|

UNIVERSITE DE YAOUNDE I UNIVERSITY OF YAOUNDE
I
FACULTE DES SCIENCES FACULTY OF
SCIENCE
DEPARTEMENT DE BIOLOGIE ET PHYSIOLOGIE VEGETALES
DEPARTMENT OF PLANT BIOLOGY
STOCKS DE CARBONE DANS LES AIRES PROTEGEES DU TCHAD :
CAS DU PARC NATIONAL DE MANDA
Mémoire présenté et soutenu en vue de
l'obtention du Master Professionnel en Sciences de
l'Environnement
Option : Assainissement et Restauration de l'Environnement
Par:
NGABA WAYE TAROUM Caleb
Licencié ès Biodiversité et Gestion de
l'Environnement Matricule: 14Y2228
Encadreur: Superviseur:
DJEKOTA N. Christophe ZAPFACK Louis
Maître Assistant (CAMES) Maître de
Conférences
Université de N'Djamena
Année académique 2016-2017

DEDICACE
A mes parents, particulièrement à ma mère
SARATOU Isabelle, ma chère tante Mota Jeannette pour ses soins, sans
oublier mes frères Emmanuel & David NGABA ainsi que Roger TOYOUM qui
n'ont cessé de me soutenir depuis le décès de notre
père... Merci !
II
REMERCIEMENTS
Ce travail est le fruit d'un stage de spécialisation et
d'une étude de terrain, dont l'aboutissement est le présent
mémoire de Master Professionnel en Sciences de l'Environnement, option
Assainissement et Restauration de l'Environnement (SEN-ARE). Il n'aurait pas
été réalisé sans la contribution et le soutien de
nombreuses personnes dont certains seront ici remerciés; il s'agit
de:
- mon Superviseur de mémoire Zapfack Louis,
Maître de Conférences à l'Université de
Yaoundé I, nous lui seront infiniment reconnaissant ;
- Chef de Département de Biologie et Physiologie
Végétales, Pr. Emmanuel Youmbi pour la formation de
qualité ;
- mon encadreur Djekota Ngarmari Christophe, Maître
Assistant (CAMES), pour sa disponibilité et ses encouragements ;
- mon coordonnateur de Master, Pr. Djocgoue François, pour
son soutien ;
- Pr. Kengne Ives, Pr. Bell Martin, Dr. Mala Armand, ainsi
qu'à tous les enseignants du Département de Biologie et
Physiologie Végétales pour leur conseil et la maîtrise des
sujets relatifs à notre formation ;
- M. Kabelong Roger pour son expertise, Madjita Matthieu pour
sa disponibilité, Arnaud Choumele, Temfack Ingrid, Madountsap
Nadège, Ntonmen Amandine, Chimi Djomo et Zekeng Jules pour leurs
multiples apports et corrections du mémoire;
- A la direction de la Conservation de la faune et des Aires
Protégées du secteur Sud à Sarh (Tchad) ;
- au personnel du détachement de Manda et aux
écogardes pour leur accueil, à Masrabé Roland et toute
l'équipe de terrain, mon cousin Ninradingam Brahim, aux chefs coutumiers
de Manda et M. Saleh Doucia de la CST pour l'accès au laboratoire
d'Agriculture ;
- à mes oncles Ngakoutou Beneloum, Rassem Naham,
Allaissem Paul et Isaac Ndotam pour leur soutien matériel mais aussi
à Marie-France Ngaba et Fidel Blague pour leur appui morale et financier
durant cette formation;
- à Christine Amagna et Abraham Tokidang pour leur soutien
financier ;
- à toute la promotion de la filière SEN-ARE
2014-2016 ;
- à toutes celles et tous ceux dont je n'ai pu ici
nommer mais qui m'ont soutenu recevez ma profonde gratitude en ces mots.
III
SOMMAIRE
DEDICACE I
REMERCIEMENTS II
SOMMAIRE III
LISTE DES FIGURES V
LISTE DES TABLEAUX VI
LISTE DES ABREVIATIONS ET ACRONYMES VII
RESUME VIII
ABSTRACT IX
CHAPITRE I. GENERALITES 1
I.1. Introduction 1
I.2. Revue de la littérature 2
I.2.1 Définition des concepts 2
I.2.2. Changements climatiques 7
I.2.3. Atténuation des changements climatiques 9
I.2.4. Forêts pour épurer l'atmosphère 9
I.2.5. Point sur la REDD+ 11
I.2.6. Mesure des stocks de carbone 12
I.3. Présentation du site d'étude 14
I.3.1. Milieu physique 14
I.3.2. Aspects socio-économiques et culturelles 16
CHAPITRE II. MATERIELS ET METHODES 18
II.1. Matériels 18
II.1.1. Localisation géographique de la zone
d'étude 18
II.1.2. Zone d'échantillonnage sur le site d'étude
18
II.1.3. Matériel technique 18
II.2. Méthodes 19
II.2.1 Collecte des données 19
II.2.2. Traitement des données 25
II.2.3. Traitement des données secondaires 27
II.2.3. Traitement des données primaires 30
CHAPITRE III - RESULTATS ET DISCUSSION 33
III.1. Résultats 33
III.1.1. Caractérisation de la diversité
floristique du PNM 33
iv
III.1.2. Stocks de carbone 38
III.1.3. Etude diachronique des TUT du PNM de la
période allant de 1986 à 2016 39
III.1.4. Discussion 43
CHAPITRE IV - CONCLUSION-RECOMMANDATIONS ET PERSPECTIVES 48
IV.1. Conclusion 48
IV.2. Recommandations 48
IV.3. Perspectives 49
BIBLIOGRAPHIE 50
ANNEXES 57
Annexe I. Tableau des coordonnées géographiques
des transects 57
Annexe II. (A) Liste des espèces et leurs effectifs (B)
Liste des genres, espèces et genre 58
Annexe III. Fiche de séchage et pesage de la biomasse
végétale 59
Annexe IV. Fiche d'inventaire 60
Annexe V. Liste des espèces-effectifs et pourcentage
par espèce 61
V
LISTE DES FIGURES
Fig. 1: Carte de localisation site d'étude 18
Fig. 2: Carte de localisation des transects du relevé
floristique 20
Fig. 3: Equipe d'échantillonnage 21
Fig. 4: Equipe de layonnage uilisé. 21
Fig. 5: dispositif d'échantillonnage du carbone
épigé et hypogé 22
Fig. 6: Disposition de quadrat 24
Fig. 7: Pointeur et prospecteur durant le comptage 26
Fig. 8: Correction image Landsat 28
Fig. 9: schéma illustrant les 03 étapes de
traitements des données SIG 29
Fig. 10: Types de végétation 30
Fig. 11: Pesage enveloppe sur la balance de précision
et mise en étuve des échantillons 31
Fig. 12: Densité absolue par transect au Nord-ouest du
PNM 34
Fig. 13: Classification modale des circonférences des
peuplements 35
Fig. 14:Structure horizontale du peuplement 36
Fig. 15: Structure verticale du peuplement du PNM 36
Fig. 16: Structure des peuplements dans le PNM 37
Fig. 17:Occupation du sol du PNM (1986) 39
Fig. 18:Occupation du sol du PNM (1998) 39
Fig. 19: Etat des TUT du PNM en 1986 ..40
Fig. 20: Etat des TUT du PNM en 1998 40
Fig. 18:Occupation du sol du PNM (1998) 40
Fig. 21: Occupation du sol du PNM (2006) 40
Fig. 20: Etat des TUT du PNM en 1998 41
Fig. 22: Etat des TUT du PNM 2006 41
Fig. 21: Occupation du sol du PNM (2006) 41
Fig. 23: Occupation du sol du PNM (2016) 41
Fig. 22: Etat des TUT au PNM en 2006 42
Fig. 24: Etat des TUT au PNM en 2016 42
Fig. 25: Pourcentage des TUT du PNM entre 1986 - 2016 42
vi
LISTE DES TABLEAUX
Tableau I: Services environnementaux du PNM 5
Tableau II: Caractéristiques des images du capteur
Landsat utilisées 27
Tableau III: Nombre d'espèces et de genres par famille
33
Tableau IV: Récapitulatif du stock total de carbone
38
VII
LISTE DES ABREVIATIONS ET ACRONYMES
AP : Aires Protégées
BM : Banque Mondiale
CDB : Convention sur la Diversité Biologique
CCNUCC : Convention Cadre des Nations contre les changements
climatiques
COMIFAC : Commission des Forêts d'Afrique Centrale
COP 21 : Conférence des Parties 21
DHP : Diamètre à Hauteur de Poitrine
FAO : Organisation des Nations Unies pour l'Agriculture
FEM : Fonds pour l'Environnement Mondial
GPS : Global Positionning System
MEA : Millenium Ecosystem Assesment
MERH : Ministère de l'Environnement et des Ressources
Halieutiques du Tchad
MNV : Mesurable, Notifiable et Vérifiable
NAMA : National Appropriate Mitigation Action
PCUCB/MC : Projet de Conservation et Utilisation Durable de la
Biodiversité dans le Moyen-
Chari
PNM : Parc National de Manda
PNUD : Programme des Nations Unie pour le
Développement
PNW : Parc National de Waza
RDC : République Démocratique du Congo
REDD+ : Réduction des émissions due à la
Déforestation et à la Dégradation des forêts et
aux
autres types d'utilisation de terres
SIG : Système d'Information Géographique
TUT : Type d'Utilisation des Terres
UICN : Union Internationale pour la Conservation de la
Nature
UNOPS : Bureau des Nations Unies pour les Services d'Appui aux
Projets
WWF : Fonds des Nations Unies pour la protection de la faune
et la flore sauvage
VIII
RESUME
Le Tchad a érigé 12,3 % de sa superficie en
aires protégées afin de conserver les grands pôles de
biodiversité dont le Parc National de Manda (PNM). Cette étude
qui s'est déroulé d'Octobre à Décembre 2016 dans le
PNM, a pour objectif d'évaluer les stocks du carbone dans les aires
protégées du Tchad. Les données ont été
collectées sur une superficie de 16,8 ha dans le Nord-Ouest du PNM.
L'inventaire floristique de tous les ligneux de diamètre à
hauteur de poitrine (DHP) = 4 cm a été réalisé sur
07 transects de 1,2 km de long et 20 m de largeur a permis de
caractériser la diversité floristique mais aussi d'estimer les
stocks de carbone des ligneux. Les stocks de carbone du sous-bois, de la
litière et des radicelles ont été collectés dans
126 quadrats (42 de 1 m de côté, 42 de 0,5 m de côté
et 42 de 0,2 m de côté). Une étude diachronique de la
dynamique de la végétation a été
réalisée pour les années 1986, 1998, 2006 et 2016. Ont
été inventorié au total 45 espèces reparties en 37
genres et 21 familles pour une densité moyenne des peuplements qui
était de 355 individus/ha. Trois classes de diamètre dominent les
peuplements : la classe de plants = 10 cm avec 51,12 % des effectifs (3049
tiges); la classe de [10-20[ cm avec 36,77 % (2193 tiges) et la classe de
[2030[ cm avec 9,03 % (539 tiges). Les classes de hauteur présentent une
structure avec : 13,29 % des effectifs appartenant à la classe des
plants = 4 m dominée par Anogeissus leiocarpus, Combretum collinum,
Gueira senegalensis, Terminalia laxiflora et Hymenocardia acida ;
19,29 % des effectifs appartiennent à la classe de [4- 7[ m,
dominée par Daniella oliveri, Piliostigma reticulatum et
Prosopis africana; et enfin 67,40 % des espèces ont une hauteur
supérieure ou égale à 7 m, les espèces qui dominent
cette classe sont : Khaya senegalensis, Parkia biglobosa et Bombax
costatum. La surface terrière est de 5,86 m2/ha.
L'équation pan-tropicale de Chave et al. (2005) a permis
d'évaluer les stocks de carbone des différents réservoirs.
Il en ressort que les ligneux stockent 23,82 #177; 0,004 tC/ha, le sous-bois
0,144 #177; 0,007 tC/ha et la litière 0,560 #177; 0,006 tC/ha.
L'étude diachronique montre que le couvert végétal occupe
78,75 % de la Superficie du PNM contre 17,46 % de sols nus et bancs de sable.
La biomasse végétale permettrait l'implémentation d'un
projet pilote REDD+, celle-ci constituerait une solution durable pour la
conservation et la réhabilitation des espaces déboisés.
Mots clés : Aires protégées, biomasse,
peuplement, stock de carbone, REDD+, PNM.
ix
ABSTRACT
Chad has erected 12.3 % of its surface area for protected
zones in order to conserve the major biodiversity poles of which the Manda
National Park (MNP) is included. The objective of this study which was done
from October to December 2016, is to assess carbon stock in Chad's protected
areas. The data were collected over an area of 16.8 ha in the NW of the MNP.
The planting inventory of all chest level diameter (DBL) trees = 4 cm was
carried out on 07 transects 1.2 km long and 20 m wide, which allowed to
characterize the floristic diversity and also to estimate the carbon stock of
the trees. Carbon stock of undergrowth, litter and rootlets were collected in
126 quadrats (42 of 1 m side, 42 of 0.5 m side and 42 of 0.2 m side). A study
Diachronic dynamics of the vegetation was carried out for the years 1986, 1998,
2006 and 2016. A total of 45 species were divided up into 37 genera and 21
families for an average stand density of 355 individuals / ha. Three classes of
diameter dominate the stands: the class of plants < 10 cm with 51.12 % of
the population (3049 stems); the class of [10-20 [cm with 36.77 % (2193 stems)
and the class of [20-30 [cm with 9.03 % (539 stems). The classes of the height
presenting the structure of: 13.29 % of the stem class = 4 m dominated by
Anogeissus leiocarpus, Combretum collinum, Gueira
senegalensis, Terminalia laxifolia and Hymenocartia acida; 19.29%
of the population belongs to the class of [4-7 [m, dominated by Daniella
oliveri, Piliostigma reticulatum and Prosopis africana; And 67.40 % of the
species have a height greater than or equal to 7 m, the species that dominate
this class are: Kaya senegalensis, Parkia biglobosa and
Bombax costatum. The basal area is 5.86 m2 / ha. The
pan-tropical equation of Chave et al. (2005) assessed the carbon
stocks of the different reservoirs. This shows that the woody stock stores
23.82 #177; 0.004 tC / ha, undergrowth 0.144 #177; 0.007 tC / ha and litter
0.560 #177; 0.006 tC / ha. The diachronic study shows that the vegetation cover
occupies 78.75 % of the MNP area against 17.46 % of bare soil and sandbanks.
Plant biomass would allow the implementation of a REDD + pilot project, which
would provide a sustainable solution for the conservation and rehabilitation of
deforested areas.
Key words: Protected areas, Plant biomass, stand, carbon
stock, sequestration, REDD+, MNP.

CHAPITRE I.
GENERALITES
X
1
CHAPITRE I. GENERALITES
I.1. Introduction
La Convention Cadre des Nations Unies sur les Changements
Climatiques (CCNUCC) adoptée en 1992 a été un pas de
géant dans la lutte contre le réchauffement climatique global
(Djomo, 2010). En son article 3, elle contraint les pays
développés visés à l'annexe I, à veiller
à ce que leurs émissions anthropiques gaz à effet de serre
(GES) ne dépassent pas les quantités qui leur sont
attribuées. Elle recommandait que le total de leurs émissions de
GES soit réduit d'au moins 5 % par rapport au niveau de 1990 durant la
période d'engagement allant de 2008 à 2012 (Anonyme, 1999). Le
protocole de Kyoto vient renforcer la CCNUCC en prescrivant l'utilisation des
écosystèmes terrestres pour diminuer la concentration de carbone
atmosphérique.
Les forêts séquestrent des quantités de
carbone plus importantes que les autres milieux terrestres ; elles le font par
le phénomène de la photosynthèse (Zapfack, 2005). La
biomasse forestière est estimée à trois échelles
spatiales successives : l'arbre, le peuplement et la région (Loubouta et
al., 2016). La forêt a trois grandes influences sur le
changement climatique : elle fournit un puit de carbone qui, lorsqu'il est
dégradé, est très sensible à l'évolution du
climat ; elle fournit un service environnemental unique en régulant le
CO2 de l'atmosphère et elle offre une alternative aux combustibles
fossiles (Alamgir et al., 2008 cit. Nguedia, 2015). Les
forêts tropicales contiennent 40 à 50 % du carbone terrestre et
jouent un rôle majeur dans le cycle global du carbone (Pan et
al., 2011 cit. Loubouta op.cit).
En Afrique centrale, à côté des
forêts tropicales du bassin du Congo, ils existent des formations
végétales plus délicates à savoir : les savanes et
les steppes. Malheureusement, ces écosystèmes ont, jusqu'ici,
reçu très peu d'attention comparé aux forêts denses
et humides (Ngomanda et al. ,2013). Toutefois, les pays d'Afrique
centrale ont intégré ces zones dans leur politique nationale de
réduction des émissions de GES du secteur forestier, notamment
par la promotion de mesures (compensation pour services environnementaux)
incitant à conserver ces espaces forestiers (Ngomanda et al.,
op cit.).
Les savanes comme les steppes sont menacées par la
désertification et les changements climatiques mais aussi par des
pratiques agricoles et environnementales non durables (Anonyme, 2003). Cela
entraîne une diminution de leur diversité biologique et
l'apparition d'espèces envahissantes (Saradoum et al. ,2012).
Quantifier les stocks de biomasse et de carbone contenus dans les forêts
tropicales et dans les savanes est devenu une
2
priorité internationale dans le cadre de la mise en
oeuvre du mécanisme REDD+ (Loubouta et al. ,2016).
Le Tchad dont plus de la moitié du territoire est
désertique, a une flore menacée par: la création des
terres agricoles, l'extension des villes, la recherche du bois de chauffe, la
pression pastorale, etc. Ces activités sont responsables de
l'avancée du désert et de l'insécurité alimentaire
(Bertrand et Lagnaba, 2011). Afin de conserver les grands pôles de
biodiversité que regorge son territoire, le Tchad a opté pour
l'aménagement d'un réseau d'aires protégées (AP)
constitué de 30 entités forestières classées et
représentant une superficie de 15 787 200 ha soit de 12,3 % du
territoire national (Worgue, 2012). Le Parc National de Manda (PNM) fait partie
de ce réseau d'AP. Cette initiative est d'autant plus utile lorsqu'il
s'avère qu'entre 1990 et 2000, le taux de déforestation annuelle
déterminée par la FAO pour le Tchad a été de 0,6 %
par an (Ouya, 2010).
Le PNM est riche en diversité faunique et floristique
(Saradoum et al., 2012). Cependant sa végétation, bien
conservée jusqu'à nos jours, est sous la menace de l'urbanisation
de la 3ème ville du Tchad : Sarh (Ouya, op.cit). Le
PNM demeure jusqu'à nos jours très peu étudié et
ses ressources naturelles très peu connue (Saradoum et al ,2012
cit. Ngavounsia, 2012). Ainsi ce thème entre en droite ligne
dans la mise en oeuvre de la Composante 2 du Projet Régional REDD+
(COMIFAC - BM - FEM) qui a pour mission de renforcer les capacités
techniques pour la mesure des stocks de carbone forestier (Anonyme, 2012 a).
D'où l'importance de ce thème pour mettre en valeur le service
écosystémique qu'est la séquestration du carbone dans le
PNM.
L'objectif général de cette étude est de
déterminer les stocks de carbone du Parc National de Manda.
Plus spécifiquement, il s'agit de :
- caractériser la diversité floristique du Parc
National de Manda ;
- déterminer la dynamique des TUT durant la
période de 1986 à 2016 ;
- évaluer les stocks de carbone
séquestrés dans le Parc National de Manda.
I.2. Revue de la littérature
I.2.1 Définition des concepts
I.2.1.1. Parc National
La loi N°14/PR/2008 portant régime des
forêts, de la faune et des ressources halieutiques au Tchad en son
chapitre 2, section 2 et article 103 définie un parc national tel une
partie du territoire national classée au nom de l'Etat où la
flore, la faune, les eaux, les
3
sites géomorphologiques, historiques et d'autres formes
de paysages qui jouissent d'une protection spéciale, et à
l'intérieur desquels le tourisme est organisé et
réglementé.
Un parc national est un périmètre d'un seul
tenant, dont la conservation de la faune, de la flore, du sol, du sous-sol, de
l'atmosphère, et en général, du milieu naturel,
présente un intérêt spécial qu'il importe de
préserver contre tout effort de dégradation naturelle, et de
soustraire à toute intervention susceptible d'en altérer
l'aspect, la composition et l'évolution (Anonyme, 1997 cit.
Banoho, 2013). Selon la classification de l'union internationale pour la
conservation de la nature (UICN) les AP de la catégorie II (Parc
National), sont de vastes aires naturelles ou quasi naturelles mises en
réserve pour protéger des processus écologiques de grande
échelle, ainsi que les espèces et les caractéristiques des
écosystèmes de la région, qui fournissent aussi une base
pour des opportunités de visites de nature spirituelle, scientifique,
éducative et récréative, dans le respect de
l'environnement et de la culture des communautés locales (Dudley, 2008
cit. Banoho, op cit.).
I.2.1.2. Ressource ligneuse
L'ensemble des végétaux occupant une même
portion de sol forestier constitue un peuplement forestier. En Afrique au Sud
du Sahara, les ressources ligneuses constituent la première source
d'énergie comestible, elles servent de bois d'oeuvres et aussi pour bien
d'autres usages par les populations riveraines des massifs forestiers (Anonyme,
2010). Dans le cas précis du PNM, l'accès à la ressource
étant formellement interdite aux populations riveraines, la ressource
est encore plus ou moins abondante et variée en fonction des sites
écologiques qu'on y rencontre (Saradoum et al., 2012). Au sein
du PNM la ressource ligneuse est considérablement diversifiée.
Elle comprend des arbres, arbustes, herbacées mais aussi des plantes
aquatiques (Ouya, 2010).
Selon le plan d'aménagement du PNM
réalisé pour la période 2011-2021, la
végétation du PNM est remarquablement conservée. Les
photographies satellitaires révèlent clairement l'absence
d'empiètement par l'agriculture ou de toutes autres activités
humaines. L'absence d'influence de l'homme est également
révélée par le nombre d'espèces
végétales présentes dans le PNM (plus de 507), alors que
la périphérie n'en comporte en moyenne que 250 par
échantillon de 1000 km2 (Antonínová et
al., 2014).
I.2.1.3. Populations riveraines et gestion de la
ressource ligneuse
Le PNM est situé dans l'une des zones les plus
peuplées du Tchad, avec une densité de 21,8 habitants / km dans
les quatre cantons périphériques au PNM (Zouglou, 2010). Cette
population en majorité rurale et pauvre a tendance à surexploiter
les ressources ligneuses en
4
périphérie du PNM de part une agriculture
extensive, la pâture du bétail en périphérie et au
sein du PNM, et une explosion démographique de plus en plus
exponentielle (Ouya, 2010).
Les relations entre la population de la
périphérie et le parc sont principalement conflictuelles, et sont
dues à l'accès aux ressources naturelles qui leur a
été interdit depuis la création du PNM (Ouya,
op.cit, Zouglou, 2010, Saradoum et al., 2012). Le
bétail transhumant et les boeufs d'attelage exercent de plus en plus une
pression importante sur la flore du PNM. Les transhumants passent
indifféremment d'un côté à l'autre du parc et
pénètrent dans les secteurs les moins surveillés. Certains
s'installent juste à sa lisière, afin de pouvoir y conduire les
troupeaux la nuit. La récolte des produits naturels (ramassage du bois,
de la paille, cueillettes de fruits, et plantes médicinales,
récolte des tubercules) par la population des habitants de la
périphérie est généralisée et clandestine
(Anonyme, 2011 a).
I.2.1.4. Historique et but de la création du
PNM
Avant la colonisation, le site du PNM actuel était une
zone naturelle gérée par les chefs traditionnels.
L'administration coloniale française classa cette zone en réserve
de faune en 1951, dénommée Réserve de Faune de Manda, car
elle débutait au village de Manda, sa superficie était alors de
104 000 ha. Elle devint une Réserve Territoriale en 1953, avec une
augmentation de sa superficie à 108 000 ha. Le 19 mars 1965, le
décret présidentiel n°56/EFC/1965, l'a érigée
en parc national « pour la propagation, la protection et la conservation
de la vie animale et végétale dans un intérêt
scientifique et éducatif » (Ouya, op.cit). Le
décret n° 56/EFC/1965 a été suivi en 1966 d'un
déguerpissement quasi-total des villages qui se trouvaient à
l'intérieur de l'AP (Koutou et Djoli y sont toutefois partiellement
restés). Parmi les villages déguerpis, on peut citer: Doguigui,
Doboro, Maïbouyou, Missi, Yanga, Mbaïnaka, Koro, Kemkian, Massa,
Gori, Kara, Ngargoli etc. Certains de ces villages ont aujourd'hui disparu
(comme Nda), d'autres ont changé de nom (Anonyme, 2011 a).
Le PNM proprement dit a été créé
sur le site de l'ancienne réserve le 19 Mars 1965 (décret
n°56/EFPC) afin de préserver la biodiversité typique de la
zone soudanienne, et plus spécifiquement :
« Ce parc national est constitué en vue de la
propagation, de la protection et de la conservation de la vie animale sauvage,
et de la végétation sauvage dans un intérêt
scientifique et éducatif au profit, à l'avantage et pour la
récréation du public » (art.2). Ses limites d'alors ont
étés modifiées par le décret n°243/PR/EFPC/PNR
du 23 Octobre 1967, portant la superficie à 114 000 ha par l'adjonction
d'une zone de collines rocheuses au Nord du parc dans le canton Niellem. La
végétation du PNM fait partie du biome « savanes et
forêts
5
claires soudaniennes » classé en danger par le WWF
et figurant parmi les 200 écorégions essentielles au monde pour
la conservation de la biodiversité globale (Worgue, 2012).
I.2.1.5. Services écosystémiques du
PNM
Les services fournis par les écosystèmes peuvent
être le maintien de la quantité et de la qualité de l'eau,
la fourniture de ressources issues de la biodiversité pour
l'alimentation, le carburant ou les médicaments, la séquestration
du carbone, la beauté du paysage et l'élevage d'espèces
sauvages favorisant le tourisme et l'écotourisme, et bien d'autres
choses. Les services fournis par les écosystèmes peuvent exister
à une échelle quelconque, depuis le niveau local ou national et
jusqu'au niveau international (Rane et Peter, 2009).
Selon le Millenium Ecosystem Assesment (2005) les services
écosystémiques sont répartis en trois grands groupes,
à savoir :
- les services d'approvisionnement,
- les services de régulation,
- les services à caractère socio-culturelles.
Les services écosystémiques offerts par le PNM,
le sont non seulement aux habitants de la périphérie, mais
également de manière plus large à la population
régionale, et jusqu'à la capitale N'Djamena située au bord
du Chari (Anonyme, 2011 a).
Tableau I: Services environnementaux du PNM
|
Groupe de service
|
Service environnemental
|
|
Services
|
- Plantes médicinales
|
|
d'approvisionnement
|
- bois de chauffe (bois mort)
|
|
- braconnage
|
|
- pêche clandestine
|
|
- produits forestiers non-ligneux
|
|
- pharmacopée
|
|
- fourniture d'eau à usage agricole et domestique
|
|
- réservoir du vivant
|
|
services de régulation
|
- Atténuation de l'effet des sécheresses
|
|
- Fonctionnement de l'écosystème naturel
d'inondation du
fleuve Chari, confluence des bassins versants du Chari (du
|
|
Bahr Sara, du Bahr Salamat)
|
|
- Régulation de l'érosion et des coulées de
boues
|
|
- zone humide (habitat d'oiseaux aquatiques), rechargement
des nappes phréatiques et prévention des
inondations
|
|
- zone de mise en défens pour les poissons du Chari et
Bahr
|
|
Sara
|
6
|
|
- purification et maintien de la qualité de l'air
- régulation du climat au niveau local, régional et
global
(stockage de carbone)
- préservation des berges du Chari et lutte contre
l'érosion des
sols par la persistance de la
végétation
|
|
services
|
socio-
|
- Sites sacrés (mares, forêts sacrées, sites
d'initiation,...)
|
|
culturelles
|
|
- témoin scientifique de la végétation
soudanienne d'origine
|
|
|
(non modifiée)
|
|
|
- Cultes traditionnels
|
|
|
- Maintien de la qualité de l'environnement olfactif et
sonore
|
|
|
- Chasse sportive et pêche de loisir en eau douce
|
|
|
- Support de recherche représentatif de la flore
soudanienne et
sahélo-soudanienne
|
|
|
- Développement des savoirs éducatifs et
excursions
|
I.2.1.6. Mode de gestion du PNM
La documentation actuelle indique que le mode de gestion du
PNM est participatif en ce sens qu'il découle d'un partenariat entre les
populations riveraines et les conservateurs du parc. Les populations de la
périphérie du parc bénéficient des avantages
procurés par le parc et apportent en retour leur soutien et leur
collaboration à la préservation des valeurs naturelles et
culturelles du parc. Elles diminuent les pressions qu'elles exercent sur les
ressources naturelles (Anonyme, 2011 a).
La stratégie de partenariat repose sur la
nécessité de s'assurer que la population de la
périphérie accorde son support aux actions de conservation.
Cependant le rôle régalien de l'administration du parc est bien
sûr d'assurer les activités de conservation, mais elle joue
également un rôle de facilitateur du développement en ce
sens qu'elle a, plus une vocation de catalyseur que de moteur du
développement (Anonyme, 2011 a). Ce mode de gestion constitue un
handicap à la mise en oeuvre du processus REDD+, celui-ci
nécessite une appropriation de la nécessité de
conservation par les populations riveraines du PNM. Cette appropriation dont il
est ici question implique une gestion participative effective des ressources
naturelles du PNM par les conservateurs conjointement avec les populations
riveraines. Pour pallier à cette situation le Plan d'aménagement
du PNM (2011-2021) prévoit de mobiliser des ressources
financières pour générer des projets de
développement à long terme pour les populations riveraines du
PNM, l'instauration des zones de gestion des ressources naturelles dans les
cantons périphériques au PNM et biens d'autres initiatives qui
7
favoriseront dans un court délai l'implantation d'un
éventuel projet pilote REDD+ dans le PNM (Anonyme, 2011 a).
I.2.1.7. Potentialité de mise en oeuvre de la
REDD+ dans le PNM
Le plan d'aménagement du PNM (2011-2021) prévoit
le montage d'un dossier REDD+. Il devra se faire sur la base des tonnes de
carbone présent dans le PNM. Sur le plan de sa diversité
floristique, le PNM est réputé pour la qualité de sa
végétation ligneuse ceci constitue un atout non
négligeable (Anonyme, 2011 a).
Le plan d'aménagement prévoit que les zones de
gestion des ressources naturelles qui seront établis dans les cantons
périphériques soient ajoutées à ce dossier, ce qui
augmentera le volume de carbone stocké et donc la
rémunération globale dont une partie bénéficiera
aux communautés gestionnaires de ces ressources naturelles.
Les retombées de la REDD+ devront être reparties
au niveau des communautés et gérer pour la conservation des
ressources naturelles (Anonyme, 2011 a). Car la possibilité de disposer
des retombées du mécanisme REDD+ nécessite de proposer,
dès la demande officielle, un mécanisme de gouvernance des fonds
indiquant leur utilisation. Le carbone stocké doit répondre aux
critères de durabilité établies par l'ONU-REDD :
Mesurable, Notifiable et Vérifiable en abrégé
système MNV. Cette mise en oeuvre sera faite dans le cadre des plans de
développement locaux successifs. Il est prévu l'instauration de
guichets du fonds fudiciaire du PNM. Ainsi le plan d'aménagement
actualisé du PNM prévoie utiliser une partie des retombées
du processus REDD+ pour alimenter ceux-ci.
I.2.2. Changements climatiques
Les changements climatiques se définissent comme :
« la modification du climat qui est attribuée directement ou
indirectement à l'activité humaine qui altère la
composition de l'atmosphère globale » (Rane et Peter, 2009).
Au début des années 1970, la question des
changements climatiques globaux a commencé à faire l'objet de
multiples conférences et recherches scientifiques avérées
pour connaître les causes de ce phénomène afin d'en trouver
des solutions. Il sera admis que les émissions de CO2 dues à la
déforestation et à la dégradation des forêts
représentaient environ
20 % des émissions annuelles de gaz à effet de
serre dans les années 1990, et environ 12 % aujourd'hui (Fayolle et
al., 2010). Cette prise de conscience aboutira à la
Conférence de Rio de Janeiro en 1992. Les Etats sortiront de cette
assise avec un plan d'action dit « AGENDA
21 ». Elle va conduire à la signature de plusieurs
autres conventions dont : la Convention Cadre des Nations Unies sur le
Changement Climatique (CCNUCC), adoptée le 9 mai 1992 et
8
le Protocole de Kyoto adopté en 1998. Sous
l'égide de la FAO en Juillet 2001, se poursuivront à Bonn
(Allemagne) des négociations sur les mécanismes de compensation
des émissions de carbone à travers les puits de carbone au cours
desquelles le rôle des forêts sera évoqué. Dans le
cadre de la CCNUCC, les bénéfices potentiels pour les parties non
visées à l'annexe I ayant diminué leurs émissions
de gaz à effet de serre seront basés sur des résultats
mesurés, reportés et vérifiés (MRV). La
précision de ces résultats aura une influence majeure sur les
compensations financières potentielles (Boulmane et al.,
2012).
I.2.2.1. Effet de serre
Pour comprendre pourquoi des changements climatiques sont en
train de se produire, il est essentiel de comprendre l'effet de serre. La Terre
reçoit la majeure partie de son énergie du soleil sous forme de
rayonnement à ondes courtes. Une grande partie de ce rayonnement solaire
incident traverse l'atmosphère pour atteindre la surface de la Terre. La
Terre absorbe une partie de cette énergie et en renvoie une partie dans
l'atmosphère sous forme de rayonnement infrarouge. Les principaux gaz
qui absorbent le rayonnement infrarouge sont le dioxyde de carbone (CO2), le
méthane (CH4), l'oxyde nitreux (N2O) et les hydrofluorocarbones (HFC).
Ces gaz piègent une partie des rayonnements infrarouges et les renvoient
à nouveau vers la surface de la Terre sous forme de chaleur, ce qui
provoque un effet de réchauffement connu sous le nom d'« effet de
serre ». L'effet de serre est nécessaire à la vie sur Terre
telle que nous la connaissons ; sans lui, la surface de la Terre aurait une
température inférieure en moyenne d'environ 35 ° C à
celle que nous connaissons. Toutefois, la combustion de combustibles fossiles
et la destruction des forêts ont entraîné un accroissement
considérable de la concentration des GES qui retiennent la chaleur dans
notre atmosphère. La proportion de ces gaz dans l'atmosphère
étant en augmentation, une plus grande partie du rayonnement est
absorbée et réémise vers la Terre sous forme de chaleur.
Au cours du XXe siècle, les températures mondiales ont
augmenté de 0,7 ° C (1,3 ° F). Si les concentrations de gaz à
effet de serre dans l'atmosphère continuent à augmenter, la
température moyenne à la surface de la Terre pourrait augmenter
de 1,8 à 4 °C (3 à 7 ° F) (Rane et Peter, 2009).
I.2.2.2. Gaz à effet de serre
Les gaz à effet de serre (GES) sont des gaz
libérés dans l'atmosphère (par l'activité humaine)
qui piègent la chaleur et contribuent ainsi au réchauffement de
la planète. Tous les GES contribuent aux changements climatiques, mais
les GES n'ont pas tous le même niveau d'impact ; le potentiel relatif de
contribution au réchauffement de la planète se base à la
fois
9
sur leur « durée de vie »
atmosphérique (le laps de temps pendant lequel le gaz restera dans
l'atmosphère) et sur leur capacité à absorber le
rayonnement infrarouge. Le potentiel de réchauffement climatique indique
le niveau d'impact relatif de chaque gaz sur le climat par rapport à
l'impact des émissions de dioxyde de carbone (CO2). Le dioxyde de
carbone est le gaz à effet de serre le plus souvent mentionné
dans le contexte des changements climatiques. Le Tonne d'équivalent
dioxyde de carbone (TeqCO2) : est l'unité de mesure utilisée pour
comparer les émissions des différents GES sur la base de leur
potentiel de réchauffement global (PRG) (Rane et Peter, 2009) Par
exemple:
? 1 tonne de CH4 a un effet équivalent à 23
tonnes de CO2 ;
? 1 tonne de N2O a un effet équivalent à 296
tonnes de CO2 (IPCC Working Group I Report cit. Rane et Peter, op.
cit.)
I.2.3. Atténuation des changements climatiques
Les mesures des stocks de carbone forestier prennent ainsi une
importance accrue pour les pays qui projettent de contribuer à
l'atténuation des changements climatiques via leurs activités
forestières (Picard et al., 2012). C'est en cela qu'en 2007, la
contribution des forêts à l'atténuation des changements
climatiques a ainsi été reconnu comme l'un des piliers de
l'agenda des changements climatiques post-2012 avec la décision sur la
réduction des émissions résultant de la
déforestation et la dégradation des forêts (REDD). La FAO
définit les forêts comme, des terres occupant une superficie de
plus de 0,5 ha avec des arbres atteignant une hauteur supérieure
à 5 mètres et un couvert arboré de plus de 10 %, ou avec
des arbres capables d'atteindre ces seuils in situ. Car les plantes vertes
absorbent le CO2 présent dans l'atmosphère par
photosynthèse, le carbone est stocké dans le feuillage, les
tiges, les systèmes racinaires et, surtout, dans le tissu ligneux des
tiges principales des arbres.
I.2.4. Forêts pour épurer
l'atmosphère
La contribution des forêts au cycle mondial du carbone
et leur rôle dans la lutte contre les changements climatiques a
été consacrée avec l'adoption en 1992 de la CCNUCC et plus
tard en 1997 du protocole de Kyoto (Antoine, 2004). Les forêts jouent un
double rôle dans les changements climatiques. Les forêts peuvent
être une source de GES, en émettant du dioxyde de carbone dans
l'atmosphère lorsqu'elles sont brûlées ou détruites,
et les forêts peuvent aussi agir comme un « puits », en
prélevant le dioxyde de carbone de l'atmosphère et en le stockant
sous forme de carbone dans leur biomasse au fur et à mesure de leur
croissance (Rane et Peter, 2009). En raison de la longue durée de vie de
la plupart des arbres et de leurs dimensions relativement importantes, arbres
et forêts sont de véritables réserves de carbone. Il
10
s'agit dans ce cas de Puits de carbone ou processus qui
extrait les gaz à effet de serre de l'atmosphère en les stockant
sous une autre forme (Zapfack, 2005). Le Groupe d'experts intergouvernemental
sur l'évolution du climat (GIEC) et d'autres comités
scientifiques estiment que jusqu'à 25 % de l'absorption du CO2 de
l'atmosphère est le fait des forêts (Anonyme, 2010). Ainsi
l'objectif poursuivi à travers le mécanisme de la REDD+, vise
à inciter les pays en développement à réduire leur
déforestation, permettant à la fois de limiter la perte de
biodiversité et d'éviter les émissions de GES (Joël
et Laurent, 2010). Dans une forêt, le carbone est stocké à
6 niveaux, que l'on appelle couramment `'réservoirs». La
conservation des forêts existantes permettra d'éviter d'envoyer
dans l'atmosphère les émissions liées à la
déforestation. La restauration des forêts réalisée
par la plantation d'arbres ou en facilitant la
régénération naturelle des arbres permettra
d'accroître la quantité de carbone que les forêts peuvent
retirer de l'atmosphère et stocker dans leur biomasse (Rane et Peter,
2009).
I.2.4.1. Paiements des services
écosystémiques
Les `'paiements des services fournis par les
écosystèmes `', également appelés Paiements pour
les services environnementaux (PSE) correspondent à un type de
disposition par lequel les bénéficiaires des services fournis par
les écosystèmes remboursent les fournisseurs de ces services. Les
services fournis par les écosystèmes concernés peuvent
être le maintien de la quantité et de la qualité de l'eau,
la fourniture de ressources issues de la biodiversité pour
l'alimentation, le carburant ou les médicaments, la séquestration
du carbone, la beauté du paysage et l'élevage d'espèces
sauvages favorisant le tourisme et l'écotourisme, et bien d'autres
choses. Le Protocole de Nagoya vient renforcer ce mécanisme de
compensation en exigeant un partage équitable des revenus issus de
l'exploitation des ressources naturelles entre avec les populations gardienne
des dites ressources. Les services fournis par les écosystèmes
peuvent exister à une échelle quelconque, depuis le niveau local
ou national et jusqu'au niveau international (les services internationaux
fournis par les écosystèmes sont souvent appelés `' biens
communs mondiaux `') et une approche de type PSE est applicable à tous
ces niveaux. Les dispositifs de paiement peuvent prendre la forme d'un accord
commercial entre des acheteurs volontaires et des vendeurs volontaires, comme
des sociétés de tourisme qui paient des communautés
africaines pour la protection de la faune locale (Rane et Peter, 2009).
11
I.2.5. Point sur la REDD+
Adoptée par la COP16 de la CCNUCC, la REDD+ comprend
des approches politiques et les incitations positives sur les questions
relatives à la réduction des émissions dues à la
déforestation et la dégradation des forêts dans les pays en
développement.
Le mécanisme REDD+ prévoit des compensations
financières pour les pays qui réduisent la déforestation
et la dégradation des forêts, et qui évitent les
émissions de gaz à effet de serre associées (Fayolle et
al., 2010).
L'idée est apparemment simple et séduisante :
financer la non-déforestation en octroyant aux pays concernés une
compensation financière calculée notamment sur les services
rendus par la forêt parmi lesquels bien évidemment
l'économie que représente le non déstockage du carbone
(Joël et Laurent, 2010).
A l'origine de l'augmentation de la température global,
il y a les GES, en premier l'élément carbone (C). C'est en cela,
que Houghton (1995) estime que les émissions correspondant au changement
de l'utilisation du sol (déboisement et augmentation des pâturages
et des terres cultivées) ont émis dans l'atmosphère un
dégagement net de 25 Pg C soit 25.106 tonnes de carbone entre 1850
à 1990. Puisque la conservation des forêts est reconnue comme une
solution de choix pour réduire les émissions de GES et augmenter
leur séquestration, la naissance d'un marché de carbone va voir
le jour, à la clé deux concepts essentiels dont :
l'évaluation du carbone séquestré et la méthode
allométrique de quantification de l'élément C.
La REDD+, en tant qu'idée, a tout d'une
réussite. Elle a suscité un grand enthousiasme quant aux
possibilités qu'elle offre de lancer le travail d'atténuation du
changement climatique de façon rapide et peu coûteuse. Nulle autre
idée pour sauver les forêts tropicales de la planète n'a
engendré un enthousiasme et un engagement de fonds comparables à
ceux suscités par la REDD+ (CIFOR, 2012).
La mise en oeuvre de la REDD+ comporte trois phases
(préparation, réformes des politiques générales et
action basée sur les résultats). De même les recherches sur
la REDD+ avancent sur trois générations (CIFOR, op.
cit). La première porte: sur la conception de la REDD+ et
capitalise les enseignements à dégager des expériences
associées passées. Elle se penche sur le contexte institutionnel
et comment gérer certains défis particuliers comme la fuite,
l'additionnalité et la permanence, et les politiques précises. La
deuxième génération quant à elle aborde: les
contours économiques, politiques et la mise en oeuvre de la REDD+. Elle
tente de répondre aux aspects qui entravent ou permettent les
décisions sur les politiques et projets de REDD+ ainsi que leur mise en
oeuvre. La troisième génération évalue: l'impact
12
de la REDD+ sur le carbone forestier et les moyens de
subsistance locaux. Elle aborde aussi les mécanismes de mesures des
résultats de la REDD+. En effet ces trois générations de
recherches se succèdent et suivent la mise en oeuvre de la REDD+
elle?même (CIFOR, op. cit).
En Afrique centrale, c'est-à-dire dans les tropiques,
très peu d'études ont étés menées sur
l'évaluation du carbone séquestré. Le constat est plus
alarmant pour les pays de la zone tropicale sèche, qui sont plus
préoccupés à combattre la sécheresse et son
corollaire (Ngomanda et al, 2013). Néanmoins les pays
forestiers du Bassin du Congo réuni au sein de la COMIFAC font
d'énormes progrès dans ce sens. C'est ainsi qu'en dehors du
derniers atelier scientifique sur les équations allométriques
tenu à Yaoundé en Avril 2013, il faut citer les travaux de :
Ibrahima et al. (2002), Zapfack (2005), Amougou (2009), Djomo (2010),
Ngomanda et al. (2013). Ces auteurs à l'unanimité
soutiennent que la capacité de séquestration d'un milieu
dépend du type d'utilisation dont il fait l'objet et de sa
diversité spécifique.
I.2.5.1. Application de la REDD+ au Tchad
Au Tchad très peu de données contribuant
à la matérialisation du processus REDD+ ont étés
collectées en dehors du dernier inventaire des GES de 1993 (CCNUCC, 2001
cit. Eco-ressources , 2012), du rapport sur l'analyse des mesures
d'atténuation appropriées au niveau national (NAMA) dans le cadre
d'un programme des Nations Unies (UN-REDD+) (Eco-ressources, op. cit.)
et de l'atelier sur la faisabilité du REDD+ d'Avril 2013 en
collaboration avec la FAO (Anonyme, 2013). L'étude la plus
récente en date est la proposition R-PP du Tchad finalisé en Juin
2014. Cela n'est pas étonnant lorsqu'il est observé que les
écosystèmes ouverts tels que les steppes, prairies et les savanes
ont souvent étés négligés dans l'évaluation
globale du Cycle de Carbone à cause soit disant de leur faible
capacité à séquestrer le Carbone (Ngomanda et
al., 2013). Il est cependant nécessaire d'estimer les stocks de
carbone pour déterminer les émissions nettes liées aux
savanes. Ces estimations étant obtenues et combinées à
l'étendue de la superficie déboisée ou
dégradée permettra de connaitre les mesures de densité du
carbone.
I.2.6. Mesure des stocks de carbone
Pour étudier l'importance de la
végétation dans le cycle du carbone, des modèles ont
été mis au point. Ces modèles tentent d'extrapoler les
résultats obtenus à l'échelle d'une parcelle à
celle d'un type d'utilisation des terres, d'un écosystème ou
alors d'une région. Il existe deux sortes d'estimation de biomasse ou de
quantité de carbone : la première utilise des
13
estimations dérivées d'échantillonnages
destructifs et la deuxième est fondée sur les bases de
données concernant des volumes de bois. Cette dernière
méthode utilise la statistique qui prend en compte les diamètres
ou la taille de l'arbre (Brown et al., 1989 cit. Zapfack,
2005).
Les stocks de carbone sont estimés à partir de
la biomasse des végétaux, c'est-à-dire de leur masse
sèche de matière organique (Sola et al., 2012). Les
approches existant pour estimer les stocks de carbone forestier dans les pays
tropicaux peuvent être regroupées en moyennes des biomes, en
mesures au sol et en mesures de télédétection (Gibbs et
al., 2007 cit. Anonyme, 2010). Des relations
allométriques sont nécessaires pour convertir les inventaires
forestiers et les données captées par les instruments de
télédétection de mesure de carbone. Un certain nombre de
relations existent au niveau mondial (Chave et al., 2005 ; GIEC, 2006
; Djomo, 2010), mais il est préférable d'élaborer des
équations spécifiques aux types de végétation du
pays dans ce domaine. Les équations allométriques sont des
modèles de régression utilisés pour estimer la biomasse
aérienne des arbres à partir d'informations sur la taille i.e.
variables facilement mesurables (diamètre et/ou hauteur) (Fayolle et
al., 2010)
Les données des inventaires forestiers sont le plus
souvent utilisées parce qu'un grand nombre de pays ont
déjà dressé au moins un inventaire. Mais peu de pays en
développement tels le Tchad disposent d'inventaires nationaux exhaustifs
(DeFries et al., 2006 cit. Anonyme, 2013 d). Ces inventaires
forestiers ou phytoécologiques effectués selon les zones
bioclimatiques permettent de déterminer le potentiel en production de
bois et de la biodiversité végétale. Pour cela, il faut
procéder d'abord à un échantillonnage stratifié qui
consiste à faire une carte de végétation sur la base des
imageries satellitaires (Landsat ou spot) afin de déterminer les
principales formations végétales (Anonyme, 2010). Dans chaque
placette à inventorier, tous les pieds d'arbre de diamètre basal
supérieur ou égal à 4 cm font l'objet de mesure et
d'identification.
Pour le calcul du volume de bois et de carbone
séquestré qui se fait après l'inventaire, le volume de
bois est déterminé sur la base de la formule mise au point par
Morel (1987) au Mali, présentée comme suit :
V= 10 * G*P
- V : volume de bois en m3
- 10: Constante ;
- G: surface terrière = C2/4ð ;
C=circonférence de l'arbre à 1,30 m au-dessus du sol ;
- P: précipitation moyenne annuelle sur le site ou la
station la plus proche exprimé en
mm.
14
Pour passer des volumes de bois aux quantités de
carbones, on utilise les relations suivantes:
- 1 m3 de bois exploité=1 tonne CO2 ;
- 1tonne de matière sèche (TMS)= 0,5 tonne de
carbone ;
- 1 tonne de carbone = 3,667tonnes CO2; soit 1tonne CO2 = 0,27
tonnes de carbone = 1 m3de bois.
En effet, les diamètres obtenus lors de l'inventaire
floristique sont utilisés pour évaluer la masse de carbone
séquestré par les individus recensés. L'équation
allométrique dite « dry » développée par Chave
et al. (2005) est une équation pantropicale adaptée aux
forêts sèches. Elle a été utilisé dans le
cadre de l'évluation des stocks de carbone au Mali (Anonyme, 2010).
Celle-ci permet de calculer la biomasse de chaque individu et de déduire
le carbone des ligneux du système étudié. Le GIEC (2006)
propose une équation pour les espèces locales. Cette
équation est la suivante: AGB=Exp [-2,134+2,530 ln(DHP)].
I.3. Présentation du site d'étude
Le PNM est situé dans la zone soudanienne au Sud-est du
Tchad, dans la Région du Moyen Chari. Il est limité à
l'ouest par la route Sarh - N'Djamena, au sud par le Bahr Sara, à l'est
par le fleuve Chari et au Nord par les rochers de Niellim (Saradoum et
al., 2012). Il est entouré par quatre cantons: Balimba au Sud,
Kokaga à l'Est, Djoli à l'Ouest et Niellim au Nord. Saradoum (op.
cit) affirment que pour faciliter l'étude de la distribution
des ressources naturelles, le PNM a été subdivisé en
quatre parties suivant les localités environnantes:
Nguéré, Djoli, Koutou et Niellim. La position géographique
du PNM, au Nord-ouest de la ville de Sarh, bordé par une route nationale
à l'ouest et le fleuve Chari à l'est, l'isole des autres AP du
pays (Anonyme, 2008 a).
I.3.1. Milieu physique
I.3.1.1. Relief
Le PNM présente un paysage varié. Ce paysage se
compose de cours d'eaux (Chari, Bahr Sara), de défluents le long
desquels on trouve des mares permanentes ou temporaires, des plaines
d'inondation sableuses, des bas plateaux cuirassés et des collines
(Anonyme, 2011 a).
I.3.1.2. Hydrographie
Le Réseau hydrographique du PNM est dominé par
le fleuve Chari et ses affluents à la limite Est du parc. A cela
s'ajoutent 37 mares dont 23 permanentes et 14 non-permanentes (Saradoum et
al., 2012). Le Bahr Sara traverse le parc dans sa partie ouest.
15
I.3.1.3. Climat
Situé dans la zone soudanienne, le PNM jouit d'un
climat de type tropical humide à deux saisons : une saison sèche
qui va de Novembre à Mars et une saison des pluies d'Avril à
Octobre. Les températures moyennes mensuelles vont de 21°C à
35°C (moyenne annuelle : 28°C) avec une humidité relative de
32 à 85 % (moyenne : 61 %). Pour les années 2000-2009, une
pluviométrie moyenne de 1 051 mm a été relevé, avec
certaines phases « sèches » comme en 2001 et 2005 avec une
moyenne de 929 mm. Une baisse moyenne de pluviométrie de 100 à
150 mm/an est donc constaté durant cette période (Anonyme, 2011
a).
I.3.1.4. Végétation
La végétation du PNM est constituée de types
écologiques mixtes. Elle regorge:
- forêt claire (en profondeur) : peuplement ouvert avec
des arbres de petite et moyenne
taille dont les cimes sont plus ou moins
jointives, l'ensemble du couvert laissant largement filtrer la lumière ;
au sol, les graminées sont peu abondantes et parfois
mélangées à d'autres plantes suffrutescentes ou
herbacées. En plus de ce type écologique propre à la zone
soudano-guinéenne, le PNM regorge plusieurs variantes de savanes dont la
succession varie en fonction du relief, du microclimat et du type de sol en
présence, ce sont les :
- savane boisée ayant des arbres et arbustes formant un
couvert clair laissant largement
passer la lumière,
- savane arborée avec des arbres et arbustes
disséminés sur le tapis graminéen,
- savane arbustive avec des arbustes uniquement, sur le tapis
graminéen,
- savane herbeuse où arbres et arbustes sont absents avec
uniquement un tapis
graminéen.
I.3.1.5. Faune
Selon une étude de l'UICN-PAPACO réalisée
en 2010, les espèces assez communes au PNM sont : l'hippotragus
(hippopotame), le guib harnaché, le cobe des roseaux, le
céphalophe de Grimm, l'ourébi, le phacochère, et les
primates (le babouin doguera, le patas et le grivet). Les espèces de
plus en plus rares (moins de 10 à 20 individus par espèces), sont
: le cobe de fassa, le cobe de Buffon, le buffle. Le bubale de Lelwel pourrait
avoir disparu du parc. La girafe (Giraffa cameloparda lisantiquorum)
aurait encore été vue en 2010. Les espèces disparues sont:
l'éland de Derby, l'éléphant, le lion et le damalisque
tiang. La zone du PNM était connue pour sa richesse faunique depuis les
années 1950, mais au début des années 1980, les
événements politico militaires qui ont frappé le pays ont
fait chuter le nombre des espèces présentes, et plus encore leurs
effectifs.
16
I.3.1.6. Sols
Le PNM est formé de plaines au sol minéral
hydromorphes de couleur beige, de même que dans le bassin alluvial du
Logone-Chari. A l'extrême nord, il y a l'érosion des sols sur
roches acides : granite, gneiss et rhyolite. Dans la zone latéritique du
Niellim, il y a formation des sols tropicaux ferrugineux avec des
concrétions de matériel sablo-argileux, beige. Mais rencontre des
vertisols, des alluvions, des sols sablo-argileux à argileux,
généralement de couleur sombre, formant des poches dans certains
endroit entre Koutou et Niellim. Au centre et au Sud du PNM, sont
exposés des sols ferralitiques rouges et des sols ferrugineux tropicaux
(Anonyme, 2011 a).
I.3.2. Aspects socio-économiques et culturelles
L'agriculture est l'activité principale qui se pratique
dans la périphérie du PNM (Ouya, 2010). Celle-ci jouit d'un
climat soudanien, caractérisé ces dernières
décennies par des irrégularités, des insuffisances et de
mauvaises répartitions spatio-temporelles des pluies. Les sols
ferralitiques et ferrugineux à dominance sablo-limoneuse sont fertiles
pour les cultures sous-pluies du coton, mil et sorgho et occupent 80 % de sa
superficie (Ouya, op.cit).
Le PNM est situé dans l'une des régions les plus
peuplées du Tchad (le Moyen-chari). En 16 années, de 1993
à 2009, la population de quatre cantons entourant le PNM a
quintuplé en passant de 17 914 habitants à 85 592 habitants. Vue
cette densité, les ressources naturelles que regorgent le PNM et sa
périphérie font l'objet de multiples sollicitations de la part
des riverains (Anonyme, 2011 a). En guise d'exemple : les besoins de bois et
charbon, principales sources d'énergies domestiques de la population
augmentent, et plus de 0,77 % des savanes arborées sont
déboisées chaque année. Cette zone
périphérique est fortement anthropisée et la
dégradation des terres croît de 0,3 % chaque année (Ouya,
op.cit).

CHAPITRE II.
MATERIELS ET
METHODES
17
18
CHAPITRE II. MATERIELS ET METHODES II.1.
Matériels
II.1.1. Localisation géographique de la zone
d'étude
Le PNM est situé à 25 km au Nord-Ouest de la
ville de Sarh, à environ 450 km au Sud-est de N'Djamena, et à 80
km de la frontière Centrafricaine. Il se situe entre les latitudes
9°20 - 9°50 Nord et les longitudes 17°45 - 18°20 Est ; son
altitude varie de 344 m à 691m (Fig. 3) (Saradoum et al.,
2012).

Chari
Fig. 1: Carte de localisation site d'étude
II.1.2. Zone d'échantillonnage sur le site
d'étude
L'échantillonnage a été
réalisé sur une superficie de 16,8 ha dans la partie Nord-ouest
du PNM. Cette phase a été réalisée sur la base
d'observations suivant la variation des types de végétation
présente le long du principal axe routier qui longe le PNM. Afin
d'éviter les effets de bord, l'échantillonnage a
été effectué à environ 2 km de l'axe routier
principal séparant le PNM des habitations.
II.1.3. Matériel technique
Pour mener à bien cette étude le matériel
suivant a été utilisé:
- une carte de la zone d'étude pour matérialiser
les transects à suivre; - des images Landsat pour évaluer la
dynamique de la végétation ;
19
- un GPS pour relever les coordonnées
géographique;
- une boussole pour orienter les transects et les quadrats;
- un appareil photo numérique pour les prises de vues;
- un décamètre pour mesuré les distances le
long du transect ;
- un ruban de 1,5 m pour la mesure des circonférences des
arbres ;
- des étiquettes et marqueurs pour préciser les
distances cumulées sur les jalons ;
- papier presse pour conserver les échantillons non
identifiées pour l'herbier ;
- un bloc note, des crayons et gommes pour les données
d'inventaires ;
- une machette pour ouvrir les transects ;
- des sachets plastiques noirs pour la collecte de la
litière, des herbacées et des fines
racines du sol ;
- des sacs de 100 kg pour la collecte globale des
échantillons afin de faciliter le
transport pour le laboratoire d'agriculture de la compagnie
sucrière du Tchad (CST-
Sarh) ;
- des enveloppes pour le séchage des échantillons
collectés à l'étuve ;
- une balance de capacité 2000 g pour peser les
échantillons avant et après séchage ;
- un tamis fin pour l'isolation des radicelles ;
II.2. Méthodes
II.2.1 Collecte des données
II.2.1.1 Collecte des données secondaires
La recherche documentaire a été faite dans la
bibliothèque du Centre National d'Appui à la Recherche (CNAR) de
N'Djamena, à la direction des AP du Tchad, à la
bibliothèque de la représentation de la FAO à N'Djamena,
à la direction des eaux et forêts et auprès du point focal
REDD+ du Tchad. A cela s'ajoutent les informations recueillies au laboratoire
d'écologie et de Systématique botanique de l'Université de
Yaoundé I, mais aussi par les recherches sur internet.
II.2.1.2 Collecte des données primaires
II.2.1.2.1. Méthodes de collecte des
données
Avant la réalisation de ce travail sur le terrain, la
carte du PNM a été consulté au préalable ; ceci
dans le but de déterminer dans le site d'étude, des points de
repère permettant d'établir les unités
d'échantillonnages (transects). Compte tenu du besoin d'obtenir des
unités d'échantillonnages représentatives et permettant
d'échantillonner le plus de formations végétales possibles
(Ntonmen, 2013).
II.2.1.2.1.1. Couverture
végétale
La couverture de la végétation a
été déterminée par la méthode des transects.
Elle a été utilisée par plusieurs auteurs en Afrique
centrale (Nkongmeneck, 1999; Hardy et Sonké, 2004 ; Zapfack, 2005 ;
Ntonmen, 2013 ; Banoho, 2013). Chacun des 07 transects prévus au plan de
sondage avait un point de départ situé à environ 2 km
après l'axe principal afin de minimiser les effets de bords et garantir
la fiabilité des données collectées. Chaque point de
départ est marqué par un piquet sur lequel sont inscrits au
marqueur les renseignements (numéro du transect, coordonnée GPS
du dit point, position du piquet par rapport à la longueur totale du
layon en mètre soit 0 m). Un inventaire des espèces est
réalisé dans tous les types de végétation
rencontrés le long de chaque transect. Le transect dégagé
par le machetteur (en coupant un minimum de végétation tout en
s'assurant de ne pas abîmer les arbres ou toutes autres plantes qui sont
comptées dans l'échantillon) est jalonné tous les 200 m et
porte les informations suivantes sur son dernier jalon : le numéro du
transect, la mention « Fin », la distance cumulée en
mètres soit 1200 m. L'identification des types de
végétation a été à l'aide de
caractéristiques discriminatives visuelles, ce sont : l'espacement entre
les plants, l'éclairage et les espèces présentes le long
du transect. L'importance de chaque TUT le long des transects permet
également d'évaluer les changements de couvert
végétal dans le Nord-ouest du PNM. Le dispositif
d'échantillonnage est présenté sur la figure 2, ci-dessous
:
20
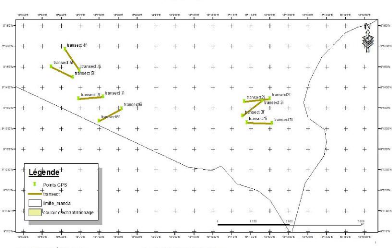
Fig. 2: Carte de localisation des transects du relevé
floristique
II.2.1.2.1.2. Méthode des transects
II.2.1.2.1.2.1. Equipe
d'échantillonnage
La collecte des données (comptage) est faite sur une
largeur de 10 m de part et d'autre de l'axe central du transect, soit une
parcelle de comptage rectangulaire totale de 2,4 ha mesurant 1,2 km de long sur
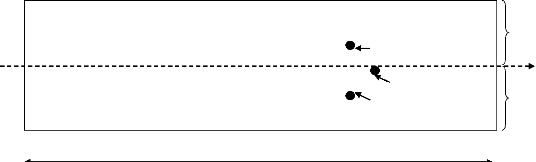
20 m de large (Fig. 2). L'équipe d'échantillonnage est
composée de trois (03) personnes dont :
- un pointeur qui enregistre les données dans les
fiches de collecte tout en consultant les prospecteurs si le nom de
l'espèce est la bonne ;
- et deux prospecteurs qui ont la charge de prospecter les
arbres dont la circonférence à 1,3 m du sol est supérieure
ou égale à 4 cm et aussi de mesurer l'arbre.
10m
Axe central du layon
Prospecteur de gauche
10m
Pointeur
Prospecteur de droite
1,2 km
Fig. 3: Equipe d'échantillonnage (comptage)
utilisé. (Adapté de: Ntonmen, 2013)
La phase d'échantillonnage est
précédée par celle du layonnage. L'équipe
responsable du layonnage pour cette étude est composée de 04
personnes (Fig.3) :
- un machetteur de tête appelé « traceur
» et aussi un boussolier qui doivent faire un dégagement sommaire
du transect tout en assurant la bonne orientation ;
- deux jalonneurs (chaîneurs) dont l'un à l'avant
tenant une extrémité du ruban de 100 m de long, coupant, plantant
et numérotant au fur et à mesure les piquets servant au
jalonnement, et le second à l'arrière tenant l'autre
extrémité du ruban et portant les échantillons
collectés.
Chaîneur

Axe central du layon Chaîneur Boussolier Traceur
Point de départ
Sens
d'évolution
21
Fig. 4: Equipe de layonnage uilisé (Adapté de :
Ntonmen, 2013).
Le dispositif déjà établi, sur chaque
arbre sélectionné, les paramètres à
considérer sont les suivants : l'essence qui est
déterminée sur la base de plusieurs caractéristiques
discriminants observés (silhouette générale, texture et
odeur de l'écorce/tranche, couleur, type et forme des feuilles...) ; la
circonférence (en cm) à 1,30 m du sol à l'aide d'un ruban
tailleur et enfin la hauteur du fût (en m). Il est à noter que
tous les prospecteurs ont des connaissances approfondies en essences
savanicoles. Les arbres inventoriés sont marqués pour ne pas
être comptés deux fois. Les espèces à contreforts
sont mesurées à 30 cm au-dessus de ces contreforts.
II.2.1.2.1.3. Estimation de la biomasse
végétale et des stocks de carbone
La collecte des données a eu lieu de Novembre à
décembre 2016 au sein du PNM. Elle a eu lieu en saison sèche,
cela explique le faible taux de biomasse végétale fraiche
prélevée. En plus à cette période sont
déclenchés les feux de brousse qui débarrassent le couvert
végétal de sa strate herbacée. A cause de cela, le long du
transect six (T6), la collecte des herbacées a été
très difficile.
Deux méthodes ont été utilisées
pour l'estimation du carbone. La méthode non destructive a servie
à l'estimation du carbone des ligneux de DHP = 4 cm (Ngomanda et
al., 2013). Elle prend en compte deux paramètres assez
accessibles à savoir : le diamètre et la densité
prédéfinie par l'équation allométrique de Chave et
al. (2005). Ces paramètres sont obtenus à l'issu des
relevés floristiques. Tandis que la méthode destructive a
été utilisée pour l'estimation du carbone
épigé et hypogé des strates inférieures (sous-bois,
litière, fines racines du sol). Les échantillons de
litière, de racines et des espèces coupées sur pied
grâce au dispositif décrit à la figure 5 et ensuite
séchés à l'étuve jusqu'à poids constant.
Lorsque le poids devient constant, cela signifie que toute l'eau contenue dans
la biomasse s'est évaporée
(Zapfack, 2005 ; Ngomanda op.cit).
0,5 m
1 m
20 cm
22
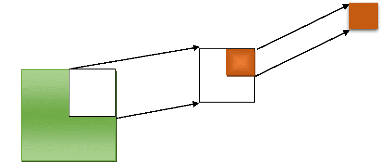
Fig. 5: dispositif d'échantillonnage du carbone
épigé et hypogé
23
Au total 126 quadras ont étés posés dont
: 42 de 1 m de côté, 42 de 0,5 m de côté et 42 de 20
cm de côté sur 50 cm de profondeur.
II.2.1.2.1.3.1. Evaluation du carbone
épigé
Les diamètres obtenus lors de l'inventaire floristique
sont utilisés pour évaluer la masse de carbone
séquestré par les individus recensés (Zapfack, 2005). Le
DHP est calculé par la formule C2/ð à l'aide du
tableur Excel 2013. L'équation allométrique
développée par Chave et al. (2005) a permis de calculer
la biomasse de chaque individu et de déduire le carbone des ligneux du
système étudié. Elle s'énonce comme suit :
Y= ñ*exp (-1,499+2,148lnD+0,207
ln(D)2)-0,0281(lnD)3)*0.001
Avec ñ=0,58 et 0,001 correspond à la conversion
en tonne/ha ; Y = biomasse (kg), ñ = densité du bois
(g/cm3), D = DHP (cm).
La quantité de carbone est obtenue en multipliant cette
biomasse par 0,47 (Hughes et al., 2000). Les arbustes dont le DHP
n'atteint pas 4 cm ainsi que les herbes (débarrassées de la
faune), sont collectés dans des quadrats de un mètre de
côté posé tous les 200 m le long de l'axe central du layon.
Ils sont coupés au niveau du sol, mis dans un sachet et refermés
hermétiquement pour éviter que le contenu des sachets ne se
mélange. Au laboratoire, ils sont transférés dans des
enveloppes, séchés à température constante et le
poids sec mesuré. Après avoir coupé la matière
verte dans les cadres d'un mètre de côté, les cadres de 0,5
m de côté sont utilisés pour collecter la litière.
Celle-ci est également enfermée dans des sachets plastiques. Ces
échantillons sont ensuite transportés jusqu'au laboratoire
où ils sont empaquetés dans des enveloppes puis
séchés dans une étuve jusqu'à poids constant. Leur
masse sèche est ensuite pesé à l'aide d'une balance de
précision (Zapfack, op.cit ; Ngomanda et al.,
2013).
III.2.1.2.1.3.2.Estimation du carbone hypogé
Pour estimer la biomasse des petites racines lignifiées
et des radicelles, le sol les contenant est extrait dans les cadres de 20 x 20
cm à 50 cm de profondeur. Les souches ainsi que les racines
maîtresses sont évitées. Le long de chaque layon, six blocs
de 20 x 20 x 50 cm ont été prélevés. Les
échantillons de sol sont prélevés respectivement tous les
10 cm de profondeur (Boulmane et al., 2013). Vu que la descente sur le
terrain a eu lieu en saison sèche, les racines sont extraites des
échantillons de sol à l'aide d'un tamis et des gants, à
l'aide d'un tamis suffisamment fin pour ne pas perdre le matériel
végétal. Après plusieurs tamisages, les racines sont
entièrement retirées et le sol est jeté (Zapfack,
2005).

24
Fig. 6: Disposition de quadrat : A) quadrat de 1 m2,
B) prélèvement des fines racines du sol
Les échantillons de litière, de radicelles et
des herbacées prélever ont été
transférés au laboratoire du département d'Agriculture de
la CST de Sarh pour les analyses. La série de pesage des emballages va
être faite à l'aide d'une balance de capacité 2000 g, de
marque Sartorius. La biomasse fraiche et chlorophylienne
conditionnée en 107 échantillons étiquetés:
d'herbacées, litière, de fines racines et radicelles ont
été empaquetés dans des enveloppes pour être
séché à l'étuve à 75°C. Avant le
séchage à l'étuve, le poids des enveloppes vides a
été mesuré, puis le poids de la biomasse fraiche
emballée est mesuré aussi. Une fois dans l'étuve, les
échantillons séjournent 48 h. Ensuite 10 échantillons
choisi au hasard du lot sont pesés afin de vérifier la variation
entre le poids frais et celui en cours de séchage. Après ce
pesage de routine, les échantillons vont encore séjournés
48 heures dans l'étuve jusqu'à l'obtention d'un poids sec
constant. Le poids constant, permet de déduire que toute l'eau contenue
dans le matériel s'est entièrement évaporée
(Ngomanda et al., 2013). La valeur des stocks de carbone est
estimée en multipliant la valeur de la biomasse sèche par le
coefficient 0,47 (Zapfack, 2005).
II.2.1.3. Collecte des données
cartographiques
Une carte du PNM réalisé en 1986 a
été fournie par la direction régionale des Parcs du
Moyen-Chari. Le Projet pour la Conservation et l'Utilisation Durable de la
Biodiversité dans le Moyen-chari (PCUDB/MC) en charge du suivi du PNM a
aussi fourni des cartes actualisées d'occupation du sol dont une carte
aérienne réalisée en 2014.
CG =
Nombre d'espèce x 100
Nombre de genre
25
II.2.2. Traitement des données
II.2.2.1. Inventaire floristique
Les fiches de comptage ont étés
dépouillées manuellement puis saisies dans un tableur Excel. La
structure horizontale des peuplements a été obtenue grâce
à l'analyse de la structure diamétrique. Les diamètres des
arbres ont été répartis en sept modalités
d'amplitude 10 cm. Les hauteurs des arbres ont aussi été
regroupées en sept modalités d'amplitude dix centimètre,
ce qui a permis d'étudier la structure verticale des espèces
inventoriées. La densité absolue pour toutes les espèces
identifiées a été déterminée par le tableur
Excel.
II.2.2.2. Densité absolue
La densité absolue indique la valeur moyenne du nombre
d'individus de l'espèce par unité d'échantillon. Dans le
cadre de cette étude, elle a été calculée par
hectare. Selon la formule (Kigomo et al., 1990 cit. Banoho,
2013) :
Da =
ni
M
Avec Da : densité absolue, ni : nombre d'individus de
l'espèce, M : superficie totale des
unités échantillonnées.
II.2.2.4. Degré de présence
Le recouvrement de l'espèce ou degré de
présence est l'expression en pourcentage de la continuité de la
couverture végétale (Guinochet, 1973 cit. Banoho,
op.cit). La formule qui a permis de le calculé est celle de
Kacoste et Salonon (1969):
Avec ni : nombre de présence, N : nombre de
relevés, Rci : recouvrement de l'espèce
II.2.2.5. Diversité générique
Une flore est diversifiée lorsqu'elle renferme moins de
grands genres plurispécifiques (Aké Assi, 1984 et Saadou, 1990
cit. Banoho, op.cit). Ainsi le coefficient
générique est le rapport entre le nombre de genre et celui des
espèces.
La capacité de séquestration d'un
écosystème étant liée à sa diversité
spécifique, ce coefficient a permis de définir la
fiabilité de la végétation du PNM en termes de
séquestration du carbone.
II.2.2.6. Régénération
naturelle
La régénération naturelle concerne les
arbres de diamètre inférieur à 20 cm. Le taux de
régénération permet d'estimer les potentialités de
renouvellement des essences. Elle se calcule par la formule ci-dessous
(Anonyme, 2007 cit. Banoho, 2013)
Nombre d'individus de circonférence
inférieur à 20 cm
|
Tr =
|
|
x 100
|
|
Nombre total d'individus
|
26
II.2.2.7. Indice de Shannon-Weaver
La diversité Alpha permet d'évaluer le poids de
l'espèce dans l'occupation du sol en utilisant l'indice de
diversité de Shannon-Weaver (1949). Cet indice varie en fonction du
nombre d'espèces présentes. Il est d'autant plus
élevé qu'un grand nombre d'espèces participe dans
l'occupation du sol. Il s'exprime en bits par individu, et varie de la plus
faible diversité (0 bit) à la plus élevée (4,5
bits) (Kabelong, 2013 cit. Frontier et al. , 1995). La
formule utilisée est la suivante :
n
H' = -? Pi Log2 Pi
i=0
Pi: la proportion relative du recouvrement moyen de
l'espèce i (valeurs comprises entre 0 et 1) ; Log2
: le logarithme à base 2.
II.2.2.8. Equitabilité de Pielou
H'
Elle tend vers 0 lorsqu'il y a dominance et vers 1 lorsqu'un
maximum d'espèces participent au recouvrement (Frontier et al.,
1995 cit. Banoho, 2013).

Fig. 7: Pointeur et prospecteur durant le comptage
27
II.2.3. Traitement des données secondaires
II.2.3.1. Données cartographiques
Sur le terrain, une descente de prospection observatoire a
été effectuée allant de l'entrée du village Manda
à celle de Niellem. Elle a permis d'obtenir les données
géographiques des deux extrémités du PNM à l'aide
d'un GPS. Ensuite lors des travaux sur le terrain, les points de départ
et final du transect ont étés relevés afin de produire une
carte de relévé floristique. Enfin pour l'obtention des cartes de
la couverture du sol, le CNAR a fourni les images landsat de la zone
d'étude. D'autres images landsat ont étés
téléchargées via internet. Les logiciels utilisés
sont Erdas Imagine 2014, ArcGIS 10.2.2, Google Earth pour le traitement d'image
et pour la partie cartographie.
Le choix de ces logiciels de traitement et des images Landsat
est justifié par les raisons suivantes :
? elles sont disponibles gratuitement ;
? elles sont indiquées pour l'analyse de la dynamique
du couvert végétal (Anonyme, 2013 c).
Tableau II: Caractéristiques des images du capteur
Landsat utilisées
|
Acquisition
d'image
|
Capteurs
|
Path-Row
|
Bande
|
Wavelength(micromètres)
|
Resolution(mètres)
|
|
1987-10
|
Enhanced Thematic Mapper Plus (ETM+)
TM
(Thématic Mapper)
OLI TIR
|
184-052
184-052
|
Band 1
|
0.45-0.52
|
30
|
|
2002-10
|
|
Band 2
|
0.52-0.60
|
30
|
|
Band 3
|
0.63-0.69
|
30
|
|
2012-11
|
Band 4
|
0.77-0.90
|
30
|
|
Band 5
|
1.55-1.75
|
30
|
|
Band 6
|
10.40-12.50
|
60 * (30)
|
|
Band 7
|
2.09-2.35
|
30
|
|
Band 8
|
.52-.90
|
15
|
|
2016-10
|
28
Le traitement se déroule en 03 phases. Une phase de
prétraitement, une autre de classification et enfin la dernière
de post-classification. Le PNM pose un problème du point de vue Image
Landsat. En effet il se trouve entre deux scènes : la scène
(182,053) et la scène (182,054). Pour avoir une image complète de
la zone d'étude, il a donc fallu effectuer une opération de
Mosaiquage qui consiste à fusionner deux images pour n'obtenir qu'une
seule. La phase de prétraitement s'effectue comme suit: avant le
processus de classification, les images subissent une correction
atmosphérique. Les valeurs numériques des images Landsat sont
transformées en réflectance puis ces dernières subissent
une correction grâce à la méthode DOS1.
En ce qui concerne la correction géométrique,
les images sont en DATA_TYPE = "L1T". Pour ce travail, il n'est pas
nécessaire de procéder à une correction
géométrique. Cependant les images Landsat pour les
périodes 2006 et 2007 présentent des défauts. En effet, le
scan-line corrector (SLC), un mécanisme de capture du satellite est
tombé en panne le 31 mai 2003. Ce qui se traduit par des rayures sur les
images. Un traitement spécial fut donc nécessaire. Pour cela, les
images de 2006 ont subi des corrections (Fill Gab), les bandes ont
été remplies de manière à permettre la suite du
travail.

A
B
Fig. 8: Correction image Landsat : A) Image Landsat avec trous
noirs, B) Remplissage
La phase de classification : la méthode
supervisée est retenue pour la classification. Ceci à cause des
regroupements naturels selon des critères statistiques. La combinaison
de bande a permis de faire ressortir les principaux éléments
constitutifs de l'image et la sélection des zones
d'intérêts afin de déterminer les différentes
classes constitutives de l'image.
La sélection des zones d'intérêt est
validée par des vérifications sur Google Earth afin de produire
des données qui se rapprochent le plus possible de la
réalité. Six (6) grandes classes ont été retenues :
1= Savane herbeuse ; 2= Savane arbustive ; 3= Savane arborée ; 4= Sols
Nus + Banc de Sable ; 5= Forêt Claire ; 6=Eau. Les bandes 2, 3, 4 et 5
sont utilisées à l'aide du module i.cluster puis du module
i.Maxlik. Ce dernier module utilise l'algorithme du maximum de
vraisemblance.
La phase de post-classification : les images
classifiées sont ensuite importés dans ArcGIS pour une analyse en
profondeur des signatures spectrales. Un rapport statistique sur l'image est
généré montrant la dynamique du couvert
végétal. Le rendu final est généré sur
ArcGIS.
Images de
départ : Landsat
Correction atmosphérique
Images Corrigé
Mosaiquage
Images
|
Découpage de la
zone d'intérêt
|
Correction
éventuelle : GAP Fill
|
|
Image 1
Classificatio
Image n
Classification Supervisée
Analyse de la signature Spectrale
Classification
Méthode du Maximum de Vraisemblance
Image 1 en
Classes
Image n en
Classes
Traitement dans un SIG :
Vectorisation, Analyse,...
Rendu
Cartographique pour
29
Post Classification
Fig. 9: schéma illustrant les 03 étapes de
traitements des données SIG (Adapté de: Theodoridisel et
al. , 2010)
II.2.3.2. Limite de la méthode
Les images satellites récentes du site d'étude
n'ont pas étés obtenues faute de financement. L'image de
l'année 1996 a été remplacée par celle de 1998 pour
la raison susmentionnée. Ainsi les images qui ont servies à
l'évaluation de la dynamique de la
30
couverture végétale du PNM sont celles de :
1986, 1998 et 2016 au lieu des années 1986, 1996 et 2016 comme
initialement prévu dans le projet de recherche. Le PNM étant
très vaste, l'équipe n'a pas pu entrer au-delà de 5 km en
profondeur par rapport à l'axe routier principal pris comme
référence. Cela n'a pas été possible faute de moyen
de déplacement et par peur d'insécurité lier à
l'insuffisance des gardes forestiers pour assurer la protection contre les
bêtes sauvages qui s'agglomèrent en profondeur du PNM pendant la
saison sèche (Saradoum et al., 2012). Les feux de brousses
sciemment déclenchés par les agents forestiers pour
défricher n'ont pas permis de prélever les échantillons
sur certains des transects.

A B C
Fig. 10: Types de végétation : A) Surface
brûlée, B) Savane arbustive à tapis herbacée, C)
Savane arborée
II.2.3. Traitement des données primaires
Toutes les données obtenues à l'issu des
relevés floristiques et de l'estimation des stocks de carbone
épigé et hypogé sont introduites et traitées
à l'aide du tableur Excel 2013. Les données provenant des
relevés floristiques sont : la circonférence des arbres à
1,3 m du sol, la hauteur des arbres de DHP = 4 cm, les coordonnées des
points de départ et final des transects, et les différents types
de végétation rencontrée le long des transects. Pour
l'estimation des stocks de carbone, les données collectées pour
le traitement sont : le poids de la biomasse fraîche des
échantillons empaquetés et leur poids constant après
séchage à l'étuve. Toutes ces données sont
introduites et traitées dans le tableur Excel 2013 (Ntonmen, 2013).

31
Fig. 11: Pesage enveloppe sur la balance de précision et
mise en étuve des échantillons (de gauche vers la droite) au
laboratoire d'agriculture de la CST-Sarh

CHAPITRE III.
RESULTATS ET
DISCUSSION
32
33
CHAPITRE III - RESULTATS ET DISCUSSION III.1.
Résultats
III.1.1. Caractérisation de la diversité
floristique du PNM
III.1.1.1. Diversité floristique
Les familles les plus représentées sont les
Rubiaceae avec 6 genres et 6 espèces, les Combretaceae avec 4 genres et
5 espèces. Le tableau III récapitule le nombre de genre,
d'espèce et de familles obtenus suite à l'inventaire
floristique.
Tableau III: Nombre d'espèces et de genres par
famille
|
Famille
|
Genres
|
%Genres
|
Espèces
|
%Espèce
|
|
Rubiaceae
|
6
|
16,21
|
6
|
13,33
|
|
Combretaceae
|
4
|
10,81
|
5
|
11,11
|
|
Mimosaceae
|
3
|
8,10
|
4
|
8,88
|
|
Caesalpiniaceae
|
3
|
8,10
|
3
|
6,66
|
|
Moraceae
|
1
|
2,70
|
3
|
6,66
|
|
Annonaceae
|
2
|
5,40
|
2
|
4,44
|
|
Ebenaceae
|
1
|
2,70
|
2
|
4,44
|
|
Euphorbiaceae
|
2
|
5,40
|
2
|
4,44
|
|
Rhamnaceae
|
1
|
2,70
|
2
|
4,44
|
|
Verbenaceae
|
1
|
2,70
|
2
|
4,44
|
|
Asclepiadaceae
|
1
|
2,70
|
2
|
4,44
|
|
Capparaceae
|
2
|
5,40
|
2
|
4,44
|
|
Anacardiaceae
|
1
|
2,70
|
1
|
2,22
|
|
Balanitaceae
|
1
|
2,70
|
1
|
2,22
|
|
Bombacaceae
|
1
|
2,70
|
1
|
2,22
|
|
Fabaceae
|
1
|
2,70
|
1
|
2,22
|
|
Loganiaceae
|
1
|
2,70
|
1
|
2,22
|
|
Malvaceae
|
1
|
2,70
|
1
|
2,22
|
|
Meliaceae
|
1
|
2,70
|
1
|
2,22
|
|
Papilionaceae
|
1
|
2,70
|
1
|
2,22
|
|
Sapotaceae
|
1
|
2,70
|
1
|
2,22
|
|
Tiliaceae
|
1
|
2,70
|
1
|
2,22
|
|
Total
|
37
|
100
|
45
|
100
|
34
III.1.1.2. Densité absolue
III.1.1.2.1. Densité absolue au Nord-Ouest du
PNM
La densité absolue varie d'un transect à un
autre. Dans les transects prospectés au Nord-Ouest du PNM (T1, T2, T3,
T4, T5, T6 et T7) comme observé sur la figure 12, la densité la
plus importante est celle du transect T1 qui est de 451 individus/ha. Ce
transect est situé à plus de 2 km après la route
séparant le village de Nguéré du PNM. Cependant la plus
faible densité est celle du transect T5 avec environ 300 individus/ha.
Ce transect se situe à moins d'un km de l'axe routier séparant le
PNM du village Nangnda. Il y a été constaté des indices de
pâturages, la trace de braconniers et une activité de vente de
bois de chauffe mais aussi de charbon en bordure de route. Il y a aussi de
grande superficie décimé au passage du feu.
La densité moyenne est de 355 individus/ha. Le constat
réalisé est que la ressource ligneuse varie à
l'intérieur du PNM en fonction de son accessibilité aux
populations riveraines. Il y a aussi les intrusions des éleveurs
transhumants en quête de pâturage et les braconniers qui souvent
utilisent le bois pour fumée leurs gibiers, mais aussi pour dresser des
tentes durant leur séjour clandestin dans le PNM.
|
500 450 400 350 300 250 200 150 100 50
0
|
|
|
|
individu/ha
|
|
|
T1 T2 T3 T4 T5 T6 T7
Densité absolue / transect
|
|
Fig. 12: Densité absolue par transect au Nord-ouest du
PNM
35
III.1.1.3. Structure du peuplement
III.1.1.3.1. Classification des circonférences
par modalité
Le traitement des données des classes de
circonférence à 1,3 m du sol (Fig. 13) montre que la
majorité des individus se retrouve dans la classe 20-40 cm. Cette classe
représente 40,59 % des individus recensés. Pour les autres
classes les effectifs sont repartis comme suit : 24,58 % pour la classe 0-20 cm
; 20,87 % pour la classe de 40-60 cm ; 7,69 % pour la classe de 60-80 cm. Pour
la classe de 80-100 cm 3,68 % ; 0,97 % pour la classe 100-120 cm; 0,75 % pour
la classe 120-140 cm et 0,33 % pour la classe 140-160 cm. Les classes qui
suivent, à savoir : la classe 160-180 cm, la classe 180-200 cm, la
classe 220-240 cm ont un pourcentage d'individus variant de 0,10 à 0,13
%. Enfin les individus supérieurs à 240 cm sont de 0,067 %. La
classification modale des circonférences montre que seulement 6,25 % des
individus inventorié atteignent des circonférences > 80 cm.

Nbre d'individus/classe
3000
2500
2000
1500
1000
500
0
Classe de ciconférence
(modalités)
45
40
35
30
5
0
25
20
15
10
% D'individus/classe
Fig. 13: Classification modale des circonférences des
peuplements
III.1.1.3.1.1. Structure horizontale du
peuplement
La structure horizontale a été obtenue à
partir des circonférences à 1,30 m du sol de chacun des 5964
ligneux inventorier. La distribution des tiges par classes de diamètres
sur les 16,8 ha prospectés dans le Nord-Ouest du PNM met en
évidence certaines divergences structurales nettement
discriminées par le nombre de tiges. En effet, il y a une dominance de
l'effectif de la classe de diamètres = 10 cm avec 51,12 % des effectifs
soit 3049 tiges. Elle montre également que la représentation des
différentes classes de diamètres a une distribution selon une
dynamique exponentielle décroissante avec 36,77 % soit 2193 individus
appartenant à la classe [10-20[cm et seulement 9,03 % pour la classe
[20-30[cm. Enfin, il faut noter que le nombre d'individus de la classe des
grands diamètres (plus de 60 cm) est très faible. Ces peuplements
sont caractéristiques de la zone bioclimatique sahelo-soudanienne.
36
La structure horizontale des peuplements permet de constater
une prédominance des tiges d'avenir (Fig. 14).

Classe ( de moda l
diamètre ités) en cm
[50;60[
[40;50[
[30;40[
[20;30[
[10;20[
[0;10[
= 60
0 500 1000 1500 2000 2500 3000 3500
Effectifs/modalités Nombre d'individus
Fig. 14:Structure horizontale du peuplement
III.1.1.3.2. Structure verticale du
peuplement
Les espèces ligneuses de la zone d'étude ont
été classé en trois (3) modalités de 10 m.La classe
de hauteur des plants = 4 m représente 13,29 % des espèces. Dans
cette classe dominent les espèces telles Anogeissus leiocarpus,
Combretum collinum, Gueira senegalensis, Terminalia laxiflora et Hymenocardia
acida. La classe de [4- 7[m représente 19,29 % des espèces,
dans cette classe dominent les espèces comme : Daniella oliveri,
Piliostigma reticulatum et Prosopis africana. Cependant plus de
67,40 % des espèces ont une hauteur supérieure ou égale
à 7 m (Fig. 15). Les espèces dominantes par leur hauteur sont :
Khaya senegalensis, Parkia biglobosa et Bombax costatum. La
classe des hauteurs = 4 m est caractérisée par plusieurs
espèces savanicoles qui, bien qu'étant matures conservent une
taille relativement petite, comme : Flugia virosa, Strychnos
spinosa, Ziziphus mauritiana et bien d'autres espèces
(Fig. 16).
80
70
60
50
40
30
20
10
0
Fréquence (%)
[0;4[ [4;7[ = 7
Classe de hauteur (m)
Fig. 15: Structure verticale du peuplement du PNM

B
37
Fig. 16: Structure des peuplements dans le PNM. A) Peuplement
d'Hymenocartia acida ; B)
Flugia virosa.
III.1.1.3. Diversité générique et
spécifique
Le coefficient générique du site est de 82,22 %.
L'indice de Shannon-Wiener est de 3,51 bits/individu, il représente la
structure spécifique de l'échantillon. Cet indice est
élevé, ce qui traduit un grand nombre d'espèces
contribuant à l'occupation du sol et la faible organisation du
peuplement. L'équitabilité de Pielou est de 0,64. Cela montre que
le peuplement est moyennement équi-repartie, c'est-à-dire que la
répartition de l'effectif des individus par espèce
représentée dans le peuplement est approximativement la
même. Elle montre la participation des espèces telles que :
Terminalia laxiflora (1351 tiges), Combretum collinum (1088
tiges), Hymenocardia acida (935 tiges) et Anogeissus leiocarpus
(588 tiges) à la réprésentativité du fond
floristique du peuplement.
III.1.1.4. Régénération des
espèces
Le taux de régénération moyen des
espèces est de 24,58 %. Ce taux est relativement faible à cause
de la courte durée de l'unique saison des pluies qui ne compte parfois
que 4 mois en moyenne (fin Mai à Août), mais aussi par une
faiblesse des réserves d'eau du sol et l'ensoleillement souvent excessif
qui est à l'origine de l'interruption du cycle des semences en dormance
dans les sols, en attente des pluies pour la germination.
III.1.1.5. Surface terrière
Elle quantifie la concurrence entre les arbres d'un peuplement
forestier. La surface terrière d'un peuplement est d'autant plus
élevé que celui-ci est dense. Dans ce cas précis, la
surface terrière du peuplement est de 5,86 m2/ha. Il en
résulte que le peuplement est clair et donne un accès plus facile
à la lumière.
38
III.1.1.6. Indices de diversité
Les indices de diversité répertoriés dans le
tableau IV donnent un aperçu des éléments ayant permis la
caractérisation des types de végétation dans le PNM.
Tableau IV : Indices de diversité
|
Indice
|
Valeur
|
|
Densité absolue
|
355 ind/ha
|
|
Taux de sondage :
|
0, 014 %
|
|
Diversité générique :
|
82, 22 %
|
|
Indice de shannon
|
3, 51 bits
|
|
Equitabilité de Pielou
|
0, 64
|
|
CG
|
84, 09
|
|
Surface terrière
|
5, 86 m2/ha
|
|
Régénération Naturelle
|
24,58 %
|
III.1.2. Stocks de carbone
III.1.2.1. Stocks de carbone épigée
En appliquant l'équation pan-tropicale de Chave et
al., (2005) les stocks carbone obtenus selon les réservoirs donnent
: un stock de 23,82 #177; 0,004 tC/ha pour les ligneux, un stock de 0,144 #177;
0,007 tC/ha pour le sous-bois et les herbacées, et un stock de 0,560
#177; 0,006 tC/ha pour la litière.
III.1.2.2. Stocks de carbone hypogée
Le stock de carbone hypogée correspond aux valeurs
obtenues par le séchage et le pesage des racines fines du sol et les
radicelles. Les grosses racines n'ont pas été
prélevées afin de ne point porter atteinte à la nutrition
des arbres. La valeur du stock obtenu est de 3,129 #177; 0,007 tC/ha.
III.1.2.3. Stock de carbone total
Le stock de carbone total obtenu équivaut à la
somme de la Carbomasse (ligneux et radicelles) et de ceux de la litière
et du sous-bois.
Tableau IV: Récapitulatif du stock total de carbone
|
|
|
Puits de carbone
|
0,144
|
Stock carbone tc/ha
|
|
sous-bois
|
0,560
|
#177; 0,007
|
|
Litière
|
3,129
|
#177; 0,006
|
|
Racine
(DHP = 4 cm)
|
23,82
|
#177; 0,007
|
|
Ligneux (ve
|
et al20)
|
27,653
|
|
39
III.1.3. Etude diachronique des TUT du PNM de la
période allant de 1986 à 2016
Cette partie présente l'apport de la
télédétection dans l'analyse diachronique de la dynamique
du couvert végétal sur le PNM à partir d'images
satellitaires Landsat. L'analyse prend en compte l'occupation des sols du PNM
au cours des années : 1986, 1998, 2006 et 2016. Elle s'étale sur
une durée de trois décennies, ceci afin de pouvoir analyser
objectivement la dynamique de la végétation du PNM au cours de 30
dernières années. III.1.3.1. Analyse des scènes
landsat du PNM et Pourcentage des TUT de 1986-2016 III.1.3.1.1. Evolution des
éléments du paysage du PNM de 1986 et 1998
Une analyse de cet intervalle de douze ans entre l'occupation du
sol du PNM en 1986 (Fig. 17 et 19) et l'occupation du sol du PNM en 1998 (Fig.
18 et 20) permet de constater :
- une régression de 18,01 % des sols nues et bancs de
sable soit 17241,74 ha entre les années 1986 et 1998. La
régression de ce TUT a fait place à l'installation des savanes
arbustives avec 41,69 % d'augmentation de superficie et une
légère perte de superficie des savanes herbeuses ;
- la savane arborée a considérablement perdu de
surface avec une perte en surface de 9,33 % de sa superficie de 1986. La
forêt claire, déjà relativement faible, a aussi perdu 0,24
% de sa superficie de 1986 ;
- le lit du fleuve Chari acquiert une plus grande densité
au détriment des poches d'eau et mayo à l'intérieur du
PNM, cette redirection du flux d'eau donne l'aspect d'une perte en surface des
points d'eau dans le PNM.
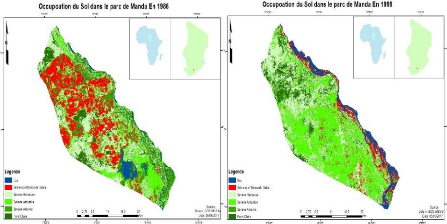
Fig. 17:Occupation du sol du PNM (1986) Fig. 18:Occupation du sol
du PNM (1998)

40
Fig. 19: Etat des TUT du PNM en 1986 Fig. 20: Etat des TUT du PNM
en 1998
III.1.3.1.2. Evolution des éléments du
paysage du PNM de 1998 et 2006
L'analyse des images landsat entre 1998 et 2006 a permis de
constater ce qui suit:
- les surfaces immergées ont perdues 2,91 % de leur
surface de 1998, cette régression a favorisé une extension des
sols nus et des bancs de sables de 6,78 % par rapport à leurs surfaces
de 1998 ;
- la savane herbeuse quant à elle a subit une augmentation
de 1,8 % de sa superficie par rapport à celle de 1998 et cela s'est fait
au détriment de la savane arbustive qui perd 7,86 % de sa surface de
1998 ;
- la savane arborée contrairement à la savane
arbustive, va s'accroitre de 4,16 % par rapport à sa superficie de 1998
tandis que la forêt perd en étendue environ 1,97 % de sa
superficie précédente ;

Fig. 18:Occupation du sol du PNM (1998) Fig. 21: Occupation du
sol du PNM (2006)

41
Fig. 20: Etat des TUT du PNM en 1998 Fig. 22: Etat des TUT du PNM
2006
Le choix de l'image landsat de 1998 au lieu de 1996 a
été opéré à cause d'une absence quasi-totale
des images satellitaires de la zone d'étude entre les années
2005-2007. Aussi, La superficie de 2006 est plus petite en termes de superficie
SIG du PNM (89894,45 ha) parce que le capteur a eu des problèmes
techniques durant la période de 2006. Les bandes noires apparaissent
dans les images et le traitement diminue donc la superficie totale de la zone
en 2006.
III.1.3.1.3. Evolution des éléments du
paysage du PNM de 2006 et 2016
L'analyse des images Landsat du PNM de 2006 et 2016 a permis de
savoir que :
- la savane herbeuse gagne en surface avec une augmentation de
9,9 % par rapport à l'année 2006, cela se fait au
détriment de la savane arbustive qui perd 7,6 % de sa surface de 2006
;
- la savane arborée gagne 3,1 % de surface au
détriment de la forêt claire qui perd 1,5 % de sa superficie par
rapport à 2006 ;
- les surfaces immergées perdent 0,9 % de leur
étendue par rapport à l'année 2006.

Fig. 21: Occupation du sol du PNM (2006) Fig. 23: Occupation du
sol du PNM (2016)
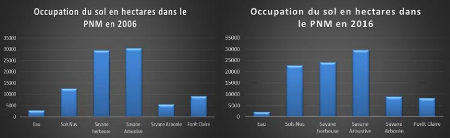
42
Fig. 22: Etat des TUT au PNM en 2006 Fig. 24: Etat des TUT au PNM
en 2016
De manière globale, l'étude diachronique permet
de constater une constante variation des TUT au sein du PNM au cours des trente
dernières années. La baisse de la pluviométrie moyenne
annuelle de plus de 100 mm/an, assortie de périodes de
sécheresses de trois à cinq ans, dans les années 70 puis
dans les années 80, et enfin de 2000 à 2005, a initiée une
« sahélisation » du biotope, plus nettement visible au Nord du
parc. Cet état de fait explique la constance de superficie de deux TUT
au sein du PNM durant les trente dernières années, à
savoir: la savane arbustive occupant en moyenne 32,3 % et la savane herbeuse
avec une moyenne de 28,4 % de la superficie totale du PNM.
Il faudrait notifier que les sols nus et bancs de sable
occupent en moyenne 17,4 % de la superficie totale du PNM sur les trente
dernières années. Ce résultat est d'autant plus
inquiétant, car les sols nues et bancs de sable constituent en termes de
superficie, le troisième TUT après la savane arbustive et la
savane herbeuse au sein du PNM. Au cours des trente dernières
années, la biomasse végétale du PNM (constituée de
la somme des différents types de végétation) occupe en
moyenne 78,75 % de la superficie totale du PNM. Cette biomasse constitue un
potentiel de séquestration de carbone non négligeable.

Fig. 25: Pourcentage des TUT du PNM entre 1986 - 2016
43
III.1.4. Discussion
III.1.4.1. Structure du peuplement
L'inventaire réalisé lors de cette étude
est systématique à transect de taille fixe. Pour Picard (2006)
(cit. Banoho, 2013) cette méthode a un double avantage : elle
est plus fiable, rapide et moins onéreuse. Ce même auteur
recommande que les transects soient de taille fixe lorsque le massif a beaucoup
d'arbre et de tailles plus réduites.
L'inventaire floristique a permis de recenser 5964 tiges dans
7 transects répartis en 45 espèces, 37 genres et 21 familles avec
un coefficient générique de 82,22 %. Les espèces les plus
représentées du point de vue de leur abondance dans le PNM sont :
Anogeissus leiocarpus, Combretum collinum, Gueira senegalensis, Terminalia
laxiflora et Hymenocardia acida. Ces résultats sont
proches de ceux de Banoho (2013) qui obtient 49 espèces ligneuses,
reparties en 42 genres et appartenant à 25 familles dans le Parc
National de Waza (PNW) situé dans la zone soudano-sahelienne au
Cameroun, avec un coefficient générique de 83, 67 %. Il en
ressort donc que le PNM est génériquement diversifié. Du
point de vue de l'abondance spécifique, les résultats obtenus
dans cette étude diffèrent de ceux de Banoho (2013) et de Jiagho
et al. (2016) qui obtiennent comme espèces dominantes au PNW:
Acacia seyal, Guiera senegalensis, Combretum molle et Balanites
aegyptiaca. Ils diffèrent aussi de ceux de Boussim (2010)
(cit. Lansina, 2013) en zone savanicole au Burkina, qui obtient des
comme espèces dominantes : Adansonia digitata, Faidherbia
albida, Lannea microcarpa, Parkia biglobosa, Vitellaria paradoxa.
Cette différence s'explique par l'indice de Shannon-Weaver qui est de 3,
51 bits pour cette étude, cet indice étant
légèrement supérieur aux 3,26 bits obtenu par Banoho
(2013) dans le PNW, montre la grande diversité spécifique du
milieu mais la faible organisation de son peuplement, ou encore le
déséquilibre de celui-ci.
Donfack (1998) conclut dans ses travaux sur la classification
des sols dans le Nord Cameroun que les sols ferrugineux sont dominés
dans les zones soudano-sahéliennes et sahélo-soudaniennes par les
espèces ligneuses telles que Combretum spp., Anogeissus
leiocarpus et Guiera senegalensis alors que les sols vertiques
sont dominés par Acacia
spp. et Combretum aculeatum. Ces
résultats démontrent que la texture et la composition chimique
des sols a une grande influence sur la diversité spécifique dans
les zones soudano-sahélienne et sahélo-soudanienne.
44
III.1.4.1.1. Densité moyenne
La densité moyenne des peuplements du PNM est de 355
individus/ha. Ces résultats sont différents des résultats
généralement obtenus dans les AP de la zone bioclimatiques
sahelo-soudanienne. C'est le cas par exemple de ceux de Banoho (2013) qui
obtient une densité moyenne comprise entre 30 à 50 individus/ha
et de Jiagho et al. (2016) qui obtient une densité moyenne de
40 tiges/ha dans le PNW. Tandis que les travaux de Sani (2009) au Niger
démontrent une densité moyenne des ligneux, de 163 individus/ha
dans un site reverdi à Marriah. La densité moyenne de cette
étude se rapproche plutôt de celle de la réserve
forestière de Lagdo dans le Nord du Cameroun en savane boisée,
avec une densité moyenne d'environ 300 tiges/ha et de Lansina (2013) qui
obtient dans le secteur sahelien strict au Burkina-faso une densité
moyenne de 305,25 individus/ha. La densité moyenne obtenue dans le PNM
s'explique par la bonne conservation de flore dans le dit parc dû
à la présence des sites de cultes et rites traditionnels mais
aussi à l'action des écogardes (Anonyme, 2011 a).
III.1.4.1.2. Surface terrière
La surface terrière des peuplements est de 5,86
m2/ha. Ces résultats sont contraires à ceux de
Boulmane et al. (2013) dans les écosystèmes secs du
Maroc, qui obtient 19,3 m2/ha dans la province d'Ifrane. Par contre
Traoré (2013) obtient dans la zone sahelo-soudanienne au Burkina une
valeur de 3,25 m2/ha, valeur qui se rapproche de celle de cette
étude. Les arbres de hauteur = 7 qui dominent les peuplements du PNM
sont assez distant les uns des autres vue la prédominance des
superficies occupées par la savane herbeuse et la savane arbustive,
d'où le manque de concurrence entre les individus expliquant la faible
surface terrière obtenue. Ces résultats permettent de conclure
que les peuplements du PNM sont clairs et donnent un accès plus facile
à la lumière. Aussi Ouaba (2006) dans la forêt
classée de Niangoloko au Burkina constate que la surface terrière
est fortement liée à la hauteur moyenne des individus.
III.1.4.1.3. Distribution du peuplement
L'étude de la distribution des circonférences
sur les 16,8 ha inventoriés démontre que 40,59 % des individus
appartiennent à la classe 20-40 cm. Banoho (2013) obtient un
résultat similaire dans le PNW au Cameroun avec 34,22 % des individus
appartenant à la classe 20-40 cm. Ce résultat est
caractéristique des écosystèmes savanicoles. Ils sont
généralement constitués de savane herbeuse, savane
arbustive et de savane arboré (Siaha, 1981 cit. Banoho,
2013).
45
Quant à la distribution diamétrique,
l'étude montre que 51,12 % des effectifs soit 3049 tiges appartiennent
à la classe de diamètres [0-10[cm. Elle montre aussi que la
représentation des différentes classes de diamètre a une
distribution à dynamique exponentielle décroissante avec 36,77 %
(soit 2193 tiges) appartenant à la classe [10-20[cm et seulement 9,03 %
à la classe [20-30[cm. Il faut noter que le nombre d'individus de la
classe de grands diamètres (plus de 60 cm) est très faible. Cette
différence dans le peuplement s'expliquent par le fait que les bosquets
forestiers se développent par un processus de succession naturelle
passant par plusieurs stade de maturité (Muller et al., 2002
cit. Ngomanda et al., 2012). Pour Jiagho et al.
(2016) ce résultat est un signe de vigueur écologique et de
garantie de la pérennité du peuplement, les jeunes individus
assurent spontanément le remplacement des individus âgés
disparus.
III.1.4.2. Estimation des stocks de carbone
Le stock de carbone total obtenu est en moyenne de 27,653
#177; 0,006 tC/ha. Il correspond à la somme de la carbomasse (ligneux et
racines), de la litière et des herbacées et du sous-bois.
L'équation de Chave et al. (2005) a été
utilisé pour les ligneux et la méthode destructive pour les
autres pools. En utilisant la même équation allométrique de
Chave et al. (2005) dans la forêt classée en zone
sahelo-soudanienne à Tiendaga au Mali, le stock de carbone obtenu est en
moyenne de 37,478 tC/ha (Anonyme, 2013 d). Tandis que Sanda (2015) à
Pitoa dans la zone sahelo-soudanienne du Cameroun obtient, en « savane
boisée à arborée » un stock de carbone de 4,70 #177;
1,09 tC/ha ; résultats largement inférieurs à ceux de
cette étude. Par contre pour un échantillonnage
réalisé sur 3700 ha dans la savane réhabilitée du
plateau de Bagombé au Gabon Ngomanda et al. (2013) obtient un
stock de carbone de 978113 tC soit une moyenne de 264,35 tC/ha, lui permettant
d'affirmer que la biomasse sèche du sous-bois des
écosystèmes savanicoles est plus considérable que ceux du
sous-bois des écosystèmes forestiers. Cette affirmation est due
au fait qu'en savanes le milieu étant très ouvert,
l'énergie lumineuse arrive directement à la strate herbeuses, ce
qui entraine une forte productivité de la biomasse en savane d'où
le fort taux de carbone stocké.
Le faible stock de carbone obtenu dans cette étude
s'explique par la taille de l'échantillon réduit de 16,8 ha, de
la saison durant laquelle l'évaluation est faite (saison sèche)
et enfin du fait que certains pools n'ont pas été estimer
à savoir : le sol, les grosses racines de plus de 2 mm de
diamètre. Ils ne l'ont pas été par manque de ressources
financières et logistiques.
46
III.1.4.3. Dynamique de la végétation du
PNM
La dynamique de la végétation du PNM entre 1986
et 2016 présente une variation moyenne d'environ 10 %. En effet la
couverture végétale est passée d'une superficie moyenne
totale de 88 % à une superficie moyenne de 78,75 % de l'étendue
du PNM. Ces résultats sont inférieurs à ceux d'Anonyme
(2015 d) à Lagdo dans l'extrême nord du Cameroun. Il obtient pour
la période de 1987 à 2014, une dégradation de près
de 30 % du massif forestier de Lagdo. Anonyme (2012 a) justifie la
densité du couvert végétal du PNM en concluant que sa
végétation est très bien préservée
jusqu'à ce jours. Toutefois, l'intrusion clandestine des populations
riveraines dans le parc pour diverses activités (pêche, chasse,
coupe de bois de chauffe et d'oeuvres) et des éleveurs à la
recherche de pâturage, sont les causes de l'augmentation de la superficie
des sols nus et bancs de sable. Banoho (2013) dans le PNW affirme que la
proximité du couvert végétal aux zones d'habitations
constitue une ménace pour la croissance de la végétation.
Le constat est le même pour cette étude. Cet état de fait
constitue une cause susceptible d'influencer la capacité de
séquestration de carbone de la végétation du PNM et de sa
périphérie, ceci par un déstockage des pools de carbone du
PNM sur le moyen terme.

CONCLUSION-
RECOMMANDATIONS
- PERSPECTIVES
CHAPITRE IV.
47
48
CHAPITRE IV - CONCLUSION-RECOMMANDATIONS ET
PERSPECTIVES
IV.1. Conclusion
L'inventaire floristique de 16,8 ha dans le Nord-Ouest du PNM
a permis d'obtenir 21 familles reparties en 37 genres et 45 espèces. Ce
résultat démontre à suffisance que le PNM regorge une
diversité spécifique considérable. L'analyse des cartes
d'occupation du sol du PNM montre que le couvert végétal occupe
78,75 % de la superficie totale du parc. Le PNM comprend différent types
de végétation dont : la savane herbeuse, la savane arbustive, la
savane arborée et les galeries forestières ou forêts
claires. Ce couvert végétal est menacé par la progression
des sols nus et bancs de sable, qui occupent 17,4 % de la superficie totale du
PNM. Cette dégradation du couvert végétal est due à
l'irrégularité des précipitations depuis le début
des années 1970 dans la zone d'étude, mais aussi à
l'intrusion clandestine des populations riveraines du PNM, des braconniers, des
éleveurs et des pécheurs qui prélèvent la biomasse
végétale. A cela s'ajoutent les feux de brousses qui sont
allumés périodiquement par les gardes forestiers eux-mêmes
occasionnant aussi une réduction du couvert végétal.
L'utilisation de la formule de Chave et al. (2005) a
donné un stock de carbone épigé de 23,82 #177; 0,004
tC/ha. Par contre dans le sous-bois elle donne un stock de 0,144 #177; 0,007
tC/ha. En ce qui concerne la litière, le stock obtenu est de 0,560 #177;
0,006 tC/ha. Pour le carbone hypogé, le stock obtenu est de 3,129 #177;
0,007 tC/ha. La biomasse sèche en savane étant plus importante
que celle du sous-bois de forêt, la carbomasse du PNM présente un
potentiel de séquestration de carbone conséquent et lui
permettant d'accéder à des fonds carbone. Cela est indispensable
pour garantir une gestion durable des ressources naturelles du PNM. Vu la forte
pression démographique que subit la biomasse végétale dans
la périphérie du PNM. L'apport d'un projet pilote REDD+
basé sur le potentiel de séquestration du carbone du couvert
végétal du PNM et de ses environs s'impose comme une solution
pour la conservation et la réhabilitation durable des espaces
déboisés.
IV.2. Recommandations Au
Gouvernement du Tchad :
· Réaliser un inventaire national de la flore
pour évaluer les stocks de carbone forestiers et définir les
moteurs de déforestation et dégradation de cette dernière
;
· créer un climat institutionnel propice à
la recherche sur le potentiel de séquestration de carbone du Tchad.
·
49
Initier un projet REDD+ pour le parc et sa
périphérie en guise de solution pour garantir une gestion durable
de la ressource ligneuse du PNM;
· augmenter les financements alloués à la
gestion des AP du Tchad. Aux partenaires nationaux et internationaux :
· Renforcer les capacités de gestion des
ressources naturelles dans les AP du Tchad ; Aux conservateurs du PNM :
· Inclure les populations riveraines dans la gestion des
ressources naturelles du parc ;
· pratiquer la méthode des feux de brousses
précoces utilisée pour régénérer la strate
herbacée et non la méthode sur brulis en saison sèche.
IV.3. Perspectives
- Réaliser une estimation des stocks de carbone pour
l'ensemble du PNM en fonction des types de végétations ;
- capitaliser les résultats de l'étude pour
réaliser un scénario de référence pour les types de
végétation présents dans le PNM et sa
périphérie ;
- lancer un projet REDD+ dans le cadre du plan
d'aménagement 2011-2021 du PNM et former les populations riveraines sur
les enjeux de la conservation de la flore face à la menace imminente des
changements climatiques globaux ;
- initier un système d'exploitation rotationnel du
massif forestier de la périphérie du PNM afin de permettre un
temps de régénération conséquent pour les surfaces
déboisées ;
- implémenter un projet de type Wood for Food
(Ngabulongo, 2012) pour le reboisement des savanes déboisées dans
la périphérie du PNM afin de répondre aux besoins de bois
énergie et bois d'oeuvre des populations riveraines tout en tirant
profit des ressources générées par le stockage de carbone
dans ses futaies régulières.

BIBLIOGRAPHIE
50
51
BIBLIOGRAPHIE
Abbo.A et Djimadjingar.A., Decembre 2011. Atelier Sur Les
Statistiques De L'environnement. Yaounde, Cameroun. 1-7p
Anonyme, 1999. Convention Cadre des Nations Unies Sur Les
Changements Climatiques (CCNUCC): inventaires des gaz à effet de serre,
Tchad 1999. 68p.
Anonyme, 2003. République du Tchad, juin 2003. Programme
d'action national de lutte contre la désertification (PAN/LCD).118p
Anonyme, 2006. Groupe International d'Expert sur le Climat.
« 2006 IPCC Guidelines for National Greenhouse Gas Inventories Volume 4
Agriculture, Forestry and Other Land Use ».
Anonyme, 2009.4ème Rapport National sur la
Diversité Biologique, Tchad. 73p.
Anonyme, 2010. Ministère de l'environnement et de
l'assainissement direction nationale des eaux et forêts manuel
d'inventaire forestier, Mali. 40p.
Anonyme, 2011 a. Plan d'aménagement du Parc National de
Manda 2011- 2021 Version provisoire Novembre 2010. Direction des Parcs
Nationaux, des Réserves de Faune et de la Chasse. 175p.
Anonyme, 2011 b. Atelier Régional d'Information et
d'Echanges sur les Forêts Modèles Et la mise en oeuvre locale de
la REDD+ dans le Bassin du Congo. 39p.
Anonyme, 2011 c. Plan d'aménagement du Parc National de
Manda pour la période 20112021. 175pp.
Anonyme, 2012 a. Etude qualitative sur les causes de la
déforestation et de la dégradation des forêts en
République Démocratique du Congo. Version finale rapport Groupe
de Travail REDD+.149p.
Anonyme, 2012 b. Composante 2 : Renforcer des capacités
techniques pour la mesure des stocks de carbone forestier. Rapport Projet
Régional REDD+. 10p.
Anonyme, 2013 a. Atelier scientifique régional sur les
équations allométriques en Afrique Centrale Yaoundé.
Cameroun. 91pp
Anonyme, 2013 b. Un atelier de sensibilisation, de formation et
de planification du projet MNV. FFBC-FAO-COMIFAC : Vers la
concrétisation du processus REDD+ au Tchad. Visité le 05/08/2015,
de
info@pfbc-cfbc.org
Anonyme, 2013 c. Guide Méthodologique estimation du
potentiel de séquestration du carbone au mali. Gestion
intégrée de la terre et de l'eau pour l'adaptation à la
variabilité et au changement climatique au Mali. 20p
Anonyme, 2014 a. Proposition pour la préparation a la
réduction des émissions liées à la
déforestation et à la dégradation des forets (R-PP -
REDD+). 156p
Anonyme, 2014 b. Méthodologie régionale
d'établissement des équations allométriques pour les
forêts du bassin du Congo. Etats des lieux des Equations
allométriques dans le bassin du Congo. 26p
52
Anonyme, 2014 c. Search for common ground. «
Prévention et gestion des conflits violents autour des ressources
naturelles partagées dans les régions du Chari-Baguirmi, Mandoul
et Moyen Chari». Rapport d'étude. 45p.
Anonyme, 2015 a. Quelles sont les types de forêts
présents sur la terre et quelles sont leurs différences ?. 6p.
Visité le 08/08/2015.
Anonyme, 2015 b. Aménagement des forêts
naturelles des zones tropicales sèches. Consulté le 05 Août
2015, de
www.fao.org/forestry.
Anonyme, 2015 c. Chapitre 5 - Les arbres et les forêts,
sources et puits de gaz à effet de serre et de carbone Le changement
climatique. Les forêts et l'aménagement forestier. Consulté
le 05 Août 2015, de
www.fao.org/forestry.
Anonyme, 2015 d. Note d'idée de projet. «
Conservation du massif de Ouro-doukoudje et reboisement des berges ouest du lac
de Lagdo ». 07 septembre 2015. 24p
Antoine C. 2004. Evaluation des stocks et des flux de carbone
liés à l'activité forestière en Aquitaine. Rapport
final. 88p
Antonínová M., Labuschagne J., Dogringar S.,
Assilek A., Dias J., 2014: Le recensement total aérien de Parc national
de Manda, Tchad, 2014, African Parks. 29p
Banoho. K. 2013. Influences des activités anthropiques
sur la ressource ligneuse dans la périphérie du Parc National de
Waza. Mémoire de Master en Biodiversité et Productions
Forestières. Université de Yaoundé I. 89p
Benoît. S., Jean. S., Mahamadou. B., &Maguette. K.
Guide d'estimation des réserves de carbone dans les
écosystèmes agrosylvopastoraux en Afrique de l'ouest.
Consulté le 27 Juillet 2015, website CILSS.
Bertrand. G. et Lagnaba. K. 2011. Potentialités et
contraintes du développement rural dans les régions du Tchad
central, oriental et méridional. Revue interne sur le secteur rural au
Tchad. Agence Française de Développement-Banque mondiale. 66p
Boulmane. M, M. Makhloufi, J.P Bouillet, L.
Saint-André, B. Satrani, M. Halim, S. El Antry, 2010. Estimation du
stock de carbone organique dans les Quercus ilex du Moyen Atlas Marocain.
Acta bot. Gallica, 157 : 451-467.
Boulmane. M., Ignacio.S-R., KHIA.K. et Hayat. O. 2012.
Estimation du stock de carbone organique dans les iliçaies du Moyen
Atlas marocain. 24pp.
Chave, J., Andalo, C., Brown, §., Cairns, M., Chambers,
J., Eamus, D., Fô1ster, H., Fromard, F., Higuchi, N., Kira, T. &
others. 2005. Tree allometry and improved estimation of carbon stocks and
balance in tropical forests. Oecologia, 145, 87 -99.
CIFOR. 2012 « Analyse de la REDD+ : Les enjeux et les
choix ». 520pp
Djimadoum. O. 1998. Identification des priorités dans
la conservation de la diversité biologique du Tchad. Rapport
d'étude. N'Djamena -Tchad. 44p.
Djomo A., 2010. Ecological management of tropical forests:
implications for climate change and carbon fluxes. Thèse de
doctarat. Buesgen-Institute, Chair of Bioclimatology, Buesgenweg. 120p
53
Dorian F., Matieu H. et Vincent M., Nicolas N. 2005.
Evaluation du stock de carbone et de la productivité en bois d'un parc
arboré à karité en zone soudanienne du Nord-Cameroun.
Ecole Nationale du Génie Rural des Eaux et des Forêts. Etude
réalisée par un groupe d'étudiants ENGREF avec l'appui de
l'IRAD et du CIRAD. 32pp
Dorisse M. D. 2005. "Etudes floristiques et ethnobotaniques
dans un village de la zone forestiere du Cameroun: cas de NKolbibanda".
Master thesis. Université de Yaoundé I, Yaoundé.
59pp
Fayolle A.G., Bouissou C., Nils B., Jean-Louis D., Picard N.
et Philippe L., 2010. Les équations allométriques pantropicales
sont-elles valides en Afrique centrale ? Journée d'étude sur le
mécanisme REDD+. Gembloux agro-bio-tech et Université de
Liège. 24p
Françis. A. 2010. Comparaison entre la forêt
tropicale et la forêt tempérée. Forêts tropicale
primaire. Episode 13, 1-7. Consulté le 08 Août 2015, de
www.wild-touch.org.
François. H., Victor. A., Kemeuze. S. et Raphaël.
N. Les espaces forestiers des savanes et steppes d'Afrique centrale. P.165-183.
Consulté le 08 Août 2015.
Houghton R. A., 2001. Counting terrestrial sources and sinks
of carbon. Climatic Change 48: 525-534.
Hughes, R.F., Kauffman, J.B. and Jaramillo, V.J. (1999).
Biomass, carbon, and nutrient dynamics of secondary forests in a humid tropical
region of México. Ecology 80: 1892-1907.
Ibrahima A., Schmidt P., Ketner P. et Mohren G. J. M., 2002.
Phytomasse et cycle des nutriments dans la forêt tropicale dense humide
du sud Cameroun. The Tropenbos-Cameroon Programme, Kribi,
Cameroon.Tropenbos - Cameroon Documents 9, XVI 81 p.
Jean. P. LANLY., 1963. Determination du taux
d'échantillonnage d'un inventaire statistique en fonction de la
densité des peuplements à inventorier. 1ère
section centrale de la station de recherches forestières de Nancy. 15
p
Jiagho, Zapfack L., Kabelong Banoho, Tsayem-Demaze M.,
Jeannine Corbonnois et Tchawa P. 2016. « Diversité de la flore
ligneuse à la périphérie du Parc national de Waza
(Cameroun) ». VertigO - la revue électronique en sciences
de l'environnement [En ligne], Volume 16 Numéro. URL :
http://vertigo.revues.org/17249
Kossi. D. 2010. Estimation de la biomasse
végétale du Sous-bois. Soutenance de Master de Biologie des
Populations et des Ecosystèmes. Université Des Sciences
et Techniques de Masuku. République du Congo. 16p
Loubota
P. G-J. et Doucet J.-L. 2016. Biomasse et
stocks de carbone des forêts tropicales africaines (synthèse
bibliographique). Bio techn. Agro. Soc. Environ. 20(4). p 508-522.
Magatte. F. D. 2010. Les consommations en Combustibles
domestiques dans la région de Fatick. Pérennisation des acquis du
PERACOD. 87pp.
54
Mangion I. 2010. « Le futur mécanisme REDD face
aux moteurs de la déforestation et de la dégradation des
forêts au Brésil, en Indonésie et en RDC ».
Mémoire de soutenance Master inédit.
Université Libre de Bruxelles, Bruxelles. 134p
Mertens B. et Boutrolle C. (2012). Diagnostic
préliminaire des centres ressources et disponibilité des
données. Composante 1'infrastructure d'accès aux données
satellitaires. UMR ESPACE-DEV, IRD. 33pp
Ngabulongo. 2012. Projet de boisement de la savane
dégradée d'Ibi Batéké pour la production de bois de
feu en RDC. Présentation scientifique. 36p
Ngavounsia T. 2012. Etude sur la diversité floristique
du Parc National de Manda au Sud-est du Tchad. Mémoire de soutenance
Master. Université de Yaoundé 1, Yaoundé.
50p
Ngomanda A., Lebamba J., Engone-Obiang N., Lepengue N. et
M'Batchi B. 2013. Caractérisation de la Biomasse sèche des
mosaïques forêt-savane des plateaux du Gabon. Journal of Applied
Bioscience. 68(1997), 5417-5425.
Nkongmeneck B.A., Tsabang N., Fongnzossie E., Balouma J.M.,
Apalo P. et Kamou E., 2003. Etude botanique du sanctuaire à gorille de
Mengame (Sud-Cameroun) : Resource ligneuses, facies de
végétation, degré de perturbation et sous-bois. The
Jane Goodall Institue-IRAD, rapport technique n 4, Yaoundé. 58
p.
Nke Ndi. J. 2008. Déforestation au Cameroun : Causes,
conséquences et solutions. Alternatives Sud. Vol 15(155).
Nguedia Darline. 2015. Evaluation des stocks de carbone dans
le massif forestier de Ngog-Mapubi-Dibang. Mémoire de Master.
Université de Yaoundé I.73pp
Ntonmen Y., 2013. Biodiversité et stock de carbone des
formations végétales d'Akomnyada et Nkologok (Mbalmayo).
Master, Université de Yaoundé I, 77 p.
Ouya Bondoro., 2010. Conservation et utilisation durable de la
biodiversité autour et dans les aires protégées du sud-est
du Tchad : cas du parc de manda et la forêt de Djoli-kera usages et
risques, quelles stratégies pour la revalorisation et le
développement local ? Thèse de doctorat en géographie
et aménagement de l'espace. Université
Paul-Valéry de Montpelier III. 276 p.
Picard. N. 2006. Méthode d'inventaire forestier. Projet de
développement rural participatif dans le moyen Atlas central (projet
Khenifra). <cirad-00147247>.43pp
Picard N., Saint-André L., Henry M., 2012.Manuel de
construction d'équations allométriques pour l'estimation du
volume et la biomasse des arbres : De la mesure de terrain à la
prédiction. FAO/CIRAD, Italie-France, 210 p.
Rane C. et Peter S. 2009. Cours d'introduction sur le
dispositif de Réduction des émissions liées à la
déforestation et à la dégradation des forêts (REDD)
Manuel de référence des participants. Rainforest- GTZ-the Nature
Conservanty-WWF. 140pp
Salifou TRAORE, 2008. Les formations à Acacia de l'Est
du Burkina Faso: typologie en relation avec les facteurs
pédoclimatiques, prédictions spatiales et fonctions sur la
dynamique du carbone et de l'azote. Thèse de Doctorat.
Université de Ouagadougou. 176 p.
55
Sani, A. R., 2009, Caractérisation biophysique des
ressources ligneuses dans un site reverdi et un site dégradé dans
le Département de MIRRIAH, Mémoire de fin de cycle de
l'obtention du Diplôme d'Ingénieur d'Eaux et forêts,
Université Abdou Moumouni de Niamey. 62p.
Sola G., Picard N., Saint-André L. et Henry M. 2012.
Résumé du manuel de construction d'équations
allométriques pour l'estimation du volume et la biomasse des arbres: de
la mesure de terrain à la prédiction. Organisation des Nations
Unies pour l'alimentation et l'agriculture, et Centre de Coopération
Internationale en Recherche Agronomique pour le Développement, Rome,
Montpellier, 20 pp.
Sonke B., 1998. Études floristiques et structurales des
forêts de la réserve de faune du Dja (Cameroun).Thèse
de doctorat : Université libre de Bruxelle (Belgique).267 p
Tabue R., Zapfack L., Noiha Noumi V., Nyeck B., Meyan-Ya
Daghela R., Ngoma L-R., Kabelong Banoho et Chimi Djomo C. Plant Diversity and
Carbon Storage Assessment in an African Protected Forest: A Case of the Eastern
Part of the Dja Wildlife Reserve in Cameroon. Journal of Plant
Sciences. Vol. 4, No. 5, 2016, pp. 95-101.
Tsopmejio T.- I. 2015. Biodiversité et stocks de
carbone dans les systèmes agroforestiers à base de cacaoyers de
Yambassa et «Evodoula-Okola» dans la Région du Centre au
Cameroun. Mémoire de Master en Sciences Forestières / Option
: Agroforesterie. Université de Yaoundé I. 46p
Tchobsala, A. Amougou et M. Mbolo, 2010, Impact of wood cut on
the structure and floristic diversity of vegetation in the peri-urban zone of
Ngaoundéré (Cameroon), Journal of Ecology and the Natural
Environment, 2(11), pp. 235-258.
Weldenson. D. 2010. Evaluation de la biomasse et des stocks de
carbone Sur des placettes forestières en forêts tropicales Humides
de Guadeloupe. Mémoire de soutenance Master inédit.
Université des Antilles et de la Guyane, Guadeloupe.
Zapfack L. 2005. Impact de l'agriculture itinérante sur
brulis sur la biodiversité végétale et la
séquestration du carbone. Thèse de doctorat.
Université de Yaoundé I, Yaoundé. 199p
Zougoulou. A. 2010. Gestion de la faune au Tchad.7p

56
ANNEXES
57
ANNEXES
Annexe I. Tableau des coordonnées
géographiques des transects
|
|
|
|
|
|
|
|
|
|
9,13,38.40
|
|
|
|
|
|
|
9,14,9.10
|
|
|
|
|
9,14,10.50
|
|
|
9,14,54.50
|
|
9,15,25.00
|
|
|
9,14,44.60
|
|
9,14,54.00
|
|
|
|
|
9,13,40.50
|
|
|
|
|
9,14,13.70
|
|

|
|
|
|
|
|
|
Noms scientifiques
|
Effectifs
|
|
Acacia ataxacanta
|
1
|
|
Acacia sieberiana
|
9
|
|
Amphiloxus pentafila
|
1
|
|
Anogeissus leiocarpus
|
587
|
|
|
|
Anona squamosa
|
32
|
|
Balanites aegyptiaca
Calotropis
|
1
|
|
|
|
Bitrospermum parkia
|
1
|
|
Bombax costatum
|
29
|
|
|
|
procera
|
1
|
|
Cedrocedrela cosqui
|
46
|
|
|
|
Combretum collinum
|
1082
|
|
Crosoperix februfurga
|
54
|
|
|
|
Daniella oliveri
|
137
|
|
Diospiros mespiliformis
|
5
|
|
Diospiros sp
|
2
|
|
Ficus glumosa
|
1
|
|
Ficus gnaphalocarpa
|
1
|
|
Ficus ingens
|
1
|
|
Flugia virosa
|
63
|
|
Gardenia erubescens
|
8
|
|
Gardenia ternifolia
|
68
|
|
Grewia venusta
|
229
|
|
Gueira senegalensis
|
45
|
|
Hexalobus monopetalus
|
14
|
|
Hibiscus squamosa
|
1
|
|
Hymenocartia acida
|
931
|
|
Kaya senegalensis
|
202
|
|
|
|
Mitracarpus gilio
|
23
|
|
Mitragyna inermis
|
1
|
|
Prosopis
|
|
|
Parkia biglobosa
|
383
|
|
Piliostigma reticulatum
|
202
|
|
|
|
africana
|
87
|
|
Sarcocephalus latifolus
|
8
|
|
|
|
Sclérocarya birrea
|
1
|
|
Strychnos spinosa
|
227
|
|
|
|
Tamarindus indica
|
12
|
|
Tephrosia bracteolata
|
1
|
|
Terminalia avicennioides
|
27
|
|
Terminalia laxifolia
|
1354
|
|
Vitellaria paradoxa
|
39
|
|
Vitex doniana
|
|
|
Vitex simplicifolia
|
|
Espèces
6
5
4
3
3
2
2
2
2
2
2
2
1
1
1
1
1
1
1
1
1
1
45
Genres
6
4
3
3
1
2
1
2
1
1
1
2
1
1
1
1
1
1
1
1
1
1
37
Famille
Rubiaceae
Combretaceae
Euphorbiaceae
Rhamnaceae
Verbenaceae
Asclepiadaceae
Capparaceae
Anacardiaceae
Balanitaceae
Bombacaceae
Fabaceae
Loganiaceae
Malvaceae
Meliaceae
Papilionaceae
Sapotaceae
Tiliaceae
Total
Mimosaceae
Caesalpiniaceae
Moraceae
Annonaceae
Ebanaceae
58
Annexe II. (A) Liste des espèces et leurs
effectifs (B) Liste des genres, espèces et genre
59
Annexe III. Fiche de séchage et pesage de la
biomasse végétale
Fiche de laboratoire
|
Ech
|
Masse du
sachet
|
Masse fraiche
|
Masse sèche
|
|
|
|
|
Date :
|
|
|
|
|
|
|
|
|
Code
|
|
|
+48h
|
+24h
|
+8h
|
+6h
|
+6h
|
+6h
|
+6h
|
+6h
|
|
LitièreL1
|
L1
|
|
|
|
|
|
|
|
|
|
|
|
L2
|
|
|
|
|
|
|
|
|
|
|
|
L3
|
|
|
|
|
|
|
|
|
|
|
|
L4
|
|
|
|
|
|
|
|
|
|
|
|
L5
|
|
|
|
|
|
|
|
|
|
|
|
L6
|
|
|
|
|
|
|
|
|
|
|
|
L...
|
|
|
|
|
|
|
|
|
|
|
|
Sous- Bois
|
SB1
|
|
|
|
|
|
|
|
|
|
|
|
SB2
|
|
|
|
|
|
|
|
|
|
|
|
SB3
|
|
|
|
|
|
|
|
|
|
|
|
SB4
|
|
|
|
|
|
|
|
|
|
|
|
SB5
|
|
|
|
|
|
|
|
|
|
|
|
SB6
|
|
|
|
|
|
|
|
|
|
|
|
SB...
|
|
|
|
|
|
|
|
|
|
|
|
Racine
|
R1
|
|
|
|
|
|
|
|
|
|
|
|
R2
|
|
|
|
|
|
|
|
|
|
|
|
R3
|
|
|
|
|
|
|
|
|
|
|
|
R4
|
|
|
|
|
|
|
|
|
|
|
|
R5
|
|
|
|
|
|
|
|
|
|
|
|
R6
|
|
|
|
|
|
|
|
|
|
|
|
R.....
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
60
Annexe IV. Fiche d'inventaire
Nom du pointeur : Date du jour :
N° du transect : Coordonnées GPS :
|
N°
|
Nom usuel
|
Nom
vernaculaire
|
Nom
scientifique
|
Circonférence
|
Hauteur
|
Usage
|
Obs
|
|
1
|
|
|
|
|
|
|
|
|
2
|
|
|
|
|
|
|
|
|
3
|
|
|
|
|
|
|
|
|
4
|
|
|
|
|
|
|
|
|
5
|
|
|
|
|
|
|
|
|
6
|
|
|
|
|
|
|
|
|
7
|
|
|
|
|
|
|
|
|
8
|
|
|
|
|
|
|
|
|
9
|
|
|
|
|
|
|
|
|
10
|
|
|
|
|
|
|
|
|
11
|
|
|
|
|
|
|
|
|
12
|
|
|
|
|
|
|
|
|
13
|
|
|
|
|
|
|
|
|
14
|
|
|
|
|
|
|
|
|
15
|
|
|
|
|
|
|
|
|
16
|
|
|
|
|
|
|
|
|
17
|
|
|
|
|
|
|
|
|
18
|
|
|
|
|
|
|
|
|
19
|
|
|
|
|
|
|
|
|
20
|
|
|
|
|
|
|
|
|
21
|
|
|
|
|
|
|
|
|
22
|
|
|
|
|
|
|
|
|
23
|
|
|
|
|
|
|
|
|
24
|
|
|
|
|
|
|
|
|
25
|
|
|
|
|
|
|
|
61
Annexe V. Liste des espèces-effectifs et
pourcentage par espèce
|
Nom d'espèce
|
Effectifs
|
% espèces
|
|
Acacia ataxacanta
|
1
|
0,01
|
|
Acacia sieberiana
|
9
|
0,15
|
|
Amphiloxus pentafila
|
1
|
0,01
|
|
Anogeissus leiocarpus
|
588
|
9,85
|
|
Annona senegalensis
|
32
|
0,53
|
|
Balanites aegyptiaca
|
1
|
0,01
|
|
Bitrospermum parkii
|
1
|
0,01
|
|
Bombax costatum
|
29
|
0,48
|
|
Calotropis procera
|
1
|
0,01
|
|
Cedrocedrela cosqui
|
46
|
0,77
|
|
Combretum collinum
|
1088
|
18,24
|
|
crossopteryx febrifuga
|
54
|
0,90
|
|
Daniellia oliveri
|
137
|
2,29
|
|
Diospyros mespiliformis
|
5
|
0,08
|
|
diospyros spp.
|
2
|
0,03
|
|
Ficus glumosa
|
1
|
0,01
|
|
Ficus gnaphalocarpa
|
1
|
0,01
|
|
Ficus ingens
|
1
|
0,01
|
|
Flugia virosa
|
63
|
1,05
|
|
Gardenia erubescens
|
8
|
0,13
|
|
Gardenia ternifolia
|
68
|
1,14
|
|
Grewia venusta
|
230
|
3,85
|
|
Gueira senegalensis
|
45
|
0,75
|
|
Hexalobus monopetalus
|
14
|
0,23
|
|
Hibiscus squamosa
|
1
|
0,01
|
|
Hymenocartia acida
|
935
|
15,67
|
|
Khaya senegalensis
|
202
|
3,38
|
|
Mitracarpus gilio
|
23
|
0,38
|
|
Mitragyna inermis
|
1
|
0,01
|
|
Parkia biglobosa
|
383
|
6,42
|
|
Piliostigma reticulatum
|
203
|
3,40
|
|
Prosopis africana
|
87
|
1,45
|
|
Sarcocephalus latifolius
|
8
|
0,13
|
|
Sclerocarya birrea
|
1
|
0,01
|
|
Strychnos spinosa
|
227
|
3,80
|
|
Tamarindus indica
|
12
|
0,20
|
|
Tephrosia bracteolata
|
1
|
0,01
|
62
Terminalia avicennioides
|
17
|
0,28
|
|
Terminalia laxifolia
|
1351
|
22,65
|
|
Vitellaria paradoxa
|
39
|
0,65
|
|
Vitex donania
|
25
|
0,41
|
|
Vitex simplicifolia
|
7
|
0,11
|
|
Ziziphus mauritiana
|
14
|
0,23
|
|
Ziziphus micronata
|
1
|
0,01
|
|
Total général
|
5964
|
100
|
| 

