|

Mémoire Universitaire présenté pour
l'obtention du diplôme de
Licence des Sciences de la Vigne
Promotion 2010 /2011
L'influence de l'alimentation hydrique de la
vigne
sur l'expression foliaire des maladies de l'Esca et du Black Dead
Arm
Réalisé par : Jean
DUFOULEUR1
Encadré par :
> Christine MONAMY2 : Responsable de la
coordination technique du Pôle Technique et Qualité > Soufiane
AYACHI2 : Chargé de missions d'études et
d'expérimentations
Responsable de formation :
> Marielle ADRIAN1 : Responsable
pédagogique et Maître de Conférences Habilité
à Diriger des Recherches
TABLE DES MATIERES
Remerciements
Liste des acronymes et abréviations
utilisées
INTRODUCTION GENERALE 1
CONTEXTE DU STAGE PROFESSIONNEL ET DU TRAVAIL PRESENTE 1
PARTIE I : RECHERCHE BIBLIOGRAPHIQUE 2
1) EPIDEMIOLOGIE DES MALADIES DE L'ESCA ET DU BLACK DEAD ARM
2
1.1 Contexte et nuisibilité 2
1.2 Eléments historiques 3
1.2.1 Historique de l'Esca 3
1.2.2 Historique du BDA 3
1.3 Symptomatologie 3
1.3.1 Caractéristiques générales et
différences entre les deux maladies 3
1.3.2 Symptomatologie de l'Esca 4
1.3.3 Symptomatologie du BDA 4
1.4 Agents pathogènes impliqués 5
1.4.1 Agents fongiques associés à la maladie de
l'Esca 5
1.4.2 Agents fongiques associés à la maladie du
BDA 5
1.5 Processus infectieux 6
1.5.1 Mécanismes menant à la dégradation du
bois 6
1.5.2 Mécanismes menant à l'expression des
symptômes foliaires 6
1.6 Interactions hôte-pathogène 7
1.6.1 Impact de l'Esca sur la vigne 7
1.6.2 Impact du BDA sur la vigne 8
1.6.3 Adaptation physiologique de la plante face à
l'agression 8
1.6.4 Complication pathologique : cavitation, embolie et
thyllose 8
1.7 Etiologie : étude des facteurs associés
à la condition pathologique 9
1.7.1 Facteurs étudiés pour les deux maladies
indistinctement (Esca/BDA) 9
1.7.2 Facteurs biotiques (Esca) 10
1.7.3 Facteurs abiotiques (Esca) 11
1.7.4 Facteurs biotiques (BDA) 11
1.7.5 Facteurs abiotiques (BDA) 11
1.8 Méthodes de lutte 12
1.8.1 Eléments d'information sur l'arsénite de
sodium 12
1.8.2 Moyens de lutte actuellement disponibles 12
1.8.3 Lutte en pépinière 12
1.8.4 Recherches en cours 13
1.8.5 Perspectives 14
2) ALIMENTATION HYDRIQUE DE LA VIGNE 14
2.1 Rôle et influence de l'eau sur la vigne 14
2.2 Notion de contrainte hydrique modérée :
entre le manque et l'excès d'eau 14
2.3 Besoins en eau de la vigne 15
2.4 Mouvements de l'eau dans le continuum
sol-plante-atmosphère (CSPA) 15
2.5 Régulation stomatique 16
2.6 Méthodes d'appréciation de la
réserve utile du sol et de l'état hydrique de la vigne 16
2.7 Techniques mises en oeuvre dans la gestion du
régime hydrique de la vigne 17
PARTIE II : MATERIEL ET METHODES 18
1) CARACTERISATION DES SITES D'ETUDES 18
1.1 Localisation et caractéristiques agronomiques
principales 18
1.2 Données géomorphologiques 18
1.3 Données pédologiques 18
1.4 Données climatiques 19
2) TECHNIQUES UTILISEES 20
2.1 Suivi du régime hydrique de la vigne 20
2.1.1 Notion de potentiel hydrique 20
2.1.2 Principe de la chambre à pression de Scholander et
techniques de mesure 20
2.1.3 Description et protocole d'utilisation de la chambre
à pression 21
2.1.4 Dispositif expérimental et mesure du potentiel
hydrique foliaire de tige (Øt) 21
2.2 Modélisation du bilan hydrique de la vigne
(modèle « Lebon etal., 2003 ») 21
2.2.1 Contexte d'utilisation 21
2.2.2 Principe de fonctionnement 22
2.2.3 Equation de modélisation 22
2.2.4 Paramètres d'entrée 22
2.2.1 Paramètres de sortie 23
2.3 Observation et notation des symptômes d'Esca/BDA
au vignoble 23
2.4 Traitement des données 23
PARTIE III: RESULTATS ET DISCUSSION 24
1) ALIMENTATION HYDRIQUE DE LA VIGNE 24
1.1 Potentiel hydrique foliaire de tige (Øt)
24
1.2 Modélisation du bilan hydrique 25
2) EXPRESSION DES MALADIES DU BOIS (ESCA/BDA) 27
3) RELATIONS ENTRE ALIMENTATION HYDRIQUE ET MALADIES DU BOIS
(ESCA/BDA) 28
CONCLUSION GENERALE 30
PERSPECTIVES ET AMELIORATIONS DU DISPOSITIF EXPERIMENTAL 30
ANNEXE I : Profils pédologiques des parcelles
d'études
ANNEXE II : Cartes de la résistivité
électrique des sols des parcelles d'études
ANNEXE III: Dénombrement des emplacements et
indication des taux de ceps jeunes, manquants et morts (non-
débourrés) sur les micro-zones d'étude
Résumé / Abstract
Remerciements
Je remercie vivement les différents organismes ainsi
que les personnes à l'origine de ce projet d'étude, et plus
particulièrement Jean Philippe GERVAIS (Directeur du pôle
Technique et Qualité au BIVB) et Christine MONAMY (responsable de la
coordination technique) avec qui j'ai été très heureux de
collaborer pendant ces quelques mois. Je remercie également
particulièrement Soufiane AYACHI (Chargé de Mission d'Etude et
d'Expérimentation) pour son implication et son soutien permanent, pour
la transmission cordiale de son vaste savoir et pour sa constante bonne humeur.
Ce travail lui est dédié : !
ÇÑß~Ô ß~á (Chokrane Jazilan
!).
J'adresse toute ma gratitude à l'ensemble des
responsables et du corps enseignant de l'Institut Universitaire de la Vigne et
du Vin de la faculté de Dijon, et plus particulièrement à
Marielle ADRIAN (Responsable de la formation & Maître de
Conférences), ainsi qu'à Sandrine ROUSSEAUX (Maître de
Conférences) et à Sophie TROUVELOT (Maître de
Conférences) pour la qualité de leur enseignement, leur
professionnalisme, leur disponibilité et leur soutien. Je remercie
également cordialement tous mes collègues de promotion que j'ai
été heureux de rencontrer et aux côtés de qui j'ai
apprécié étudier pendant cette année
universitaire.
Je tiens enfin à remercier l'ensemble des personnes
(dont je ne peux dresser ici une liste exhaustive) qui m'ont aidé de
près ou de loin à m'accomplir dans le monde de la vigne et du
vin, avec une attention particulière envers ma soeur Marie DUFOULEUR
ainsi qu'envers Alix JOJOT (OEnologue) et Romain DELILLE (OEnologue) pour le
soutien qu'ils m'ont apporté lors de mon projet de reprise
d'études.
Liste des acronymes utilisés
BIVB : Bureau Interprofessionnel des Vins de Bourgogne
DGAL : Direction Générale de l'Alimentation
DRAF : Direction Régionale de l'Alimentation, de
l'Agriculture et de la Forêt
ENITAB : Ecole Nationale d'Ingénieur en Travaux Agricoles
de Bordeaux
ENTAV : Etablissement National Technique pour
l'Amélioration de la Viticulture
IARC : International Agency for Research on Cancer
ICGTD : International Council on Grapevine Trunk Disease
INRA : Institut National de la Recherche Agronomique
ITV : Institut Technique de la Vigne et du Vin
IUVV : Institut Universitaire de la Vigne et du Vin
JORF. : Journal Officiel de la République
Française
MSA : Mutualité Sociale Agricole
ONIVINS : Office National Interprofessionnel des Vins
SDQPV : Sous-Direction de la Qualité et de la Protection
des Végétaux
SRPV : Services Régional de la Protection des
Végétaux
VINIFLHOR : Office national interprofessionnel des fruits, des
légumes, des vins et de l'horticulture
Liste des abréviations utilisées
ATSW : Available Transpirable Soil Water BDA : Black Dead Arm
CODIT : Compartmentalization Of Decay In Trees CSPA : Continuum
Sol-Plante-Atmosphère
El : Eutypa lata
Es : Evaporation du Sol
ETP : Evapotranspiration potentielle ETR : Evapotranspiration
réelle
Fm : Fomitiporia mediterranea
Fp : Fomitiporia punctata
FTSW : Fraction of Transpirable Soil Water Isv : Indice de Stress
de la Vigne
P : précipitations
Pal : Phaeoacremonium aleophilum
Pch : Phaeomoniella chlamydospora PRD : Partial Rootzone
Drying
R : Ruissellement de surface
R.U. : Réserve Utile
RDI : Regulated Deficit Irrigation
TDR : Time Domaine Reflectometry TTSW : Total Transpirable Soil
Water Tv : Transpiration de la Végétation
Introduction générale
Les maladies du bois de la vigne concernent les principales
zones de culture viticoles dans le monde et affectent toutes les régions
de production françaises. L'Esca et le Black Dead Arm sont deux acteurs
majeurs de ces maladies de dépérissement. Appelées
syndromes en raison de leur symptomatologie complexe, ces maladies sont
associées au développement lent et systémique de
champignons pathogènes dans le bois. Elles engendrent une perte de
rendement, affectent la qualité des moûts et entraînent une
mort prématurée des ceps, mettant en péril la
pérennité des vignobles. Elles constituent donc une lourde
menace, d'autant plus que l'arsénite de sodium, seul moyen de lutte
chimique considéré comme efficace vis-à-vis de l'Esca, a
été interdit d'utilisation en 2001. La définition de
nouvelles méthodes de protection suppose une meilleure connaissance de
l'étiologie et de l'épidémiologie. L'objectif de cette
étude est de préciser l'influence d'un facteur abiotique majeur :
l'alimentation en eau. Une première partie sera consacrée
à une recherche bibliographique sur les maladies du bois (Esca et Black
Dead Arm) et sur l'alimentation hydrique de la vigne. La présentation
des parcelles d'étude, les données météorologiques
et les méthodes mises en oeuvre au cours de ce travail feront l'objet
d'une deuxième partie. La troisième partie sera consacrée
à la présentation et à la discussion des résultats
obtenus.
Contexte du stage professionnel et du travail
présenté
Le stage professionnel clôture la formation en Licence
des Sciences de la Vigne, dispensée par l'Institut Universitaire de la
Vigne et du Vin (Dijon ; région Bourgogne ; France). Il s'est
déroulé du 14 juin au 30 septembre 2011, au Bureau
Interprofessionnel des Vins de Bourgogne (BIVB), dont le siège social
est installé à Beaune.
Le BIVB est une organisation reconnue par l'Etat
français et soumise à son contrôle économique et
financier. Sa vocation est de représenter et de défendre les
intérêts des vins de Bourgogne et des professionnels de la
filière (vignerons, coopérateurs et
négociants-éleveurs). Différentes actions sont
menées par cet organisme sur les plans techniques et économique,
ainsi qu'au niveau du marketing et de la communication. Il est financé
par les professionnels qui versent des cotisations individuelles et son budget
global est d'environ 10 millions d'euros. Chaque année, environ 15% de
ce montant est alloué au Pôle technique et Qualité.
Courant 2006, le BIVB a initié une augmentation
importante des travaux menés sur les maladies du bois en Bourgogne. Ces
études régionales sont complémentaires de celles qui sont
menées par le groupe national des maladies du bois1. Les
principaux objectifs sont de faire avancer les connaissances en
épidémiologie parcellaire de l'Esca et du Black Dead Arm, en
étudiant les poids respectifs des facteurs intrinsèques à
la plante et des facteurs environnementaux. Le travail présenté
dans ce mémoire est relatif à ce dernier point. Il porte sur
l'étude de l'influence d'un facteur abiotique sur ces maladies :
l'alimentation hydrique de la vigne.
1 Le groupe national des maladies du bois a été
constitué sous l'égide de l'Onivins (Viniflhor) suite au retrait
de l'arsénite de sodium en novembre 2001. Il implique les principaux
organismes techniques et de recherche (chambres d'agriculture, ENITAB,
ENTAV-ITV France, INRA, interprofessions, Service Régional de la
Protection des Végétaux, universités, et vignoble
Moët et Chandon) et représente les différentes
régions viticoles françaises. Une étroite collaboration a
été menée avec un groupe international créé
en 1998 : l'International Council on Grapevine Trunk Disease (ICGTD).

LES MALADIES DU BOIS DANS LE MONDE
British Columbia
Washington
Oregon
Californie ETATS-UNIS
Sud de la
Californie
MEXIQUE
?
Québec
EUROPE
?
Côte Est
?
?
?
TURQUIE
IRAN
?
?
EGYPTE
?
?
?
?
BRÉSIL
AUSTRALIE
CHILI
?
AFRIQUE DU SUD
Hunter Valley
ARGENTINE
Margaret River
Victoria
NOUVELLE ZELANDE
Présence d'eutypiose, d'esca et/ou de BDA et d'autres
maladies de dépérissement
Présence d'eutypiose et d'autres maladies de
dépérissement notamment liées à
Botryosphaeria
Australie méridionale
Pas d'eutypiose (zone semi-désertique,
précipitations inférieures à 300 mm / an) mais existence
d'autres maladies de dépérissement
? Pas de données précises sur les
maladies du bois dans ces pays
Les Maladies du Bois en Midi-Pyrenees, decembre 2004
42
PARTIE I : RECHERCHE BIBLIOGRAPHIQUE
1) Epidémiologie des maladies de l'Esca et du
Black Dead Arm 1.1 Contexte et nuisibilité
L'Esca, le Black Dead Arm (BDA) et l'Eutypiose2,
sont considérés comme les principales maladies du bois de la
vigne (Larignon, 2004-a). Ces maladies de dépérissement sont
associées à la présence de différents champignons,
capables de dégrader les tissus ligneux. Très dommageables pour
la pérennité du patrimoine viticole, elles concernent la
charpente des souches et compromettent le maintien et la
longévité de l'outil de production (Larignon, 2004-b). Elles sont
présentes dans les principales régions viticoles du monde (Figure
1). A l'étranger, d'autres maladies du bois proche de l'Esca se
manifestent : maladie de Petri, Black Goo et Esca «juvénile»
(Guérin, 2004). En France, selon Dubos (2002), ces maladies concernent
toutes les régions viticoles et affectent une part grandissante du
vignoble. Cet auteur estime que l'Esca est en passe de devenir le
problème phytosanitaire le plus important des vignobles
européens.
Selon l'observatoire national des maladies du bois mis en
place en 2003 sur 612 parcelles, 65% des parcelles présentent au moins
un symptôme d'Esca et/ou de BDA (Herlemont, 2004-a). Les
conséquences économiques et financières sont
particulièrement graves : coût de la recherche et de la lutte
préventive, pertes liées à la diminution du rendement,
pertes liées à la mort des ceps. Cette mortalité au
vignoble peut nécessiter un renouvellement des plants pouvant atteindre
plus de 10% (Grosmann, 2008). Cet aspect n'est pas ignoré des
investisseurs qui exigent aujourd'hui un bilan sanitaire avant toute
transaction patrimoniale (Dubos, 2002).
Les répercussions sur la qualité oenologique
sont également importantes : production excessive à
l'échelle du pied (si les ceps détruits ne sont pas
remplacés et que le rendement à la parcelle est maintenu au
même niveau), ou au contraire rajeunissement des parcelles (si des
remplacements importants sont effectués), entrainant dans les deux cas
une dépréciation de la qualité des vins. On note
également le risque d'abandon des cépages les plus sensibles, ce
qui pourrait entrainer une perte de la typicité des vins des
régions concernées (Larignon, 2009).
Depuis l'interdiction de l'utilisation de l'arsénite de
sodium en 2001 dans tous les pays viticoles pour des raisons de toxicité
humaine et environnementale (Spinosi et al., 2009), les viticulteurs
redoutent une recrudescence de ces maladies, notamment vis-à-vis de
l'Esca, étant donné l'absence de méthodes de lutte
alternative efficace actuellement disponible (Larignon, 2009).
2 Identifiée en France à la fin des
années 70, l'Eutypiose est la maladie la mieux connue mais reste encore
difficile à combattre en dépit des travaux réalisés
par de nombreuses équipes portant sur différents thèmes :
épidémiologie, relation hôte-parasite, méthodes de
lutte (Larignon, 2004-a). On note que BIVB a restreint le champ de cette
étude aux maladies de l'Esca et du BDA.
1.2 Eléments historiques
1.2.1 Historique de l'Esca
L'Esca est une maladie cryptogamique connue depuis très
longtemps. Les écrits de la période gréco-romaine en
faisaient déjà mention (Dubos, 2002). Son origine a longtemps
été indéterminée et attribuée à un
accident d'ordre physiologique, désigné sous le nom de folletage.
(Rolland, 1873 ; Ravaz, 1898). En 1905-1906, Ravaz semble être le premier
à avoir réalisé, qu'une maladie possédant les
mêmes caractéristiques que le folletage était
déjà connue (Surico, 2009). Celle-ci était
désignée auparavant sous le nom de << iska >> en
Grèce (Pavlou, 1906). Ce n'est qu'à la fin du
XIXème siècle que cette forme fut attribuée
à un champignon et de ce fait distinguée du folletage (Larignon,
2005) En 1922, le terme << Esca >> a été introduit
par Viala pour désigner cette maladie (Larignon, 2004-b).
1.2.2 Historique du BDA
Ce dépérissement est encore peu connu en raison
de sa récente identification, et peu d'équipes travaillent
actuellement sur cette maladie (Larignon, 2004-a). Le terme << Black Dead
Arm >> a été introduit par Lehoczky en 1974 pour
désigner la maladie dans le vignoble de Tokaj en Hongrie. L'expression a
ensuite été reprise par Cristinzio (1978) et Rovesti et
Montermini (1987) lors de leurs études respectives en Italie. La maladie
a été identifiée en France en 1999 dans le vignoble du
Médoc (Larignon et al., 2000). Elle s'est étendue
rapidement aux autres régions viticoles françaises, et il semble
qu'elle soit présente dans les vignobles des zones à climats
méditerranéens et tempérés. Le BDA a souvent
été confondu avec l'Esca en raison d'une symptomatologie
convergente au niveau du feuillage. Son émergence est probablement
liée au respect de l'interdiction de l'arsénite de sodium
prononcée en 2001 (Dubos, 2002).
1.3 Symptomatologie
1.3.1 Caractéristiques
générales et différences entre les deux
maladies
Selon Larignon (2007), l'Esca et le BDA sont des syndromes qui
présentent une grande similitude de symptômes foliaires,
observables en général à partir de juin pour le BDA, et
juillet pour l'Esca. Il existe une très grande variabilité
d'expression de ces maladies. De façon générale, les
symptômes les plus visibles sont caractérisés par des
colorations du feuillage et des dessèchements (formes lentes : photos
2-3 et 8-9), alors que les symptômes les plus graves conduisent à
d'importants flétrissements (formes foudroyantes : photos 1 et 7).
D'après les travaux de Larignon et Dubos (2001-a), on
peut distinguer l'Esca et le BDA en étudiant sur les feuilles le
détail des expressions symptomatologiques de leurs formes lentes
respectives. Pour les symptômes d'Esca sur cépage noir, les
digitations rouges observées entre les nervures sont bordées d'un
liseré jaune alors que celui-ci est absent dans le cas du BDA (photos 2
et 8). On note également que les symptômes de BDA sont d'un rouge
vineux alors que ceux de l'Esca sont plus clairs. Sur cépage blanc,
seule la couleur des digitations permet de différencier les deux
maladies (absence du liséré jaune caractéristique de
l'Esca autour des digitations). Les digitations sont d'un jaune plutôt
blanc pour l'Esca alors que le jaune est plus vif pour le BDA (photo 3 et 9).
Il est également possible d'observer les deux faciès sur la
même plante, sur le même rameau, voire sur la même feuille
(photo 4).
Un suivi des feuilles tout au long de la période
végétative a montré que les tissus de couleur rouge vineux
(typique du BDA) s'éclaircissaient pour devenir rouge clair, puis
présentaient à leur

1988 1989 1990 1991 1992 1993
A nnées d'observation

60
50
Souches avec symptOmes (%)
40
30
20
10
0
30,4
21,5
30,8
26,8
51
14,1
Photo 1 : Forme sévère foudroyante de
l'Esca (apoplexie)
Source : Larignon (2009) -
Figure 2 : Taux de souches présentant des
symptômes de la forme lente de l'Esca en septembre Document
non publié
(Etude menée en Charentes de 1988
à 1993 sur des parcelles d'Ugni blanc non traitées)
Source : Desaché etal. (1994)

Photo 2 : Evolution de l'expression symptomatologique de
l'Esca Photo 3 : Evolution de l'expression symptomatologique de
l'Esca
sur cépage noir (forme lente) sur cépage
blanc (forme lente)
Source : Larignon (2009) - Document non publié Source
: Larignon (2009) - Document non publié

(I)
(III)
(V)
Photo 4 : Double faciès Esca / BDA sur feuille
Source : Larignon (2009) - Document non publié
Symptômes d'Esca
(forme lente)
Symptômes de BDA
(forme lente)
Photo 5 : Symptômes d'Esca sur fruit Photo 6 :
Symptômes d'Esca sur bois
Source : Larignon (2009) - Document non publié
(I: nécrose caractéristique ; II: stade précoce ; III :
nécrose sectorielle)
Source : Larignon (2009) - Document non publié
périphérie des jaunissements (Larignon et Dubos,
2001-a). Cette observation suggère à Lecomte et al.,
(2006-a) que le faciès BDA peut être une phase transitoire de
l'expression de la forme lente de l'Esca. Des études sur le profil
anthocyanique de feuilles atteintes par le BDA ou par l'Esca, à des
stades précoces d'expression foliaire, montrent qu'ils sont
différents et signes de dérèglements spécifiques du
métabolisme foliaire (Larignon et al., 2003).
1.3.2 Symptomatologie de l'Esca
D'après Larignon (2009), l'Esca est
généralement observé sur des vignes âgées
d'au moins 8 ans et les symptômes sont très fluctuants d'une
année sur l'autre. En effet, au vignoble, un cep infecté une
année peut très bien se comporter de façon asymptomatique
l'année suivante (Dubos, 2002). La figure 2 illustre cette
variabilité interannuelle dans l'expression de la maladie. Les premiers
symptômes apparaissent, selon les régions, de fin juin à
mi-juillet. Ils se manifestent régulièrement durant toute la
période végétative et touchent soit toute la plante, soit
un seul bras, soit un ou quelques rameaux.
Cette maladie se présente dans le vignoble sous deux
formes. La forme foudroyante (ou « apoplectique ») est
caractérisée par un dessèchement brutal et total de la
végétation -fruits, feuilles et rameaux- (photo 1). La forme
lente se manifeste selon deux modalités, suivant la couleur du
cépage impliqué (photos 2 et 3). Sur cépage noir, on
observe à la surface du limbe de petites taches jaunâtres, qui
vont rougir et prendre une teinte rouge clair, puis griller. Sur cépage
blanc, la forme lente se manifestera sous la forme de petites taches de couleur
jaune, plutôt blanche, à la surface du limbe. Ces taches vont
s'agrandir, fusionner et griller, pour ne laisser qu'une bande verte le long
des nervures principales. Sur les fruits, un retard dans la maturation des
raisins est observé, il peut s'accompagner d'un flétrissement et
d'un dessèchement (photo 5) (Larignon, 2009). Ces symptômes sont
en association ou non avec les symptômes foliaires (Chiarappa, 1959).
Au niveau du bois, la lésion caractéristique de
l'Esca est une nécrose claire et tendre en position centrale qui est
constituée de plusieurs zones (photo 6-I). Au centre, on trouve un bois
clair et friable (A), plus connu sous le nom d'amadou ou pourriture blanche.
Cette zone est délimitée par un liseré noir (B). Entre le
bois sain et le liseré, on observe une nouvelle zone de couleur
brun-rose, de faible épaisseur (C). Cette nécrose
caractéristique est précédée par un autre type de
symptôme : une nécrose brune et dure en position centrale (photo
6-II). Il est parfois également possible d'observer une nécrose
sectorielle de couleur brune -caractéristique de l'Eutypiose- (photo
6-III). Ces différents types de lésion peuvent tout à fait
apparaitre sur un même cep. On peut également observer des
ponctuations noires (correspondant à des vaisseaux
nécrosés) (Larignon et Dubos, 1997).
1.3.3 Symptomatologie du BDA
Le BDA peut être observé sur des vignes
âgées d'au moins trois ans. Les symptômes apparaissent
tôt en saison, de fin mai à mi-juillet selon les régions
viticoles, et se manifestent de façon régulière tout au
long de la période végétative. Ils peuvent toucher soit
toute le plante, soit un seul bras ou encore un ou plusieurs rameaux (Larignon,
2009).
Il existe deux formes de cette maladie : une forme
sévère -apoplexie- (photo 7) et une forme lente, qui conduisent
à une chute prématurée des feuilles. La forme
sévère (défoliation brutale) est différente de la
forme apoplectique de l'Esca. Il peut subsister quelques feuilles à
l'extrémité des rameaux (Larignon et Dubos, 2001-a). La forme
lente se manifeste selon deux modalités, suivant la couleur du
cépage impliqué (photo 8 et photo 9). Sur cépages noirs,
on observe des petites taches d'un rouge vineux à la surface des limbes
des feuilles. Ces taches vont s'agrandir, fusionner, puis prendre


P
(
P
S
|
Photo 14 : Nécrose sectorielle du BDA
Source : Larignon (2009) - Document non publié
|
|
Photo 11 : Symptômes du BDA sur inflorescence et
fruit Photo 13 : Coupe transversale au niveau de la bande brune
Source : Larignon (2009) - Document non publié Source
: Larignon (2009) - Document non publié
une teinte feuille morte en ne laissant qu'une bande verte le
long des nervures principales (Larignon, 2009). Sur cépages blancs, On
observe tout d'abord la formation de taches jaunes de couleur vive, qui vont
griller. Ces taches vont s'agrandir, fusionner pour ne laisser qu'une bande
verte le long des nervures principales. On note que d'autres faciès
peuvent également être aperçus (photo 10) : on peut ainsi
observer une perte de la turgescence de la feuille, qui va conduire à la
formation de nécroses plutôt orange, ne laissant progressivement
qu'une bande verte le long des nervures principales. A un stade très
évolué, lorsque les feuilles ne sont pas tombées, les
derniers tissus qui restaient verts commencent à jaunir. Selon la
gravité de la maladie et la période à laquelle elle se
manifeste, elle peut toucher les inflorescences ou les fruits (photo 11),
conduisant à leur dessèchement (Larignon, 2009).
Au niveau du bois, à partir d'un rameau malade,
l'enlèvement de l'écorce (qui se détache facilement
à la main) révèle la présence d'une bande brune qui
part du rameau malade et qui descend jusqu'au niveau de la soudure, voire dans
le porte-greffe (photo 12). Cette bande brune peut se former de part et d'autre
d'une nécrose sectorielle et se traduit ensuite soit par la
présence de taches noires, soit par la formation d'un
chancre3 (Larignon et Dubos, 2001-a). En coupe transversale, on
observe la bande brune (photo 13). Si on agrandit cette zone, on observe
à côté de la bande brune, un secteur dans lequel les
vaisseaux du bois sont obstrués par une substance jaunâtre. La
quantité de vaisseaux obstrués est vraisemblablement responsable
de la forme plus ou moins sévère de la maladie. On peut
également observer des nécroses sectorielles (photo 14). Elles
peuvent être confondues avec la nécrose de l'Eutypiose, mais il
existe toutefois quelques différences, notamment en ce qui concerne la
couleur. Pour le BDA, la nécrose est un peu plus grise et ne
présente pas de stries brunes (Larignon, 2009).
1.4 Agents pathogènes impliqués
1.4.1 Agents fongiques associés à la
maladie de l'Esca
Selon Larignon (2004-a), les champignons impliqués dans
le syndrome de l'Esca se répartissent en deux séquences dans le
processus menant à la dégradation du bois. Une colonisation
primaire du bois par Phaeomoniella chlamydospora (Pch) et
Phaeoacremonium aleophilum (Pal) semble être une étape
nécessaire pour l'installation secondaire de Fomitiporia
mediterranea (Fm), responsable de la dégradation du bois
caractéristique de l'Esca (pourriture blanche ou « amadou »).
Un autre champignon, Eutypa lata4 (El), est
également considéré comme pionnier dans ce processus.
Viret (2004) ajoute qu'un relevé mycologique précis sur des ceps
atteints d'apoplexie a permis d'isoler plus de 60 champignons
différents, dont Pch, Pal, Fm et El, mais également Phomopsis
viticola, Botryosphaeria obtusa, et un grand nombre d'autres
champignons saprophytes ou endophytes de la vigne. Selon cet auteur, la
biologie et l'épidémiologie des différents champignons
liés à l'Esca restent lacunaires et sont difficiles à
étudier.
1.4.2 Agents fongiques associés à la
maladie du BDA
Concernant le Black Dead Arm, plusieurs espèces de la
famille des Botryosphaeriaceae semblent être associées
(Larignon, 2004-b). On considère que les principaux champignons
associés au BDA sont les suivants : Botryosphaeria obtusa et
Neofusicoccum parvum (Larignon, 2009).
3 Nécrose de l'écorce qui s'accompagne souvent de
crevasses, de boursouflures, de bourrelets et d'autres déformations, et
qui contamine le bois sous-jacent.
4 Agent fongique par ailleurs responsable de la maladie de
l'Eutypiose.

Figure 3 : Premier processus de dégradation du
bois (Esca)
Source : Larignon (2009)

Figure 4 : Second processus de dégradation du
bois (Esca)
Source : Larignon (2009)

Figure 5 : Combinaison des deux processus de
dégradation du bois (Esca)
Source : Larignon
(2009)
1.5 Processus infectieux
1.5.1 Mécanismes menant à la
dégradation du bois
> Esca
Il est certain que les infections ne sont possibles que par
les plaies de taille ou autres blessures (Viret, 2004). Concernant l'Esca, deux
processus de dégradation du bois menant à la formation de la
nécrose caractéristique ont été décrits
(Larignon et Dubos, 1997 ; Larignon, 1991) Le premier processus aboutit
à la formation d'une nécrose claire et tendre en position
centrale et fait intervenir trois champignons : Phaeomoniella
chlamydospora (Pch), Phaeoacremonium aleophilum (Pal) et
Fomitiporia mediterranea (Fm) (Figure 3). Dans un premier temps, de
petites ponctuations noires vont apparaître, dans lesquelles on retrouve
Phaeomoniella chlamydospora. Ensuite, il y a formation d'une petite
nécrose à partir de la moelle, de couleur rose, due à
Phaeoacremonium aleophilum. Cette nécrose va se
développer et se mélanger avec les ponctuations noires, pour
former la nécrose brune et dure en position centrale. Ce bois est
ensuite colonisé par Fomitiporia mediterranea, pour former une
nécrose claire et tendre, qui va progressivement s'étendre. Dans
ce cas, on observe des symptômes de la forme lente de l'Esca, quel que
soit le stade de ces nécroses dans le cep (Larignon, 2004-c ; Larignon
2009).
Le second processus (Figure 4) conduit à la formation
d'une nécrose claire et tendre en position sectorielle et implique
Eutypa lata (El), responsable de la nécrose brune en position
sectorielle, et Fomitiporia mediterranea qui dégrade ce bois
brun en une pourriture blanche et tendre. Cette nécrose peut
évoluer jusqu'à occuper la quasi-totalité du bois. A un
stade avancé, des symptômes d'apoplexie pourront être
observés car il ne reste alors que très peu de bois fonctionnel.
On note que les deux processus peuvent être observés sur la
même plante (Figure 5) (Larignon et Dubos, 1997 ; Larignon, 2009).
Mugnai et al., (1999) ont également
schématisé la colonisation du bois d'un tronc de vigne par les
champignons de l'Esca (figure 6). Sur cette représentation, trois
espèces fongiques ont été isolées et
quantifiées pour chaque portion du tronc : Phaeomoniella
chlamydospora (Pch), Phaeoacremonium aleophilum (Pal) et
Fomitiporia punctata (Fp)5. Ce dernier est majoritaire dans
le bois pourri et sa présence diminue vers le bois sain, alors que Pch
et Pal deviennent dominants. Ceux-ci se trouvent principalement dans les stries
noires et dans le bois brun-rouge, dans les parties du bois autour de la moelle
et en marge des tissus du bois carié.
> BDA
Le processus de dégradation du bois n'a pas encore
été décrit pour le BDA (Larignon et al., 2009).
Au niveau du bois, le champignon n'est pas isolé sur toute la longueur
de la bande brune caractéristique. On ne sait pas si ces champignons
sont à l'origine de cette bande brune ou s'ils colonisent cette
nécrose (Larignon, 2004-b).
1.5.2 Mécanismes menant à l'expression
des symptômes foliaires
Ces mécanismes sont méconnus car il est
notamment difficile de reproduire les symptômes foliaires de l'Esca et du
BDA par simple inoculation artificielle des champignons en question
(Guérin, 2004). Concernant l'Esca, l'expression des symptômes au
niveau des feuilles et des baies n'est pas reliée à la
présence de mycélium des champignons dans le rameau :
l'hypothèse qu'ils produisent des molécules agissant à
distance (toxines) a donc été émise (Mugnai et
al., 1999). Certaines d'entre elles
5 Fomitiporia Mediterranea a été
distinguée en tant que nouvelle espèce distincte de
Fomitiporia punctata. (Fisher, 2002)

Pch : 33% Pal : 44% Fp : 11%
Pch : 27% Pal : 11% Fp : 44%
Pch : 46% Pal : 10% Fp : 1%
Pch : 7% Pal : 12% Fp : 14%
Pch : 83% Pal : 9% Fp : 1%
Pch : 18% Pal : 54% Fp : 0%
Pch : 2% Pal : 8% Fp : 62%
Figure 6 : Schéma de la colonisation du bois d'un
tronc de vigne par les champignons de l'Esca (Coupes transversales
à gauche, coupe longitudinale à droite)
(Pch : Phaeomoniella chlamydospora ; Pal : Phaeoacremonium
aleophilum ; Fp : Fomitiporia punctata)

- : Plaies de taille
- BN : Brown Necrosis (nécrose brune)
- BRW : Brown-Red Wood (bois brun-rouge)
- AP : Altered Pith (moelle
altérée)
Source : Mugnai etal. (1999)
- BS : Black Streaks (stries noires)
- DP : Decayed Pith (moelle cariée)
- BL : Black Line (liséré noir)
- DW : Decayed wood (Bois carié)
ont été identifiées. Fkyera et al.,
(2000) ont décrit de nombreuses molécules chez
Phaeomoniella chlamydospora (Pch) dont l'isosclérone et la
scytalone. Ces deux dernières molécules sont également
produites par Phaeoacremonium aleophilum (Pal). Le fait qu'elles
soient produites à la fois par Pch et par Pal justifie qu'elles aient
été très étudiées quant à leur
implication dans l'expression des symptômes foliaires (Bruno et al.,
2007). Par comparaison, Fomitiporia mediterranea produit peu de
molécules dont les deux principales sont la 6-methoxymelleine et le
2,2-dimethyl-4-oxochoman-6- carbaldehyde (Fkyera et al., 2000). La
fonction de ces différentes molécules n'est pas clairement
déterminée.
1.6 Interactions hôte-pathogène
Au-delà de l'identification et de la connaissance des
différents bioagresseurs impliqués, on sait que les maladies de
dépérissement font intervenir l'aptitude de la plante à
répondre à l'agression (Goutouly, 2010). A propos de l'Esca,
Magnin-Robert et al., (2010) expliquent que la proposition de nouveaux
moyens de lutte ne pourra se faire que suite à une compréhension
plus fine des mécanismes mis en oeuvre, passant notamment par une
reproduction des symptômes foliaires et une caractérisation de son
impact sur la physiologie de la plante.
1.6.1 Impact de l'Esca sur la vigne
Depuis quelques années, des études
s'intéressent à l'évolution de la physiologie de la plante
au cours du développement de la maladie, notamment vis-à-vis du
métabolisme carboné. Celui-ci a été choisi car son
bon fonctionnement est vital pour la pérennité de la plante : les
perturbations précoces sont donc de bons indicateurs de stress
(Magnin-Robert et al., 2010).
> Forme lente
En 2006, Petit et al., ont étudié
l'altération de l'appareil photosynthétique en fonction de
l'intensité de l'expression des symptômes foliaires de la forme
lente de l'Esca. Cette altération se traduit par une diminution de
l'activité photosynthétique et de la transpiration, une
perturbation du fonctionnement des photosystèmes et une diminution de la
concentration en pigments chlorophylliens. La conséquence directe de
cette diminution de la photosynthèse est une chute des réserves
carbonées dans les sarments. L'année suivante, les faibles
réserves peuvent entraîner une baisse de vigueur de la plante,
affectant son développement végétatif et reproducteur. En
complément de ces travaux, Christen (2006) a observé que cette
modification de la photosynthèse pouvait être
détectée 2 mois avant l'expression des symptômes foliaires,
chez le Cabernet-Sauvignon.
> Forme apoplectique
Selon Letousey et al., (2010), l'étude de la
forme foudroyante de l'Esca montre que les mécanismes
photosynthétiques sont fortement perturbés 7 jours avant que les
feuilles ne deviennent apoplectiques (dessèchement). Visuellement, aucun
signe ne permet de dire que la plante va prochainement exprimer des
symptômes foliaires. Cette altération est détectable au
niveau physiologique avec une chute brutale de l'assimilation du CO2, une
fermeture des stomates, une baisse de l'activité du photosystème
II et une diminution de la transcription de certains gènes
associés à la photosynthèse. A l'inverse, certains
gènes de défense sont surexprimés dans les feuilles 7
jours avant et pendant l'expression visibles des symptômes sur la plante.
La plante perçoit donc un stress, avant même l'expression visible
de la maladie, et active une réponse de type défense. Cette
perception est maximale juste un jour avant le dessèchement total des
feuilles.
Comme on associe souvent la forme apoplectique à un
excès d'eau dans le sol avec des températures
élevées (Surico et al., 2006), l'altération du
transport hydrique dans la plante liée à des dysfonctionnements
du xylème a également été étudiée.
Ainsi, Edwards et al., (2006) ont observé une diminution de la
conductance stomatique entraînant un déficit hydrique chez des
plantes infectées par

Figure 7 : Les quatre types de barrières de
compartimentation du modèle CODIT décrits par Shigo et Marx en
1977 (1 : développement de thylles et
sécrétion de substances gommeuses dans les vaisseaux du
xylème ; 2 : caractères morphologiques des dernières
cellules en fin de chaque anneau de croissance ; 3 : réactions de
défense de la plante impliquant les rayons parenchymateux ; 4 :
formation d'une zone de barrage par le cambium)
Source : Rioux (2011)

Faisceaux conducteurs
Phloème secondaire (liber)
Vaisseau du bois
Xylème secondaire (bois)
Parenchyme cortical
Écorce
Sclérenchyme
Assise subéro-phellodermique
Assise libéro-ligneuse (cambium)
Parenchyme de rayon ligneux
Parenchyme médullaire (moelle)
Photo 15 : Coupe transversale d'un rameau principal de
vigne, sain et aoûté (sarment) Source : -
Crédit photo: Trouvelot (Communication personnelle)
- Légende : Dufouleur (2011)
Phaeomoniella chlamydospora. De plus, une
étude comparative de la fluorescence chlorophyllienne d'une plante
exprimant la maladie (forme lente ou forme foudroyante non
précisée) avec celle subissant un stress hydrique a
révélé deux fonctionnements différents du
photosystème II (Christen, 2006). Il a donc été conclu que
l'expression des symptômes de l'Esca ne pouvait pas simplement être
corrélée à un stress hydrique, mais qu'elle était
plus complexe.
1.6.2 Impact du BDA sur la vigne
Aucune étude spécifique caractérisant
l'impact du BDA sur différents métabolismes de la plante
(métabolisme carboné, hydrique ou encore la mise en place de
défenses) n'a été publiée (Larignon et
al., 2009).
1.6.3 Adaptation physiologique de la plante face
à l'agression
En réponse à une agression par des
pathogènes (ou suite à une blessure), la plante met
généralement en place un mécanisme de défense
complexe afin de limiter leur progression. Celui-ci consiste en une
série de modifications anatomiques se traduisant par la formation de
compartiments étanches qui isolent la zone atteinte (Shigo et Marx,
1977). Dans leur modèle CODIT (Compartmentalization Of Decay In Trees),
ces auteurs décrivent quatre types de barrières (ou « murs
») dont les trois premières servent à restreindre la
progression des microorganismes dans le bois présent au moment de la
blessure (figure 7). La formation de la première barrière
correspond au développement de thylles et à la
sécrétion de substances gommeuses dans les vaisseaux du
xylème. Ce premier mur est discontinu et constitue la plus faible des
barrières limitant la propagation verticale des microorganismes. La
seconde barrière est attribuée aux caractères
morphologiques des dernières cellules en fin de chaque anneau de
croissance. Ce mur est continu sur toute la longueur de l'arbre et limite
l'expansion parallèle aux rayons des microorganismes (expansion
radiale). La troisième barrière implique les rayons
parenchymateux, tissus vivants du bois, qui peuvent déclencher des
réactions de défense de la plante. Il s'agit de la plus forte
barrière des trois que nous avons déjà citées, mais
elle peut être discontinue. Elle limite la progression des
microorganismes perpendiculaire aux rayons (expansion tangentielle). La
quatrième barrière, la plus forte et la plus importante, consiste
en une zone de barrage formée par le cambium après la blessure.
Ce mur isole le bois attaqué de celui qui sera formé par la suite
et empêche la progression des champignons. La formation de ces
barrières demande beaucoup d'énergie à la plante. La
multiplication des zones lésées entrave fortement le flux de
sève et peut entraîner un affaiblissement
généralisé de la plante, qui devient alors incapable de se
défendre efficacement contre les champignons pathogènes.
Afin de mieux visualiser les processus en jeu et les tissus
concernés, on pourra également se reporter à la photo 15.
Il s'agit d'une photographie d'une coupe transversale d'un rameau principal de
vigne, sain et aoûté. On peut y découvrir l'organisation du
système vasculaire dit « secondaire » (car
élaboré à partir d'un méristème secondaire,
le cambium). Les faisceaux conducteurs sont constitués de xylème
(ou « bois », impliqué dans le transport de la sève
brute) et de phloème (ou « liber », véhiculant la
sève élaborée). On note la taille importante des vaisseaux
du bois. Les différents parenchymes ont généralement un
rôle de réserve. Le sclérenchyme (tissu de soutien) et
l'assise subéro-phellodermique (exclusivement «
subérogène » dans le rameau, à la différence
de la racine) ont été notés à titre indicatif.
1.6.4 Complication pathologique : cavitation,
embolie et thyllose
On sait que les maladies du bois entrainent la formation de
nécroses qui altèrent le fonctionnement vasculaire de la plante,
en diminuant la quantité de xylème fonctionnel. Quand on connait
les mécanismes de la cavitation, de l'embolie et de la thyllose, on
comprend qu'une plante
infectée par l'Esca ou le BDA va être encore plus
sensible à ces accidents. Ceux-ci sont décrits ciaprès.
D'après Carbonneau et al. (2007), le flux de
sève brute est, d'une façon générale,
extrêmement sensible aux variations brusques de la pression
intravasculaire. Ils expliquent que ces variations de pression peuvent
être dues à une forte augmentation de la demande hydrique
atmosphérique et qu'elles peuvent provoquer l'apparition de bulles d'air
dans les vaisseaux du xylème (phénomène de cavitation). La
rupture du courant de sève par ces bulles peut ainsi complètement
bloquer la circulation (embolie). Sous certaines conditions
atmosphériques (température élevée et faible
hygrométrie), Adrian et Fournioux (2011) indiquent qu'un accident encore
plus grave peut survenir. L'évapotranspiration très importante au
niveau du feuillage peut provoquer une telle dépression dans le
système circulatoire qu'elle mène à la formation de
thylles6. L'issue de cet accident est fatale si elle concerne un
nombre trop important de vaisseaux (fanaison très rapide de tout le
feuillage avec mort du cep en quelques heures, ou « apoplexie »).
On comprend donc que, sous certaines conditions climatiques
critiques, une plante qui dispose d'un nombre limité de vaisseaux
fonctionnels (nécroses associées aux maladies du bois) pourra
moins bien s'adapter à son environnement. Le fonctionnement vasculaire
sera moins « souple » (baisse de la conductivité hydraulique
de la plante) et on observera davantage d'à-coups en cas d'augmentation
subite de la demande évaporatoire. Ce mode de fonctionnement
prédispose la plante aux accidents climatiques
précédemment décrits (embolie et thyllose).
1.7 Etiologie : étude des facteurs associés
à la condition pathologique
Les maladies de dépérissements impliquent des
pathosystèmes complexes pour lesquels il est nécessaire d'avoir
une approche globale de la plante dans son environnement. Cette
complexité est liée au développement systémique des
champignons dans les tissus du bois, à la similitude existant entre les
symptômes de l'Esca et du BDA et à leur non-permanence, ainsi
qu'à la difficulté d'identifier et de hiérarchiser les
facteurs biotiques et abiotiques impliqués dans ces maladies
(Guérin, 2004).
1.7.1 Facteurs étudiés pour les deux maladies
indistinctement (Esca/BDA)
> L'âge
Selon Morvan (2009), l'âge des ceps infectés
semble être le principal paramètre influençant l'expression
des symptômes. S'appuyant sur une étude de l'observatoire national
des maladies du bois, cet auteur constate que l'âge optimal d'expression
se situerait entre 15 et 20 ans. Il indique également que les
observations faites sur le réseau de parcelles de
référence du BIVB tendent à confirmer cette information,
avec toutefois un léger décalage : plutôt 20-25 ans. Il
conclut qu'il est hasardeux de vouloir comparer des parcelles qui n'ont pas le
même âge, car les différences d'âge sont susceptibles
de cacher l'influence des autres paramètres. D'après Larignon
(2009), l'Esca est généralement observé sur des vignes
âgées d'au moins 8 ans alors que le BDA peut être
observé sur des vignes âgées d'au moins trois ans. A propos
de l'Esca, Dubos (2002) note que l'apparition des symptômes a lieu sur
des vignes de plus en plus jeunes (dès l'âge de 4 ans).
6 Selon Adrian et Fournioux (2011), la thyllose consiste en la
formation de thylles, qui sont des invaginations de la membrane plasmique des
cellules bordant les vaisseaux du bois (parenchyme périvasculaire) dans
la lumière de ces derniers. Ces vaisseaux sont ainsi «
condamnés ». On note que l'origine de la thyllose peut être
naturelle (obstruction des vaisseaux les plus anciens) ou accidentelle
(accident climatique).

15%
14%
12%
11%
9%
8%
0%
6%
5%
3%
2%
(712 parcelles de référence)
(15 à 40 parcelles par cépages)
(300 ceps observés par parcelle)
Moyenne nationale = 4,84%
Figure 8 : Taux d'expression global des symptômes
des maladies de l'Esca et du BDA selon le cépage
Source
: DGAL - SDQPV (2007) - Document non publié

2002 2003 2004 2005 2006 2007
(300 ceps observés par parcelle)

RVATI ON
PAS D'OBSE
Côte d'Or
Chardonnay
(29 parcelles)
Côte d'Or
Pinot Noir
(40 parcelles)
2,5% 2,0% 1,5% 1,0% 0,5% 0,0%
Figure 9 : Suivi des taux d'expression global de l'Esca
et du BDA en Côte-d'Or
Source : DRAF - SRPV Bourgogne (2007) - Document non
publié
> La sensibilité variétale
D'après l'observatoire national des maladies du bois,
4,7 % des ceps (0,2 à 20 % selon les cépages) expriment un ou
plusieurs symptômes de maladies du bois (Herlemont, 2004-a). Selon la
même étude, les cépages semblant les plus sensibles
à l'Esca et au BDA sont le Sauvignon, le Cabernet Sauvignon, le Chenin,
l'Ugni blanc et l'Auxerrois. Une étude épidémiologique
(non publiée) de la DGAL (Direction Générale de
l'Alimentation) et de la SDQPV (Sous-Direction de la Qualité et de la
Protection des Végétaux) menée au niveau national en 2007
montre que le taux moyen d'expression des symptômes observé sur le
réseau de parcelles de références (712 unités) est
de 4,84% (figure 8). On note qu'il existe d'importantes variations selon le
cépage concerné, avec un taux d'expression minimum de 0,46%
(Pinot Noir) et un taux maximum à 15,11% (Trousseau). En Bourgogne,
d'après l'étude (non publiée) menée par la DRAF
(Direction Régionale de l'Alimentation, de l'Agriculture et de la
Forêt) et le SRPV (Services Régional de la Protection des
Végétaux) en 2007, le Pinot Noir et le Chardonnay de
Côte-d'Or comportent des taux d'expression relativement faibles et la
situation semble stable (figure 9). Le Chardonnay de l'Yonne, quant à
lui, présente un taux d'expression de symptômes foliaires plus
important, proche de la moyenne nationale (figure 8).
> Le rapport Carbone/Azote dans les tissus
végétaux
Selon Goutouly (2010), ce rapport interagit fortement avec la
propagation des pathogènes, en orientant le métabolisme de la
plante. En effet, un C/N faible (nutrition azotée forte) favorise le
métabolisme de croissance. A l'inverse, un C/N élevé
favorise le métabolisme secondaire avec la production de composés
phénoliques. Ces conditions apparaissent lors de stress oxydatifs comme
les stress hydrique, azoté ou thermique. Les voies de biosynthèse
de ces composés sont communes à celles que la vigne met en place
pour limiter la progression de pathogène dans le bois
(biosynthèse de molécules de défense). Selon ce
raisonnement, les années aux conditions climatiques favorables à
la croissance conduiraient à une baisse des capacités de
résistances de la vigne vis-à-vis des maladies de
dépérissement. Ceci se traduirait alors par une augmentation des
symptômes foliaires, le complexe de pathogènes rencontrant moins
de résistance de la part de la plante. Inversement, les années
défavorables à la croissance favoriseraient la résistance
et entraîneraient une baisse de l'expression foliaire. Ce fonctionnement
écophysiologique pourrait expliquer, en partie du moins, les
fluctuations interannuelles des symptômes.
> Le sol
D'après Goutouly et al. (2007), les parcelles
à forte réserve utile, dont le sol est caractérisé
par une alimentation en eau non limitante, seraient plus atteintes par les
maladies du bois.
1.7.2 Facteurs biotiques (Esca)
> L'âge
D'après Larignon (2009), l'Esca est
généralement observé sur des vignes âgées
d'au moins 8 ans. Toutefois, Dubos (2002) note que l'apparition des
symptômes a lieu sur des vignes de plus en plus jeunes (dès
l'âge de 4 ans).
> La sensibilité variétale
Les enquêtes réalisées dans le vignoble
ont permis de classer les cépages selon la sensibilité à
l'expression des symptômes foliaires de l'Esca (Grosmann, 2008). Ainsi,
des cépages tels que le Cabernet Sauvignon, le Sauvignon blanc, le
Mourvèdre, l'Ugni blanc, le Cinsault ou le Trousseau, expriment plus
facilement les symptômes de la forme lente que le Merlot, le Carignan, la
Roussane ou le Pinot noir. Larignon et al., (2009) soulignent
qu'aucune étude n'a été réalisée pour
classer les cépages selon l'importance des nécroses dans le bois
et qu'aucune donnée n'est disponible sur les
espèces autres que Vitis. Ils indiquent
également que les vignes mères de porte-greffe ne
présentent pas de symptômes foliaires sur la partie
herbacée, et que seules des nécroses caractéristiques de
l'Esca y sont observées dans le bois.
> Les pratiques culturales (interventions humaines)
Selon Dubos (2002), d'une façon générale,
toutes les opérations entrainant des blessures (suppression des bras
mort, modification du mode de conduite pour la mise en oeuvre de vendanges
mécaniques, etc.) sont favorables à la maladie. Certains
systèmes de taille, notamment la taille guyotpoussard, semble limiter
l'Esca (Geoffrion et Renaudin, 2002). Toutefois, les premières
observations de Dumot et Ménard (2005) sur l'Esca montrent l'absence
d'effet du mode de conduite, de la fertilisation et de la protection des plaies
de taille. Selon ces mêmes auteurs, cette maladie semble
indépendante des différences engendrées par les
modalités, que ce soit sur les plaies de taille (surface des plaies,
protection, date de taille) ou sur la vigueur.
1.7.3 Facteurs abiotiques (Esca)
Bien que le complexe de champignons associés au
dépérissement soit présent de façon très
générale au vignoble, l'expression des symptômes est
pourtant variable en fonction du contexte écophysiologique (facteurs
climatiques et édaphiques), matérialisant un effet parcelle et
même un effet année (Goutouly, 2010). Selon cet auteur, la
progression du complexe de pathogènes dans le bois accentuerait la
vulnérabilité de la plante, la rendant plus sensible aux
variations de l'environnement.
D'après Surico et al., (2000), les
étés doux et pluvieux semblent favorables à l'expression
des symptômes de la forme lente alors que les étés chauds
induiraient davantage l'apoplexie : cette dernière est observée
selon certaines conditions climatiques, notamment lors de périodes de
sécheresse ou de vent fort. Elle est due au fait que
l'évapotranspiration trop importante n'est plus compensée par un
afflux en eau suffisant car il ne reste qu'une faible quantité de bois
fonctionnel. Une étude des bilans hydriques réalisée
pendant trois années consécutives (2004 à 2006) dans le
vignoble bordelais a montré que le stress hydrique de la vigne
contribuerait à inhiber l'expression foliaire (Goutouly et al.,
2007). Toutefois, Christen (2006) montre que l'expression des
symptômes de l'Esca ne peut pas simplement être
corrélée à un stress hydrique (voir paragraphe 1.5.1 :
Impact de l'Esca sur la vigne - Forme apoplectique).
1.7.4 Facteurs biotiques (BDA)
> L'âge
D'après Larignon (2009), le BDA peut
généralement être observé sur des vignes
âgées d'au moins trois ans.
> La sensibilité variétale
Il est connu que les cépages Cabernet Sauvignon, Cabernet
franc, Sauvignon blanc sont plus sensibles à ce
dépérissement que le Merlot (Larignon et Dubos, 2001-b).
1.7.5 Facteurs abiotiques (BDA)
Peu d'informations spécifiques sont disponibles sur les
facteurs environnementaux ayant un rôle majeur dans le
développement de la maladie et de son expression. Selon Larignon et
Vigues (2010) certains facteurs climatiques interviennent sur l'expression des
symptômes, mais leurs rôles ne sont pas connus. Ces auteurs pensent
que la connaissance du climat (pluie, évapotranspiration et
températures maximales pour l'expression des premiers symptômes)
pourrait aider à prédire le pourcentage annuel d'expression.
1.8 Méthodes de lutte
1.8.1 Eléments d'information sur
l'arsénite de sodium
Du fait de la toxicité avérée de ce
produit, son emploi est interdit en France depuis le 8 novembre 2001 (avis au
J.O.R.F. du 23 novembre 2001). Les dérivés minéraux de
l'arsenic sont employés depuis très longtemps comme insecticide,
fongicide et défoliant en agriculture. Ces produits sont des agents
cancérogènes reconnus (Harmandon, 2004). Ils appartiennent au
groupe 1 des agents cancérogènes classés par l'Agence
Internationale de Recherche sur le Cancer pour l'Union Européenne (IARC,
1987). Des études récentes menées par la Mutualité
Sociale Agricole (MSA) ont montré que l'arsénite de soude
était dangereux pour la santé des utilisateurs en dépit
des précautions prises lors de son application (Herlemont, 2004-b). Il
était capable de tuer la plupart des champignons associés aux
maladies du bois dans le xylème comme le montrent les travaux de
Larignon et al., (2008). Il n'avait toutefois aucune efficacité
sur l'Eutypiose (Larignon, 2009), et sa réintroduction serait une
aberration, tant au niveau écologique qu'au niveau pratique (Viret,
2004).
1.8.2 Moyens de lutte actuellement
disponibles
Il n'existe plus de méthodes curatives depuis
l'interdiction d'utilisation de l'arsénite de sodium. La lutte est donc
essentiellement préventive à l'heure actuelle (Larignon, 2009).
Les mesures prophylactiques ont pour objectif de réduire la
quantité d'inoculum et de diminuer les risques de contamination. Bien
que considérées comme nécessaires, il est impossible
d'affirmer que ces mesures soient suffisantes comme moyen de lutte efficace
contre l'Esca et le BDA (Sentenac et al., 2004). Contrairement
à l'Eutypiose, la taille tardive, de même que la
désinfection des plaies de taille (par des mastics7 ou par
l'escudo8), s'avèrent inefficace sur l'expression des
symptômes foliaires de l'Esca ou du BDA (Dumot et Ménard, 2005).
On note également qu'aucune expérimentation n'a permis de montrer
l'efficacité de la restauration des souches par le recépage
vis-à-vis de l'Esca et du BDA, cette pratique étant par ailleurs
efficace vis-à-vis de l'Eutypiose (Creaser et Wicks, 2004).
D'après Larignon et al., (2009), le rôle des sarments
laissés au sol (pouvant héberger les sources d'inoculum des
champignons associés aux maladies du bois) sur la propagation de ces
maladies n'est pas connu. Enfin, selon Lecomte et al., (2006-b), le
compostage des sarments permet cependant d'éradiquer les champignons
associés aux maladies du bois.
1.8.3 Lutte en
pépinière
Des recherches sur les méthodes de lutte sont
menées également au niveau de la production des plants en
pépinières car les champignons pionniers de l'Esca et du BDA se
propagent par le matériel végétal (Larignon et
al., 2006). Parmi les traitements effectués sur le matériel
de propagation, le traitement à l'eau chaude réduit l'incidence
de Phaeomoniella chlamydospora lorsqu'il est effectué à
une température de 50°C pendant 30 minutes (Edwards et al.,
2004). Fourié et Halleen (2006) montrent également que des
bains de bénomyl, de captane ou de didecylméthylammonium
chloride, réduisent sa présence dans les tissus ligneux au niveau
de la soudure et à la base du plant. L'utilisation d'autres substances
actives comme l'eau de Javel, l'oxyquinoléine, des Trichoderma
ne montre pas d'efficacité vis-à-vis des champignons
associés aux maladies du bois (Larignon et al., 2008). Ces
mêmes auteurs indiquent que des traitements sont également
effectués sur les plants à la sortie de la
pépinière. Le traitement à l'eau chaude effectué
à une température de 50°C pendant 45 minutes montre
7Il existe des mastics cicatrisants contenant des
agents fongicides comme le Ramag C (carbendazime + thirame) ou le Nectec
(imazalil + azaconazol) (Viret, 2004).
8 Association de flusilazole et de carbendazime
une bonne efficacité vis-à-vis de Phaeomoniella
chlamydospora et de Botryosphaeria obtusa, mais il ne
présente pas d'efficacité à l'égard de
Phaeoacremonium aleophilum et de Neofusicoccum parvum.
1.8.4 Recherches en cours
Les recherches menées par différents
laboratoires dans le monde ont porté sur l'évaluation de
l'efficacité de molécules chimiques et de produits biologiques
aussi bien au laboratoire (Jaspers, 2001) que dans le vignoble,
appliqués sous différentes formes (sécateur traitant,
injection de matières actives dans le tronc, protection des plaies par
badigeonnage et par pulvérisation, traitement sur toute la souche,
peinture) et à différents stades phénologiques de la vigne
(Di Marco et al., 2000 ;, Laukart et al., 2001 ; Sentenac
et al., 2004 ; Dula et al., 2007).
> Stimulation des Défenses Naturelles
L'activation des systèmes de défenses naturelles
de la plante pourrait être une piste de lutte sérieuse. Les «
défensines » sont des protéines produites de façon
spontanée par la vigne (entre autre) pour lutter contre diverses
bactéries et champignons. On les rencontre
préférentiellement au niveau des baies (protection contre le
Botrytis par exemple) et malheureusement assez peu ou pas dans les vaisseaux du
bois. Pour le moment, les éliciteurs étudiés (Stimulateurs
de Défense Naturelle comme l'ergostérol, qui induisent dès
leur application des réactions de défense naturelles)
n'augmentent pas la sécrétion de ces défensines. Les
études entreprises actuellement au niveau génétique visent
à étudier les plantes produisant naturellement beaucoup de ces
composés, à en étudier la mobilité dans le
xylème et à chercher à stimuler la sécrétion
de ces protéines (Goulet, 2008).
> Lutte biologique
Les recherches portent de plus en plus sur l'effet de
micro-organismes utilisés en lutte biologique, notamment les
Trichoderma. Ces champignons présentent une activité
antagoniste et d'hyperparasitisme à l'égard d'un grand nombre de
microorganismes (particulièrement ceux du sol), et sont utilisés
en lutte biologique à l'égard de diverses maladies sur
différentes cultures (Chet, 1987). Ils peuvent jouer également un
rôle dans l'élicitation des mécanismes de défense de
la plante (Shigo et al., 1977). Selon Hunt et al., (2001),
l'application de Trichoderma sur les plaies de taille réduit
fortement la colonisation du bois par Eutypa lata (-85%). De
même, Di Marco (2003) montrent que l'application d'une souche de
Trichoderma harzianum sur les plaies de taille empêche le
développement de Phaeomoniella chlamydospora.
> Lutte chimique
La lutte chimique est également une voie
étudiée, la difficulté résidant dans le fait que
les antifongiques traditionnels ne sont pas mobiles dans tout le système
de la plante (phloème et xylème)9. On cherche donc
à élaborer des fongicides systémiques phloémiens
doté d'une propriété particulière :
l'ambimobilité. De tels composés peuvent passer du liber
(phloème) vers le bois (xylème), et ainsi atteindre les
champignons parasites. Bien qu'il soit difficile d'effectuer les modifications
de structures moléculaires pour obtenir cette propriété
tout en conservant les propriétés fongicides de la
molécule initiale, les premiers résultats démontrent qu'il
est possible de créer de tels composés. Le fenpiclonil en
particulier, offre en effet des possibilités pour effectuer les
modifications de structure souhaitées (Jousse, 2004).
9 Les fongicides classiques sont soit de contact, auquel cas,
ils restent en surface des plantes traitées, soit systémiques
ascendants, c'est-à-dire véhiculés avec la sève
brute depuis les racines vers le feuillage. C'est ce qui explique que le seul
remède proposé jusqu'ici consiste en un badigeonnage des plaies
de taille, ceci afin de constituer une barrière pour stopper
l'envahissement par les spores de champignons parasites (Jousse, 2004).
1.8.5 Perspectives
Les essais menés jusqu'à maintenant par les
différents laboratoires dans le monde n'ont pas permis de trouver de
méthodes de lutte curatives ou préventives satisfaisantes
utilisant des produits chimiques, soit parce que les produits testés ne
sont pas efficaces, soit parce que leur mode d'application n'est pas
réalisable par le viticulteur (Larignon et al., 2009). Il
apparaît clairement qu'il sera très difficile de trouver un seul
et unique produit efficace (Larignon, 2009).
2) Alimentation hydrique de la vigne 2.1 Rôle et
influence de l'eau sur la vigne
D'après Zufferey (2007), l'alimentation hydrique de la
vigne est déterminante pour le fonctionnement physiologique de base de
la plante (croissance végétative et photosynthèse). Le
rôle de l'eau est primordial durant tous les stades de
développement végétatif et reproductif de la vigne, et sa
disponibilité détermine les aspects quantitatifs et qualitatifs
de la production (développement des baies, teneur en sucres,
acidité, polyphénols et arômes) (Crespy, 2003 et 2009).
La précocité du cycle phénologique
dépend essentiellement de la température du sol, qui
ellemême est en relation avec sa teneur en eau (Morlat, 1989). Van
Leeuwen (2001) précise que la contrainte hydrique joue un rôle
essentiel sur la précocité et que son effet est plus important
que celui de la température du sol. Par ailleurs, la vitesse de
maturation est en grande partie déterminée par le régime
hydrique de la vigne (Van Leeuwen et Seguin, 1994).
D'après Serrano (2001), la présence d'eau dans
le sol est globalement favorable au potentiel végétatif de la
vigne. Si elle en dispose suffisamment tout au long de son cycle, elle
privilégie sa croissance végétative : la
photosynthèse augmente et elle utilise alors son énergie à
la production de feuilles et tiges. La vigne offre par ailleurs une très
bonne résistance à la sécheresse qui est liée,
entre autres, à sa grande capacité d'exploration du sol
(Zufferey, 2007). L'implantation du système racinaire, sa morphologie et
son fonctionnement, sont des éléments importants qui
déterminent l'offre en eau pour la plante (Deloire et al.,
2005). L'utilisation de l'eau par la plante dépend aussi de la
disponibilité de cet élément dans le sol (variable selon
sa teneur et sa force de liaison avec les particules), et bien
évidemment du climat (précipitations et demande
évaporatoire) (Soltner, 2005). Cet auteur rappelle que le stock d'eau du
sol est un paramètre qui varie en fonction des propriétés
pédologiques (texture, structure, charge en éléments
grossiers et matière organique du sol).
2.2 Notion de contrainte hydrique modérée :
entre le manque et l'excès d'eau
A la différence de la plupart des productions
végétales, la vigne est généralement
cultivée en dessous de l'optimum agronomique. En effet, un régime
hydrique modérément limitant induit généralement
des conséquences bénéfiques sur la qualité des vins
produits. On parle dans ce cas de « contrainte hydrique »,
l'expression « stress hydrique » devant être
réservée à des situations critiques, où un manque
d'eau excessif altère la qualité du raisin ou met en péril
la pérennité de la vigne (Van Leeuwen et Vivin, 2008).
Les excès comme les manques d'eau sont
généralement néfastes à l'obtention d'une vendange
qualitative. Une alimentation hydrique non limitante tout au long de la saison
induit un excès de
vigueur qui altère la qualité de la
récolte : hausse des rendements, compétition accrue pour la
répartition des sucres entre partie végétative et
reproductive, phénomènes de dilution, réduction de la
coloration des vins, altération du microclimat de la zone des grappes,
développement de parasites... (Payan et Salançon, 2002). Au
contraire, en cas de déficit hydrique sévère, la
photosynthèse se trouve trop fortement limitée par la fermeture
des stomates (phénomène de régulation stomatique limitant
les échanges gazeux) et des blocages de la maturation peuvent se
produire. (Tregoat et al., 2002).
L'apparition d'une contrainte hydrique au cours de la saison
modifie profondément le fonctionnement physiologique de la vigne.
Celle-ci favorise l'arrêt de croissance des rameaux de la vigne et limite
le grossissement des baies, notamment lorsqu'elle intervient avant la
véraison (Becker et Zimmermann, 1984). L'idéal pour une
production de raisins de qualité est de se retrouver dans une situation
de contrainte hydrique modérée dans le stade
pré-véraison ou bien entre la véraison et la
récolte, ce qui permet d'atteindre des teneurs optimales des baies en
sucres, polyphénols et arômes (Van Leeuwen et Seguin 1994). Cet
état se traduit au niveau édaphique par un épuisement
quasiment complet des réserves en eau au cours de la maturation (Payan
et Salançon, 2002). On note que lorsque les racines se trouvent dans un
sol en cours d'assèchement, leurs extrémités produisent de
l'acide abscissique. Cette hormone est considérée comme
étant favorable à la maturation du raisin (Stoll et al.,
2000 ; Stoll et al., 2001).
2.3 Besoins en eau de la vigne
Il est admis que la vigne nécessite un minimum de 250
mm de pluie durant la période végétative et la maturation
des baies, même si la vigne peut subsister dans des conditions encore
plus sèches (Galet, 2000). Sous un climat où la
pluviométrie annuelle est inférieure à 350 mm, la plupart
des vignobles nécessitent une irrigation, quel que soit le type de sol.
L'évapotranspiration moyenne annuelle (ETM) des vignobles est
estimée de 350 à 600 mm (Deloire, 2008). On considère que
la culture de la vigne avec des rendements économiquement viables est
difficile lorsque la pluviosité annuelle est inférieure à
400 mm. Cette valeur doit cependant être modulée en fonction de la
répartition des précipitations au cours de l'année et de
la capacité de rétention d'eau des sols (Choné et al.,
2001-a).
2.4 Mouvements de l'eau dans le continuum
sol-plante-atmosphère (CSPA)
Du point de vue des transferts hydriques, le sol, la plante et
l'atmosphère forment un continuum (Cruiziat, 1997). L'eau en provenance
du sol est absorbée au niveau des racines et elle transite par la plante
avant de rejoindre l'atmosphère en sortant par les stomates des
feuilles. L'auteur explique que la transpiration au niveau des feuilles induit
une tension qui est le moteur principal du transfert de l'eau à travers
la plante. Elle crée un appel qui est retransmis dans le système
vasculaire grâce aux forces de cohésion de l'eau.
Laberche (2010) ajoute par ailleurs qu'un certain nombre de
plantes (dont la vigne) génèrent également une pression ou
« poussée radiculaire » au niveau de la racine. L'action de
cette poussée est annulée le jour dès que la transpiration
prend le relais, en induisant une tension dans les vaisseaux du xylème
(qui se comportent comme des capillaires).
D'après Zufferey et Murisier (2004), le continuum
sol-plante-atmosphère est parcouru par des résistances au
transfert de l'eau (résistance hydraulique de la part du sol et de
l'interface sol-racine, résistances dans la plante, depuis la racine
jusqu'aux feuilles et résistance stomatique). Ces auteurs
indiquent que les plantes tentent de maintenir la
différence de potentiel entre les feuilles et les racines
en-deçà du seuil de cavitation (voir paragraphe 1.6.4 :
Complication pathologique : cavitation, embolie et thyllose). La
régulation stomatique constitue en effet un moyen de réguler les
flux d'eau à travers la plante pour permettre le métabolisme et
la croissance du végétal, mais aussi pour éviter la
cavitation.
2.5 Régulation stomatique
Les stomates jouent un rôle clé dans la
régulation des flux d'eau et de CO2 des feuilles. Ils constituent le
moyen de contrôler les pertes d'eau par la transpiration et le
prélèvement de CO2 nécessaire à la
photosynthèse. Les mouvements stomatiques (ouverture et fermeture)
dépendent des variations de turgescence des cellules de garde et des
cellules épidermiques voisines. La turgescence résulte soit d'une
variation du potentiel hydrique des cellules de garde (phénomène
passif), soit d'une variation active du potentiel osmotique10 de
celles-ci. De nombreux facteurs externes (présence de lumière,
taux de CO2, température, hygrométrie, état hydrique du
sol, etc...) et internes (hormones végétales et état
hydrique du végétal notamment) influent sur le fonctionnement des
stomates (Laberche, 2010 ; Adrian et Fournioux, 2011).
2.6 Méthodes d'appréciation de la
réserve utile du sol et de l'état hydrique de la vigne
De nombreuses techniques sont utilisées pour suivre le
régime hydrique de la vigne (Van Leeuwen et al., 2001-a ; Van
Leeuwen et al., 2003). Les plus anciennes sont basées sur des
analyses d'échantillons de sol pour estimer la réserve en eau du
sol accessible à la vigne : soit directement (mesure de la teneur en eau
du sol par méthode gravimétrique), soit indirectement
(prédiction par les classes de pédotransfert à partir de
certaines données physiques du sol : texture, structure et type
d'horizon) (Ayachi, 2010). D'autres techniques, également
utilisées depuis longtemps, permettent de mesurer directement
l'évolution du stock d'eau du sol ou sa disponibilité pour la
plante (sondes à neutrons, sondes TDR- Time Domaine
Reflectometry- ou tensiométrie) (Seguin, 1970).
Le bilan hydrique théorique constitue une nouvelle
approche du régime hydrique de la vigne, au moyen de la
modélisation (Riou et Lebon, 2000 ; Lebon et al., 2003). Il
propose de simuler en continu le stock d'eau restant dans le sol (à
partir de données concernant le stock d'eau au début de la
saison), auquel on ajoute les apports (précipitations) et retranche les
pertes (évapotranspiration). Il présente l'avantage de
discriminer le degré de contrainte hydrique à travers trois
composantes : l'époque, la durée et l'intensité. Mais la
principale difficulté de cette approche concerne l'estimation du stock
d'eau au début de la saison, qui est extrêmement difficile
à estimer à cause de particularités propres à la
culture de la vigne (profondeur d'enracinement, sols caillouteux) (Van Leeuwen
et Vivin, 2008).
D'autres pratiques désormais plus répandues en
viticulture reposent sur l'utilisation de la vigne comme indicateur de son
propre état hydrique11. On parle dans ce cas «
d'indicateurs physiologiques », et ceux-ci sont nombreux : utilisation de
la chambre à pression de Scholander pour mesurer le potentiel hydrique
de la vigne (potentiel foliaire, potentiel de base ou potentiel tige)
(Choné et al.,, 2001-b), suivi des micro-variations de
diamètre d'organes de la vigne (Van Leeuwen et al.,, 2000),
mesures du
10 L'ouverture du stomate est provoquée par l'accumulation
de potassium dans la vacuole des cellules concernées (Laberche,
2010).
11 Un déficit hydrique provoque en effet un certain nombre
de modifications dans le fonctionnement physiologique de la vigne qui sont
mesurables (variations de la tension de la sève dans le xylème,
fermeture stomatique, déroulement de la photosynthèse...).
débit de la sève (Valancogne et Nasr, 1989) ou
encore mesure de la température des feuilles (Payan et Salançon,
2003).
La discrimination isotopique du carbone 13
(ÄC13) est un autre indicateur physiologique du régime
hydrique. Le C13 représente environ 1,108% du CO2
atmosphérique, contre environ 98,89% pour le C12. Ce dernier,
plus léger, est préférentiellement utilisé lors de
la photosynthèse. La contrainte hydrique, en provoquant la fermeture des
stomates pendant une partie de la journée, ralentit les échanges
de CO2 entre la feuille et l'atmosphère, et limite ainsi la
discrimination isotopique. Le rapport C13 /C12 se
rapproche, dans ces conditions, de celui du CO2 atmosphérique. La mesure
du ÄC13 sur les sucres du moût à maturité
constitue alors un indicateur global de la contrainte hydrique subie par la
vigne au cours de la période de maturation (Van Leeuwen et al.,
2001-b, Gaudillère et al., 2002).
2.7 Techniques mises en oeuvre dans la gestion du
régime hydrique de la vigne
La perte de qualité par un déficit hydrique
insuffisant est beaucoup plus fréquente que la perte de qualité
par un stress hydrique excessif, même si elle passe
généralement inaperçue. Lorsque la pluviosité
estivale et la réserve en eau des sols ne permettent pas d'atteindre de
façon régulière un déficit hydrique suffisant, il
faut augmenter la surface foliaire par hectare pour augmenter
l'évapotranspiration réelle et choisir un porte-greffe qui
valorise peu les réserves en eau du sol (Riparia Gloire de Montpellier
par exemple). Ces situations peuvent également être
valorisées par le choix de l'encépagement (cépages
précoces) (Van Leeuwen, 2001). A l'inverse, dans les situations
où on constate une perte de qualité due à un stress
hydrique excessif (climat très sec, sols avec de faibles réserves
en eau), il est possible de limiter les effets négatifs sur la vigne par
une adaptation de sa conduite et par un choix adapté du matériel
végétal (Choné et al., 2001-a). La meilleure
protection contre les effets néfastes d'un stress hydrique est une
limitation du rendement. Un rendement modéré permet d'avoir une
surface foliaire moyennement faible sans altérer le rapport
feuille/fruit. Le choix de porte-greffes résistants à la
sécheresse (par exemple le 110 Richter) est sûrement la forme
d'adaptation à la sécheresse la plus répandue. On peut
également signaler qu'une faible alimentation en azote de la vigne
réduit ses besoins en eau, par une limitation de la vigueur et de la
surface foliaire (Van Leeuwen et Vivin, 2008).
Dans des situations extrêmes, et lorsque la
législation le permet, le viticulteur peut également avoir
recours à l'irrigation. Lorsqu'elle est nécessaire, l'irrigation
doit progressivement amener la vigne à une situation de contrainte
hydrique modérée, tout en lui évitant de basculer dans une
situation de stress hydrique. Ce pilotage fin de l'irrigation peut être
réalisé grâce au suivi régulier de l'état
hydrique de la vigne par la mesure du potentiel tige (Choné et al.,
2001-b).
Les techniques d'irrigation les plus récentes
consistent à alimenter la vigne avec une quantité d'eau
volontairement réduite. Dans le cas du RDI (« Regulated Deficit
Irrigation »), une contrainte hydrique est volontairement
provoquée après la floraison par une période sans
irrigation, notamment pour réduire la taille des baies. Le PRD («
Partial Rootzone Drying ») fait appel à une irrigation
alternée : avec une périodicité d'environ deux semaines,
chaque côté du rang est irrigué de façon
alternée. Ainsi, une partie du système racinaire se trouve en
permanence dans un sol qui est en train de s'assécher. Un effet
très net est constaté sur le potentiel oenologique du raisin, qui
peut en partie être expliqué par une plus grande synthèse
d'acide abscissique en comparaison avec des vignes irriguées et ne
subissant aucune contrainte hydrique (Stoll et al., 2001).
Tableau 1 : Principales caractéristiques
agronomiques des parcelles d'étude Source : Ayachi
(2010)
|
Parcelle Région Code Appellation Cépage Clone PG
Date de
plantation
|
Distance
inter-rang
|
Distance
inter-cep
|
Mode de
conduite
|
|
St-Martin-Sous-Montaigu
|
Côte
Chalonnaise SL4 Les Montaigus Pinot Noir 115 161-49 1986 1.05 m
0.9 m Guyot simple
|
Volnay Côte de
Beaune CD8 Poissots Pinot Noir 115 161-49 1987 1 m 1 m Guyot
simple
|
Chassagne-
Montrachet
|
Côte de
Beaune CD9 Chassagne Pinot Noir 115 3309C 1983 1 m 0.9 m
Cordon
|
Tableau 2 : propriétés
géomorphologiques des parcelles étudiées
Source : Ayachi (2010)
Région Département Code Pierrosité de la
surface
(%) Formation géologique Pente
|
St-Martin Sous - Montaigu
|
Côte
Chalonnaise
(71)
|
Formation sur calcaire
SL 4 5% (graviers et cailloux) type bajocien supérieur
1%
|
Volnay Côte d'Or
(21) CD 8 1% (graviers) Formation sur cône de
déjection Surface plane
Chassagne-Montrachet Côte d'Or
(21) CD 9 1% (graviers et cailloux) Formation sur cône
déjection 1%
Tableau 3: Propriétés physico-chimiques et
hydriques des différents horizons des parcelles
d'étude
(MO : teneur en matière organique; CaCO3 : teneur en
carbonate de calcium ; EG : charge en éléments grossiers ; RU :
réserve utile totale exploitable par les racines). La texture est
déterminée à partir de diagramme de texture de Jamagne
(1967). (Horizons : ALO : argile lourde, A : argileuse, SA : sablo-argileux.
LAca : horizon organo-minéral (labouré) carbonaté ; Sca :
horizon structural carbonaté ; S/Cca : horizon de transition
carbonaté ; Cca : horizon d'altération calcaire ; LA : horizon
organominéral (labouré) ; S : horizon structural ; Sg : horizon
structural avec traces d'hydromorphie ; IICk : horizon d'altération
d'origine colluviale avec fortes précipitation de CaCO3 ; A : horizon
organo-minéral ; IIS : horizon structural d'origine colluviale ; Sca(g)
: horizon structural carbonaté avec des traces d'hydromorphie ; IICca :
horizon d'altération carbonaté d'origine colluviale ; Ccag :
horizon d'altération carbonaté avec traces d'hydromorphie.
Source : Ayachi (2010)
|
Parcelle (Code)
|
Horizon (cm)
|
Granulométrie
(en %)
|
Texture
|
MO
(%)
|
CaCO3 total (g/kg)
|
pH
|
EG
(%)
|
RU (mm)
|
|
Argile
|
Limon
|
Sable
|
|
|
LAca (0-18)
|
51,45
|
43,24
|
5,31
|
ALO
|
4,91
|
79
|
8,47
|
5
|
|
|
St-Martin sous
|
Sca (18-30)
|
51,80
|
44,00
|
4,20
|
ALO
|
3,26
|
90
|
8,43
|
2
|
|
|
|
|
|
|
|
|
|
|
|
45,68
|
|
Montaigu (SL4)
|
S/Cca (30-40)
|
52,05
|
43,44
|
4,50
|
ALO
|
3,89
|
97
|
8,42
|
10
|
|
|
Cca (40-80)
|
49,50
|
35,60
|
14,90
|
ALO
|
1,33
|
218
|
8,50
|
90
|
|
|
LA (0-25)
|
45,35
|
47,05
|
7,59
|
ALO
|
1,93
|
14
|
8,19
|
1
|
|
|
S (25-40)
|
45,90
|
46,10
|
8,00
|
ALO
|
1,37
|
10
|
8,12
|
1
|
|
|
Volnay (CD8)
|
|
|
|
|
|
|
|
|
|
133,55
|
|
Sg (40-100/130)
|
52,75
|
41,54
|
5,71
|
ALO
|
0,55
|
12
|
8,32
|
2
|
|
|
IICk (100/130-140)
|
43,76
|
25,17
|
31,07
|
A
|
0,43
|
266
|
8,36
|
80
|
|
|
A (0-25/35)
|
34,23
|
29,63
|
36,14
|
A
|
1,17
|
8
|
7,98
|
2
|
|
|
S (25/35-50)
|
35,04
|
27,83
|
37,14
|
A
|
1,23
|
7
|
7,87
|
1
|
|
|
Chassagne
|
IIS (50-70/80)
|
46,00
|
27,40
|
26,60
|
ALO
|
1,02
|
9
|
8,14
|
50
|
|
|
|
|
|
|
|
|
|
|
|
127,33
|
|
Montrachet (CD9)
|
Sca(g) (70/80-105)
|
30,90
|
39,00
|
30,10
|
A
|
2,25
|
258
|
8,70
|
3
|
|
|
IICca (105-125)
|
21,18
|
22,98
|
55,84
|
SA
|
0,42
|
288
|
8,62
|
30
|
|
|
Ccag (125-140)
|
33,33
|
24,82
|
41,84
|
A
|
0,56
|
20
|
8,54
|
10
|
|
PARTIE II : MATERIEL ET METHODES
Le BIVB réalise un suivi des maladies du bois
(Esca/BDA) depuis l'année 2006. On maîtrise encore mal
l'étiologie pour ces maladies. Parmi les différents axes de
recherches qui ont été définis, l'étude de
l'influence des facteurs abiotiques constitue une partie importante des
investigations. L'objectif de ce travail est de préciser l'influence de
l'alimentation hydrique de la vigne sur l'expression foliaire des
symptômes des maladies de l'Esca et du BDA.
1) Caractérisation des sites
d'études
1.1 Localisation et caractéristiques agronomiques
principales
L'étude a été menée dans la
région de Bourgogne (département de la Côte d'Or -21- et de
la Saône et Loire -71-). Trois parcelles ont été suivies,
elles font partie du réseau de suivi de la maturité mis en place
par le BIVB depuis 1988. Le tableau 1 en résume les principales
caractéristiques agronomiques.
Les parcelles sont plantées avec le même
cépage (Pinot-Noir) et le même clone (115) ; leurs densités
de plantation sont similaires. On note que SL4 et CD8 ont à peu
près les mêmes dates de plantation, les mêmes porte-greffes,
et le même mode de conduite. La parcelle CD9 a été
plantée quelques années plus tôt, avec un porte-greffe
différent. Elle est conduite en Cordon de Royat.
1.2 Données géomorphologiques
Les propriétés géomorphologiques des
parcelles étudiées sont présentées dans le tableau
2. Les données ont été obtenues à partir des
travaux d'Ayachi (2010).
Les parcelles CD8 et CD9 se situent en bas de coteau et
résultent de la formation de cônes de déjection. La
pierrosité observable en surface est faible (1%). On note la
présence d'une pente légère de 1% sur CD9. La parcelle SL4
est installée sur un substrat d'altération de nature calcaire
(bajocien supérieur). Elle est située en bas d'un mont et les
éléments grossiers représentent 5% de la surface. Cette
parcelle est quasiment plane, avec toutefois une faible pente observable sur
les 3 derniers entrepiquets, côté mont.
1.3 Données pédologiques
Au cours de l'année 2009/2010 une fosse a
été creusée dans chaque parcelle et une
caractérisation pédologique précise a été
effectuée par Ayachi (2010). Le tableau 3 regroupe les
propriétés physico-chimiques et hydriques de chaque parcelle. Les
photos détaillées des profils sont présentées en
annexe I.
On constate d'après le tableau que la profondeur du sol
varie d'une parcelle à l'autre. Elle est faible dans la parcelle SL4 et
très profonde dans les deux autres parcelles. Le nombre des horizons
varie de 4 à 6, chacun présente des caractéristiques
différentes des autres. On note une homogénéité
texturale du sol dans la parcelle SL4 (texture argile lourde), la même
texture pour la parcelle CD8 (à l'exception du dernier horizon de
texture argileuse). La parcelle CD9 est caractérisée par une
variabilité texturale, trois types de texture différentes ont
été observées (argileuse, argile lourde et


Figure 10 : Données climatiques - saison 2011
(janvier à septembre)
Données obtenues à
partir de la station météo de Rully (71) pour la parcelle SL4
età partir de la station météo de
Beaune (21) pour les
parcelles CD8 et CD9
sablo-argileuse). Les sols sont calcaires avec des teneurs en
carbonates de calcium variables d'une parcelle à l'autre et d'un horizon
à l'autre. La teneur en matière organique dans les horizons de
surface est plus forte par rapport aux horizons sous-jacents. La réserve
utile accessible à la vigne (calculée par rapport à la
profondeur du sol) est très variable, elle est plus forte dans les
parcelles à sols profonds (133,55 et 127,33 mm respectivement pour les
parcelles CD8 et CD9) et plus faible dans la parcelle la moins profonde (45,68
mm pour SL4).
1.4 Données climatiques
Les données climatiques de la saison 2011 sont obtenues
à partir des stations météorologiques du BIVB se situant
le plus proche possible des parcelles suivies. On note que si la parcelle SL4
est équipée d'une station, il n'en va pas de même pour les
parcelles CD8 et CD9, pour lesquelles les données de la station
météorologiques de Beaune ont été utilisées.
Les précipitations, la température et l'évapotranspiration
sont représentées dans la figure 10. La différence entre
les apports et les pertes cumulatifs nous permet de calculer le bilan
climatique de la saison (de janvier jusqu'à la récolte).
Comme les données climatiques sont identiques pour CD8
et CD9 (même source de données), ces deux parcelles ont
été regroupées dans le même graphique. Concernant le
calcul de leur bilan climatique (P-ETP), la différence entre ces deux
parcelles se fait au niveau de l'ETP. Cependant, les données pour ce
paramètre étant très proches pour les deux parcelles
(gabarit de végétation comparable et sol non enherbé), on
a considéré que cette différence était
négligeable, et seule une courbe moyenne a été
tracée. (En fin de saison, le 25/09/11, la valeur exacte du bilan
hydrique final calculé pour CD8 est de -343,58mm contre -344,04mm pour
CD9).
Sur ces graphiques, on constate que SL4 a reçu une
quantité de pluie plus importante que CD8/CD9 (somme des
précipitations du 01/01/11 au 25/09/11 pour SL4=528mm et pour
CD8/CD9=464mm, soit une différence de 64mm entre les deux stations). Par
ailleurs, on observe une répartition irrégulière des
précipitations au cours de la saison. Les quantités de pluies les
plus importantes sont tombées pendant les mois de juin, juillet et
août (somme des précipitations mensuelles pour SL4 : juin=100mm ;
juillet=141mm ; août=114mm et pour CD8/CD9 : juin=66mm ; juillet=130mm ;
août=101mm). On note également qu'à la date du 07/06/11, il
est tombé une quantité d'eau particulièrement importante
sur SL4 (60,8mm, contre 22,4mm pour CD8/CD9 le même jour).
A partir du mois de mai, la température a
commencé à augmenter progressivement, avec un maximum
observé le 22/08/11 pour les deux stations (Pour cette journée,
température moyenne sur SL4=27,5°C et pour CD8/CD9=27,3°C).
Sur l'ensemble de la période étudiée, la
température moyenne est à peu près identique pour les deux
stations (environ 13,8°C).
En calculant le bilan climatique (P-ETP) de la période
allant du mois de janvier jusqu'au 25 septembre, il paraît clairement que
le bilan climatiques est déficitaire pour les deux stations. Les
parcelles CD8 et CD9 enregistrent un déficit hydrique plus
élevé (environ -344mm) par rapport à la parcelle SL4
(-287,76 mm), soit une différence de 56mm entre les deux stations.
2) Techniques utilisées
2.1 Suivi du régime hydrique de la vigne
2.1.1 Notion de potentiel hydrique
Selon Riou et al., (1997), la notion de potentiel
hydrique permet de comparer les états hydriques des milieux physiques
(sol et ses différentes strates) et biologiques (plante et ses
différents organes), et de déterminer le sens de circulation de
l'eau. Le potentiel hydrique (exprimé en mégapascals ou en bars)
est une grandeur négative, qui représente l'énergie de la
liaison de l'eau avec un milieu (le potentiel de l'eau libre étant
égal à zéro). L'eau se déplace dans le sens des
potentiels décroissants. Le potentiel hydrique de l'air est presque
toujours plus bas que celui du sol, ce qui explique que les plantes soient
traversées par un courant d'eau ascendant. Le potentiel hydrique dans
les feuilles dépend de celui du sol et des
résistances12 au transfert de l'eau à travers la
plante.
2.1.2 Principe de la chambre à pression de
Scholander et techniques de mesure
D'après Payan et Salançon (2003), la
méthode de référence utilisée actuellement pour
étudier la contrainte hydrique de la vigne se réalise à
l'aide de la chambre à pression (ou « bombe ») mise au point
par Scholander et al. en 1965. Le principe consiste en l'application
d'une pression sur les parois du limbe d'une feuille insérée dans
une chambre hermétique, via un gaz inerte, jusqu'à ce que la
sève contenue dans la feuille exsude par la section du pétiole.
La pression exercée, lue sur un manomètre, correspond alors au
« potentiel hydrique régnant dans la feuille. Selon le mode
opératoire (heure de la mesure, ensachage de la feuille au
préalable ou non), la chambre à pression permet de suivre
plusieurs indicateurs physiologiques. Ainsi, plusieurs applications ont
été définies (Van Leeuwen et al., 2003) :
- le potentiel hydrique foliaire (ØF) :
mesuré sur une feuille au cours d'une journée ensoleillée,
sa valeur ne représente que le potentiel hydrique de la feuille
prélevée, à l'instant de la mesure.
- le potentiel hydrique foliaire de base (ØB) :
il correspond au même potentiel hydrique foliaire, mais il est
mesuré cette fois juste avant le lever du soleil. En fin de nuit les
stomates sont fermés, la transpiration est bloquée, et la mesure
reflète l'état d'hydratation de la plante entière, en
relation avec l'état de l'eau dans le sol (teneur en eau).
- le potentiel hydrique tige (ØT) : il est
mesuré au cours d'une journée ensoleillée, sur une feuille
qui a été recouverte par un sachet étanche et opaque
pendant au moins une heure avant la mesure : à l'obscurité les
stomates de la feuille se ferment, et le potentiel hydrique de la feuille
s'équilibre avec le potentiel hydrique du rameau entier (tige) qui la
porte. Cette mesure permet d'approcher l'état hydrique de la plante
entière en cours de journée (qui est sous la dépendance de
de la capacité de fourniture racinaire autant que de l'intensité
de la demande climatique au moment de la mesure). Selon Choné et
al., (2000), il s'agit de l'indicateur le plus précis dans le cadre
d'un suivi de la contrainte hydrique subie par la vigne, et c'est celui-ci qui
a été utilisé dans le cadre de cette étude.
12 Voir paragraphe 2.4 : Mouvements de l'eau dans le continuum
sol-plante-atmosphère (CSPA)

Figure 11 : Schéma du principe de fonctionnement
d'une chambre à pression
Un gaz sous pression (a) injecté dans une chambre
hermétique (b) contenant une feuille permet une exsudation de
sève à l'extrémité du pétiole (c). Lecture
de la pression (d). Source : Payan et Salançon (2003)

C D
A B
E
Photo 16 : Mesure du potentiel foliaire de
tige
A : chambre à pression de Scholander, avec bouteille de
gaz ; B : feuille ensachée ; C : découpe de
l'extrémité du pétiole ; D : mise en place de la feuille
dans la chambre et injection du gaz ; E : observation à la loupe du
ménisque de sève. Source : Dufouleur (2011)
Tableau 4 : Etat hydrique de la vigne selon la valeur
des potentiels foliaires de tige (Øt)
Source : Sibille
etal. (2007)
Øt (bars) Etat hydrique de la vigne
0 à -5 contrainte hydrique absente à
légère
-5 à -8 contrainte hydrique légère à
moyenne
-8 à -11 contrainte hydrique moyenne à forte
--11 à -14 contrainte hydrique forte à
sévère
< -14 contrainte hydrique sévère
2.1.3 Description et protocole d'utilisation de la
chambre à pression
La chambre à pression de Scholander est une enceinte
fermée permettant d'exercer une pression sur une feuille13.
Reliée à une bouteille d'azote liquide, elle est composée
d'un cylindre permettant d'enfermer la feuille dont le pétiole
dépasse par le couvercle. Le couvercle de la chambre comprend un joint
torique. Celui-ci, serré autour du pétiole, permet d'enfermer la
feuille de façon hermétique dans la chambre. Deux vannes
permettent de contrôler le débit de gaz entrant et sortant de la
chambre. Un manomètre indique l'évolution de la pression à
l'intérieur de celle-ci (Figure 11). Quand la pression du gaz
appliquée équilibre la pression de la sève dans la
feuille, la section du pétiole dépassant du couvercle s'humecte
de sève. Il est très important de régler finement la
montée en pression du gaz. Dans le cas contraire, on risque de
surestimer la mesure en relevant trop tard la pression sur le manomètre,
par rapport au moment de l'apparition de la sève.
2.1.4 Dispositif expérimental et mesure du
potentiel hydrique foliaire de tige (Øt)
Pour chacune des parcelles suivies et de façon à
limiter les effets de bordures, il a été défini une
micro-zone d'étude, à l'intérieur de laquelle 12 ceps de
référence ont été choisi sur 2 rangs
côtes-àcôtes. Les ceps ont été
sélectionnés sur les critères suivants : âge adulte
(> 4 ans) ; expression végétative correcte ; absence visible
de maladie ou de carence. Ils ont été marqués et
numérotés.
Les mesures de potentiel de tige ont été
effectuées par deux équipes différentes sur un
réseau de parcelles de référence géré par le
BIVB et par l'IUVV. Elles ont eu lieu en milieu de journée (midi
solaire), période pendant laquelle la demande évaporatoire est
maximale (ØTIGE MIN). On les a réalisées sur les ceps de
références, en sélectionnant des feuilles saines issues de
la partie médiane d'un rameau principal. L'ensachage a été
réalisé pendant la matinée, au minimum une à deux
heures avant la mesure, dans des doubles sacs plastiques opaques et
aluminisés en surface. Les feuilles ont été
prélevées une par une et juste avant la mesure, en arrachant le
pétiole au niveau de son insertion sur le noeud. Le pétiole a
été amputé de son extrémité à l'aide
d'un cutter bien aiguisé pour ne pas écraser les tissus. Il a
ensuite été introduit dans l'orifice du couvercle de la chambre
prévu à cet effet. L'application du gaz a été
effectuée une fois le couvercle refermé, avec une montée
en pression très progressive. Dès l'apparition du ménisque
de sève (observé à l'aide d'une loupe), la valeur de la
pression régnant dans la chambre a été notée (photo
16).
A partir des résultats obtenus, une valeur moyenne des
potentiels a été calculée pour chaque parcelle et pour
chaque date de mesures. Pour interpréter ces résultats et
caractériser le niveau de la contrainte hydrique subie au cours de la
saison, nous avons utilisé les valeurs seuils définies par
Sibille et al., (2007) pour les potentiels hydriques foliaires de tige
(Øt) (tableau 4).
2.2 Modélisation du bilan hydrique de la vigne
(modèle « Lebon et al., 2003 »)
2.2.1 Contexte d'utilisation
L'application des outils de diagnostic du stress hydrique
à la parcelle (mesure des potentiels hydriques de tige, dans cette
étude) fournit une information précise mais momentanée. La
multiplication des mesures étant difficilement envisageable sur
l'ensemble de la saison (nombre de feuilles limité et conditions
climatiques parfois défavorables), un modèle de bilan hydrique
(Lebon et al., 2003) a été utilisé en
complément, pour évaluer l'état hydrique de la vigne et la
disponibilité en eau offerte par le milieu. Le bilan hydrique
présente l'avantage de discriminer le degré de contrainte
hydrique à travers trois composantes: l'époque, la durée
et l'intensité.
13 Idéalement, cette feuille aura été
prélevée quelques dizaines de secondes avant d'être
placée dans l'enceinte (Choné et al., 2001-b).

Figure 12 : Schématisation des flux et des
quantités d'eau considérés par le bilan
hydrique.
ATSW = quantité d'eau accessible dans le sol à un
moment donné ; TTSW = quantité maximale d'eau utilisable
dans
le sol ; P = Précipitations ; R = Ruissellement de surface ; Tv =
Transpiration de la végétation ;
Es = Evaporation du sol ; Ru
= Réserve utile
Source : Payan et Salançon (2003)
Tableau 5 : Niveau de la contrainte hydrique de la
vigne
Source : Riou et Payan (2001)
|
Niveau de la contrainte
|
FTSW (%)
|
|
Absence de contrainte
|
> 40
|
|
Contrainte faible
|
21 - 40
|
|
Contrainte modérée
|
7 - 21
|
|
Contrainte forte
|
< 7
|
2.2.2 Principe de fonctionnement
Le modèle de bilan hydrique repose sur l'acquisition de
données simples et accessibles, caractérisant la parcelle sur les
plans climatiques, édaphiques et agro-physiologiques (paramètres
d'entrée). On considère le sol comme un réservoir qui se
rempli grâce aux précipitations, et qui se vide continuellement en
fonction de la demande climatique (évapotranspiration réelle, ou
ETR). Le modèle délivre une estimation quotidienne de la
fraction d'eau du sol utilisable par la plante, et il permet, de calculer un
indice caractérisant le vécu de la plante par rapport a cette
disponibilité en eau.
2.2.3 Equation de
modélisation
La quantité d'eau disponible a un instant «
t >> (Available Transpirable Soil Water, ou
ATSWt) est égale a la quantité d'eau accessible dans le
sol en début de saison (Total Transpirable Soil Water,ou
TTSW), a laquelle on additionne des gains issus des
précipitations (P) et retranche les pertes par transpiration de
la végétation (Tv), évaporation du sol
(Es) et ruissellement de surface (R) (figure 12). D'une
façon générale, la TTSW décrit les
potentialités hydriques de la parcelle.
ATSWt = TTSW + P - Tv - Es - R
On note ici que le modèle que nous utilisons (Lebon et
al., 2003), ne prend pas en compte le ruissellement de surface
(R) évoqué précédemment.
ATSWt = TTSW + P - Tv - Es
Le rapport entre la quantité d'eau du sol utilisable a
un instant « t >> (ATSWt) et la quantité
totale d'eau, lorsque le sol est au maximum de son état d'hydratation
(TTSW), représente la fraction d'eau du sol utilisable par la
plante a cet instant (Fraction of Transpirable Soil Water, ou
FTSWt). Elle reflète l'évolution du stock d'eau qui se
vide suite a des prélèvements par évapotranspiration et
qui se remplit par des apports extérieurs (précipitations).
FTSWt = ATSW / TTSW
2.2.4
Paramètres d'entrée
Il est nécessaire d'initialiser le modèle avec un
certain nombre de données relatives a la parcelle et a la période
pour laquelle la modélisation va être effectuée.
> Données cimatiques
Ce groupe comprend les paramètres suivants (une valeur
moyenne par jour) : précipitations ; température maximum et
minimum ; évapotranspiration potentielle (ETP). (Il est
possible de calculer ce dernier, a partir des températures minimales et
maximales en utilisant les formules empiriques de Hargreaves fournies dans le
modèle).
> Données édaphiques
La TTSW a été remplacée par la
valeur de la réserve utile (RU). Celle-ci a été
calculée a partir d'échantillons de sol prélevés
sur l'ensemble des horizons constituants le profil pédologique de chaque
parcelle, en utilisant des techniques de mesures des propriétés
hydriques du sol en laboratoire (teneur en eau volumique au point de
flétrissement et a la capacité au champ) (Ayachi, 2010).
> Données agro-physiologiques
Ce jeu de données comprend des informations relatives a
la position géographique de la parcelle, a la densité de
plantation, au gabarit de la végétation, a la phénologie
spécifique du cépage concerné (somme des
températures actives) et a la surface du sol (albédo,
enherbement...). Le modèle tient compte de la croissance de la
végétation et d'une régulation de la transpiration lorsque
le sol s'assèche.
2.2.1 Paramètres de sortie
Le modèle simule l'évolution du bilan hydrique
de la vigne, représenté par la FTSW (%) et
délivre un indice de stress de la vigne (ISv : taux de
conductance stomatique de la vigne variant en fonction de FTSW). Pour
caractériser le niveau de la contrainte hydrique subie au cours de la
saison, nous avons utilisé une classification proposée par Riou
et Payan (2001) (tableau 5).
2.3 Observation et notation des symptômes
d'Esca/BDA au vignoble
Selon le protocole mis en place pour ce suivi
(réalisé depuis 2006), seuls 10 rangs des parcelles
concernées ont été analysés (notion de micro-zone
au sein de la parcelle de référence). On note que les micro-zones
définies pour la mesure des potentiels hydrique foliaire de tige
(Øt) sont inclues dans celles qui ont été définies
pour le suivi des maladies du bois.
L'observation et la notation des symptômes ont eu lieu
en fin de cycle végétatif (début septembre), une date
tardive permettant de maximiser le nombre de symptômes exprimés.
Les différentes formes de l'Esca et du BDA ont été
discriminées (voir paragraphe 1.3 : Symptomatologie). Le nombre maximum
de ceps implantables sur la micro-zone (notion d' « emplacements »,
ce chiffre étant relatif à la densité de plantation
utilisée sur la parcelle), ainsi que le nombre de jeunes plants (exclu
de la population sensible aux maladies du bois), de ceps manquants et de ceps
morts (c'est-à dire n'ayant pas débourré :
différent de l'apoplexie totale) ont également été
comptés. Les taux d'expression des différentes formes des
maladies du bois ont été exprimés par rapport à la
population de ceps considérés comme sensibles aux maladies du
bois (nombre total de ceps vivants sur la micro-zone, hors jeunes).
2.4 Traitement des données
Les données ont été traitées en
utilisant le logiciels statistique Statbox v.7.1.9 et le tableur Excel 2010.

SL4


CD8
Figure 13 : Représentation graphique des
résultats de mesures du potentiel hydrique foliaire de tige
(Øt)
et de l'état hydrique de la vigne pour chaque date de
mesure.
(Lignes à l'extrémité des segments:
valeur maximale et minimale ;
ligne discontinue : médiane ; ligne en
gras : moyenne ; cercles : valeurs extrêmes)
PARTIE III: RESULTATS ET DISCUSSION
1) Alimentation hydrique de la vigne 1.1 Potentiel
hydrique foliaire de tige (Øt)
Les mesures de potentiel de tige ont été
réalisées sur les parcelles SL4 et CD8. Les résultats sont
présentés dans la figure 13. A la suite d'une
extériorisation massive de symptômes foliaires d'enroulement viral
touchant la quasi-totalité des ceps de la parcelle CD9, aucune mesure de
potentiel hydrique n'a été réalisée sur cette
dernière (absence de feuilles saines pour réaliser les
mesures).
En utilisant pour référence les valeurs seuils
définies par Sibille et al., (2007) pour interpréter ces
résultats, on constate que la vigne de la parcelle SL4 n'a pas subi une
contrainte hydrique importante durant l'ensemble de la période pendant
laquelle les mesures ont été effectuées (quatre mesures,
de juillet à août). Les valeurs obtenues sont plutôt
homogènes (écart-type = 0,92), ce qui indique que les
résultats sont cohérents et que l'on peut en tirer des
conclusions à l'échelle de la micro-zone d'étude.
A l'inverse, les résultats concernant CD8 sont plus
dispersés (écart-type = 1,91) et donc plus difficile à
interpréter. Bien que cette parcelle dispose d'une réserve utile
bien plus élevée que SL4 (respectivement 133,55mm contre
45,65mm), CD8 pourrait globalement avoir subi une contrainte hydrique
légèrement plus prononcée. Cette différence peut
s'expliquer par le fait que le volume des précipitations a
été plus important sur SL4 (somme des précipitations de
janvier à septembre=528mm) que sur CD8 (somme des précipitations
de janvier à septembre=464mm).
Résultats de Øt sur la totalité
de la période de mesure (bars)14 :
|
Parcelle
|
Maximum
|
Minimum
|
Moyenne
|
Médiane
|
Ecart-type
|
|
SL4
CD8
|
-3
-1,5
|
-6
-11,2
|
-4,47
-5,1
|
-4,1
-4,7
|
0,92
1,91
|
Sur la première moitié de la période de
mesures (période << P1 >> : du 1er au 31
juillet), les valeurs des résultats relatives à la parcelle CD8
sont groupées et comparables à celles observées pour SL4.
En revanche, à partir du mois d'août (période << P2
>> : du 1er au 17 août), les valeurs pour CD8 sont
nettement plus dispersées (écart-type : 2,43) : il est donc plus
hasardeux d'extrapoler ces résultats à l'ensemble de la
micro-zone pour P2.
Résultats de Øt sur la période P1
(bars)14:
|
Parcelle
|
Maximum
|
Minimum
|
Moyenne
|
Médiane
|
Ecart-type
|
|
SL4
CD8
|
-3
-3,3
|
-5
-5,9
|
-3,8
-4,4
|
-4
-4,3
|
0,51
0,72
|
14 Valeur extrême (-7 bars) exclue pour CD8

SL4
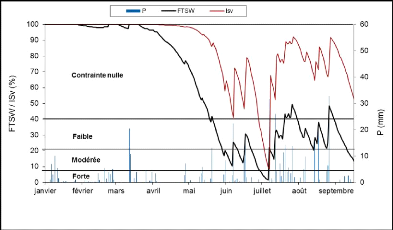
CD8

CD9
Figure 14 : Simulation du bilan hydrique de la vigne
(modèle « Lebon et al., 2003 ») - saison 2011
(janvier à septembre) Données obtenues à
partir de la station météo de Rully (71) pour la parcelle
SL4
et à partir de la station météo de
Beaune (21) pourles parcelles CD8 et CD9
|
Résultats de Øt sur la période P2
(bars) :
|
|
|
|
|
|
Parcelle
|
Maximum
|
Minimum
|
Moyenne
|
Médiane
|
Ecart-type
|
|
SL4
CD8
|
-4
-1,5
|
-6
-11,2
|
-5,1
-5,7
|
-5,3
-5,3
|
0,74
2,43
|
A l'avenir, afin d'améliorer la comparatibilité
des résultats, il serait intéressant de mieux caractériser
les parcelles au niveau pédologique. En effet, nous n'avons pas tenu
compte de l'hétérogénéité des sols au sein
des micro-zones étudiées. En intégrant à notre
approche des mesures de résistivité électrique des
sols15, la pertinence de nos comparaisons au niveau micro-local
serait significativement améliorée.
On note que le BIVB fait actuellement réaliser de
telles mesures par la société Géocarta®. Pour chaque
parcelle, cette société établit une cartographie en
couleur du proche sous-sol en trois dimensions qui permet d'étudier la
dynamique hydrique dans le sol et la variabilité spatiale de la
couverture géo-pédologique. En exploitant les cartes de
résistivité électrique, il est possible de subdiviser
chaque parcelle en plusieurs sous-zones distinctes, d'un point de vue
pédologique et hydrodynamique. A titre indicatif et en guise
d'illustration, les cartes de résistivité des parcelles SL4, CD8
et CD9 figurent en annexe II. Afin de réaliser des comparaisons de
potentiel hydrique foliaire de tige (Øt) intra et inter parcellaires qui
soient moins hasardeuses, il serait utile de redéfinir les micro-zones
d'étude en fonction des caractéristiques des sous-zones
parcellaires observées sur les cartes de résistivité.
1.2 Modélisation du bilan hydrique
L'utilisation du modèle « Lebon et al.
» (2003) nous a permis de simuler en continu le vécu hydrique
des parcelles d'études, de janvier à septembre. Sur la figure 14,
trois graphiques ont été tracés. Ils comportent les
précipitations et l'évolution de la FTSW (Fraction
of Transpirable Soil Water) relatives à chaque site. L'ISv
(Indice de Stress de la vigne) délivré par le modèle y est
également reporté, mais à titre indicatif seulement,
puisque cet indice est difficilement interprétable (absence de valeur
seuils d'interprétation).
La classification proposée par Riou et Payan (2001)
nous a permis de caractériser le niveau de la contrainte hydrique subie,
en fonction de la FTSW. On a pu ainsi définir 4 niveaux de
contraintes au sein même des graphiques (Contrainte nulle, faible,
modérée et forte).
En considérant globalement les trois graphiques, on
observe que la parcelle SL4 semble avoir subi des périodes de contrainte
hydriques plus importante que les deux autres parcelles. En effet, au cours de
la saison, la valeur de FTSW est nulle à trois reprises pour
SL4, alors qu'elle reste constamment positive pour CD8 et CD9. (On note que la
contrainte maximale pour ces deux parcelles est observée le 06/07/11,
avec une valeur de FTSW de 1,62% pour CD8 et de 1,23% pour CD9).
15 La résistivité électrique d'un sol
reflète sa capacité à résister au passage d'un
courant électrique. Elle varie en fonction de plusieurs facteurs
relatifs à la composition pédologiques du sol, parmi lesquels :
la texture, la profondeur du sol, la porosité, la pierrosité
(charge en éléments grossiers), l'humidité du sol, la
teneur en carbone du sol, l'origine du matériel parental et la teneur en
carbonates de calcium (CaCO3) (Goulet et Barbeau 2004; Goutouly etal.
2006).
Par ailleurs, on observe également que la parcelle SL4
semble reconstituer plus rapidement ses réserves en eau disponible pour
la plante. En effet, a plusieurs reprises, la valeur de FTSW atteint
100% en peu de temps. Ces variations brutales s'expliquent par la faible valeur
calculée de la réserve utile pour la parcelle SL4 : sa valeur
(45,65mm) est environ trois fois plus faible que la réserve utile des
parcelles CD8 et CD9 (respectivement 133,55mm et 127,33mm).
Il convient toutefois de nuancer les résultats
observés pour SL4. En effet, il est important de rappeler ici que la
valeur de la réserve utile, calculée a l'échelle de la
parcelle et assimilées a la TTSW dans le modèle, est une
estimation difficile a réaliser et dont l'incertitude n'est pas
négligeable. Comme on aura pu le constater précédemment
sur les cartes de résistivité des sols, la situation est complexe
au niveau micro-local, car hétérogène. Cette
complexité biaise forcément l'estimation de la réserve
utile (difficulté de réaliser une fosse pédologique
représentative de la profondeur d'enracinement,
hétérogénéité de la distribution des
systèmes racinaires a l'échelle de la parcelle, présence
de radicelles arrivant a s'insinuer dans les moindres fractures de la roche
mère...). Dans ces conditions, on comprend que le tracé de la
FTSW ne soit pas exactement représentatif de la fraction d'eau
réellement disponible pour la plante (sous-estimation des
réserves hydriques, le plus souvent).
En examinant les valeurs calculées par le
modèle, on constate une période de contrainte hydrique maximale
au début du mois de juillet, et ce pour les trois parcelles
étudiées. En effet, pour SL4, CD8 et CD9, les valeurs respectives
de la FTSW sont particulièrement basses et toutes
inférieures au seuil de 7% (contrainte forte). Si on compare ces
résultats a ceux que nous avons obtenus en mesurant les potentiels
hydriques foliaires de tige (Øt) a la même période, on
s'aperçoit qu'il y a contradiction entre les valeurs mesurée a la
vigne et les valeurs obtenues grâce au modèle. En effet, pour SL4,
les valeurs mesurées le 01/07/11 avec la chambre a pression indiquent
clairement que la contrainte hydrique est faible a légère (valeur
moyenne = -3,63 bars ; médiane = -3,5 bars ; écart-type = 0,57).
Au contraire, pour la même parcelle et a la même date, le
modèle indique une contrainte hydrique maximale, avec une valeur de
FTSW de 0%. De la même manière, les valeurs
mesurées le 27/06/11 sur CD8 reflètent une contrainte
légère (valeur moyenne = -4,95 bars ; médiane = -5,1 bars
; écart-type = 0,54) et s'opposent aux valeurs obtenues avec le
modèle qui indique une contrainte modéré pour la
même date (FTSW = 10,23%).
Ainsi, en comparant les valeurs des potentiels hydriques
foliaires de tige (Øt) dont nous disposons aux valeurs calculées
de la FTSW, nous avons mis en évidence des contradictions
avérées entre la réalité et le modèle
(sous-estimation de la FTSW). Ce dernier comporte donc des limites. En
plus de l'estimation difficile et parfois approximative de la réserve
utile (correspondant a la TTSW), on note également qu'il est
fait abstraction dans le modèle des flux hydriques liés au
ruissellement, des phénomènes de remontées capillaires
et/ou des mouvements d'eau souterrains. De plus, la transpiration d'une
éventuelle culture intercalaire (enherbement par exemple) y est mal
estimée (aucune spécification par rapport a l'espèce
concernée, sa densité, son cycle...).
Finalement, on retiendra que, si le modèle constitue un
moyen pratique et utile pour représenter la contrainte hydrique en
continu sur un millésime, on n'oubliera pas qu'il ne constitue qu'une
simplification parfois abusive de la réalité. Les valeurs
obtenues par le calcul a l'aide du modèle « Lebon et al.
» (2003) sont donc a interpréter avec recul et prudence.


A
B
Figure 15 : Suivi des taux d'expression globaux des
maladies de l'Esca et du BDA (tous symptômes confondus) (A :
Evolution annuelle de l'expression des maladies par parcelles
étudiées ;
B : Comparaison de l'expression des maladies dans les
parcelles étudiées par année)
Source : données SL4, 2006 à 2010 : Chedozeau,
2010 ; données CD8 et CD9, 2006 à 2010 : Reviron, 2010
2) Expression des maladies du bois (Esca/BDA)
En reprenant les données de Chedozeau (2010) et de
Reviron (2010)16, nous avons pu représenter
l'évolution des taux d'expression globaux (c'est-à-dire tous
symptômes confondus) pour chaque parcelle étudiée et pour
chaque année, de 2006 à 2011 (Figure 15). Concernant
l'année 2011, nous avons discriminé les différentes formes
d'expression symptomatologiques de l'Esca et du BDA pour chacune des parcelles
(Figure 16). Les taux d'expression ont été exprimés par
rapport à la population de ceps considérés comme sensibles
aux maladies du bois (nombre total de ceps vivants, hors jeunes). A titre
indicatif, le tableau fourni en annexe III rassemble pour chaque micro-zone de
comptage des maladies du bois le nombre d'emplacement ainsi que les taux de
ceps jeunes, morts et manquants (exprimés par rapport au nombre
d'emplacement sur la micro-zone de comptage des maladies du bois).
> Analyse globale (2006 - 2011)
On constate tout d'abord qu'au cours des 6 dernières
années, la parcelle CD8 semble avoir été la moins
touchée, avec un taux d'expression global moyen (Esca et BDA confondu)
de 2,7% sur toute cette période (contre 10,9% pour SL4 et 15,7% pour
CD9). En 2011, avec un taux annuel global de 3,5%, c'est également CD8
qui exprime le moins de symptômes (contre 8% pour SL4 et 4,6% pour CD9).
Pour les trois parcelles étudiées, en comparaison avec des
années comme 2006, 2007 ou 2008, on considère que le
millésime 2011 ne constitue pas une année à forte
expression symptomatique pour les maladies du bois. On note toutefois que nos
valeurs sont largement supérieures au taux d'expression global moyen
d'environ 0,5% défini pour le pinot noir (figure 8).
> Analyse pour SL4 (2011)
Avec 8% d'expression tout symptômes confondus (Esca et
BDA), cette parcelle est la plus touchée des trois zones que nous avons
étudiées cette année. On compte 0,7% de ceps
frappés d'apoplexie (Esca et BDA confondus). La répartition des
symptômes d'Esca et de BDA semble à peu près
équivalente pour les deux maladies. On remarque toutefois que le BDA
semble être à peine plus dominant (formes d'apoplexie partielle
plus importante).
> Analyse pour CD8 (2011)
Comme nous l'avons déjà remarqué dans
notre analyse globale, la parcelle CD8 est la moins touchée des trois
sites d'études (taux annuel global de 3,5%). Nous n'avons trouvé
aucuns cep apoplectiques, qu'il s'agisse de la forme totale ou partielle. Avec
davantage de formes lentes observées, le BDA semble
légèrement dominer dans cette parcelle.
> Analyse pour CD9 (2011)
La parcelle CD9 possède un taux annuel global de 4,6%.
On note que c'est sur cette zone que l'on a rencontré le plus de ceps
manifestant l'apoplexie totale (2%). Dès lors, on peut se demander si
l'âge moyen de la population de ceps, significativement plus
élevé pour CD9 que pour SL4 et CD817, n'a pas
favorisé l'expression de ce symptôme critique. Quoi qu'il en soit,
il conviendra de garder à l'esprit cette différence d'âge
en comparant les parcelles entre elles, les ceps plus âgés
exprimant plus facilement les symptômes des maladies du bois. Sur CD9,
les formes d'apoplexie partielle liées au BDA sont observées plus
fréquemment.
16 Pour chaque année et pour chaque parcelle, les
données sont relatives aux mêmes micro-zones que celles que nous
avons établies.
17 SL4 et CD8 possèdent des dates de plantation qui sont
comparables (respectivement 1986 et 1987), alors que CD9 a été
plantée plus précocement, en 1983.



SL4
CD8
CD9
3) Relations entre alimentation hydrique et maladies du
bois (Esca/BDA)
Pour chacune des parcelles étudiées, nous allons
maintenant essayer de trouver des liens de cause à effet entre
l'alimentation hydrique et l'expression foliaire des symptômes des
maladies du bois.
> Analyse pour SL4
Rappelons tout d'abord que la parcelle SL4 possède la
plus faible réserve utile des trois sites que nous avons
étudiés (45,65mm, contre 133,55mm pour CD8 et 127,33mm pour CD9).
On comprend donc que la probabilité d'observer une contrainte, voire un
stress hydrique sur cette parcelle est plus élevée que pour les
deux autres. Pourtant, si on se réfère à nos valeurs de
potentiel hydrique foliaire de tige (Øt), on constate que le niveau de
la contrainte hydrique pour SL4 était plutôt léger. Les
précipitations abondantes sur cette parcelle expliquent en partie ce
résultat (somme des précipitations de janvier à
septembre=528mm). Même si cette constatation est en contradiction avec
nos résultats issus de la modélisation du bilan hydrique, on
considérera donc que SL4 n'a globalement pas subi de contrainte
importante au niveau hydrique en 2011. En effet, comme nous l'avons
expliqué précédemment, le modèle Lebon et al.
(2003) comporte des limites, et il est fort probable qu'il ait
surestimé l'intensité de cette contrainte.
Dès lors, nous pouvons tenter de rapprocher ce statut
hydrique (contrainte globalement légère) du fait que, cette
année, la parcelle SL4 semble avoir été la plus
touchée par les maladies du bois (8% d'expression globale, tout
symptômes confondus pour l'Esca et le BDA). Bien que disposant d'une
réserve utile plutôt faible (45,65mm), la parcelle SL4 a
bénéficié d'une alimentation en eau non limitante. En
s'appuyant sur les travaux de Goutouly (2010), on peut penser que cette
situation a favorisé un métabolisme orienté vers la
croissance végétative des ceps, ce qui a entrainé un
faible rapport Carbone/Azote dans les tissus végétaux et donc une
baisse des capacités de résistance de la vigne vis-à-vis
de l'Esca et du BDA (faible biosynthèse de molécules de
défense, comme les composés phénoliques).
> Analyse pour CD8
Concernant la parcelle CD8, nous avons constaté qu'il
était plus difficile de définir un statut hydrique global pour la
saison à partir de nos seules valeurs de potentiel hydrique foliaire de
tige (Øt), vu la dispersion des résultats à partir du mois
d'août. On note cependant que cette parcelle possède une
réserve utile importante (133,55mm), ce qui, a priori, ne la
prédispose pas à des niveaux de contrainte hydrique
sévères. De plus, en reprenant l'évolution de la
FTSW (dont on peut penser qu'elle a été
sousestimée par le modèle), on considèrera que CD8 n'a
globalement pas subi de contrainte importante au niveau hydrique en 2011.
Rappelons malgré tout que la somme des précipitations de janvier
à septembre observée pour cette parcelle (464mm) a
été moins importante que pour SL4 (528mm sur la même
période).
Le statut hydrique de CD8 (contrainte globalement plutôt
légère), combiné à son faible taux annuel
d'expression globale (3,5% tous symptômes confondus pour l'Esca et le
BDA, avec aucun cep apoplectique), constituent une situation qu'il est
difficile d'expliquer. En effet, CD8 ne subissant pas de contrainte hydrique
prononcée, on aurait pu s'attendre à ce que cette parcelle soit
au moins aussi touchée que SL4. Or il n'en est rien : même en
reprenant les résultats des années antérieures (2006
à 2011), on constate que la parcelle CD8 (qui comporte une
réserve utile importante) a été
systématiquement et significativement moins
touchée par les maladies du bois (Esca et BDA) que la parcelle SL4 (qui
comporte pourtant une réserve utile relativement faible). Ce
résultat va à l'encontre des explications proposées par
Goutouly (2010) vis-à-vis du rapport C/N dans les tissus
végétaux.
> Analyse pour CD9
Vu que cette parcelle n'a fait l'objet d'aucune mesure de
potentiel hydrique foliaire de tige (Øt), vu l'extériorisation
massive de symptômes foliaires d'enroulement viral et vu la date de
plantation significativement plus précoce (comparée à SL4
et CD8), nous pensons qu'il n'est ni pertinent, ni utile de réaliser une
analyse pour cette parcelle. Ajoutons que celle-ci devrait
impérativement être écartée du réseau de
suivi des maladies du bois, en raison de sa nonreprésentativité
(taux de ceps atteint de virose proche de 100%).
Conclusion générale
L'Esca et le Black Dead Arm sont des maladies graves,
complexes et multifactorielles qui impliquent plusieurs agents
pathogènes. Les interactions entre le climat, le sol, le cépage
et l'équilibre physiologique de la plante rendent l'étude de ces
pathologies particulièrement difficile. On peut penser que de nombreuses
investigations seront nécessaires afin de mieux comprendre ces syndromes
de dépérissement. L'alimentation hydrique de la vigne est un
facteur étiologique difficile à étudier, notamment parce
qu'il suppose un niveau de connaissances géo-pédologiques
approfondi des sites d'études. L'utilisation de cartes de
résistivité des sols nous permettra à l'avenir de
définir des zones d'études homogènes et comparables. Par
ailleurs, il serait impératif de mieux connaître les sources
d'infection et de transmission des différents agents pathogènes
afin de pouvoir prendre des mesures sanitaires adéquates. Rappelons
qu'une réintroduction de l'arsénite de soude serait une
aberration, tant au niveau environnemental qu'en matière
d'efficacité, puisque les pays qui ne l'ont jamais appliqué sont
touchés au même degré que les pays qui l'ont utilisé
régulièrement (Viret, 2004).
Perspectives et améliorations du dispositif
expérimental
Nous avons vu que le millésime 2011 ne constituait pas
une année à forte expression symptomatologique pour les parcelles
étudiées. Nous considérons donc qu'il serait important et
intéressant de poursuivre les observations dans le but d'obtenir des
résultats qui soient plus significatifs. Dans cette optique il
conviendrait d'améliorer le dispositif expérimental en tenant
compte des remarques suivantes.
Tout d'abord, il semble primordial de définir des zones
d'études qui soient homogènes d'un point de vue hydrodynamique,
et si possible géo-pédologique. L'utilisation de cartes
parcellaires de résistivité des sols permettrait même de
délimiter plusieurs micro-zones d'étude par parcelle afin de
réaliser des comparaisons intra-parcellaires. Par ailleurs, il serait
particulièrement intéressant de disposer de stations
météorologiques supplémentaires afin de pouvoir travailler
avec des jeux de données climatiques précis et individuels pour
chacune des parcelles concernées. En outre, il serait également
judicieux d'intégrer plus de parcelles à cette étude (si
possible avec une date de plantation qui soit comparable), afin de lui
conférer plus de poids.
Enfin, il serait utile d'intégrer la notion d' «
itinéraire hydrique » pour caractériser plus
précisément et comparer plus facilement les niveaux de contrainte
hydrique au cours d'une saison. Cette méthode utilisée pour
imager le vécu hydrique d'une parcelle viticole au cours de la saison a
été proposée par Codis (2000). Elle présente
l'avantage de discriminer les conditions d'alimentation en eau à deux
niveaux : par la date d'apparition et par l'intensité de la contrainte.
Le cycle végétatif est scindé en cinq classes
représentatives des principaux stades phénologiques de la vigne.
A chaque classe est attribuée une note de contrainte hydrique de 0
à 3 (ou « note de stress ») en fonction des réserves en
eau sur la période considérée. Un «
itinéraire hydrique » est ainsi défini par un
quintuplé pour chaque parcelle et par cycle végétatif. Ce
quintuplé constitue l'image du régime hydrique qui a
prévalu sur une parcelle en cours d'année.
ANNEXE I :
Profils pédologiques des parcelles
d'études
Source : Ayachi (2010)
- Profil pédologique de la parcelle
Saint-Martin-sous-Montaigu SL4 - (Calcosol)
- Profil pédologique de la parcelle Volnay CD8 - (Brunisol
rédoxique sur colluvion calcaire)
- Profil pédologique de la parcelle Chassagne Montrachet
(CD9) (Colluviosol rédoxique complexe)

0 cm
LAca
18 cm
Sca
30 cm
S/Cca
40 cm
Cca
80 cm

0 cm
LA
25 cm
S
40 cm
Sg
100 cm
IICk
140 cm
Profil pédologique de la parcelle Volnay
CD8
(Brunisol rédoxique sur colluvion calcaire)

35 cm
S
50 cm
IIS
80 cm
Sca(g)
105 cm
IICca
125 cm
Ccag
140 cm
Profil pédologique de la parcelle Chassagne
Montrachet (CD9)
(Colluviosol rédoxique complexe)
ANNEXE II :
Cartes de la résistivité
électrique des sols des parcelles d'études,
avec
identification de différentes sous-zones parcellaires
(Mesures réalisées par la
société GEOCARTA®, Données BIVB 2011)

CD8
CD9
SL4
ANNEXE III:
Dénombrement des emplacements et indication des
taux de ceps jeunes,
manquants et morts (non-débourrés) sur
les micro-zones d'étude
(Données Dufouleur 2011)
|
Parcelle
|
Code
|
Nombre
d'emplacements
|
Jeunes ceps
|
Pied manquant
|
Pied mort
(non-débourré)
|
|
St-Martin-
|
|
|
|
|
|
|
Sous-
|
SL4
|
1 100
|
39,9%
|
0,9%
|
2,6%
|
|
Montaigu
|
|
|
|
|
|
|
Volnay
|
CD8
|
1 070
|
12,2%
|
2,3%
|
5,4%
|
|
Chassagne-
|
|
|
|
|
|
|
Montrachet
|
CD9
|
990
|
0,1%
|
7,3%
|
11,3%
|
(Pourcentages établis par rapport au nombre
d'emplacements sur la micro-zone de comptage des maladies du bois)
BIBLIOGRAPHIE
Adrian M. et Fournioux J.C., 2011, <
Morphologie et anatomie de la vigne », Ed. Féret, 143
p.
Ayachi S., 2010, < Caractérisation
de la réserve utile des sols viticoles bourguignons dans le
réseau de suivi des maladies du bois», Mémoire de fin
d'étude (Master professionnel « Vigne et terroir » ; Institut
Universitaire de la Vigne et du Vin de Dijon et Agrosup Dijon), pp. 30
Becker N. et Zimmermann A., 1984, <
Influence de divers apports d'eau sur des vignes en pots sur la maturation des
sarments, le développement des baies et la qualité du vin »,
Bulletin O.I.V. 641-642, pp. 584- 596
Bruno G., Sparapano L., Graniti A., 2007,
<Effects of three esca-associated fungi on Vitis vinifera L.: IV.
Diffusion through the xylem of metabolites produced by two tracheiphilous fungi
in the woody tissue of grapevine leads to esca-like symptoms on leaves and
berries », Physiological and Molecular Plant Pathology 71, pp.
106-124
Carbonneau A., 1998, < Irrigation, vignoble
et produits de la vigne », Traité d'irrigation, Editions TEC
DOC Lavoisier, pp. 257-276
Carbonneau A., Deloire A., Jaillard B., 2007,
< La vigne : physiologie, terroir, culture », Ed.
Dunod, 442 p.
Chedozeau, C., 2010, < Rôle de
l'alimentation hydrique sur l'expression des maladies du bois : Esca et Black
Dead Arm », Mémoire de fin d'étude (Licence des Sciences
de la Vigne), Institut Universitaire de la Vigne et du Vin de Dijon, 30
p.
Chet I., 1987, < Trichoderma -
Application, mode of action, and potential as biocontrol of soilborne plant
pathogenic fungi », Innovative Approaches to Plant Disease
Control, pp. 137-160.
Chiarappa L., 1959, <Wood decay of the
grapevine and its relationships with black measles disease »,
Phytopathology 49, pp. 510-519 (in : Larignon P., Fontaine
F., Farine S., Clément C. et Bertsch C., 2009, < Esca et
Black Dead Arm : deux acteurs majeurs des maladies du bois chez la Vigne
», C. R. Biologies 332, pp. 765- 783)
Chone X., Tregoat O., Van Leeuwen C. et Dubourdieu D.,
2000, < Déficit hydrique modéré de la vigne:
parmi les trois applications de la chambre à pression, le potentiel tige
est l'indicateur le plus précis », Journal International des
Sciences de la Vigne et du Vin 34-n°4, 169-176
Choné X., Tregoat O., Van Leeuwen C., 2001-a,
< Fonctionnement hydrique des terroirs, base de l'irrigation
raisonnée de la vigne », Journal International des Sciences de
la Vigne et du Vin, N° Hors-série: « Un raisin de
qualité : de la vigne à la cuve », pp. 47-51
Choné X., Van Leeuwen C., Dubourdieu D.,
Gaudillère J.P., 2001-b, <Stem water potential is a
sensitive indicator for grapevine water status », Annals of Botany
87, pp. 477-483
Christen D., 2006, <Towards an integrative
management of eutypa dieback and esca disease of grapevine »,
thèse doctorat, Swiss Federal Institute of Technology, of Zurich
(ETH)
Codis S. 2000, <Elaboration d'un outil de
diagnostic du stress hydrique de la vigne à partir d'un modèle de
bilan hydrique et de la température foliaire. Application à la
typologie d'un réseau de parcelles », Mémoire de fin
d'études, E.N.S.A. Montpellier-E.N.E.S.A. Dijon, 65 p
Crespy A., 2003, « Fonctionnement des
terroirs et savoir-faire viticole : les clés de la qualité
», Ed. Avenir OEnologie, 191 p.
Crespy A., 2009, « De la vigne à la
cave - Evaluation de la quantité et de la qualité d'une
récolte », Ed. Avenir OEnologie, 159 p.
Cristinzio G., 1978, « Gravi attacchi di
Botryosphaeria obtusa su vite in provincia de Isernia »,
Informatore Fitopatologico 28, pp. 21-23 (in : Larignon P.,
Fontaine F., Farine S., Clément C. et Bertsch C., 2009,
« Esca et Black Dead Arm : deux acteurs majeurs des maladies du bois chez
la Vigne », C. R. Biologies 332, pp. 765-783)
Cruiziat P. 1997, « Les relations
plante-eau, de la cellule à la plante entière », pp. 1-40
(In : Riou C., Bonhomme R., Chassin P., Neveu A., Papy F.,
1997, « L'eau dans l'espace rural.
Production végétale et qualité de l'eau »,
INRA-Editions, 424 p.
Deloire A., 2008, « Irrigation de la vigne
: synthèse », SupAgro - Montpellier, 24 p.
Deloire A., Zebic O., Bernard N., Brenon E., Hunter J.J.,
2005, « Influence de l'état hydrique de la vigne sur le
style de vin », Revue française d'oenologie 215, pp. 11-15
Desaché F., Courlit Y. et Ménard E.,
1994, «Evaluation de la nuisibilité économique de
l'Esca de la vigne », Annales ANPP, 4ème Conférence
Internationale sur les Maladies des Plantes, tome 3, pp. 1341-1348
(in : Dubos B., 2002 - « Maladies cryptogamiques
de la vigne », Ed. Féret, 160 p.)
Di Marco S., 2003, « Experiments on the
control of esca by Trichoderma », Phytopathologia Mediterranea
43, pp. 108-115
Di Marco S., Mazzulo A., Calzarano F., Cesari A., 2000,
« The control of esca: Status and perspectives »,
Phytopathologia Mediterranea 39, pp. 232-240.
Dubos B., 2002, « Maladies cryptogamiques
de la vigne », Ed. Féret, 160 p.
Dula T., Kapes M.E., Horvath A., Rabai A., 2007, «
Preliminary trials on treatment of esca-infected grapevines with trunk
injection of fungicides », Phytopathologia Mediterranea 46, pp.
91-95
Dumot V. et Ménard E., 2005, «
Résultats d'un essai de longue durée sur les maladies du bois en
Charentes », cahier technique EUROVITI 15em éd., Montpellier,
30 nov- 1er déc. 2005, pp. 71-78
Edwards J., Pascoe I.G., Salib S., Laukart N., 2004,
« Hot water treatment of grapevine cuttings reduces incidence of
Phaeomoniella chlamydospora in young vines », Phytopathologia
Mediterranea 43, 158 p.
Edwards J., Salib S., Thompson F., Pascoe I.G., 2007,
«Effect of Phaeomoniella chlamydospora infection on the
grapevine's physiological response to water stress », Phytopathologia
Mediterranea 46, pp. 68-86
Fisher M., 2002, « A new wood-decaying
basidiomycete species associated with esca of grapevine: Fomitiporia
mediterranea (Hymenochaetales) », Mycological Progress
1, pp. 315-324 (in : Abou-Mansour E., Polier J., Pezet R. and
Tabacchi R., 2009, « Purification and partial
characterisation of a 60 KDa laccase from Fomitiporia mediterranea
», Phytopathologia Mediterranea 48, p. 447)
Fourié P.H., Halleen F., 2006,
«Chemical and biological protection of grapevine propagation
material from trunk disease pathogens », European Journal of Plant
Pathology 116, pp. 255-265.
Galet P., 2000, « Précis de
Viticulture », Ed. Déhan, 584 p.
Gaudillère J.P., Van Leeuwen C., Ollat N.,
2002, «Carbon isotope composition of sugars in grapevine,
integrated indicator of vineyard water status », Journal of
Experimental Botany 53, pp. 757-763
Geoffrion R. et Renaudin I., 2002, «
Tailler contre l'Esca de la vigne. Une mesure utile face à la menace de
recrudescence de cette ancienne maladie du bois », Phytoma 554,
pp. 23-27
Goulet E., 2008, « Les Maladies du Bois au
vignoble : Chercheurs et Techniciens autour de la table », Compterendu
de journée thématique sur les maladies du bois (IFV,
Toulouse), 3 p.
Goulet E. et Barbeau G., 2004, « Apports
des mesures de résistivité électrique du sol dans les
études sur le fonctionnement de la vigne et dans la spatialisation
parcellaire », Congrès « Viticultural Terroir Zoning
», Cape Town (Afrique du Sud), 9p.
Goutouly J.P., 2010, « Influence de la
physiologie de la plante sur le développement de l'Esca »,
Compte-rendu de journée thématique « Les maladies du
bois de la vigne », 16-17 novembre 2010, Villefranche/Saône, p.
20
Goutouly J.P., Destrac-Irvine A., Laveau C.,
Guérin-Dubrana L., 2007, « L'écophysiologie de la
vigne. Mieux comprendre les maladies de dépérissement »,
L'Union Girondine 1035, pp. 28-32
Goutouly JP, Rousset D, Perroud H, et
Gaudillère JP, 2006, « Caractérisation de la
variabilité spatiale et temporelle de l'état hydrique d'un sol de
vigne par résistivité électrique », VIem
Congrès International des terroirs viticoles, 6p.
Grosmann J., 2008, « Observatoire national
et régional des maladies du bois : bilan et perspectives de 4
années d'observations », Euroviti, pp. 8-17
Guérin L., 2004, « Quelles approches
épidémiologiques des maladies du bois ? », Rencontre
Technique: Les maladies du bois en Midi-Pyrénées (Station
Régionale ITV Midi-Pyrénées), pp. 34-35)
Harmandon F., 2004, « Effets toxiques de
l'arsénite de sodium sur la santé humaine et exposition des
utilisateurs », Rencontre Technique: Les maladies du bois en
Midi-Pyrénées (Station Régionale ITV
MidiPyrénées), pp. 8-12
Herlemont B., 2004-a, « L'observatoire
national des maladies du bois et la situation en France », Rencontre
Technique: Les maladies du bois en Midi-Pyrénées (Station
Régionale ITV Midi-Pyrénées), p. 14
Herlemont B., 2004-b, « Contexte
réglementaire de l'interdiction de l'arsénite de sodium »,
Rencontre Technique: Les maladies du bois en Midi-Pyrénées
(Station Régionale ITV Midi-Pyrénées), p. 13
Hunt J.S., Gale D.S.J. et Harvey I.C., 2001,
«Evaluation of Trichoderma as biocontrol for protection against
wood invading fungi implicated grapevine trunk diseases »,
Phytopathologia Mediterranea 40, 485 p.
International Agency for Research on Cancer (IARC),
1987, «Monographs on the evaluation of carcinogenic risks to
humans », Supplement 7, IARC, Lyon (in : Harmandon F.,
2004, « Effets toxiques de l'arsénite
de sodium sur la santé humaine et exposition des utilisateurs »,
Rencontre Technique: Les maladies du bois en Midi-Pyrénées
(Station Régionale ITV Midi-Pyrénées), pp. 8-12)
Jaspers M.V., 2001, <Sensibility of
Phaeomoniella chlamydospora to fungicides in vitro », New Zealand
Plant Protection 54, pp. 225-228
Jousse C., 2004, < La recherche de
molécules ambimobiles pour lutter contre les maladies du bois »,
Rencontre Technique: Les maladies du bois en Midi-Pyrénées
(Station Régionale ITV Midi-Pyrénées), pp. 37
Laberche J.C., 2010, < Biologie
végétale », Ed. Dunod, 305 p.
Larignon P. et Dubos B., 2001-a, < Le Black
Dead Arm. Maladie nouvelle à ne pas confondre avec l'esca Phytoma
538, pp. 26-29.
Larignon P. et Dubos B., 2001-b, < The
villainy of Black Dead Arm », Wines and Vines, pp. 86-89
Larignon P. et Dubos B., 1997, < Fungi
associated with esca disease in grapevine », European Journal of Plant
Pathology 103, pp.147-157
Larignon P. et Vigues V., 2010,
<Identification des facteurs climatiques favorables à
l'expression des symptômes de black dead arm de la vigne »,
Compte-rendu de journée thématique « Les maladies du
bois de la vigne », 16-17novembre 2010, Villefranche/Saône, p.
18
Larignon P., 1991, ((Contribution à
l'identification et au mode d'action des champignons associés au
syndrome de l'Esca de la vigne », thèse doctorat,
université de Bordeaux II, 238 p.
Larignon P., 2004-a, < La constitution
d'un groupe international de travail sur les maladies du bois et les premiers
résultats des expérimentations menées par l'ITV en
laboratoire et en pépinières », Rencontre Technique: Les
maladies du bois en Midi-Pyrénées (Station Régionale ITV
Midi-Pyrénées), pp. 24-27
Larignon P., 2004-b, < Les maladies de
dépérissement : symptomatologies et éléments de
biologie des champignons associés », Rencontre Technique: Les
maladies du bois en Midi-Pyrénées (Station Régionale ITV
Midi-Pyrénées), pp. 15-17
Larignon P., 2004-c, < Réflexions sur
l'Esca », Phytoma 576, pp. 28-31.
Larignon P., 2005, < Les maladies de
dépérissement : le programme national)), Euroviti, p. 47
Larignon P., 2007, < Note nationale : Les maladies du
bois)), rencontre IFV, 5 p.
Larignon P., 2009, < Le point sur les
maladies du bois : symptomatologie, cycles biologiques, influence des facteurs
environnementaux, bilan des recherches et perspectives)), Matinées
Techniques du BIVB : Mieux comprendre les maladies du bois, pp. 1-30
Larignon P., Berud F., Girardon K., Dubos B., 2006,
< Maladies du bois de la vigne. Quelques éléments sur
la présence des champignons associés, leur localisation dans le
bois et les moments de contaminations », Phytoma 592, pp.
14-17
Larignon P., Coustou I., Darné G., Dubos B.,
2003, < Les différences précoces de la composition
anthocyanique foliaire de Cabernet Sauvignon (Vitis vinifera L.)
atteint par l'Esca ou par le Black Dead Arm », (in :
7ème symposium international d'oenologie, Editions Tec
& Doc Lavoisier, pp. 33-35)
Larignon P., Fontaine F., Farine S., Clément C. et
Bertsch C., 2009, « Esca et Black Dead Arm : deux acteurs majeurs
des maladies du bois chez la Vigne », C. R. Biologies 332, pp.
765-783
Larignon P., Fulchic R., Céré L., et Dubos
B., 2000, « Le Black Dead Arm: une maladie à prendre au
sérieux », Union Girondine, pp. 30-31 (in :
Dubos B., 2002, « Maladies cryptogamiques de la vigne »,
Ed. Féret, 160 p.)
Laukart N., Edwards J., Pascoe I., Nguyen N.K., 2001,
«Curative treatments trailed on young grapevines infected with
Phaeomoniella chlamydospora », Phytopathologia Mediterranea
40, S459-S463.
Lebon E., Dumas V., Pieri P., Schultz H.R., 2003,
« Modelling the seasonal dynamics of the soil water balance of
vineyards », Functional Plant Biology 30, pp. 699-710
Lecomte P., Darrieutort G., Defives A., Louvet G.,
Liminana J.M., Blancard D., 2006-a, « Observations of Black Dead
Arm symptoms in Bordeaux vineyards : Evolution of foliar symptoms, localisation
of longitudinal necroses, questions, hypotheses, Integrated Protection in
Viticulture », !OBC/wprs Bulletin 29 (11), pp. 93-94
Lecomte P., Louvet G., Vacher B., Guilbauld P.,
2006-b, « Survival of fungi associated with grapevine decline in
pruned wood after composting », Phytopathologia Mediterranea 45,
pp. 127-130
Lehoczky J., 1974, « Black Dead-arm
disease of grapevine caused by Botryosphaeria stevensii infection »,
Acta Phytopathologica Academiae Scientiarum Hungaricae vol. 9, pp.
319-327 (in : Larignon P., Fontaine F., Farine S., Clément C.
et Bertsch C., 2009, « Esca et Black Dead Arm : deux acteurs
majeurs des maladies du bois chez la Vigne », C. R. Biologies
332, pp. 765-783)
Letousey P., Baillieul F., Perrot G., Rabenoelina F.,
Boulay M., Vaillant-Gaveau N., Clément C., Fontaine F., 2010,
«Early events prior to visual symptoms in the apoplectic form of
grapevine esca disease », Phytopathology, 100-5, pp. 424-431
Magnin-Robert M., Letousey P., Spagnolo A., Clement C.
et Fontaine F., 2010, « Forme apoplectique de l'Esca :
altération précoce de la physiologie de la plante »,
Compte-rendu de journée thématique « Les maladies du
bois de la vigne », 16-17 novembre 2010, Villefranche/Saône, p.
22
Morlat R., 1989, « Le terroir viticole :
contribution à l'étude de sa caractérisation et de son
influence sur les vins. Application aux vignobles rouges de la moyenne
vallée de la Loire », Thèse de Doctorat d'Etat,
Université Bordeaux!!, 289 p.
Morvan G., 2009, « Les maladies du bois
en Bourgogne: résultats des travaux 2005/2008 et projet Bourgogne
2009/2010 », Matinées Techniques du B!VB : Mieux comprendre les
maladies du bois, pp. 35-46
Mugnai L., Surico G., Graniti A., 1999,
« Esca (Black Measles) and brown wood-streaking: two old and elusive
diseases of grapevines », Plant Disease 83-5, pp. 404-418.
Pavlou C.H., 1906, « Lettre à
Ravaz », Progrès Agricole et Viticole 46, pp. 690
(in Surico G., 2009, «Towards a redefinition of
the diseases within the esca complex of grapevine», Phytopathologia
Mediterranea 48, pp. 5- 10)
Payan J.C. et Salançon E. 2002, «
Définition de la contrainte hydrique : incidences sur les
caractéristiques de la vendange », Les Entretiens
Viti-Vinicoles Rhône-Méditerranée, 17 avril 2002,
Gruissan, pp. 6-9
Petit A.N., Vaillant N., Boulay M., Clément C.,
Fontaine F., 2006, «Alteration of photosynthesis in grapevines
affected by esca », Phytopathology 96, pp. 1060-1066
Ravaz L., 1898, « Sur le folletage
», Revue de Viticulture 10, pp. 184-186 (in Surico G.,
2009, «Towards a redefinition of the diseases within the esca
complex of grapevine», Phytopathologia Mediterranea 48, pp. 5-
10)
Reviron A. S., 2010, « Les maladies du
bois : Esca / BDA - Influence de l'alimentation hydrique sur l'expression des
symptômes », Mémoire de fin d'étude (Licence des
Sciences de la Vigne), Institut Universitaire de la Vigne et du Vin de
Dijon, 30 p.
Riou C. et Payan J.C., 2001, « Outils de
gestion de l'eau en vignoble méditerranéen. Application du bilan
hydrique au diagnostic du stress hydrique de la vigne », Compte Rendu
du Groupe Européen d'Etudes des Systèmes de Conduite de la vigne
(GESCO), Montpellier, 3-7juillet, 12ème journée,
pp.125-133
Riou C., Bonhomme R., Chassin P., Neveu A., Papy F.,
1997, « L'eau dans l'espace rural. Production
végétale et qualité de l'eau »,
INRA-Editions, 424 p.
Rioux D., 2011, « Le compartimentage :
un moyen de défense des arbres », L'éclaircie du service
canadien des forêts (Centre de foresterie des Laurentides)
n°65, p. 1
Riou C. et Lebon E., 2000, «Application
d'un modèle de bilan hydrique et de la mesure de la température
de couvert au diagnostic du stress hydrique de la vigne à la parcelle
», Bulletin de l'O.I.V. 73, pp. 755-764
Rolland G., 1873, «Considérations
physiologiques sur l'apoplexie de la vigne », Bulletin de la
Société d'Agriculture de France 5, pp. 539-545
(in Surico G., 2009, «Towards a redefinition of
the diseases within the esca complex of grapevine», Phytopathologia
Mediterranea 48, pp. 5-10)
Rovesti L. et Montermini A., 1987, « Un
deperimento della vite causato da Sphaeropsis malorum diffuso in
provincia di Reggio Emilia », Informatore Fitopatologico 1, pp.
59-61 (in : Larignon P., Fontaine F., Farine S., Clément C.
et Bertsch C., 2009, « Esca et Black Dead Arm : deux acteurs
majeurs des maladies du bois chez la Vigne », C. R. Biologies
332, pp. 765-783)
Scholander P.F., Hammel H.T., Bradstreet E.D.,
Hemmingzen E.A. 1965, « Sap Pressure in Vascular Plants »,
Science 148, pp. 339-346 (In : Tregoat O., Van Leeuwen C.,
Choné X., Gaudillère J.P., 2002, « Etude du
régime hydrique et de la nutrition azotée de la vigne par des
indicateurs physiologiques. Influence sur le comportement de la vigne et la
maturation du raisin », Journal International des Sciences de la Vigne
et du Vin 36, pp. 133-142
Seguin G., 1970, « Les sols de vignobles du
Haut-Médoc. Influence sur l'alimentation en eau de la vigne et sur la
maturation du raisin », Thèse de Doctorat d'Etat,
Université de Bordeaux, 141 p.
Sentenac G., Larignon P., Molot B., Viguès V.,
Kuntzmann P., 2004, « Evaluation de l'efficacité de
fongicides et d'agents biologiques utilisés dans la lutte contre les
maladies du bois Esca et BDA. Premiers résultats
d'expérimentations menées sur le terrain », Rencontre
Technique: Les maladies du bois en Midi-Pyrénées (Station
Régionale ITV Midi-Pyrénées), pp. 28-33
Serrano E., 2001, « Régime hydrique
et minéral de la vigne - Eléments de potentialité d'un
terroir », compterendu V'innopôle - ITV, 6 p.
Shigo A.L., Pottle H.W., Blanchard R.O., 1977,
« Biological control of wood hymenomycetes by Trichoderma
», Plant Disease Report 61, pp. 687-690.
Sibille I, Ojeda H, Prieto J, Maldonado S, Lacapere
JN, Carbonneau A, 2007, « Relation between the values of three
pressure chamber modalities (midday leaf, midday stem and predawn water
potential) of 4 grapevine cultivars in drought situation of the southern of
France », Applications for the irrigation control, Proceedings of XV
International Symposium of GESCO, Porec, Croatia, pp. 685-695
Spinosi J., Févotte J., Vial G., 2009,
« Eléments techniques sur l'exposition professionnelle aux
pesticides arsenicaux. Matrice cultures - expositions aux pesticides arsenicaux
», Institut de veille sanitaire, 22 p.
Soltner D., 2005, « Les bases de la
production végétale - Tome 1 : le sol », Collection
Sciences et Technique 472 p.
Station Régionale ITV Midi-Pyrénées,
2004, « Les maladies du bois en Europe et dans le monde »,
Rencontre Technique: Les maladies du bois en
Midi-Pyrénées, p. 42
Stoll M., Dry P., Loveys B., McCarthy M., 2001,
«Strategic irrigation management in Australian vineyards »,
Journal International des Sciences de la Vigne et du Vin 35, pp.
129-139
Stoll M., Loveys B., Dry P., 2000,
«Hormonal changes induced by partial rootzone drying of irrigated
grapevine », Journal of Experimental Botany 51, pp. 1627-1634
Surico G., 2009, «Towards a redefinition of
the diseases within the esca complex of grapevine», Phytopathologia
Mediterranea 48, pp. 5-10
Surico G., Marchi G., Braccini P., Mugnai L., 2000,
«Epidemiology of Esca in some vineyards in Tuscany (Italy)
», Phytopathologia Mediterranea 39-1, pp. 190-205
Surico G., Mugnai L., Marchi G., 2006,
«Older and more recent observations on esca: A critical review
», Phytopathologia Mediterranea 45 (supplement), pp. 68-86
Tregoat O., Van Leeuwen C., Choné X.,
Gaudillère J.P., 2002, «Etude du régime hydrique et
de la nutrition azotée de la vigne par des indicateurs physiologiques.
Influence sur le comportement de la vigne et la maturation du raisin
»,Journal International des Sciences de la Vigne et du Vin 36,
pp. 133-142
Valancogne C. et Nasr Z., 1989, « Measuring
sap flow in the stem of small trees by a heat balance Method »,
HortScience 24, pp. 383-385
Van Leeuwen C., 2001, « Choix du
cépage en fonction du terroir dans le Bordelais », Journal
International des Sciences de la Vigne et du Vin, N° Hors-série:
« Un raisin de qualité : de la vigne à la cuve »,
pp. 97-102
Van Leeuwen C., Choné X., Tregoat O.,
Gaudillère J.P., 2001-a, « The use of physiological
indicators to assess
vine water uptake and to manage vineyard irrigation
», The Australian Grapegrower and Winemaker 449, pp.
18-24
Van Leeuwen C. et Seguin G., 1994, «
Incidences de l'alimentation en eau de la vigne, appréciée par
l'état hydrique du feuillage, sur le développement de l'appareil
végétatif et la maturation du raisin (Vitis vinifera
variété Cabernet franc, Saint-Emilion, 1990) », Journal
International des Sciences de la Vigne et du Vin 28, pp. 81-110
Van Leeuwen C. et Vivin P, 2008, «
Alimentation hydrique de la vigne et qualité des raisins
», Innovatio Agronomiques 2, pp. 159- 167
Van Leeuwen C., Gaudillère J.P., Tregoat O.,
2001-b, « Evaluation du régime hydrique de la vigne
à partir du rapport isotopique 13C/12C », Journal International
des Sciences de la Vigne et du Vin 35, pp. 195-205
Van Leeuwen C., Lerich O., Renard R., Tregoat O., Alla
P.L., 2000, « Micromorphometric changes in trunk diameter in
relation to mild water stress in field grown vines », Journal
International des Sciences de la Vigne et du Vin 34, pp. 41-47
Van Leeuwen C., Tregoat O., Choné X., Jaeck
M.-E., Rabusseau S., Gaudillère J.-P., 2003, « Le suivi du
régime hydrique de la vigne et son incidence sur la maturation du raisin
», Bulletin O.I.V. vol. 76, n° 867-868, pp. 367 378
Viret O., 2004, « L'Esca en Suisse :
situation en 2001 et évolution 2004 », Rencontre Technique: Les
maladies du bois en Midi-Pyrénées (Station Régionale ITV
Midi-Pyrénées), pp. 46-48
Zufferey et Murisier, 2004, « Etude des
terroirs viticoles vaudois : comportement de la vigne en fonction des
conditions pédo-climatiques », travail de recherche viticole -
Station fédérale de recherches agronomiques de Changins
(Agroscope), 221 p.
Zufferey V., 2007, « Alimentation en eau et
irrigation de la vigne », Revue suisse Vitic. Arboric. Hortic. Vol. 39
(1), pp. 77-78
L'influence de l'alimentation hydrique de la vigne sur
l'expression foliaire des maladies de l'Esca et du Black Dead Arm
RESUMÉ (250 mots)
Les maladies du bois de la vigne sont très dommageables
pour le patrimoine viticole mondial. L'Esca et le Black Dead Arm sont les deux
principaux acteurs impliqués dans cette nouvelle crise sanitaire. Les
agents fongiques qui en sont responsables attaquent des organes pérennes
et provoquent une mort prématurée des ceps, à plus ou
moins long terme. Les symptômes s'expriment au niveau du bois par la
formation de nécroses sectorielles et/ou centrales, par la
présence de bandes brunes ou de chancres, et au niveau foliaire par des
décolorations et des dessèchements qui peuvent être
foudroyants. Les maladies de dépérissements impliquent des
pathosystèmes complexes pour lesquels il est nécessaire d'avoir
une approche globale de la plante dans son environnement. L'alimentation
hydrique est un facteur abiotique important qui conditionne le
développement global de la vigne. Au cours de ce travail, nous avons
tenté d'estimer les niveaux de contraintes hydriques sur des micro-zones
d'étude définies au sein de trois parcelles différentes.
Pour ce faire nous avons réalisé des mesures de potentiel
hydrique foliaire de tige (Øt). Nous avons également eu recours
au modèle Lebon et al. (2003) afin de simuler en continu le
bilan hydrique de la vigne. Les résultats obtenus ont été
confrontés aux relevés de symptômes que nous avons
effectués sur les mêmes sites. Le lien entre l'expression des
maladies et l'alimentation hydrique est difficile à établir.
L'utilisation des cartes de résistivité électrique des
sols permettront à l'avenir de tenir compte de
l'hétérogénéité des sols et de
réaliser des comparaisons plus pertinentes.
Mots clés : Esca ; Black Dead Arm ;
étiologie ; alimentation hydrique ; sol
The influence of water supply of the vine on the foliar
expression of Esca and Black Dead Arm diseases
ABSTRACT (227 words)
Grapevine trunk diseases are very harmful to the wine-growing
heritage. Esca and Black Dead Arm are the two major actors involved in this new
sanitary crisis. The responsible fungi agents attack long-lasting organs and
provoke a premature death of the stocks in the shorter or longer term. The
infection can be diagnosed by the presence in the wood forming tissues of
sectorial and/or central necrosis, by the presence of brown stripes or cankers,
and by discoloration and dryings on the leaves which can occur very fast. Those
diseases imply complex pathosystems for which it is necessary to have a global
approach of the plant in its environment. Water supply is an important abiotic
factor wich determines the global development of the vine. Throughout this
study, we tried to give an estimation of the levels of hydric constraints on
micro-zones defined within three different plots. We thus realized measures of
stem water potential (Øt). We also used the Lebon et al. model
(2003) to simulate continuously the water balance of the vine. The results were
confronted with the symptoms observations which we made on the same sites. The
link between the expression of the diseases and the water supply is difficult
to establish. The use of soils resistivity maps will allow in the future to
take into account the heterogeneousness of soils and to realize more relevant
comparisons.
Keywords : Esca ; Black Dead Arm ; etiology ;
water supply ; soil
| 

