, and interfacial tension coefficient), for the first geometry, and the mean-separation (function of temperature and interaction strength between the adjacent membranes), for the second one. Finally, we give the expression of the confinement free energy, as a function of the polymer size, and discuss the effects of external pressure or lateral tension on the radius expression for two confining parallel membranes. Key words: D-Polymeric fractals, Biomembranes, Vesicles, Confinement. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||