UNIVERSITE AIX-MARSEILLE I - UNIVERSITE DE PROVENCE
UFR Civilisations et Humanités
Département
d'Histoire de l'Art et
d'Archéologie
Mémoire de Master 1
Archéologie et Histoire de
l'Art
Option Préhistoire
|
Etude des dents de caprinés de
l'ensemble moustérien de l'abri Pié
Lombard (Alpes Maritimes)
|
Mémoire présenté par Cindy
CAUSSE
Suivi scientifique assuré par Jean-Philippe Brugal et
Philippe Fernandez Sous la direction de Jean-Pierre Bracco
UNIVERSITE AIX-MARSEILLE I - UNIVERSITE DE PROVENCE
UFR Civilisations et Humanités
Département
d'Histoire de l'Art et
d'Archéologie
Mémoire de Master 1
Archéologie et Histoire de
l'Art
Option Préhistoire
|
Etude des dents de caprinés de
l'ensemble moustérien de l'abri Pié
Lombard (Alpes Maritimes)
|
Mémoire présenté par Cindy
CAUSSE
Suivi scientifique assuré par Jean-Philippe Brugal et
Philippe Fernandez Sous la direction de Jean-Pierre Bracco
Remerciements
En préambule à ce mémoire, je souhaite
adresser ici tous mes remerciements aux personnes qui m'ont apporté leur
aide et qui ont ainsi contribué à l'élaboration de ce
mémoire.
Tout d'abord M. Jean-Philippe Brugal et M.Philippe Fernandez
pour l'aide et le temps qu'ils ont bien voulu me consacrer ainsi que pour leur
bibliographie précieuse, sans qui ce mémoire n'aurait jamais vu
le jour.
Je remercie particulièrement M.Olivier Bignon pour ses
nombreux conseils forts utiles, sa gentillesse et pour l'expérience que
j'ai acquise grâce à lui.
Je tiens aussi à remercier mes amis pour leur soutien et
leur présence.
Enfin je dédie ce modeste travail à ma famille,
et plus particulièrement à mes parents et à ma
grand-mère à qui je dois énormément. Pour leur
soutien sans faille et leur encouragement, merci.
INTRODUCTION ET PROBLEMATIQUES 6
1. PRESENTATION DU GISEMENT 7
1.1 Situation géographique 8
1.2 Historique du site 9
1.3 Etat des recherches 10
1.3.1 Les données géologiques 10
1.3.2 Les analyses paléoenvironnementales 11
1.3.3 Le matériel lithique 12
1.3.4 Datation de l'ensemble moustérien 14
1.4 Contexte régional 15
2. PALEONTOLOGIE : CARACTERISATION GENERIQUE ET
SPECIFIQUE 17
2.1 Caractérisation des générique
et spécifique (bouquetin/chamois/tahr) 18
2.1.1 Le bouquetin 18
2.1.2 Le chamois 19
2.1.3 Le tahr 20
2.1.4 Capra et/ou
Hemitragus ? 21
2.1.4.1 Dentition supérieure 22
2.1.4.2 Dentition inférieure 23
2.2 Détermination des dents de C. ibex
25
2.2.1 Présentation du matériel 25
2.2.2 Critères morphologiques et discriminations
morphométriques 27
2.2.2.1 Distinction entre dentition supérieure et
dentition inférieure chez Capra ibex 28
2.2.2.2 Distinction M1-M2 ;
DP3-DP4 ; P3-P4 et
M1-M2 28
3. ARCHEOZOOLOGIE : STRUCTURE DE LA POPULATION
33
3.1 Taphonomie 34
3.2 Quantification des restes dentaires 36
3.2.1 Nombre de restes (NR) 36
3.2.2 Nombre minimum d'individus (NMI) 37
3.3 Estimation de l'âge 37
3.3.1 Définition de la méthode 37
3.3.2 Les classes d'âges 38
3.3.2.1.1 PHASE I : les juvéniles 39
3.3.2.1.2 PHASE II : les subadultes 40
3.3.2.1.3 PHASE III : les adultes et PHASE IV : les vieux 49
3.4 Estimation de la saison de mortalité
50
3.4.1 NMI de combinaison 50
3.4.2 Les profils de mortalités 51
3.4.3 Ethologie du bouquetin. 54
3.4.3.1 Etude de ses exigences écologiques et
détermination de son environnement 54
3.4.3.2 Etude de son comportement social au rythme des saisons :
d'après des études menées sur des
populations actuelles de bouquetin des Alpes et des
Pyrénées. 55
3.5 Discussion 56
CONCLUSION 59
ILLUSTRATIONS 61
TABLE DES ILLUSTRATIONS 79
TABLE DES ANNEXES 82
ANNEXES 83
BIBLIOGRAPHIE 105
Introduction et problématiques
Le matériel dentaire étudié dans ce
travail provient de fouilles du gisement moustérien de Pié
Lombard dirigées par P.J. Texier dans les années soixante-dix et
quatre-vingt. Les restes fauniques sont conservés dans la réserve
du laboratoire de la MMSH (Maison Méditerranéenne des Sciences de
l'Homme) sous la responsabilité de J.P. Brugal. Cette collection a
déjà fait l'objet d'une analyse paléontologique par J.P.
Gerber (1973), puis archéozoologique par Jean-Philippe Brugal
(carnivores notamment) et C. Diez (taphonomie) ; ces dernières
études ne sont pas publiées.
Ce travail concerne plus précisément
l'étude des restes dentaires des caprinés de l'ensemble
moustérien du site : bouquetin, chamois et tahr sont les trois
espèces qui peuvent être identifiées à Pié
Lombard et qui correspondent avec les cervidés à la famille
dominante dans l'assemblage faunique. Les nombreuses perturbations
sédimentaires liées principalement aux changements climatiques
rendent impossible la mise en évidence de différents niveaux de
dépôts. Pié Lombard présente cependant les
caractéristiques d'un gisement pléistocène à
indices de fréquentation humaine (Brugal, Jaubert, 1991) avec une faible
proportion d'industrie lithique surtout composée d'outils qui attestent
une fréquentation surtout moustérienne, ainsi que des passages
plus récents épipaléolithique. Nous n'avons donc pas tenu
compte des indications stratigraphiques qui ne peuvent correspondre à
une éventuelle évolution de la composition faunique, nous
parlerons d'ensemble moustérien et d'ensemble
épipaléolithique.
Ce travail repose sur deux principaux axes de recherche :
d'abord une partie paléontologique consacrée à la
caractérisation générique et spécifique des
différentes espèces de caprinés présente à
Pié Lombard d'après des critères morphologiques et
biométriques dans le but de préciser la présence ou
l'absence d'Hemitragus dans la collection. Puis une partie
archéozoologique et taphonomique afin de déterminer la structure
de la population étudiée pour l'apport d'informations
complémentaires sur l'origine de l'assemblage qui reste encore
incertaine et la part de l'homme dans l'accumulation osseuse.
Les méthodes développées pour cette
étude reposent avant tout sur l'association d'analyses quantitatives
(métrique, statistique) et qualitatives (table d'usure, stade
d'éruption dentaire, observation des caractéristiques
morphologique) de ces restes.
1. Présentation du gisement
1.1 Situation géographique
L'abri de Pié Lombard est situé dans les
Alpes- Maritimes à environ 2 km au Sud-ouest
de la commune de Tourette- sur-
Loup. La cavité s'ouvre au pied d'une falaise
à environ 200
mètres d'altitude absolue
entre l'actuelle route de Pié Lombard et la
rivière du Loup. Sa
t les
position est assez particulière puisqu'il se situe
entre le rivage méditerranéen à 9 km e chaînons
alpins les plus méridionaux 13 km plus au nord. (fig. 1)
Le gisement
est situé à l'entrée de
l'abri
occupant un fossé de 3,30 m de long sur
un
-
mètre de large environ, bordé par deux
lèvres rocheuses et orienté Nord sud. (Texier, 1996).


Figure 1 : Localisation de l'abri Pié Lombard,
d'après Google Map http://maps.google.fr/ (consulté le 1 juin
2009)
1.2 Historique du site
Le site fut découvert dans les années
soixante par Armand Mellira, un prospecteur amateur.
Suite à cela, un premier sondage, au cours duquel furent
découverts quelques ossements et outils, et qui a
permit à Henri de Lumley (1965) de distinguer
deux stades et deux interstades du Würm ancien,
soit stade isotopique 5, dans le remplissage de Pié
Lombard. Quatre campagnes de fouille dirigées par Pierre Jean
Texier furent réalisées en 1971, 1975, 1982 et 1983.
L'étude des outils réalisée par ce
dernier soutient l'attribution à un gisement
moustérien. De plus la présence d'une
petite série lithique composée de lamelles, de pointes
à dos, de grattoirs prouve aussi le passage plus récent
d'un groupe sans doute épipaléolithique (Texier,
1974) . Quant à la faune, l' étude des grands
mammifères fut d'abord réalisée par J.P Gerber au
début des années 70 (Gerber, 1973) , et
plus récemment par J.P Brugal et C. Diez. Celle des
micromammifères par M. Abbassi et E. Desclaux (1996). Enfin des
analyses complémentaires palynologiques
(Renault-Miskovsky, 1980 ) et anthracologiques (Bazile-Robert,
1979) des niveaux moustériens et
épipaléolithiques viennent renforcer les
données paléoenvironnementales.
1.3 Etat des recherches
1.3.1 Les données géologiques
Le sédiment quaternaire des niveaux
archéologiques est composé d'un cailloutis anguleux
altéré à matrice argilo-sableuse de couleur brun rouge
à brun foncé (De Lumley, 1965). Ce dépôt est aussi
constitué de fragments de stalactites et de plancher stalagmitique. Deux
ensembles de couches on été distingués : l'ensemble I
moustérien avec des niveaux d'accumulations importants et qui rassemble
les couche C4 d, e, e1 et f et l'ensemble II épipaléolithique des
couches C4 a, b, c qui reposent en discordance sur les couches
moustériennes qu'elles ravinent. Une partie de ce dépôt a
subi des remaniements constituant une zone R dans laquelle a été
retrouvé du matériel archéologique sans doute
majoritairement contemporain des niveaux moustérien. (fig. 2)

Figure 2 : Coupe stratigraphique (Texier,
1996)
1.3.2 Les analyses paléoenvironnementales
L'analyse pollinique (Renault-Miskovsky, 1980) a
été réalisée sur les sédiments
moustériens et épipaléolithiques, ainsi que sur les
concrétions formées par le plancher stalagmitique et stalactite
à la base du remplissage dans les couches e et f. Les
échantillons récoltés dans le remplissage
moustérien suggèrent un climat froid et sec pour le OIS 5.
Composé d'un couvert arboréen relativement faible et
principalement constitué par les pins accompagnés de quelques
taxons méditerranéens. En revanche l'étude pollinique des
couches épipaléolithiques indique un couvert forestier un peu
plus fourni, toujours dominé par les pins mais accompagnés
d'espèces thermophiles et méditerranéennes un peu plus
nombreuses.
L'analyse anthracologique (Bazile-Robert, 1979)
effectuée sur les charbons de bois récoltés dans les
parties concrétionnées des couches f et e indique
également un paysage ouvert constitué de Pins sylvestre ainsi que
plus rarement de Chênes et d'Erables. Cette association résulte
sans doute de la situation géographique du site et de ses alentours :
proche du littoral mais soumis aux influences montagnardes.
Le remaniement des sédiments suite à un
phénomène de « suçoir » a compromis
l'étude sédimentologique et archéozoologique couche par
couche qui aurait pu indiquer des variations climatiques plus précises
et permettre une meilleure compréhension de l'origine de l'accumulation
qui, nous le verrons, provient de différentes phases de
dépôts. L'analyse a donc été effectuée sur
les couches de l'ensemble moustérien qui correspond au stade isotopique
5 et à une période plutôt tempérée. Les
résultats de l'étude paléontologique entrepris par J.P.
Gerber (1973) indique une faune à paysage presque exclusivement
forestier
tel que le chat sauvage, le lynx, le cerf ou le sanglier. La
quasi absence du cheval, peut être düà la topographie du
milieu, ainsi que l'absence du renne, animal typique d'un climat froid ;
semblent attester la présence d'une phase relativement
humide du OIS 5, propice au développement forestier. La touche
montagnarde est apportée par la présence de la marmotte, du
bouquetin et du chamois. Cette association faunique ne semble donc pas
conforter les résultats obtenus par les analyses palynologiques. Les
deux espèces d'herbivores dominantes à Pié Lombard
étant le cerf avec 35,4% (data P. Fernandez inédit) et le
bouquetin, deux animaux qui ne fréquentent pas le même biotope. Le
gisement de Pié Lombard, situé à 200 mètres
d'altitude absolue, se trouve à la charnière entre un milieu
ouvert très escarpé constitué de falaises et de plateaux
où régnaient le bouquetin, et un milieu à tendance plus
forestier
situé en fond de vallée et habité par le
cerf . Cette topographie très contrastée forme une sorte de
cloisonnement de ces deux différents biotopes. Le bouquetin souvent
assimilé à un climat froid et aux milieux de haute altitude est
en réalité une espèce plutôt souple sachant
s'adapter à divers environnements et climats.
Enfin la présence de nombreux carnivores (fig.3), soit
12, 7 % sur l'ensemble des espèces de grands mammifères des
couches moustériennes (sur un ensemble de 1760 vestiges du
postcrânien dont 7,6% d'indéterminables taxonomiquement, data J.P.
Brugal inédit), souligne l'éventuelle participation de ces
animaux dans l'origine de l'accumulation osseuse.
1.3.3 Le matériel lithique
Le site de Pié Lombard a livré une faible
quantité de vestiges lithiques qui rendent compte d'activités de
production très limitées qui concerne principalement les phases
de transformation et d'entretien des produits retouchés avec un fort
pourcentage d'outils soit 38,1% (Texier, 1974). Les produits arrivent donc
déjà retouchés ou non dans l'abri, ils sont ou non
modifiés sur place puis abandonnés ou repartent avec les hommes,
ne faisant donc que transiter par l'abri (Porraz, 2004). Le site semble donc
avoir été occupé à plusieurs reprises pendant des
périodes assez brèves. L'industrie est de débitage
Levallois, sur 316 pièces récoltées : 126 outils (surtout
des racloirs), 29 éclats levallois, 24 éclats non levallois, 161
petits éclats et éclats de retouches, 5 nucléus dont 3
réutilisés comme outils (Texier, 1974).
|
Carnivores : 12,7%
- Canidae : Canis lupus (loup)
Vulpes vulpes (renard)
Cuon sp. (dhole)
- Ursidae : Ursus spelaeus (ours des cavernes) ?
Ursus arctos (ours brun)
- Felidae : Felis sylvestris (chat sauvage)
Lynx speleae (lynx)
Panthera pardus (panthère)
- Mustelidae : Meles meles (blaireau)
Artiodactyles :
- Suidae : Sus scrofa (sanglier)
- Cervidae : Cervus elaphus (cerf élaphe) :
35,4%
Dama dama (daim)
- Caprinae : Capra ibex (bouquetin) : 43,2%
Rupicapra rupicapra (chamois) - Bovinae : 1,2 %
Périssodactyles :
- Equidae : Equus caballus (cheval) : 1 reste
- Elephantidae : Elephas sp. (éléphant)
Hominidae :
- Homo neandertalensis : dents isolées
|
Figure 3: Liste faunique des grands mammifères des
couches moustériennes de Pié Lombard, pourcentage basé sur
l'étude du postcrânien (d'après Gerber, 1973 ; Brugal et
Fernandez inédit)
1.3.4 Datation de l'ensemble moustérien
Des datations biostratigraphiques sont fournies d'après
l'étude des dents d'Arvicola (Abbassi ; Desclaux, 1996) à partir
de l'indice d'épaisseur de l'email dentaire des M1. Les travaux
menés sur l'analyse de l'émail du genre Arvicola en Europe
centrale et du nord durant le Quaternaire, ont permis d'établir une
chronologie pertinente des séquences pléistocène
permettant la mise en évidence de variations temporelles et
géographiques.
L'analyse de l'indice d'épaisseur effectuée sur
les quatres individus du genre Arvicola identifiés à Pié
Lombard est comparable à celui des séquences du
pléistocène supérieur en France et en Europe centrale.
Plus précisément, des mesures
radiométriques ont été effectuées sur le plancher
stalagmitique qui se trouve à la base du remplissage et qui se situe
entre 147 Ka #177; 10 Ka et 130 Ka #177; 20 Ka. Trois échantillons de
silex brûlés ont aussi permis d'établir une datation ATL
s'échelonnant de 67.0 Ka #177; 8.1 ; 73.8 Ka #177; 9.6 à 108.4 Ka
#177; 9.8 (Texier, 1996).
La durée de l'accumulation reste encore
indéfinie. Les données paléoenvironnementales, et
notamment paléontologiques avec la présence majoritaire de taxons
typiques d'un climat tempéré, situent l'assemblage durant une
période d'interstade. Dans ses travaux Henri De Lumley (1965) identifie
deux stades et deux interstades entre le stade isotopique 4 et 5, ce qui
correspond à la fourchette de dates obtenues par la datation
thermoluminescence. L'accumulation a donc vraisemblablement eu lieu durant une
période climatique plutôt tempéré,
entrecoupée de relatives phases froides qui n'ont pas réellement
modifiées le paysage ni la composition faunique. On pourrait alors
suggérer une attribution empirique de la durée de l'accumulation
qui se situerait entre le stade isotopique 5c et 4, c'està-dire pendant
une période du Würm relativement clémente.
1.4 Contexte régional
Les gisements archéologiques datant de l'OIS 4 et 5 qui
correspondent au début de la phase glaciaire du Würm sont assez
nombreux dans le Sud est de la France. Cependant la grande majorité de
ces gisements est située en grotte ou cavité ; les forts
ruissellements résultant de l'interstade du OIS 5a, dû à
l'augmentation de l'humidité, ont provoqué une vidange partielle
ou totale de certaines grottes comme à Pié Lombard
(Jourdain-Annequin, 2004).
La situation particulière de la Provence qui
présente un relatif cloisonnement dû à son caractère
montagneux, en a fait une région « refuge » où le
peuplement humain n'a pas connu d'interruption marquée en comparaison
aux régions alpines plus septentrionales. Ce peuplement étant
tributaire des variations climatiques et donc des bouleversements
écologiques. Les Préalpes et les zones de piémont,
où se situe le gisement de Pié Lombard, n'ont pas
été recouvertes par les glaciers qui se sont formés dans
le domaine alpin durant la dernière glaciation. Les installations
moustériennes paraissent nombreuses dans ces zones de Préalpes et
particulièrement durant le stade isotopique 5, période
relativement clémente du début de la glaciation. Malgré
tout le caractère contraignant que représente le milieu
montagneux : caprices du climat et dangerosité de l'espace, la
présence humaine atteste clairement la réalité d'un
attrait spécifique à ces régions, probablement lié
à l'exploitation de ressources particulières. Pié Lombard
étant situé à une altitude de 200 mètres, il peut
être considéré comme un gisement de montagne. L'occupation
du site est attesté pour la période moustérienne,
après aucun indice ne révèlent une activité
anthropique jusqu'au passage épipaléolithique. C'est en effet
à partir du Tardiglaciaire que l'on assiste à un
réchauffement climatique qui engendre la fonte des glaciers. Les
régions montagneuses jusqu'alors timidement visitées apparaissent
comme des espaces neufs et de découverte. Les alpes du nord sont
peuplés en premiers car elles présentent de nombreux avantages
liées à la déglaciation : création de lacs et de
rivières, facteur déterminant dans le choix d'installation
à plus ou moins long terme. Les conditions
paléoécologiques ne sont cependant pas les mêmes entre les
Alpes du nord et du sud. On remarque une densité de peuplement beaucoup
moins forte dans les Alpes du sud durant ces périodes plus
clémentes (Jourdain-Annequin, 2004). Les intenses phases
d'érosion sont sans doute en partie responsables de cette carence, et on
connaît moins bien l'histoire du peuplement du paléolithique
récent dans cette région.
Différents modèles d'exploitations peuvent
être mis en évidence, ils concernent principalement
l'approvisionnement en matériau siliceux et l'exploitation de la
biomasse. A partir de là un modèle reposant sur trois
schémas d'exploitation peut être envisagé (Tillet, 2000).
Les stations de plein-air d'altitude, plutôt liées à
l'approvisionnement en matière première siliceuse ; les gisements
plus ou moins anthropisés à l'entrée des grottes et sous
abri qui semblent attester de stratégies saisonnières
d'exploitation des ressources animales ; et les gisements en cavité
à indice de passage qui sont surtout des gisements
paléontologiques, les rares outils constituant des indices de passages,
ou bien un apport naturel depuis le porche d'entrée de la cavité.
Le gisement de Pié Lombard appartient probablement à l'un de ces
deux derniers modèles.
2. Paléontologie : caractérisation
générique et spécifique
2.1 Caractérisation des générique
et spécifique (bouquetin/chamois/tahr)
L'objectif de cette étude paléontologique est de
déterminer précisément les espèces
présentent à Pié Lombard à l'aide de données
comparatives morphologiques et biométriques. La première
étude du matériel effectuée par J.P. Gerber a
révélé la présence de C. ibex ibex et de
R. rupicapra dans la collection avec une large dominance de
Capra. Pour aller plus loin il serait intéressant de
vérifier la présence d'une autre espèce de capriné
qui a peuplé ces régions durant le pléistocène
moyen, il s'agit d'Hemitragus. Il a été reconnu dans
plusieurs gisements du Sud Est de la France et des Pyrénées : au
Bau de l'Aubesier dans le Vaucluse (Fernandez, 2006), à la grotte des
Cèdres dans le Var (Defleur et al, 1995), à la Caune de L'Arago
dans les Pyrénées Orientales (Rivals, 2004) ou encore à
l'Escale dans les Bouches-du-Rhône (CrégutBonnoure, 1995).
2.1.1 Le bouquetin
En France le bouquetin est représenté par trois
espèces principales : Capra ibex (Bouquetin des Alpes),
Capra pyrenaica (bouquetin des Pyrénées) et Capra
caucasica (bouquetin du Causase) (Crégut-Bonnoure, 2002). On pense
que c'est au cours du stade isotopique 6 que C.ibex
pénètre en Europe en suivant l'arc alpin. On le retrouve en
France à partir de l'interglaciaire OIS 5e (Riss-Würm) où il
s'installe véritablement au cours de l'OIS 5 dans la région
alpine. Durant cette période Capra ibex est identifié
sur l'ensemble des bordures de l'arc Alpin ainsi que dans la Massif Central et
en Italie. A la même période, C. caucasica est aussi
présent dans la région provençale et occupe une aire de
répartition assez large : il est identifié dans le Languedoc, les
Pyrénées Orientales, en Provence et dans la Péninsule
ibérique (Gibraltar). Enfin on peut éliminer l'éventuelle
présence de C. pyrenaica à Pié Lombard, cette
espèce émerge à partir de C. caucasica
praepyrenaica dans les Pyrénées au cours de l'OIS 2
(Crégut-Bonnoure, 2002).
2.1.2 Le chamois
L'arrivée en France de Rupicapra date du
début du Pléistocène moyen au stade isotopique 6,
probablement au même moment que Capra ibex
(Crégut-Bonnoure, 1979). Deux espèces sont identifiées :
R. rupicapra de forme alpine et qui est celle identifiée
à Pié Lombard (Gerber, 1973) et R. pyreneica de forme
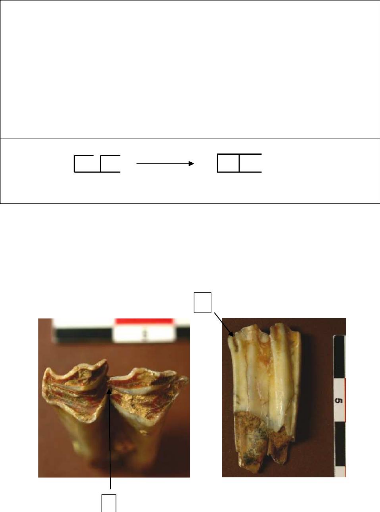
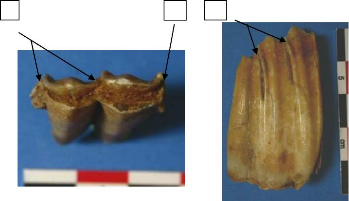
pyrénéenne. Très peu de restes de chamois ont
été identifié à Pié Lombard, le NRDT (nombre
de restes de dents total) est de 40 pour un NMI de fréquence de 6 avec 2
dP4 et 4 M3 (fig. 4 et 5)
|
Dents supérieures
|
Dents en série
|
Dents isolées
|
|
G
|
D
|
G
|
D
|
|
D2
|
0
|
0
|
0
|
0
|
|
D3
|
0
|
0
|
0
|
0
|
|
D4
|
0
|
0
|
0
|
0
|
|
P2
|
0
|
1
|
0
|
0
|
|
P3
|
0
|
1
|
0
|
0
|
|
P4
|
0
|
1
|
1
|
0
|
|
M1
|
0
|
1
|
0
|
1
|
|
M2
|
1
|
0
|
0
|
1
|
|
M3
|
1
|
0
|
0
|
0
|
Figure 4: Répartition anatomique de la dentition
supérieure de R. rupicapra de Pié Lombard.
|
Dents inférieures
|
Dents en série
|
Dents isolées
|
|
G
|
D
|
G
|
D
|
|
D2
|
1
|
0
|
0
|
0
|
|
D3
|
1
|
0
|
1
|
1
|
|
D4
|
1
|
0
|
1
|
1
|
|
P2
|
0
|
0
|
0
|
0
|
|
P3
|
0
|
0
|
0
|
2
|
|
P4
|
1
|
0
|
0
|
1
|
|
M1
|
1
|
0
|
0
|
2
|
|
M2
|
0
|
0
|
0
|
0
|
|
M3
|
0
|
0
|
1
|
4
|
Figure 5: Répartition anatomique de la dentition
inférieure de R. rupicapra de Pié Lombard.
2.1.3 Le tahr
Les deux espèces fossiles qui ont peuplé la
France sont Hemitragus cedrensis et Hemitragus bonali. Au
début du Pléistocène moyen c'est H. jemlahicus
qui migre depuis l'Asie (Tibet) donnant naissance à une nouvelle
espèce H. bonali. La période du Riss constitue une
période de transition avec la disparition d'H. bonali et
l'apparition d'H. cedrensis. C'est au début du Riss III que
l'on identifie Hemitragus cedrensis. Au cours de l'interglaciaire du
OIS 5e on assiste à une raréfaction de cette espèce,
à peu près à la même époque où C.
ibex commence à s'installer dans la région
(Crégut-Bonnoure, 2002). Les deux espèces seraient-elles
rentrées en compétition ? Le stade isotopique 5 marque la
disparition du tahr dans la région provençale et en France. Il
semble donc peu probable que celui-ci soit présent dans l'assemblage du
gisement de Pié Lombard qui se situe durant la période
d'expansion du genre Capra et d'extinction du genre
Hemitragus. Cependant si cette espèce est identifiée
à Pié Lombard il s'agirait vraisemblablement d'H.
cedrensis.
2.1.4 Capra et/ou Hemitragus ?
Nous allons d'abord vérifier à l'aide de
critères morphologiques si Hemitragus est présent dans
la collection. En prenant en compte la situation géographique du site
(les AlpesMaritimes) et la période (OIS 5) pendant laquelle a eu lieu
l'accumulation osseuse, nous allons effectuer une comparaison morphologique
entre Capra ibex et Hemitragus cedrensis.
Il n'est cependant pas impossible que Capra caucasica
est aussi fait partie du cortège faunique de cette région
à cette époque, mais là aussi certains critères de
distinction morphologique permettent d'écarter cette
éventualité. En effet chez C. ibex la P2 est
biradiculée et le métaconide de la P3 dilaté ; alors que
chez C. caucasica la P2 est triradiculée et le
métaconide de la P3 rarement dilatée (Crégut-Bonnoure,
2002). A Pié Lombard toute les P2 sont biradiculée à
l'exception d'une seule dent triradiculée, probablement une
originalité individuelle et le métaconide de la P3 est
dilaté.
En synthétisant les données publiés par
P. Fernandez (2006) et E. Crégut-Bonnoure (1992, 2002, 2005-2006)
concernant les critères morphologiques de distinction entre Capra
ibex et Hemitragus cedrensis, nous sommes en mesure de juger de
l'absence ou de la présence d'Hemitragus dans la collection
(fig. 6 et 7)
2.1.4.1
Dentition supérieure

Hemitragus cedrensis :
P2/
M1/
- Racine en position mésio -linguale plus
développ
é et avance du côté linguale
- Racine de la face distale
peu étendue
M3/
- Dent parfois triradiculée.
- Echancrure vestibulo-
linguale diagonale
marquée (1)
-
Face vestibulaire le parastyle et le
mésostyle dessinent un « V » (3)
- Présence d'un renflement baso-linguale
du lobe distal
- Obliquité baso-linguale
de la surface du
lobe distal marquée
3
Face vestibulaire
Face occlusale
1
Capra ibex :
P2/
M1/
-
Racine linguale en position nettement
mésiale et un peu moins de la moitié du lobe
distal
-
Racine distale volumineuse
M3/
-
Développement important du lobe de l'aile
métastylaire (4)
-
Obliquité de la surface du lobe distal
modérée
- Dent biradiculée.
-
Face mésiale très échancrée
au niveau de la couronne (2)
2
Face linguale
Face linguale
4
Figure 6
: Critères de distinction morphologique entre
Capra ibex et Hémitragus cedrensis pour la dentition
-Bonnoure, 2002 et 2005-2006).
supérieure (d'après Fernandez 2006 et
Crégut
2.1.4.2 Dentition inférieure
|
Hemitragus cedrensis :
|
Capra ibex :
|
|
P/3
-
-
|
Sillon vestibulo- linguale marqué
Présence d'un renflement basal
modéré de
l'entoconide et du paraconide du lobe distal en
face vestibulaire
|
P/3
- Sillon vestibulo- linguale peu profond
- Présence d'un renflement basal sur le
bord
distal marqué
|
|
P/4
-
-
-
-
|
Absence d'un renflement basal du lobe distal
Dilatation du métaconide limitée
Entoconide très peu visible
Sillon distal peu visible
|
P/4
-
-
-
-
|
Présence d'un renflement basal du lobe
distal Dilatation du métaconide
marquée
Dilatation de l'entoconide réduit
Sillon distal large et obliquité marquée
de la surface linguale du lobe distal v ers le milieu
de la dent
|
|
M/1
-
|
Sur la face linguale présence d'un
creusement vers la base de la couronne (1)
1
Face linguale
|
M/1
-
|
Absence de creusement en face linguale
Face linguale
|
|
M/3
-
|
Le bord distal du dernier lobe est étroit
dans sa partie supérieure (2) et présente un
renflement dans sa partie inférieure (3)
2
3
Face vestibulaire
|
M/3
-
|
Le bord distal du dernier lobe est épais
sur toute sa hauteur (4)
4
Face vestibulaire
|
Figure 7
: Critères de distinction morphologique entre
Capra ibex et Hémitragus cedrensis pour la dentition
-Bonnoure, 2002 et 2005-2006).
inférieure (d'après Fernandez 2006 et
Crégut
Les vestiges dentaires de Pié Lombard sont clairement
de type Capra ibex. Notamment d'après l'observation des
particularités morphologiques des P2 et M3 supérieures et des P4,
M1 et M3 inférieures. Les clichés photographiques des tableaux
ci-dessus représentent le matériel dentaire des caprinés
de Pié Lombard. Certains critères ont été plus
discriminants que d'autre, comme par exemple la présence chez Capra
ibex d'une aile métastylaire plus développée sur les
M3, d'un sillon distal plus large sur les P4, ou encore de l'absence
d'un creusement vers la base de la couronne sur la face linguale des M1.
Cependant durant le OIS 5 de part et d'autre du Rhône,
deux sous espèces de C. ibex ont été
identifiées (Crégut-Bonnoure, 1992) : C. ibex cebennarum
et C. ibex ibex. Le premier a été identifié
à l'ouest du Rhône dans les gisements des Pêcheurs
(Ardèche), à la Sartanette (Gard) et à Mialet (Gard) et
présente un morphotype dentaire différent de celui de son
contemporain C. ibex ibex. La différence morphologique se situe
au niveau des molaires avec un raccourcissement des deux premières
molaires et un développement de la troisième chez C. ibex
ibex. Cette différence prouve certainement l'absence
d'échange est-ouest de l'axe Rhodanien. (Crégut-Bonnoure,
2002)
Les comparaisons métriques effectuées entre les
dents de Capra (sur des individus adultes) de l'abri Pié
Lombard et de la grotte des Pêcheurs montrent que la longueur et la
largeur occlusale maximale sont nettement supérieures chez Capra
ibex cebennarum de la grotte des Pêcheurs que chez Capra
ibex de l'abri Pié Lombard pour les dents jugales
définitives supérieures (fig.35 et 36). Pour les dents jugales
définitives inférieures on observe que la longueur occlusale est
supérieure chez Capra cebennarum mais elle est beaucoup moins
marquée que sur les dents supérieures alors que la largeur
occlusale est quasiment comparable avec cependant une très
légère supériorité pour les dents de la grotte des
Pêcheurs (fig. 37 et 38). En d'autre terme, cette comparaison soutient
l'existence à cette époque (OIS 5) de deux morphotypes :
Capra cebennarum qui occupe un territoire à l'ouest du
Rhône et qui, d'après les études morphologiques et
métriques, semble posséder une dentition plus robuste que son
voisin Capra ibex ibex qui évolue à l'Est du
Rhône. Le bouquetin de Pié Lombard est vraisemblablement Capra
ibex ibex.
2.2 Détermination des dents de C. ibex
Après avoir déterminé
spécifiquement les restes dentaires de caprinés du site de
Pié Lombard, et exclu la présence du tahr dans la collection ; la
suite de ce travail aura pour objet d'étude le bouquetin qui est
l'espèce qui domine dans l'assemblage osseux et dentaire.
Les spécificités morphologiques dentaires de
Capra ibex ont déjà été
étudiées et développées dans de nombreux ouvrages
(Couturier, 1962 ; Crégut-Bonnoure, 1992, 2002,2005 ; Fernandez, 2006 ;
Lavocat, 1966).
2.2.1 Présentation du matériel
Le NRDT (nombre de dents total) est de 759 avec 52% (NRD :
398) de dents isolées dont 70% (NRD : 281) de jugales et 30% d'incisives
(NRD : 117). Ainsi que 14% (NDR : 104) de dents en série et un taux de
fragmentation de 33% (NRD : 250) dont 26% (NRD : 65) de fragments
déterminables et 74% (NRD : 185) d'indéterminables. Le travail
qui suit est effectué sur un NRD (nombre de dents déterminables)
de 502 spécimens (dents isolées et en série
déterminables) qui représentent 66% du NRDT de 759.
On observe un déséquilibre entre la
représentation de la dentition supérieure (NRD : 160) et
inférieure (NRD : 225), cette dernière étant mieux
représentée (fig.8). De même qu'entre la
représentation des dents gauches et droites pour les dentitions
inférieure et supérieure (fig.9 et 10). Pour la dentition
supérieure en particulier avec un fort déséquilibre entre
la représentation des séries dentaires gauches (NRD : 8) et
droites (NRD : 35).
Nous avons aussi remarqué que les fragments de
mandibules et de maxillaires, ainsi que les séries dentaires de dents
isolées appartiennent toutes à l'ensemble moustérien ou
à la zone R, vraisemblablement plus moustérienne
qu'épipaléolithique (fig.2).
La méthodologie développée dans ce
travail repose sur l'étude des dents jugales. La détermination et
l'analyse des stades d'usure et d'éruption nous semblent plus
pertinentes sur les dents jugales que sur les incisives. Pour l'ensemble
moustérien nous avons déterminé (ann.18):
|
Incisives
|
Gauche
|
Droite
|
|
déciduales
|
6
|
10
|
|
I1
|
4
|
5
|
|
I2
|
8
|
10
|
|
I3
|
7
|
12
|
|
I4
|
9
|
6
|
|
Total
|
34
|
43
|
|
77
|
Figure 8: Répartition anatomique des incisives de
C. ibex en NRD (nombre de dents déterminées)
|
Dents inférieures
|
Dents en série
|
Dents isolées
|
|
G
|
D
|
Total
|
G
|
D
|
Total
|
|
D2
|
1
|
0
|
1
|
3
|
4
|
7
|
|
D3
|
5
|
2
|
7
|
5
|
4
|
9
|
|
D4
|
5
|
1
|
6
|
12
|
5
|
17
|
|
P2
|
0
|
0
|
0
|
9
|
10
|
19
|
|
P3
|
1
|
4
|
5
|
8
|
7
|
15
|
|
P4
|
3
|
4
|
7
|
8
|
13
|
21
|
|
M1
|
5
|
9
|
14
|
20
|
14
|
34
|
|
M2
|
4
|
8
|
12
|
11
|
15
|
26
|
|
M3
|
3
|
6
|
9
|
8
|
8
|
16
|
|
Total
|
27
|
34
|
61
|
84
|
80
|
164
|
|
225
|
Figure 9: Répartition anatomique de la dentition
inférieure de C. ibex en NRD (nombre de dents
déterminées)
|
Dents supérieures
|
Dents en série
|
Dents isolées
|
|
G
|
D
|
Total
|
G
|
D
|
Total
|
|
D2
|
1
|
3
|
4
|
5
|
5
|
10
|
|
D3
|
2
|
5
|
7
|
5
|
3
|
8
|
|
D4
|
2
|
5
|
7
|
4
|
8
|
12
|
|
P2
|
0
|
4
|
4
|
7
|
1
|
8
|
|
P3
|
1
|
4
|
5
|
7
|
6
|
13
|
|
P4
|
1
|
4
|
5
|
6
|
7
|
13
|
|
M1
|
1
|
3
|
4
|
5
|
13
|
18
|
|
M2
|
0
|
4
|
4
|
7
|
9
|
16
|
|
M3
|
0
|
3
|
3
|
10
|
9
|
19
|
|
Total
|
8
|
35
|
43
|
56
|
61
|
117
|
|
160
|
Figure 10: Répartition anatomique de la dentition
supérieure de C. ibex en NRD (nombre de dents
déterminées)
2.2.2 Critères morphologiques et discriminations
morphométriques
La distinction des différentes dents se fait
d'après les caractères morphologiques spécifiques de
chacune. Dans cette partie seule les dents jugales, c'est-à-dire les
molaires et prémolaires seront prises en compte. Jusqu'à
présent les deux espèces de caprinés
repérées à Pié Lombard sont le bouquetin
(espèce dominante du site avec le cerf) et le chamois
représenté en proportion très faible. Nous allons donc
mener cette étude sur la dentition du bouquetin (Capra ibex
ibex). La formule dentaire définitive de ce dernier totalise trente
deux

dents et peut s'exprimer ainsi : I ~ ~ + C ~ ~ + P ~ ~ + M ~ ~ =
32 dents.
2.2.2.1 Distinction entre dentition supérieure
et dentition inférieure chez Capra ibex
Commençons par la distinction entre les dents jugales
supérieures et inférieures. On reconnaît la molaire ou
prémolaire supérieures par la présence sur la face
vestibulaire de la dent d'une colonnette interlobaire, ainsi qu'un
diamètre vestibulo-lingual beaucoup plus large que sur la jugale
inférieure offrant une section plus quadrangulaire. Pour les dents peu
usées, on remarque sur les molaires inférieures la
présence d'une ouverture en position distolinguale dans la partie
supérieure de la dent. Sur la face mésiale de la molaire
inférieure, la présence du pli caprin beaucoup plus
prononcé que sur la face distale et qui peut servir pour la
latéralisation de la dent. Celui-ci très pincé sur les
dents peu usées, s'élargit en même temps que la surface
occlusale s'aplanie et s'use. Il est beaucoup moins remarquable sur les
molaires supérieures.
2.2.2.2 Distinction M1-M2 ; DP3-DP4 ; P3-P4 et
M1-M2
Nous avons choisis de traiter de manière plus
approfondie la distinction entre certaines dents jugales. Cette distinction se
fait d'abord par discrimination morphologique, les mesures biométriques
(prise sur des spécimens adultes) viennent ensuite appuyer les
résultats obtenus.
- Distinction M1-M2 : on remarque de
manière générale que la M1 est moins hypsodonte que la M2.
Les bords mésiaux et distaux de la M1 ont une tendance plus
marquée à s'élargir et à diverger au fur et
à mesure que l'on s'approche de la surface occlusale de la dent
(Fernandez, 2006). Les bords mésiaux et distaux de la M2 paraissent plus
rectilignes. Mais le caractère le plus distinctif se trouve au niveau de
la limite cément émail, on observe sur le bord mésial de
la M1 une limite beaucoup plus haute que sur le bord distal de la dent, ce qui
permet aussi de distinguer une M1 gauche d'une M1 droite. Cette
particularité ne se rencontre pas sur les M2 (fig.11 et 39)

|
1
|
3
|
7
|
|
M1
|
12
|
13
|
9
|
|
12,88 - 15,94
|
6,83 - 10,12
|
10,87 - 28,56
|
|
14,7
|
8,62
|
21,87
|
|
0,89
|
0,88
|
5,08
|
|
M2
|
6
|
6
|
6
|
|
15,82 - 17,51
|
8 - 9,85
|
23,36 - 36,81
|
|
16,7
|
8,78
|
31,93
|
|
0,77
|
0,71
|
5,08
|
M / 1 M / 2
12 13 14 15 16 17 18
l o n g u e u r
M / 2
M / 1

10
9,5
9
8,5
8
7,5
7
6,5
6
Moyenne hauteur de couronne M/1-M/2
30
25
20
15
10
5
0
M1 M2

35






Figure 11: Dimensions comparées des dents M/1
et M/2 isolées de l'abri Pié Lombard en mm; 1 : diamètre
mesio-distal occlusal, 3 : diamètre vestibulo-lingual occlusale du lobe
mesial, 7 : hauteur de couronne face linguale.
- Distinction DP3-DP4 : le
meilleur critère de distinction se trouve au niveau des
racines, triradiculées. Sur la DP3 la racine
mésiale est large et supporte tout le lobe mésiale de la dent. Le
lobe distal est lui supporté par deux racines plus petites qui se
situent de part et d'autre en face vestibulaire et en face linguale. Sur la
DP4, deux racines en face vestibulaire qui supportent chacune un
lobe. La troisième racine qui tend vers la face linguale et qui supporte
les deux lobes mésial et distal de la dent (Lavocat, 1966). Ce qui fait
que les deux lobes distinctement séparés sur les DP3
au niveau des racines, se rejoignent et convergent à la hauteur du
collet sur les DP4. L'allure générale de la dent s'en
trouve modifiée, les deux lobes de la DP4 rassemblés
par la racine semblent s'étirer vers l'extérieur sur la face
vestibulaire de la dent (fig.12 et 40)
|
1
|
3
|
7
|
|
dP3
|
8
|
6
|
7
|
|
11,93 - 14,19
|
7 - 8,60
|
6 - 10,87
|
|
12,86
|
7,65
|
9,12
|
|
0,75
|
0,72
|
1,85
|
|
dP4
|
12
|
11
|
9
|
|
12,19 - 15,05
|
7,51 - 9,88
|
9,92 - 15,40
|
|
14,14
|
8,74
|
11,73
|
|
0,81
|
0,77
|
1,56
|
dP3/ - dP4/
10
9,5
9
8,5
8
7,5
7
6,5
6

dP3/
dP4/
12 13 14 15 16
l
Moyenne hauteur de couronne dP3/ -
dP4/

14

12
10
8
6
4
2
0
dP3 dP4





Figure 12: Dimensions comparées des dP3/ et des
dP4/ isolées de l'abri Pié Lombard ; 1 : diamètre
mesiodistal occlusal, 3 : diamètre vestibulo-lingual maximal occlusal, 7
: hauteur de couronne face vestibulaire.
- Distinction P3-P4 :
d'une manière générale la P4 est de
couronne plus haute que la P3. Mais il n'existe pas
réellement de critères de distinctions très claires entre
ces deux prémolaires. Nous nous sommes surtout appuyés sur
l'allure générale de la dent suivant son stade d'usure. Mais la
détermination ne s'en trouve pas nécessairement biaisée ;
la P4 étant nettement plus « robuste » que la
P3, notamment avec une couronne beaucoup plus haute ainsi qu'un
diamètre mésio-distal occlusale et vestibulo-lingual (DVL) au
collet plus élevé. La détermination peut s'effectuer en
fonction de ces critères. De plus les données biométriques
sont là pour conforter cette observation (fig.13 et 41)
|
1
|
3
|
7
|
|
P3
|
13
|
12
|
10
|
|
7,91 - 8,95
|
8,08 - 9,10
|
13,06 - 22,27
|
|
8,43
|
8,58
|
16,19
|
|
0,35
|
0,31
|
2,83
|
|
P4
|
8
|
8
|
8
|
|
9,13 - 11,40
|
8,33 - 9,94
|
19,74 - 32,02
|
|
9,98
|
8,93
|
24,86
|
|
0,94
|
0,62
|
4,35
|

P 3! - P 4!
10
9,5
P4/
9
8,5
P3/
8
9 10 11 12
l ongueur
Moyenne hauteur de couronne P3/
- P4/
20
15
10
5
0
P3 P4




Figure 13: Dimensions comparées des P3/ et des P4/
isolées de l'abri Pié Lombard ; 1 : diamètre mesio-distal
occlusal, 3 : diamètre vestibulo-lingual maximal occlusal, 7 : hauteur
de couronne face vestibulaire.
- Distinction M1-M2 :
le principal critère distinctif se trouve au niveau des styles
mésio et disto-lingual. On remarque en effet que les styles de la
M1 ont tendance à diverger du collet vers
l'extrémité libre de la dent (Fernandez, 2006). Alors que les
styles de la M2 restent dans l'ensemble plus rectilignes même si on
observe sur le style mésiolinguale une convexité qui s'accentue
vers le collet. Ce caractère est beaucoup moins prononcé sur les
M1. Enfin de manière plus générale la
M1 est nettement moins haute de couronne que la M2
(fig.14 et 42)

|
1
|
3
|
7
|
|
M1
|
5
|
5
|
4
|
|
14,5 - 15,82
|
10,25 - 12,16
|
13,67 - 23,15
|
|
15,14
|
11,58
|
17,07
|
|
0,51
|
0,78
|
4,27
|
|
M2
|
2
|
2
|
2
|
|
18,15 -
|
|
|
|
18,81
|
11,19 - 11,42
|
30,53 - 32,31
|
|
18,48
|
11,31
|
31,42
|
|
0,47
|
0,16
|
1,26
|
Moyenne hauteur de couronne M1/ - M2/
30
25
20
15
10
5
0
M1 M2
35
M1/ - M2/
14 15 16 17 18 19 20
l o n g u e u r
13
12,5
12
11,5
11
10,5
10
M 2
M1
Figure 14: Dimensions comparée des M1/ et des M2/
isolées de l'abri Pié Lombard ; 1 : diamètre mesio-distal
occlusal, 3 : diamètre vestibulo-lingual occlusale du lobe mesial, 7 :
hauteur de couronne face linguale.
3. Archéozoologie : Structure de la
population
3.1 Taphonomie
L'objectif principal de l'étude taphonomique du
matériel dentaire de Pié Lombard est l'examen de traces
éventuelles d'activités anthropiques. Le matériel dentaire
n'est pourtant pas le support le plus diagnostic pour déterminer le
degré et la fréquence de l'action humaine sur l'assemblage
faunique. Les ossements se prêtent plus volontiers à cette
étude : traces de découpes, types de fracturations, taux de
fracturation, action du feu...
L'activité potentielle qui peut être
repérée sur le matériel dentaire est, entre autre, la
récupération de la moelle osseuse contenue dans le canal
mandibulaire. Dans ce cas on peut observer des traces d'impacts et une
fracturation typique de forme hélicoïdale sur la mandibule. Les
racines des dents sont en générales fracturées au moment
de la percussion. Mais cette activité n'est pas pratiquée de
manière systématique, elle dépend à la fois des
besoins nutritionnels humains, de l'état de santé
de l'animal tué ainsi que du moment oücelui-ci a
été exploité par l'homme. Mais à Pié
Lombard, rien n'indique que les animaux ont
été chassés par les humains puis
transportés à l'abri. Les hommes ont pu profiter de la
présence d'animaux déjà morts pour récupérer
certaines parties encore comestibles.
L'étude des mandibules (11) et des maxillaires (11), ne
révèle pas de traces explicites de fracturation ou de
découpe. La partie incisive du corps mandibulaire n'est jamais
présente et seule une mandibule a été retrouvée
avec sa branche mandibulaire (fig.43). Cette séparation n'est pas
forcément d'origine anthropique, la partie incisive du corps
étant plus cartilagineuse et donc plus fragile. Lorsque la partie
molaire du corps a disparu on ne remarque aucune trace de fracturation due
à une percussion, ni aucun point d'impact. De plus les racines des dents
semblent plus endommagées par des processus post dépositionels
que par une réelle fracturation. Pour les fragments de maxillaires,
seule une partie du processus palatin et du corps maxillaire sont encore
présents. Comme pour les mandibules, l'os incisif a disparu (fig. 44)
Les racines des dents isolées ne portent pas de traces
évidentes de fracturation. Elles sont souvent entières et
semblent indiquer que la dent s'est déchaussée de manière
naturelle (ni fracturation anthropique, ni piétinement). Enfin de
nombreuses séries dentaires ont été retrouvées sans
corps mandibulaire.

Taux de fragmentation du
NRDT
9%
24%
67%
Pour résumé : peu de séries dentaires
retrouvées avec leurs corps mandibulaire ou maxillaire, un assemblage
composé presque exclusivement de dents isolées, des racines peu
ou pas endommagées (fig.45) ; suggèrent une longue exposition
à l'air libre. En effet l'action du weathering (Behrensmeyer, 1978) qui
désigne les altérations liées aux conditions
météorologiques, semble avoir eu un impact sur la conservation du
matériel (fissuration longitudinale). Même si on ne peut pas
estimer de manière précise la durée de l'exposition,
celle-ci a été suffisante pour que le dessèchement du
crâne entraîne le déchaussement des dents.
Une fois la dent isolée de son corps osseux elle
devient plus sensible aux processus post-dépositionels. Le pourcentage
de fragmentation est de 33% du NRDT (nombre de dents total). Il prend en compte
les fragments déterminables et non déterminables. Nous avons
séparé en deux catégories les restes fragmentés.
Une première catégorie regroupent les fragments
déterminables soit 26% du nombre de reste fragmenté et 9% du NRDT
(molaire ou prémolaire, dent inférieure ou supérieure),
avec dans la plupart des cas une séparation de la dent au niveau de la
zone interlobaire, la structure du lobe reste entière ou quasi
entière. La deuxième catégorie est composée des
restes non déterminables soit 74% du nombre de reste fragmenté et
24% du NRDT, dont la structure qui perd sa cohésion se débite en
plaque. Nous avons aussi intégré dans cette catégorie les
fragments de moins de 1cm, qui ne sont pas nombreux (fig.15).
La fragmentation dépend aussi de l'âge et du
stade d'usure. On remarque donc sur le matériel de Pié Lombard un
degré de fragmentation inégale entre les jeunes individus dont la
structure dentaire est plus fragile (pas de racine et cohésion moins
dense entre les lobes due à une faible usure et qui peuvent se
séparer plus facilement) et les adultes possédant une racine
fermée et une structure dentaire plus compacte. Ce biais taphonomique
peut avoir des répercussions importantes pour l'étude des profils
de mortalité. Même si une sousreprésentation des jeunes
individus ne peut pas être uniquement due à un biais taphonomique,
il convient de rester prudent sur les interprétations.
3.2 Quantification des restes dentaires
3.2.1 Nombre de restes (NR)
|
M
|
R
|
E
|
Total
|
|
NRDT
|
759
|
136
|
60
|
955
|
|
NRD
|
502
|
53
|
49
|
604
|
Figure 16: Tableau du NRD et NRDT de l'ensemble du
matériel du gisement de Pié Lombard M : ensemble
moustérien
R : matériel provenant de la zone remaniée qui
appartient aux couches moustériennes ? E : ensemble
épipaléolithique
NRD : nombre de dents déterminables : jugales, incisives
et fragments
NRDT : nombre de restes de dents total avec fragments
indéterminables
Même si les vestiges récupérés dans
la zone R (fig.16) semblent provenir plutôt de l'ensemble
moustérien que de l'ensemble épipaléolithique, seuls les
spécimens de l'ensemble moustérien seront pris en compte dans
cette étude.
3.2.2 Nombre minimum d'individus (NMI)
La première méthode utilisée ici est le
NMI de fréquence. En prenant en compte les M3 et les DP4, tout en
faisant attention à ne pas comptabiliser les M3 en stade
d'éruption car la période de formation de la M3 définitive
coïncide avec le remplacement de la DP4 par la P4 ; nous avons alors
déterminé un NMI f de 29 individus : 17 individus
juvélines (0 à 4 mois) et subadultes (4 à 30 mois) et 12
individus adultes et vieux. Avec 17 DP4 gauches et 12 M3 droites, ainsi que 12
M3 droites.
3.3 Estimation de l'âge
3.3.1 Définition de la méthode
Les méthodes développées pour
l'estimation de l'âge à partir des restes dentaires reposent pour
la plupart sur des observations menées sur des populations domestiques
actuelles (Wilson, 1982 ; Habermehl, 1985 ; Greenfield et al, 2008). Permettant
ainsi l'étude de l'évolution dentaire sur des individus
d'âges connus. On peut ainsi connaître les dates d'éruptions
dentaires et travailler sur les degrés d'usures de chaque dent suivant
l'âge de l'individu. Malheureusement ces caractéristiques ne sont
stables ni dans le temps ni dans l'espace et varient selon plusieurs facteurs.
Par exemple les animaux domestiques soumis à un régime
alimentaire différent ainsi qu'à un mode de vie plus stressant
ont subis des modifications biologiques (génétique et hormonaux)
et morphologiques liés à leurs captivités. On observe par
exemple l'absence chez certain individus domestiques de la P2. C'est pourquoi
nous avons choisi d'établir les classes d'âges à partir
d'études réalisées majoritairement sur des populations
sauvages actuelles (Couturier, 1962) ou fossiles (Perez Ripoll, 1988).
Pour les juvéniles et les subadultes qui
possèdent encore leurs dentitions lactéales nous avons
établi des classe d'âges en fonction du stade d'éruption
dentaire (Couturier, 1962) et du stade d'usure (Perez Ripoll, 1988 ; Rivals
2004). Pour les individus adultes et vieux les classes d'âges sont
déterminées à partir des stades d'usures (Perez Ripoll,
1988 ; Rivals
2004). Cette étude a d'abord été
effectuée sur les fragments de mandibules et de maxillaires du gisement.
Les dents isolées ont ensuite été classées en
fonction de ces résultats.
Chaque classe d'âge est ensuite assimilée
à une phase (de I à IV) qui prend en compte à la fois le
stade d'éruption et le stade d'usure. Ces phases sont elles
divisées en plusieurs états (a, b, c) qui désignent de
manière plus précise les différentes classes d'âges:
pour les juvéniles (phase I) et les subadultes (phase II) ces
états sont déterminés à partir des éruptions
dentaires successives, pour les adultes (phase III) cela correspond à
deux catégories d'usures distinctes séparant les adultes jeunes
des adultes vieillissants. Ce découpage en phase va nous permettre
ensuite d'optimiser le nombre d'individu à l'aide de la méthode
du NMI de combinaison, ainsi que d'évaluer plus
précisément les saisons de mortalité.
Nous allons faire la description de ces phases d'après
les molaires (fig.17). Seulement certaines prémolaires ont pu être
classées par phase car l'étude de leur niveau d'usure est moins
évidente que celle des molaires, elles seront cependant prises en compte
pour le calcul du NMI de combinaison.
3.3.2 Les classes d'âges
FACE
VESTIBULAIRE
Paracône Métacône
|
|
|
Mésostyle
|
|
|
FACE
DISTALE
|
|
|
|
Parastyle
|
Métastyle
|
FACE
MESIALE
|
|
|
|
|
|
|
|
|
1.
Protocône Métoconule
FACE
LINGUALE
métaconide
2.
Figure 17 : Nomenclature des molaires : 1. Molaire
supérieure gauche ; 2. Molaire inférieure gauche
3.3.2.1.1 PHASE I : les juvéniles
Cette phase regroupe les individus
possédant uniquement leurs dentition donc âgé de 0 à
4 mois.
L e décompte a été effectué
à partir des D4 dont la surface occlusale n'est
pas
développée : fermeture des cuspides,
ouverture en position disto-linguale (o) dans la partie
nt la soudure n'est pas achevée
supérieure de la dent et décalage entre les
trois lobes do
(fig.18)

Figure 18: dP/4 en phase I, vue occlusale
3.3.2.1.2 PHASE II : les subadultes
Selon Couturier (1962) les individus dont la M1 est en stade
d'éruption est âgé de 4 à 6 mois et de 5 à 6
mois pour la M1. On observe en effet une éruption plus
tardive de la dentition définitive supérieure de quelques
semaines par rapport à la dentition définitive inférieure.
Jusqu'à 12 mois l'éruption de la première molaire reste le
seul évènement. La M2 n'apparaît qu'à partir du
15ème mois pour la dentition inférieur et au
17ème mois pour la dentition supérieure jusqu'à
24 mois. En résumé un individu âgé de deux ans
possède encore sa dentition lactéale (D2, D3, D4) ainsi que ses
deux premières molaires définitives. Enfin la troisième et
dernière molaire apparaît à l'âge de 28 mois pour les
M3 et de 29-30 mois pour les M3. C'est aussi durant cette
période que s'effectue le remplacement de la dentition lactéale
par la définitive, avec d'abord l'éruption de la P2, puis de la
P3 et enfin de la P4. Passé le 30ème mois l'individu
possède donc toute sa dentition jugale définitive. C'est aussi
à cet âge que l'animal acquiert sa maturité sexuelle. Cette
limite marque le passage à l'âge adulte.
Nous avons donc divisé cette phase en trois états
définis en fonction des stades d'éruptions dentaires qui
correspondent à des âges précis :
- Phase IIa : individus âgés de 4 à 12 mois
et éruption de la M1
- Phase IIb : individus âgés de 15 à 24 mois
et éruption de la M2
- Phase IIc : individus âgés de 28 à 30 mois
et éruption de la M3
Les stades d'usures par lesquels passe une dent en
éruption se révèlent être identiques pour les
M1-M2 et les M1-M2. C'est pourquoi ces
différents stades ne seront détaillés que pour les M1, les
M2 suivant le même processus d'usure.
41
Première molaire
inférieure
Phase IIa : éruption de la M1

des lobes mésia l et distal (o) ainsi
qu'ouverture mésio
couronne (fig.19)
-linguale.
1. Décalage de hauteur et de niveau entre le lobe
mésial et distal.
Début de l'usure du
lobe mésial avec ouverture progressive du
métaconide et du
protoconid e.
Ouverture disto- linguale
linguale du lobe mésial jusqu'à mi hauteur
de la
2. Formation du parastylide et donc fermeture
mésio
Ouverture complète du métaconide et du
protoconide.
4. Ouverture progressive de l'entoconide et de
l'hypoconide.
1 2 3
-
o

Figure 19: fragment de mandibule droite, éruption
de la M/1 en phase IIa, vue occlusale et vestibulaire
Phase IIb : éruption de la M2
1. Ouverture complète de l'entoconide et de
l'hypoconide. L'ouverture disto- linguale (o) des deux lobes
se rétrécie à couronne (fig.20)
Le protoconide et l'entoconide se joignent
(j)
2. Fermeture disto- linguale du lobe
mésial.
la partie haute de la
o
j
L'ouverture disto- linguale du lobe distal est toujours
présente. Formation du métastylide.
1
2
|
Avec la fermeture du lobe distal sur la partie disto-linguale, la
dent est complètement formée (fig.21)
|

Figure 21: M/1 droite en phase IIc, vue
occlusale.
44
Première molaire
supérieure
Phase IIa : éruption de la M1
1. Les deux lobes ne sont pas encore soudés sur
la partie haute de la couronne en face vestibulaire, formant
une fossette (f).
ouverture progressive du métacône et
Début de l'usure du lobe mésial
avec
métaconule.
En position disto-
vestibulaire pas d'ouverture mais la couronne remonte
moins haut formant un sorte de dépression qui
s'étire vers le bas (d).
2. En face linguale, la partie interlobaire n'est pas
soudée (i) (fig.22)
Ouverture complète du métacône et du
métaconule.
Début de l'usure du lobe distal avec ouverture du
paracône et du protocône.
Formation du métastyle.
|


Figure 22
: fragment de maxillaire avec M1/ droite en
éruption, vue linguale et M1/ en phase IIa, vue
occlusale
1. Ouverture complète du paracône et du
protocône
2. La fossette disparaît et les deux lobes se
soudent : formation du mésostyle (m). Formation du
parastyle.
En face linguale la partie interlobaire n'est
pas encore tout à fait soudée (i) (fig.23)
|


1
Figure 23: M1/ droite en phase IIb, vue
occlusale
|
Avec la fermeture en face linguale de la partie interlobaire la
dent est complètement formée (fig.24)
|

Figure 24: M1/ droite en phase IIc, vue
occlusale
1.
Début d'usure du lobe mésial avec
ouverture progressive du métaconide et du
protoconide.
Ouverture en position disto-linguale sur le
lobe
mésial et médian jusqu'à
mi
hauteur de la couronne (o)
Ouverture en position mésio-
linguale sur le lobe mésiale sur la partie
haute
de la couronne (m)
2. Ouverture de l'entoconide et de l'hypoconide du lobe
médian.
3. Ouverture complète de l'entoconide et de l'h
ypoconide du lobe médian ainsi que de l'hypoconulide du
lobe distal.
Formation du métastylide.
Fermeture mésio- linguale du lobe
mésial avec formation du parastylide. L'ouverture
disto-
linguale se rétrécie, d'abord sur le lobe
mésial puis sur le lobe médian où elle
remonte sur la partie haute de la couronne (o) (fig.25)
o
m
o

du métacône et du métaconule.
du paracône et du protocône.
: formation du mésostyle.
Ouverture en position disto-
vestibulaire du lobe distal, qui descend jusqu'à l'a
ile
métastylaire (o) (fig.26)
2. Ouverture progressive
Ouverture complète
4. La fossette disparaît et les deux lobes se
soudent
o
f
1. O uverture progressive du paracône et du
protocône. Présence de la fossette interlobaire
en face vestibulaire (f)
1.
2.
3.
3.3.2.1.3 PHASE III : les adultes et PHASE IV : les vieux
La phase III est aussi divisée en deux états:
- Phase IIIa : les adultes jeunes, âgés de plus de
30 mois
- Phase IIIb : les adultes vieillissants
Cependant ces états ne permettent pas d'établir
une classe d'âge précise comme pour les phases I et II mais de
distinguer les adultes jeunes des adultes vieillissants en fonction du
degré d'usure de la dent et de comparaisons métriques.
Les critères de déterminations pour les phases III
et IV étant identiques, elles seront traitées dans une même
partie.
Tout d'abord les critères morphologiques liés au
stade d'usure dentaire sont déterminés à partir de
l'observation des racines, du degré d'usure des cuspides et de
l'ouverture et de l'usure de la fossette (fig.27, 47 et 51). Nous avons ensuite
établi des comparaisons métriques pour chaque molaire selon
chaque phase (fig. 46, 48, 49, 50, 52 et 53)
|
Racine
|
Cuspides
|
fossettes
|
|
Phase II
|
Pas de racine, la dent est ouverte au collet
|
saillantes
|
Ouvertes et saillantes
|
|
Phase IIIa
|
Formation de la racine
|
Légère usure
|
Ouvertes et légèrement usée
|
|
Phase IIIb
|
Racine formée
|
Forte usure, aplanies
|
Fermées et fortement usées
|
|
Phase IV
|
Racine formée
|
Très forte usure, les cuspides sont complètement
polies
|
Très forte usure jusqu'à disparition de la
fossette
|
De manière générale c'est aux niveaux du
diamètre mesio-distal au collet et de la hauteur de couronne que
l'écart des dimensions est le plus important. En effet le degré
d'hypsodontie marqué chez les jeunes individus, s'attenue au fur
à mesure que l'animal vieillit, la hauteur de couronne diminue donc
proportionnellement à l'usure et devient donc un critère de
distinction majeur. L'usure qui aplanie la surface occlusale de la dent modifie
son aspect général (fig.27), et élargit la dent
particulièrement au niveau de la partie haute de la couronne. Les
diagrammes suggèrent en effet clairement une augmentation du
diamètre mésio-distal et de manière plus aléatoire
une augmentation du diamètre vestibulo-lingual.
3.4 Estimation de la saison de mortalité
3.4.1 NMI de combinaison
Ce NMI de combinaison est donc obtenu à partir des
critères d'âges que nous avons développés dans la
partie précédente. Cette méthode permet à la fois
d'optimiser le nombre d'individu de fréquence ainsi que
d'appréhender le nombre d'individu par classe d'âge afin d'estimer
les saisons de mortalités. Certains spécimens n'ont pu être
classés dans une phase précise, notamment ceux de la phase III.
En réponse à ce problème nous avons choisi de
pondérer les phases (fig.28 et 29) afin d'augmenter le NMIc. On obtient
ainsi un NMIc de 36 (fig.30).
|
INF
|
dP2
|
dP3
|
dP4
|
P2
|
P3
|
P4
|
M1
|
M2
|
M3
|
NMI
|
|
Juvéniles
|
Phase I
|
-
|
2/0
|
2/1
|
-
|
-
|
-
|
-
|
-
|
-
|
2
|
|
Subadultes
|
Phase IIa
|
-
|
1/2
|
1/2
|
-
|
-
|
-
|
3/1
|
-
|
-
|
3
|
|
Phase IIb
|
1/0
|
1/0
|
1/1
|
-
|
-
|
-
|
4/3
|
1/3
|
-
|
4
|
|
Phase IIc
|
-
|
-
|
-
|
-
|
0/2
|
1/1
|
0/2
|
1/7
|
2/2
|
7
|
|
Adultes
|
Phase IIIa
|
-
|
-
|
-
|
-
|
1/2,5
|
5/9
|
9/9
|
7/4
|
6/2
|
9
|
|
Phase IIIb
|
-
|
-
|
-
|
-
|
6/5,5
|
3/5
|
6/4
|
4/4
|
2/5
|
6
|
|
Vieux
|
Phase IV
|
-
|
-
|
-
|
-
|
1/1
|
1/1
|
1/2
|
2/3
|
1/2
|
3
|
Figure 28: tableau de répartition de la dentition
inférieure par classe d'âge et par phase et nombre minimum
d'individu par phase
|
SUP
|
dP2
|
dP3
|
dP4
|
P2
|
P3
|
P4
|
M1
|
M2
|
M3
|
NMI
|
|
Juvéniles
|
Phase I
|
-
|
1/0
|
1/0
|
-
|
-
|
-
|
-
|
-
|
-
|
1
|
|
Subadultes
|
Phase IIa
|
1/1
|
1/1
|
1/1
|
-
|
-
|
-
|
4/4
|
-
|
-
|
4
|
|
Phase IIb
|
0/1
|
0/1
|
0/1
|
-
|
-
|
-
|
0/2
|
0/3
|
-
|
3
|
|
Phase IIc
|
-
|
-
|
1/0
|
-
|
-
|
-
|
-
|
1/2
|
-
|
2
|
|
Adultes
|
Phase IIIa
|
-
|
-
|
-
|
3,5/2,5
|
3,5/5
|
3/5,5
|
1/1
|
2/6
|
5/5
|
6
|
|
Phase IIIb
|
-
|
-
|
-
|
3,5/2,5
|
3,5/5
|
3/4,5
|
0/7
|
2/1
|
1/5
|
7
|
|
Vieux
|
Phase IV
|
-
|
-
|
-
|
-
|
-
|
-
|
1/2
|
1/1
|
2/2
|
2
|
Figure 29: tableau de répartition de la dentition
supérieure par classe d'âge et par phase et nombre minimum
d'individu par phase
|
JUVENILES
Phase I
|
SUBADULTES
|
ADULTES
|
VIEUX
Phase IV
|
Total
|
|
Phase IIa
|
Phase IIb
|
Phase IIc
|
Phase IIIa
|
Phase IIIb
|
|
NMI c
|
2
|
4
|
4
|
7
|
9
|
7
|
3
|
36
|
|
NMI c %
|
6
|
11
|
11
|
19
|
25
|
19
|
8
|
|
Figure 30 : tableau récapitulatif du NMIc pour
chaque phase
3.4.2 Les profils de mortalités
Le profil de mortalité permet d'estimer la structure
d'âge d'une population abattue et d'en déduire d'éventuelle
préférence de chasse (Lyman, 1984). Pour caractériser la
mortalité d'une population on utilise différents types de profils
théoriques :
- Le profil de type catastrophique (Lyman, 1987) qui
caractérise la structure d'une population vivante dans lequel le nombre
d'individu dans les classes d'âges successives est décroissant. Ce
type de profil peut se rencontrer lorsqu'une population a été
décimée en masse par exemple suite à un
évènement catastrophique comme une éruption volcanique, un
incendie ou peu refléter le tableau de chasse d'un prédateur ou
l'abattage en masse d'individus au cours d'un épisode de chasse.
- Le profil de type attritionnel (Lyman, 1987) avec une
sur-représentation des jeunes et des individus âgés
caractéristique d'une courbe de mortalité naturelle suite aux
maladies ou à la malnutrition. Dans un contexte
archéologique ce type de profil peut indiquer une chasse orientée
sur les individus les plus faibles.
- Le profil de sélection des adultes typique «
force de l'âge » (Stiner, 1991) dans lequel les classes de jeunes et
d'individus âgés sont sous-représentées. En contexte
archéologique ce type de profil peut suggérer une chasse
sélective axée sur les individus adultes.
Cependant l'utilisation de ces profils théoriques sur
un assemblage fossile peut poser quelques problèmes d'ordre
méthodologiques notamment liés à la conservation
différentielle des vestiges dentaires. En ce qui concerne la collection
du gisement de Pié Lombard nous avons remarqué que les fragments
de dents non déterminables appartiennent en grande partie à de
jeunes individus, ce qui peut avoir comme conséquence une sous
représentation de cette classe d'âge. De plus, la population
étudiée provient d'un ensemble de couches dans lesquelles il
n'est pas possible de distinguer plusieurs épisodes de
dépôts. Les profils reflètent donc une globalité
à travers laquelle nous allons tenter de distinguer l'origine de
l'accumulation : acquisition humaine, carnivore ou lieu de mort naturelle.
|
Juvéniles
|
Subadultes
|
Adultes
|
Vieux
|
|
NMIc
|
2
|
15
|
16
|
3
|
|
NMIc %
|
6
|
42
|
44
|
8
|
|
50 40 30 20 10 0
|
|
|
|
|
|
|
% NMIc
|
Juvéniles Subadultes Adultes Vieux
Figure 31: pourcentage du NMIc par classe
d'âge
30
Stade I Stade IIa
Stade IIbStade IIc
Stade IV
Stade
IIIa
Stade
IIIb
JUVENILES SUBADULTES
ADULTES
VIEUX
25 20 15 10 5 0
Figure 33: profil de mortalité d'après le
NMIc par phase et par classe d'âge
Nous remarquons donc ici un profil de type sélection
des adultes qui peut être typique d'une acquisition sélective des
animaux dans la force de l'âge (fig 30 à 33). Néanmoins ce
résultat ne peut clairement illustrer une acquisition biologique du
gibier. En effet la prédominance des subadultes et adultes par rapport
aux jeunes et aux individus âgés regroupent une large
catégorie d'âge qui correspond aux conditions naturelles de
mortalités (Lyman, 1984).
De plus ce profil ne rend pas en compte des différents
modes d'agrégations des groupes : groupes familiaux et groupes de
célibataires (Bignon, 2008). La composition des hardes diffère
aussi selon les saisons. Un résumé des exigences
écologiques et du comportement social de bouquetin peut contribuer
à une meilleure compréhension de ces profils de
mortalité.
3.4.3 Ethologie du bouquetin.
3.4.3.1 Etude de ses exigences écologiques et
détermination de son environnement
« Le rôle fondamental de l'altitude est un «
préjugé issu d'un artefact », sa présence
dépend de celle de rocher, du bord de la mer jusqu'aux plus hautes
cimes, les hautes altitudes étant considérées comme des
secteurs marginaux, dans lesquelles les bouquetins ont été
repoussés » (Raye, 1994)
En effet, en tentant de définir les exigences
écologiques du bouquetin; on se rend compte que cet animal aujourd'hui
symbole du milieu de haute montagne, est en réalité parfaitement
adapté à des milieux de plus basse altitude et à des
conditions écologiques différentes, pourvu qu'il y ait des
rochers (étage subalpin et alpin, gorge calcaire, bord de mer). En fait
la structure molle et adhérente de ces sabots en fait l'animal rupicole
par excellence (Couturier, 1958)
L'espérance de vie du bouquetin est de vingt ans, la
sénescence apparait vers l'âge de quinze seize ans. C'est alors
que l'individu a tendance à s'isoler du reste du groupe. Les bouquetins
adultes et les femelles suitées forment des hardes distinctes. C'est
à partir de trois ans que les mâles sont rejetés par leurs
mères et trouvent refuges auprès des hardes de mâles
célibataires.
Dès mi novembre les mâles entrent en rut. La
maturité sexuelle est atteinte vers l'âge de deux ans mais les
possibilités de reproduction ne sont offertes qu'à partir de
trois ans. Durant cette période on assiste à un rassemblement des
hardes, femelles/cabris et mâles, ainsi qu'à de nombreux combats
de mâles. Début janvier marque la fin du rut mais aussi le
début de l'hiver alors que les animaux sont affaiblis. La gestation dure
de cinq à six mois, les femelles sont alourdis et fatiguées par
les aléas hivernaux. Les naissances se font à partir de
début juin jusqu'à début juillet. Une étagne ne met
bas que d'un seul cabri tous les deux ans et les jumeaux sont rares (Couturier,
1962 ; Toigo et al, 1996).
3.4.3.2 Etude de son comportement social au rythme des
saisons : d'après des études menées sur des populations
actuelles de bouquetin des Alpes et des Pyrénées.
Des biologistes et zoologues (Gonzalez, 1994 ; Raye, 1994 ;
Toigo et al, 1996) se sont intéressés au comportement saisonnier
du bouquetin afin de définir ses préférences
écologiques. On remarque d'une façon générale qu'au
printemps l'habitat du bouquetin est plus bas : besoin de nourriture, mise
à bas, fatigue et danger d'avalanche. C'est à cette
période que l'on peut les rencontrer à la lisière des
forêts. Cependant ils ne s'aventurent que très rarement dans ses
environnements fermés préférant les espaces ouverts et
escarpés, contrairement aux chamois. Durant l'été le
bouquetin a tendance à remonter vers les hauteurs à la recherche
de fraicheur. Bien qu'habitué à fréquenter les
environnements montagneux, l'hiver reste une saison très difficile pour
cette espèce avec la menace du vent et du verglas. Le manque de
nourriture et le verglas obligent les bouquetins à descendre à
des altitudes plus basses et à trouver refuge sur les pentes
dégagées. Il arrive parfois que les hardes se réfugient
dans des cavités rocheuses afin de s'abriter du vent, faisant d'elles
des proies vulnérables. On peut penser que ce comportement
conditionné en grande partie par l'environnement et le climat est une
réponse d'ordre biologique et peut être pris en compte pour
appréhender les choix effectués par les chasseurs
paléolithiques. Même si le facteur humain a eu des impacts au
niveau de l'étendue du territoire occupé par les bouquetins. En
atteste la présence de cet animal dans des gisements
archéologiques situés en périphérie des zones
montagneuses.
En règle générale, le bouquetin n'est pas
un animal très farouche. Il est possible de s'en approcher de
très près lorsque celui-ci se repose au soleil par exemple. En
revanche les femelles sont plus craintives lorsqu'elles sont
accompagnées de leurs cabris. Il reste cependant très important
pour le chasseur de connaitre le milieu et le comportement de l'animal à
chasser. La chasse à l'affut ou à l'approche sont deux tactiques
qui se prêtaient sans doute bien à la chasse au bouquetin.
3.5 Discussion
L'étude de l'estimation de l'âge des individus
indique que tous les stades d'éruptions sont présents dans la
série dentaire de Pié Lombard. Pour chaque stade une fourchette
d'âge a été déterminée (Couturier, 1962)
à partir des mois de naissance de juin - juillet (fig.34). On remarque
ainsi que toutes les saisons sont représentées dans l'assemblage.
Cela indique que plusieurs épisodes de dépôts ont eu lieu,
mais ne nous renseigne pas sur l'origine ni la durée de l'accumulation.
L'étude de la saisonnalité de l'ensemble moustérien qui
rassemble plusieurs niveaux de dépôts ne nous éclaire pas
sur l'origine de l'accumulation osseuse car elle ne permet pas la mise en
évidence d'un comportement d'acquisition saisonnier.
|
Classe d'âge
|
0 - 4 mois
|
4 - 12 mois
|
15 - 24 mois
|
28 - 30 mois
|
|
Mois
|
Juin - octobre
|
Octobre - juin
|
Septembre - juin
|
octobre - décembre
|
|
Saison
|
Eté -automne
|
Automne - hiver - printemps
|
Automne - hiver- printemps
|
Automne - hiver
|
|
NMIc
|
2
|
4
|
4
|
7
|
Figure 34: NMIc des juvéniles et des subadultes en
relation avec les mois et saisons (d'après Couturier, 1962)
Nous avons vu que la sous-représentation des classes
d'individus jeunes et âgés peut être liée à
des problèmes méthodologiques et taphonomiques. Néanmoins
si les vieux individus sont si peu représentés c'est peut
être aussi parce qu'ils ne fréquentent plus les hardes familiales
et de célibataires, choisissant une vie solitaire. En supposant que
l'accumulation soit d'ordre biologique humain ou carnivore, l'acquisition
aurait donc été axée plutôt sur les hardes que sur
des individus faibles et isolés. On peut même aller plus loin si
on prend en compte la nombre important d'individus subadultes qui n'ont pas
encore atteint leurs maturités sexuelles qui composent les hardes
familiales de femelles suitées, et qui pourrait alors suggérer
une chasse préférentielle de ces groupes. Cependant au vu des
nombreuses contraintes liées à la nature de l'accumulation, ces
interprétations ne peuvent rester que dans le domaine du
théorique.
L'origine de l'accumulation reste donc incertaine. Elle peut
être naturelle, les animaux peuvent avoir été pris au
piège à cet endroit puis exploités dans un second temps
par les carnivores et les humains. Nous avons l'exemple à la grotte de
l'Hortus (Hérault) datée de l'OIS 3 qui présente un
assemblage faunique très riche en bouquetin avec un NR (nombre de reste)
de 1766 qui représentent 103 individus (Pillard, 1970). Certaines
couches présentent un assemblage de squelettes de bouquetins complets et
en places qui indique que ces animaux sont morts naturellement dans un
piège. Cependant ces squelettes ne présentent aucune trace d'une
exploitation postérieure par les hommes ou par les carnivores. En
Ardèche, la grotte des Pêcheurs (paléolithique moyen et
supérieure) présente aussi un assemblage très riche en
bouquetin (Capra ibex) avec 1536 restes identifiés et un NMI de
39 individus et en carnivores (320 restes) principalement composé de
loup et d'hyène (Moncel et al, 2008). Les traces d'une exploitation par
les hommes (présence d'une zone de combustion et d'une industrie
lithique) et par les carnivores (traces de crocs, absence de petits os comme
les carpes et tarses) posent la question de l'origine de l'accumulation.
L'étude de la topographie de la grotte et de ces alentours
complétée par l'analyse archéozoologique des restes
d'herbivores permet d'avancer l'hypothèse d'un piégeage naturelle
comme à la grotte de l'Hortus. Les restes de bouquetin ont
été retrouvés dans ce qui semblait être une sorte
d'aven (secteur 4 : paléolithique moyen) avec toutes les parties du
squelette représentés. Nous avons vu que parfois les hardes se
réfugient durant les intempéries dans des abris rocheux, à
la grotte des Pêcheurs les bouquetins du secteur 4 ont pu vouloir se
réfugier dans cette cavité et s'aventurer trop
profondément dans la grotte au point de se retrouver
piégés. Les hommes et les carnivores auraient donc eu
accès secondairement à ces carcasses. Les restes de carnivores
appartiennent majoritairement à de vieux individus qui se sont
probablement isolés pour mourir en paix. La grotte ne semble pas avoir
servie de tanière. La présence humaine est attestée par la
présence d'une zone de combustion et de rares vestiges lithiques,
ceux-ci ont pu aussi trouver refuge dans la grotte. Ces deux gisements
présentent quelques similitudes avec l'abri Pié Lombard. Ils sont
situés à environ 200 mètres d'altitude dans un
environnement très escarpé composé de plateaux et de
falaises, idéal pour les bouquetins qui domine dans l'assemblage osseux
; les vestiges lithiques sont peu nombreux et surtout composés d'outils.
Cependant à Pié Lombard l'exploitation des ossements d'herbivores
par les hommes et les carnivores semblent plus intense et la présence du
cerf dans l'assemblage (35,4%) peut sous entendre une acquisition biologique du
gibier. En effet si on considère la position géographique de la
grotte et sa difficulté d'accès, il est peu probable que ce lieu
ait été visité par les cerfs, leurs présences
serait donc du à une acquisition biologique ? Cela ne prouve pas
de manière évidente que les agents biologiques
sont seuls responsables de l'accumulation osseuse, mais semblent jouer un
rôle plus important que dans la grotte de l'Hortus et des
Pêcheurs.
Une étude de la représentation anatomique du
squelette du bouquetin et du cerf pourrait apporter des informations
complémentaires sur ce sujet, elle pourrait mettre en évidence
d'éventuelles stratégies de transports ou l'exploitation
différentielle des carcasses. Ce qui constituerait un témoin de
la contribution humaine dans l'accumulation osseuse. Les restes dentaires de
capriné n'apportent pas de réelles informations quant au
rôle et à l'impact humain sur l'assemblage. L'étude
archéozoologique du postcrânien : représentation
squelettique, étude de la fragmentation des os longs et des fractures,
des traces de découpes, des traces de crocs etc. pourrait nous
éclaircir sur la contribution humaine et carnivore concernant l'origine
de l'assemblage et la fréquence de l'exploitation.
Conclusion
Rupicapra rupicapra et Capra ibex ibex sont
les deux espèces identifiées à Pié Lombard.
D'après l'étude des caractères morphologiques distinctifs
nous avons démontré l'absence d'Hemitragus cedrensis
dans la collection. La période de l'OIS 5 dans laquelle se situe le
gisement de Pié Lombard est celle d'une transition qui correspond
à l'augmentation de la population de Capra ibex et à la
disparition d'Hemitragus cedrensis dans le Sud est de la France. Le
bouquetin (espèce dominante avec le cerf dans l'assemblage faunique de
Pié Lombard) est une espèce essentielle de la faune de l'OIS 4 et
5 du Sud Est de la France où il a trouvé un milieu favorable
à son développement : un climat froid et sec dans une
région rocheuse située entre la mer méditerranée et
le domaine alpin. Ainsi on le retrouve très souvent dans les assemblages
fauniques des gisements moustériens du Sud Est de la France où il
domine comme à l'Hortus, à la grotte des Pêcheurs et
à Pié Lombard. Ces deux premiers sites étant d'avantage
des gisements paléontologiques. L'impact humain sur l'assemblage
faunique de Pié Lombard semble plus important même si la
fréquence, la durée et le degré de cet impact ne sont pas
encore définis.
Il n'y a pas de traces évidente d'une activité
anthropique sur les vestiges dentaires des caprinés: pas de
fracturations typiques d'une activité de récupération de
la moelle osseuse, pas de traces de boucheries et un fort pourcentage de dents
isolées possédant encore leurs racines indiquant qu'elles se sont
déchaussées suite à l'action du weathering. Nous n'avons
pas non plus observé de traces de dents de carnivores sur les fragments
de mandibules et de maxillaires, le rôle de ces animaux dans
l'accumulation osseuse ainsi que dans le traitement des carcasses est à
considérer puisque ce type de traces s'observent sur les ossements de
Pié Lombard.
L'étude de la structure de la population de
capriné, menée ici sur les vestiges dentaires doit être
complétée par l'analyse archéozoologique du post-cranien
des caprinés mais aussi par celle des cervidés, afin d'apporter
des informations plus précises quant au rôle de l'homme dans
l'assemblage. Nous rappelons cependant qu'en raison des nombreuses
perturbations géologiques qui rendent impossible une lecture
stratigraphique précise, l'étude de la structure de la population
ne peut indiquer une éventuelle évolution dans la composition
faunique qui est considérée dans son ensemble. Et donc la mise en
évidence de
comportements distinctifs d'acquisition et de traitement des
carcasses par divers agents biologiques.
Enfin pour conclure nous pouvons ajouter que l'exploitation
des milieux d'altitudes définis par une topographie et un biotope
particuliers témoigne d'un comportement économique singulier et
de l'attrait spécifique de cet environnement qui semble tenir une place
tout à fait originale au sein de l'organisation territoriale. Sans doute
lié à la fois à des activités
cynégétiques, comme cela semble être le cas à
Pié Lombard, et à la recherche de bonne matière
première siliceuse. Car même si nous ne somme pas encore en mesure
de définir l'origine de l'accumulation osseuse, l'occupation ponctuelle
de ce gisement semble néanmoins liée à des
activités de boucherie.
Illustrations
|
P2/
|
Pié Lombard
|
Pêcheurs (Crégut, 2002)
|
|
1
|
11
|
1
|
|
5,42 - 7,31
|
|
|
6,77
|
7,5
|
|
0,54
|
|
|
3
|
9
|
1
|
|
6,13 - 7,20
|
|
|
6,66
|
6,9
|
|
0,37
|
|
|
P3/
|
|
|
|
1
|
18
|
3
|
|
7,78 - 9,04
|
8,5 - 9,8
|
|
8,38
|
9,26
|
|
0,36
|
0,68
|
|
3
|
18
|
3
|
|
7,98 - 9,10
|
8,6 - 9,6
|
|
8,56
|
9,16
|
|
0,31
|
0,51
|
|
P4/
|
|
|
|
1
|
17
|
9
|
|
7,47 - 14,16
|
9 - 12,7
|
|
10,01
|
10,62
|
|
1,57
|
1,14
|
|
3
|
17
|
8
|
|
6,96 - 10,77
|
9,1 - 10,8
|
|
8,90
|
10,13
|
|
1,07
|
0,56
|
|
M1/
|
|
|
|
1
|
9
|
15
|
|
13,11 - 16,46
|
16 - 19
|
|
14,94
|
17,90
|
|
1,20
|
0,99
|
|
3
|
8
|
12
|
|
9,78 - 12,16
|
10 - 13,7
|
|
11,38
|
12
|
|
0,88
|
1,26
|
|
M2/
|
|
|
|
1
|
11
|
14
|
|
16,87 - 19,41
|
17 - 21,2
|
|
18,05
|
19,60
|
|
0,73
|
1,15
|
|
3
|
11
|
14
|
|
9,73 - 12,76
|
12,1 - 14,2
|
|
11,25
|
13,22
|
|
0,87
|
0,71
|
|
M3/
|
|
|
|
1
|
11
|
26
|
|
16,24 - 22,61
|
17 - 26,5
|
|
19,75
|
21,08
|
|
2,29
|
2,37
|
|
3
|
14
|
23
|
|
9,15 - 12,34
|
11,7 - 15,6
|
|
10,74
|
13,45
|
|
1,04
|
1,13
|
Figure 35: Dimensions comparées de Capra ibex
cebennarum de la grotte des Pêcheurs et de Capra ibex de
Pié Lombard pour la dentition supérieure, d'après la
longueur (1) et la largeur maximale (3) occlusale.
63

P3/
Pêcheur
Pié Lombard
8 8,5 9 9,5 10
longueur
10
9,5
9
8,5
8
14
Pêcheur
M1/
Pié Lombard
13
12
11
10
12 14 16 18 20
longueur
12
11
10
9
largeu r
8
7
P4/
Pêcheur
Pié Lombard
7 8 9 10 11 12
longueur
M 2/
17 17,5 18 18,5 19 19,5 20 20,5 21
l o n g u e u r
14
13,5
13
12,5
12
11,5
11
10,5
10
Pêcheur
Pi é Lombard
M 3 /
Pié Lombar d
9
8
15 17 19 21 23 25
l ongueur
16
15
14
13
12
11
10
Pêcheur
Figure 36: Diagrammes des dimensions comparées de
Capra ibex cebennarum de la grotte des Pêcheurs et de Capra
ibex de Pié Lombard pour la dentition supérieure,
d'après la longueur et la largeur maximal occlusale.
|
P/3
|
Pié Lombard
|
Pêcheurs
|
|
1
|
22
6,74 - 9,33
8,01
0,60
|
24
7 - 10,4
8,74
0,73
|
|
3
|
21
|
22
|
|
4,74 - 6,77
|
5,4 - 7,2
|
|
5,72
|
6,35
|
|
0,5
|
0,48
|
|
P/4
|
|
|
|
1
|
24
|
28
|
|
8,93 - 11,37
|
8,9 - 12,3
|
|
10,08
|
10,82
|
|
0,75
|
0,92
|
|
3
|
22
|
23
|
|
4,89 - 7,83
|
6,9 - 8,3
|
|
7,14
|
7,44
|
|
0,63
|
0,37
|
|
M/1
|
|
|
|
1
|
25
|
32
|
|
11,06 - 15,94
|
12,6 - 18,6
|
|
13,90
|
15,67
|
|
1,38
|
1,54
|
|
3
|
25
|
31
|
|
6,83 - 10,12
|
6,9 - 8,3
|
|
8,52
|
8,72
|
|
0,7
|
0,82
|
|
M/2
|
|
|
|
1
|
17
|
30
|
|
15,71 - 18
|
16,9 - 22,6
|
|
16,72
|
18,63
|
|
0,65
|
1,19
|
|
3
|
17
|
29
|
|
8 - 10,33
|
8 - 11,8
|
|
9,26
|
9,64
|
|
0,69
|
0,89
|
|
M/3
|
|
|
|
1
|
10
|
22
|
|
23,31 - 28,15
|
22,7 - 31,9
|
|
25,29
|
27,00
|
|
1,46
|
2,17
|
|
3
|
11
|
24
|
|
8,05 - 10,39
|
8,2 - 11,1
|
|
9,24
|
9,49
|
|
0,77
|
0,66
|
Figure 37: Dimensions comparées de Capra ibex
cebennarum de la grotte des Pêcheurs et de Capra ibex de
Pié Lombard pour la dentition inférieure, d'après la
longueur (1) et la largeur maximale (3) occlusale.
65

Pêcheur
Pié Lombard
P/3
7
6,5
6
5,5
5
4,5
4
6 6,5 7 7,5 8 8,5 9 9,5 10
longeur

8
Pêcheur
7,5
7
Pié Lombar d
6,5
6
P / 4
8 8,5 9 9,5 10 10,5 11 11,5 12
l ongueur

10
Pêcheur
M/1
Pié Lombard
9,5
9
8,5
8
7,5
7
6,5
6
10 12 14 16 18
longue ur
M/ 2
15 17 19
l ongueur

11
10,5
10
9,5
9
8,5
8
7,5
7
Pié Lombard
Pêcheur
M/3

Pié Lombard
Pêcheur
20 22 24 26 28 30
longueur
10,5
10
9,5
9
8,5
8
Figure 38 : Diagrammes des dimensions comparées de
Capra ibex cebennarum de la grotte des Pêcheurs et de Capra
ibex de Pié Lombard pour la dentition inférieure,
d'après la longueur et la largeur maximale occlusale.

66
Figure 39: Distinction M1 et M2: à gauche M2 et M1
en vue linguale ; à droite M2 et M1 en vue mésiale
Figure 40: Distinction dP3 et dP4:
à gauche dP4 et dP3 en vue vestibulaire; à
droite racines des dP4 et dP3

Figure 41: Distinction P3 et P4 ,
vue vestibulaire

Figure 42: Distinction M2 et M1 ,
vue vestibulaire

Figure 43: Mandibule gauche de Capra ibex
retrouvée presque entière (sauf la P2) avec sa branche
mandibulaire

Figure 44: Maxillaire de Capra ibex
retrouvée presque entière, avec conservation processus palatin et
corps maxillaire


M/1
25
20
phase IIIa phase IIIb phase IV
0








1 2 3 4 7
variables
|
M1
|
1
|
2
|
3
|
4
|
7
|
|
Phase IIIa
|
12
12,88 - 15,94
|
9
10,35 - 13,18
|
13
6,83 - 10,12
|
11
6,71 - 10,17
|
9
10,87 - 28,56
|
|
14,7
|
11,37
|
8,62
|
8,48
|
21,87
|
|
0,89
|
0,90
|
0,88
|
0,92
|
5,08
|
|
Phase
|
7
|
4
|
7
|
7
|
4
|
|
IIIb
|
12,5 - 14,92
|
10,87 - 12,2
|
8,14 - 9,13
|
8,23 - 9,97
|
9,92 - 17,27
|
|
13,49
|
11,57
|
8,57
|
9,08
|
13,39
|
|
0,88
|
0,58
|
0,40
|
0,62
|
3,40
|
|
Phase
|
1
|
1
|
1
|
2
|
2
|
|
IV
|
11,77
|
10,73
|
7,24
|
7,46 - 8,63
|
6,1 - 12,6
|
|
|
|
|
8,05
|
9,35
|
|
|
|
|
0,83
|
4,60
|
Figure 46: dimensions comparées de la M/1 en
phase III et IV ; 1 : diamètre mesio-distal occlusal, 2 :
diamètre mesio-distal au collet, 3 : diamètre vestibulo-lingual
occlusale du lobe mesial, 4 : diamètre vestibulo-lingual du lobe distal,
7 : hauteur de couronne face linguale.


20
15
10
en mm
5
0




1 2 3 4 7
variables
phase IIIa phase IIIb phase IV



|
M2
|
1
|
2
|
3
|
4
|
7
|
|
Phase IIIa
|
6
15,82 - 17,51
|
5
11,17 - 14,53
|
6
8 - 9,85
|
6
7,53 - 10,17
|
6
23,36 - 36,81
|
|
16,70
|
13,10
|
8,78
|
8,48
|
31,93
|
|
0,77
|
1,32
|
0,71
|
1,53
|
5,08
|
|
Phase
|
5
|
4
|
5
|
5
|
4
|
|
IIIb
|
16,29 - 18
|
13,64 - 14,39
|
9,03 - 10,3
|
8,5 - 9,65
|
26,5 - 29,33
|
|
17,01
|
14,2
|
9,48
|
9,14
|
28,03
|
|
0,71
|
0,37
|
0,51
|
0,45
|
1,22
|
|
Phase
|
3
|
1
|
3
|
3
|
2
|
|
IV
|
13,95 - 16,43
|
15,25
|
9,23 - 9,72
|
9,28 - 10,28
|
10,94 - 16,79
|
|
15,22
|
|
9,51
|
9,86
|
13,87
|
|
1,24
|
|
0,25
|
0,52
|
4,14
|
Figure 48: dimensions comparées des M/2 en
phase III et IV ; 1 : diamètre mesio-distal occlusal, 2 :
diamètre mesio-distal au collet, 3 : diamètre vestibulo-lingual
occlusale du lobe mesial, 4 : diamètre vestibulo-lingual du lobe distal,
7 : hauteur de couronne face linguale.
|
|
|
|
M3
|
1
|
2
|
3
|
4
|
5
|
7
|
|
Phase IIIa
|
3
23,31 - 26,46
24,76
1,59
|
2
27,82 - 29,99
28,91
1,53
|
3
8,05 - 9,54
8,92
0,78
|
5
7,68 - 8,93
8,12
0,56
|
3
4,90 - 5,89
5,31
0,43
|
4
30,94 - 35,96
33,34
2,63
|
|
Phase
|
3
|
3
|
4
|
5
|
3
|
3
|
|
|
IIIb
|
24,2 - 26,05
|
26,55 - 29,31
|
8,87 - 10,39
|
7,19 - 10,15
|
5,12 - 5,94
|
22,61 - 38,93
|
|
|
|
25,42
|
27,58
|
9,56
|
8,61
|
5,52
|
31,55
|
|
|
|
1,05
|
1,50
|
0,81
|
1,08
|
0,41
|
8,27
|
|
|
Phase
|
|
|
|
1
|
1
|
1
|
|
|
IV
|
|
|
|
10,32
|
5,90
|
14,35
|
|
|
|
|
|
|
|
40
|
M/3
|
|
35
|
|
|
|
30
|
|
|
|
25
|
|
|
|
20
|
phase IIIa
|
|
|
15
|
phase IIIb
|
|
|
10
|
phase IV
|
|
|
5
|
|
|
|
0
|
|
|
|
|
|
|
1 2 3 4 5 7
variables
|
|
|
Figure 49 : dimensions comparées des M/3 en
phase III et IV ; 1 : diamètre mesio-distal occlusal, 2 :
diamètre mesio-distal au collet, 3 : diamètre vestibulo-lingual
occlusale du lobe mesial, 4 : diamètre vestibulo-lingual du lobe
médian, 5 : diamètre vestibulo-lingual du lobe distal, 7 :
hauteur de couronne face linguale.

18
16
14
12
10
8
en mm
6
4
M1/
Phase IIIa Phase IIIb Phase IV
2
0
1 2 3 4 7
variables
|
M1
|
1
|
2
|
3
|
4
|
7
|
|
Phase
|
2
|
1
|
1
|
|
|
|
IIIa
|
16,1 - 16,46
|
11,55
|
9,78
|
|
|
|
16,28
|
|
|
|
|
|
0,25
|
|
|
|
|
|
Phase
|
5
|
4
|
5
|
4
|
4
|
|
IIIb
|
14,5 - 15,82
|
10,6 - 12,87
|
10,25 - 12,16
|
9,39 - 12,05
|
13,67 - 23,15
|
|
15,14
|
11,40
|
11,58
|
10,64
|
17,07
|
|
0,51
|
1,2
|
0,78
|
1,09
|
4,27
|
|
Phase
|
3
|
|
3
|
3
|
2
|
|
IV
|
13,24 - 14,80
|
|
11,65 - 12,70
|
10,47 - 10,92
|
12,55 - 13,49
|
|
13,76
|
|
12,20
|
10,74
|
13,02
|
|
0,90
|
|
0,53
|
0,24
|
0,66
|
Figure 50 : dimensions comparées des M1/ en
phase III et IV ; 1 : diamètre mesio-distal occlusal, 2 :
diamètre mesio-distal au collet, 3 : diamètre vestibulo-lingual
occlusale du lobe mesial, 4 : diamètre vestibulo-lingual du lobe distal,
7 : hauteur de couronne face vestibulaire.


30
25

phase IIIa phase IIIb phase IV
en mm
20
15
10
5
0







1 2 3 4 7
variables
|
M2
|
1
|
2
|
3
|
4
|
7
|
|
Phase IIIa
|
5
16,87 - 19,41
|
3
11,83 - 15,65
|
5
9,73 - 11,69
|
4
8,57 - 10,39
|
4
32,39 - 38,19
|
|
17,89
|
13,74
|
10,60
|
9,14
|
35,73
|
|
0,94
|
1,91
|
0,73
|
0,85
|
2,70
|
|
Phase
|
2
|
2
|
2
|
2
|
2
|
|
IIIb
|
18,15 - 18,81
|
14,24 - 15,07
|
11,19 - 11,42
|
9,21 - 10,09
|
30,53 - 32,31
|
|
18,48
|
14,66
|
11,31
|
9,65
|
31,42
|
|
0,47
|
0,59
|
0,16
|
0,62
|
1,26
|
|
Phase
|
2
|
1
|
2
|
2
|
1
|
|
IV
|
17 - 17,92
|
13,78
|
11,76 - 13,03
|
10,88 - 11,34
|
20,75
|
|
17,46
|
|
12,40
|
11,11
|
|
|
0,65
|
|
0,90
|
0,33
|
|
Figure 52: dimensions comparées des M2/ en
phase III et IV ; 1 : diamètre mesio-distal occlusal, 2 :
diamètre mesio-distal au collet, 3 : diamètre vestibulo-lingual
occlusale du lobe mesial, 4 : diamètre vestibulo-lingual du lobe distal,
7 : hauteur de couronne face vestibulaire.

45
40
35
30
en mm
25
20
15
10
M3/
phase IIIa phase IIIb phase IV
5
0
1 2 3 4 7
variable
|
M3
|
1
|
2
|
3
|
4
|
7
|
|
Phase IIIa
|
3
16,24 - 17,19
|
2
24,53 - 27,69
|
6
9,15 - 11,16
|
5
6,62 - 9,70
|
3
39,3 - 42,09
|
|
16,70
|
26,11
|
9,88
|
8,07
|
40,88
|
|
0,48
|
2,23
|
0,77
|
1,32
|
1,43
|
|
Phase
|
5
|
3
|
5
|
6
|
5
|
|
IIIb
|
18,83 - 22,10
|
21,44 - 23,10
|
10,90 - 12,34
|
8,79 - 11,38
|
27,13 - 36,75
|
|
20,98
|
22,42
|
11,57
|
9,76
|
29,87
|
|
1,46
|
0,87
|
0,53
|
0,91
|
3,94
|
|
Phase
|
3
|
2
|
3
|
3
|
3
|
|
IV
|
21,49 - 24,02
|
21,7 - 24,01
|
11,02 - 13,24
|
9,81 - 11,56
|
12,29 - 19,17
|
|
23,14
|
22,86
|
12,13
|
10,72
|
16,85
|
|
1,43
|
1,63
|
1,11
|
0,88
|
3,95
|
Figure 53: dimensions comparées des M3/ en
phase III et IV ; 1 : diamètre mesio-distal occlusal, 2 :
diamètre mesio-distal au collet, 3 : diamètre vestibulo-lingual
occlusale du lobe mesial, 4 : diamètre vestibulo-lingual du lobe distal,
7 : hauteur de couronne face vestibulaire.
Table des illustrations
FIGURE 1 : LOCALISATION DE L'ABRI PIE LOMBARD, D'APRES GOOGLE MAP
HTTP://MAPS.GOOGLE.FR/
(CONSULTE LE 1 JUIN 2009) 9
FIGURE 2 : COUPE STRATIGRAPHIQUE (TEXIER, 1996) 10
FIGURE 3: LISTE FAUNIQUE DES GRANDS MAMMIFERES DES COUCHES
MOUSTERIENNES DE PIE LOMBARD, POURCENTAGE BASE SUR L'ETUDE DU POSTCRANIEN
(D'APRES GERBER, 1973 ; BRUGAL ET FERNANDEZ INEDIT) 13
FIGURE 4: REPARTITION ANATOMIQUE DE LA DENTITION SUPERIEURE DE
R. RUPICAPRA DE PIE LOMBARD. _ 19
FIGURE 5: REPARTITION ANATOMIQUE DE LA DENTITION INFERIEURE DE
R. RUPICAPRA DE PIE LOMBARD. 20
FIGURE 6: CRITERES DE DISTINCTION MORPHOLOGIQUE ENTRE
CAPRA IBEX ET HEMITRAGUS
CEDRENSIS POUR LA DENTITION SUPERIEURE (D'APRES FERNANDEZ 2006 ET
CREGUT-BONNOURE, 2002 ET 2005-2006). _ 22
FIGURE 7: CRITERES DE DISTINCTION MORPHOLOGIQUE ENTRE
CAPRA IBEX ET HEMITRAGUS
CEDRENSIS POUR LA DENTITION INFERIEURE (D'APRES FERNANDEZ 2006 ET
CREGUT-BONNOURE, 2002 ET 2005-2006). 23
FIGURE 8: REPARTITION ANATOMIQUE DES INCISIVES DE C. IBEX EN
NRD (NOMBRE DE DENTS DETERMINEES)26
FIGURE 9: REPARTITION ANATOMIQUE DE LA DENTITION INFERIEURE DE
C. IBEX EN NRD (NOMBRE DE DENTS DETERMINEES) 26
FIGURE 10: REPARTITION ANATOMIQUE DE LA DENTITION SUPERIEURE DE
C. IBEX EN NRD (NOMBRE DE DENTS DETERMINEES) 27
FIGURE 11: DIMENSIONS COMPAREES DES DENTS M/1 ET M/2 ISOLEES DE
L'ABRI PIE LOMBARD EN MM; 1 : DIAMETRE MESIO-DISTAL OCCLUSAL, 3 : DIAMETRE
VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL, 7 : HAUTEUR DE COURONNE FACE
LINGUALE. 29
FIGURE 12: DIMENSIONS COMPAREES DES DP3/ ET DES DP4/ ISOLEES DE
L'ABRI PIE LOMBARD ; 1 : DIAMETRE
MESIO-DISTAL OCCLUSAL, 3 : DIAMETRE VESTIBULO-LINGUAL MAXIMAL
OCCLUSAL, 7 : HAUTEUR DE COURONNE FACE VESTIBULAIRE. 30
FIGURE 13: DIMENSIONS COMPAREES DES P3/ ET DES P4/ ISOLEES DE
L'ABRI PIE LOMBARD ; 1 : DIAMETRE MESIO-DISTAL OCCLUSAL, 3 : DIAMETRE
VESTIBULO-LINGUAL MAXIMAL OCCLUSAL, 7 : HAUTEUR DE COURONNE FACE VESTIBULAIRE.
31
FIGURE 14: DIMENSIONS COMPAREE DES M1/ ET DES M2/ ISOLEES DE
L'ABRI PIE LOMBARD ; 1 : DIAMETRE MESIO-DISTAL OCCLUSAL, 3 : DIAMETRE
VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL, 7 : HAUTEUR DE COURONNE FACE
LINGUALE. 32
FIGURE 15: POURCENTAGE DES DEUX CATEGORIES DE FRAGMENTATION :
CATEGORIE 1 : 9% ; CATEGORIE 2 :
24% ; ET DU NOMBRE DE DENT TOTAL DETERMINEE
: 67% SUR L'ENSEMBLE DU NRDT (NOMBRE DE
DENTS TOTAL) 35
FIGURE 16: TABLEAU DU NRD ET NRDT DE L'ENSEMBLE DU MATERIEL DU
GISEMENT DE PIE LOMBARD 36
FIGURE 17: NOMENCLATURE DES MOLAIRES : 1. MOLAIRE SUPERIEURE
GAUCHE ; 2. MOLAIRE INFERIEURE
GAUCHE 39
FIGURE 18: DP/4 EN PHASE I, VUE OCCLUSALE 39
FIGURE 19: FRAGMENT DE MANDIBULE DROITE, ERUPTION DE LA M/1 EN
PHASE IIA, VUE OCCLUSALE ET
VESTIBULAIRE 41
FIGURE 20: M/1 GAUCHE EN PHASE IIB, VUE OCCLUSALE ET LINGUALE
42
FIGURE 21: M/1 DROITE EN PHASE IIC, VUE OCCLUSALE. 43
FIGURE 22: FRAGMENT DE MAXILLAIRE AVEC M1/ DROITE EN ERUPTION,
VUE LINGUALE ET M1/ EN PHASE IIA,
VUE OCCLUSALE 44
FIGURE 23: M1/ DROITE EN PHASE IIB, VUE OCCLUSALE 45
FIGURE 24: M1/ DROITE EN PHASE IIC, VUE OCCLUSALE 46
FIGURE 25: M/3 DROITE EN PHASE IIC, VUE OCCLUSALE ET LINGUALE
47
FIGURE 26 : M3/ EN PHASE IIC, VUE OCCLUSALE ET VESTIBULAIRE 48
FIGURE 27: CRITERES MORPHOLOGIQUES DISTINCTIFS ENTRE LES PHASES
II, IIIA, IIIB ET IV 49
FIGURE 28: TABLEAU DE REPARTITION DE LA DENTITION INFERIEURE PAR
CLASSE D'AGE ET PAR PHASE ET NOMBRE MINIMUM D'INDIVIDU PAR PHASE 50
FIGURE
29: TABLEAU DE REPARTITION DE LA DENTITION SUPERIEURE PAR CLASSE D'AGE ET PAR
PHASE ET
NOMBRE MINIMUM D'INDIVIDU PAR PHASE 51
FIGURE 30 : TABLEAU RECAPITULATIF DU NMIC POUR CHAQUE PHASE
51
FIGURE 31: POURCENTAGE DU NMIC PAR CLASSE D'AGE 52
FIGURE 32: PROFIL DE MORTALITE D'APRES LE NMIC PAR CLASSE D'AGE
52
FIGURE 33: PROFIL DE MORTALITE D'APRES LE NMIC PAR PHASE ET PAR
CLASSE D'AGE 53
FIGURE 34: NMIC DES JUVENILES ET DES SUBADULTES EN RELATION AVEC
LES MOIS ET SAISONS (D'APRES COUTURIER, 1962) 56
FIGURE 35: DIMENSIONS COMPAREES DE CAPRA IBEX
CEBENNARUM DE LA GROTTE DES PECHEURS ET DE CAPRA
IBEX DE PIE LOMBARD POUR LA DENTITION SUPERIEURE, D'APRES LA
LONGUEUR (1) ET LA LARGEUR MAXIMALE (3) OCCLUSALE. 62
FIGURE 36: DIAGRAMMES DES DIMENSIONS COMPAREES DE
CAPRA IBEX CEBENNARUM DE LA GROTTE DES PECHEURS ET DE
CAPRA IBEX DE PIE LOMBARD POUR LA DENTITION
SUPERIEURE, D'APRES LA LONGUEUR ET LA LARGEUR MAXIMAL OCCLUSALE. 63
FIGURE 37: DIMENSIONS COMPAREES DE CAPRA IBEX
CEBENNARUM DE LA GROTTE DES PECHEURS ET DE CAPRA
IBEX DE PIE LOMBARD POUR LA DENTITION INFERIEURE, D'APRES LA
LONGUEUR (1) ET LA LARGEUR MAXIMALE (3) OCCLUSALE. 64
FIGURE 38 : DIAGRAMMES DES DIMENSIONS COMPAREES DE
CAPRA IBEX CEBENNARUM DE LA GROTTE DES PECHEURS ET DE
CAPRA IBEX DE PIE LOMBARD POUR LA DENTITION
INFERIEURE, D'APRES LA LONGUEUR ET LA LARGEUR MAXIMALE OCCLUSALE. 65
FIGURE 39: DISTINCTION M1 ET M2: A GAUCHE M2 ET M1 EN VUE
LINGUALE ; A DROITE M2 ET M1 EN VUE MESIALE 66
FIGURE 40: DISTINCTION DP3 ET DP4: A GAUCHE
DP4 ET DP3 EN VUE VESTIBULAIRE; A DROITE RACINES DES
DP4
ET DP3 66
FIGURE 41: DISTINCTION P3 ET P4 , VUE
VESTIBULAIRE 67
FIGURE 42: DISTINCTION M2 ET M1 , VUE
VESTIBULAIRE 67
FIGURE 43: MANDIBULE GAUCHE DE CAPRA
IBEX RETROUVEE PRESQUE ENTIERE (SAUF LA P2) AVEC SA BRANCHE
MANDIBULAIRE 68
FIGURE 44: MAXILLAIRE DE CAPRA IBEX
RETROUVEE PRESQUE ENTIERE, AVEC CONSERVATION PROCESSUS PALATIN ET CORPS
MAXILLAIRE 68
FIGURE 45: FRAGMENT DE MANDIBULE AVEC M/3 ET M/2: LE CORPS
MANDIBULAIRE EST FRAGMENTE MAIS LES RACINES SONT INTACTES 70
FIGURE 46: DIMENSIONS COMPAREES DE LA M/1 EN PHASE III ET IV ; 1
: DIAMETRE MESIO-DISTAL OCCLUSAL, 2 : DIAMETRE MESIO-DISTAL AU COLLET, 3 :
DIAMETRE VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL, 4 : DIAMETRE
VESTIBULO-LINGUAL DU LOBE DISTAL, 7 : HAUTEUR DE COURONNE FACE
LINGUALE. 71
FIGURE 47: DE GAUCHE A DROITE: M/1 PHASE IIIA, M/1 PHASE IIIB ET
M/1 PHASE IV; EN HAUT VUE VESTIBULAIRE ET EN BAS VUE LINGUALE 72
FIGURE 48: DIMENSIONS COMPAREES DES M/2 EN PHASE III ET IV ; 1 :
DIAMETRE MESIO-DISTAL OCCLUSAL, 2 : DIAMETRE MESIO-DISTAL AU COLLET, 3 :
DIAMETRE VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL,
4 : DIAMETRE VESTIBULO-LINGUAL DU LOBE DISTAL, 7 : HAUTEUR DE
COURONNE FACE LINGUALE. 73
FIGURE 49 : DIMENSIONS COMPAREES DES M/3 EN PHASE III ET IV ; 1 :
DIAMETRE MESIO-DISTAL OCCLUSAL, 2 : DIAMETRE MESIO-DISTAL AU COLLET, 3 :
DIAMETRE VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL,
4 : DIAMETRE VESTIBULO-LINGUAL DU LOBE MEDIAN, 5 : DIAMETRE
VESTIBULO-LINGUAL DU LOBE DISTAL, 7 : HAUTEUR DE COURONNE FACE LINGUALE. 74
FIGURE 50 : DIMENSIONS COMPAREES DES M1/ EN PHASE III ET IV ; 1 :
DIAMETRE MESIO-DISTAL OCCLUSAL, 2 : DIAMETRE MESIO-DISTAL AU COLLET, 3 :
DIAMETRE VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL,
4 : DIAMETRE VESTIBULO-LINGUAL DU LOBE DISTAL, 7 : HAUTEUR DE
COURONNE FACE VESTIBULAIRE. 75
FIGURE 51: DE GAUCHE A DROITE: M1/ EN PHASE IIIA, M1/ EN PHASE
IIIB ET M1/ EN PHASE IV; EN HAUT VUE LINGUALE ET EN BAS VUE VESTIBULAIRE 76
FIGURE 52: DIMENSIONS COMPAREES DES M2/ EN PHASE III ET IV ; 1 :
DIAMETRE MESIO-DISTAL OCCLUSAL, 2 : DIAMETRE MESIO-DISTAL AU COLLET, 3 :
DIAMETRE VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL,
4 : DIAMETRE VESTIBULO-LINGUAL DU LOBE DISTAL, 7 : HAUTEUR DE
COURONNE FACE VESTIBULAIRE. 77
FIGURE 53: DIMENSIONS COMPAREES DES M3/ EN PHASE III ET IV ; 1 :
DIAMETRE MESIO-DISTAL OCCLUSAL, 2 : DIAMETRE MESIO-DISTAL AU COLLET, 3 :
DIAMETRE VESTIBULO-LINGUAL OCCLUSALE DU LOBE MESIAL,
4 : DIAMETRE VESTIBULO-LINGUAL DU LOBE DISTAL, 7 : HAUTEUR DE
COURONNE FACE VESTIBULAIRE. 78
Table des annexes
ANNEXE 1: NOMENCLATURE DES DENTS JUGALES : MOLAIRE SUPERIEURE
GAUCHE, VUE OCCLUSALE ET M3
GAUCHE, VUE OCCLUSALE (D'APRES CREGUT-BONNOURE, 2002)
84
ANNEXE 2: REPRESENTATION SCHEMATIQUE D'UNE DP4
DROITE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006
85 ANNEXE 3 : REPRESENTATION SCHEMATIQUE D'UNE P3 DROITE D'H.
CEDRENSIS, D'APRES FERNANDEZ 2006 85 ANNEXE 4: REPRESENTATION SCHEMATIQUE D'UNE
P4 DROITE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006_ 85
ANNEXE 5: REPRESENTATION SCHEMATIQUE D'UNE M1 GAUCHE D'H.
CEDRENSIS, D'APRES FERNANDEZ 2006
86
ANNEXE 6: REPRESENTATION
SCHEMATIQUE D'UNE M2 GAUCHE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006
86
ANNEXE 7: REPRESENTATION
SCHEMATIQUE D'UNE M3 GAUCHE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006
87
ANNEXE 8: REPRESENTATION
SCHEMATIQUE D'UNE DP3/4 DROITE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006
87 ANNEXE 9: REPRESENTATION
SCHEMATIQUE D'UNE P2 DROITE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006_
88 ANNEXE 10: REPRESENTATION SCHEMATIQUE D'UNE P3 P4
DROITE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006
88
ANNEXE 11: REPRESENTATION
SCHEMATIQUE D'UNE M1 DROITE D'H. CEDRENSIS, D'APRES FERNANDEZ 2006
88
ANNEXE 12: REPRESENTATION D'UNE M2 DROITE D'H. CEDRENSIS, D'APRES
FERNANDEZ 2006 89
ANNEXE 13: REPRESENTATION SCHEMATIQUE D'UNE M3 DROITE D'H.
CEDRENSIS, D'APRES FERNANDEZ 2006
89
ANNEXE 14: TABLEAU D'INVENTAIRE ET DE MESURES DES DENTS JUGALES
ISOLES DE C. IBEX DE L'ENSEMBLE
MOUSTERIEN 90
ANNEXE 15: TABLEAU
D'INVENTAIRE ET DE MESURES DES SERIES DENTAIRES DE C. IBEX DE L'ENSEMBLE
MOUSTERIEN 96 ANNEXE 16: TABLEAU
D'INVENTAIRE ET DE MESURES DES INCISIVES DE C. IBEX DE L'ENSEMBLE MOUSTERIEN
99 ANNEXE 17: TABLEAU D'INVENTAIRE ET DE MESURES DES
DENTS JUGALES ISOLEES DE C. IBEX DE L'ENSEMBLE
EPIPALEOLITHIQUE 102
ANNEXE 18: TABLEAU D'INVENTAIRE ET DE
MESURES DES INCISIVES DE C. IBEX DE L'ENSEMBLE
EPIPALEOLITHIQUE 103
ANNEXE 19: TABLEAU D'INVENTAIRE ET DE
MESURES DES DENTS JUGALES DE C. IBEX DE LA ZONE R
(MOUSTERIEN?) 103
ANNEXE 20 : TABLEAU D'INVENTAIRE ET DE
MESURES DES SERIES DENTAIRES DE C. IBEX DE LA ZONE R
(MOUSTERIEN?) 104
ANNEXE 21 :
TABLEAU D'INVENTAIRE ET DE MESURE DES INCISIVES DE C. IBEX DE LA ZONE R
(MOUSTERIEN ?)
104
Annexes
1 : diamètre mésio-
distal occlusal
2 : diamètre mésio- distal au
collet
3 : diamètre vestibulo-
lingual occlusale du lobe mésial ou maximum pour
les P2, P3 et P4
4 : diamètre vestibulo- lingual du lobe distal ou
médian (M/3 et D/4)
5 : diamètre vestibulo-
lingual du lobe distal des M/3 et P/4
6 : diamètre vestibulo- lingual au
collet
7 : hauteur de couronne maximum (
prise en face linguale pour les dents inférieures
et en face ).
vestibulaire pour les dents
supérieures


Annexe 1
: Nomenclature des dents jugales : Molaire
supérieure gauche, vue occlusale et M3 gauche, vue occlusale
(d'après Cregut- Bonnoure, 2002)

3
4
5

6
Annexe 4: Représentation schématique d'une
P4 droite d'H. cedrensis, d'après Fernandez 2006
Annexe 2: Représentation schématique d'une
dP4 droite d'H. cedrensis, d'après Fernandez 2006

Annexe 3 : Représentation schématique d'une
P3 droite d'H. cedrensis, d'après Fernandez 2006

6
Annexe 5: Représentation schématique d'une
M1 gauche d'H. cedrensis, d'après Fernandez 2006

6
Annexe 6: Représentation schématique d'une
M2 gauche d'H. cedrensis, d'après Fernandez 2006

Annexe 7: Représentation schématique d'une
M3 gauche d'H. cedrensis, d'après Fernandez 2006

3
6
Annexe 8: Représentation schématique d'une
dP3/4 droite d'H. cedrensis, d'après Fernandez 2006

Annexe 9: Représentation schématique d'une
P2 droite d'H. cedrensis, d'après Fernandez 2006

6
3
Annexe 10: Représentation schématique d'une
P3 P4 droite d'H. cedrensis, d'après Fernandez 2006

Annexe 11: Représentation schématique d'une
M1 droite d'H. cedrensis, d'après Fernandez 2006


6
Annexe 12: Représentation d'une M2 droite d'H.
cedrensis, d'après Fernandez 2006
Annexe 14: Tableau d'inventaire et de mesures des dents
jugales isolés de C. ibex de l'ensemble
moustérien
|
N
|
Dents
|
Couche
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
467
|
M/3 G
|
E4 E4 i
|
21.30
|
28.53
|
7.43
|
7.72
|
4.95
|
9.65
|
43.62
|
|
1856
|
M/3 G
|
C5 e
|
21.53
|
X
|
7.24
|
5.89
|
4.74
|
8.61
|
38.81
|
|
357
|
M/3 G
|
C4 C4 f
|
24.52
|
27.82
|
9.18
|
8.50
|
5.15
|
7.74
|
31.21
|
|
2124
|
M/3 G
|
C3 e
|
23.31
|
X
|
8.05
|
7.73
|
4.90
|
X
|
35.24
|
|
2031
|
M/3 G
|
B4 MA
|
26.00
|
29.31
|
9.53
|
8.64
|
5.94
|
9.23
|
38.93
|
|
3472
|
M/3 G
|
B5 MA3
|
X
|
X
|
X
|
7.78
|
X
|
X
|
35.96
|
|
194
|
M/3 G
|
F4
|
24.20
|
26.55
|
8.47
|
8.16
|
5.12
|
8.44
|
33.11
|
|
3131
|
M/3 G
|
B5 MA
|
X
|
X
|
X
|
7.68
|
X
|
X
|
X
|
|
17.1
|
M/3 D
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3080
|
M/3 D
|
F3
|
X
|
X
|
6.10
|
4.66
|
X
|
X
|
33.79
|
|
15.1
|
M/3 D
|
F3
|
26.46
|
29.99
|
9.54
|
8.93
|
5.89
|
10.27
|
30.94
|
|
978
|
M/3 D
|
C4 C4 G
|
X
|
X
|
10.39
|
10.15
|
X
|
X
|
X
|
|
IV
|
M/3 D
|
F3
|
X
|
X
|
X
|
7.19
|
X
|
X
|
X
|
|
1004
|
M/3 D
|
C4 C4 G
|
26.05
|
26.89
|
9.83
|
8.93
|
5.49
|
9.65
|
22.61
|
|
2092
|
M/3 D
|
B4 MA
|
X
|
X
|
X
|
10.32
|
5.90
|
X
|
14.35
|
|
1599
|
M/3 D
|
C5 e
|
X
|
X
|
X
|
8.37
|
X
|
X
|
X
|
|
3
|
M/2 G
|
F3
|
16.29
|
X
|
6.06
|
5.92
|
X
|
X
|
X
|
|
3057
|
M/2 G
|
F3
|
17.89
|
X
|
X
|
8.37
|
X
|
X
|
X
|
|
216
|
M/2 G
|
D4 D4 f
|
X
|
13.18
|
9.85
|
10.17
|
X
|
8.33
|
23.36
|
|
3103
|
M/2 G
|
B4 M
|
17.33
|
X
|
8
|
7.53
|
X
|
9.22
|
36.81
|
|
76
|
M/2 G
|
D5 D5 f
|
15.83
|
X
|
9.27
|
9.26
|
X
|
X
|
X
|
|
2088
|
M/2 G
|
MA B4
|
15.82
|
12.58
|
8.52
|
8.06
|
X
|
7.37
|
33.57
|
|
1600
|
M/2 G
|
C5 e
|
16.43
|
11.17
|
8.10
|
7.62
|
X
|
8.41
|
29.59
|
|
48
|
M/2 G
|
C4 C4 b
|
16.67
|
13.77
|
9.20
|
8.50
|
X
|
7.95
|
27.83
|
|
2087
|
M/2 G
|
MA B4
|
16.29
|
13.64
|
9.03
|
9.12
|
X
|
8.55
|
28.66
|
|
707
|
M/2 G
|
C5e
|
17.49
|
14.39
|
9.22
|
8.94
|
X
|
8.26
|
29.33
|
|
13.1
|
M/2 G
|
F3
|
16.43
|
15.25
|
9.72
|
10.28
|
X
|
10.05
|
10.94
|
|
365
|
M/2 D
|
C4 E1
|
16.21
|
X
|
6.52
|
5.44
|
X
|
X
|
X
|
|
3432
|
M/2 D
|
B4 MA
|
18
|
X
|
7.51
|
6.37
|
X
|
X
|
X
|
|
480
|
M/2 D
|
C4 C4 ei
|
19.51
|
13.18
|
7.61
|
7.30
|
X
|
8.18
|
40.91
|
|
3453
|
M/2 D
|
B4 MA
|
18.77
|
13.38
|
8.40
|
7.75
|
X
|
8.93
|
40.88
|
|
10.2
|
M/2 D
|
F3
|
18.01
|
X
|
8.48
|
8.69
|
X
|
X
|
39.51
|
|
56
|
M/2 D
|
F4 F4 e
|
18.55
|
X
|
8.67
|
8.59
|
X
|
X
|
X
|
|
3014
|
M/1 D
|
F3
|
15.94
|
X
|
7.93
|
X
|
X
|
X
|
28.56
|
|
3488
|
M/2 D
|
B5 MA3
|
17.27
|
14.06
|
X
|
X
|
X
|
8.48
|
31.48
|
|
3550
|
M/2 D
|
B5 MA4
|
17.51
|
14.53
|
8.94
|
8.25
|
X
|
8.33
|
36.76
|
|
38
|
M/2 D
|
E4E4 h
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
14
|
M/1 D
|
F3
|
14.23
|
X
|
10.12
|
9.24
|
X
|
X
|
X
|
|
214
|
M/1 D
|
E4 EB1
|
14.83
|
X
|
8.91
|
8.80
|
X
|
X
|
X
|
|
3539
|
M/2 D
|
B5 MA3
|
16.58
|
X
|
9.63
|
9.48
|
X
|
X
|
X
|
|
247
|
M/2 D
|
C4 C4 e
|
18.00
|
14.26
|
10.30
|
9.65
|
X
|
9.31
|
26.50
|
|
7
|
M/2 D
|
F3
|
15.28
|
X
|
9.58
|
9.28
|
X
|
X
|
16.79
|
|
14
|
M/2 D
|
F4
|
13.95
|
X
|
9.23
|
10.02
|
X
|
X
|
X
|
|
3141
|
M/2 D
|
B4 M
|
16.87
|
13.68
|
9
|
X
|
X
|
8.52
|
31.54
|
|
2355
|
M/1 G
|
C3 e
|
16.82
|
13.48
|
5.90
|
X
|
X
|
X
|
25.79
|
|
36
|
M/1 G
|
C4 C4 a
|
16.40
|
12.51
|
6.56
|
6.38
|
X
|
8.34
|
32.17
|
|
3504
|
M/1 G
|
B5 MA3
|
16.14
|
X
|
6.54
|
6.57
|
X
|
X
|
X
|
|
8
|
M/1 G
|
D4 D4 f
|
15.94
|
X
|
6.88
|
6.24
|
X
|
X
|
X
|
|
216
|
M/1 G
|
D4 D4 f
|
X
|
13.18
|
9.85
|
10.17
|
X
|
8.33
|
23.36
|
|
VIII
|
M/1 G
|
F3
|
15.59
|
11.60
|
8.85
|
8.60
|
X
|
7.97
|
24.88
|
|
541?
|
M/1 G
|
B4 f
|
14.53
|
10.61
|
8.77
|
8.96
|
X
|
7.82
|
23.79
|
|
3525
|
M/1 G
|
B5 MA3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
2088
|
M/1 G
|
MA B4
|
14.03
|
10.35
|
8.56
|
8.39
|
X
|
7.54
|
18.76
|
|
2314
|
M/1 G
|
C3 e
|
14.16
|
X
|
8.31
|
8.41
|
X
|
X
|
X
|
|
80
|
M/1 G
|
D5 D5 f
|
14.70
|
10.85
|
7.53
|
7.80
|
X
|
7.36
|
24.13
|
|
346
|
M/1 G
|
E4 E4 I
|
12.50
|
X
|
8.14
|
8.55
|
X
|
X
|
X
|
|
?
|
M/1 G
|
B4 e
|
13.05
|
11.36
|
8.17
|
8.23
|
X
|
X
|
11.25
|
|
3265 bis
|
M/1 G
|
B5 MA3
|
14.92
|
X
|
9.13
|
9.97
|
X
|
X
|
X
|
|
939
|
M/1 G
|
C4 C4 G
|
13.15
|
X
|
8.56
|
9.16
|
X
|
X
|
X
|
|
752
|
M/1 G
|
C4 C4 ei
|
12.69
|
12.20
|
8.46
|
9.23
|
X
|
8.41
|
9.92
|
|
3497
|
M/1 G
|
B5 MA3
|
11.77
|
10.73
|
7.24
|
7.46
|
X
|
7.13
|
6.10
|
|
7.2
|
M/1 G
|
F3
|
13.08
|
9.72
|
6.96
|
6.69
|
X
|
7.07
|
16.33
|
|
11.2
|
M/1 G
|
F3
|
12.98
|
X
|
6.83
|
6.91
|
X
|
X
|
14.49
|
|
3134
|
M/1 D
|
B4 M
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3087
|
M/1 D
|
B5 MA
|
16.62
|
X
|
6.20
|
5.55
|
X
|
X
|
X
|
|
3307
|
M/1 D
|
B4 MA
|
14.66
|
11.89
|
7.67
|
7.14
|
X
|
8.58
|
27.66
|
|
305
|
M/1 D
|
F4 F4 ?
|
12.88
|
10.96
|
6.83
|
6.71
|
X
|
7.17
|
10.87
|
|
3278
|
M/1 D
|
B5 M3
|
14.24
|
11.55
|
8.42
|
8.70
|
X
|
7.74
|
19.33
|
|
2074
|
M/1 D
|
MA B4
|
15.55
|
11.89
|
X
|
X
|
X
|
8.25
|
X
|
|
61
|
M/1 D
|
D4 D4 f
|
15.72
|
11.56
|
8.94
|
7.51
|
X
|
8.37
|
23.95
|
|
10.2 bis
|
M/1 D
|
F3
|
X
|
X
|
9.06
|
X
|
X
|
7.95
|
X
|
|
14.2
|
M/1 D
|
F3
|
13.98
|
10.87
|
9.10
|
9.70
|
X
|
8.46
|
17.27
|
|
3374
|
M/1 D
|
B4 MA
|
14.14
|
11.83
|
8.42
|
8.72
|
X
|
8.15
|
15.11
|
|
328
|
M/1 D
|
C4 C4 f
|
X
|
X
|
X
|
8.63
|
X
|
X
|
12.60
|
|
352
|
P/4 D
|
C4 C4 f
|
9.46
|
8.63
|
7.43
|
5.94
|
X
|
6.52
|
14.34
|
|
3030
|
P/4 D
|
F3
|
10.27
|
9.62
|
6.98
|
5.75
|
X
|
5.97
|
17.46
|
|
246
|
P/4 D
|
C4 C4 e
|
11.03
|
9.63
|
7.82
|
6.81
|
X
|
6.87
|
16.40
|
|
14.2
|
P/4 D
|
F3
|
10.85
|
9.33
|
7.83
|
6.80
|
X
|
X
|
X
|
|
2097
|
P/4 D
|
MA B4
|
10.40
|
8.53
|
6.97
|
5.37
|
X
|
6.41
|
18.31
|
|
10.1
|
P/4 D
|
F3
|
11.37
|
X
|
7.77
|
5.98
|
X
|
X
|
20.23
|
|
3276
|
P/4 D
|
B5 M3
|
9.08
|
8.24
|
6.68
|
5.76
|
X
|
5.33
|
10.71
|
|
170
|
P/4 D
|
C4 C4 j
|
9.79
|
9.59
|
7.21
|
6.24
|
X
|
6.51
|
14.26
|
|
3276
|
P/4 D
|
B5 M3
|
9.30
|
8.80
|
6.69
|
5.73
|
X
|
5.24
|
11.08
|
|
281
|
P/4 D
|
D4 D4 f
|
9.37
|
X
|
7.43
|
6.98
|
X
|
X
|
X
|
|
13
|
P/4 D
|
F3
|
10.54
|
9.47
|
X
|
6.91
|
X
|
6.28
|
8.85
|
|
T 20
|
P/4 D
|
E4 EB1
|
X
|
X
|
X
|
4.58
|
X
|
X
|
X
|
|
16
|
P/4 D
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3.4
|
P/4 G
|
MA
|
10.96
|
9.20
|
X
|
5.58
|
X
|
6.17
|
X
|
|
?
|
P/4 G
|
C5 e
|
9.47
|
9.34
|
7.31
|
6.21
|
X
|
6.34
|
14.74
|
|
1.5
|
P/4 G
|
F4
|
11.20
|
X
|
7.63
|
6.31
|
X
|
X
|
17.18
|
|
7.2
|
P/4 G
|
F3
|
9.64
|
9.02
|
7.41
|
6.23
|
X
|
5.68
|
10.50
|
|
49
|
P/4 G
|
D4 D4 f
|
9.82
|
8.16
|
6.75
|
6.17
|
X
|
5.84
|
10.14
|
|
3247
|
P/4 G
|
B4 M
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3483
|
P/4 G
|
B5 MA3
|
9.54
|
8.65
|
7.48
|
5.91
|
X
|
5.13
|
9.42
|
|
1718
|
P/3 G
|
C5 e
|
7.97
|
X
|
5.20
|
X
|
X
|
X
|
X
|
|
14.2
|
|
F3
|
7.86
|
7.72
|
5.79
|
X
|
X
|
5.91
|
11.86
|
|
209
|
P/3 G
|
C4 C4 ei
|
7.45
|
7.50
|
6.41
|
X
|
X
|
5.57
|
10.47
|
|
16.1
|
P/3 G
|
F3
|
7.89
|
8.17
|
5.70
|
X
|
X
|
5.81
|
11.43
|
|
15.2
|
P/3 G
|
F3
|
8.22
|
8.03
|
6.19
|
X
|
X
|
5.92
|
12
|
|
1.5 bi
|
P/3 G
|
F4
|
8.52
|
X
|
5.16
|
X
|
X
|
X
|
12.73
|
|
240
|
P/3 G
|
F4 F4 f
|
7.50
|
X
|
5.86
|
X
|
X
|
X
|
10.72
|
|
10.1
|
P/3 D
|
F3
|
8.93
|
7.63
|
X
|
X
|
X
|
X
|
13.31
|
|
5
|
P/3 D
|
F4
|
8.34
|
8.32
|
6.01
|
X
|
X
|
6.52
|
11.72
|
|
1
|
P/3 D
|
F3
|
7.58
|
7.90
|
6.38
|
X
|
X
|
5.53
|
10.84
|
|
16
|
P/3 D
|
F3
|
8.30
|
X
|
5.95
|
X
|
X
|
X
|
X
|
|
3074
|
P/3 D
|
B5 MA
|
8.01
|
X
|
5.04
|
X
|
X
|
X
|
10.57
|
|
12
|
P/3 D
|
F3
|
7.45
|
7.66
|
5.71
|
X
|
X
|
5.31
|
9.50
|
|
T.19
|
P/2 G
|
E4 EB1
|
5.72
|
4.44
|
3.11
|
X
|
X
|
3.23
|
8.57
|
|
/
|
P/2 G
|
B4 f
|
5.94
|
5.45
|
2.55
|
X
|
X
|
4.22
|
9.80
|
|
/
|
P/2 G
|
B5 MA3
|
5.83
|
5.09
|
3.76
|
X
|
X
|
4.14
|
9.28
|
|
1627
|
P/2 G
|
C5f
|
5.81
|
5.09
|
5.79
|
X
|
X
|
3.96
|
9.72
|
|
/
|
P/2 G
|
C3 e
|
6.29
|
5.06
|
3.78
|
X
|
X
|
3.61
|
8.58
|
|
13.2
|
P/2 G
|
F3
|
5.25
|
4.81
|
4.12
|
X
|
X
|
4.27
|
8.57
|
|
/
|
P/2 G
|
C4 f
|
6.11
|
4.90
|
3.76
|
X
|
X
|
4.44
|
8.92
|
|
4
|
P/2 G
|
F4
|
6.58
|
5.47
|
3.91
|
X
|
X
|
4.85
|
9.06
|
|
17.1
|
P/2 G
|
F3
|
5.77
|
4.77
|
4.68
|
X
|
X
|
4.10
|
7.84
|
|
435
|
P/2 D
|
C4 C4 h
|
5.42
|
4.04
|
2.44
|
X
|
X
|
3.33
|
8.96
|
|
13
|
P/2 D
|
F3
|
5.62
|
4.74
|
3.21
|
X
|
X
|
3.65
|
9.73
|
|
1.2
|
P/2 D
|
F4
|
6.08
|
5.25
|
4.19
|
X
|
X
|
3.95
|
9.90
|
|
11.2
|
P/2 D
|
F3
|
5.69
|
4.76
|
3.16
|
X
|
X
|
3.69
|
9.83
|
|
T 214
|
P/2 D
|
C4 e
|
6.19
|
4.91
|
3.16
|
X
|
X
|
4.65
|
9.75
|
|
16
|
P/2 D
|
F3
|
6.71
|
5.84
|
4.04
|
X
|
X
|
4.43
|
9.75
|
|
14.2
|
P/2 D
|
F3
|
6.02
|
4.66
|
3.04
|
X
|
X
|
4.32
|
9.32
|
|
T.217
|
P/2 D
|
C4 e
|
5
|
4.27
|
3.07
|
X
|
X
|
4.20
|
9.16
|
|
12.1
|
P/2 D
|
F3
|
5.87
|
4.88
|
3.26
|
X
|
X
|
3.66
|
9.19
|
|
3309
|
P/2 D
|
MA B4
|
6.03
|
4.96
|
3.69
|
X
|
X
|
3.50
|
8.86
|
|
3213
|
M3/
|
B4 M
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
16.2
|
M3/ G
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
5w
|
M3/ G
|
B4 e
|
16.24
|
X
|
9.15
|
6.62
|
X
|
14.22
|
41.24
|
|
2058
|
|
B4 MA
|
X
|
23.53
|
X
|
X
|
X
|
X
|
X
|
|
IV
|
M3/ G
|
F3
|
X
|
X
|
10.34
|
9.22
|
X
|
X
|
X
|
|
2096
|
M3/ G
|
MA B4
|
17.19
|
24.53
|
9.98
|
7.46
|
X
|
15.38
|
42.09
|
|
33
|
M3/ G
|
MA A4
|
X
|
27.69
|
11.16
|
9.70
|
X
|
X
|
39.30
|
|
3081
|
M3/ G
|
B4 M
|
X
|
X
|
10.90
|
9.25
|
X
|
12.50
|
27.55
|
|
164
|
M3/ G
|
C4 C4 ei
|
24.02
|
X
|
12.14
|
10.79
|
X
|
12.58
|
19.09
|
|
17.2
|
M3/ G
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3134
|
M3/ G
|
F4
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3108
|
M3/ G
|
B5 MA
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
17.1
|
M3/ D
|
F3
|
16.67
|
X
|
9.37
|
7.33
|
X
|
X
|
X
|
|
257
|
M3/ D
|
C4 C4 e
|
X
|
X
|
9.30
|
X
|
X
|
X
|
X
|
|
12
|
M3/ D
|
F3
|
22.10
|
X
|
11.44
|
10.15
|
X
|
12.69
|
X
|
|
2078
|
M3/ D
|
MA B4
|
22.01
|
X
|
12.34
|
11.38
|
X
|
13.45
|
27.13
|
|
3178
|
M3/ D
|
B5 MA2
|
21.86
|
22.71
|
11.70
|
9.59
|
X
|
13.24
|
29.10
|
|
3293
|
M3/ D
|
B4 MA
|
18.83
|
21.44
|
X
|
8.79
|
X
|
13.96
|
36.75
|
|
1960
|
M3/ D
|
C4 e
|
20.09
|
23.10
|
11.36
|
9.40
|
X
|
13.63
|
28.80
|
|
3411
|
M3/ D
|
B5 MA3
|
23.91
|
24.01
|
13.24
|
11.56
|
X
|
13.71
|
12.29
|
|
124
|
M3/ D
|
E4E4 g
|
21.49
|
21.70
|
11.02
|
9.81
|
X
|
12.61
|
19.17
|
|
3100
|
M1-2/
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
10
|
M1-2/
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
406
|
M2/ G
|
C4 C4 h
|
18.34
|
X
|
9.67
|
7.54
|
X
|
X
|
X
|
|
2022
|
M2/ G
|
MA B4
|
17.71
|
11.83
|
10.22
|
8.57
|
X
|
13.20
|
34.71
|
|
3322
|
M2/ G
|
B4 MA
|
17.99
|
13.73
|
10.78
|
8.92
|
X
|
13.36
|
37.63
|
|
85
|
M2/ G
|
D5 D5 f
|
18.15
|
14.24
|
11.19
|
9.21
|
X
|
14.03
|
32.31
|
|
2098
|
M2/ G
|
MA B4
|
18.81
|
15.07
|
11.42
|
10.09
|
X
|
14.28
|
30.53
|
|
1.0
|
M2/ G
|
F4 F4 d
|
17.92
|
13.78
|
11.76
|
10.88
|
X
|
13.10
|
20.75
|
|
3527
|
M2/ G
|
B5 MA3
|
16.98
|
11.49
|
X
|
X
|
X
|
12.43
|
28.50
|
|
VIII
|
M2/ D
|
F3
|
18.26
|
X
|
7.46
|
5.97
|
X
|
X
|
X
|
|
1716
|
M2/ D
|
C4 e
|
19.21
|
X
|
8.16
|
6.19
|
X
|
X
|
X
|
|
2158
|
M2/ D
|
C3 e1
|
19.40
|
X
|
9.44
|
6.73
|
X
|
X
|
X
|
|
3155
|
M2/ D
|
B5 MA2
|
20.61
|
14.03
|
9.94
|
8.49
|
X
|
13.94
|
X
|
|
1717
|
M2/ D
|
C4 e
|
17.44
|
11.39
|
9.89
|
7.88
|
X
|
11.92
|
37.55
|
|
3318
|
M2/ D
|
B4 MA
|
17.47
|
X
|
10.60
|
X
|
X
|
X
|
38.19
|
|
15.2
|
M2/ D
|
F3
|
19.41
|
15.65
|
11.69
|
10.39
|
X
|
15.14
|
32.39
|
|
1006
|
M2/ D
|
C4 G
|
16.87
|
X
|
9.73
|
8.68
|
X
|
X
|
X
|
|
16.1
|
M2/ D
|
F3
|
17
|
X
|
13.03
|
11.34
|
X
|
X
|
X
|
|
1515
|
M1/ D
|
D4 f
|
16.44
|
X
|
7.61
|
6.39
|
X
|
X
|
X
|
|
52
|
M1/ D
|
E4 E4 h
|
16.21
|
X
|
8.05
|
6.27
|
X
|
X
|
X
|
|
2074
|
M1/ D
|
B4 MA
|
16.66
|
11.44
|
8.79
|
7.28
|
X
|
12.26
|
34.25
|
|
2417
|
M1/ D
|
B5 MA3
|
16.46
|
11.55
|
9.78
|
X
|
X
|
X
|
X
|
|
255
|
M1/ D
|
C4 C4 ei
|
15.82
|
11.26
|
11.60
|
9.39
|
X
|
12.16
|
23.15
|
|
16
|
M1/ D
|
A5 MA
|
15.19
|
X
|
11.77
|
X
|
X
|
X
|
|
|
3496
|
M1/ D
|
B5 MA3
|
X
|
12.87
|
10.25
|
X
|
X
|
X
|
13.67
|
|
2029
|
M1/ D
|
C4 e
|
15.38
|
10.86
|
12.10
|
12.05
|
X
|
13.09
|
14.59
|
|
6
|
M1/ D
|
F4
|
14.50
|
X
|
X
|
10.48
|
X
|
X
|
X
|
|
289
|
M1/ D
|
C4 C4 e
|
14.81
|
10.60
|
12.16
|
10.62
|
X
|
12.56
|
16.85
|
|
/
|
M1/ D
|
B5 MA3
|
13.24
|
X
|
11.65
|
10.47
|
X
|
X
|
12.55
|
|
3415
|
M1/ D
|
B5 MA3
|
14.80
|
X
|
12.70
|
10.92
|
X
|
X
|
13.49
|
|
coupe
|
M1/ G
|
C5 e
|
17.22
|
X
|
7.04
|
5.91
|
X
|
X
|
X
|
|
195
|
M1/ G
|
C4 C4 e1
|
17.51
|
19.45
|
6.46
|
4.98
|
X
|
13.40
|
26.76
|
|
VIII
|
M1/ D
|
F3
|
16.98
|
X
|
7.87
|
6.01
|
X
|
X
|
X
|
|
3076
|
M1/ G
|
B5 MA
|
X
|
14.86
|
X
|
6.27
|
X
|
X
|
X
|
|
3007
|
M1/ G
|
B4 M
|
16.10
|
X
|
X
|
X
|
X
|
X
|
X
|
|
R691?
|
M1/ G
|
B4 BRM
|
13.24
|
X
|
12.24
|
10.84
|
X
|
X
|
X
|
|
3147
|
P4/
|
B4 M
|
11.16
|
9.39
|
6.96
|
X
|
X
|
8.25
|
X
|
|
138
|
P4/ G
|
F4 F4 e
|
8.98
|
7.90
|
7.33
|
X
|
X
|
8.98
|
26.51
|
|
268
|
P4/ G
|
C4 C4 f
|
9.13
|
8.33
|
9.94
|
X
|
X
|
10.94
|
19.74
|
|
?
|
P4/ G
|
C4 C4 b
|
11.27
|
X
|
X
|
X
|
X
|
X
|
X
|
|
/
|
P4/ G
|
C3 e
|
X
|
X
|
9.66
|
X
|
X
|
11.03
|
25.24
|
|
3530
|
P4/ G
|
B5 MA3
|
9.34
|
7.80
|
8.35
|
X
|
X
|
8.62
|
21.14
|
|
2350
|
P4/ G
|
C3 e
|
9.30
|
6.87
|
8.33
|
X
|
X
|
5.95
|
26.03
|
|
P4/ D
|
F3
|
|
6.75
|
|
|
|
8.79
|
|
|
313
|
P4/ D
|
C4 C4 f
|
11.40
|
7.94
|
8.71
|
X
|
X
|
11.08
|
32.02
|
|
1.3
|
P4/ D
|
F4
|
9.56
|
6.50
|
8.79
|
X
|
X
|
8.69
|
25.33
|
|
3312
|
P4/ D
|
B4 MA
|
9.29
|
6.18
|
9.25
|
X
|
X
|
9.38
|
20.25
|
|
32
|
P4/ D
|
B5 MA
|
10.54
|
6.37
|
8.42
|
X
|
X
|
9.02
|
29.11
|
|
368
|
P3/ G
|
D4 E1
|
8.95
|
X
|
X
|
X
|
X
|
X
|
16.02
|
|
IV
|
P3/ G
|
F3
|
8.31
|
8.21
|
8.88
|
X
|
X
|
X
|
15.57
|
|
T.94
|
P3/ G
|
C4 C4 f R
|
8.49
|
X
|
8.32
|
X
|
X
|
X
|
X
|
|
3114
|
P3/ G
|
B4 M
|
8.89
|
6.65
|
8.33
|
X
|
X
|
7.12
|
13.28
|
|
R
|
P3/ G
|
F4 F4 ?
|
8.26
|
7.26
|
8.56
|
X
|
X
|
8.93
|
X
|
|
66
|
P3/ G
|
E4 E4 e
|
8.11
|
8.36
|
8.85
|
X
|
X
|
8.39
|
13.06
|
|
3070
|
P3/ G
|
B4 M
|
8.17
|
7.64
|
8.53
|
X
|
X
|
8.83
|
16.28
|
|
1795
|
P3/ D
|
C4 e
|
8.79
|
8.03
|
8.88
|
X
|
X
|
9.61
|
17.17
|
|
14.1
|
P3/ D
|
F3
|
8.13
|
7.23
|
8.61
|
X
|
X
|
8.33
|
15.62
|
|
3042
|
P3/ D
|
F3
|
8.20
|
X
|
9.10
|
X
|
X
|
X
|
X
|
|
1
|
P3/ D
|
F3
|
8.85
|
7.95
|
8.08
|
X
|
X
|
8.52
|
19.05
|
|
R
|
P3/ D
|
C5 e
|
7.91
|
7.09
|
8.17
|
X
|
X
|
9.23
|
22.27
|
|
2151
|
P3/ D
|
C4 e1
|
8.58
|
7.12
|
8.62
|
X
|
X
|
8.34
|
13.59
|
|
201
|
P2/
|
C4 C4 ei
|
6.22
|
4.75
|
4.56
|
X
|
X
|
2.44
|
16.31
|
|
119
|
P2/ G
|
F4 F4 f
|
7.13
|
6.18
|
6.49
|
X
|
X
|
5.86
|
11.68
|
|
1.3
|
P2/ G
|
F3
|
7.10
|
5.83
|
7.20
|
X
|
X
|
5.80
|
11.43
|
|
?
|
P2/ G
|
E4 E4 e
|
7.10
|
5.69
|
6.68
|
X
|
X
|
5.38
|
13.97
|
|
13.2
|
P2/ G
|
F3
|
6.56
|
6.39
|
X
|
X
|
X
|
X
|
13.93
|
|
/
|
P2/ G
|
B5 MA3
|
7.12
|
X
|
X
|
X
|
X
|
5.47
|
13.15
|
|
V
|
P2/ G
|
F3
|
7.31
|
6.14
|
7.13
|
X
|
X
|
5.58
|
10.41
|
|
/
|
P2/ D
|
B4 MA
|
6.83
|
6.32
|
6.94
|
X
|
X
|
6.37
|
X
|
|
3
|
D/2 D
|
F3
|
5.72
|
4.71
|
X
|
X
|
X
|
3.28
|
6.85
|
|
15.2
|
D/2 G
|
F3
|
5.12
|
4.53
|
X
|
X
|
X
|
3.14
|
|
|
11.2
|
D/2 D
|
F3
|
5.62
|
4.47
|
X
|
X
|
X
|
3.09
|
7.80
|
|
D/2 G
|
F4 h
|
5.18
|
3.93
|
X
|
X
|
X
|
2.52
|
6.26
|
|
D/2 G
|
D3-4 F
|
5.66
|
4.58
|
X
|
X
|
X
|
3.05
|
6.55
|
|
20
|
D/2 D
|
F4 F4 d
|
5.63
|
4.82
|
X
|
X
|
X
|
2.94
|
6.30
|
|
3
|
D/3 G
|
F3
|
10.28
|
8.19
|
4.61
|
X
|
X
|
4.97
|
7.13
|
|
D/3 G
|
B5
|
9.99
|
8.13
|
4.47
|
X
|
X
|
5.15
|
9.34
|
|
D/3 G
|
B5 MA3
|
8.97
|
7.49
|
5.14
|
X
|
X
|
4.10
|
6.74
|
|
15.1
|
D/3 G
|
F3
|
9.10
|
7.91
|
5.31
|
X
|
X
|
4.42
|
5.07
|
|
R
|
D/3 G
|
E4
|
8.55
|
7.19
|
4.18
|
X
|
X
|
3.83
|
6.02
|
|
D/3 D
|
B5 MA3
|
9
|
7.65
|
4.59
|
X
|
X
|
4.21
|
7.19
|
|
510
|
D/3 D
|
C4 e1
|
8.70
|
7.72
|
4.58
|
X
|
X
|
4.53
|
6.96
|
|
17.1
|
D/3 D
|
F3
|
9.61
|
7.42
|
4.71
|
X
|
X
|
X
|
7.49
|
|
D/3 D
|
B5 MA3
|
9.16
|
8.31
|
5.12
|
X
|
X
|
X
|
5.24
|
|
D/3
|
C4 e
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3252
|
D/3
|
B5 MA2
|
8.77
|
X
|
4.42
|
X
|
X
|
X
|
7.10
|
|
3
|
D/4 G
|
F3
|
18.43
|
X
|
5.12
|
6.35
|
6.55
|
X
|
9.06
|
|
2
|
D/4 G
|
F3
|
19.30
|
13.39
|
5.43
|
6.58
|
6.71
|
X
|
X
|
|
2355
|
D/4 G
|
C3 e
|
18.34
|
|
5.24
|
5.98
|
X
|
X
|
8.33
|
|
3329
|
D/4 G
|
B4 MA
|
19.19
|
15.37
|
5.79
|
6.91
|
7.12
|
7.55
|
9.75
|
|
3076
|
D/4 G
|
B5 MA
|
17.46
|
13.80
|
4.92
|
6.37
|
X
|
X
|
X
|
|
3429
|
D/4 G
|
B4 MA
|
17.44
|
14
|
5.28
|
6.81
|
6.91
|
6.85
|
7.59
|
|
R
|
D/4 G
|
E4
|
X
|
X
|
5.44
|
7.05
|
X
|
X
|
X
|
|
173
|
D/4 G
|
C4 C4 e1
|
X
|
X
|
5.34
|
X
|
X
|
X
|
X
|
|
1.2
|
D/4 G
|
F4
|
X
|
X
|
5.54
|
6.68
|
X
|
X
|
X
|
|
D/4 G
|
C5 e
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
15.2
|
D/4 G
|
F3
|
X
|
X
|
5.06
|
6.32
|
X
|
X
|
X
|
|
160
|
D/4 D
|
C4 C4 a
|
17.82
|
14.15
|
5.80
|
7.08
|
7.20
|
6.18
|
8.88
|
|
IV
|
D/4 D
|
F3
|
16.25
|
14.05
|
4.66
|
6.54
|
6.36
|
6.30
|
8.70
|
|
3397
|
D/4 D
|
B5 MA3
|
18.41
|
13.99
|
5.29
|
6.24
|
6.36
|
X
|
9.92
|
|
132
|
D/4 D
|
C4 C4 e
|
15.90
|
X
|
5.34
|
6.40
|
6.77
|
X
|
7.46
|
|
?
|
D/4 D
|
C4 C4 e1
|
20.30
|
15
|
5.25
|
5.77
|
5.31
|
X
|
11.70
|
|
D2/ G
|
C5 e
|
7.49
|
7.60
|
6.14
|
X
|
X
|
5.10
|
4.01
|
|
R
|
D2/ G
|
C4 e1
|
7.97
|
7.14
|
5.32
|
X
|
X
|
4.66
|
6.60
|
|
10.1
|
D2/ G
|
F3
|
7.67
|
7.16
|
5.63
|
X
|
X
|
4.27
|
6.27
|
|
D2/ G
|
B5 MA3
|
7.56
|
7.52
|
5.91
|
X
|
X
|
5.27
|
5.15
|
|
D2/ G
|
C3 e
|
8.48
|
7.71
|
5.31
|
X
|
X
|
5.07
|
7.53
|
|
718
|
D2/ D
|
C4 e1
|
7.77
|
7.50
|
5.47
|
X
|
X
|
4.60
|
6.78
|
|
VIII
|
D2/ D
|
F3
|
7.15
|
X
|
5.62
|
X
|
X
|
X
|
X
|
|
3254
|
D2/ D
|
B5 MA2
|
8.96
|
X
|
6.04
|
X
|
X
|
X
|
8.61
|
|
1825
|
D2/ D
|
C5 e
|
8.04
|
7.28
|
5.36
|
X
|
X
|
4.90
|
6.47
|
|
D2/ D
|
B4 MA
|
7.78
|
X
|
5.26
|
X
|
X
|
X
|
5.23
|
|
43
|
D3/ G
|
C4 C4 b
|
11.93
|
X
|
8.60
|
X
|
X
|
X
|
6
|
|
D3/ G
|
C3 e
|
13.09
|
10.35
|
7.28
|
X
|
X
|
9.54
|
10.60
|
|
184
|
D3/ G
|
E4 E4 K
|
14.19
|
9.41
|
7
|
X
|
X
|
9.61
|
10.40
|
|
D3/ G
|
MA 3-4
|
12.84
|
X
|
|
X
|
X
|
X
|
9.70
|
|
1756
|
D3/ G
|
C5 e
|
12.50
|
X
|
|
X
|
X
|
X
|
X
|
|
14 bis
|
D3/ D
|
F4
|
12.30
|
X
|
7.15
|
X
|
X
|
X
|
9.04
|
|
VIII
|
D3/ D
|
F3
|
12.39
|
10.15
|
8.54
|
X
|
X
|
9.93
|
7.25
|
|
3071
|
D3/ D
|
B4 M
|
13.60
|
|
7.34
|
X
|
X
|
X
|
10.87
|
|
1549
|
D4/ G
|
C4 e
|
14.13
|
10.82
|
X
|
X
|
X
|
11.51
|
11.60
|
|
3238
|
D4/ G
|
F3
|
14.64
|
X
|
9.34
|
X
|
X
|
X
|
X
|
|
IV
|
D4/ G
|
F3
|
15.05
|
11.21
|
9.88
|
X
|
X
|
12.21
|
12.46
|
|
183
|
D4/ G
|
E4 E4 K
|
14.91
|
X
|
7.97
|
X
|
X
|
X
|
15.40
|
|
3307
|
D4/ D
|
B5 MA3
|
12.19
|
9.89
|
9.77
|
X
|
X
|
11.75
|
10.51
|
|
9
|
D4/ D
|
C4 C4 b
|
14.13
|
X
|
8.80
|
X
|
X
|
X
|
11.31
|
|
VIII
|
D4/ D
|
F3
|
13.55
|
X
|
8.06
|
X
|
X
|
11.11
|
11.45
|
|
314
|
D4/ D
|
C4 C4 f
|
14.78
|
10.56
|
8.90
|
X
|
X
|
X
|
11.81
|
|
2007
|
D4/ D
|
MA B4
|
13.57
|
10.47
|
8.52
|
X
|
X
|
X
|
11.12
|
|
VII
|
D4/ D
|
F3
|
14.24
|
X
|
8.18
|
X
|
X
|
X
|
X
|
|
1715
|
D4/ D
|
C4 e
|
13.71
|
X
|
9.26
|
X
|
X
|
X
|
9.92
|
|
3425
|
D4/ D
|
B4 MA
|
14.83
|
X
|
7.51
|
X
|
X
|
X
|
X
|
|
D3 ou 4/
|
B5 MA3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
1520
|
D3 ou 4/
|
D4 f
|
X
|
X
|
8.59
|
X
|
X
|
X
|
X
|
|
14 bis
|
D3 ou 4/
|
F4
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
Annexe 15: Tableau d'inventaire et de mesures des
séries dentaires de C. ibex de l'ensemble
moustérien
|
N
|
Dents
|
Couche
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
1683
|
M/3 D
|
C5 e
|
23.17
|
26.23
|
7.08
|
5.94
|
X
|
8.77
|
43.79
|
|
1683 BIS
|
M/2 D
|
C5 e
|
17.17
|
14.18
|
8.17
|
7.69
|
X
|
8.87
|
36.12
|
|
1683
|
M/1 D
|
C5 e
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
1683 TER
|
P/4 D
|
C5 e
|
10.24
|
X
|
7.05
|
4.86
|
X
|
X
|
X
|
|
1684 TER
|
P/3 D
|
C5 e
|
8.54
|
X
|
4.74
|
X
|
X
|
X
|
X
|
|
2
|
M/2 G
|
F3
|
16.29
|
X
|
9.41
|
9.17
|
X
|
9.38
|
26.73
|
|
2
|
M/1 G
|
F3
|
13.49
|
11.52
|
8.47
|
9.24
|
X
|
7.19
|
11.95
|
|
fouille clandestine
|
M/3 D
|
C3 e
|
24.38
|
23.93
|
8.23
|
7.74
|
4.44
|
8.36
|
35.42
|
|
fouille clandestine
|
M/2 D
|
C3 e
|
16.54
|
13.16
|
8.97
|
8.75
|
X
|
8.23
|
27.99
|
|
fouille clandestine
|
M/1 D
|
C3 e
|
13.12
|
10.66
|
8.32
|
8.46
|
X
|
7.35
|
15
|
|
fouille clandestine
|
P/4 D
|
C3 e
|
9.53
|
7.69
|
6.96
|
5.49
|
X
|
5.71
|
15.96
|
|
fouille clandestine
|
P/3 D
|
C3 e
|
7.14
|
7.16
|
5.41
|
X
|
X
|
5.07
|
11.74
|
|
17.1
|
M/2 D
|
F3
|
15.84
|
X
|
7.76
|
7.22
|
X
|
X
|
X
|
|
17.2
|
P/4 D
|
F3
|
9.74
|
X
|
7.39
|
6.23
|
X
|
X
|
12.98
|
|
17.1
|
P/3 D
|
F3
|
7.69
|
X
|
5.70
|
X
|
X
|
X
|
11.12
|
|
17.1
|
P/3 G
|
F3
|
7.98
|
7.99
|
6.77
|
X
|
X
|
5.60
|
6.91
|
|
17.2
|
X
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
379
|
M/1 G
|
C4 C4 E1
|
13.83
|
14.22
|
5.21
|
4.78
|
X
|
7.97
|
24.03
|
|
379
|
D/4 G
|
C4 C4 E1
|
18.31
|
14.05
|
5.04
|
5.87
|
5.74
|
6.94
|
10.61
|
|
379
|
D/3 G
|
C4 C4 E1
|
9.31
|
7.61
|
4.56
|
X
|
X
|
5.01
|
7.02
|
|
II
|
M/2 D
|
F3
|
20.03
|
X
|
6.22
|
6.11
|
X
|
X
|
X
|
|
II
|
M/1 D
|
F3
|
17.06
|
13.67
|
7.05
|
6.14
|
X
|
9
|
29.83
|
|
II
|
D/4 D
|
F3
|
X
|
X
|
X
|
6.74
|
6.82
|
X
|
X
|
|
II
|
P/4 G
|
F3
|
11.35
|
9.48
|
7.64
|
5.92
|
X
|
7.01
|
14.28
|
|
21
|
M/3 G
|
F3
|
X
|
X
|
X
|
X
|
5.10
|
X
|
X
|
|
21
|
M/2 G
|
F3
|
16.74
|
12.67
|
9.06
|
8.77
|
X
|
X
|
X
|
|
21
|
M/1 G
|
F3
|
12.77
|
X
|
8.30
|
8.53
|
X
|
X
|
X
|
|
21
|
P/4 G
|
F3
|
9.84
|
8.80
|
7.04
|
5.75
|
X
|
6.21
|
16.06
|
|
16.2
|
M/3 D
|
F3
|
23.99
|
26.73
|
8.73
|
8.01
|
5.45
|
9.79
|
32.32
|
|
16.2
|
M/2 D
|
F3
|
16.94
|
9.71
|
9.33
|
X
|
X
|
X
|
X
|
|
16.2
|
M/1 D
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
195-7
|
D/4 G
|
F3
|
20.39
|
14.51
|
5.57
|
5.85
|
5.76
|
7.30
|
16.70
|
|
195-7
|
P/3 D
|
F3
|
8.80
|
8.57
|
5.71
|
X
|
X
|
6.06
|
X
|
|
195-7
|
D4/ G
|
F3
|
14.70
|
X
|
7.13
|
X
|
X
|
X
|
X
|
|
195-7
|
D3/ G
|
F3
|
13.28
|
X
|
5.82
|
5.69
|
X
|
X
|
X
|
|
195-7
|
P4/ D
|
F3
|
10.11
|
9.02
|
9.12
|
X
|
X
|
X
|
24.68
|
|
195-7
|
P2/ G
|
F3
|
6.94
|
6.45
|
6.62
|
X
|
X
|
X
|
10.44
|
|
195-7
|
I
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
195-7
|
I
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
195-7
|
I
|
F3
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3165
|
M/1 G
|
B5 MA2
|
17.14
|
12.58
|
6.56
|
6.64
|
X
|
8.13
|
34.55
|
|
3165
|
D/4 G
|
B5 MA2
|
17.69
|
13.99
|
4.88
|
6.24
|
6.64
|
6.77
|
7.77
|
|
3165
|
D/3 G
|
B5 MA2
|
9.18
|
X
|
5.22
|
X
|
X
|
X
|
6.56
|
|
3165
|
D/2 G
|
B5 MA2
|
5.66
|
4.75
|
3.48
|
X
|
X
|
3.02
|
6.43
|
|
8.2
|
P4/ D
|
F3
|
8.13
|
X
|
9.38
|
X
|
X
|
X
|
X
|
|
8.2
|
P3/ D
|
F3
|
7.78
|
X
|
8.57
|
X
|
X
|
X
|
X
|
|
8.2
|
P2/ D
|
F3
|
6.72
|
X
|
6.53
|
X
|
X
|
X
|
X
|
|
197-200
|
M/1 G
|
F3 I
|
16.88
|
X
|
6.08
|
5.45
|
X
|
X
|
29.24
|
|
197-200
|
P/3 G
|
F3 I
|
6.74
|
7.54
|
5.77
|
X
|
X
|
5.75
|
10.18
|
|
197-200
|
D/2 D
|
F3 I
|
5.34
|
X
|
2.73
|
X
|
X
|
X
|
6.63
|
|
197-200
|
P4/ D
|
F3 I
|
8.36
|
7.06
|
10.15
|
X
|
X
|
10.39
|
16.72
|
|
197-200
|
|
F3 I
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
197-200
|
|
F3 I
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
2021
|
P4/ G
|
MA B4
|
8.47
|
X
|
9.78
|
X
|
X
|
X
|
X
|
|
2021
|
P3/ G
|
MA B4
|
8.47
|
X
|
8.60
|
X
|
X
|
X
|
X
|
|
215
|
P/4 G
|
E4
|
9.74
|
X
|
6.94
|
6.36
|
X
|
X
|
11.65
|
|
215
|
P/3 G
|
E4
|
8.05
|
X
|
5.75
|
X
|
X
|
X
|
10.91
|
|
333
|
P4/ D
|
C4 C4 F
|
10.82
|
7.90
|
9.30
|
X
|
X
|
10.21
|
30.85
|
|
P3/ D
|
C4 C4 F
|
9.04
|
7.42
|
7.98
|
X
|
X
|
8.23
|
24.68
|
|
P2/ D
|
C4 C4 F
|
5.42
|
5.18
|
6.13
|
X
|
X
|
5.76
|
16.66
|
|
3153
|
M/1 D
|
B4 M
|
14.74
|
X
|
5.73
|
5.52
|
X
|
X
|
X
|
|
3153
|
D/4 D
|
B4 M
|
17.47
|
X
|
5.17
|
6.27
|
6.45
|
X
|
X
|
|
3153
|
D/3 D
|
B4 M
|
9.88
|
X
|
4.58
|
X
|
X
|
X
|
X
|
|
1814
|
M/1 D
|
C5 e
|
|
X
|
X
|
X
|
X
|
X
|
X
|
|
1814
|
D/4 D
|
C5 e
|
19.60
|
X
|
5.48
|
6.12
|
X
|
X
|
X
|
|
1814
|
D/3 D
|
C5 e
|
11.01
|
X
|
4.86
|
|
X
|
X
|
X
|
|
35
|
M1/ D
|
MA A4
|
18.49
|
X
|
7.79
|
5.33
|
X
|
X
|
X
|
|
35
|
D4/ D
|
MA A4
|
15.37
|
X
|
8.64
|
7.01
|
X
|
X
|
X
|
|
35
|
D3/ D
|
MA A4
|
|
X
|
6.92
|
7.39
|
X
|
X
|
X
|
|
35
|
D2/ D
|
MA A4
|
8.66
|
X
|
5.85
|
X
|
X
|
X
|
X
|
|
1789
|
M1/ G
|
C5 e
|
X
|
X
|
7.71
|
6.25
|
X
|
12.20
|
30.23
|
|
1789
|
D4/ G
|
C5 e
|
14.49
|
X
|
8.25
|
7.50
|
X
|
X
|
X
|
|
1789
|
D3/ G
|
C5 e
|
13.01
|
X
|
X
|
X
|
X
|
X
|
X
|
|
1789
|
D2/ G
|
C5 e
|
8.45
|
X
|
6.08
|
X
|
X
|
X
|
X
|
|
845
|
M1/ D
|
C4 C4 F
|
17.48
|
11.19
|
9.14
|
6.79
|
X
|
11.47
|
38.81
|
|
845
|
D4/ D
|
C4 C4 F
|
15.66
|
X
|
8.80
|
8.26
|
X
|
X
|
11.80
|
|
845
|
D3/ D
|
C4 C4 F
|
12.57
|
X
|
7.54
|
7.97
|
X
|
X
|
X
|
|
845
|
D2/ D
|
C4 C4 F
|
8.70
|
X
|
5.75
|
X
|
X
|
X
|
X
|
|
3509
|
M/2 D
|
B4 MA
|
18.26
|
X
|
5.87
|
5.03
|
X
|
X
|
X
|
|
3509
|
M/1 D
|
B4 MA
|
15.92
|
X
|
6.95
|
6.44
|
X
|
X
|
X
|
|
3089
|
M/3 D
|
MA B5
|
X
|
X
|
5.16
|
4.64
|
X
|
X
|
X
|
|
3089
|
M/2 D
|
MA B5
|
18.83
|
13.98
|
7.51
|
6.51
|
X
|
8.11
|
42.72
|
|
2067
|
M3/ D
|
MA B4
|
22.61
|
X
|
12.25
|
10.93
|
X
|
X
|
X
|
|
2067
|
M2/ D
|
MA B4
|
18.36
|
X
|
12.76
|
11.10
|
X
|
X
|
X
|
|
1887
|
M3/ D
|
C5 e
|
19.86
|
X
|
10.42
|
9.06
|
X
|
X
|
X
|
|
1887
|
M2/ D
|
C5 e
|
17.64
|
X
|
11.37
|
9.73
|
X
|
X
|
X
|
|
1887
|
P4/ D
|
C5 e
|
14.16
|
X
|
10.77
|
9.74
|
X
|
X
|
X
|
|
1887
|
P3/ D
|
C5 e
|
8.39
|
X
|
9.05
|
8.99
|
X
|
X
|
X
|
|
1887
|
P2/ D
|
C5 e
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3352
|
M3/ D
|
B4 MA
|
19.83
|
22.57
|
10.71
|
9.53
|
X
|
X
|
37.71
|
|
3352
|
M2/ D
|
B4 MA
|
18.72
|
X
|
12.03
|
10.32
|
X
|
X
|
34.84
|
|
SW
|
M/3 G
|
B4 f
|
25.79
|
X
|
9.68
|
9.57
|
6.65
|
X
|
X
|
|
SW
|
M/2 G
|
B4 f
|
15.71
|
X
|
10.33
|
9.91
|
X
|
X
|
X
|
|
SW
|
M/1 G
|
B4 f
|
11.06
|
X
|
8.03
|
8.31
|
X
|
X
|
X
|
|
SW
|
P/4 G
|
B4 f
|
8.93
|
X
|
4.89
|
6.92
|
X
|
X
|
X
|
|
142
|
M/3 D
|
D4 D4 e1
|
28.15
|
X
|
10.01
|
X
|
6.54
|
X
|
X
|
|
142
|
M/2 D
|
D4 D4 e1
|
16.88
|
X
|
10.27
|
X
|
X
|
X
|
X
|
|
142
|
M/1 D
|
D4 D4 e1
|
11.40
|
X
|
X
|
9.23
|
X
|
X
|
X
|
|
205
|
M/3 G
|
C4 C4 e
|
X
|
X
|
9.65
|
9.67
|
X
|
X
|
X
|
|
205
|
M/2 G
|
C4 C4 e
|
15.87
|
X
|
9.83
|
9.97
|
X
|
X
|
X
|
|
3137
|
M2/ D
|
B5 MA
|
17.46
|
X
|
11.94
|
11.06
|
X
|
X
|
X
|
|
3137
|
M1/ D
|
B5 MA
|
13.13
|
X
|
11.57
|
10.50
|
X
|
X
|
X
|
|
3137
|
P4/ D
|
B5 MA
|
7.47
|
X
|
9.81
|
X
|
X
|
X
|
X
|
|
3137
|
P3/ D
|
B5 MA
|
7.93
|
X
|
8.46
|
X
|
X
|
X
|
X
|
|
3137
|
P2/ D
|
B5 MA
|
6.29
|
X
|
6.25
|
X
|
X
|
X
|
X
|
|
3153
|
M/3 D
|
B5 MA2
|
28.26
|
X
|
10.09
|
9.80
|
5.89
|
X
|
X
|
|
3153
|
M/2 D
|
B5 MA2
|
15.64
|
X
|
9.99
|
10.03
|
X
|
X
|
X
|
|
3153
|
M/1 D
|
B5 MA2
|
10.27
|
X
|
7.64
|
8.56
|
X
|
X
|
X
|
|
3153
|
P/4 D
|
B5 MA2
|
9.95
|
X
|
6.98
|
7.15
|
X
|
X
|
X
|
|
3153
|
P/3 D
|
B5 MA2
|
7.91
|
X
|
6.04
|
X
|
X
|
X
|
X
|
|
138
|
D/4 G
|
F4 F4 e
|
17.88
|
X
|
5.04
|
5.64
|
5.76
|
X
|
X
|
|
138
|
D/3 G
|
F4 F4 e
|
10.07
|
X
|
4.54
|
X
|
X
|
X
|
7.49
|
|
3518
|
D/4 G
|
B4 MA
|
19.81
|
X
|
4.70
|
5.32
|
5.15
|
X
|
X
|
|
3518
|
D/3 G
|
B4 MA
|
X
|
X
|
X
|
X
|
X
|
X
|
7.81
|
|
9.1
|
D/4 G
|
F3
|
19.65
|
X
|
4.91
|
4.66
|
5.26
|
X
|
X
|
|
9.1
|
D/3 G
|
F3
|
10.86
|
8.58
|
5.06
|
X
|
X
|
4.72
|
9.40
|
|
2110
|
D4/ D
|
C4 e
|
14.69
|
X
|
X
|
X
|
X
|
X
|
X
|
|
2110
|
D3/ D
|
C4 e
|
12.90
|
X
|
X
|
X
|
X
|
X
|
X
|
|
2110
|
D2/ D
|
C4 e
|
7.63
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3196
|
D4/ D
|
F3
|
14.44
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3196
|
D3/ D
|
F3
|
12.59
|
X
|
X
|
X
|
X
|
X
|
X
|
|
208
|
D4/ D
|
E4 EB1
|
15.92
|
X
|
8.14
|
X
|
X
|
X
|
X
|
|
208
|
D3/ D
|
E4 EB1
|
13.74
|
X
|
7.37
|
X
|
X
|
X
|
X
|
Annexe 16: Tableau d'inventaire et de mesures des
incisives de C. ibex de l'ensemble moustérien
|
Numéro
|
Dents
|
Couche
|
DMD
|
DVL
|
|
17.1
|
I1 G
|
F3
|
7.70
|
|
|
202
|
I1 G
|
C4 C4 e
|
7.64
|
4.85
|
|
56
|
I1 G
|
E4 E4 h
|
7.50
|
4.66
|
|
3338
|
I1 G
|
B5 MA3
|
7.62
|
5.24
|
|
3312
|
I1 D
|
B5 MA3
|
8.67
|
4.84
|
|
1591
|
I1 D
|
D4 f
|
7.38
|
|
|
II
|
I1 D
|
F3
|
9.21
|
|
|
3442
|
I1 D
|
B4 MA
|
8.66
|
4.94
|
|
248
|
I1 D
|
C4 C4 e
|
7.15
|
4.97
|
|
17.43
|
I2 D
|
C5 e
|
7.26
|
5.13
|
|
3347
|
I2 D
|
B5 MA3
|
8.20
|
4.80
|
|
158
|
I2 D
|
E4 E4 j
|
6.45
|
4.48
|
|
16.2
|
I2 D
|
F3
|
7.15
|
|
|
191
|
I2 D
|
E4 E4 k
|
|
5.08
|
|
1689
|
I2 D
|
C5 e
|
|
5.33
|
|
III
|
I2 D
|
F3
|
7.15
|
4.39
|
|
269
|
I2 D
|
E4 E4 j
|
7.27
|
4.92
|
|
4
|
I2 D
|
F4
|
6.83
|
4.88
|
|
T.223
|
I2 G
|
C4 e
|
6.64
|
4.66
|
|
3519
|
I2 G
|
B5 MA3
|
6.04
|
4.21
|
|
2134
|
I2 G
|
C5 e
|
7.19
|
|
|
155
|
I2 G
|
F4 F4 f
|
6.92
|
4.46
|
|
3170
|
I2 G
|
B4 M
|
7.13
|
4.85
|
|
14 bis
|
I2 G
|
F4
|
7.40
|
4.64
|
|
3469
|
I2 G
|
B5 MA3
|
7.39
|
4.39
|
|
12.2
|
I2 G
|
F3
|
6.48
|
4.97
|
|
16
|
I3 G
|
D4 D4 f
|
8.20
|
3.99
|
|
3399
|
I3 G
|
B4 MA
|
5.64
|
|
|
905
|
I3 G
|
C4 C4 f
|
5.86
|
4.06
|
|
1643
|
I3 G
|
C5 e
|
6.61
|
3.36
|
|
546
|
I3 G
|
C4 C4 ei
|
6.56
|
3.88
|
|
2354
|
I3 G
|
C3 e
|
6.07
|
5.61
|
|
166
|
I3 G
|
E4 E4 j
|
7.18
|
4.08
|
|
73
|
I3 D
|
E4 E4 d
|
6.29
|
4.34
|
|
3395
|
I3 D
|
B4 MA
|
6.94
|
4.68
|
|
1.3
|
I3 D
|
F4
|
6.28
|
3.46
|
|
536
|
I3 D
|
E4 E4 I
|
6.63
|
4.44
|
|
I3 D
|
B5 MA3
|
6.37
|
4.59
|
|
sw
|
I3 D
|
B4 f
|
6.24
|
4.41
|
|
I3 D
|
B4 e
|
|
|
|
1598
|
I3 D
|
C5 e
|
5.74
|
5.21
|
|
3133
|
I3 D
|
F4
|
7.15
|
2.84
|
|
I
|
I4 G
|
F5
|
5.75
|
3.29
|
|
1797
|
I4 G
|
C5 e
|
5.95
|
3.42
|
|
1803
|
I4 G
|
C5 e
|
5.70
|
3.45
|
|
sw
|
I4 G
|
B4 e
|
5.92
|
3.52
|
|
16
|
I4 G
|
F3
|
6.67
|
3.28
|
|
II
|
I4 G
|
F3
|
6
|
3.55
|
|
3380
|
I4 G
|
B4 MA
|
6.52
|
3.67
|
|
1.3
|
I3 D
|
F4
|
6.44
|
4
|
|
1522
|
I4 D
|
C4 f
|
6.04
|
3.71
|
|
3323
|
I4 D
|
B4 MA
|
5.92
|
4.03
|
|
130
|
I4 D
|
C4 C4 e
|
7.10
|
3.69
|
|
13.1
|
I4 D
|
F3
|
6.51
|
3.59
|
|
1.3
|
I4 D
|
F4
|
6.45
|
4.10
|
|
I4 D
|
C5 e
|
6.28
|
|
|
14
|
I G
|
F3
|
6.13
|
|
|
3310
|
I G
|
B4 MA
|
6.15
|
4.05
|
|
I
|
I G
|
F3
|
5.89
|
5.14
|
|
28
|
I G
|
D4 D4 f
|
5.96
|
4.70
|
|
1540
|
I G
|
C4 e
|
5.25
|
4.53
|
|
IV
|
I G
|
F3
|
5.90
|
3.97
|
|
11
|
I G
|
E4 E4 e
|
6.47
|
5.19
|
|
3022
|
I G
|
B5 MA
|
4.82
|
4.79
|
|
I G
|
MA 3-4
|
7.15
|
4.07
|
|
282
|
I G
|
C4 C4 e
|
5.86
|
3.45
|
|
368
|
I G
|
E4 E4 I
|
5.70
|
4.25
|
|
17.1
|
I4 G
|
F3
|
6.46
|
|
|
I G
|
B4 MA
|
6.34
|
|
|
3188
|
I G
|
F3
|
5.65
|
4.40
|
|
I D
|
B5 MA3
|
6.33
|
4.23
|
|
I D
|
B5 MA3
|
5.09
|
4.49
|
|
184
|
I D
|
C4 C4 e1
|
4.56
|
4.88
|
|
82
|
I D
|
E4 E4 e
|
5.02
|
4.21
|
|
I D
|
B5 MA3
|
5.31
|
5.43
|
|
7.2
|
I D
|
F3
|
4.72
|
4.31
|
|
I D
|
B5 MA3
|
5.29
|
4.79
|
|
?
|
I D
|
C4 C4 f
|
5.61
|
|
|
T.28
|
DI G
|
D4
|
4.04
|
2.69
|
|
3179
|
DI G
|
B5 MA2
|
4.79
|
2.45
|
|
DI G
|
MA 3-4
|
4.96
|
3.59
|
|
sw
|
DI G
|
B4 e
|
5.05
|
3.03
|
|
sw
|
DI G
|
B4 f
|
5.88
|
3.43
|
|
3327
|
DI G
|
B5 MA3
|
6.35
|
3.35
|
|
3199
|
DI D
|
B4 M
|
5.94
|
3.42
|
|
1.2
|
DI D
|
F4
|
4.33
|
2.97
|
|
1.2
|
DI D
|
F4
|
4.39
|
|
|
DI D
|
MA 3-4
|
5.09
|
3.08
|
|
113
|
DI D
|
AM A5
|
5.40
|
3.01
|
|
3058
|
DI D
|
B4 M
|
4.78
|
3.51
|
|
3258
|
DI D
|
B5 MA2
|
4.63
|
3.03
|
|
3311
|
DI D
|
B5 MA3
|
4.57
|
2.89
|
|
13.1
|
DI D
|
F3
|
5.04
|
3.49
|
|
3223
|
DI D
|
B5 MA
|
4.46
|
2.72
|
|
XI
|
DI
|
F3
|
|
3.55
|
|
V
|
I G
|
F3
|
6.14
|
4.86
|
|
1.2
|
I G
|
F4
|
6.23
|
5.26
|
|
1898
|
I G
|
C5 e
|
|
|
|
I4 G
|
A5 AM
|
5.69
|
|
|
940
|
I G
|
C4 C4 g
|
6.11
|
|
|
498
|
I G
|
C4 C4 ei
|
|
|
|
1601
|
I G
|
C5 e
|
|
|
|
3136
|
I2 D
|
B5 MA
|
7.24
|
4.08
|
|
497
|
I3 D
|
F4 c
|
5.96
|
4.28
|
|
3379
|
I3 D
|
B4 MA
|
6.93
|
3.81
|
|
1529
|
I
|
C4 f
|
5.95
|
|
|
1.6
|
I
|
F4
|
|
|
|
372
|
I
|
D4 e1
|
6.25
|
|
|
V
|
I
|
F3
|
|
5.25
|
|
64
|
I
|
E4 E4 1
|
4.99
|
3.78
|
|
882
|
I
|
C4 C4 f
|
4.18
|
3.62
|
|
I
|
C4 e
|
4.35
|
4.40
|
|
891
|
I
|
C4 C4 f
|
5.14
|
4.04
|
|
5400
|
I
|
B5 MA 4
|
4.83
|
5.06
|
|
IV
|
I
|
F3
|
4.96
|
3.89
|
|
I
|
B4 MA
|
5.03
|
4.53
|
|
82
|
I
|
F4 F4 f
|
5.50
|
4.15
|
Annexe 17: Tableau d'inventaire et de mesures des dents
jugales isolées de C. ibex de l'ensemble
épipaléolithique
|
N°
|
Dents
|
Couche
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
23
|
M/3 D
|
C4 C4 d
|
X
|
X
|
6.96
|
6.38
|
X
|
X
|
48.44
|
|
?
|
M/2 G
|
C4 C4 d
|
19.13
|
X
|
X
|
7.87
|
X
|
X
|
X
|
|
25
|
M/2 G
|
C4 C4 d
|
20.28
|
X
|
7.85
|
7.83
|
X
|
X
|
X
|
|
33
|
M/2 D
|
C4 C4 d
|
18.28
|
14.45
|
8.39
|
7.76
|
X
|
8.52
|
41.09
|
|
101
|
M/1 G
|
D5 D5 d
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
3222
|
M/1 D
|
B4 a
|
13.42
|
X
|
8.19
|
8.42
|
X
|
X
|
X
|
|
24
|
P/4 G
|
B4 a
|
10.84
|
9.87
|
7.10
|
6.20
|
X
|
6.23
|
19.39
|
|
1
|
P/2 G
|
B5
|
5.76
|
5.04
|
3.19
|
X
|
X
|
3.49
|
10.16
|
|
1
|
P/2 G
|
B5
|
5.93
|
5.05
|
5.06
|
X
|
X
|
3.93
|
8.89
|
|
1543
|
P/2 G
|
C5 d
|
5.95
|
4.52
|
5.09
|
X
|
X
|
4.18
|
8.90
|
|
2
|
P/2 D
|
B5
|
5.89
|
4.92
|
2.42
|
X
|
X
|
3.89
|
8.46
|
|
623
|
M3/ G
|
C3 C3 c
|
15.56
|
22.48
|
8.01
|
5.80
|
X
|
13.06
|
X
|
|
28
|
M3/ G
|
C4 C4 d
|
19.99
|
X
|
10.86
|
9.45
|
X
|
13.20
|
32.46
|
|
63
|
M3/ D
|
A5 A
|
17.95
|
X
|
10.20
|
8.45
|
X
|
X
|
X
|
|
41
|
M3/ D
|
A5 A
|
20.08
|
19.93
|
10.60
|
9.63
|
X
|
13.39
|
29.02
|
|
405
|
M2/ G
|
C3 C3 c
|
17.98
|
17.07
|
8.77
|
5.90
|
X
|
12.81
|
32.44
|
|
622
|
M2/ G
|
C3 C3 c
|
18.17
|
14.92
|
10.30
|
8.51
|
X
|
13.78
|
35.62
|
|
210
|
M2/ D
|
A4 R
|
17.75
|
X
|
8.03
|
5.76
|
X
|
X
|
X
|
|
1842
|
M2/ D
|
C5 d
|
19.28
|
13.59
|
10.94
|
9.77
|
X
|
12.41
|
33.49
|
|
210
|
M2/ D
|
A4 R
|
18.53
|
X
|
11.90
|
10.26
|
X
|
X
|
X
|
|
620
|
P4/ G
|
C3 C3 c
|
10.30
|
7.95
|
8.53
|
X
|
X
|
10.43
|
23.87
|
|
59
|
P4/ D
|
C4 C4 d
|
9.86
|
7
|
8.20
|
X
|
X
|
10.03
|
26.33
|
|
608
|
P4/ D
|
C3 C3 c
|
10.64
|
8.31
|
9.34
|
X
|
X
|
12.23
|
29.21
|
|
3046
|
P3/ D
|
B5 A
|
8.14
|
7.84
|
9.07
|
X
|
X
|
9.04
|
9.87
|
|
210
|
P2/ D
|
A4 R
|
7.76
|
X
|
6.25
|
X
|
X
|
X
|
X
|
|
164
|
P2/ D
|
D5 D5 d
|
6.20
|
5.94
|
5.88
|
X
|
X
|
4.59
|
8.19
|
|
2
|
D/2 G
|
B5
|
5.46
|
4.76
|
X
|
X
|
X
|
3.06
|
5.76
|
|
2
|
D/2 D
|
B5
|
5.07
|
4.48
|
X
|
X
|
X
|
2.77
|
6
|
|
104
|
D/3 G
|
C4 C4 d
|
10.22
|
X
|
5.21
|
X
|
X
|
X
|
7.55
|
|
11
|
D/3 D
|
A4 A
|
9.85
|
|
5.49
|
X
|
X
|
X
|
7.21
|
|
74
|
D/4 D
|
C4 d
|
17.94
|
X
|
X
|
6.03
|
X
|
X
|
X
|
|
1
|
D2/ D
|
B5
|
7.72
|
X
|
5.80
|
X
|
X
|
X
|
3.63
|
|
1732
|
D3/ G
|
C5 d
|
12.69
|
10.14
|
8.43
|
X
|
X
|
9.75
|
7.24
|
|
2
|
D3/ D
|
B5
|
12.30
|
X
|
8.45
|
X
|
X
|
X
|
6.79
|
|
1
|
D4/ D
|
B5
|
14.93
|
10.16
|
8.48
|
X
|
X
|
X
|
14.26
|
|
3004
|
D3 ou 4/
|
B5
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
Annexe 18: tableau d'inventaire et de mesures des
incisives de C. ibex de l'ensemble
épipaléolithique
|
Numéro
|
Dents
|
Couche
|
DMD
|
DVL
|
|
3819
|
I1 G
|
B4 A
|
|
4.98
|
|
sw
|
I2 D
|
B4 a
|
7.40
|
4.64
|
|
210
|
I3 D
|
A4 R
|
6.83
|
4.07
|
|
I4 D
|
B5
|
6.43
|
|
|
163
|
I G
|
D5 D5 d
|
5.16
|
4.77
|
|
3000
|
I D
|
B5 A
|
5.30
|
4.55
|
|
4
|
DI G
|
A5 A
|
5.22
|
2.80
|
|
701
|
DI G
|
C4 C4 c
|
4.71
|
3.69
|
|
3009
|
DI
|
B4 A
|
|
|
|
210
|
I3
|
A4 R
|
|
4.55
|
|
8
|
I
|
D4 D4 c
|
4.96
|
3.64
|
Annexe 19: tableau d'inventaire et de mesures des dents
jugales de C. ibex de la zone R (moustérien?)
|
N
|
Dents
|
Couche
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
422
|
M/2 G
|
R
|
15.99
|
13.37
|
9.06
|
8.35
|
X
|
8.95
|
26.86
|
|
?
|
M/2 D
|
R
|
16.89
|
X
|
5.52
|
4.89
|
X
|
X
|
X
|
|
361
|
M/2 D
|
R 72
|
19.21
|
15.46
|
8.51
|
7.62
|
X
|
9.45
|
38.26
|
|
1512
|
M/2 D
|
F6 c
|
17.29
|
13.90
|
9.43
|
8.75
|
X
|
8.05
|
32.17
|
|
384
|
M/1 G
|
R 72
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
363
|
M/1 G
|
R 72
|
14.45
|
X
|
8.64
|
8.81
|
X
|
X
|
X
|
|
429
|
M/1 G
|
R
|
11.84
|
10.73
|
8.69
|
9.74
|
X
|
6.69
|
6.77
|
|
T55
|
P/4 G
|
F4 d R
|
11.21
|
10.04
|
6.13
|
4.86
|
X
|
6.69
|
9.90
|
|
?
|
P/4 G
|
R?
|
9.75
|
X
|
6.86
|
4.67
|
X
|
X
|
X
|
|
P/3 G
|
R
|
8.64
|
7.38
|
3.81
|
X
|
X
|
5.13
|
15.40
|
|
P/3 D
|
C4 C4 f
|
8.13
|
X
|
X
|
X
|
X
|
X
|
X
|
|
1511
|
P/2 D
|
F6 c
|
5.69
|
4.63
|
3.67
|
X
|
X
|
3.50
|
8.88
|
|
R 24
|
M3/ G
|
Remanié
|
14.92
|
21.12
|
7.22
|
6.04
|
X
|
13.93
|
X
|
|
R 23
|
M2/ G
|
|
17.35
|
14.02
|
9.75
|
8.18
|
X
|
12.78
|
41.34
|
|
372?
|
M2/ D
|
R 72
|
18.22
|
11.09
|
11.16
|
9.50
|
X
|
13.45
|
35.75
|
|
1515
|
M1/ D
|
F6 c
|
15.76
|
X
|
7.79
|
6.35
|
X
|
X
|
X
|
|
1523
|
M1/ G
|
F6 c
|
17.73
|
14.89
|
9.33
|
6.95
|
X
|
12.70
|
27.93
|
|
?
|
M1/ G
|
F4 R
|
16.59
|
11.34
|
10.55
|
9.70
|
X
|
12.80
|
29.53
|
|
22
|
M1/ G
|
R
|
16.46
|
10.38
|
10.41
|
9.69
|
X
|
13
|
30.35
|
|
/
|
M1/ G
|
|
16.38
|
12.67
|
10.83
|
10.13
|
X
|
13.30
|
24.01
|
|
T.58
|
P4/ G
|
C4 R
|
10.56
|
X
|
8.41
|
X
|
X
|
X
|
X
|
|
R 20
|
P4/ G
|
|
8.41
|
6.74
|
9.04
|
X
|
X
|
9.09
|
20.81
|
|
R 21
|
P4/ G
|
|
9.39
|
X
|
8.06
|
X
|
X
|
X
|
X
|
|
R.18
|
P3/ G
|
|
7.67
|
7.22
|
7.71
|
X
|
X
|
8.41
|
22.46
|
|
R.5
|
P2/ G
|
|
6.39
|
5.80
|
6.53
|
X
|
X
|
6.20
|
10.31
|
|
R.7
|
P2/ G
|
|
8
|
7.26
|
5.60
|
X
|
X
|
7.30
|
6.84
|
|
D/2 D
|
R4
|
4.90
|
4.36
|
X
|
X
|
X
|
2.90
|
6.76
|
|
T.54
|
D/3 D
|
C4 e1 R
|
9.40
|
7.95
|
4.95
|
X
|
X
|
4.43
|
6.68
|
|
T.42
|
D/3
|
E4 R
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
393
|
D/4 D
|
R 72
|
16.89
|
X
|
4.85
|
6.05
|
6.06
|
X
|
8.89
|
|
R.19
|
D3/ G
|
|
13.02
|
X
|
8.89
|
X
|
X
|
X
|
8.36
|
|
T.52
|
D3/ D
|
C4 a R
|
11.71
|
9.73
|
7.65
|
X
|
X
|
9.05
|
8.68
|
Annexe 20 : tableau d'inventaire et de mesures des
séries dentaires de C. ibex de la zone R
(moustérien?)
|
Numéro
|
Dents
|
Couche
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
R
|
M/2 D
|
/
|
15.34
|
X
|
7.32
|
7.30
|
X
|
X
|
30.31
|
|
R
|
M/1 G
|
/
|
13.32
|
10.62
|
6.28
|
6.09
|
X
|
7.51
|
18.54
|
|
R
|
P/4 D
|
/
|
11.68
|
9.91
|
5.80
|
4.98
|
X
|
7.13
|
21.49
|
|
R
|
P4/ G
|
/
|
9.83
|
6
|
7.37
|
X
|
X
|
6.68
|
24.61
|
|
210
|
M/1 D
|
R4R
|
16
|
|
6
|
5.37
|
|
|
|
|
210
|
D/4 D
|
R4R
|
18.24
|
|
5.37
|
6.44
|
6.59
|
|
|
|
210
|
D/3 D
|
R4R
|
9.68
|
|
4.79
|
|
|
|
|
Annexe 21 : tableau d'inventaire et de mesure des
incisives de C. ibex de la zone R (moustérien ?)
|
Numéro
|
Dents
|
Couche
|
DMD
|
DVL
|
|
198
|
I1 G
|
F6 c
|
8.10
|
|
|
T.60
|
I1 G
|
C4 R
|
7.91
|
|
|
1522
|
I1 G
|
F6 c
|
7.80
|
4.96
|
|
1514
|
I1 D
|
F6 c
|
8.26
|
|
|
392
|
I1 D
|
R
|
8.31
|
|
|
386
|
I2 D
|
R
|
6.81
|
4.69
|
|
387
|
I2 D
|
R 72
|
6.76
|
4.90
|
|
424
|
I3 D
|
R
|
7.27
|
4.50
|
|
I4 G
|
R
|
6.62
|
3.71
|
|
1506
|
I4 G
|
F6 c
|
6.58
|
3.20
|
|
395
|
I G
|
R
|
6.66
|
4.75
|
|
T.62
|
DI G
|
C4 R
|
4.94
|
3.29
|
|
I G
|
C5 e R
|
5.91
|
4.39
|
|
1502
|
I D
|
F6 c
|
5.61
|
|
Bibliographie
- Abassi M., Desclaux E., 1996, Arvicola
lacepede, 1799 (Rodentia, Mammalia) de
quatre séquences du Sud-est de la France et de Ligurie datant de la fin
du Pléistocène moyen et du début du
Pléistocène supérieur,
Quaternaire, t. 7 (1), p. 29-37
- Annequin-Jourdain C., 2004, Atlas culturel des
Alpes occidentals: de la Préhistoire à la fin du Moyen
Age, Paris, édition Picard, 439 p.
- Bazile-Robert E., 1979, Flore et
végétation du sud de la France pendant la dernière
glaciation d'après l'analyse anthracologique, Thèse
de 3e cycle, Université des sciences et techniques du Languedoc,
Montpellier, 154 p
- Behrensmeyer, A.K, 1978, Taphonomic and ecologic information
from bone weathering, Paleobiology, vol.4 (2), p.
150-162
- Bignon O., Chasser les chevaux à la fin du
Paléolithique dans le Bassin parisien : Stratégie
cynégétique et mode de vie au Magdalénien et à
l'Azilien ancien, Oxford, édition Archeopress, 170 p.
(British archaeological Reports International, 1747)
- Brugal J.-P, Jaubert J., 1991, Les gisements
paléontologiques pléistocènes à indices de
fréquentation humaine : un nouveau type de comportement de
prédation ?, Paléo, n°3, p.
15-41
- Couturier M., 1962, Le bouquetin des
Alpes, Grenoble, édition Arthaud, 1564 p.
- Couturier M., 1958, Parallèle anatomique physiologique
et écologique entre le pied du
bouquetin des Alpes (Capra aegagrus ibex ibex) et celui du
chamois (Rupicrapra rupicapra) en rapport avec l'adaptation à la
montagne de ces deux espèces, Mammalia, t. 22
(1), p.76-89
- Crégut-Bonnoure E., 1979, La faune de
mammifères du Pléistocène moyen de la Caune de l'Arago
à Tautavel (Pyrénées Orientales, Marseille,
Travaux du laboratoire de Paléontologie humaine et de
Préhistoire, 3 p.
- Crégut-Bonnoure E., 1992, Intérêts
biostratigraphique de la morphologie dentaire de Capra
(Mammalia, Bovidae), Annales Zoologici
Fennici, n°28, p.273-290
- Crégut-Bonnoure E, 2002, Les Ovibovini,
Caprini et Ovini (Mammalia, Artiodactyla, Bovidae, Caprinae) du
Plio-Pléistocène d'Europe : systématique, évolution
et biochronologie, Mémoire de Doctorat, Université
Claude-Bernard-Lyon I, 1 vol. 429 p. texte, 1 vol. 686
fig. et XXXIII pl., 1 vol. 212 tabl.
- Crégut-Bonnoure E., 2005/2006, Nouvelles données
paléogéographiques et chronologiques sur les Caprinae
(Mammalia, Bovidae) du Pléistocène moyen et
supérieur d'Europe, Antropologia, vol.57 (1),
p. 205-219
- Defleur A., Crégut-Bonnoure E. (dir.), 1995,
Le gisement Paléolithique moyen de la grotte des
Cèdres (Var), Paris, édition Maisons des Sciences
de l'Homme, 192 p. (Documents d'Archéologie Française, 49)
- De Lumley H., 1965, Le paléolithique
ancien et moyen du midi méditerranéen dans son cadra
géologique (Ligurie, Provence, Bas-Languedoc, Rousillon,
Catalogne), Mémoire de doctorat, Faculté des
Sciences de l'Université de Paris, vol.1, 391 p.
- Fernandez P., 2009, De l'estimation de l'âge individuel
dentaire au modèle descriptif des
structures d'âge des cohortes fossiles,
Bulletin de la société préhistorique
française, t. 106 (1), p. 5-14
- Fernandez P., 2006, Etude paléontologique
des ongulés du gisement moustérien du Bau de l'Aubesier
(Vaucluse, France) : morphométrie et contexte
biochronologique, Lyon, Editions de l'U.F.R des Sciences de la
Terre, 231p., 86 fig., 213 tabl., 6 pl. (Documents des Laboratoires de
Géologie, 161)
- Gerber J.-P., 1973, La faune des grands mammifères du
Würm ancien dans le Sud-Est de la France, Université de Provence,
Marseille Centre Saint Charles, 310 p. (Travaux du laboratoire de
Géologie historique et de Paléontologie ; 5)
- Gonzalez G., 1994, Variations saisonnières dans
l'utilisation du milieu par le bouquetin
ibérique selon l'âge et le sexe,
Travaux scientifique du Parc de la Vanoise, XVIII,
p.151-174
- Greenfield H. J., Arnold E.R, 2008, Absolute age and tooth and
wear sequences in sheep and goat : determining age-at-death in zooarchaeology
using a modern control sample, Journal of Archaeological
Science, vol.35, p. 836-849
- Habermehl K.H., 1985, Altersbestimmung bei Wild -
und Pelztieren : Moglichkeiten und
Methoden : ein praktischer Leitfaden fur
Jager, Hambourg, édition Paul Parey, 223p.
- Lavocat R., 1966, « Faune et flores préhistorique
de l'Europe occidentale », Atlas de
Préhistoire, Paris,
édition N. Boubée, 486p., 137pl., 33 tabl. (« L'Homme et ses
origine », 3)
- Lyman R. L., 1984, Bone density and differential survivorship
of fossil classes, Journal of
Anthropological Archaeology, vol.3, p.
259-299
- Lyman R.L, 1987, On the analysis of vertebrate mortality
profiles : sample size, mortality
type, and hunting preasure, American
Antiquity, vol.16, p. 125-142
- Moncel M.-H, Brugal J.-P, Prucca A., Lhomme G., 2008, Mixed
occupation during the Middle
Palaeolithic : Case of a small pit-cave_site of Les
Pêcheurs (Ardèche, south-eastern France),
International journal of osteoarchaeology, vol.27,p.
382-398
- Perez Ripoll M., 1988, Estudio de la sequencia del desgaste de
los molars de la «capra
pyrenaica» de yacimientos prehistoricos, Archivo
de Prehistoria Levantina, vol. XVIII, p.83-127
- Pillard B., 1970, La faune de grands
mammifères de la grotte moustérienne de l'Hortus
(Valfaunès, Hérault)
Interprétations paléontologique et
palethnographique, Mémoire de doctorat, Faculté des
Sciences de l'Université de Paris, 147 p.
- Porraz G., 2004, Dans l'ombre des plus grands : les sites
moustériens de l'abri Pié Lombard
(Alpes-Maritimes, France) et de la grotte du Broion
(Vénitie, Italie). Présentation de leurs industries lithiques,
in : Un siècle de construction du discours scientifique en
préhistoire. Volume III « ... Aux conceptions d'aujourd'hui »,
Avignon, 21-25 septembre 2004, J. Evin (Dir.), Paris,
Société préhistorique française, p.237-249
- Renault-Miskovsky J., Texier P.J., 1980, Intérêt
de l'analyse pollinique détaillée dans les
concrétions de grotte, Application à l'Abri
Pié-Lombard (Tourettes-sur-Loup, Alpes-
Maritimes), Bulletin de l'Association
française pour l'Etude de Quaternaire, t. 3, p. 129-134
- Rivals F., 2004, Les petits bovidés (Caprini
et Rupicaprini) pléistocène dans le bassin
méditérranéen et le Caucas : Etude paléontologique,
biostratigraphique, archéozoologique et
paléoécologique, Oxford, Editions Archeopress, 252
p. (British archaeological Reports International, 1327)
- Stiner M.C, 1991, Human predators and prey
mortality, Boulder, Westvieew Press, 276.p
- Texier P.-J., 1974, L'Industrie moustérienne de l'abri
Pié-Lombard, Tourettes-sur-Loup, Alpes-
Maritimes, Bulletin de la Société
préhistorique française, t. 71 (2), p. 429-448
- Texier P.-J., 1996, Abri Pié Lombard, Tourettes-sur-Loup
(Alpes-Maritimes), Opération de
sondage du 8 au 14 juin 1996, Autorisation DRACAR, n° 108,
10 p.
- Tillet T., 2000, Le moustérien dans l'arc Alpin :
complémentarité entre sites en grotte et sites
en plein-air, in : Les Paléoalpins - hommage
à Pierre Bintz, Grenoble, édition Université
Joseph Fourier, p.99-106 (Géologie alpine - Mémoire h.s, 31)
- Toigo C., Michallet J., Blanc D., 1995, Structure des groupes
chez le bouquetin des Alpes
(Capra ibex ibex) dans le massif de Belledonne-sept-Laux
(Isère), Gibier Faune Sauvage, Game WIldlife,
vol. 12, p.133-146
- Toigo C., Gaillard J.-M., Michallet J., 1996, La taille des
groupes : un bioindicateur de l'effectif
des populations de bouquetin des Alpes (Capra ibex ibex) ?,
Mammalia, t. 60 (3), p. 463-472
- Raye G., 1994, L'utilisation de l'espace en hiver par le
bouquetin des Alpes (Capra ibex ibex)
dans le parc de la Vanoise, Travaux scientifique du
Parc de la Vanoise, XVIII, p.205-238
- Wilson R., Grigson C., Payne S., 1982, Ageing and
sexing animal bones from archaeological
sites, Oxford,
édition Archeopress, 268 p. (British archaeological Reports
International)


