|
BURKINA FASO
UNITE-PROGRES-JUSTICE
MINISTERE DES
ENSEIGNEMENTS SECONDAIRE,
SUPERIEUR ET DE LA RECHERCHE
SCIENTIFIQUE
UNIVERSITE POLYTECHNIQUE DE BOBO-DIOULASSO
INSTITUT DU DEVELOPPEMENT RURAL

MEMOIRE
en vue de l'obtention
du
DIPLOME DE MASTER RECHERCHE EN ANALYSE DES POPULATIONS
DES
ESPACES FAUNIQUES ET HALIEUTIQUES
SPECIALITE :
Analyse
des populations des espaces halieutiques
|
THEME : ETUDE COMPARATIVE DES MACROINVERTEBRES ET
LEUR
IMPACT SUR LE REGIME ALIMENTAIRE DE Clarias
gariepinus
(BURCHELL, 1822) DE DEUX COURS D'EAU PERMANENTS DE
LA
REGION OUEST DU BURKINA FASO
|
|
Présenté par :
SANOGO Souleymane
Devant le jury composé de :
Pr. Guenda WendengoudiPr. Kabré T. André
Dr Hien Mipro
N° : -2010/MFH2 (Halieutique, Faune) DECEMBRE
2010

DEDICACE
Je dédie ce mémoire
A ma famille
qui ne cesse de croire en moi,
A mes amis
qui n'hésitent aucunement à me
faire
confiance,

TABLE DES MATIERES
LISTE DES TABLEAUX v
LISTE DES FIGURES vii
SIGLES ET ABREVIATIONS viii
REMERCIEMENTS ix
Résumé: x
Abstract: xi
INTRODUCTION 1
I. METHODOLOGIE 4
1.1 Présentation du milieu 4
1.2 Stations d'étude et périodes de
récolte 5
1.3 Matériel 7
1.3.1 Matériel de laboratoire 7
1.3.2 Matériel de terrain 7
1.4 PROTOCOLE 9
1.4.1 Méthode d'échantillonnage
9
1.4.2 Récolte des macroinvertébrés
10
1.4.3 Capture des poissons et récoltes des
contenus stomacaux 10
1.4.4 Mesure de caractéristiques physico-chimiques
au niveau des stations 11
1.4.5 Identification des macroinvertébrés
et analyses des données 11
1.4.5.1 Identification des macroinvertébrés
11
1.4.5.2 Analyse des données 11
II RESULTATS 13
2.1 Caractéristiques physico-chimiques des deux
cours d'eau le Kou et le Wé 13
2.2 Etude comparative des macroinvertébrés
sur le Kou et le Wé 14
2.2.1 Les populations de macroinvertébrés
inventoriées au niveau des deux cours
d'eau 14
2.2.2 Analyse des résultats des études AFC
18
2.2.2.1 Etude sur la répartition
saisonnière des macroinvertébrés immatures au niveau des
deux cours d'eau 18 2.2.2.2 Etude sur la répartition saisonnière
des macroinvertébrés adultes au
niveau des deux cours d'eau 22
2.2.2.3 Etude de l'habitat des
macroinvertébrés immatures 26
2.2.2.4 Etude de l'habitat des
macroinvertébrés adultes 30
2.3 Densité du benthos sur les 2 cours d'eau
33
2.4 Etude de l'alimentation de C. gariepinus au niveau du
Kou 33
2.4.1 Caractéristiques des poissons 33
2.4.2 Habitude alimentaire des poissons 34
2.4.3 Impact de la densité des
macroinvertébrés benthiques sur C. gariepinus
36
III- DISCUSSION 37
3.1 Les populations de macroinvertébrés
37
3.1.1 Macroinvertébrés et qualité de
l'eau 37
3.1.2 Répartition saisonnière des
macroinvertébrés 38
3.1.2.1 Répartition saisonnière des
immatures 38
3.1.2.2 Répartition saisonnière des adultes
38
3.1.3 Habitats des macroinvertébrés
39
3.1.3.1 Habitats des immatures 39
3.1.3.2 Habitats des adultes 39
3.2 Régime alimentaire de C. gariepinus
39
CONCLUSION 41
BIBLIOGRAPHIE 42
WEBOGRAPHIE 45
ANNEXES 46

LISTE DES TABLEAUX
Tableau 1: Liste du matériel utilisé au
laboratoire 7
Tableau 2: Liste exhaustive du matériel
utilisé sur le terrain 8
Tableau 3: Caractéristiques physico-chimiques des
rivières Kou et Wé 13
Tableau 4: Captures d'Arthropodes (Insectes, Arachnides
et Crustacés) au niveau du Kou 15
Tableau 5: Captures totales des Annélides au
niveau du Kou 16
Tableau 6: Captures totales de Mollusques au niveau du
Kou 16
Tableau 7: Captures totales de Mollusques au niveau du
Wé 16
Tableau 8: Captures des Annélides au niveau du
Wé 16
Tableau 9: Captures totales d'Arthropodes (Insectes,
Arachnides et Crustacés) au niveau du Wé 17
Tableau 10: Résultat AFC réalisé sur
la variation saisonnière des immatures de macroinvertébrés
(Valeurs propres et pourcentage de variance) 18
Tableau 11: Résultat AFC réalisé sur
la variation saisonnière des immatures de macroinvertébrés
(Résultats relatifs aux colonnes: contributions et qualité de
représentation) 19
Tableau 12: Résultat AFC réalisé sur
la variation saisonnière des immatures de macroinvertébrés
(Résultats relatifs aux lignes: contributions et qualité de
représentation) 19
Tableau 13: Résultat AFC réalisé sur
la variation saisonnière des adultes de macroinvertébrés
(Valeurs propres et pourcentage de variance) 22
Tableau 14: Résultat AFC réalisé sur
la variation saisonnière des adultes de macroinvertébrés
(Résultats relatifs aux colonnes: contributions et qualité de
représentation) 22
Tableau 15: Résultat AFC réalisé sur
la variation saisonnière des adultes de macoinvertébrés
(Résultats relatifs aux lignes: contributions et qualité de
représentation) 23
Tableau 16: Résultat AFC réalisé sur
les habitats des immatures de macroinvertébrés (Valeurs propres
et pourcentage de variance) 26
Tableau 17: Résultat AFC réalisé sur
les habitats des immatures de macroinvertébrés (Résultats
relatifs aux colonnes: contributions et qualité de
représentation) 26
Tableau 18: Résultat AFC réalisé sur
les habitats des immatures de macroinvertébrés (Résultats
relatifs aux lignes: contributions et qualité de représentation)
27
Tableau 19: Résultat AFC réalisé sur
les habitats des adultes de macroinvertébrés (Valeurs propres et
pourcentage de variance) 30
Tableau 20: Résultat AFC réalisé sur
les habitats des adultes de macroinvertébrés (Résultats
relatifs aux colonnes: contributions et qualité de
représentation) 30
Tableau 21: Résultat AFC réalisé sur
les habitats des adultes de macroinvertébrés (Résultats
relatifs aux lignes: contributions et qualité de représentation)
31
Tableau 22: Variation saisonnière du facteur de
condition de Fulton de C. gariepinus au niveau du Kou 34
Tableau 23: Indice d'occurrence des différentes
proies 35
LISTE DES FIGURES
Figure1 :Bassin du Kou indiquant les deux cours et les
stations d'investigation (Source: VREO-2009) 6
Figure 2: Délimitation de la zone de
récolte sur un cours d'eau 9
Figure 3: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés
immatures au niveau du Kou et du Wé en période d'étiage,
en début de saison pluvieuse et pendant les hautes eaux (axes 1 et 2)
20
Figure 4: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés
immatures au niveau du Kou et du Wé en période d'étiage,
en début de saison pluvieuse et pendant les hautes eaux (axes 1 et 3)
20
Figure 5: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés
immatures au niveau du Kou et du Wé en période d'étiage,
en début de saison pluvieuse et pendant les hautes eaux (axes 2 et 3)
21
Figure 6: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés adultes
au niveau du Kou et du Wé en période d'étiage, en
début de saison pluvieuse et pendant les hautes eaux (axes 1 et 2)
24
Figure 7: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés adultes
au niveau du Kou et du Wé en période d'étiage, en
début de saison pluvieuse et pendant les hautes eaux (axes 1 et 3)
24
Figure 8: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés adultes
au niveau du Kou et du Wé en période d'étiage, en
début de saison pluvieuse et pendant les hautes eaux (axes 2 et 3)
25
Figure 9: Résultat AFC; distribution spatiale des
populations de macroinvertébrés immatures au niveau des habitats
(axes 1 et 2) 28
Figure 10: Résultat AFC; distribution spatiale des
populations de macroinvertébrés immatures au niveau des habitats
(axes 1 et 3) 28
Figure 11: Résultat AFC; distribution spatiale des
populations de macroinvertébrés immatures au niveau des habitats
(axes 2 et 3) 29
Figure 12: Résultat AFC; distribution spatiale des
populations de macroinvertébrés adultes au niveau des habitats
(axes 1 et 2) 32
Figure 13: Résultat AFC; distribution spatiale des
populations de macroinvertébrés adultes au niveau des habitats
(axes 1 et 3) 32
Figure 14: Evolution de la densité du benthos au
niveau du Kou et du Wé 33
Figure 15: Variation de la densité des
macroinvertébrés benthiques et du facteur de condition de Fulton
de C. gariepinus au niveau du Kou 36
SIGLES ET ABREVIATIONS
AEED : Association Eau Environnement et
Développement
AFC : Analyse Factorielle des correspondances
CMSB : Centre Météo Secondaire de Bobo
Dioulasso
LaRFPF : Laboratoire de Recherche et de
Formation en Pêche et Faune
NEPAD : New Partenership for Africa's
Development (Nouveau Partenariat pour le Développement de l'Afrique)
ONEA : Office National de l'eau et de
l'Assainissement VREO : Valorisation des Ressources en Eau de
l'Ouest WFD: Water Framework Directive
REMERCIEMENTS
Ce travail qui est le fruit d'un engagement personnel à
la recherche du savoir, n'aurait pu être mené à bien sans
le concours et la franche collaboration de certaines personnes. C'est le lieu
ici pour moi de leur signifier ma très grande reconnaissance.
J'adresse mes sincères remerciements au Pr.
Kabré T. André, mon maître de stage et directeur du LaRFPF,
pour tous ses conseils, sa disponibilité et sa marque de sympathie
à mon égard. Sa très grande confiance en moi m'a
donné une direction dans la recherche scientifique. Je lui suis
reconnaissant pour m'avoir fait connaître la vie des
macroinvertébrés que nous menaçons tous les jours par nos
actes.
Au Pr. Guenda Wendengoudi, président du jury et au Dr.
Hien Mipro, j'adresse mes sincères remerciements pour n'avoir
ménagé aucun effort pour que ma soutenance se tienne dans de
bonnes conditions. Merci pour vos critiques franches et sincères pour
l'amélioration de la qualité scientifique de ce document.
Mes remerciements vont à l'endroit de Messieurs,
Sawadogo Issouf technicien supérieur de l'environnement à la
direction régionale de l'environnement et du cadre de vie à
BoboDioulasso, Sanou Adama responsable du laboratoire d'analyse de la
qualité de l'eau à l'ONEA Bobo, Oumarou Issouf et Zongo
Mahamoudou, tous deux techniciens de météorologie au CMSB,
Valentin Iogo ingénieur de travaux en informatique au programme VREO,
Midekor Akoly Agbevi chargé de mission de l'observatoire de l'eau
à l'AEED., merci d'avoir accordé à ma modeste personne un
peu de votre temps. Chacun de vous verra son empreinte dans ce document.
Je remercie mes amis Rouamba W. Eric Parfait et
Ouédraogo Abdoulaye pour la relecture et la mise en page de mon
document.
A tous les enseignants qui ont accordé une importance
majeure à cette formation, que Dieu vous récompense de la
meilleure des manières.
Aux autres étudiants de cette première promotion
Master Recherche en Analyse des Populations des Espaces Fauniques et
Halieutiques, je souhaite bon courage et que Dieu nous donne la force de
continuer.
Résumé:
La rivière Kou et le marigot Wé sont deux cours
d'eau de la région ouest du Burkina Faso. Leur présence contribue
à l'amélioration de l'économie locale.
A travers une étude comparative sur ces deux cours
d'eau, l'inventaire des populations de macroinvertébrés a
été réalisé pendant la période
d'étiage, en début de saison pluvieuse et pendant les hautes
eaux. Sur le Kou, 33 familles d'Insectes, 1 famille d'Arachnides, 1 famille de
Crustacés, 2 familles d'Annélides et 5 familles de Mollusques ont
été identifiées tandis que sur le Wé, 22 familles
d'Insectes, 1 famille d'Annélides, 2 familles de Mollusques ont
été identifiées. On observe sur ces deux cours d'eau, une
faible quantité de EPT
(Ephèméroptères-Plécoptères-Trichoptéres),
avec 0,016% sur le Kou et 0.008% sur le Wé. Les espèces
appartenant à ces ordres d'Insectes, très sensibles à la
pollution témoignent de par leur abondance dans un cours d'eau, une
bonne santé écologique (Moisan et Pelletier, 2008). A l'instar
des autres cours d'eau du pays, ces deux cours d'eau se caractérisent
par la forte présence d'Insectes (90,70% sur le Kou et 92,94% sur le
Wé) et leur benthos est dominé par les Chironomidae
(23,69% au niveau du Kou et 50,37% au niveau du Wé). La
densité des macroinvertébrés benthiques sur ces deux cours
ne dépasse pas les 500 individus au m2 pendant les périodes
d'étude.
Cette étude a aussi permis d'analyser le régime
alimentaire de Clarias gariepinus (Burchell, 1822) au niveau du Kou.
Cette espèce de poisson, malgré son régime omnivore
consomme majoritairement les Insectes (indice d'occurrence est de 76,47%) et
son poids augmente pendant les périodes de forte densité
benthiques (période d'étiage et pendant les hautes eaux). Pour
une stabilité des écosystèmes liés à ces
cours d'eau, cette étude préconise leur suivi permanent afin de
déceler toute perturbation écologique qui affectera toute la
région. Ce suivi devrait prendre en compte la sensibilisation des
populations riveraines qui semblent ignorer les méfaits des effluents
domestiques.
Mots clés : macroinvertébrés -
Chironomidae - Clarias gariepinus - effluents domestiques.
Abstract:
Rivers «Kou» and «Wé» are two rivers
in the west region of Burkina Faso. They contribute to the improvement of local
economy.
In a comparative study of these two rivers, the inventory of
macro-invertebrates population was realized during low water period, the
beginning of rainy season and high water period.
On the «Kou» river 33 families of Insects, one
family of Arachnid, one family of shellfish, two families of Annelid and five
families of Mollusc have been identified, while on the «Wé»
river 22 families of Insects, one family of Annelid, two family of Mollusc have
been described. We notice on these two rivers a small quantity of EPT
(Ephemeroptera-PlecopteraTrichoptera) with 0.016% on the «Kou» river
and 0.008% on the «Wé» river. The species belonging to these
orders of Insects, very sensitive to pollution show by their abundance in a
waterway a good ecological well being (Moisan & Pelletier, 2008).
Like observed in most waters bodies of the country, the two
rivers are characterized by high presence of Insects (90.70% on the river
«Kou» and 92.94% on the river «Wé»), and their
benthic populations are dominated by Chironomidae (23.69% on the river
«Kou» and 50.37% on the river «Wé»). The density of
macro-invertebrates in the benthos of both rivers does not go beyond 500
individuals per square meter (m2) during the period of study.
This study has allowed us to analyse the diet of Clarias
gariepinus (Burchell, 1822), in the river «Kou». This species of
fish, despite its omnivorous diet, eats in majority Insects (occurrence index
is 76.47%) and its weight increases during high benthic density period
(low water period and high water period).
For the stability of the ecosystem of the two waterways, this
study recommends their permanent survey in order to depict any ecological
disturbance that will affect the whole region. This survey must include the
education of the riverside population on the impact of domestic effluents on
rivers waters quality.
Key words: macro-invertebrates - Chironomidae -
Clarias gariepinus - domestic effluents
xii
INTRODUCTION
Les questions environnementales constituent aujourd'hui des
défis majeurs pour la survie de l'humanité. Les récents
bouleversements climatiques à travers le monde avec leurs corollaires de
destructions des biens matériels et même de perte en vie humaine
ont contribué à renforcer la recherche sur la conservation de
l'équilibre des écosystèmes.
Les écosystèmes aquatiques,
précisément ceux continentaux, font l'objet d'un
intérêt particulier à travers de nombreuses conventions
inter-états.
La convention de Ramsar (Iran) du 2 février 1971 relative
aux zones humides mettait déjà en exergue la sensibilité
de ces zones qui par conséquent devraient bénéficier d'un
suivi.
Moisan et Pelletier (2008) proposent à cet effet un
guide de suivi des cours d'eau du Québec, basé sur les
communautés biologiques. Selon eux, l'intégrité
écologique d'un cours d'eau est une combinaison des
intégrités chimiques, physiques et biologiques et
généralement, toute dégradation d'une ou de plusieurs de
ces composantes se reflète dans la communauté biologique. Citant
Hellawel (1986), Barbour et al. (1999) et WFD (2003), et en abordant
dans le même sens qu'eux, Moisan et Pelletier (2008) relatent les
différentes étapes du suivi des cours d'eau en accordant une
place de choix aux macroinvertébrés benthiques. La
diversité de ces êtres dans un cours d'eau témoigne d'une
bonne santé écologique.
En Afrique, les connaissances sur ces êtres sont de plus
en plus diversifiées grâce aux travaux remarquables de chercheurs.
La clé d'identification de Levêque et Durand (1981) et
l'iconographie des insectes aquatiques de Dejoux et al. (1983) en sont
des exemples. Ces auteurs ont inspiré Diomandé et al.
(2009) qui, après une évaluation de la macrofaune benthique de la
rivière Agnéby, cours d'eau au sud de la Côte d'Ivoire, ont
identifié les paramètres abiotiques structurant ces
peuplements.
Une des importances de ces macroinvertébrés est
leur présence dans « le bol alimentaire » de nombreux
poissons. Cette ressource halieutique contribue pour une bonne partie à
la sécurité alimentaire et aux revenus des ménages, ainsi
qu'à la réduction de la pauvreté en Afrique et participe
ainsi à son développement économique (NEPAD, 2005).
Le Burkina Faso à l'instar des autres pays
sahéliens est confronté au problème de l'eau. Très
peu arrosé, ses cours d'eau connaissent pour la plupart des
périodes de tarissement dont la durée dépend des
précipitations. De plus, l'exploitation de cette ressource naturelle par
les populations riveraines perturbe très souvent l'équilibre des
écosystèmes qui y sont liés.
De nombreuses études sur les
macroinvertébrés en relation avec la qualité biologique de
l'eau ont été réalisées, étoffant ainsi la
littérature au plan national. Ces études, pour la plupart,
étaient menées dans des eaux lacustres. On peut citer entre
autres les travaux de Dejoux (1977) sur les Chironomes du lac Bam dans la
région du centre- nord, Kabré et al. (2000) dont les
travaux sur les barrages de Bagré au centre-est ont abouti à un
inventaire et une quantification de la densité benthique de ces
barrages, Kabré et al. (2002) qui ont analysé
l'évolution saisonnière des macroinvertébrés du lac
du barrage de la Comoé au sud-ouest et Ouéda et al.
(2009) qui ont fait une étude sur l'abondance, la diversité
et la dynamique saisonnière des zooplanctons dans le barrage de Loumbila
au centre du pays.
Dans les milieux lotiques, on peut citer les travaux de Guenda
(1996), sur le réseau hydrographiques du Mouhoun (ex Volta noire, figure
1). Cette étude a permis une approche de la distribution des Insectes et
fourni une clé d'identification de nymphes de Simuliidae.
La rivière Kou et le marigot Wé, situés
à l'ouest du pays sont deux cours d'eau permanents dont la
présence contribue à une croissance de l'économie locale.
En effet, l'approvisionnement en eau potable, en poissons et produits vivriers
de Bobo-Dioulasso, seconde ville du pays, vient du Kou ; l'eau du marigot
Wé qui sert à l'agriculture et aux activités domestiques
des populations se particularise par la présence de silures
sacrés.
Les travaux de recherche relatifs aux
macroinvertébrés benthiques en relation avec la qualité de
l'eau sur ces deux cours d'eau n'existent pas. L'objectif de la
présente, tout en contribuant à combler ce déficit
d'information, est de faire l'inventaire comparatif des populations de
macroinvertébrés sur ces deux cours d'eau. Elle a permis
d'établir l'impact de ces macroinvertébrés sur le
régime alimentaire de Clarias gariepinus (Burchell, 1822),
poisson dont la résistance aux adversités du milieu a
été relatée par Corbet (1961).
Les objectifs spécifiques qui s'en dégagent sont
les suivants :
- inventorier les populations de macroinvertébrés
sur ces deux cours d'eau par saison ; - déterminer les espèces
(ou traits biologiques) indicatrices de pollution ;
- analyser l'impact des macroinvertébrés dans la
nutrition de C. gariepinus.
Il est sans doute nécessaire de poser
l'hypothèse suivante : Les macroinvertébrés benthiques
sont des traits biologiques permettant de caractériser la qualité
physicochimique et la diversité biologique de ces deux cours d'eau.
Ce document est structuré en trois parties. Dans la
première partie, une présentation du matériel et de la
méthodologie sera faite. La deuxième partie traitera des
différents résultats obtenus et la dernière partie sera
basée sur la discussion.
I. METHODOLOGIE
1.1 Présentation du milieu
La zone d'étude, située à l'ouest du
pays, est l'une des régions les plus arrosées. On y trouve des
cours d'eau permanents, des lacs, des mares et des sources d'eau souterraine.
Le climat de type tropical soudano-sahélien est
caractérisé par l'alternance de deux saisons distinctes: une
saison des pluies de mi-mai à octobre et une saison sèche le
reste de l'année.
Selon les informations reçues du Centre
Météorologique Secondaire de Bobo-Dioulasso (CMSB) la
pluviométrie annuelle varie de 900 à 1500 mm sur une moyenne de
85 jours de pluies; les températures minimales varient entre 18° C
et 26° C, et les maximales de 30° C à 38° C. Les mois
chauds sont les mois de février, mars, avril et mai ;
l'évapotranspiration réelle est estimée à 860 mm.
Les cours d'eaux permanents de la région bénéficient de la
contribution des sources aquifères.
L'infiltration est seulement de 10% des pluies, car la zone est
dominée par une texture peu perméable (Wellens et al.,
2009).
La rivière Kou située entre le 11°05' et
11°25' de latitude nord et de 4°20' et 4°30' de longitude ouest,
part de Péni au sud de Bobo-Dioulasso et se jette dans le Fleuve Mouhoun
(figure 1). La rivière connaît à certains endroits la
destruction de berges et l'ensablement. Le marigot Wé, prend sa source
à 8 km au sud de Bobo-Dioulasso et rejoint le Kou au nord de la ville
après l'avoir traversée. La plupart des plantes aquatiques sur
ces deux cours d'eau sont de la famille des Poacae
(représentée par Eulodina indica, Panicum sp,
Panicum
anabatistum), des Convolvulacae (avec
Ipomoea eriocarpa, Ipomoea pescapre) et des
Onagracae (avec Ludwizia sp).
1.2 Stations d'étude et périodes de
récolte
Trois stations ont été retenues sur le Kou et une
sur le Wé (figure 1). Le respect des croyances
locales ne permettait pas de nombreuses duplications de
l'échantillonnage sur le marigot Wé. La capture des
macroinvertébrés s'est effectuée en trois périodes
à savoir la période d'étiage, le début de la saison
des pluies et la période des hautes eaux. Les captures ont donc
débuté en avril et elles ont pris fin en juillet avec une sortie
par mois, soit 4 sorties au total. La sortie d'avril étant la
période d'étiage, mai-juin correspond à la période
de début de saison pluvieuse et juillet à la période des
hautes eaux.

Station d'étude
Figure1 : Bassin du Kou indiquant les
deux cours et les stations d'investigation (Source: VREO-2009)
1.3 Matériel
1.3.1 Matériel de laboratoire
Ce matériel se compose d'instruments et documents
utilisés au laboratoire pour
l'identification, la conservation des
macroinvertébrés et d'instruments servant à l'analyse des
poissons et de leurs contenus stomacaux (tableau 1).
1.3.2 Matériel de terrain
Ce matériel est utilisé sur le terrain pour la
reconnaissance et la capture des macroinvertébrés
ainsi que la capture des poissons. Le tableau 2 donne une
liste de tout le matériel utilisé lors des différents
échantillonnages et même du matériel servant de protection
sur le terrain comme les gants et les bottes car certains endroits sont
très pollués.
Tableau 1: Liste du matériel utilisé au
laboratoire
|
Matériel
|
Rôle
|
Nombre
|
|
Microscope
|
Observation des macroinvertébrés
|
1
|
|
Alcool 900
|
Fixation et conservation des macroinvertébrés
|
1 litre
|
|
Boite de Pétri
|
Fixation des macroinvertébrés pour l'observation
|
2
|
|
Etiquettes
|
Identification des échantillons
|
1
|
|
Trousse à dissection
|
Dissection des poissons
|
1
|
|
Ichtyomètre
|
Mesure de la taille des poissons
|
1
|
|
Balance
|
Mesure du poids des poissons et du bol alimentaire
|
1
|
|
Documents (cf. bibliographie)
|
Identification des macroinvertébrés
|
|
Tableau 2: Liste exhaustive du matériel
utilisé sur le terrain
|
Matériel
|
Rôle
|
Nombre
|
|
Pinces
|
Capture des macroinvertébrés
|
2
|
|
Gants
|
Protection
|
1 paire
|
|
Bottes
|
Protection
|
1 paire
|
|
Crayon à papier
|
Prise de notes
|
1
|
|
Calepin
|
Prise de notes
|
1
|
|
Loupe de poche
|
Repérage des macroinvertébrés
|
1
|
|
Appareil photo
|
Prise de vue des endroits
|
1
|
|
Thermomètre
|
Prise de la température de l'eau
|
1
|
|
Piquets
|
Délimiter la zone de récolte
|
4
|
|
ph-mètre
|
Mesure du pH de l'eau
|
1
|
|
Ficelles
|
Délimitation de la zone de récolte
|
1
|
|
Conductimètre
|
Mesure de la conductivité de l'eau
|
1
|
|
Mètre
|
Mesure de la zone de récolte
|
1
|
|
Etiquettes
|
Identification des échantillons
|
142
|
|
Flacons
|
Récolte des macroinvertébrés
|
142
|
|
bocaux
|
Récolte du fond de l'eau
|
4
|
|
Montre
|
Prise du temps
|
1
|
|
Alcool 900
|
Fixation des macroinvertébrés
|
2 litres
|
|
Sac
|
Transport du matériel
|
1
|
|
Benne géologique
|
Récolte du benthos
|
1
|
|
Epuisette de maille 2 mm
|
Récolte des macroinvertébrés dans l'eau
|
1
|
|
Seau
|
Tri du benthos
|
2
|
|
Tamis de maille 0,2 mm
|
Tamisage du benthos
|
1
|
|
Tamis de maille 0,6 mm
|
//
|
1
|
|
Tamis de maille 2 mm
|
//
|
1
|
|
Tamis de maille 4 mm
|
//
|
1
|
|
Couteau de poche
|
|
1
|
|
Disque de Secchi
|
Mesure de la transparence de l'eau
|
1
|
|
Sachets plastiques
|
Etouffer les poissons
|
8
|
|
Nasse
|
Capture des poissons
|
1
|
1.4 PROTOCOLE
1.4.1 Méthode d'échantillonnage
Pour cette étude, nous avons choisi l'approche
multihabitat comme décrit par Moisan et
Pelletier (2008). Les habitats échantillonnés
sont les racines des plantes, les pierres, les branches mortes flottantes, la
surface de l'eau et la vase (fond de l'eau). En effet selon ces auteurs, ce
protocole est réalisé dans les cours d'eau à substrat fin
et à écoulement lent. Ces caractéristiques se retrouvent
au niveau de ces deux cours d'eau.
La récolte sur le terrain a nécessité la
délimitation d'une surface de travail de 15 m2. La zone a
été balisée par les ficelles et les piquets (figure 2).
Cours d'eau

Zone de récolte
Figure 2: Délimitation de la zone de
récolte sur un cours d'eau
1.4.2 Récolte des macroinvertébrés
- récolte sur la vase
Le prélèvement du benthos s'est effectué
à l'aide de la benne géologique de 300 cm2 de surface
de balayage. Deux coups de benne sont portés sur le fond de l'eau soit
une surface de 600 cm2. La vase obtenue, mélangée avec
une quantité suffisante d'eau est passée à travers les
mailles des différents tamis des plus grandes aux plus petites. Le tri
se fait directement sur les différents tamis. Les
macroinvertébrés sont récoltés et introduits dans
les flacons contenant une quantité suffisante d'alcool. A l'aide de la
loupe, les plus petits sont récupérés sur le tamis
à petites mailles. Pour un échantillonnage plus complet, les
vases retenues sur les deux derniers tamis sont conservées dans des
bocaux avec un peu d'alcool, pour une fouille au laboratoire sous
microscope.
- récolte à la surface
Cette récolte s'est effectuée avec
l'épuisette, qui est promenée sur toute la surface
d'échantillonnage. Ceux qui sont visibles sont traqués
automatiquement.
Pour ne pas perdre les individus capturés,
l'épuisette est mise dans le seau et le contenu est trié hors de
l'eau. L'épuisette a aussi permis de capturer les individus nageant en
profondeur.
- récolte sur les racines des plantes
Les plantes arrachées dans la zone sont rincées et
l'eau de rinçage est passée entre les mailles du tamis de 0,2 mm.
Les individus retenus sont recueillis.
- récolte sur les pierres
Les pierres en parties inondées de la surface sont
recueillies et rincées et l'eau de rinçage est passée sur
le tamis de 0,2 mm de maille. Les individus visibles sont préalablement
recueillis.
- récolte sur les branches mortes
Cette récolte se fait juste après la
délimitation de la zone de récolte. Toutes les branches mortes
sur l'eau sont automatiquement recueillies dans l'épuisette. Les
macroinvertébrés visibles sont récupérés
directement. A l'aide de pinces, les écorces des branches sont
écartées et les éventuels individus vus sont recueillis
dans des flacons.
1.4.3 Capture des poissons et récoltes des contenus
stomacaux
Les C. gariepinus ont été
capturés à l'aide de nasse très tôt le matin. Une
fois capturés, ils sont tués par étouffement pour
arrêter les processus de digestion. Au laboratoire, les poissons sont
pesés et leur longueur totale est mesurée. Les estomacs sont
recueillis avec le contenu et le poids du bol alimentaire des estomacs non
vides est déterminé.
1.4.4 Mesure de caractéristiques physico-chimiques
au niveau des stations Ces mesures concernent la température,
la profondeur, la conductivité, la transparence, le pH
et la vitesse d'écoulement de l'eau. Elles se sont
réalisées sur chaque station pendant les différentes
sorties.
Les mesures de la température et de la profondeur ont
concerné deux endroits différents à chaque station et une
moyenne a été retenue.
Les mesures de la conductivité et du pH ont
été réalisées sur des échantillons d'eau au
laboratoire d'analyse de qualité de l'eau de l'ONEA.
En l'absence de matériel adéquat, la vitesse
d'écoulement de l'eau a été estimée par la vitesse
de déplacement d'un objet sur l'eau (Moisan et Pelletier, 2008). Quatre
mesures ont été réalisées et une moyenne a
été retenue par station. Les valeurs trouvées ont permis
de comparer la vitesse d'écoulement des deux cours d'eau.
1.4.5 Identification des macroinvertébrés et
analyses des données
1.4.5.1 Identification des macroinvertébrés
L'identification s'est limitée à la famille pour tous
les individus.
L'identification des Insectes et leur classification
taxonomique ont été faites au laboratoire sous un microscope
binoculaire avec l'aide de clé d'identification de Merrit et Cummins
(1984) et de Levêque (1981), de Roger et Jacques (1978) et du guide
d'identification de Moisan (2006). Le guide de Moisan (2006) a aussi permis
d'identifier les Mollusques et les Annélides. Le recours à des
sites web a été profitable, car ces sites avaient assez
d'illustrations sur les macroinvertébrés (confère
webographie).
1.4.5.2 Analyse des données
* analyse des macroinvertébrés
capturés par station
Pour comparer ces deux cours d'eaux, pendant les 3
périodes sus citées, nous avons considéré les
moyennes à chaque période sur chaque cours d'eau. Pendant le
traitement des données, les adultes ont été
séparés des immatures (Nymphes et larves)
Deux études AFC ont été
réalisées avec le logiciel R 2.10.0:
- une première qui a permis de comparer les deux cours
d'eau pendant les trois périodes. - une deuxième pour montrer
l'affinité des macroinvertébrés aux différents
substrats.
La densité numérique (nombre d'individus au
m2) du benthos a été calculée au niveau de
chaque rivière durant les trois périodes :
|
Ó des organismes benthiques
échantillonnés
Densité (au mètre carré) = x 10000
cm2
600 cm2
|
Il est à noter que le total des organismes benthiques
utilisé pour ce calcul est la moyenne considérée plus
haut.
* analyse du régime alimentaire des
poissons
Les estomacs des poissons ont été
prélevés chez les individus en bon état. L'identification
des proies s'est réalisée avec les documents cités plus
haut.
Nous avons ainsi calculé :
- l'indice d'occurrence de Hyslop (1980) :
Io = (Nx/Nt)*100
Io : indice d'occurrence
Nx : nombre d'estomacs contenant une catégorie
d'aliments
Nt : nombre total d'estomacs non vides
Cet indice permet de montrer la préférence
alimentaire des poissons dans un lot de proies. Ainsi : % Io<10 proies
accidentelles
10 = % Io = 50 : proies secondaires
50 = % Io : proies préférées (Hureau,
1970)
- le facteur de Fulton (1911) :
f= P/ L3
f: facteur de Fulton; P: poids (g) ; L: longueur (cm)
Cette relation implique que plus le facteur est
élevé, plus le poisson est gros et il se trouve ainsi dans de
meilleures conditions environnementales.
* calcul des constances a et b dans la relation
poids-taille
Il existe une relation entre la longueur L et le poids P des
poissons de la forme P=aLb a et b étant des constances, b est
généralement compris entre 2,5 et 3,5 (Moreau, 1979, cité
par Baijot et al. 1994).
La recherche de ces paramètres a nécessité
une transformation logarithmique. Les limites de confiances de b ont
été calculées comme proposé par Sparre et Vanema
(1996).
II RESULTATS
2.1 Caractéristiques physico-chimiques des deux
cours d'eau le Kou et le Wé
Le tableau 3 indique les différentes
caractéristiques physico-chimiques trouvées au niveau de ces deux
cours d'eau. L'estimation de la vitesse de l'eau permet de constater un
écoulement plus rapide de l'eau au niveau du Kou par rapport au
Wé. La transparence de l'eau est restée bonne sur ces deux cours
d'eau pendant les périodes d'échantillonnage.
Tableau 3: Caractéristiques physico-chimiques des
rivières Kou et Wé
|
Périodes
|
Caractéristiques
|
Kou
|
Wé
|
|
pH
|
7.25
|
6.40
|
|
Température (°C)
|
28
|
27
|
|
étiage
|
Profondeur (cm)*
|
26
|
23
|
|
Conductivité (us/cm)
|
240
|
310
|
|
pH
|
7.40
|
7.00
|
|
Température (°C)
|
27
|
28
|
|
Début des pluies
|
Profondeur (cm)*
|
30
|
27
|
|
Conductivité (us/cm)
|
124
|
286
|
|
pH
|
7.60
|
7.10
|
|
Température (°C)
|
27
|
28
|
|
Hautes eaux
|
Profondeur (cm)*
|
45
|
33
|
|
Conductivité (us/cm)
|
77
|
243
|
* profondeur de visibilité du disque de Secchi
2.2 Etude comparative des
macroinvertébrés sur le Kou et le Wé
2.2.1 Les populations de macroinvertébrés
inventoriées au niveau des deux cours d'eau
Les tableaux 4, 5, 6, 7, 8 et 9 indiquent les
différentes familles de macroinvertébrés capturées
au niveau des différents habitats pendant les trois périodes
d'étude. Au total, 803 individus dont 746 Arthropodes (Insectes,
Arachnides et Crustacés), 41 Annélides et 16 Mollusques ont
été identifiés sur les deux cours d'eau. Ces individus se
repartissent en 36 familles d'Insectes dont les importants sont les
Carabidae, les Dytiscidae, les Elmidae, les
Belostomatidae, les Calopterygidae et les
Chironomidae, une famille d'arachnides (les Acaridae), une
famille de Crustacés, les Potamidae (tableau 4 et 9), 2
familles de d'Annélides qui sont les Hirudinae et les
Oligocheta (tableau 5 et 8) et en 5 familles de Mollusques à
savoir les Ancylidae, les Physidae, les Lymnaeidae,
les Planorbidae et les Valvatidae (tableau 6 et 7). Ces
différentes familles se retrouvent au niveau du Kou à l'exception
de 3 familles d'Insectes que sont les Ptilodactylidae, les
Muscidae et les Tanyderidae (tableau 4, 5 et 6). Au niveau du
Wé, 22 familles d'Insectes sont représentées dont la plus
importante est la famille des Chironomidae, une famille
d'Annélides, la famille des Oligocheta et 2 familles de
Mollusques à savoir les Physidae et les Lymnaeidae
(tableau 7, 8 et 9).
Parmi les Arthropodes, les Insectes sont les plus nombreux
(90,70% sur le Kou et 92,94% sur le Wé). Les Mollusques
représentent 3,82% et les Annélides 4,78% au niveau du Kou. Au
niveau du Wé, on a 1,9% de Mollusques et 7,32% d'Annélides.
Le benthos de ces deux cours d'eau est dominé par les
Chironomidae avec 23,69% sur le Kou et de 50,37% sur le Wé.
Tableau 4: Captures d'Arthropodes (Insectes, Arachnides
et Crustacés) au niveau du Kou
|
Familles
|
code
|
Etiage (avril)
|
Début de saison
(mai-
juin)
|
Hautes eaux (juillet)
|
|
EA PI VA TR RA
|
EA PI VA TR RA
|
EA PI VA TR RA
|
|
Coléoptères
|
|
Carabidae
|
Cb
|
|
|
|
1
|
2
|
|
4
|
|
|
5
|
|
3
|
|
|
1
|
|
Chrysomelidae
|
Cy
|
|
|
|
|
|
2
|
|
|
1
|
4
|
|
|
|
|
|
|
Cucurlionidae
|
Cu
|
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
|
Dytiscidae
|
Dy
|
7
|
|
3
|
1
|
1
|
1+1
|
1+1
|
1
|
1+3
|
6
|
1
|
4+1
|
|
2
|
|
|
Elmidae
|
El
|
1
|
|
2
|
1
|
|
|
4+2
|
|
4
|
2+1
|
|
2
|
1
|
2
|
|
Gyrinidae
|
Gy
|
|
|
|
|
|
2
|
|
|
|
|
2
|
|
|
|
|
|
Histeridae
|
Hi
|
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
|
Hydraenidae
|
Hd
|
1
|
1
|
|
3
|
|
|
|
|
3
|
4
|
|
1
|
|
|
|
|
Hydrophilidae
|
Hy
|
|
|
3
|
5
|
|
|
1
|
|
3
|
|
|
|
|
1
|
|
|
Ptilodactylidae
|
Pt
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Spercheidae
|
Sp
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Staphilinidae
|
St
|
|
1
|
|
|
|
|
1
|
|
1
|
|
|
|
|
|
|
|
Hémiptères
|
|
Belostomatidae
|
Be
|
108
|
|
|
|
1
|
8
|
|
|
1
|
3
|
|
|
|
|
1+4
|
|
Cydnidae
|
Cn
|
|
|
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
Gerridae
|
Gr
|
1
|
|
|
|
|
11
|
|
|
|
|
7
|
|
|
|
|
|
Hebidae
|
He
|
|
|
|
|
|
1
|
|
|
|
|
|
|
|
|
|
|
Naucoridae
|
Na
|
2
|
|
|
|
1
|
1
|
2
|
|
2
|
1
|
|
|
|
|
|
|
Nepidae
|
Np
|
|
|
|
|
2
|
|
1
|
|
|
|
|
|
|
|
|
|
Notonectidae
|
No
|
|
|
|
|
|
15
|
|
|
|
|
|
|
|
|
|
|
Odonates
|
|
Calopterygidae
|
Cp
|
|
|
3
|
|
1
|
1
|
2
|
|
2
|
11
|
|
1
|
|
|
|
|
Gomphidae
|
Gd
|
|
|
1
|
|
1
|
|
|
|
|
1
|
|
|
|
|
|
|
Libellulidae
|
Ll
|
|
|
2
|
|
|
|
|
|
1
|
2
|
|
|
1
|
|
|
|
Diptères
|
|
Ceratopogonidae
|
Ce
|
|
|
|
|
|
|
1
|
1
|
|
|
|
1
|
|
|
|
|
Chironomidae
|
Ch
|
|
|
7
|
|
|
|
7
|
35
|
12
|
1
|
|
|
81
|
4
|
|
|
Culicidae
|
Cl
|
|
|
|
|
|
|
|
|
|
|
2
|
|
|
|
|
|
Dolichopogonidae
|
Dp
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
|
|
|
Muscidae
|
Mu
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tabanidae
|
Ta
|
|
|
|
|
|
|
|
2
|
|
|
|
|
|
|
|
|
Tanyderidae
|
Td
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Simuliidae
|
Sm
|
|
|
|
|
|
|
8
|
|
|
|
|
|
|
|
|
|
Stratiomyidae
|
Sd
|
|
|
|
|
|
1
|
|
|
1
|
|
|
|
|
|
|
|
Syrphidae
|
Sy
|
|
|
3
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ephéméroptères
|
|
Ephemerellidae Ep 1 2 2
|
|
Trichoptères
|
|
Hydropsychidae Hp 4
|
|
Lépidoptères
|
|
Cossidae Cs 1
|
|
Orthoptères
|
|
Gryllotalpidae Gl 1
|
|
Arachnides
|
|
Acaridae Ac 1 1
|
|
Crustacés
|
|
Potamidae Pm 3
|
Tableau 5: Captures totales des Annélides au
niveau du Kou
|
Familles
|
Code
|
Etiage (avril)
|
Début de saison (mai-
juin)
|
Hautes eaux (juillet)
|
|
|
EA
|
PI
|
VA
|
TR
|
RA
|
EA
|
PI
|
VA
|
TR
|
RA
|
EA
|
PI
|
VA
|
TR
|
RA
|
|
Hirudinae
|
Hr
|
6
|
1
|
|
|
|
|
1
|
|
|
|
|
|
|
2
|
|
|
Oligocheta
|
Og
|
1
|
1
|
6
|
|
1
|
|
8
|
|
|
|
|
2
|
|
|
|
Tableau 6: Captures totales de Mollusques au niveau du
Kou
|
Familles
|
Code
|
Etiage (avril)
|
Début EA
|
de PI
|
saison juin) VA
|
TR
|
(mai-
RA
|
EA
|
Hautes
PI
|
eaux VA
|
(juillet) TR
|
RA
|
|
EA
|
PI
|
VA
|
TR
|
RA
|
|
Ancylidae
|
An
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
|
|
Physidae
|
Py
|
|
|
2
|
|
|
|
3
|
|
1
|
|
|
|
|
|
|
|
Planorbidae
|
Pb
|
|
1
|
2
|
|
|
|
1
|
|
|
|
|
|
|
|
|
|
Lymnaeidae
|
Ly
|
|
|
|
|
|
|
1
|
|
1
|
|
|
|
|
|
|
|
Valvatidae
|
Vv
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
|
Tableau 7: Captures totales de Mollusques au niveau du
Wé
|
Familles
|
Code
|
Etiage (avril)
|
Début EA
|
de PI
|
saison juin) VA
|
TR
|
(mai-
RA
|
EA
|
Hautes
PI
|
eaux VA
|
(juillet) TR
|
RA
|
|
EA
|
PI
|
VA
|
TR
|
RA
|
|
Ancylidae
|
An
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Physidae
|
Py
|
|
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Planorbidae
|
Pb
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Lymnaeidae
|
Ly
|
|
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Valvatidae
|
Vv
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tableau 8: Captures des Annélides au niveau du
Wé
|
Familles
|
Code
|
Etiage (avril)
|
Début EA
|
de PI
|
saison juin) VA
|
TR
|
(mai-
RA
|
Hautes EA
|
PI
|
eaux VA
|
(juillet) TR
|
RA
|
|
EA
|
PI
|
VA
|
TR
|
RA
|
|
Hirudinae
|
Hr
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Oligocheta
|
Og
|
|
|
|
|
|
|
3
|
|
1
|
3
|
|
2
|
|
2
|
1
|
Tableau 9: Captures totales d'Arthropodes (Insectes,
Arachnides et Crustacés) au niveau du Wé
|
Familles
|
code
|
Etiage (avril)
|
Debut de saison (mai-
juin)
|
Hautes eaux (juillet)
|
|
EA PI VA TR RA
|
EA PI VA TR RA
|
EA PI VA TR RA
|
|
Coléoptères
|
|
Carabidae
|
Cb
|
|
|
|
|
|
|
1
|
|
3
|
2
|
|
|
|
|
|
|
Chrysomelidae
|
Cy
|
|
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
Cucurlionidae
|
Cu
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Dytiscidae
|
Dy
|
1
|
|
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
Elmidae
|
El
|
|
|
|
|
|
|
|
|
1
|
2
|
|
|
|
|
1
|
|
Gyrinidae
|
Gy
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Histeridae
|
Hi
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Hydraenidae
|
Hd
|
1
|
|
|
1
|
|
|
|
|
|
1
|
|
|
|
|
|
|
Hydrophilidae
|
Hy
|
|
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
Ptilodactylidae
|
Pt
|
|
|
|
|
|
|
|
|
2
|
|
|
|
|
|
|
|
Spercheidae
|
Sp
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Staphilinidae
|
St
|
|
|
|
|
|
|
|
|
1
|
2
|
|
|
|
|
|
|
Hémiptères
|
|
Belostomatidae
|
Be
|
3
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cydnidae
|
Cn
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Gerridae
|
Gr
|
|
|
|
|
|
4
|
|
|
|
|
1
|
|
|
|
|
|
Hebidae
|
He
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Naucoridae
|
Na
|
2
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Nepidae
|
Np
|
|
|
|
|
3
|
|
|
|
|
|
|
|
|
|
|
|
Notonectidae
|
No
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Odonates
|
|
Calopterygidae
|
Cp
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Gomphidae
|
Gd
|
|
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Libellulidae
|
Ll
|
|
|
|
|
|
|
|
1
|
|
2
|
|
|
|
|
|
|
Diptères
|
|
Ceratopogonidae
|
Ce
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Chironomidae
|
Ch
|
|
12
|
|
|
|
7
|
78
|
37
|
32
|
4
|
4
|
4
|
1
|
2
|
1
|
|
Culicidae
|
Cl
|
|
|
|
|
|
|
|
|
|
|
7
|
|
|
|
|
|
Dolichopogonidae
|
Dp
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Muscidae
|
Mu
|
|
|
|
|
|
|
|
|
|
1
|
|
|
|
|
|
|
Tabanidae
|
Ta
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tanyderidae
|
Td
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1
|
|
|
Simuliidae
|
Sm
|
|
|
|
|
|
|
|
|
6
|
|
|
|
|
|
|
|
Stratiomyidae
|
Sd
|
|
|
|
|
|
|
|
|
|
|
|
2
|
|
|
|
|
Syrphidae
|
Sy
|
|
|
8
|
|
|
|
|
2
|
|
|
|
|
|
|
|
|
Ephéméroptères
|
|
Ephemerellidae Ep 2
|
|
Trichoptères
|
|
Hydropsychidae Hp
|
|
Lépidoptères
|
|
Cossidae Cs
|
|
Orthoptères
|
|
Gryllotalpidae Gl
|
|
Arachnides
|
|
Acaridae Ac
|
|
Crustacés
|
|
Potamidae Pm
|
· Les cases vides correspondent à
0
· Les nombres soulignés correspondent aux
immatures
· habitats : EA : surface de l'eau ; PI : pierre ;
VA : vase ; TR : tronc d'arbre ; RA : racine de plantes
2.2.2 Analyse des résultats des études
AFC
Dans cette étude, nous avons ressorti les
différents axes permettant d'avoir le maximum d'information de
départ. Les variables et individus retenus pour l'interprétation
au niveau de chaque axe sont ceux dont la valeur de la contribution est
supérieure à la moyenne de contribution à la formation de
l'axe, et qui ont une qualité de représentation supérieure
ou égale à 0,5 (nombres en gras dans les tableaux 11, 12, 14, 15,
17, 18, 20, et 21). Les axes considérés ont permis de
représenter dans un plan les variables et individus (figures 3, 4, 5, 6,
7, 8, 9, 10, 11, 12 et 13).
2.2.2.1 Etude sur la répartition
saisonnière des macroinvertébrés immatures au niveau des
deux cours d'eau
Nous considérerons dans cette étude, les 3
premiers axes qui fournissent 78,60% des informations pour
l'interprétation des résultats (tableau 10). Les
interprétations au niveau de l'axe 1 concerneront les variables K1 (Kou
pendant l'étiage) et W2 (Wé en début de saison) ainsi que
les familles des Belostomatidae et des Chironomidae.
Au niveau de l'axe 2, seules les familles de
macroinvertébrés, à savoir les Dytiscidae, les
Calopterygidae, les Gomphidae, les Ceratopogonidae,
les Dolichopogonidae, les Tabanidae et les Simulidae
sont bien représentées.
Au niveau de l'axe 3, seulement la famille des Syrphidae
est bien représentée.
Les axes 1 et 2 dans cette étude sont bien
intéressants car ils opposent les périodes d'étiage aux
périodes de pluies au niveau des deux cours d'eau (figures 4 et 5).
Tableau 10: Résultat AFC réalisé sur
la variation saisonnière des immatures de macroinvertébrés
(Valeurs propres et pourcentage de variance)
|
Valeur propre
|
Pourcentage de variance
|
Pourcentage cumulé des variances
|
|
axe 1
|
5.151337e-01
|
3.569915e+01
|
35.69915
|
|
axe 2
|
3.269230e-01
|
2.265601e+01
|
58.35516
|
|
axe 3
|
2.921790e-01
|
2.024823e+01
|
78.60339
|
|
axe 4
|
2.228584e-01
|
1.544425e+01
|
94.04764
|
|
axe 5
|
8.589176e-02
|
5.952363e+00
|
100.00000
|
|
axe 6
|
6.541928e-33
|
4.533605e-31
|
100.00000
|
Tableau 11: Résultat AFC réalisé sur
la variation saisonnière des immatures de macroinvertébrés
(Résultats relatifs aux colonnes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
K1
|
43.8267700
|
22.6679311
|
1.3304094
|
0.69069846
|
0.226718187
|
0.011892232
|
|
K2
|
8.0049528
|
35.2477462
|
27.7045363
|
0.16412329
|
0.458636109
|
0.322174654
|
|
K3
|
3.4392436
|
0.4544536
|
0.3029379
|
0.18329092
|
0.015370676
|
0.009157158
|
|
W1
|
0.5461718
|
0.4854839
|
41.7845656
|
0.01207877
|
0.006813869
|
0.524129289
|
|
W2
|
30.8830200
|
1.7841009
|
7.6312054
|
0.67756713
|
0.024841478
|
0.094963072
|
|
W3
|
13.2998418
|
39.3602844
|
21.2463454
|
0.22800556
|
0.428235483
|
0.206591406
|
Tableau 12: Résultat AFC réalisé sur
la variation saisonnière des immatures de macroinvertébrés
(Résultats relatifs aux lignes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe 3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
Dy
|
0.23834535
|
8.5784666
|
3.29621510
|
0.043166400
|
0.70213742
|
0.252726334
|
|
El
|
1.09332768
|
3.8461821
|
0.06777939
|
0.335255397
|
0.53300168
|
0.008798708
|
|
Pt
|
0.19905005
|
0.3126200
|
0.28021215
|
0.206375113
|
0.14648228
|
0.122992265
|
|
Be
|
69.73373814
|
3.0771676
|
0.54951105
|
0.977038148
|
0.01948467
|
0.003259424
|
|
Cp
|
0.45670558
|
19.3494915
|
0.15576302
|
0.048275099
|
0.92433568
|
0.006970229
|
|
Gd
|
0.23834535
|
8.5784666
|
3.29621510
|
0.043166400
|
0.70213742
|
0.252726334
|
|
Ll
|
0.29861298
|
1.5248093
|
0.38462693
|
0.226497673
|
0.52268964
|
0.123506708
|
|
Ce
|
0.04044802
|
14.4234450
|
0.06340166
|
0.006135607
|
0.98878661
|
0.004071521
|
|
Ch
|
18.49683244
|
9.1192346
|
5.37500059
|
0.736640949
|
0.16413084
|
0.090621827
|
|
Dp
|
0.02022401
|
7.2117225
|
0.03170083
|
0.006135607
|
0.98878661
|
0.004071521
|
|
Mu
|
0.19905005
|
0.3126200
|
0.28021215
|
0.206375113
|
0.14648228
|
0.122992265
|
|
Ta
|
0.02022401
|
7.2117225
|
0.03170083
|
0.006135607
|
0.98878661
|
0.004071521
|
|
Td
|
0.15273026
|
0.3367476
|
0.70800105
|
0.018138820
|
0.01807431
|
0.035597022
|
|
Sm
|
0.51925370
|
6.7819873
|
0.75061780
|
0.109299519
|
0.64516222
|
0.066888786
|
|
Sd
|
0.06784051
|
0.3897160
|
0.72588207
|
0.008063727
|
0.02093477
|
0.036526542
|
|
Sy
|
2.70663510
|
0.9620399
|
83.88348761
|
0.069897969
|
0.01122797
|
0.917079808
|
|
Ep
|
1.32380913
|
0.6191316
|
0.03871843
|
0.718849703
|
0.15193890
|
0.008900744
|
|
Hp
|
4.17460364
|
0.1527068
|
0.04925341
|
0.973876449
|
0.01609980
|
0.004864304
|
|
Cs
|
0.02022401
|
7.2117225
|
0.03170083
|
0.006135607
|
0.98878661
|
0.004071521
|

Cp
Gd Dy
K2
Dpe Cs Ta
Sm
El
Ep
HK1
fBe
Ll
Sd
Td Mu Pt
K3 W1
3 W2
Ch Sy
|
CI m2 (21.83P/4
|
-0.5 0.0 0.5 1.0 1.5 20 25 30
|
|
-2 -1 0 1
Dim 1 (48.45%)
Figure 3: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés
immatures au niveau du Kou et du Wé en période d'étiage,
en début de saison pluvieuse et pendant les hautes eaux (axes 1 et
2)
-1 0 1 2 3
Di m3 (2a51%)
CA factor map

Sy
W1
Gd
Dy
HpBeK1
EpEl
CpLl
Sd
K2 e Dp Ch Cs
Ta K3Sm W2
W3 Mu
Pt
Td
-3 -2 -1 0 1
Dim 1 (48.45%)
Figure 4: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés
immatures au niveau du Kou et du Wé en période d'étiage,
en début de saison pluvieuse et pendant les hautes eaux (axes 1 et
3)

Sy
W1
Gd Dy
Be Hp K
1
Sd
Mu
Pt W3
Td
Ep El Cp
K2
Ch Ll e
Cs
Dp
Ta
K3
W2 Sm
-1 0 1 2 3
Di m3 (2:151N
-1 0 1 2 3
Dim 2 (21.89%)
Figure 5: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés
immatures au niveau du Kou et du Wé en période d'étiage,
en début de saison pluvieuse et pendant les hautes eaux (axes 2 et
3)
2.2.2.2 Etude sur la répartition
saisonnière des macroinvertébrés adultes au niveau des
deux cours d'eau
Les trois premiers axes dans cette étude fournissent
78,12% des informations de départ (tableau 13). Nous considérons
ces axes pour l'interprétation de nos résultats. Au niveau de
l'axe 1, les variables W2 (Wé pendant l'étiage) et W3 (Wé
en hautes eaux) sont bien représentées ainsi que les familles des
Belostomatidae, des Naucoridae des Physidae et des
Oligocheta.
Au niveau de l'axe 2, la variable K3 (Kou en hautes eaux) et
les familles des Elmidae, des Gyrinidae, des
Cydnidae, des Gerridae, des Ceratopogonidae, des
Culicidae et des Acaridae sont bien
représentées.
Les familles des Chrysomelidae, des
Cucurlionidae, des Histeridae, des Hebidae, des
Notonectidae, des Gryllotalpidae, des Potamidae, des
Ancylidae et des Valvatidae sont bien
représentées par l'axe 3. Cet axe oppose les périodes de
début de saison pluvieuse des deux cours d'eaux aux autres
périodes (figures 7 et 8).
Tableau 13: Résultat AFC réalisé sur
la variation saisonnière des adultes de macroinvertébrés
(Valeurs propres et pourcentage de variance)
|
Valeur propre
|
Pourcentage de variance
|
Pourcentage cumulé des variances
|
|
axe 1
|
4.492465e-01
|
3.641748e+01
|
36.41748
|
|
axe 2
|
2.982156e-01
|
2.417439e+01
|
60.59187
|
|
axe 3
|
2.162705e-01
|
1.753164e+01
|
78.12351
|
|
axe 4
|
1.502710e-01
|
1.218148e+01
|
90.30499
|
|
axe 5
|
1.195977e-01
|
9.695008e+00
|
100.00000
|
|
axe 6
|
3.380905e-33
|
2.740679e-31
|
100.00000
|
Tableau 14: Résultat AFC réalisé sur
la variation saisonnière des adultes de macroinvertébrés
(Résultats relatifs aux colonnes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe 3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
K1
|
0.001204995
|
9.351969
|
6.670239
|
4.266855e-05
|
0.21982210
|
0.11370423
|
|
K2
|
16.849640680
|
3.488594
|
38.757159
|
4.258369e-01
|
0.05852598
|
0.47153853
|
|
K3
|
6.088432209
|
58.618990
|
16.792264
|
1.130872e-01
|
0.72275647
|
0.15015158
|
|
W1
|
25.295807147
|
12.439101
|
25.805524
|
4.603605e-01
|
0.15027409
|
0.22608656
|
|
W2
|
29.073681510
|
9.518742
|
9.940614
|
5.521034e-01
|
0.11999014
|
0.09087537
|
|
W3
|
22.691233459
|
6.582603
|
2.034200
|
5.004878e-01
|
0.09637818
|
0.02159940
|
Tableau 15: Résultat AFC réalisé sur
la variation saisonnière des adultes de macoinvertébrés
(Résultats relatifs aux lignes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe 3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
Cb
|
1.104036e+01
|
0.93360373
|
5.4438398
|
0.4020625
|
0.02256931
|
0.09543950
|
|
Cy
|
1.134216e-01
|
0.00228377
|
13.621695
|
0.0156998
|
0.0002098
|
0.9077028
|
|
Cu
|
7.654377e-01
|
0.23873935
|
3.6572828
|
0.25727408
|
0.05326670
|
0.59177597
|
|
Dy
|
1.691501e+00
|
0.03014421
|
0.6185792
|
0.22555312
|
0.00266824
|
0.03970860
|
|
El
|
5.084137e+00
|
9.34884530
|
0.4358089
|
0.41215608
|
0.50309272
|
0.01700799
|
|
Gy
|
1.453530e-01
|
13.3792433
|
1.0352326
|
0.01496079
|
0.91412949
|
0.05129571
|
|
Hi
|
7.654377e-01
|
0.23873935
|
3.6572828
|
0.25727408
|
0.05326670
|
0.59177597
|
|
Hd
|
1.113987e+00
|
1.75706075
|
3.1538238
|
0.23225185
|
0.24317071
|
0.31654023
|
|
Hy
|
4.784205e-01
|
0.30649800
|
0.1427430
|
0.09718460
|
0.04132957
|
0.01395903
|
|
Sp
|
8.128054e-05
|
0.95029581
|
0.9346093
|
0.00001569
|
0.12183299
|
0.08689668
|
|
St
|
9.343841e-01
|
1.49945381
|
7.5563136
|
0.12261503
|
0.13061617
|
0.47735430
|
|
Be
|
8.933094e+00
|
0.24034454
|
2.0006293
|
0.69166324
|
0.01235300
|
0.07457135
|
|
Cn
|
5.892410e-01
|
8.54634025
|
3.3758576
|
0.07265188
|
0.69948711
|
0.20037827
|
|
Gr
|
7.041246e-01
|
7.00403898
|
0.4516169
|
0.08508126
|
0.56179593
|
0.02627043
|
|
He
|
7.654377e-01
|
0.23873935
|
3.6572828
|
0.25727408
|
0.05326670
|
0.59177597
|
|
Na
|
7.303663e+00
|
1.60002434
|
0.0147150
|
0.86138848
|
0.12526522
|
0.00083547
|
|
Np
|
1.010879e+01
|
6.86789586
|
13.262226
|
0.40049713
|
0.18062158
|
0.25294690
|
|
No
|
2.296313e+00
|
0.71621806
|
10.971848
|
0.25727408
|
0.05326670
|
0.59177597
|
|
Ce
|
5.892410e-01
|
8.54634025
|
3.3758576
|
0.07265188
|
0.69948711
|
0.20037827
|
|
Cl
|
5.892410e-01
|
8.54634025
|
3.3758576
|
0.07265188
|
0.69948711
|
0.20037827
|
|
Gl
|
7.654377e-01
|
0.23873935
|
3.6572828
|
0.25727408
|
0.05326670
|
0.59177597
|
|
Ac
|
5.753711e-03
|
5.82094722
|
0.0028163
|
0.00144738
|
0.97201734
|
0.00034106
|
|
Pm
|
7.654377e-01
|
0.23873935
|
3.6572828
|
0.25727408
|
0.05326670
|
0.59177597
|
|
An
|
7.654377e-01
|
0.23873935
|
3.6572828
|
0.25727408
|
0.05326670
|
0.59177597
|
|
Py
|
3.651832e+00
|
0.80001217
|
0.0073575
|
0.86138848
|
0.12526522
|
0.00083547
|
|
Pb
|
3.748718e-01
|
0.11820570
|
0.4471273
|
0.14941755
|
0.03127539
|
0.08579513
|
|
Ly
|
2.895055e+00
|
1.72896808
|
1.4476222
|
0.60724318
|
0.24073456
|
0.14617520
|
|
Vv
|
7.654377e-01
|
0.23873935
|
3.6572828
|
0.25727408
|
0.05326670
|
0.59177597
|
|
Hr
|
1.287240e-03
|
0.04753681
|
1.5963557
|
0.00018008
|
0.00441468
|
0.10751443
|
|
Og
|
3.599778e+01
|
19.5381831
|
1.1264856
|
0.64043012
|
0.23074155
|
0.00964793
|

Np
W1
Be
Ly
Na Py
CuAn He Pm No Vv Gl Hi
K2
Pb
Hd
Dy Cy H
Hy
Ac
Sp
K1
r
Gy
Gr
St
K3
El
eCl Cn
Cb
W2
Og
W3
|
Clm2(24.17°/4
|
-20 -1.5 -1.0 -Q5 QO Q5 1.0
|
|
-1 0 1 2
Dim 1 (36.42%)
Figure 6: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés adultes
au niveau du Kou et du Wé en période d'étiage, en
début de saison pluvieuse et pendant les hautes eaux (axes 1 et
2)
CA factor map
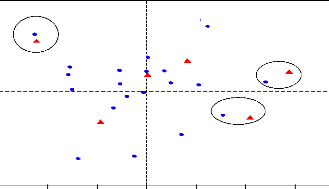
eCl Cn
Np W1
Sp
K3
Ly
Gy
Hd
K1
Hr
W3
Be
Og
Na
Py
Ac
Hy
Pb
Cb
W2
K2
St
Cy
An Pm Cu Vv No He Gl Hi
Dy Gr El
|
rri3(17.53'/4
|
-1.0 -Q5 QO Q5 1.0
|
|
-1.0 -0.5 0.0 0.5 1.0 1.5
Dim 1 (36.42%)
Figure 7: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés adultes
au niveau du Kou et du Wé en période d'étiage, en
début de saison pluvieuse et pendant les hautes eaux (axes 1 et
3)

Di m3 (17.53X
-1.5 -1.0 -0.5 0.0 0.5 1.0
K3
Sp
LyW3
Gy
Hr
Be
Hd K1
GrEl
Og
Dy
Ac
Na
Py
Hy
eCl Cn
Np
W1
Pb
Cb
K2
St
PmAnCu Cy He No Vv Gl Hi
W2
-2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0
Dim 2 (24.17%)
Figure 8: Résultat AFC; comparaison de la
distribution spatiale des populations de macroinvertébrés adultes
au niveau du Kou et du Wé en période d'étiage, en
début de saison pluvieuse et pendant les hautes eaux (axes 2 et
3)
2.2.2.3 Etude de l'habitat des
macroinvertébrés immatures
Les deux premiers axes nous fournissent 79,49% des informations
(tableau 16). Mais pour plus de précision, les investigations se
porteront au niveau du troisième axe.
La variable EA (eau) et les familles des Belostomatidae
et des Chironomidae sont bien représentées au niveau de
l'axe 1 (tableau17). Au niveau de l'axe 2, les familles des Elmidae,
des Stratiomyidae, des Syrphidae et des
Hydropsychidae sont bien représentées (tableaux 17 et
18) tandis que sur l'axe 3, le famille des Gomphidae et celle des
Libellulidae ont une bonne représentation.
L'axe 1 oppose les supports (pierre, racine, vase tronc) à
la surface de l'eau (figures 9 et 10).
Tableau 16: Résultat AFC réalisé sur
les habitats des immatures de macroinvertébrés (Valeurs propres
et pourcentage de variance)
|
Valeur propre
|
Pourcentage de variance
|
Pourcentage cumulides variances
|
|
axe 1
|
7.592157e-01
|
6.365448e+01
|
63.65448
|
|
axe 2
|
7.592157e-01
|
1.583969e+01
|
79.49417
|
|
axe 3
|
1.713634e-01
|
1.436752e+01
|
93.86169
|
|
axe 4
|
7.321244e-02
|
6.138308e+00
|
100.00000
|
|
axe 5
|
1.708076e-32
|
1.432092e-30
|
100.00000
|
Tableau 17: Résultat AFC réalisé sur
les habitats des immatures de macroinvertébrés (Résultats
relatifs aux colonnes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe 3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
EA
|
7.838086e+01
|
0.4364121
|
0.7593135
|
9.964025e-01
|
0.0013805
|
0.0021787
|
|
PI
|
8.322239e+00
|
1.3831737
|
12.3242112
|
5.159247e-01
|
0.0213373
|
0.1724479
|
|
VA
|
9.123611e+00
|
41.1320678
|
11.5283495
|
4.019834e-01
|
0.4509616
|
0.1146466
|
|
TR
|
4.172664e+00
|
23.8984651
|
15.4810577
|
2.352977e-01
|
0.3353450
|
0.1970414
|
|
RA
|
6.302116e-04
|
33.1498813
|
59.9070681
|
2.881163e-05
|
0.3771214
|
0.6181763
|
Tableau 18: Résultat AFC réalisé sur
les habitats des immatures de macroinvertébrés (Résultats
relatifs aux lignes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe 3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
Dy
|
0.46008323
|
5.64352982
|
5.8588458
|
0.1135020
|
3.4644e-01
|
3.2623e-01
|
|
El
|
0.63836952
|
6.35845295
|
0.0569709
|
0.2745178
|
6.8040e-01
|
5.5297e-03
|
|
Pt
|
0.15702913
|
3.61425726
|
2.5811577
|
0.0483649
|
2.7700e-01
|
1.7943e-01
|
|
Be
|
67.6718566
|
0.01469313
|
0.0191706
|
0.9998189
|
5.4018e-05
|
6.3929e-05
|
|
Cp
|
0.00347242
|
14.8653588
|
16.688641
|
0.0004555
|
4.8527e-01
|
4.9416e-01
|
|
Gd
|
0.20397365
|
1.41720844
|
7.6068842
|
0.0807594
|
1.3962e-01
|
6.7979e-01
|
|
Ll
|
0.36394433
|
7.30248228
|
35.044472
|
0.0350523
|
1.7501e-01
|
7.6182e-01
|
|
Ce
|
0.44630660
|
0.36458834
|
0.3534820
|
0.3294030
|
6.6959e-02
|
5.8886e-02
|
|
Ch
|
11.8514250
|
3.64033424
|
3.3165394
|
0.8610355
|
6.5812e-02
|
5.4386e-02
|
|
Cl
|
13.8975910
|
0.31096292
|
0.5964826
|
0.9845871
|
5.4820e-03
|
9.5381e-03
|
|
Dp
|
0.14615502
|
0.09761860
|
0.9589144
|
0.1178983
|
1.9594e-02
|
1.7459e-01
|
|
Ta
|
0.15406606
|
2.79127765
|
0.8624908
|
0.1314418
|
5.9258e-01
|
1.6608e-01
|
|
Td
|
0.15702913
|
3.61425726
|
2.5811577
|
0.0483649
|
2.7700e-01
|
1.7943e-01
|
|
Sm
|
0.90925983
|
7.34977016
|
10.029852
|
0.1755311
|
3.5306e-01
|
4.3703e-01
|
|
Sd
|
0.34123855
|
8.83937845
|
0.3985099
|
0.1207432
|
7.7829e-01
|
3.1827e-02
|
|
Sy
|
1.54066062
|
27.9127765
|
8.6249086
|
0.1314418
|
5.9258e-01
|
1.6608e-01
|
|
Ep
|
0.60325204
|
0.18287930
|
1.7376220
|
0.5728528
|
4.3214e-02
|
3.7243e-01
|
|
Hp
|
0.30813212
|
5.58255530
|
1.7249817
|
0.1314418
|
5.9258e-01
|
1.6608e-01
|
|
Cs
|
0.14615502
|
0.09761860
|
0.9589144
|
0.1178983
|
1.9594e-02
|
1.7459e-01
|

Di m2 (15.84°A)
- 1.5 -1.0 -0.5 0.0 Q5 1.0 1.5
S
Ddy
SmEl
TdPt
EABe
Ch Cl
Ce
TaHp Sy
EpDp Cs PI
VA
TRGd
Ll
RA Cp
-1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5
Dim 1 (63.65%)
Figure 9: Résultat AFC; distribution spatiale des
populations de macroinvertébrés immatures au niveau des habitats
(axes 1 et 2)
CA factor map

Ci m3 (14.37°/4
- 1.0 -Q5 OD 0.5 1.0 1.5
Ll
RA
Gd
Cp
Hp
Ta Sy
VASd
El
Cl
CeP
I
EpTR
Dp
Cs
Sm Dy
TdPt
Ch EABe
-1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5
Dim 1 (63.65%)
Figure 10: Résultat AFC; distribution spatiale des
populations de macroinvertébrés immatures au niveau des habitats
(axes 1 et 3)
CA factor map

Ll
RA
Gd
Cp
TR
Sm
Dy
Td Pt
Hp
Ta Sy
VA
Be Ch EA
Ce Cl PI
Ep
Dp
Cs
-1.0 -0.5 0.0 0.5 1.0 1.5
Di m3 (14.37Y4
Sd
El
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5
Dim 2 (15.84%)
Figure 11: Résultat AFC; distribution spatiale des
populations de macroinvertébrés immatures au niveau des habitats
(axes 2 et 3)
2.2.2.4 Etude de l'habitat des
macroinvertébrés adultes
Les trois premiers axes dans cette étude qui fournissent
88,51% des informations seront considérés pour l'analyse (tableau
19).
L'axe 1 donne une bonne représentation de la variable EA
(eau) et les familles des Gyrinidae et des Belostomatidae.
Au niveau de l'axe 2, la variable VA (vase) et les familles des
Carabidae, des Dytiscidae, des Ancylidae, des
Physidae, des Lymnaeidae et des Valvatidae sont bien
représentées.
Les familles des Gryllotalpidae, des Acaridae,
des Potamidae et des Hirudinae ont une bonne
représentation sur l'axe 3.
Tableau 19: Résultat AFC réalisé sur
les habitats des adultes de macroinvertébrés (Valeurs propres et
pourcentage de variance)
|
Valeur Propre
|
Pourcentage de variance
|
Pourcentage cumulé des variance
|
|
axe 1
|
5.187083e-01
|
4.223212e+01
|
42.23212
|
|
axe 2
|
3.645813e-01
|
2.968343e+01
|
71.91554
|
|
axe 3
|
2.038501e-01
|
1.659703e+01
|
88.51257
|
|
axe 4
|
1.410922e-01
|
1.148743e+01
|
100.00000
|
|
axe 5
|
8.683623e-33
|
7.070019e-31
|
100.00000
|
Tableau 20: Résultat AFC réalisé sur
les habitats des adultes de macroinvertébrés (Résultats
relatifs aux colonnes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe 3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
EA
|
64.308301
|
5.55488512
|
0.2467263
|
0.94025370
|
0.0570852990
|
0.001417689
|
|
PI
|
8.693417
|
5.84485271
|
38.8120896
|
0.24308311
|
0.1148706407
|
0.426499900
|
|
VA
|
18.105641
|
61.69121352
|
7.4640042
|
0.27614389
|
0.6613266491
|
0.044738459
|
|
TR
|
6.956849
|
0.00442874
|
6.5829067
|
0.25289797
|
0.0001131577
|
0.094045513
|
|
RA
|
1.935793
|
26.90461990
|
46.8942732
|
0.04894006
|
0.4780833910
|
0.465921811
|
Tableau 21: Résultat AFC réalisé sur
les habitats des adultes de macroinvertébrés (Résultats
relatifs aux lignes: contributions et qualité de
représentation)
|
Contributions
|
Qualité de représentation
|
|
Axe 1
|
Axe 2
|
Axe 3
|
Axe 1
|
Axe 2
|
Axe 3
|
|
Cb
|
0.463249
|
4.312971e+00
|
0.0940670
|
0.1295949
|
8.4804e-01
|
0.0103418
|
|
Cy
|
0.033390
|
1.450791e+00
|
4.1118212
|
0.0103668
|
3.1659e-01
|
0.5017002
|
|
Cu
|
0.462478
|
4.188782e-04
|
1.1135477
|
0.0874223
|
5.5653e-05
|
0.0827232
|
|
Dy
|
0.816959
|
3.792478e+00
|
0.2534608
|
0.2117208
|
6.9080e-01
|
0.0258143
|
|
El
|
2.785342
|
1.361025e+00
|
0.0607894
|
0.7396313
|
2.5402e-01
|
0.0063438
|
|
Gy
|
8.855554
|
1.088310e+00
|
0.0864523
|
0.9131778
|
7.8879e-02
|
0.0035035
|
|
Hi
|
0.462478
|
4.188782e-04
|
1.1135477
|
0.0874223
|
5.5653e-05
|
0.0827232
|
|
Hd
|
0.086495
|
1.298146e+00
|
1.4696689
|
0.4477592
|
3.8646e-01
|
0.2446345
|
|
Hy
|
4.424020
|
2.770505e-01
|
0.9216746
|
0.4477592
|
1.9708e-02
|
0.0366600
|
|
Sp
|
2.951851
|
3.627700e-01
|
0.0288174
|
0.9131778
|
7.8879e-02
|
0.0035035
|
|
St
|
2.180837
|
1.930468e+00
|
4.7675665
|
0.3873403
|
2.4099e-01
|
0.3327773
|
|
Be
|
5.292386
|
2.483914e-01
|
4.0223808
|
0.7441247
|
2.4547e-02
|
0.2222618
|
|
Cn
|
2.951851
|
3.627700e-01
|
0.0288174
|
0.9131778
|
7.8879e-02
|
0.0035035
|
|
Gr
|
23.61481
|
2.902160e+00
|
0.2305394
|
0.9131778
|
7.8879e-02
|
0.0035035
|
|
He
|
2.951851
|
3.627700e-01
|
0.0288174
|
0.9131778
|
7.8879e-02
|
0.0035035
|
|
Na
|
2.781114
|
2.405768e-01
|
0.0882355
|
0.9276746
|
5.6402e-02
|
0.0115666
|
|
Np
|
0.922801
|
9.158097e+00
|
12.340126
|
0.0689214
|
4.8075e-01
|
0.3622042
|
|
No
|
8.855554
|
1.088310e+00
|
0.0864523
|
0.9131778
|
7.8879e-02
|
0.0035035
|
|
Ce
|
0.644605
|
6.166033e-01
|
7.3228955
|
0.1064194
|
7.1549e-02
|
0.4751135
|
|
Cl
|
2.951851
|
3.627700e-01
|
0.0288174
|
0.9131778
|
7.8879e-02
|
0.0035035
|
|
Gl
|
0.113089
|
2.236241e+00
|
6.9709991
|
0.0252185
|
3.5050e-01
|
0.6109162
|
|
Ac
|
0.226178
|
4.472481e+00
|
13.941998
|
0.0252185
|
3.5050e-01
|
0.6109162
|
|
Pm
|
0.113089
|
2.236241e+00
|
6.9709991
|
0.0252185
|
3.5050e-01
|
0.6109162
|
|
An
|
2.908770
|
1.410093e+01
|
3.0512640
|
0.1977693
|
6.7385e-01
|
0.0815298
|
|
Py
|
5.987697
|
1.123769e+01
|
0.0179237
|
0.4231405
|
5.5817e-01
|
0.0004977
|
|
Pb
|
1.289210
|
1.233207e+00
|
14.645791
|
0.1064194
|
7.1549e-02
|
0.4751135
|
|
Ly
|
5.578981
|
1.869890e+01
|
1.9818109
|
0.2851698
|
6.7179e-01
|
0.0398105
|
|
Vv
|
2.908770
|
1.410093e+01
|
3.0512640
|
0.1977693
|
6.7385e-01
|
0.0815298
|
|
Hr
|
0.264677
|
2.985179e-02
|
9.2668633
|
0.0638497
|
5.0615e-03
|
0.8785410
|
|
Og
|
6.120045
|
4.362360e-01
|
1.9025899
|
0.6918662
|
3.4662e-02
|
0.0845279
|
CA factor map

CnrHeSp Gy No Cl
EA
BeNa
Dy
Hy
Hr TR Cu
Hi
Cy
Hd
Cb
RA
Np GlAcPm
og El PI S
t Ce Pb
An
Vv
Ly VA
Py
-1 0 1 2 3
m2 029.68°A
-2 -1 0 1 2
Dim 1 (42.23%)
Figure 12: Résultat AFC; distribution spatiale des
populations de macroinvertébrés adultes au niveau des habitats
(axes 1 et 2)
CA factor map

Ci m3 (166N
-1.5 -1.0 -Q5 QO Q5 1.0 1.5
GlAcPm
An
Vv
Np
RA
Cy
VALy
Be
Na
Py
EA
CnrHeSp No Gy Cl
Hd Hy
TR og
Cu St PI Hi
Hr
Ce
Pb
Dy
Cb El
-2 -1 0 1 2
Dim 1 (42.23%)
Figure 13: Résultat AFC; distribution spatiale des
populations de macroinvertébrés adultes au niveau des habitats
(axes 1 et 3)
2.3 Densité du benthos sur les 2 cours d'eau
Sur le Kou, les hautes eaux sont favorables au benthos, alors
qu'au niveau du Wé la densité
est élevée en début de saison pluvieuse
(figure 14).
étiage début de hautes saison eaux
pluvieuse
nombre
d'individus
au m2
|
500 450 400 350 300 250 200 150 100 50
0
|
|
|
|
|
Kou Wé
|
|
|
Figure 14: Evolution de la densité du benthos au
niveau du Kou et du Wé
2.4 Etude de l'alimentation de C. gariepinus au niveau
du Kou
2.4.1 Caractéristiques des poissons
Au total 181 poissons ont été capturé au
niveau de la station de Bama : 2 pendant la période
d'étiage, 17 en début de saison pluvieuse et 162
pendant les hautes eaux.
Il existe une bonne corrélation entre la taille et le
poids de ces poissons (coefficient de corrélation est de
0,947)
La relation Longueur-poids des poissons est : P =
0,019L3,015
Les limites de confiance de b (3,015) pour un risque d'erreur de
5% sont 2,918 et 3,112 : soit 3,015 #177; 0,097.
Le facteur de Fulton traduisant les conditions des poissons
à chaque période est consigné dans le tableau 10.
Tableau 22: Variation saisonnière du facteur de
condition de Fulton de C. gariepinus au niveau du Kou
|
Périodes
|
Etiage
|
Début de saison
|
Hautes eaux
|
|
Facteur
|
0,0077
|
0,0059
|
0,0088
|
2.4.2 Habitude alimentaire des poissons
Le coefficient de vacuité (pourcentage de l'effectif des
poissons ayant l'estomac vide) est à
71,82%. La moyenne du bol alimentaire des poissons ayant au moins
une proie dans l'estomac est de 0,91 g avec un écart à la moyenne
de 0,75 g.
L'indice d'occurrence calculé pour chaque type d'aliment
est consigné dans le tableau 11.
Tableau 23: Indice d'occurrence des différentes
proies
|
Proies
|
Indice (%)
|
|
Insectes
|
76.47
|
|
Odonates
|
23.52
|
|
* Libellulidae
|
9.80
|
|
* Calopterygidae
|
7.84
|
|
* Gomphidae
|
1.96
|
|
* ni
|
7.84
|
|
Coléoptères
|
35.29
|
|
* Dytiscidae
|
19.60
|
|
* Carabidae
|
5.88
|
|
* Hydrophilidae
|
1.96
|
|
* ni
|
13.72
|
|
Trichoptères
|
1.96
|
|
* Hydropsychidæ
|
1.96
|
|
Hyménoptères
|
7.84
|
|
* Formicidae
|
7.84
|
|
Plécoptères (ni)
|
1.96
|
|
Diptères
|
5.88
|
|
* Stratiomyidae
|
1.96
|
|
* Simuliidae
|
1.96
|
|
* Chironomidae
|
1.96
|
|
Orthoptères
|
1.96
|
|
* Tettigoniidae
|
1.96
|
|
Hétéroptères
|
21.56
|
|
* Belostomatidae
|
9.80
|
|
* Notonectidae
|
3.92
|
|
* ni
|
7.84
|
|
Insectes (ni)
|
17.64
|
|
Mollusques
|
7.84
|
|
* Planorbidae
|
3.92
|
|
* Physidae
|
7.84
|
|
* Lymnaeidae
|
1.96
|
|
* Sphaeriidae
|
1.96
|
|
Poisson
|
11.76
|
|
Batraciens (grenouilles)
|
1.96
|
|
Plantes
|
35.29
|
|
* débris
|
33.33
|
|
* grains de riz
|
3.92
|
|
Terre
|
1.96
|
* ni : non identifié
2.4.3 Impact de la densité des
macroinvertébrés benthiques sur C. gariepinus
Le facteur de Fulton calculé avec les moyennes des tailles
et des poids évolue dans le même sens que la densité
benthique (Figure 15).
|
0.01
|
|
600
|
|
|
|
|
0.009
0.008
0.007
|
500
400
|
densit6 du benthos (nombre d'individus au m2)
|
|
|
|
facteur de Fulton
|
0.006
|
|
|
|
|
0.005
|
300
|
|
facteur de Fulton densité du benthos
|
|
0.004
0.003
0.002
0.001
0
|
200 100 0
|
|
|
étiage Début de Hautes
saison eaux
pluieuse
Figure 15: Variation de la densité des
macroinvertébrés benthiques et du facteur de condition de Fulton
de C. gariepinus au niveau du Kou
III- DISCUSSION
3.1 Les populations de
macroinvertébrés
Les populations de macroinvertébrés sur ces deux
cours d'eau sont dominées par les Insectes (tableau 4 et 9), et parmi
ces derniers les Chironomidae sont les plus nombreux surtout au niveau
du benthos. Ces remarques, en adéquation avec les résultats de
Kabré et al. (2000) et de Kabré et al. (2002),
confirment ceux de Guenda (1996) réalisé sur le réseau
hydrographique du Mouhoun. Le Kou est en effet un affluent de ce fleuve. En
outre, des 8 ordres d'insectes inventoriés par Guenda (1996), 7 ont des
représentants au niveau du Kou à savoir les Diptères, les
Trichoptères, les Ephéméroptères, les
Coléoptères, les Odonates, les Hétéroptères
et les Lépidoptères.
Les Chironomidae étant très nombreux au
niveau du benthos de ces 2 cours d'eau, ils influenceront donc la
densité des individus benthiques. C'est ainsi que les fortes
concentrations des Chironomidae en début de saison pluvieuse
sur le Wé relève considérablement la densité du
benthos sur ce cours d'eau par rapport au Kou. Mais pendant les hautes eaux, la
densité sur le Wé baisse (Figure 14).
Les valeurs de la densité benthique au niveau de ces deux
cours d'eau sont en deçà de celles obtenues par Guenda (1996).
3.1.1 Macroinvertébrés et qualité de
l'eau
Les caractéristiques physico-chimiques sur ces deux
cours d'eau ne permettent pas d'évaluer leur qualité. Le pH tend
a augmenté de l'étiage aux hautes eaux tandis que la
conductivité diminue (tableau3). Mais ces valeurs sont en
deçà des valeurs critiques. L'utilisation des traits biologiques
des espèces vivants dans ces cours d'eau s'avère
nécessaire.
Le niveau taxonomique atteint dans la détermination des
individus dans cette étude ne permet de faire qu'une analyse partielle
sur la qualité de ces deux cours. En effet, ils se caractérisent
par une faible quantité de EPT (Ephéméroptères,
Plécoptères, Trichoptères), avec 0,016% sur le Kou et
0.008% sur le Wé (tableaux 4 et 9). Les espèces appartenant
à ces ordres d'Insectes, très sensibles à la pollution
témoignent de par leur abondance dans un cours d'eau, une bonne
santé écologique (Moisan et Pelletier, 2008). Ces données
relatent une mauvaise qualité de ces deux cours d'eau pendant notre
étude.
3.1.2 Répartition saisonnière des
macroinvertébrés
La répartition des macroinvertébrés
observés dans cette étude est un phénomène naturel
qui est une expression du cycle biologique des espèces au niveau des
cours d'eau. Dejoux et al. (1971) affirmaient en ce sens que les
macroinvertébrés sont soumis périodiquement au rythme des
variations saisonnières.
3.1.2.1 Répartition saisonnière des
immatures
Les résultats de l'AFC relatives aux immatures (figure
3) indiquent une attirance des trois périodes au niveau du Wé et
le Kou pendant les hautes eaux. Ces périodes sont regroupées dans
un même quadrant avec les immatures des Muscidae, des
Syrphidae et des Chironomidae. Ces
macroinvertébrés se retrouvent donc pendant ces périodes
au niveau de ces cours d'eau.
La disposition des trois périodes du Kou, indique une
variation des macroinvertébrés immatures. Les
Hydropsychidae et les Belostomatidae sont présents
pendant l'étiage, les Simuliidae et les Dytiscidae
plus nombreux en début de saison et les Chironomidae
prolifèrent pendant la période des hautes eaux.
Les périodes d'étiage se caractérisent par
une faible quantité de Diptères. Ce constat avait
été relaté par Kabré et al. (2002).
3.1.2.2 Répartition saisonnière des
adultes
Les figures 6, 7 et 8 présentent des oppositions et des
affinités entre les périodes d'étude au niveau des deux
cours d'eau. Il existe ainsi une affinité entre les périodes de
début de saison pluvieuse et les hautes eaux au niveau du Wé
(figure 6). Ces deux périodes se caractérisent par une
présence d'Oligocheta. La figure 7 indique les
macroinvertébrés retrouvés pendant les périodes
d'étude au niveau du Wé. Ainsi les Nepidae se retrouvent
pendant l'étiage, les Oligocheta en début de saison et
les Carabidae pendant les hautes eaux. Il existe une affinité
entre ces deux périodes du Wé avec la période
d'étiage au niveau du Kou. Ces périodes se situent dans un
même quadrant. Les Nepidae se retrouvent au niveau du Wé
pendant la période d'étiage, les Lymnaeidae et les
Oligocheta sont plus fréquents au niveau du Wé pendant
les hautes eaux et les Hydraenidae au niveau du Kou en période
d'étiage.
3.1.3 Habitats des macroinvertébrés
3.1.3.1 Habitats des immatures
Cette étude ressort l'affinité des
macroinvertébrés immatures pour les supports dans les cours
d'eaux. En effet, la grande majorité se retrouve
accrochée (figures 9 et 10). L'axe 1 oppose les supports (pierre,
racine, vase tronc) à la surface de l'eau (figures 9 et 10). Les
Odonates (Gomphidae, Libellulidae et Calopterygidae)
ont une affinité pour les racines (figure 11).
Les Belostomatidae et Culicidae se rencontrent
à la surface de l'eau (figure 9 et 10).
3.1.3.2 Habitats des adultes
Les adultes de macroinvertébrés dans ces deux cours
d'eau se repartissent aussi en deux
groupes. En effet l'axe 3 dans cette étude oppose la
surface de l'eau et les substrats (figure 12 et 13). Les familles des
Gyrinidae, des Gerridae, des Belostomatidae, des
Naucoridae, des Dytiscidae et des Hirudinae se
trouvent à la surface ou nageant dans l'eau. Au niveau des substrats,
les Nepidae sont accrochés aux racines, les Lymnaeidae
et les Physidae se retrouvent au niveau de la vase.
3.2 Régime alimentaire de C. gariepinus
Selon Corbet (1958, 1961), C. gariepinus a un
régime omnivore. Les jeunes se nourrissent d'insectes divers, de
graines, de débris de végétaux ; les adultes consomment
des poissons et des mollusques. Ce régime se retrouve chez ce poisson au
niveau du Kou. Les Arthropodes, précisément les Insectes sont les
proies préférées de C. gariepinus. Les
débris de plantes et les petits poissons constituent leur alimentation
secondaire, tandis que les batraciens sont leurs proies accidentelles. Ces
batraciens ont été retrouvés au niveau des estomacs des
poissons avant la saison des pluies. Cette période se caractérise
par un faible taux de macroinvertébrés adultes au niveau de la
rivière.
Pendant les hautes eaux, les proies des poissons sont
constitués d'adultes d'Insectes avec une préférence pour
les Coléoptères. Ce constat ne met nullement en doute les
résultats de Coffman et Ferrington (1994) qui affirmaient que la plupart
des prédateurs aquatiques se nourrissent de Chironomidae a un
moment donné de leur développement. En effet la bonne
corrélation entre la densité des macroinvertébrés
benthiques et le facteur de condition de Fulton (figure 15), indique que ces
poissons sont bien nourris pendant les périodes de prolifération
des Chironomidae. Cependant, la chute de la valeur en début de
saison serait liée
au stress éventuel lié à tout début
de saison des pluies dans les plans d'eau au Burkina Faso comme l'indiquait
Kabré (1994).
Selon Paugy et Levêque (1999), les espèces de
poissons se nourrissent des proies qu'elles trouvent dans le milieu où
elles vivent et le régime alimentaire peut changer d'un endroit à
un autre ou d'une saison à l'autre. Toujours selon ces mêmes
auteurs, il y a une relation entre la forme de la tête et le
régime alimentaire. Donc, la forme aplatie de la tête de ce
poisson (annexe 4) lui permet de racler les fonds de rivière donc de
consommer en majorité les Chironomidae pendant les
périodes d'étiage et les périodes de hautes eaux.
La constante b (3,015#177; 0,097) de la relation longueur-poids
confirme les travaux de Sanon (1995) qui avait trouvé 3,09 pour ces
mêmes poissons. Ce résultat implique chez ce poisson, une
croissance isométrique c'est-à-dire qu'il conserve à peu
près son embonpoint au cours de sa vie.
CONCLUSION
L'évaluation de la qualité biologique des
écosystèmes liés à ces deux cours d'eau
périurbains révèle une population de
macroinvertébrés dominée par les Insectes (90,70% sur le
Kou et 92,94% sur le Wé), avec une forte concentration de larves de
Chironomidae (23,69% sur le Kou et de 50,37% sur le Wé) au
niveau du benthos. La rivière Kou dont le trajet évite
l'agglomération de Bobo Dioulasso est caractérisée par une
diversité de macroinvertébrés plus importante que le
marigot Wé qui traverse la ville. En effet, ce marigot est un
dépotoir d'ordures ménagées et d'autres types de produits
dangereux pour l'écosystème aquatique. Ces sources de pollution
d'origine anthropique sur le Wé se retrouvent en certains endroits au
niveau du Kou.
Une meilleure qualité de l'eau induit une
diversification des macroinvertébrés, par conséquent une
abondance de proies pour les prédateurs aquatiques comme C.
gariepinus qui se nourrit en majorité d'insectes.
Ces deux cours d'eau étant rattachés au fleuve
Mouhoun, cours d'eau plus important, leur suivi s'avère
nécessaire afin de déceler toute perturbation écologique
qui aura des conséquences sur l'écosystème de ce
réseau hydrographique. Ce suivi devrait prendre en compte la
sensibilisation des populations riveraines, principales
bénéficiaires de la santé écologique des cours
d'eau.
Les différentes familles inventoriées devraient
constituer le point de départ de toute action de suivi sur ces deux
cours d'eau. En effet l'utilisation des macroinvertébrés
benthiques comme outil permettant la caractérisation de la
qualité physicochimique des cours d'eau, nécessite une
étude plus poussée sur les traits biologiques relatifs à
certaines espèces dont la présence ou l'absence laisse
présager une pollution ou une bonne santé écologique.
BIBLIOGRAPHIE
Baijot, E., Moreau, J. et Bouda S., 1994.
Aspects hydrobiologiques et piscicoles des retenues d'eau en zone
soudano-sahélienne. Cas du Burkina Faso. CTA, 349p
Barbour, M. T., J. Gerritsen, B. D. Snyder et J. B.
Stribling, 1999. Rapid Bioassessment protocols for use in Wadeable
Rivers: Peryphyton, Benthic Macroinvertebrates, and Fish, 2e édition,
Washington, D.C., U.S. Environmental Protection Agency, Office of Water,
EPA841- B.99-002, 11 chapters, 4 annexes.
Coffman, W. P. et Ferrington L. C.
Jr, 1984. Chironomidae. In: «An introduction to the aquatic
insects of North America», Merrit R. W., Cummins K.W., Kendall, Hunt
Publishing Company. Second edition, Dubuque, Iowa, USA, p. 551-652.
Corbet, P.S., 1958. Effects of Simulium control
on insectivorious fisches. Nature lond. 181, pp 570.571
Corbet, P.S., 1961. The Biological significance
of the attachment of immature stages of Simulium to mayflies and crabs.
Bulletin of Entomological Research, 52: 695-699 Cambridge University press
Dakouré, J. Derouane et B. Tychon, 2009.
Elaboration du bilan d'eau du bassin versant du Kou et analyse critique des
principaux composants.
Dejoux, C., Lauzanne L. et Levêque C.,
1971. Nature des fonds et répartition des organismes benthiques
dans la région du Bol (Archipel Est du lac Tchad). Cahier ORSTROM,
série Hydrobiologie, 3-4 :213-223.
Dejoux C., 1977. Chironomes du lac Bam (Haute
Volta). Cah. ORSTOM, sér..Hydrobiol.., 11,4 :291-295.
Dejoux, C., J.M. Elouard, P.Forge et J.L. Maslin,
1983. Catalogue iconographique des insectes aquatiques de Côte
d'Ivoire. ORSTROM, OMS, OCP, VCU, Hybio.
Diomande, D., Y. K. Bony, E. O. Edia, K. F. Konan et G.
Gourene, 2009. Diversité des macroinvertébrés de
la Rivière Agnéby (Côte d'Yvoire ; Afrique de l'Ouest).
European Journal of Scientific Research. ISSN 1450-216X Vol 35 No. 3 (2009),
pp. 368-377
Fulton, T.W., 1911. The sovereignty of the sea.
Edinburgh and London
Guenda, W., Thèse. 1996. Etude
faunistique, écologique et de la distribution des insectes d'un reseau
hydrographique de l'ouest africain : le Mouhoun (Burkina Faso) ; Rapport avec
Simulium damnosum Théobald, vecteur de l'onchocercose
Guenda, W., 1997. Nouvelles espèces du
genre Catoxyethira Ulmer du Burkina Faso,
(Trichoptera, Hydroptilidae).
Bulletin de la société entomo. De France, 102 : 217-224.
Guenda, W., 1997. Premières
données sur le genre Psychomyiellodes Mosely (Trichoptera : Echonidae)
et description d'une nouvelle espèce du Burkina Faso (Afrique
Occidentale). Ann. Limnol., 36 : 163-167
Hellawell, J. M., 1986. Biologicol Indicators of
Freshwater Pollution Environmental Hureau J.C., 1970. Biologie
comparée de quelques poissons antarctiques (Nototheniidae). Bull. Inst.
Océanogr. Monaco, 58 (1393), 244 p.
Hyslop, E.J. 1980. Stomac content analysis. a
review of methods and their application. J. Fish. Biol., 17, 411-429
Kabré T. A. 1994. Impact de la saison sur
le facteur de condition et la croissance des juveniles de tilapia au barrage de
Loumbila. Revue Sc. Tech. 21 (2), CNRST, Burkina Faso.
Kabré T. A., Illé A. et Guenda W.,
2000. Inventaire et étude de la densité de
distribution des insectes du benthos des deux lacs de barrage de Bagré.
Science et Technique, Vol. 24 (2). Kabré T. A., Diguingué
D et Bouda S., 2002. Effet du
rétrécissement de la superficie d'eau sur les
macroinvertébrés du lac de barrage de la Comoé, Sud-Ouest
du Burkina Faso. Science et Technique, série Sciences Naturelles et
Agronomie, vol. 26(1).
Lévêque, C., J. R. Durand, 1981.
Flore et Faune aquatiques de l'Afrique Sahelo- Soudanienne, Tome II. Editeurs
scientifiques hydrobiologiques, O.R.S.T.O.M
Merrit, R. W., K. W. Cummins, 1984. An
introduction to the aquatic insects of north America, second édition,
Dubuque (Iowa), Kendall/Hunt Publishing Company, 722 p Moisan, J.,
2006. Guide d'identification des principaux
macroinvertébrés benthiques d'eau douce du Québec,
Surveillance volontaire des cours d'eau peu profonds, Direction du suivi de
l'état de l'environnement, ministère du développement
durable, de l'environnement et des parcs, ISBN-13 :978-2-550-48518-6(PDF),
ISBN-10 : 2-550-48518-1 (PDF)-82 p
Moisan, J. et L. Pelletier, 2008. Guide de
surveillance biologique basée sur les macroinvertébrés
benthiques d'eau douce du Québec - Cours d'eau peu profonds à
substrat grossier, 2008. Direction du suivi de l'état de
l'environnement, ministère du Développement durable, de
l'Environnement et des parcs, ISBN : 978-2-550-53591-1 (version
imprimée),
86 p. (incluant 6 ann.)
NEPAD., 2005. Plan d'action du NEPAD pour le
développement des pêcheries et de l'Aquaculture en Afrique. NEPAD
- Sommet Du Poisson pour tous. Abuja, Nigeria. 32 p. Oueda,
A., W. Guenda, A.T. Kabré, F. Zongo et G.B. Kabré, 2007.
Diversity, abundance and seasonal dynamic of zooplankton community in a
south-saharan reservoir (Burkina Faso). J. Boil., 7:1-9.
Paugy, D. et C. Levêque, 1999.
Régime alimentaire et réseaux trophiques. In : « Les
poissons des eaux continentales africaines ; diversité, écologie,
utilisation par l'homme », Paugy D. et Levêque C., Institut de
Recherche pour le Développement, Paris, France. p. 157- 190
Roger, G. B., H. E. Jaques, 1978. How to know
the insects. Thirst Edition, 409p
Sanon, Z.L., 1995.
Inventaire et dynamique de quelques espèces de poissons dans la
réserve de la biosphère de la mare aux hippopotames et dans le
lac de la vallée du Kou. Mémoire de fin d'étude.
Spare, P., S.C. Venema, 1996. Introduction
à l'évolution des stocks de poissons tropicaux. Première
partie : Manuel
Sib, S., T. Paré, et H. Belo, 2003. Etude
de la dégradation des berges et de l'occupation des terres dans le
bassin versant du Kou. Rapport, 31p.
WFD (Water Framework Directive), 2003. Overall
Approach to the Classification of Ecological Status and Ecological Potential,
Water Framework Directive Common Implementation Strategy Working Group 2, A
Ecological Status (ECOSTAT),
28p. et 2 annexes
Wellens, J., F. Traoré, M.-J. Niamian, H.
Karambiri, M. Diallo, N.F. Compaoré, D.
2009. Renforcement structurel de la
capacité de gestion des ressources en eau pour l'agriculture dans le
bassin du Kou. Rapport technique 2007-2008
WEBOGRAPHIE
http://www.cnrs.fr/cw/dossiers/doseau/decouv/ecsysMenu.html
le 10-08-2010*
http://www.eurojournals.com/ejsr.htm
le 15-08-2010*
http://www.rpn.ch/.../cle_de_determination_desinvertebres_d_eau_douce.pdf
le 15-08-2010*
http://insects.about.com/od/onlineidentificationkeys/Online_keys_for_identifying_insects.htm
le 15-08-2010*
http://www.ecifm.rdg.ac.uk/pesticides.htm
le 29-08-2010*
http://www.laease.com/conductivite.html
le 29-08-2010*
http://cwss.www.de/workshop/wfd/Ecological-Classification-Guidance.pdf
le 11-09-2010*
http://www.epa.gov/owow/monitoring/rbp/wp61pdf/rbp.pdf
le 23-09-2010*
http://www.reynier.com/ciaf/Pays-mono/Burkina/Geographie.html
le 24-09-2010*
http://www.ambaburkina.dk/reliefethydrologie.html
le 24-09-2010*
http://fr.wikipedia.org/wiki/chironomidae
le 29-09-2010*
* date de dernière consultation
ANNEXES
Annexe 1 : pluviométrie (en mm) pendant
l'échantillonnage
|
Mois
dates
|
février
|
Mars
|
Avril
|
Mai
|
Juin
|
juillet
|
|
1
|
|
|
|
30.9
|
4.1
|
|
|
2
|
|
|
|
0.9
|
|
|
|
3
|
|
|
|
|
12.6
|
|
|
4
|
|
|
|
|
|
|
|
5
|
|
|
|
1.8
|
|
16.8
|
|
6
|
|
|
|
|
|
|
|
7
|
|
|
|
28.8
|
|
2.8
|
|
8
|
|
|
|
|
|
9.5
|
|
9
|
|
|
|
|
0.2
|
0.6
|
|
10
|
|
|
20.4
|
22.6
|
34.4
|
2.6
|
|
11
|
|
|
|
3.3
|
|
12.3
|
|
12
|
|
|
|
|
14.4
|
|
|
13
|
1.1
|
|
2.3
|
|
|
|
|
14
|
|
|
|
|
0.9
|
|
|
15
|
|
|
|
|
|
|
|
16
|
|
|
|
|
1.8
|
20.6
|
|
17
|
|
|
|
4.2
|
0.1
|
|
|
18
|
|
|
|
|
|
|
|
19
|
|
|
8.8
|
3.5
|
|
|
|
20
|
|
|
|
|
|
|
|
21
|
|
|
|
30.8
|
1.1
|
20.7
|
|
22
|
|
|
|
|
|
15.0
|
|
23
|
|
|
|
|
|
13
|
|
24
|
|
|
|
25.9
|
8.6
|
0.2
|
|
25
|
|
|
|
|
|
|
|
26
|
|
|
3.4
|
|
|
|
|
27
|
|
|
|
|
|
|
|
28
|
|
|
|
0.2
|
|
|
|
29
|
|
|
23.5
|
11
|
|
2
|
|
30
|
|
|
0.4
|
|
0.2
|
4.7
|
|
31
|
|
|
|
9.3
|
|
4.8
|
|
total
|
1.1
|
0
|
58.9
|
173.2
|
78.4
|
124.6
|
Source : Centre Météo Secondaire de
Bobo-Dioulasso
Annexe 2: Principaux types de pollution des eaux
continentales, d'après C. Lévêque.
Écosystèmes aquatiques (Hachette, 1996)
|
Type de pollution
|
Nature
|
Source
|
|
Physique
|
Pollution thermique
|
Rejets d'eau chaude
|
Centrales thermiques
|
|
Pollution radioactive
|
Radio isotopes
|
Installations nucléaires
|
|
Matières organiques
|
Glucides, lipides, protides
|
Effluents domestiques agricoles
|
|
Ammoniac, nitrates
|
Elevage et piscicultures
|
|
Chimiques
|
fertilisants
|
Nitrates, phosphates
|
Agricultures, lessives
|
|
Métaux et métalloïdes
|
Mercure, cadnium, plomb, aluminium, arsenic
|
Industries, agricultures, pluies acides, combustion
|
|
Pesticides
|
Insecticides, herbicides, fongicides
|
Agricultures, industries
|
|
Organochlorés
|
PCB, solvants
|
Industries
|
|
Composés organiques de synthèses
|
Nombreuses molécules
|
industries
|
|
détersifs
|
Agents tensio-actifs
|
Effluents domestiques
|
|
Hydrocarbures
|
Pétrole et dérivés
|
Industries pétroliers, transports
|
|
Microbiologique
|
Bactéries, virus, champignons
|
Effluents urbains et élevage
|
Source :
http://www.cnrs.fr/cw/dossiers/doseau/decouv/ecsysMenu.html
le 10-08-2010
Annexe 3 : Evolution de la densité benthique sur
le Kou et le Wé
|
Rivières
|
Densités du benthos (nombres d'individus au
m2)
|
|
Etiage
|
Début de saison
|
Hautes eaux
|
|
Kou
|
250
|
183.33
|
500
|
|
wé
|
183.33
|
366.66
|
16.66
|
Annexe 4: Clarias gariepinus

Clarias gariepinus (Burchell, 1822). Photo :
Wikipédia 20-12-10

C. gariepinus capturés au niveau du Kou (Photo :
Sanogo Souleymane, juillet 2010)
Règne : Animalia Embranchement
:Chordata
Classe: ActinopterygiiOrdre:
Siluriformes
Famille: Clariidae
Genre: Clarias
Espèce: C. gariepinus
Nom binomial : Clarias gariepinus (Burchell, 1822)
Annexe 5 : Tailles, poids et contenus stomacaux des
poissons ayant l'estomac non vide
|
Taille
(cm)
|
Poids
(g)
|
Poids bol
alimentaire
(g)
|
Contenus stomacaux
|
|
Arthropodes
|
Mollusques
|
Autres
|
|
23.4
|
95.1
|
3.1
|
Odonates
* 1 Libellulidae
* 1 Calopterygidae
* 2 ni
Coléoptères
* 3 Dytiscidae * 1 Carabidae
Trichoptères
* 6 Hydropsychidae
Hyménoptères
* 1 Formicidae
Plécoptères
* 1 mue
|
|
* 1 Poisson ni
* 3 batraciens (grenouilles)
|
|
25
|
125.6
|
3.2
|
Hétéroptères
* 1 ni Coléoptères
* 4 Dytiscidae
* 1 ni Hyménoptères
* 4 Formicidae Odonates
* 2 ni Orthoptères
* 1 Tettigoniidae
|
|
* 5 poissons ni
|
|
38
|
362.4
|
1.3
|
|
|
Terre
|
|
41.3
|
402.6
|
3.1
|
Odonates
* 3 Libellulidae
|
|
Débris de plante
|
|
25.2
|
151.9
|
0.5
|
Coléoptères
* 3 Dytiscidae Odonates
* 1 Calopterygidae
Hétéroptères
* 1 Notonectidae
|
|
|
|
31
|
210.7
|
0.3
|
Odonates
* 1 Calopterygidae
Hétéroptères
* 1 Notonectidae
|
|
|
|
27
|
140.2
|
1
|
Odonates
* 1 Libellulidae
Coléoptères
*1 Dytiscidae Diptères
* 2 Chironomidae
|
|
|
|
43.1
|
510.8
|
3.6
|
Hétéroptères
* 2 Belostomatidae
|
|
Débris de plante
|
|
|
|
Coléoptères
* 1Dytiscidae
|
|
|
|
31
|
200.3
|
1.1
|
|
|
Débris de plante
|
|
36
|
365.1
|
0.4
|
Hétéroptères
* 1 Belostomatidae
Coléoptères
* 1 Carabidae
|
|
|
|
20
|
61.1
|
0.6
|
Odonates
* 1 ni (la) Coléoptères
* 1 Dytiscidae
|
|
Débris de plantes
|
|
15.4
|
26.1
|
0.1
|
Hétéroptères
* 1 Belostomatidae
|
|
|
|
22
|
73.1
|
0.4
|
Coléoptères
* 1 ni
Hétéroptères
* 1 Belostomatidae
|
|
|
|
19.6
|
56.8
|
0.9
|
Diptères
* 1 Simuliidae
|
|
|
|
24
|
86.9
|
0.8
|
Odonates * 1 ni
|
2 Physidae, 2 Sphaeriidae
|
|
|
26
|
103.2
|
0.9
|
Hétéroptères
* 1 ni Coléoptères
* 1 ni
|
|
Débris de plantes
|
|
16.1
|
31
|
0.5
|
Insecte (ni)
|
|
|
|
21.3
|
78.6
|
0.4
|
Insectes (ni)
|
|
|
|
24
|
98.4
|
0.8
|
Insecte (ni)
|
|
Grains de riz
|
|
18.5
|
47.8
|
0.2
|
|
|
Débris de plantes
|
|
22.4
|
84.3
|
0.3
|
Coléoptères
* 1 Dytiscidae * 1 ni
|
|
|
|
39
|
230.2
|
0.9
|
Coléoptères
* 1 Dytiscidae
|
|
|
|
30
|
149.8
|
0.8
|
Hétéroptère * 1 ni
|
|
|
|
23.7
|
93.2
|
1.2
|
|
|
Poisson (ni)
|
|
23.4
|
96
|
0.2
|
Insecte (ni)
|
|
|
|
19.4
|
74.1
|
0.9
|
|
5 Physidae
2 Planorbidae
|
Débris de plante
|
|
24
|
98.4
|
1.2
|
Odonates
* 1 Libellulidae (ad)
|
|
Débris de plante
|
|
28
|
150.2
|
0.8
|
Coléoptères
* 1 Dytiscidae Odonates
* 1 Gomphidae (ad)
|
|
|
|
23.5
|
99.5
|
0.7
|
Coléoptères
* 1 Dytiscidae (ad) Insecte (ni)
|
|
|
|
18.5
|
44.5
|
0.4
|
Hétéroptères
* 1 Belostomatidae
|
|
|
|
22.9
|
80.3
|
0.3
|
|
|
2 grains de riz Débris de plante
|
|
15.5
|
27.8
|
1.3
|
|
|
Poissons (ni)
|
|
18.6
|
55.8
|
0.8
|
Hétéroptères * 1 ni
|
|
Débris de plante
|
|
20
|
58.3
|
0.4
|
|
|
Débris de plante
|
|
19
|
57.1
|
1
|
Coléoptères
* 1 Hydrophilidae Diptères
* 1 Stratiomyidae (la)
|
|
Débris de plante
|
|
20
|
53.2
|
0.8
|
Hyménoptères
* 3 Formicidae (ad)
|
|
|
|
19.8
|
49.6
|
0.8
|
Insectes (ni)
|
|
Débris de plante
|
|
27
|
131.4
|
0.3
|
Insectes (ni)
|
|
|
|
20.7
|
71
|
1.1
|
|
2 Physidae
|
Débris de plante
|
|
19.6
|
54.7
|
0.3
|
Coléoptère * 1 ni
|
|
|
|
20.5
|
61.2
|
0.6
|
Insectes (ni)
|
|
|
|
22.7
|
80.4
|
0.8
|
Odonates
* 1 Calopterygidae (la)
|
|
|
|
23.5
|
110.1
|
1.3
|
|
|
Poisson (ni)
|
|
21.6
|
87.4
|
1.1
|
|
|
Poisson (ni)
|
|
22
|
79.6
|
0.9
|
Odonates
* 1 Lubelliludae
|
1 Planorbidae
1 Physidae
2 Lymnaeidae
|
|
|
18.7
|
49.7
|
0.3
|
Coléoptères * 1 ni
|
|
|
|
20.1
|
56.3
|
1.3
|
Hyménoptères
* 1
Formicidae
|
|
Poisson (ni)
|
|
17.9
|
45.1
|
0.7
|
|
|
Débris de plante
|
|
21
|
56.8
|
0.8
|
Coléoptères * 1 ni
|
|
Débris de plante
|
|
18.7
|
50.2
|
0.3
|
Insecte (ni)
|
|
|
|
18.6
|
49.7
|
0.9
|
Coléoptères
* 1 Carabidae
|
|
Débris de plante
|
* ni : non identifié
Annexe 6 : fiche de sortie
FICHE DE SORTIE
Mois de :

Date
Heure d'arrivée
Heure de départ
Profondeur de l'eau
Conductivité de l'eau
Transparence de l'eau
Station
pH
Koumi
Nasso
Bama
Bobo
Vitesse d'écoulement de l'eau :
Distance : 5 m

Koumi
Nasso
Bama
Bobo
No expérience
Station
Vitesse (m/s)
Vitesse moyenne (m/s)
Vitesse (m/s)
Vitesse moyenne (m/s)
Vitesse (m/s)
Vitesse moyenne (m/s)
Vitesse (m/s)
Vitesse moyenne (m/s)
Temps (s)
Temps (s)
Temps (s)
Temps (s)
1
2
3
4
5
Annexe 7 : total des poissons capturés à la station
de Bama (taille en cm ; poids en g)
|
taille
|
poids
|
taille
|
poids
|
Taille
|
poids
|
taille
|
poids
|
|
28.5
|
172.7
|
21
|
69.6
|
19
|
49.2
|
16.9
|
37.8
|
|
21.5
|
74.2
|
20
|
66.5
|
17.8
|
42.5
|
19.6
|
55.9
|
|
19.4
|
74.1
|
21.6
|
82.4
|
16.5
|
31.4
|
14.3
|
22.8
|
|
27
|
131.4
|
18.5
|
47.8
|
13.6
|
20.8
|
18
|
45.3
|
|
22.3
|
77.5
|
19.5
|
51.7
|
20
|
58.1
|
18.5
|
44.3
|
|
20.5
|
66.9
|
21.8
|
75
|
18.7
|
51.2
|
12.9
|
15.6
|
|
18.5
|
44.7
|
21.4
|
65.1
|
24
|
106.4
|
14.2
|
24.9
|
|
20.6
|
60
|
22
|
79.6
|
19.8
|
48.9
|
19.4
|
68.4
|
|
22
|
75.8
|
22.1
|
75.9
|
19
|
47.1
|
19
|
45.2
|
|
20
|
53.2
|
19.6
|
55.2
|
20
|
51.9
|
14.5
|
22
|
|
24
|
109.2
|
25.5
|
124.6
|
19
|
54
|
17.4
|
36.1
|
|
23.5
|
99.5
|
21
|
72.7
|
17.5
|
41.8
|
15.5
|
25.8
|
|
28
|
150.2
|
21.2
|
62.2
|
20
|
57.7
|
23.4
|
95.1
|
|
16.8
|
32.2
|
18
|
44.5
|
22
|
77.5
|
25
|
125.6
|
|
18.8
|
55.8
|
18.8
|
53.9
|
22.7
|
80.4
|
38
|
362.4
|
|
29
|
176
|
17
|
35.1
|
22.5
|
85.9
|
41.3
|
402.6
|
|
20
|
58.3
|
20.5
|
63.8
|
19.8
|
49.6
|
40
|
403.2
|
|
21
|
74.8
|
23.5
|
104.5
|
24
|
97.2
|
31.6
|
220
|
|
19
|
57.1
|
21.6
|
87.4
|
21.5
|
65.5
|
30
|
196.4
|
|
18.7
|
50.2
|
19.2
|
62.5
|
16
|
33.3
|
25.2
|
151.9
|
|
25
|
113.1
|
18
|
45
|
18
|
49.5
|
31
|
210.7
|
|
19.6
|
54.7
|
22.5
|
78
|
21
|
82.1
|
27
|
140.2
|
|
19.5
|
60.4
|
18.5
|
51.7
|
20
|
56.1
|
43.1
|
510.8
|
|
18.1
|
42.8
|
18.6
|
49.7
|
18.5
|
46.1
|
31
|
200.3
|
|
19
|
50.3
|
26
|
126.8
|
17
|
39.2
|
36
|
365.1
|
|
15.4
|
26.1
|
17.9
|
45.1
|
19
|
51.8
|
39.4
|
363.3
|
|
23.5
|
96
|
20.7
|
71
|
19.5
|
57.2
|
32
|
189.9
|
|
24
|
86.9
|
20.1
|
56.3
|
15
|
27.7
|
30.7
|
149.8
|
|
20.7
|
67
|
19.2
|
60.7
|
22
|
73.1
|
39.3
|
230.2
|
|
19.6
|
56.8
|
15.9
|
26.5
|
16
|
28.5
|
31
|
173.4
|
|
22.2
|
89.5
|
24
|
101
|
21.8
|
79
|
26.5
|
103.2
|
|
19.4
|
54.9
|
15.5
|
27.8
|
20
|
53
|
|
|
|
24.3
|
116.2
|
19.6
|
68.9
|
18.5
|
50.6
|
|
|
|
16.1
|
31
|
19
|
46.5
|
22.5
|
77.1
|
|
|
|
21.3
|
78.6
|
26
|
127.3
|
16
|
28.6
|
|
|
|
24
|
110.2
|
20
|
59.5
|
17.1
|
39.3
|
|
|
|
20
|
62.1
|
14.6
|
22.1
|
20.6
|
57.8
|
|
|
|
24.2
|
112.1
|
14.5
|
24.5
|
21
|
56.8
|
|
|
|
23.7
|
93.2
|
19.6
|
50.5
|
21.5
|
55
|
|
|
|
24
|
98.4
|
23.8
|
95
|
18
|
51.4
|
|
|
|
24
|
98.7
|
23.5
|
101.1
|
23.5
|
103.5
|
|
|
|
23.5
|
109.3
|
22.9
|
80.3
|
19.8
|
50
|
|
|
|
20.2
|
57.3
|
20.5
|
61.2
|
18.5
|
47.5
|
|
|
|
18.2
|
47.6
|
18.5
|
52.8
|
20.2
|
69.3
|
|
|
|
18.6
|
53.1
|
24
|
91.3
|
17
|
47.6
|
|
|
|
25.3
|
119.1
|
21.5
|
71.8
|
23.5
|
85.8
|
|
|
|
19.8
|
56.8
|
21
|
77.7
|
18.7
|
49.7
|
|
|
|
21.7
|
79.6
|
22
|
81.4
|
16
|
34
|
|
|
|
22.4
|
84.3
|
14.2
|
19.1
|
16.8
|
33.7
|
|
|
|
14.5
|
24
|
15.6
|
31.6
|
15.5
|
29.2
|
|
|
| 

