|
REPUBLIQUE DE CÔTE D'IVOIRE Union -
Discipline - Travail
|
Ministère de l'Enseignement Supérieur et de la
Recherche Scientifique
|
Université Nangui Abrogoua
|
|
|
UFR des Sciences de la Nature
Année Académique 2012-2013
DIPLÔME D'ETUDES APPROFONDIES EN GESTION ET
VALORISATION DES RESSOURCES NATURELLES
Option : BIODIVERSITE ET GESTION DURABLE DES ECOSYSTEMES
|
Thème :
EFFETS DE L'INOCULATION DES VERS DE TERRE SUR LA
PRODUCTION DU MAÏS : EXPERIMENTATIONS EN MILIEUX PAYSANS
|
Présenté par
GOLLI Siagbé
(Maître es Sciences)
Soutenu le 22 Janvier 2013 devant le jury composé de
:
Président : Pr N'DA Konan, Maître de
conférences
Directeur scientifique : Pr TONDOH Ebagnerin
Jérôme, Maître de conférences
Rapporteur : Pr DAGNOGO Mamadou, Professeur titulaire
Examinateur: Dr TIHO Seydou, Maître Assistant
I
TABLE DES MARTIERES
DEDICACE III
REMERCIEMENTS IV
RESUME V
ASBTRACT VI
INTRODUCTION 1
I. REVUE BIBLIOGRAPHIQUE 3
1. Ecologie de Millsonia omodeoi et de
Hyperiodrilus africanus 3
1.1 Millsonia omodeoi 3
1.2 Hyperiodrilus africanus 3
2. Notions de groupe fonctionnel et de type écologique de
vers de terre 4
3. Vers de terre et agriculture 5
II. MILIEU D'ETUDE 7
1. Localisation 7
2. Climat 10
2.1 Pluviométrie 10
2.2 Température 10
3. Sols et végétation 9
4. peuplement de vers de terre 12
III. MATERIELS ET METHODES 14
1.Choix des espèces animales et végétale
14
2. Dispositif expérimental 13
3. Culture de maïs et inoculation des vers de terre 14
31 Culture de maïs 15
3.2 Inoculation des vers de terre 16
4. Mesures effectuées à la fin de
l'expérimentation 18
4.1 Au niveau du maïs 16
4.2 Au niveau des vers de terre 16
5. Perception des paysans des effets des vers de terre sur la
fertilité du sol et sur la
production agricole 18
6. Analyses statistiques 19
IV. RESULTATS 20
1. Maïs 22
II
1 1 Croissance du maïs 23
1 2 Production du maïs 23
1 3 Efficience de l'utilisation de l'eau 23
1 4 Paramètres de production du maïs sensibles
aux différents traitements 23
2. Impact de l'inoculation sur le peuplement de vers de terre
26
3 Perception des paysans des effets des vers de terre sur la
fertilité du sol et sur la
production des cultures 30
3.1 Connaissance générale des paysans sur les
vers de terre 30
3.2 Observations des paysans propriétaires de
parcelles sur les parcelles expérimentales
30
3.3 Perception des paysans des effets des vers de terre sur
la fertilité du sol 31
3.4 Perception des paysans des effets des vers de terre sur
le rendement des cultures 31
3.5 Avis des paysans sur l'utilisation des vers de terre
comme biofertilisants 31
V DISCUSSION 33
1. Impact des vers de terre sur la croissance et la production
d'une culture de maïs 33
2. Impacts de l'inoculation des populations de M. omodeoi
et H. africanus sur les
communautés de vers de terre 34
3. Perception des paysans des effets ddes vers de terre sur la
fertilité du sol et la production
agricole 35
CONCLUSION 36
REFERENCES BIBLIOGRAPHIQUES 37
ANNEXES 47
DEDICACE
Ce travail est dédié à
Monsieur et Madame Houra Delatour,
Mon oncle Ziboh,
Mes frères et soeurs,
III
Tous mes parents et amis.
IV
REMERCIEMENTS
Je remercie profondément le Professeur TONDOH Ebagnerin
Jérôme, Maître de Conférences à
l'Université Nangui Abrogoua, Chercheur au Centre de Recherche en
Ecologie et Coordonnateur National du projet "Conservation and Sustainable
Management of Below-Ground Biodiversity" « CSM-BGBD », pour m'avoir
initié à l'étude des vers de terre et pour la confiance
qu'il m'a accordé au long de ces deux années de travail.
Je suis également reconnaissant au Professeur Tano Yao,
Directeur dudit projet, pour sa disponibilité et sa gentillesse.
Un grand merci au Docteur Koné Armand, Assistant
à l'Université Nangui Abrogoua, pour son assistance dans la
rédaction de ce mémoire.
Je voudrais aussi remercier Docteur Tahoux Martine et tout le
personnel du CRE pour leur soutien.
J'adresse un remerciement particulier à Guéi
Arnauth Martinez, Doctorant à l'Université Nangui Abrogoua, pour
son aide et sa rigueur sur le terrain, l'analyse des données et la
rédaction de ce document.
Je tiens également à remercier les amis que sont
Gbapa Ange Edgar et Baidai Yannick avec qui j'ai effectué les quatre
mois de travaux de terrain. Que le ciel vous accorde sa protection sans faille
afin que nous puissions goutter ensemble au fruit de ces efforts fournis !
Les travaux de terrain requièrent toujours la
participation importante des personnes. Je voudrais ainsi remercier Kablan
Aurore, Marie Noëlle, Charly, Pierre, Eric, Olivier, Blaise, Gohan-Ni et
tous les habitants de Goulikao. Je n'oublie pas tous mes amis du CRE, en
l'occurrence, Assi Jules, Kassi Yéboua et Bosso Marcel.
Enfin, un merci particulièrement chaleureux à
mes parents, à toute ma famille et à mes amis Ouattara
Fétigué, Kouadio Julien, Dangbé Fréjus, Wondji
Pamphile et Kouamé Alexis sans lesquels ce travail n'aurait pu avoir
lieu.
Résumé
L'effet de l'introduction de deux espèces de vers de
terre de groupes fonctionnels à effet contrastés, Millsonia
omodeoi (ver compactant) et Hyperiodrilus africanus (ver
décompactant) sur la croissance, la production du maïs (Zea
mays) a été étudié en milieu paysan, dans la
région du Centre-Ouest de la Cote d'Ivoire (Goulikao, Oumé).
Cette étude qui a été réalisée avec la
participation des paysans dont les jachères ont été
utilisées comme parcelles expérimentales, comprend au total cinq
traitements : (i) maïs + M. omodeoi (M+Mo), (ii) maïs +
H. africanus (M+Ha), (iii) maïs + M. omodeoi+ H.
africanus (M+Mo+Ha), (iv) maïs + urée + superphosphate triple
(M+U+SPP), et (v) le témoin (M) sans ajouts de vers de terre ni
d'engrais. Dans chaque parcelle, une quantité de vers de terre 90 g a
été inoculée par mètre carré, en trois
fractions : 25 g les 15è et 30è jour
après semi et 40 g le 45ème jour après semis.
La perception des paysans de l'impact des vers de terre sur la fertilité
du sol et la production du maïs ont été également
évalués à partir d'un échantillon de 95 paysans.
Les résultats ont montré que seules les parcelles
caractérisées par l'introduction des vers de terre et
l'application d'engrais inorganiques ont montré une amélioration
significative de l'utilisation efficiente de l'eau et de la production du
maïs. Le poids des grains de maïs a augmenté de 21,5%, 21,2%,
19,4% et 13,9%, respectivement, dans les traitements M+Mo, M+Mo+Ha, M+U+SPP et
M+Ha. L'amélioration de l'utilisation efficiente de l'eau et
l'augmentation de la production du maïs seraient essentiellement dues
à l'association des deux espèces de vers de terre à effet
contrastés. Par ailleurs, l'enquête auprès des paysans a
révélé qu'une proportion de 49,3% pense que les vers de
terre ont un effet positif sur la fertilité tandis que 23% pensent le
contraire. Quant aux rendements des cultures, 57% de ces paysans croient qu'ils
sont améliorés par ces organismes alors que 31% disent le
contraire. En conséquence, 80% des paysans se disent favorables à
l'utilisation des vers de terre pour améliorer la production agricole.
Cependant, le coût financier d'une telle technologie serait une
contrainte majeure.
V
Mots clés: vers de terre, groupe
fonctionnels, agriculture durable, culture de maïs.
Abstract
The impact of two earthworms species with contrasting impact
on soil functioning, Millsonia omodeoi (compacting) and
Hyperiodrilus africanus (decompacting), on maize (Zea mays)
growth and production were investigated in field conditions in the Centre-West
region of Ivory Coast (Goulikao, Oumé). The treatments included: (i)
maize + M. omodeoi (M+Mo), (ii) maize + H. africanus (M+Ha),
(iii) maize + M. omodeoi+ H. africanus (M+Mo+Ha), (iv) maize
+ urea + super phosphate triple (M+U+SPP), and (v) the control (M). A total of
90 g of worms were inoculated per meter square at tree times: 25 g at 15 and 30
days after planting and 40 g, 45 days after planting. The perception of farmers
about the effect of earthworms on soil fertility and crop yields was assessed
through a survey of 95 farmers from the study site. Results maize production
and water use efficiency was increased in inorganic fertilizer and worm-based
treatments. Grain yields were enhanced by 21.5%, 21.2%, 19.4% and 13.9% in
M+Mo, M+Mo+Ha, M+U+SPP and M+Ha treatments, respectively. The inoculation of
earthworms along with the application of inorganic fertilizer have
significantly enhanced cobs production and improve the water use efficiency of
maize crops, which is likely due to the association of the two earthworm
species. A survey undertaken among farmers in the study site showed that 49.3%
farmers believed that earthworms positively influence soil fertility whereas
23% think the contrary. Regarding agricultural outputs, 57% of farmers think
that earthworms increased crops yields contrarily to 31% of the farmers. As a
result, 80% of farmers are willing to use earthworms for improving agricultural
production. However, the economic cost of such technology is likely to be a
bottleneck.
VI
Keywords: earthworms, functional groups, sustainable agriculture,
maize crop
1
INTRODUCTION
L'agriculture itinérante sur brûlis, la
principale pratique agricole dans les régions tropicales,
génère de faibles rendements à l'hectare. Elle favorise
une perte importante d'éléments nutritifs et par
conséquent, n'est pas considérée comme une pratique
agricole durable (Brown et al., 1994). L'agriculture intensive est
caractérisée par la pratique de la monoculture
(variétés améliorées), l'utilisation de pesticides
et d'engins lourds. Ce type d'agriculture a contribué à faire
reculer la famine dans le monde, mais elle est également
considérée comme une forme de pratique agricole non durable
(relativement instable selon les principes écologiques). Elle est
à l'origine d'une grande perte de nutriments susceptibles de polluer les
eaux, une pullulation de mauvaises herbes et une invasion de maladies et de
pestes des plantes (Tilman, 1999).
Dans un contexte d'agriculture durable, le maintien de la
fertilité du sol et partant l'amélioration de la production
agricole par la manipulation des organismes dits ingénieurs du sol
constitue une voie à explorer (Lavelle et al., 1989; Lavelle et
al., 1997). Les vers de terre agissent sur la production
végétale qu'ils améliorent par l'intermédiaire de
cinq processus (Scheu, 2003; Brown et al., 2004): (i) l'augmentation
de la vitesse de minéralisation de la matière organique rendant
ainsi les nutriments disponibles à la plante, (ii) le contrôle des
parasites et les pestes des plantes, (iii) la modification de la structure du
sol, (iv) la stimulation des activités des symbiontes, et (v) la
production de substances (hormones) de croissance de la plante par la
stimulation des activités microbiologiques. Ces organismes peuvent, de
ce fait, être considérés comme une ressource naturelle
à intérêt agronomique pouvant être utilisés
pour accroître la production agricole de manière durable (Lee,
1985; Lavelle et al., 1998).
L'impact des vers de terre sur la production
végétale a été largement étudiés en
mésocosmes (pots soumis aux conditions naturelles) et en microcosme
(laboratoire) (Derouard et al.1997 ; Scheu et al., 1999 ;
Wurst & Jones, 2003 ; Kreuzer et al., 2004 ; Blouin et al.,
2006 ; Laossi et al., 2009; 2010). Dans ces expériences
où les variables environnementales (température, humidité
du sol et la disponibilité des ressources) sont plus ou moins
contrôlées et où la densité de vers de terre
ajoutés ne reflète pas la réalité, leurs
activités sont parfois exagérées (Eriksen-Hamel &
Whalen, 2007). Souvent, lorsqu'ils sont réalisés en conditions
naturelles, les organismes préexistants sont supprimés
préalablement (Gilot et al. 1996 ; Pashanasi et al.
1996 ; Gilot, 1994; 1997 ; Ouédraogo et al., 2006 ;
Eriksen-Hamel & Whalen, 2007). Ceci ne traduit pas non plus la
réalité car des interactions existent entres les
différents organismes du
2
sol et même entre espèces de vers de terre
(compactants et décompactants). Aussi, les travaux de recherche dont les
résultats sont destinés au monde paysan ont le plus souvent
été conduits sans la participation de ces paysans. Par ailleurs,
la manipulation des vers de terre en milieux paysans (plantations de
thé) a abouti à la mise au point d'une technologie appelée
Fertilisation Bio-Organique dans les Plantations Arborées (FBO)
(Senapati et al. 1999 ; 2002). La FBO initialement
expérimentée pendant 3 ans en Inde a permis d'accroître
jusqu'à 240% la production de thé, une rentabilité des
exploitations augmenter jusqu'à 260% et amélioration des
propriétés physico-chimiques et biologiques du sol (Senapati
et al. 1999). Cette technologie dont les détails de son
application n'ont pas été publiés par les auteurs est
protégée par un brevet (ref. PCT/FR 97/01363) (Senapati et
al. 2002 ; Dash et al. 2009). La FBO a cependant
été transférée dans d'autres pays du monde tels que
la Chine et l'Australie. En Côte d'Ivoire, des travaux sur l'impact des
vers de terre sur la production agricole se sont tous déroulés
dans les environs de la réserve de Lamto et concernent
généralement l'espèce M. omodeoi (Gilot et
al., 1996; Gilot, 1997; Derouard et al., 1997; Blouin et
al., 2006). Malgré le succès relatif de ces études,
aucune initiative visant à promouvoir l'utilisation de ces organismes
comme bio-fertilisant n'a été initié.
La présente étude est réalisée
dans la phase finale du projet « Conservation and Sustainable Management
of Below-Ground BioDiversity » (CSM BGBD) dont l'un des objectifs est de
valoriser les services écosystémiques des organismes du sol.
L'objectif particulier de cette étude est d'utiliser les vers de terre
comme bio-fertilisants pour améliorer la production agricole (culture de
maïs), en collaboration avec les paysans. Elle consistera à tester
l'hypothèse selon laquelle l'inoculation simultanée de vers de
terre à rôles fonctionnels contrastés (compactants contre
décompactants) favorise une augmentation de la production agricole.
I. 3
REVUE BIBLIOGRAPHIQUE
1. Ecologie des vers de terre utilisés
1.1. Millsonia omodeoi
Millsonia omodeoi appartient à la famille des
Acanthodrilidae. C'est une espèce non pigmentée de couleur rose
avec un clitellum orange (Annexe 1C). A l'âge adulte, les individus de
cette espèce mesurent 15 à 20 cm de long et pèsent en
moyenne (poids frais) 5 à 6 g (Lavelle, 1978). Bien qu'étant
hermaphrodite, leur reproduction est croisée comme tous les vers de
terre (Lavelle, 1978). L'espérance de vie des juvéniles à
la naissance est de 6 mois. Mais ils peuvent vivre plus de 20 mois. En
années normalement humides, la maturité sexuelle est atteinte
vers 10 mois. La fécondité moyenne est de 6 cocons par adulte et
par an; ceux-ci éclosent 21 jours en moyenne après la ponte et le
taux d'éclosion est proche de 100% (Lavelle, 1978).
Les vers de cette espèce vivent dans les vingt premiers
centimètres du sol et se nourrissent de terre moyennement riche en
matière organique provenant de l'horizon superficielle (0 à 20
cm), d'où leur appartenance au groupe des endogées
mésohumiques (Lavelle, 1981).
Les vers juvéniles peuvent ingérer 20 à
30 fois leur poids frais de terre par jour et produisent de gros turricules
compacts (diamètre > 2 mm), formés d'unités arrondies
et coalescentes qui contribuent à la compaction du sol, d'où leur
appellation de "vers compactant" (Blanchart et al., 1997).
Il faut également mentionner que cette espèce a
fait l'objet de plusieurs études en milieux contrôlés comme
semi contrôlés (Gilot et al.1996 ; Gilot et al
1997 ; Derouard et al.1997 ; Blouin et al.2006).
1.2. Hyperiodrilus africanus
H. africanus appartient à la famille des
Eudrilidae. Il représente l'un des rares vers de grande taille
appartenant à cette famille; les espèces de cette famille
étant généralement filiformes (Annexe 1B). C'est un ver
légèrement pigmenté qui mesure en moyenne, à
l'âge adulte, 8 à10 cm et pèse plus de 1 g. Les populations
de cette espèce se trouvent dans les régions humides de savanes
et de forêts perturbées (Omodeo, 1954 ; Madge, 1969) de l'Afrique
occidentale (Côte d'Ivoire, Nigeria) et centrale (RDC, Angola). La
durée maximale de vie des individus est de 8 mois (Tondoh, 1998). Ce
vers est caractérisé par une importante fécondité
(un adulte peut produire en moyenne 30 cocons par an), un nombre
élevé d'embryon par cocon (1 à 5) et une reproduction
parthénogénétique (Tondoh & Lavelle, 1997). Toutes ces
caractéristiques
4
démographiques font d'elle, une espèce à
grande capacité d'expansion et donc de profile démographique de
type « r » stratège « r » (Tondoh & Lavelle,
2005).
Les individus appartenant à cette espèce vivent
dans l'horizon superficiel du sol (0-20 cm) et s'y nourrissent. Ils exploitent
des zones plus riches en matière organique (mélange de
litière et de terre) en délaissant partiellement les sables: ce
sont des endogées polyhumiques (Tondoh, 1998). Ils produisent des
turricules friables (diamètre < 2 mm). Et au travers cet effet de
désagrégation du sol, ils sont appelés "vers
décompactant" (Blanchart et al., 1997).
2. Notion de groupe fonctionnel et de type
écologique des vers de terre
Un groupe fonctionnel est un ensemble d'espèces qui
exerce un effet similaire sur un processus biogéochimique
spécifique de l'écosystème (Swift et al. 2004).
Ainsi, en se focalisant sur l'impact des vers de terre sur la structure du sol,
on distingue des vers compactants et décompactants:
- les vers du groupe des compactants (gros vers) provoquent la
macro agrégation du sol à travers la production de gros
turricules compacts et réduisent ainsi la macroporosité du sol
;
- le groupe des décompactants renferme les vers de
petites tailles qui ont des effets opposés à ceux du groupe
précédent. Ils découpent les larges agrégats (>
5 mm) en petites pièces (0,25-2 mm) réduisant du coup, la
densité des gros agrégats (Blanchart et al., 1997 ;
Derouard et al., 1997; Lavelle et al., 2006).
La classification des vers de terre en types
écologiques est basée sur le mode d'utilisation des ressources,
de l'espace et d'adaptation aux conditions difficiles du milieu souvent
difficiles et imprévisibles. Ces types écologiques sont :
- les épigées: dans ce groupe, les
espèces vivent dans la litière et s'en nourrissent. Le cocon est
la forme de résistance de ces espèces aux conditions
défavorables ;
- les anéciques: les espèces de ce groupe vivent
la plupart du temps dans des réseaux de galerie enfoncés parfois
très profondément (1 à 6 m) et se nourrissent de
litière à la surface du sol. Leur forme de résistance est
une diapause vraie;
- quant au dernier groupe de cette classification, c'est -
à - dire les endogées, les individus se nourrissent
essentiellement de terre (matière organique). Ils sont appelés
oligo-, méso- ou polyhumiques suivant qu'ils se nourrissent d'une terre
énergiquement pauvre, moyennement riche ou riche. La forme de
résistance de ces espèces aux conditions défavorables se
fait par quiescence
5
Cependant, certains auteurs (Sheehan et al., 2006 ;
Sheehan et al., 2007 ; Laossi et al., 2009 ; Laossi et
al., 2010) prennent les types écologiques des vers de terre pour
des groupes fonctionnels.
3. Vers de terre et agriculture
Depuis la fin du XIX siècle (1881), Darwin avait
reconnu l'amélioration de la production végétale par les
vers de terre, invertébrés qualifiés d'ingénieurs
du sol (Lavelle et al., 1997). Depuis cette date jusqu'aujourd'hui, de
nombreuses études sur l'effet de ces organismes sur la production de
diverses espèces végétales ont été
effectuées (Lavelle et al., 2006).
La plupart des ces études confirment les travaux de
Darwin (Brown et al., 1999; Scheu, 2003; Brown et al., 2004;
Lavelle et al., 2006), pendant que quelques uns ont décrits des
effets nuls ou négatifs de ces organismes sur la production
végétale (Spain et al., 1992; Brown et al.,
1999; Joshi et al.,1999; Scheu, 2003).
Les vers de terre en tant que décomposeurs facilitent
la libération des éléments minéraux par la
décomposition de la matière organique. Avec les travaux
récents, le constat est que les vers n'agissent pas seulement sur la
disponibilité des nutriments pour la plante, mais ils influencent la
rhizosphère toute entière. Les mécanismes par lesquels les
vers agissent sur la croissance englobent des effets aussi bien directs
qu'indirects. Les effets directs se traduisent par la levée de la
dormance des semences par les turricules, le transport de petits grains en
surface ou à l'intérieur du sol et la création de galeries
qui favorisent la croissance des racines (Darwin, 1881; Ayanlaja et
al., 2001; Scheu, 2003). La production végétale est en
grande partie modifiée indirectement par les activités de ces
organismes à travers cinq processus (Scheu, 2003; Brown et al.,
2004) à savoir:
(i) la libération accélérée des
nutriments pour la plante ; la minéralisation de la matière
organique est accélérée au cours du transit de la
nourriture dans l'estomac des ver de terre et aussi à travers le rejet
des turricules contenant une proportion importante de minéraux
facilement assimilables par la plante (Subler et al., 1997 ;
Haynes et al., 1999).
(ii) la stimulation des symbiotes (micro-organismes
mutualistes et mycorhizes) ; ils augmentent les activités
microbiologiques par ce que leurs turricules (contenant des composés
facilement métabolisables), les galeries et les middens associés
constituent un véritable microenvironnement (conditions
physico-chimiques) favorable au développement des microorganismes
(bactéries) (Brown, 1995).
(iii)
6
la protection des plantes contre les pestes et parasites; les
activités des vers de terre rendent la plante plus vigoureuse et la
permettent ainsi de résister aux maladies et parasites (Lavelle et
al., 2006).
(iv) l'amélioration de la structure physique du sol ;
ces organismes modifient la porosité et l'agrégation du sol
(Shipitalo & Le Bayon, 2004) lors de leurs activités de
creusés et de rejet de turricules conduisant par la suite à une
augmentation de la disponibilité de l'eau et l'oxygène (Doube
et al., 1997) pour la plante.
(v) la production par des micro-organismes de certaines
substances (hormones) stimulatrices de la croissance végétale.
Les turricules des vers de terre contiennent des substances humiques (auxine)
qui influencent positivement la croissance des plantes (Muscolo et al.
1999 ; Nardi et al., 2002). Cette activité hormonale est due
à la présence dans l'estomac des vers de terre de microorganismes
qui accélèrent le processus d'humification et améliorent
ainsi la qualité de l'humus (Dell Agnola & Nardi, 1987).
Cependant, selon Blouin et al. (2006), les
mécanismes responsables de ces effets ne sont généralement
pas connus de manière précise.
Les relations entre les vers de terre, les sols et les
plantes, ont été largement étudiées à
travers les travaux réalisés à court terme en microcosme
ou au laboratoire. Cependant, l'extrapolation de ces résultats à
l'échelle de la parcelle et la quantification de l'influence des
activités des vers dans les expériences à cours terme sont
difficiles (Carpenter, 1996). L'hypothèse selon laquelle les
activités des vers sont exagérées dans les
expériences à petites échelles à cause du
contrôle des paramètres environnementaux (température,
humidité du sol et la disponibilité des ressources) ou de
l'introduction de nombres irréalistes de vers dans les
mésocosmes, serait l'une des raisons. Par ailleurs, les
expériences à court terme ne reproduisent pas fidèlement
les effets à long terme de ces animaux sur la dynamique de la
matière organique du sol et la croissance végétale (Barot
et al., 2007).
Ceci étant, quelle densité ou biomasse de vers
est susceptible d'accroître la production végétale? La
manipulation des vers de terre à grande échelle en milieux
paysans consistant à inoculer de larges populations de ces organismes,
pourrait apporter des éléments de réponse (Bohlen et
al., 2004). Ainsi, d'après Lavelle et al., (1994) et Gilot
(1994), une augmentation de la production végétale survient
après l'inoculation d'une biomasse supérieure à 40 g
m-2 ou lorsque la densité de vers de terre est
supérieure à 300 ind.m-2 (Eriksen-Hamel & Whalen,
2007).
Toutefois, la réussite des travaux sur la manipulation
des vers de terre en milieux paysans dépend des espèces
utilisées (Derouard et al., 1997; Eriksen-Hamel & Whalen,
2007), du type
7
de sol (Doube et al., 1997; Laossi et al.,
2010), de l'espèce végétale (Pashanasi et al.,
1996; Brown et al., 2004; Laossi et al., 2009), de sources de
nourriture et surtout de la pluviométrie (Pashanasi et al.,
1996; Eriksen-Hamel & Whalen, 2007). L'introduction de vers compactants
peut entraîner non seulement une compaction du sol qui à long
terme est néfaste pour les plantes, mais aussi pour les vers eux
même (Derouard et al., 1997). En plus, même si les vers
compactants et décompactants sont inoculés, il ne peut y avoir de
bons résultats en absence de pluies et de sources de nourriture
(Eriksen-Hamel & Whalen, 2007). En outre, certaines espèces
végétales ne réagissent pas de la même
manière aux activités des vers de terre car elles ne
possèdent pas les mêmes facteurs limitants au niveau des
ressources et utilisent différemment la même ressource (Laossi
et al., 2009). Par exemple, les légumineuses, n'étant
pas limitées par l'azote du milieu, auront une réaction
différente de celle des graminées aux activités des vers
(Brown et al., 2004).
8
II. MILIEU D'ETUDE
1. Localisation
L'étude a été conduite dans la
région du Centre Ouest de la Côte d'Ivoire, dans le
département d'Oumé (région du fromager),
précisément dans les environs du village Goulikao (6°17'N
5°31'W). Situé à 27 km de la ville d'oumé, le village
Goulikao est constitué de trois groupes socio ethniques : les
autochtones Gban (Gagou), les allochtones (Baoulé, Gouro, Wan,
Sénoufo, Malinké, etc.) et les allogènes
(Burkinabé, Béninois, Maliens, etc.). Il est entouré de 4
campements (Mafia, N'kroïdjo, Djèkoffikro et Petit Bouaké)
situés à des distances variables, chacun autour du village noyau
(FIG. 1). Ces campements sont constitués principalement d'allochtones
Baoulé.
Les parcelles expérimentales ont été
installées dans des jachères de 1 à 2 ans appartenant
à 10 paysans volontaires pour héberger l'étude et servir
d'intermédiaire auprès des autres paysans du village Goulikao et
campements associés (Tableau I).

Yamoussoukro
Abidjan
9
Figure 1: Localisation et situation du site d'étude.
10
Tableau I : Localisation géographique des parcelles
sélectionnées pour l'expérimentation
|
Noms des paysans
|
Altitude (m)
|
Latitude Nord (°min s)
|
Longitude Ouest (°min s)
|
|
Komenan Kacou
|
175.7
|
6°31'48.7»
|
5°32'53.2»
|
|
Djabla Sylvanus
|
170
|
6°31'23.9»
|
5°32'51.6»
|
|
Guény Christophe
|
167.2
|
6°31'17.7»
|
5°32'44.5»
|
|
Guény Henriette
|
177.5
|
6°30'98.3»
|
5°32'19.1»
|
|
N'guessan Emile
|
175.3
|
6°30'91.4»
|
5°32'15.5»
|
|
Komenan Béli
|
179.6
|
6°30'94.8»
|
5°32'10.9»
|
|
Tizié Mangoua
|
177.9
|
6°30'81.8»
|
5°31'87.4»
|
|
Zéli André
|
183
|
6°30'90.7»
|
5°31'80.3»
|
|
Gohan Golé
|
182.3
|
6°30'69.9»
|
5°32'12.4»
|
|
Yao K. Evrard
|
181.7
|
6°31'58.5»
|
5°31'71.0»
|
2. Climat
|
|
|
|
Le climat de cette région est du type
subéquatorial attiéen à pluviométrie bimodale. Il
est caractérisé par deux saisons pluvieuses dont la grande va de
Mars à Juin et la petite, de Septembre à Octobre. Elles sont
suivies de deux saisons sèches dont la grande est comprise entre
Novembre et Février, et la petite part du mois de juillet au mois
d'Août.
2.1 Pluviométrie
La pluviométrie moyenne au cours des 20
dernières années s'élève à 1371,88 mm. Elle
est de 1599,6 mm pour l'année 2008 et de 1356,09 mm pour l'année
d'étude (2009). La pluviométrie moyenne de 2009 donc est faible
par rapport à celles de l'année précédente et des
20 dernières années. Les précipitations mensuelles de 2009
sont comprises entre 1,53 mm en Décembre et 364,74 mm de pluies en
Avril, alors qu'elles sont entre 0 mm (Janvier) et 234,3 mm (Mai) pour
l'année 2008 (Fig. 2).
2.2 Température
Les températures sont constamment élevées
et leurs variations sont de faible amplitude sur les 22 ans. Les valeurs
moyennes annuelles sur les années 2008 et 2009 sont respectivement de
25,8°C et 26,4°C. Les températures mensuelles varient entre
24,1°C et 29,6°C en 2008 et enfin entre 25,1°C et 27,3°C en
2009.
Pour ce qui concerne la période d'étude
(Août à Décembre 2009), tous les mois, sauf le mois de
Septembre, sont considérés comme des mois secs, la
pluviométrie de chacun de ces mois étant inférieure au
double de leur température (Fig. 2).

Température (°C)
Pluviométrie (mm)
400
350
300
250
200
150
100
50
0
200
175
150
125
100
75
50
25
0
11
Jan Fev Mar Avr Mai Jui Jui Aou Sep OCT Nov Dec
Oct
Mois
|
Pluviométrie 2009 Pluviométrie 2008
Pluviométrie 88-07
Température 2009 Température 2008
Température 88-07
|
Figure 2: Diagramme ombrothermique sur la période
1987-2007 et au cours des années 2008 et 2009.
3. Sols et végétation
Les sols de la région d'Oumé, sols sont issus
des formations précambriennes composées de granites, de schistes
et d'intrusions basiques. Ils sont caractérisés de sols
ferrallitiques moyennement désaturés (Perraud, 1971; Lecomte,
1990). Distribués de façon homogène, avec toutefois, des
différences topographiques prononcées (Angui et al.,
2005), ils présentent généralement un horizon
gravillonnaire épais et un concrétionnement important. Les sols
d'Oumé sont parfois indurés sur le sommet et à mi-versant
avec des caractéristiques chimiques qui se présentent de la
manière suivante: un pli légèrement acide (6,5); un taux
de saturation en bases supérieur à 50%; une somme de bases
échangeables de 13,5 cmolc.kg-1;
12
une carence en phosphore assimilable (0,48%); une teneur en
argile supérieure à 30%; un taux de matière organique
voisin de 5%. L'argile dominante est la kaolinite (Lecomte, 1990).
La végétation naturelle est une forêt
dense humide de type mésophile ou sémi-décidue appartenant
au secteur ombrophile du domaine guinéen (Monnier, 1983). Cette
forêt, avec une superficie de 58580 ha au départ,
représente actuellement qu'une superficie de 7232 ha et n'existe que
sous forme d'îlots dans le domaine rural (Assié et al.,
2008). La plus grande partie de cette forêt (presque toute la forêt
restante) est protégée par le gouvernement sous forme de
forêt classée (forêt classée de la
Téné).
4. Peuplements de vers de terre
Le peuplement de vers de terre de la région
d'Oumé est composé de 20 espèces dont 4
indéterminée (Dichogaster sp 1, Dichogaster sp
2 et Dichogaster sp 3 et Agastrodrilus sp). Ces vingt
espèces se repartissent entre deux familles (
Acanthodrilidae.et Eudrilidae)
et 7 genres (Tondoh et al., 2007; Baidai, 2009; Bosso, 2010) (Tableau
II).
La biomasse des vers de cette région est dominée
par Millsonia omodeoi et Dichogaster terrae-nigrae.
L'espèce Hyperiodrilus africanus domine en termes de
densité dans les cultures vivrières. Les jachères
récurrentes à C. odorata constituent les milieux qui
hébergent une abondance et une diversité plus grandes de ces
organismes (Monin, 2005; Guéi, 2006; Tondoh et al., 2007).
13
Tableau II : Espèces de vers de terre présentes
dans la région d'Oumé
Espèces Familles
Millsonia omodeoi (Sims, 1986) Acanthodrilidae
Millsonia lamtoiana (Omodeo & Vaillaud, 1967)
Acanthodrilidae
Dichogaster baeri (Sciacchitano, 1952)
Acanthodrilidae
Dichogaster eburnea Acanthodrilidae
Dichogaster ehrhardti (Michaelsen, 1898)
Acanthodrilidae
Dichogaster saliens (Beddard, 1893)
Acanthodrilidae
Dichogaster mamillata Acanthodrilidae
Dichogaster terrae nigrae (Omodeo & Vaillaud,
1967) Acanthodrilidae
Dichogaster papillosa (Omodeo, 1958)
Acanthodrilidae
Dichogaster sp 1 Acanthodrilidae
Dichogaster sp 2 Acanthodrilidae
Dichogaster sp 3 Acanthodrilidae
Agastrodrilus multivesiculatus (Omodeo &
Vaillaud, 1967) Acanthodrilidae
Agastrodrilus opisthogynus Acanthodrilidae
Agastrodrilus sp Acanthodrilidae
Hyperiodrilus africanus (Beddard, 1891) Eudrilidae
Lavellea composita (Omedeo, 1985 Eudrilidae
Scolecillus compositus (Omodeo, 1958), Eudrilidae
Stuhlmannia palustris (Omodeo & Vaillaud, 1967),
Eudrilidae
Stuhlmannia zielae (Omodeo, 1963) Eudrilidae
14
III. MATERIELS ET METHODES
1. Choix des espèces animales et
végétale
Les espèces M. omodoei et H. africanus
ont été choisies pour être utilisées comme
bio-fertilisants pour plusieurs raisons: (i) ces deux espèces de vers de
terre appartiennent à des groupes fonctionnels différents dont
l'action antagoniste de compaction (M. omodeoi) et de
décompaction (Eudrilidae) conduit au maintien d'une structure grumeleuse
du sol (Blanchart, 1990; Derouard et al., 1997) ; (ii) la biologie et
la démographie de ces deux espèces sont relativement bien connues
(Lavelle, 1983; Gilot, 1994; Tondoh & Lavelle, 1997; Tondoh, 1998; Tondoh
& Lavelle, 2005) ainsi que leur impact sur la structure du sol (Blanchart,
1990; Blanchart et al., 1997 & 1999) et la disponibilité en
éléments minéraux du sol (Martin, 1991; Blouin et
al., 2006) ; (iii) ces deux espèces sont résistantes aux
conditions difficiles des milieux (faible fertilité, forte
évapotranspiration, etc.).
Le maïs (Zea mays), variété jaune
de type pioneer issue du croisement entre la variété jaune
Bouaké et la variété blanche IRAT 8, a été
choisi pour son cycle court (3 mois), mais aussi par ce que les paysans de la
région d'Oumé l'utilisent pour leur subsistance et comme source
de revenu.
2. Dispositif expérimental
Le dispositif expérimental est constitué d'un
ensemble de 10 parcelles (20 m x 10 m) situées dans le domaine de
paysans différents. Ces parcelles sont espacées d'au moins 1 km
dans le terroir de Goulikao. On considère que ces parcelles jouissent
d'un environnement agro-pédologique similaire et peuvent par
conséquent être considérées comme des
répétitions. Chacune des dix parcelles a été
délimitées dans des jachères (1 à 2 ans)
dominées par l'adventice Chromolaena odorata (Fig. 3A). Chaque
parcelle comprend cinq placettes carrées de 3 m de côté
séparées par des allées d'un mètre de large (Fig.
3B). Pour prévenir les fuites des vers de terre, des tranchées
d'environ 10 cm de largeur ont été creusées autour des
différentes placettes (Fig.3C). Ces tranchées ont servi de lieux
d'enfouissement des bâches noires (30 cm de hauteur dont 20 cm en dessous
et 10 cm au dessus du sol). Une palissade renforcée par des feuilles de
palmes a été posée autour des différentes parcelles
pour réduire la pression des animaux (Aulacodes et rats palmistes) sur
les pieds de maïs (Fig. 3D).

15
A) Jachère à C. odorata B) Parcelle
prête pour le semis

C) Tranchée autour des placettes D)
Palissade autour d'une parcelle
Figure 3: Parcelles d'étude et les
différentes étapes de sa mise en place pour le semis (Photos
Arnauth Guéi)
16
Cinq traitements ont été mis en place dans
chacune des dix parcelles sélectionnées (Fig. 4):
- témoin sans vers de terre et sans engrais (M);
- maïs + M. omodeoi (M+Mo);
- maïs + H. africanus (M+Ha);
- maïs + M. omodeoi + H. africanus
(M+Mo+Ha);
- maïs+ urée + superphosphate triple (M+U+SPP).
|
Maïs
|
|
Maïs
|
|
Maïs + M.
|
|
Maïs
|
|
Maïs+urée
|
|
+
|
1 m
|
+
|
|
omodeoi +
|
|
|
|
+superphos
|
|
M. omodeoi
|
|
H. africanus
|
|
H.
africanus
|
|
|
|
phate triple
|
|
|
|
|
|
|
|
|
|
|
3 m
|
Figure 4: Disposition des différentes
placettes au sein des parcelles
2. Culture de maïs et inoculation des vers de
terre
3.1. Culture de maïs
Le semis du maïs a été
réalisé à l'aide de machettes pour établir des
poquets de faible profondeur (environ 3 à 4 cm). Les poquets
étaient séparés de 0,4 m et établis sur trois
lignes distantes de 0,8 m par carré de dimension 3 m x 3 m. Deux
à trois grains de maïs ont été placés dans ces
poquets. Une semaine après le semis, un autre semis a été
effectué dans le but de remplacer les grains qui n'ont pas germé.
Le démariage s'est déroulé 25 jours après les semis
pour ne laisser qu'un seul pied par poquet. Cela correspond à un total
de 21 pieds par placette, soit 31250 pieds ha-1 (Koné,
2009).
Le désherbage des parcelles a été
effectué à la main afin d'éviter les blessures des vers.
Le premier nettoyage a eu lieu quinze jours après les semis. Les deux
derniers nettoyages sont survenus à un et deux mois après
semis.
L'urée (46% d'azote) a été
appliquée à la dose de 50 kg.ha-1, soit 98,44 g par
carré de 9 m2. Le tiers de cette quantité a
été appliqué juste après le semis. Les deux tiers
restants ont été appliqués 40 jours après le semis
(Kang, 1997). Le superphosphate triple (45% de phosphore) a été
entièrement appliqué après le semis à la dose de 30
kg.ha-1, soit 60,28 g par parcelle de 9 m2 (Kang, 1997).
Les fertilisants chimiques ont été appliqués autour des
pieds de maïs.
17
3.2. Inoculation des vers de terre
La mortalité élevée des vers de terre
constitue la principale contrainte pour la réussite des
expériences avec introduction de ces organismes (Tondoh &
Konaté, 2005). Pour réduire la mortalité des vers et pour
rendre leurs effets plus durable, l'inoculation s'est effectuée en 3
phases : (i) la première inoculation a eu lieu 15 jours après le
semis où 225 g, soit 25 g.m-2 de M. omodeoi (Mo) et
225 g de H. africanus (Ha) ont été introduits dans les
placettes de 3 m x 3 m; (ii) 30 jours après semis, cette même dose
a été appliquée, et (iii) la dernière inoculation a
eu lieu le 45ème jour après le semis où la
biomasse de 40 g.m-2 (360 g par carré) a été
appliquée.
Les vers utilisés pour l'inoculation ont
été récoltés à la veille des jours
d'inoculation dans les bas-fonds ou les parcelles laissées en
jachère situées aux alentours des différents milieux
d'expérimentation. Les individus utilisés sont des stades adultes
et sub-adultes, plus résistants, ont été
récoltés à l'aide de houes (Annexe 1A) et directement
inoculés le soir ou conservés dans des bassines contenant de la
terre de jachères pour la prochaine inoculation. L'introduction des vers
au champ s'est effectuée au coucher du soleil dans des tranchées
de 5 cm de profondeur, préalablement humidifiées. Ces
tranchées ont été creusées entre les lignes de
maïs (Annexe 1D). Pour éviter que ces animaux soient
dévorés par des prédateurs, il a fallu attendre que tous
les individus appliqués pénètrent totalement dans le
sol.
Au total, c'est une biomasse de 90 g.m-2 qui a
été inoculée dans les différents traitements
à base de vers de terre. La moitié de la biomasse de chacune des
espèces a été utilisée pour les traitements mixtes
(M+Mo+Ha) (Tableau III). Cette dose de vers de terre appliquée est en
rapport avec les propos de Lavelle et al. (1994 et 1997) et de Gilot
(1994). Selon eux, une augmentation de la production végétale
survient en milieux paysans lorsqu'une biomasse de vers de terre
supérieure à 40 g.m-2 est inoculée. Le nombre
de vers introduits par enclos n'a pas été contrôlé.
Aucune élimination des vers préexistants n'a été
effectuée au préalable.
Par ailleurs, les travaux de Guéi (2006) et Tondoh
et al. (2011) réalisés dans les agro
écosystèmes de Goulikao (Oumé) ont montré que la
biomasse globale de M. omodeoi des agro écosystèmes
d'Oumé (Goulikao) oscille entre 0,8 et 14,1 g.m-2 tandis que
celle de H. africanus est entre 0,2 et 3,9 g.m-2. Les
biomasses de M. omodeoi et H. africanus de terre dans les
cultures vivrières selon ces auteurs sont respectivement de 14,1
g.m-2 et 3,7 g.m-2.
18
Tableau III : Biomasses des deux espèces inoculées
et les différentes périodes d'inoculations
|
Traitements
|
Espèces introduites
|
Biomasse (g.m-2)
|
|
|
15 jours
semis
|
après 30 jours après
semis
|
45 jours après
semis
|
|
Témoin (M)
|
M. omodeoi
|
0
|
0
|
0
|
|
H. africanus
|
0
|
0
|
0
|
|
M+Mo
|
M. omodeoi
|
25
|
25
|
40
|
|
H. africanus
|
0
|
0
|
0
|
|
M+Ha
|
M. omodeoi
|
0
|
0
|
0
|
|
H. africanus
|
25
|
25
|
40
|
|
M+Mo+Ha
|
M. omodeoi
|
12,5
|
12,5
|
20
|
|
H. africanus
|
12,5
|
12,5
|
20
|
|
M+U+SPP
|
M. omodeoi
|
0
|
0
|
0
|
|
H. africanus
|
0
|
0
|
0
|
3. Mesures effectuées à la fin de
l'expérimentation
4.1. Au niveau du maïs
? Croissance du maïs
La croissance du maïs a été mesurée
à travers la hauteur des plants aux 30ème,
50ème et 75ème jours après semis sur
9 pieds de maïs par traitement. Les mesures se sont effectuées sur
les mêmes pieds de maïs à tous les stades. Un
décamètre a été utilisé pour la mesure de ce
paramètre (Annexe 2). Pour tenir compte de
l'hétérogénéité probable des conditions du
sol sur une même parcelle, les 9 pieds de maïs ont été
sélectionnés de sorte à couvrir toute la parcelle (Fig.
5). La hauteur du plant de chaque traitement est la moyenne des hauteurs des 9
pieds du traitement ayant fait l'objet de mesures.
? Production du maïs
La production de maïs a été
évaluée sur les pieds de maïs parvenus à
maturité (90 jours). Ils ont été dessouchés dans
deux rectangles de dimension 0,8 m x 0,4 m placées selon la diagonale
des placettes, ce qui correspond à 4 pieds par rectangle, soit un total
de 8 pieds par placette (Fig. 5). Ces pieds de maïs, séchés
d'abord à l'air libre pendant 2 à 5 jours, ont
été
ramenés au laboratoire pour être
séchés à l'étuve pendant 48 heures à 65
°C avant d'être pesés. Cinq paramètres ont permis
d'estimer la production du maïs. Il s'agit de :
1- la biomasse épigée,
2- le nombre des épis,
3- le poids des épis,
4- le poids des grains,
5- le poids des racines et
Le ratio biomasse épigée/biomasse racinaire a
été également évalué.
|
Pied de maïs non sélectionné
Pied de maïs sélectionné
Rectangle pour prélever les pieds
|
19
Figure 5: Dispositions des pieds de maïs
sélectionnés pour la mesure du paramètre de croissance et
les rectangles pour leur prélèvement.
? Efficience de l'utilisation de l'eau (EUE)
L'efficience de l'utilisation de l'eau a été
estimée en utilisant la formule de Ouédraogo et al .
(2006) en vue de savoir si la présence des vers de terre favorise une
meilleure utilisation de l'eau par le maïs. Ce paramètre a
été calculé à partir de la formule suivante :
EUE = (Biomasse épigée du traitement - Biomasse
épigée du témoin) (kg.ha-1) /
Pluviométrie totale (mm).
4.2. Au niveau des vers de terre
Après avoir procédé à la
récolte du maïs (décembre 2009), les vers de terre ont
été échantillonnés à l'aide de deux petits
monolithes de terre (TSBF) de dimension 25 x 25 x 30
20
cm dans chacune des placettes d'étude. Les vers
récoltés furent conservés dans des piluliers contenant du
formol dilué à 4 % afin d'être identifiés au niveau
de l'espèce et pour la détermination de l'abondance et la
diversité des espèces. Cette identification a été
réalisée à partir des spécimens de
référence validés par le Muséum d'Histoire
Naturelle de Budapest à Hongrie.
? Abondance des vers de terre
La densité (nombre d'individus au mètre
carré) et la biomasse (masse (g) d'individus par mètre
carré) ont été mesurées pour estimer l'abondance du
peuplement des vers de terre au sein des différentes parcelles.
? Diversité des vers de terre
La diversité du peuplement de vers de terre a
été analysée en mesurant 3 paramètres.
- Le nombre moyen d'espèces; il a été
obtenu à partir du nombre d'espèces par monolithe et a permis de
mesurer la richesse spécifique des peuplements de vers de terre.
- L'indice de Shannon-Weaver (H) permet de mesurer la
diversité spécifique d'un peuplement à partir du nombre
d'espèces et de leur abondance relative (Legendre & Legendre, 1984).
Cet indice a été calculé à partir de la formule
suivante :
n
? ?? Pi . log 2 Pi où ni N
H Pi ? correspond à la proportion de
l'espèce i dans le peuplement, ni
i
l'effectif de l'espèce i et N l'effectif total.
- L'équitabilité (E) est calculée
parallèlement à H afin de montrer la répartition des
effectifs entre les espèces. Elle est calculée à partir de
la formule ci-dessous :
H
log2
E ? avec H qui correspond à l'indice de Shannon et S le
nombre moyen d'espèces.
S
L'équitabilité varie de 0 à 1: elle tend
vers 0 quand la quasi-totalité des effectifs est concentrée sur
une espèce ; elle est de 1 lorsque toutes les espèces ont la
même abondance.
5. Perception des paysans des effets des vers de terre
sur la fertilité du sol et sur la production agricole
L'enquête s'est déroulée dans les mois de
Novembre et Décembre 2009. Au total, 95 paysans dont les âges
varient de 21 à 75 ans ont été interviewés. Ils
sont composés des 10 paysans propriétaires des parcelles
expérimentales, 40 autres paysans de Goulikao, 20 paysans du
21
campement "Petit Bouaké" et 25 paysans du campement
"Djèkoffikro". L'interview des 10 propriétaires des parcelles a
débuté chez eux et s'est achevé sur les parcelles de
maïs. Quant aux autres paysans, ils ont été tous
questionnés chez eux, à leur domicile. Les personnes
interrogées appartiennent à toutes les couches socio-ethniques
présentent dans la zone d'étude (Gban, Baoulé, Gouro, Wan,
Mossi, Béninois, Malinké, Sénoufo, etc.).
Le questionnaire réalisé à l'aide d'une
fiche d'enquête comprenait les informations suivantes : la
présentation, les cultures vivrières, les connaissances à
propos des vers de terre, les organismes indicateurs de la qualité du
sol et les effets des vers de terre sur la fertilité du sol et sur le
rendement des cultures (Annexe 5).
6. Analyses statistiques
Après vérification de la normalité
à l'aide du test de Shapiro-Wilk, le test de Levene, au seuil de 5% a
été utilisé pour vérifier
l'homogénéité des variances. Le test paramétrique
ANOVA 1 (analyse de variances à un facteur) a permis de comparer les
moyennes des données issues des placettes (50 placettes) lorsque les
variances étaient homogènes. Dans le cas contraire, le test non
paramétrique de Kruskall-Wallis, au seuil de 5%, a été
utilisé. Toutes ces analyses statistiques ont été
réalisées à l'aide du logiciel Statistica (1999).
Par ailleurs, une Analyse en Composantes Principales (A.C.P)
regroupant les paramètres de production du maïs a été
effectuée afin d'identifier ceux qui sont sensibles à
l'introduction des vers de terre et à l'application des engrais. Cette
analyse multivariée a été réalisée à
l'aide du logiciel ADE-4 (Thioulouse et al., 1997).
Les données de l'enquête socio-économique
ont été analysées à l'aide du test Khi2
au seuil de 5%.
22
IV. RESULTATS
1. Maïs
1.1 Croissance du maïs
Au stade 30 jours après semis, la hauteur maximale des
plants de maïs traités à l'aide du vers M. omodeoi
était de 45,9 cm alors que le témoin avait une hauteur de
58,21 cm. Cependant, le test ANOVA 1 ne montre aucune différence
significative entre ces différentes hauteurs (p > 0,05) (Tableau
IV).
Au 50ème jour, le témoin a
enregistré la plus grande valeur d'hauteur des plants (150,36 #177;
10,33 cm). Venaient ensuite les traitements M+Mo+Ha (147,71 #177; 10,09 cm),
M+Ha (135,42 #177; 11,89 cm), et M+U+SPP (131,60 #177; 14,50 cm). Le traitement
M+Mo produit encore les pieds de maïs les plus courts (117,71 #177; 10,48
cm). Toutefois, la hauteur des plants ne varie pas significativement d'un
traitement à un autre (p > 0,05) à ce stade (Tableau IV).
Au dernier stade, le traitement à base de fertilisants
chimiques présentait les plus grands pieds de maïs (242,40 #177;
11,16 cm) alors que les pieds issus du traitement M+Mo étaient toujours
les plus courts (231,15 #177; 11,61 cm). Le témoin et les traitements
M+Mo+Ha et M+Ha ont affichés des hauteurs de plant
intermédiaires. Comparés une fois de plus au témoin, les
hauteurs des plants ne sont pas significativement affectées par
l'activité des vers (p > 0,05) (Tableau IV).
Tableau IV: Influence des vers de terre et des engrais
inorganiques sur la hauteur des plants de maïs aux stades 30, 50 et 75
jours après semis.
|
Traitements
|
Hauteur du plant (cm)
|
|
|
|
|
|
Stade 30 jours
|
Stade 50 jours
|
Stade 75 jours
|
|
Maïs
|
58,21
|
#177; 4,33
|
150,36
|
#177; 10,33
|
240,46
|
#177; 7,14
|
|
Maïs + Mo
|
45,90
|
#177; 2,9
|
117,77
|
#177; 10,48
|
231,15
|
#177; 11,61
|
|
Maïs + Ha
|
50,59
|
#177; 3,49
|
135,42
|
#177; 11,89
|
239,58
|
#177; 9,52
|
|
Maïs + Mo + Ha
|
55,90
|
#177; 3,45
|
147,71
|
#177; 10,09
|
239,73
|
#177; 8,34
|
|
Maïs + U + SPP
|
54,63
|
#177; 3,27
|
131,60
|
#177; 14,50
|
242,40
|
#177; 11,16
|
23
1.2. Production du maïs ? Biomasse
épigée
La biomasse de la partie aérienne (feuilles, tiges et
épis) ne varie pas significativement d'un traitement à un autre
(p > 0,05). Néanmoins des augmentations de+15,4%, +12,1% et +0,8% ont
été respectivement observées au niveau des traitements
M+Mo+Ha, M+U+SPP et M+Ha. Quant au traitement M+Mo, une baisse de -16,1% de la
biomasse a été enregistrée (Fig. 6a).
? Nombre d'épis
Le traitement avec association des vers a produit plus
d'épis (135937,50 épis.ha-1) pendant que le
témoin en a produit le plus faible (129687,50
épis.ha-1). Les traitements M+Mo et M+U+SPP ont, quant
à eux, produit le même nombre d'épis (131250,00
épis.ha-1). Ce dernier est légèrement
inférieur au nombre d'épis obtenu avec l'ajout de l'espèce
H. africanus (134375,00 épis.ha-1). Toutefois,
l'importance de ces différences entre les différents traitements
n'est pas significative (p > 0,05) (Fig. 6b).
? Poids des épis
Le poids le plus élevé provient du traitement
où les deux vers ont été associés (21,18 #177; 1,26
t.ha-1). La biomasse des épis obtenue dans le traitement
à base de H africanus (17,73 #177; 1,65 t.ha-1) est
sensiblement égale à celle avec l'application d'engrais chimiques
(17,18 #177; 1,06 t.ha-1). Le traitement M+Mo a produit un faible
poids d'épis (15,18 #177; 1,48 t.ha-1). Comparé au
témoin (16,69 #177; 1,39 t.ha-1), le poids des épis a
augmenté de +26,9% au niveau du traitement avec association des deux
vers, de +6,2% avec l'inoculation de H africanus et de +2,9% avec
l'application des engrais chimiques. Contrairement aux autres traitements, une
réduction de - 9% est constatée avec l'introduction de M.
omodeoi. La biomasse des épis est significativement
rehaussée avec l'application des vers de terre et des intrants
inorganiques (p < 0,05) (Fig. 6c).
? Poids total des grains
La production des grains a également augmenté
avec l'ajout des vers de terre et des engrais chimiques. L'augmentation du
poids des grains est de 21,5%; 21,2%; 19,4% et 13,9%, respectivement, dans les
traitements M+Mo, M+Mo+Ha, M+U+SPP et M+Ha. Toutefois, cette différence
entre ces traitements n'est pas significative (p > 0,05) (Fig. 6d).
24
? Poids des racines
La production de racines est forte (4,93 t.ha-1
soit une augmentation de +30,77%) lorsque les deux vers sont
associés, moyenne avec l'application des engrais (4,36 t.ha-1
soit + 15,65%) et de H. africanus (4,13 t.ha-1 soit +
9,55%) alors qu'elle est réduite avec l'introduction de M. omodeoi
(3,61 t.ha-1 soit -4,24%), le témoin ayant
affiché un poids égale à 3,77 t.ha-1.
Toutefois, cette différence observée entre les poids des racines
issus des différents traitements n'est pas significative (p > 0,05)
(Fig. 6e).
? Ratio biomasse épigée/racine
L'inoculation des vers de terre a occasionné une
réduction du ratio biomasse épigée/racine du maïs
tandis que l'application des engrais chimiques donne un résultat (10,09
#177; 0,79) sensiblement égale à celui du témoin (10,28
#177; 0,32 ; le plus grand ratio biomasse épigée/racine). La
réduction de la valeur du ratio est beaucoup plus marquée lorsque
les deux espèces sont associées (-10,02%) alors qu'elle est
respectivement de -7,78% et -5,45% avec l'introduction de H. africanus
et M. omodeoi. La diminution de la valeur du ratio biomasse
épigée/racine du maïs sous l'effet des engrais et des vers
de terre n'est cependant pas significative (p > 0,05) (Fig. 6f).

Biomasse épigée (t.hã1)
40
20
60
50
30
10
0
a)
Nombre d'épis (épis.hã1)
145000
140000
135000
130000
125000
120000
b)

Traitements
6
Poids des grains (t.hã1)
5
25
c)
Poids des épis (t.hã1)
4
20
3
15
2
10
1
5
0
e)
0
6
Poids des racines (t.ha-1)
5
4
3
Ratio
2
1
0
11,5
11
10,5
10
9,5
9
8,5
8
f)
d)
Traitements
25
Figure 6: Effets des vers de terre et des engrais chimiques
sur la production du maïs; (a) biomasse épigée, (b) nombre
d'épis, (c) poids des épis, (d) poids des grains, (e) poids des
racines et (f) ratio biomasse épigée/racine.
1.3. Efficience de l'utilisation de l'eau (EUE)
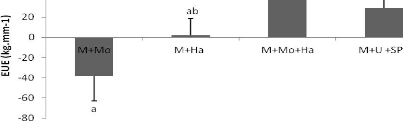
Les valeurs de l'EUE par le maïs sont plus
élevées dans deux des traitements avec vers (M+Ha et M+Mo+Ha) et
celui à base d'intrants inorganiques par rapport au troisième
traitement avec vers (M+Mo) où une baisse de -38,23 kg.mm-1 a
été observée. L'augmentation de l'EUE est plus importante
au niveau du traitement M+Mo+Ha (36,61 kg.mm-1). Le traitement
à base d'intrants inorganiques vient en seconde position avec une valeur
de 28,83kg. mm-1, puis M+Ha avec une valeur de 1,99 kg.
mm-1. Cette variation dans l'efficience de l'utilisation de l'eau
par le maïs entre les traitements est significative (p < 0,05) (Fig.
7).
26
Figure 7: Effets des traitements sur l'efficience de
l'utilisation de l'eau
27
1.4. Paramètres de production du maïs sensibles
aux différents traitements
Les deux premiers axes du cercle de corrélation
(Fig.11a) expriment 83,3% de l'information dont 67,2% détenues par l'axe
1 et 16,1% par l'axe 2. Toutes les analyses ont été
réalisées selon l'axe 1 qui contient 4 fois plus d'information
que le second. L'analyse du cercle de corrélation a
révélé que tous les paramètres de production (la
biomasse épigée, le nombre des épis, le poids des
épis et le poids des racines) sont positivement corrélés
à l'axe 1. Parmi ces paramètres seuls le poids des épis,
le poids des racines et la biomasse épigée sont fortement
corrélés à cet axe. Le poids des grains est quant à
lui positivement corrélé au deuxième axe, alors que c'est
tout à fait le contraire pour le nombre des épis.
La projection des 5 traitements dans le plan factoriel 1-2
montre une ordination des traitements selon trois groupes : (i) le traitement
M+Mo+Ha, (ii) les traitements M+U+SPP et M+Ha et, (iii) les traitements M+Mo et
M. Toutefois, cette ordination des traitements n'est pas significative (p >
0,05).

a)
Axe 1 (67.2%)
Axe 2 (16.1%)
Nombre des épis
Poids des grains
Biomasse épigée
Poids des racines
Poids des épis
1
-1 1
-1
b)
M
M+Mo
M+Ha
P < 0.145
M+U+SPP
M+Mo+Ha
-6.5 4
-3
3
Figure 8: Analyse en Composantes Principales des
paramètres de production du maïs; (a) Cercle de corrélation
des paramètres de production du maïs, (b) Distribution des
différents traitements dans le plan factoriel 1-2.
28
2. Impact de l'inoculation sur le peuplement de vers de
terre
Le peuplement de vers de terre sur l'ensemble du site est
riche de 13 espèces dont une non identifiée (Dichogaster
sp). Ces espèces échantillonnées sont
réparties en 5 genres et 2 familles (Tableau V).
Par comparaison à la biomasse introduite, la biomasse
de H. africanus et celle de M. omodeoi ont
considérablement diminuées. La diminution de la biomasse de
H. africanus est plus importante au niveau du traitement M+Ha (-95%)
par rapport au traitement M+Mo+Ha (84%). Quant au traitement M+Mo où
l'espèce M. omodeoi a été introduite, on a
enregistré une réduction de -60% de sa biomasse, tandis qu'au
niveau du traitement avec association des deux espèces, la biomasse a
été réduite de -67%. Les traitements M+U+SPP et M
(témoin), où les vers de terre n'ont pas été
introduits, ont montré les plus faibles biomasses (Tableau VI).
29
Tableau V: Espèces de vers de terre
échantillonnées sur l'ensemble des parcelles
Espèces Familles
Millsonia omodeoi (Sims, 1986) Acanthodrilidae
Millsonia lamtoiana (Omodeo & Vaillaud, 1967)
Acanthodrilidae
Dichogaster ehrhardti (Michaelsen, 1898)
Acanthodrilidae
Dichogaster baeri (Sciacchitano, 1952)
Acanthodrilidae
Dichogaster saliens (Beddard, 1893)
Acanthodrilidae
Dichogaster mamillata Acanthodrilidae
Dichogaster terrae nigrae (Omodeo & Vaillaud,
1967) Acanthodrilidae
Dichogaster papillosa (Omodeo, 1958)
Acanthodrilidae
Dichogaster sp Acanthodrilidae
Agastrodrilus multivesiculatus (Omodeo &
Vaillaud, 1967) Acanthodrilidae
Agastrodrilus opisthogynus Acanthodrilidae
Hyperiodrilus africanus (Beddard, 1891) Eudrilidae
Stuhlmannia zielae (Omodeo, 1963) Eudrilidae
Tableau VI: Biomasses des vers de terre introduits et celles
récoltées des différents traitements à la fin des
travaux
Traitements Biomasse totale introduite (g.m-2)
Biomasse totale récoltée (g.m-2)
|
|
|
|
|
|
M omodeoi
|
H africanus
|
M. omodeoi
|
H. africanus
|
|
M
|
0
|
0
|
5,63 #177; 3,06
|
1,48 #177; 0,92
|
|
M+Mo
|
90
|
0
|
35,68 #177; 12,91
|
1,64 #177; 0,75
|
|
M+Ha
|
0
|
90
|
2,40 #177; 1,12
|
4,52 #177; 1,31
|
|
M+Mo+Ha
|
45
|
45
|
14,61 #177; 2,46
|
7,10 #177; 1,63
|
|
M+U+SPP
|
0
|
0
|
2,28 #177; 1,20
|
0,96 #177; 0,52
|
30
Comparativement au témoin, en considérant le
peuplement global des vers de terre, la biomasse et la densité moyenne
des vers de terre ont augmenté dans les traitements où M.
omodoei et H. africanus ont été introduits. Le
traitement M+Mo a enregistré la plus forte densité moyenne (144,8
#177; 41,3 ind.m-2) et la plus forte biomasse moyenne (37,4 #177;
19,95 g.m-2), ce qui correspond à une hausse de 207 % de la
densité et de 183% de la biomasse. On a noté également une
augmentation de 127% de la densité au niveau des traitements M+Ha+Mo et
M+Ha, alors que pour la biomasse moyenne, elle est de 159% pour M+Mo+Ha et de
57% pour M+Ha (Fig.9). Le traitement à base d'engrais a
entraîné une augmentation de la biomasse de 12% mais une
réduction de -24% de la densité des vers. L'augmentation de la
densité dans les traitements à base vers de terre est
significative (p < 0,01) tandis que celle de la biomasse ne l'est pas (p
> 0,05).
Toujours en comparant les autres traitements au témoin,
la richesse spécifique, l'indice de Shannon et l'Equitabilité
sont également plus élevés au niveau des traitements
à base de vers de terre, avec un effet plus marqué pour le
traitement M+Mo+Ha. Ce dernier traitement a montré la plus grande
richesse spécifique (3,9 #177; 0,49 espèces.m-2), la
plus forte valeur d'indice de Shannon (1,61 #177; 0,2) et de même pour
l'équitabilité (0,8 #177; 0,09). A l'opposé, le traitement
à base de fertilisants chimiques héberge le plus petit nombre
d'espèces (1,7 #177; 0,37 espèces.m-2) et
possède le plus petit indice de Shannon (0,74 #177; 0,21) et la plus
petite valeur de l'équitabilité (0,58 #177; 0,16). Les
traitements M+Ha, M+Mo et le témoin (M) affichent les richesses
spécifiques, les indices de Shannon et d'équitabilités
intermédiaires. Les richesses spécifiques pour ces trois
traitements sont: 3,7 #177; 0,56 espèces.m-2 (M+Ha); 3,4
#177; 0,34 espèces.m-2 (M+Mo) et 2,6 #177; 0,4
espèces.m-2 (témoin). Pour l'indice de Shannon et
l'Equitabilité, ils sont respectivement de 1,52 #177; 0,21 et 0,77 #177;
0,09 (M+Ha); 1,21 #177; 0,07 et 0,76 #177; 0,06 (M+Mo) et 1,05 #177; 0,2 et
0,71 #177; 0,12 (témoin). L'augmentation de la richesse
spécifique et de l'indice de Shannon Weaver (H) du traitement M+USPP au
traitement avec association M. omodeoi et H. africanus est
significative (p < 0,01 et p < 0,05respectivement) (Fig. 10).

a)
b)
31
Figure 9 : Abondance des vers de terre à travers les
traitements; (a) densité et (b) biomasse.

2
b
b
b)
1,5
ab
a
a
1
0,5
0
Shannon Equitabilté
a)
M M+Mo M+Ha M+Mo+Ha M+U+SPP
Traitements
Figure 10: Diversité spécifique des vers de terre;
(a) richesse spécifique, (b) indice de Shannon et
l'Equitabilité.
32
3. Perception des paysans des effets des vers de terre
sur la fertilité du sol et sur la production des cultures
3.1. Connaissance générale des paysans sur
les vers de terre
Toutes les personnes interrogées (100%) disent que les
vers de terre sont beaucoup plus actifs pendant la saison pluvieuse et que leur
présence dans le sol est marquée par les dépôts de
turricules à la surface du sol. Au total, 77% de ces personnes
prétendent que les turricules sont nombreux pendant la saison des pluies
alors que c'est le contraire pour 23% d'entre elles. Selon une partie de ces
paysans (79%), il existe plusieurs types de turricules (petits ou gros, dures
ou friables) tandis que pour 16%, il n'y a qu'un seul type (gros). Les autres
(5%) n'ont eu d'avis à donner. Pour 79% de ces paysans, plusieurs
espèces de vers de terre existent dans la nature, 6% pensent qu'il
existe une seule espèce et le reste (15%) dit ne pas faire
d'observation. Ces paysans s'appuient sur les formes des turricules, la taille
et la couleur des vers de terre pour établir une différence entre
eux.
Tous les paysans interviewés laissent leurs parcelles
en jachère dans le but de rétablir la fertilité du sol
et/ou pour lutter contre les mauvaises herbes (adventices). Pour 55% de ces
paysans, les vers de terre dont leur présence est marquée par les
turricules, représentent les meilleurs indicateurs de la
fertilité du sol. Les autres (45%) pensent plutôt aux termites
et/ou aux fourmis.
3.2. Observations des paysans propriétaires des
parcelles sur les parcelles expérimentales
A la question de savoir dans lequel des traitements l'on
trouve d'abondants, moyennement abondants et peu abondants de turricules, 90%
des propriétaires de parcelles ont désigné le traitement
M+Ha pour le traitement ayant d'abondants turricules, 100% pour moyennement
abondants au niveau du traitement M+Mo+Ha, 100% pour peu abondants dans les
traitements M+Mo, témoin et M+U+SPP. De la même manière que
précédemment, 60% des personnes intérrogées ont
considéré le traitement M+Mo+Ha comme le meilleur (la meilleure
production de maïs). Une partie (20%) a désigné le
traitement M+Ha et les autres (20%), le traitement M+U+SPP. Ce qui fait que la
majorité des propriétaires de parcelles (70%) pense qu'il n'y a
pas de lien entre la meilleure production de maïs et l'abondance des
turricules. Cependant, 60% des paysans pensent que ce sont les vers de terre
qui sont responsables de l'augmentation de la production du maïs alors que
20% disent le contraire, le reste (20%) est sans avis.
33
3.3 Perception des paysans des effets des vers de terre sur
la fertilité du sol
La moitié des paysans (49,25%) affirme que
l'augmentation de la fertilité du sol est liée à la
présence des vers de terre alors que 23% pensent le contraire. Certains
(15,5%) prétendent que certains vers de terre augmentent cette
fertilité pendant que d'autres la réduisent. Pour 4,12 % de ces
personnes, ces organismes n'ont aucun effet sur la fertilité du sol. Les
paysans n'ayant pas d'avis à donner sur ce sujet occupent une proportion
de 8,12%. Il y a une différence hautement significative entre les points
de vue des groupes de paysans (p < 0,00001).
3.4 Perception des paysans des effets des vers de terre sur
le rendement des cultures
La proportion des paysans qui pensent que les activités
des vers provoquent une augmentation du rendement des cultures est de 57% alors
que 31% des paysans croient que les vers affectent négativement le
rendement des cultures. Seulement 4% prétendent qu'ils n'exercent aucun
effet et 8% n'ont pas d'avis sur cette question. En considérant les avis
des groupes de paysans, 90% des paysans propriétaires de parcelles
associés aux travaux et 55% des paysans de Goulikao reconnaissent
l'effet bénéfique des vers sur les rendements. Cependant, ceux
des deux campements sont beaucoup pessimistes sur ce point (45% à Petit
Bouaké et 38% à Djèkoffikro reconnaissent l'effet
bénéfique des vers). Le test Khi2 montre qu'il y a une
différence très significative entre les réponses des
paysans propriétaires de parcelle et celles des paysans de
Djèkoffikro et Goulikao (respectivement p < 0,000001 ; p <
0,000001), entre celles des paysans de Djèkoffikro et de Petit
Bouaké (p < 0,00001), entre les paysans de Goulikao et Petit
Bouaké (p = 0,0001) mais elle n'est pas significative entre les avis des
paysans de Goulikao et ceux des paysans de Djèkoffikro (p > 0,05)
(Tableau VII).
3.5 Avis des paysans sur l'utilisation des vers de terre
comme bio-fertilisants
Les paysans propriétaires de parcelles de maïs
sont tous unanimes sur le fait que les vers de terre améliorent la
production agricole alors que ceux n'ayant pas été directement en
contact avec les parcelles de démonstration (Goulikao,
Djékoffikro et Petit Bouaké) ont des réponses
nuancées de manière à révéler une
différence très significative entre les réponses des
propriétaires de parcelle et celles des paysans de Goulikao, de Petit
Bouaké et de Djèkoffikro (p < 0,000001 ; p < 0,000001 et p
< 0,00001 respectivement). Toutefois, à part la différence
significative observée entre les réponses des paysans de Goulikao
et celles de ceux de Petit Bouaké (p = 0,04), il n'y a pas de
différence significative entre les réponses des paysans de
Goulikao et Djèkoffikro (p > 0,05), de Djèkoffikro et Petit
Bouaké (p > 0,05) (Tableau VIII).
34
Tableau VII: avis des paysans de l'impact des vers de terre sur
le rendement des cultures. (Gkao : Goulikao, PBké : Petit Bouaké,
Dkk : Djé Koffikro)
|
Groupes de
paysans
|
élève
(%)
|
Diminue (%)
|
Aucun effet
(%)
|
Pas d'avis
(%)
|
Niveau de significativité entre les groupes (test
Khi2)
|
|
Contre Gkao
|
Contre PBke
|
Contre DKK
|
|
Pp
|
90
|
0
|
10
|
0
|
< 0,00001
|
< 0,0001
|
< 0,00001
|
|
Gkao
|
55
|
30
|
2,5
|
12,5
|
-
|
< 0,00001
|
< 0,0001
|
|
PBke
|
45
|
55
|
0
|
0
|
|
-
|
0,106
|
|
Dkk
|
38
|
38
|
4
|
20
|
|
|
-
|
Tableau VIII : Avis des paysans sur l'adoption ou non de
l'utilisation des vers de terre comme bio-fertilisants. Gkao : Goulikao,
PBké : Petit Bouaké, Dkk : Djé Koffikro
Niveau de significativité entre les groupes (test
Khi2)
|
Groupes de paysans
|
Avis positif (%)
|
Avis négatif (%)
|
Contre Gkao
|
Contre PBke
|
Contre Dkk
|
|
Pp
|
100
|
0
|
< 0,000001
|
< 0,000001
|
< 0,00001
|
|
Gkao
|
83
|
17
|
-
|
0,04
|
0,670
|
|
PBke
|
70
|
30
|
|
-
|
0,142
|
|
Dkk
|
80
|
20
|
|
|
-
|
.
35
V. DISCUSSION
1. Impact des vers de terre sur la croissance et la
production de maïs
L'expérimentation a montré que l'inoculation des
espèces M. omodeoi et/ou H. africanus et l'application
des engrais chimiques n'engendrent pas une augmentation significative de la
croissance du maïs. L'espèce H. africanus quant à
elle, a une influence positive sur la production de maïs. Cette production
est encore meilleure lorsque les deux espèces de vers de terre sont
associées, au point d'être supérieure à celle
obtenue avec ajout d'intrants inorganiques. En plus de l'amélioration de
la porosité et de l'agrégation du sol par les vers de terre
(Edwards & Lofty, 1980; Blanchart et al., 1999; Shipitalo & Le
Bayon, 2004), l'amélioration de la production de maïs pourrait
s'expliquer par une meilleure utilisation des éléments
minéraux du sol en présence des vers de terre (Gilot, 1994). Ces
derniers sont reconnus pour leur capacité à augmenter la
minéralisation de la matière organique du sol, rendant ainsi les
éléments nutritifs disponibles à la plante (Barois et
al., 1987; James, 1991; Lavelle et al., 1992; Subler et
al., 1997). L'efficience de l'utilisation de l'eau par le maïs est
également meilleure dans les traitements avec H. africanus et
avec l'association des deux espèces de vers de terre combinées.
Ceci est probablement une conséquence de l'amélioration des
propriétés physiques. Ces résultats sont conformes
à ceux de Ouédraogo et al. (2006) qui ont
rapporté une amélioration significative de l'EUE par le sorgho
grâce aux macro-invertébrés du sol.
D'après Lavelle et al., (1994) et Gilot
(1994), l'augmentation significative de la production du maïs survient en
milieu paysan lorsqu'une biomasse de vers de terre supérieure à
40 g m-2 est inoculée. Cette hypothèse semble
être confirmée pour H. africanus dans notre étude,
pas pour l'espèce M. omodeoi.
Sur les parcelles où M. omodeoi a
été inoculée seule, le maïs a connu la vitesse de
croissance la plus faible. L'EUE a également été
réduite. Ces résultats pourraient s'expliquer par la
réduction de la biomasse racinaire, elle-même occasionnée
par la compaction du sol sous l'effet de ce vers de terre. L'importante
quantité inoculée (90 g m-2) peut être à
l'origine de ce résultat contraire à ceux des travaux
antérieurs effectués avec cette espèce. En effet, Gilot
et al. (1994 et 1997) avait utilisé que 25 g.m-2. A
l'instar de Pontoscolex corethrurus responsable de la compaction des
sols en Amazonie brésilienne (Chauvel et al., 1999), M.
omodeoi, introduite en forte la biomasse a certainement contribué
à compacter le sol au point d'avoir un effet néfaste sur la
croissance du maïs. La dominance de vers de terre compactants dans un
milieu empêche une bonne croissance des racines à cause de la
réduction de la
36
porosité et de l'infiltration de l'eau du sol. Selon
Derouard et al. (1997) pour obtenir une structure de sol proche de la
réalité ou d'éviter la compaction du sol et donc
d'améliorer la production végétale, les vers de groupes
antagonistes devraient être associés. Ce qui est confirmé
dans notre étude
Selon Scheu et al. (1999), Wurst & Jones (2003)
et Kreuzer et al. (2004), la présence des vers de terre
occasionne une réduction du ratio biomasse épigée/racine.
Laossi et al. (2010) ont montré le contraire. Dans notre
étude, même si la différence entre les traitements n'est
pas significative, l'inoculation des vers de terre a provoqué une
réduction de ce ratio alors que l'application des engrais chimiques a eu
un effet contraire.
2. Impact de l'inoculation des populations de M. omodeoi et
H. africanus sur les communautés de vers de terre
A la fin des travaux, les biomasses des deux espèces
introduites, seule ou associées, ont drastiquement baissé. Le
même constat a été fait dans plusieurs études du
même genre (Gilot et al., 1996; Gilot, 1997 ; Subler et
al., 1997; Boyer et al.,1999; Muys et al., 2003; Chan
et al., 2004; Eriksen-Hamel & Whalen, 2007). La conservation des
vers dans du formol avant d'être pesés réduit de 25%
environ le poids de ces organismes (Senapati, 1980). Les mesures n'étant
pas corrigées, cela a dû causer, à un certain degré,
cette réduction de la biomasse des espèces introduites. En outre,
les vers ont été échantillonnés au début de
la saison sèche (mi-décembre), une période
défavorables au cours de laquelle certains vers vont en profondeur ou
entrent en quiescence et d'autres meurent. Par ailleurs, à part les
débris organiques laissés sur place après le
défrichage des parcelles, il n'y a pas eu d'apport de résidus
organiques utilisés comme sources de nourriture pour les vers de
terre.
Comparativement au témoin, l'augmentation de la
diversité du peuplement des vers de terre dans les trois traitements
à base de vers de terre pourrait s'expliquer par l'amélioration
du micro-environnement des parcelles. En effet, ces organismes ont la
capacité de créer des habitats et de réguler directement
ou indirectement les ressources pour d'autres organismes qui sont ici d'autres
espèces de vers, d'où le nom d'ingénieurs
d'écosystèmes (Jones et al., 1994, Lavelle et
al., 1997). Par contre, une réduction de la diversité et la
densité des vers a été observée au niveau du
traitement à base d'engrais chimiques. En effet, la conséquence
de l'application des engrais azotés est l'acidification du sol,
condition néfaste aux organismes du
37
sol (Bünemann et al., 2006). Au niveau des vers
de terre, l'acidification du sol est susceptible d'entraîner une baisse
de leur densité et de leur diversité (Edwards & Bohlen,
1996).
3. Perceptions des paysans par rapport aux effets des vers
de terre sur la fertilité du sol et la production agricole
Plus de la moitié des paysans considèrent les
vers de terre ou d'autres groupes d'organismes du sol comme indicateurs de la
qualité des sols. Par exemple, pour les Gban, la présence
abondante de turricules dans un milieu est synonyme de sols pauvres en raison
de l'effet compactant de certaines espèces dans les plantations de cacao
(Tondoh, non publié) . Au Cameroun, les paysans utilisent aussi ces
organismes comme un indicateurs de la qualité du sol (Birang et
al., 2003).
Il y a une divergence des avis sur l'effet des vers de terre
sur la fertilité du sol. Ces invertébrés, pour certains,
augmentent la fertilité du sol à travers l'amélioration de
la porosité du sol, la production de turricules qui enrichiraient le sol
lorsqu'ils s'érodent ou se fragmentent sous l'action de la pluie. La
conséquence de tout ceci serait que là où
l'activité de vers de terre est intense, le rendement des cultures
(maïs, riz, banane, etc.) est élevé. Pour d'autres, le taux
élevé de gros turricules à la surface du sol engendre une
compaction de ce dernier, réduisant ainsi sa fertilité, et
partant une réduction des rendements. Cette dernière observation
est conforme aux résultats que nous avons obtenus sur les parcelles
expérimentales et dans la littérature. Cet avis des paysans de
Goulikao est en contradiction avec ceux du Cameroun qui pensent que le niveau
élevé de turricules à la surface du sol est associé
à une augmentation du rendement des cultures (Birang et al.,
2003).
Moins de la moitié des paysans pensent que les vers
exercent des effets néfastes sur la fertilité du sol et la
production agricole. Il y a donc une évolution de la perception des
paysans de Goulikao des effets de ces organismes sur la fertilité et la
production agricole par rapport aux paysans du Cameroun où plus de 70%
des paysans les considèrent comme des pestes des plantes (Birang et
al., 2003). Le fait d'associer les paysans aux travaux sur l'introduction
des vers de terre en milieux paysans a été très
apprécié des paysans au point où tous les paysans qui ont
été associés aux travaux sur l'inoculation des vers de
terre au champ et plus de 80% des autres paysans sont d'accord pour
s'approprier cette technique agricole. Toutefois, la récolte ou la
production de la quantité nécessaire de vers de terre
représente une contrainte majeure à la promotion de cette
technologie.
38
CONCLUSION
Cette étude suggère que l'inoculation de M.
omodeoi (compactant) ou de H. africanus (décompactant)
seule ou en association ne favorise pas une croissance significative du
maïs. Cependant, la production du maïs et l'efficience de
l'utilisation de l'eau par cette plante sont meilleures lorsque les deux
espèces de vers de terre sont associées.
L'inoculation des vers de terre représente une
alternative sérieuse à l'utilisation d'intrants inorganiques qui
ne sont pas souvent accessibles aux petits paysans. Toutefois, une étude
poussée sur l'évaluation économique de cette technologie
semble incontournable avant toute promotion. Les paysans de Goulikao dans leur
majorité reconnaissent les effets bénéfiques des vers de
terre sur la fertilité du sol et sur le rendement des cultures et
prétendent être prêts à s'approprier la technique de
bio-fertilisation à base de vers de terre.
L'obtention des vers pour les inoculations étant
difficile, pour des futures études de ce genre, il faudrait songer
à établir des unités de production massive des vers de
terre ou favoriser la multiplication des espèces
préexistantes.
39
REFERENCES BIBLIOGRAPHIQUES
Angui P.K.T., Assié K.H., Djeke D.M., Tié B.T.,
2005. Morphological, physical and chemical characteristics of soil under
different land use systems in Center-West Côte d'Ivoire. Mid-term report.
CSM-BGBD Project. Phase 1: 21p.
Assié K.H., Angui K.T.P., Tamia A.J., 2008. Effets de la
mise en culture et des contraintes naturelles sur quelques
propriétés physiques d'un sol ferrallitique au centre ouest de la
Côte d'Ivoire: conséquences sur la dégradation des
sols. European Journal of Scientific Research Vol. 23, N°1,
149-166.
Ayanlaja S.A., Owa S.O., Adigun M.O., Senjobi B.A., Olaleye
A.O., 2001. Leachate from earthworm castings breaks seed dormancy and
preferentially promotes radicle growth in jute. HortScience 36,
143-144.
Baidai Y.D., 2009. Rôle du vers de terre Dichogaster
terrae nigrae (Acanthodrilidae) dans la modification du niveau de
compaction d'un sol dans une plantation de cacaoyers. Mémoire de
Maîtrise de l'Université d'Abobo-Adjamé, Côte
d'Ivoire. 28 p.
Barois I., Verdier B., Kaiser P., Mariotti A., Rangel P.,
Lavelle P., 1987. Influence of tropical earthworm Pontoscolex corethrurus
(Glossoscolecidae) on fixation and mineralization of nitrogen. In:
Bonvicini A. M., Omodeoo P. (Eds), On Earthworms. Mucchi, Bologna,
Italy pp 151-158.
Barot S., Ugolini A. Brikci F.B., 2007. Nutrient cycling
efficiency explains the long- term effect of ecosystem engineers on primary
production. Functional ecology 21, 1-10.
Barrion A.T., Litsinger J.A., 1997. Dichogaster nr.
Curgensis Michaelsen (Annelida: Octochaetidae): an earthworm pest of
terraced rice in the Philippine Cordilleras. Crop Protection 16-1,
89-93.
Blanchart E., 1990. Rôle des vers de terre dans la
formation et la conservation de la structure des sols de la savane de Lamto
(Côte d'Ivoire). Thèse de l'Université de Rennes I.
Blanchart E., Albrecht A., Alegre J., Duboisset A., Gilot C.,
Pashanasi B., Lavelle P., Brussard L., 1999. Effects of earthworms on soil
structure and physical properties. In Lavelle P., Brussard L., Hendrix P.
(Eds), Earthworm Management in Tropical Agroecosystems. CAB International,
Wallingford, pp 149-172.
Blanchart E., Lavelle P., Braudeau E., Le Bisonnais Y.,
Valentin C., 1997. Regulation of soil structure by geophagous earthworm
activities in humid savannas of Côte d'Ivoire. Soil Biology and
Biochemistry 29, 431-439.
40
Blouin M., Barot S., Lavelle P., 2006. Earthworms
(Millsonia anomala, Megascolecidae) do not increase rice growthly
through enhanced nitrogen mineralization. Soil Biology and Biochemistry
38, 2063-2068.
Bongoua A.J., 2002. Caractérisation de l'état de
fertilité des jachères plantées en légumineuses
fixatrices d'azote dans la région d'Oumé. Mémoire de DEA
de Pédologie, Université de Cocody-Abidjan, Côte d'Ivoire,
p 59.
Bosso A.M., 2010. Communautés de
macroinvertébrés comme indicateurs de la qualité du sol
en zone de forêt semi-decidue. Mémoire de DEA.
Université d'Abobo-Adjamé 29P. Bouché M.B., 1977.
Stratégies lombriciennes: Soils Organisms as Components of
Ecosystems. Ecol. Bull. 25, 122-132.
Boyer J., Michellon R., Chabanne A., Reversat G., Tibere R.,
1999. Effects of trefoil cover crop and earthworm inoculation on maize crop and
soils organisms in Reunion Island. Biology and Fertility of Soils 28,
364-370.
Brown G.G., 1995. How do earthworms affect microfloral and
faunal diversity? Plant and soil 170, 209-231.
Brown G.G., Edwards C.A., Brussard L., 2004. How earthworms
effect plant growth: burrowing into the mechanisms. Earthworm Ecology (Ed C.A.
Edwards), pp. 441. St Lucie Press, Boca Raton, Fl.
Brown G.G., Pashanasi B., Villenave C., Patron J.C., Senapati
B.K., Giri S., Barois I., Lavelle P., Blanchart E., Blakemore R.J., Spain A.V.,
Boyer J., 1999. Effects of earthworms on plant production in the tropics.
Earthworm Management in Tropical Agroecosystems (eds P. Lavelle, L.
Brussard & P. Hendrix), pp. 87-137 CABI Publishing, New York.
Brown S., Anderson J.M., Woomer P.L., Swift M.J., Barrios E.,
1994. Soil biological processes in tropical ecosystems. In The Biology
Management of tropical soil fertility (Woomer P.L., and Swift M.J., Eds),
pp. 15-45. John Wiley, Chichester.
Bünemann E.K., Schwenke G.D., Van Zwieten L., 2006.
Impacts of agricultural inputs on soil organisms - a review. Australian
Journal of Soil Research 44, 379-406.
Carpenter S.R., 1996. Microcosm experiments have limited
relevance for community and ecosystem ecology. Ecology 77, 677-680.
Chan K.Y., Baker G.H., Conyers M.K., Scott B., Munro K., 2004.
Complementary ability of three European earthworms (Lumbricidae) to bury lime
and increase pasture production in acidic soils of south eastern Australia.
Applied Soil Ecology 26, 257271.
41
Chauvel A., Grimaldi M., Barros E., Blanchart E., Sarrazin M.,
Lavelle P., 1999. Pasture degradation by an Amazonian earthworm. Nature
389, 32-33.
Csuzdi C., Tondoh E.J., 2007. New and little known earthworm
species from Ivory Coast (Oligochaeta: Acanthodrilidae: Benhamiinae and
Eudrilidae). J. Nat. Hist. 41, (41) 2551-2567.
Darwin C., 1881. The formation of vegetable mould through the
action of worms with observation of their habits. Murray, London, 158 p.
Dash M. C., Saxena K. G., Giri S., 2009. Vermitechnology for
wasteland reclamation, plant productivity and composting: A review in Indian
context. International Journal of Ecology and Environmental Sciences
35 (2-3), 163-185.
Dell'Agnola G., Nardi S., 1987. An overview of earthworm
activity in the soil. In: Bonvicini Pagliai A.M., Omodeo P. (Eds), On
Earthworms. Mucchi, Modena, PP. 103-112.
Derouard L., Tondoh J., Vilcosqui L., Lavelle P., 1997.
Effects of earthworm introduction on soil processes and plant growth. Soil
Biology and Biochemistry Vol. 29 3/4. 541-547.
Doube B.M., Williams P.M.J.,Willmott P.J., 1997. The influence
of two species of earthworm (Aporrectodea trapezoides and
Aporrectodea rosea) on the growth of wheat, barley and faba beans in
the three soiltyupes in the greenhouse. Soil Biology and Biochemistry
29, 503-509.
Edwards C.A., 2004. Earthworm ecology. CRC, Boca Raton, p
441
Edwards C.A., Bohlen P.J., 1996. Biology and Ecology of
earthworms. Chapman and Hall, London, 426 p.
Edwards C.A., Lofty J. R., 1980. Effects of earthworm
inoculation upon the root growth of direct drilled cereals. Journal of
Applied Ecology 17, 533-543.
Eriksen-Hamel N.S., Whalen J.K., 2007. Impacts of earthworms
on soil nutrients and plant growth in soybean and maize agroecosystems.
Agriculture Ecosystems and Environment 120, 442-448.
Gilot C., 1994. Effets de l'introduction du vers
géophage tropical Millsonia anomala omodeoi en systèmes
cultivés sur les caractéristiques des sols et la
productivité végétale en moyenne Côte d'Ivoire.
Thèse de l'Université, Institut National agronomique de
Paris-Grignon.
Gilot C., 1997. Effects of a tropical geophageous earthworm,
M. anomala (Megascolicidae), on soil characteristics and production of
a yam crop in Ivory Coast. Soil Biology and Biochemistry 29 (3/4),
353-359.
42
Gilot C., Lavelle P., Ganry F., 1996. Effects of a tropical
geophageous earthworm, Millsonia anomala (Megascolicidae), on some
soil characteristics, on maize-residue decomposition and on maize production in
Ivory Coast. Applied Soil Ecology 4, 201211.
Gohoré Bi G.L., 2007. Savoirs paysans et qualité
des sols: cas du terroir Goulikao (Oumé, Côte d'Ivoire).
Mémoire de DUT, Université de Bouaké, URES de Kohrogo. 21
p.
Guéi A.M., 2006. Variation interannuelle des
peuplements de vers de terre le long d'un gradient de perturbation des
forêts en Côte d'Ivoire. Mémoire de DEA de
l'Université d'Abobo-Adjamé, Côte d'Ivoire. 25 P.
Haynes R. J., Fraser P. M., Tregurtha R. J., Piercy J. E.,
1999. Size and activity of the microbial biomass and N, S and P availability in
earthworm casts derivied from arable and pastoral soil and arable soilamended
with plant residues. Pedobiologia 43, 568573.
James S.W., 1991. Soil nitrogen, phosphorus and organic matter
processing by earthworms in tallgrass prairie. Ecology 72,
2101-2109.
Jones C.G., Lawton J.H., Shachak M., 1994. Organisms as
ecosystem engineers. Oikos 69, 373-386.
Joshi R.C., Matchoc O.R.O., Cabigat J.C., James S.W., 1999.
Survey of earthworm in the Ifugao rice terraces, Philippines. Journal of
Environmental Science and Management 2, 1-12.
Kang B.T., 1997. Les engrais : définitions et calculs.
Guide de phytotechnie de l'IITA, n°1. Ibadan, Nigeria: Internationale
Institutes of Tropicale Agriculture (IITA).
Koné A.W., 2009. Qualité des sols en zone de
savane humide de Côte d'Ivoire: utilisation de légumineuses
herbacées comme alternative pour une valorisation des terres marginales
et une agriculture durable. Thèse de Doctorat Unique, Université
d'Abobo-Adjamé. 194 p.
Kreuzer K., Bonkowski M., Langel R., Scheu S., 2004.
Decomposer animals (Lumbricidae, Collembola) and organic matter distribution
affect the performance of Lolium Perenne (Poacae) and Trifolium
repens (Fabacae). Soil Biology and Biochemistry 36,
20052011.
Laossi K.R., Noguera D.C., Bartolomé-Lasa A., Mathieu
J., Blouin M., Barot S., 2009. Effects of an endogeic and an anecic earthworm
on the competition between four plants and their reletive fecundity. Soil
Biology and Biochemistry 41, 1668-1673.
43
Laossi K.R., Ginot A., Noguera D.C., Blouin M., Barot S.,
2010. Earthworm effects on plant growth do not necessarily decrease with soil
fertility. Plant and soil 328, 109-118.
Lavelle P., 1978. Les vers de terre de la savane de Lamto
(Côte d'Ivoire) : peuplements, populations et fonction dans
l'écosystème. PhD, dissertation,Université Paris VI, Publ
Lab Zool Ecole Normale Supérieure, 12 : 30 P.
Lavelle P., 1981. Stratégies de reproduction chez les
vers de terre. Acta OEcologia/ OEcol.Gener. Vol. 2, n°2,
117-133.
Lavelle P., Barois I., Martin A., Zaidiz Z., Schaefer R.,
1989. Management of earthworm populations in agrosystems: A possible way to
maintain soil quality? In: Clarholm M., Bergström L. (eds ) Ecology of
arable land. Kluwer Acdemic Publishers pp 109-122.
Lavelle P., Bignell D., Lepage M., Wolters V., Roger P.,
Ineson P., Heal O.W., Dhillon S., 1997. Soil function in changing world: the
role of invertebrate ecosystem engineers. European Journal of Soil Biology
33(4), 159-193.
Lavelle P., Decaëns T., Aubert M., Barot S., Blouin M.,
Bureau F., Margerie P., Mora P., Rossi J. P., 2006. Soil invertebrates and
ecosystem services. European Journal of Soil Biology 42, S3-S15.
Lavelle P., Gilot C., Fragoso C., Pashanasi B., 1994. Soil
fauna and sustainable land use in the humid tropics. In Soil Resilience and
Sustainable Land Use (I. Szaboles and D. Greenland, Eds), pp. 291-308. CAB
International, Wallingford, UK.
Lavelle P., Melendez G., Pashanasi B., Schaefer R., 1992.
Nitrogen mineralization and reorganization in casts of the geophagous tropical
earthworm Pontoscolex corethrurus (Glossoscolecidae). Biology and
Fertility of soils 14, 49-53.
Lavelle P., Pashanasi B., Charpentier F., Gilot C., Rossi
J.-P., Derouard L., Andre J., Ponge J-F., Bernier N., 1998. Large-scale effects
of earthworms on soil organic matter and nutrient dynamics, in: C.A. Edwards
(Ed.), Earthworm Ecology, CRC press, Boca Raton, FL, pp. 103-122.
Lecomte P., 1990. Place et intégration de l'arbre dans
l'exploitation agricole ivoirienne du centre ouest. Cas de la région
d'Oumé. Mémoire de fin de d'étude CNEARC. Montpellier,
France, 109 p.
Lee K., 1985. Earthworm: their ecology and relationships with
soil and land use. Academic Press, London. 411 p.
Madge D.S., 1969. Fieldand laboratories studies on the
activities of two species of tropical earthworms. Pedobiologia 9,
188-214.
44
Martin A., 1991. Short- and long-term effects of the endogeic
earthworm Millsonia anomala (Omodeo) (Megascoolecidae, Oligochaeta) of
tropical savannas on soil organic matter. Biology and Fertility of Soils
11, 234-238.
Monin M.L., 2005. Peuplements de vers de terre le long d'un
gradient d'utilisation des sols en zone de forêt semi décidue
d'Oumé (Côte D'Ivoire): abondance et diversité.
Mémoire de DEA. Université d'Abobo-Adjamé. 46 p.
Monnier Y., 1983. Les sols. In Atlas Côte
d'Ivoire, 2e Edition, Jeune Afrique, Paris, pp 2021.
Muscolo A., Bovalo F., Gionfriddo F., Nardi S., 1999.
Earthworm humic matter produces auxin-like effects on daucus carota cell growth
and nitrate metabolism. Soil Biology and Biochemistry 31,
1303-1311.
Muys B., Beckers G., Nachtergale L., Lust N; Merckx R.,
Granval P., 2003. Medium-term evaluation of a forest soil restoration trial
combining tree species change, fertilisation and earthworm introduction.
Pedobiologia 47, 772-783.
Nardi S., Pizzeghello D., Muscolo A., Vaniello A., 2002.
Physiological effects of humic substances on higher plants. Soil Biology
and Biochemistry 34, 1527-1536.
Omodeo P., 1954. Eudrilidae e octochaetina delle Costa
d'Ovorio (Oligochaeta). Momorie del Museo Civico di Storia Naturale di
Verona 4, 162-166.
Ouédraogo E., Mando A., Brussaard L., 2006. Soil
macrofauna affect crop and water use efficiencies in semi-arid West Africa.
European journal of soil biology 42, S275-S277.
Pashanasi B., Lavelle P., Alegre J., Charpentier F., 1996.
Effect of the endogeic earthworm Pontoscolex corethrurus on soil
chemical characteristics and plant growth in a low-input tropical agrosysterm.
Soil Biology and Biochemistry 28, 801-810.
Perraud A., 1971. Les sols. In le milieu naturel de
la Côte d'Ivoire. Mémoire ORSTOM N°50, ORSTOM, Paris, France.
PP 265-391.
Rossi J.P., 2003. The spatiotemporal pattern of a tropical
earthworm species assemblage and its relationship with soil structure.
Pedobiologia (Jena) 47, 497-503.
Scheu S, Theenhaus A., Jones H.T., 1999. Links between the
detritivore and the herbivore system: effects of earthworm and collembolan on
plant growth and aphid development. Oecologia 119, 541-551.
Scheu S., 2003. Effects of earthworm on plant growth: patterns
and perspectives. Pedobiologia 47, 846-856.
45
Senapati B.K., 1980. Aspects of ecophysiological studies on
tropical earthworms (Distribution, Populations dynamics, Production, Energetics
and their role in the decomposition process). Ph.D, Sambalpur.
Senapati B.K., Lavelle P., Giri S., Pashanasi B., Alegre J.
Decaëns T., Jiménez J.J., Albrecht A., Blanchart E., Mahieux M.,
Rousseaux L., Thomas R., Panigrahi P.K., Venkatachalan M., 1999. In-soil
technologies for tropical ecosystems. In Earthworm management in tropical
agroecosystems, eds Lavelle P., Brussaard L. and Hendrix P.F., pp. 199-237.
CAB International, Wallingford.
Senapati B.K., Lavelle P., Panigrahi P.K., Gri S., Brown G.G.,
2002. Restoring soil fertility and enhancing productivity in Indian tea
planatations with earthworms and organic fertilizers. International Technical
Workshop on Biological Management of Soil ecosystems for Sustainable
Agriculture, Brazilian Agril. Research Corp, Brazil. 182, 172-190.
Sheehan C., Kirwan L., Connolly J., Bolger T., 2006. The
effects of earthworm functional group diversity on nitrogen dynamics in soils.
Soil Biology and Biochemistry 38, 2629-2636.
Sheehan C., Kirwan L., Connolly J., Bolger T., 2007. The
effects of earthworm functional group diversity on earthworm community
structure. Pedobiologia 50, 479-491.
Shipitalo M.J., Le Bayon R.C., 2004. Quantifying the effects
of earthworm on aggregation and porosity. In: Edwards C. A. (Ed), Earthworm
Ecology. CRC Press, Boca Raton p. 441.
Spain A.V., Lavelle P., Mariotti A., 1992. Stimulation of
plant growrh by tropical earthworms. Soil Biology and Biochemistry 24,
1629-1633.
Subler S., Baranski C.M., Edwards C.A., 1997. Earthworm
additions increased short-term nitrogen availability and leaching in two grains
crop agroecosystems. Soil Biology and Biochemistry 29, 413-421.
Swift M.J., Izac A.M.N, Van Noordwijk M., 2004. Biodiversity
and ecosystem services in agricultural landscape: Are we asking the right
questions? Agriculture Ecosystems and Environment 104, 113-134.
Tilman D., 1999. Global environmental impacts of agricultural
expansion: the need for sustainable and efficient practises. Proc. Natl.
Acad. Sci. USA 96, 5995-6000.
Tondoh E. J., 1998. Démographie et fonctionnement des
populations de vers de terre Hyperiodrilus africanus (Eudrilidae) dans
une savane protégée du feu en moyenne Côte d'Ivoire.
Thèse de Doctorat Unique, Université Paris 6, 209 p.
46
Tondoh E. J., Guéi A. M., Csuzdi C., Okoth P., 2011.
Effect of land-use on the earthworm assemblages in semi-deciduous forests of
Central-West Ivory Coast. Biodivers conserv 20 : 169-184.
Tondoh E.J., Konaté S., 2005. Diversité et
rôles écologiques des vers de terre en Côte d'Ivoire.
Bioterre Rev. Inter. Sci. de la vie et de la terre Vol.5 N°1,
90-108.
Tondoh E. J., Lavelle P., 1997. Effect of two organic residues on
growth and cocoon production of the earthworm Hyperiodrilus africanus
(Eudrilidae). European Journal of Soil Biology 33: 13-18.
Tondoh E.J, Lavelle P., 2005. Population dynamics of
Hyperiodrilus africanus (Oligochaeta, Eudrilidae) in Ivory Coast.
Journal of Tropical Ecology 21: 1-8.
Tondoh E.J., Monin M.L., Tiho S., Csuzdi C., 2007. Can
earthworm be used as bioindicators of land-use perturbation in semi-deciduous
forest? Biology and Fertility of Soils 43, 584-592.
Wurst S., Jones H.T., 2003. Indirect effects of earthworms
(Aporrectodea caliginosa) on an above-ground interaction.
Pedobiologia 47, 91-97.

A) Collecte de vers de terre B) Hyperiodrilus
africanus
C) Millsonia omodeoi D) Introduction des vers
au champ
D) Maïs + Millsonia omodeoi
F) Maïs + Hyperiodrilus africanus
ANNEXES
Annexe 1
Récolte de vers de terre, spécimens de vers
de terre, inoculation de vers de terre et des
exemples de traitements
Annexe 2
Mesure de la hauteur du plant du maïs

Hauteur du plant
Annexe 3
Densité moyenne (ind.m-2) des vers de
terre par traitement en fin d'étude
|
Espèces
|
Traitements
|
|
|
|
|
|
M
|
M+Mo
|
M+Ha
|
M+Mo+Ha
|
M+U+SPP
|
|
M. omodeoi
|
13,60#177;3,78
|
69,60#177;20,90
|
12,80 #177; 4,95
|
32,80 #177; 5,52
|
6,40 #177; 3,33
|
|
H .africanus
|
6,40 #177; 3,33
|
8,00 #177; 3,96
|
28,80 #177; 7,27
|
32,00 #177; 7,82
|
4,00 #177; 2,15
|
|
D. baeri
|
5,60 #177; 3,38
|
5,60 #177; 2,08
|
21,60 #177; 6,75
|
12,80 #177; 5,23
|
5.60 #177; 4.14
|
|
D. terrae nigrae
|
4,80 #177; 2,72
|
20,80#177;13,97
|
14,40 #177; 7,52
|
11,20 #177; 5,99
|
4,00 #177; 2,73
|
|
D. saliens
|
0,80 #177; 0,80
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
|
D. ehrhardti
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,80 #177; 0,80
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
|
D. papillosa
|
0,00 #177; 0,00
|
3,20 #177; 3,20
|
3,20 #177; 2,44
|
0,80 #177; 0,80
|
0,00 #177; 0,00
|
|
D. mamillata
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,80 #177; 0,80
|
0,00 #177; 0,00
|
|
Dichogaster. sp
|
5,60 #177; 2,68
|
13,60 #177; 5,60
|
11,20 #177; 3,99
|
9,60 #177; 2,00
|
4,80 #177; 3,41
|
|
A.multivesiculatus
|
0,80 #177; 0,80
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
|
A. opisthogynus
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,80 #177; 0,80
|
0.00 #177; 0,00
|
0,00 #177; 0,00
|
|
M. lamtoiana
|
0,00 #177; 0,00
|
0,80 #177; 0,80
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,80 #177; 0,80
|
|
S. zielae
|
9,60 #177; 6,06
|
23,20#177;22,33
|
13,60#177;11,19
|
7,20 #177; 6,36
|
10,40 #177; 4,63
|
Annexe 4
Biomasse moyenne (g.m-2) des vers de terre
par traitement en fin d'étude
|
Espèces
|
Traitements
|
|
|
|
|
|
M
|
M+Mo
|
M+Ha
|
M+Mo+Ha
|
M+U+SPP
|
|
M. omodeoi
|
5,63 #177; 3,06
|
35,68#177;12,91
|
2,40 #177; 1,12
|
14,61 #177; 2,46
|
2,28 #177; 1,40
|
|
H. africanus
|
1,49 #177; 0,92
|
1,64 #177; 0,75
|
4,52 #177; 1,31
|
7,10 #177; 1,63
|
0,96 #177; 0,52
|
|
D. baeri
|
0,69 #177; 0,44
|
0,74 #177; 0,28
|
1,87 #177; 0,79
|
1,56 #177; 0,68
|
0,71 #177; 0,54
|
|
D. terrae nigrae
|
4,93 #177; 3,07
|
16,06#177;11,59
|
10,93 #177; 7,97
|
10,64 #177; 5,32
|
2,78 #177; 1,93
|
|
D. saliens
|
0,01 #177; 0,01
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
|
D. ehrhardti
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,02 #177; 0,02
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
|
D. papillosa
|
0,00 #177; 0,00
|
0,06 #177; 0,06
|
0,04 #177; 0,03
|
0,03 #177; 0,03
|
0,00 #177; 0,00
|
|
D. mamillata
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,04 #177; 0,04
|
0,00 #177; 0,00
|
|
Dichogaster sp
|
0,10 #177; 0,05
|
17,63#177;17,41
|
0,22 #177; 0,11
|
0,17 #177; 0,04
|
0,06 #177; 0,04
|
|
A.multivesiculatus
|
0,18 #177; 0,18
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
|
A. opisthogynus
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,52 #177; 0,52
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
|
M. lamtoiana
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
0,00 #177; 0,00
|
7,89 #177; 7,89
|
|
S. zielae
|
0,23 #177; 0,16
|
0,47 #177; 0,45
|
0,33 #177; 0,31
|
0,16 #177; 0,14
|
0,29 #177; 0,17
|
NB : La question n° 7 concerne exclusivement les
propriétaires de parcelles qui ont été associés aux
travaux de manipulations de vers de terre en milieux paysans.
Annexe 5
Questionnaire :
Nom prénoms âge sexe ethnie cultures
vivrières
1) Avez-vous déjà vu un ver de terre ?
2) Pendant quelle saison de l'année les vers de terre sont
abondants dans le sol ?
3) Quels sont les structures produites par les vers de terre et
qui signalent leur présence dans le sol ?
4) Pendant quelle saison de l'année les turricules des
vers de terre sont abondants dans le sol ?
5) Tous les turricules ont-ils le même aspect ?
6) Selon vous, ces différents types de turricules sont
produits par la même espèce de ver de terre ?
7) Quelles sont les parcelles ("parcelles paysannes") dans
lesquelles les turricules des vers de terre sont abondants, moyennement
abondants, peu abondants, absents ?
7a) Selon vous, existe-il un lien entre l'abondance des
turricules et la production de maïs ?
7b) Quelle est la parcelle dans laquelle de maïs qui la
meilleure production ?
7c) Pensez vous que les vers de terre sont responsables de cette
production épigée ?
8) Pourquoi pratiquez vous souvent la mise en jachère de
vos parcelles ?
9) Quels organismes (vers de terre, fourmis, termites...) ou
structure biogénique (turricules, termitière...) sont abondants
dans les parcelles en jachère ?
10) Lequel de ces organismes pensez-vous être indicateur de
la fertilité des sols ?
11) Les vers de terre affectent-ils la fertilité des sols
(dans le sens d'une augmentation ou d'une diminution)?
12) Pensez vous que les vers de terre agissent sur le rendement
des cultures (maïs, cacao, arachides, riz, banane, manioc (augmentation,
diminution, aucun effet)?
13) Etes vous prêts à utiliser les vers de terre
comme engrais biologique dans vos champs ?
a) Si oui, comment compter vous le faire ;
b) Si non, pourquoi ?
|
|


