|

REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE
L'ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE
UNIVERSITE CHEIKH LARBI TEBESSI DE TEBESSA
oie
)1'
INSTITUT DES SCIENCES DE LA VIE ET DE LA NATURE
EPARTE DES SCIENCES DE IA ET DE L'
TERRITOIRE
MENT DU
Mémoire présenté en vue de l'obtention du
diplôme de
MAGISTER
41" Spécialité : Géologie

Biostratigraphie et Paléoenvironnement du
Crétacé moyen
des Hameimats à partir de l'étude
de la
faune et de la microfaune - Coupe du Djebel
Chemla
(Morsott, NE Al érie .
Soutenu publiquement le 23 novembre 2008
MT BOLTMEZBEUR Abderrahmane, Maître de Conférences
(Tébessa) Président «J-
MT HERKAT Missoum, Professeur (USTHB, Alger)
ii Examinateur ',
Mme SALMI-LAOUAR Sihem, Maître de Conférences
(Annaba) )Examinateur
Mme KECHID-BENKHEROUF Fatiha, Maître
de Conférences (USTHB, Alger) Rapporteur
Option : Géologie des formations sédimentaires
REMERCIEMENTS
Je tiens à remercier tous ceux qui, de près ou
de loin, m'ont aidé lors de la réalisation de ce Magister, et en
premier lieu, mon encadreur, Mme F. KECHID-BENKHEROUF, qui m'a apporté
une aide précieuse dès qu'elle le pouvait.
Je remercie également mon mari, Mr A. DJERRAB, pour son
soutien sans faille et son aide de tous les instants ; ainsi que tous les
chercheurs que j'ai sollicités, et qui ont accepté de m'aider,
que ce soit en me faisant parvenir des documents, toujours utiles et
précieux, ou en m'épaulant pour la détermination des
fossiles. Que soient ainsi remerciés :
Mme M. CARON (Fribourg, Suisse) et Mr R. WERNLI (Genève,
Suisse), tous deux
spécialistes en foraminifères et qui m'ont
gracieusement offert des documents,
Mr H. GEBHARDT (Vienne, Autriche), et surtout Mr B. ANDREU
(Toulouse, France), qui m'ont aidé pour la détermination des
ostracodes et m'ont communiqué des documents,
Mr F.BAUDIN (Paris, France), qui a accepté de
réaliser dans son laboratoire des analyses sur certains de mes
échantillons,
Mr T. SAOS (Tautavel, France) qui a réalisé
quelques clichés au MEB au sein du CERP de Tautavel (Université
de Perpignan).
Je suis bien entendu très reconnaissante envers les
membres de mon jury, qui ont accepté de juger mon travail : Mr M. HERKAT
(Alger), Mme S. SALMI-LAOUAR (Annaba) et Mr A. BOUMEZBEUR (Tébessa).
Enfin, merci à tous ceux qui m'ont côtoyé
au Centre Universitaire de Tébessa, et notamment les techniciens du
laboratoire de Géologie, grâce à qui j'ai pu travailler
dans de bonnes conditions, ainsi que mes collègues de Magister, avec qui
j'ai eu, pour certains, des échanges fructueux.
RÉSUME
L'étude paléontologique de formations
sédimentaires datées du Crétacé moyen
(essentiellement du Cénomanien) de la région de Tébessa,
dans l'est algérien, a permis la récolte d'une centaine
d'échantillons, et parmi eux de nombreux fossiles (surtout des bivalves,
gastéropodes et échinodermes). La plupart des
échantillons, marneux, ont été lavés au travers de
deux tamis. Le tri et le comptage des microfossiles récoltés dans
le résidu fin a permis la mise en oeuvre d'études statistiques,
tant sur les populations d'ostracodes que de foraminifères. Par
ailleurs, des lames minces ont été confectionnées dans
quelques échantillons indurés.
Les deux objectifs clairement définis ont
été d'une part de déterminer la biostratigraphie des
niveaux, et d'autre part de préciser leur milieu de dépôt
(paléoenvironnement).
Trois zones ont ainsi été reconnues au cours du
Cénomanien (zone à Rotalipora brotzeni, à R. cushmani
et à Whiteinella archeocretacea).
L'évènement anoxique majeur marquant la fin du
Cénomanien (l'OAE2) a également bien été
caractérisé.
L'étude paléoenvironnementale
révèle un milieu de dépôt marin, plutôt calme
et relativement profond, de type plate-forme externe (-- 200 m). Les conditions
sur le fond semblent avoir été fréquemment
caractérisées par un manque plus ou moins prononcé
d'oxygène. L'étude révèle cependant une certaine
hétérogénéité de ces conditions,
marquée par une grande variabilité des assemblages
(foraminifères et ostracodes) : se succèdent ainsi quatre cycles
de dépôt.
Malgré quelques imprécisions, notamment dans la
détermination des (micro)fossiles, cette étude a donc globalement
bien atteint ses objectifs.
MOTS CLES : Cénomanien -- Algérie
-- Foraminifère -- Ostracode -- OAE2.
4c. 100
`_;à1.:».113.
4:"x1911 c4...:; Zijaa.,11".44:4fill
*À÷ày.,,
`:J
·134:111`:à
·le*-1-.11:Lul.
41)1
9~ .a11çrà
`:à
· 441 41>A v mil `:à*.11
Sal.(`:à
· 43.111 j
`:à
·Lià J.:)L.11 *.L.e.1A)
`:à
·te*-1..A 41, 41,
e,Let)"441.4cv1 .a1.1.4 je. 4.Là..11.39
Id 1...L 4.52 Zjà
jt,11
j9i6eti1 çrà
`:à
· 441 eL.41.4 c31.).1*
.(`:à
· LLA:u 9 `:à
·
141:5Luil ) *À,LA÷.411 `:à
·Lu1111
·
· .14 4_3-1.) `Làtii+U.4-x+0=
Zul J.111 bl+1aut-...11 ae..4
.04.)41:j1._ei5.1.4.,b
rt..e.o
*À÷à:I.A.j..4.
" (zone à Rotalipora brotzeni,
à R. cushmani et à Whiteinella
archeocretacea)."
,0111 j..à.fi11
.5.; Lek. ,0111 sJJail *Z
1. à )14 *Àii1:1...11 "41,11 Ls.k.
Là."111
. (0AE2)40-11-.
4,e_)1- 415 t.93 (DA .4-"3
CiAle
· C°95
di°4-n÷11 4..à11
`1,1,?m 4eJ tta
Ls.c u.,rem cs9jà
.`,.11sJI.. 200
(:21-.15 495
LD1- (1).° `e39
`".119.)1.1 " (12.14
4÷ (A
j)elv`f+1. 9S1j1..,
c.11111 `J*1_11 (.>:cipq tià
.41A ilij.411
Là1àtàti1
.(OAE2)
«:J.I.LA.u11`.):11_».11
ABSTRACT
Approximately 100 samples have been collected
essentially on Cenomanian levels in the area of Tébessa (east Algeria).
A number of fossils (bivalve, gastropod, echinoderm...) have also been found.
Our samples, mainly consisting of marls, have been washed through two sieves.
Then, we have performed statistical studies by sorting out and counting
microfossils (ostracods and foraminifera).
At the beginning, two objectives were defined, that is
to clarify biostratigraphy and palaeoenvironment.
· Biostratigraphy : three zones have been
distinguished during Cenomanian times (Rotalipora brotzeni, R.
cushmani and Whiteinella archeocretacea
zones). Oceanic anoxic event (OAE) of terminal Cenomanian has also
been clearly identified.
· Palaeoenvironment : our study reveal a
peaceful marine environment, quite deep (about 200 m), probably located on
distal ramp. Anoxic conditions seem to have been quite frequent. However those
conditions also seem highly heterogeneous, indeed microfossilistic asssemblages
are extremely variable. As a result we have differentiate four deposit
sequences.
KEY-WORDS : Cenomanian - Algeria - Foraminifera -
Ostracod - OAE2.
TABLE DES MATIERES
INTRODUCTION 1
I - Généralités 2
1. Contexte géographique 2
2. Contexte géologique 3
3. Présentation du site d'étude 5
4. Précision sur les terrains étudiés 6
II- Matériel et méthodes 8
1. Description des prélèvements 8
2. Méthodes utilisées 8
a- Calcimétrie 8
b- Dosage du carbone organique total (COT) 8
c- Traitement des échantillons meubles ou faiblement
indurés 8
d- Paramètres déterminés à l'issue
du tri 9
III -- Résultats et interprétations 12
1. Généralités 12
a- Historique des travaux existants 12
b- Aperçu général sur le monde au
Crétacé moyen 12
c- Le log stratigraphique 14
2. Résultats 16
a- Les fossiles 16
b- Aspects minéralogiques et géochimiques 21
c- Résultats du tri des microfossiles 23
c-1 -- Les ostracodes 24
c-2 -- Les foraminifères 29
· Ratio benthique /planctonique 29
· Evolution de la diversité spécifique
29
· Richesse microfossilistique apparente 29
· Foraminifères planctoniques : variation et
caractérisation 30
· Foraminifères benthiques 32
d- Etude des lames minces 45
3. Interprétations 52
a- Axe Biostratigraphie 52
b- Axe Paléoenvironnement / Paléogéographie
54
· Etude des microfossiles
· 54
· Caractéristiques générales et
implications paléoenvironnementales : 55
· Comment
l'hétérogénéité des résultats
observée mène à une interprétation
d'ordre séquentiel
· 57
c- La crise cénomano-turonienne 63
c-1 - Généralités 63
c-2- L'OAE2 sur la coupe du Dj. Chemla 64
d- Le passage Vraconnien-Cénomanien 66
e- Questions en suspens 67
CONCLUSION et PERSPECTIVES 69
BIBLIOGRAPHIE 71
1. Auteurs cités dans le texte 71
2. Auteurs cités dans les parties concernant la
systématique 75
LISTE des TABLEAUX, PLANCHES et
FIGURES
Tableau 1 : Type de microfossiles observés 10
Tableau 2 : Détail de la répartition et de
l'abondance des foraminifères benthiques. 40
Tableau 3 : Résumé des caractéristiques
générales de la coupe étudiée. 56
Planche 1 : Bivalves (ostréidés). 19
Planche 2 : Bivalves (A à F) et Gastéropodes (G
à H) 20
Planche 3 : Oursins (A, C) et Ammonites (B, D) 21
Planche 4: Ostracodes. 27
Planche 5 : Foraminifères benthiques agglutinés.
34
Planche 6 : Foraminifères benthiques calcaires. 37
Planche 7 : Foraminifères benthiques calcaires (suite).
38
Planche 8 : Illustrations de quelques microfossiles au MEB. 42
Planche 9 : Lames minces (Vraconnien -- Cénomanien moyen)
47
Planche 10 : Lames minces (Cénomanien sup.) 49
Planche 11 : Lames minces (Cénomanien sup. -- Turonien
inf.) 51
Figure 1 : Localisation géographique du site. 2
Figure 2 : Schéma des unités structurales d'Afrique
du Nord 3
Figure 3 : Carte géologique de la région de Morsott
(d'après la carte géologique de Morsott au
1/50 000) et localisation de la coupe étudiée. 4
Figure 4 : Coupe du Djebel Chemla. 5
Figure 5 : Echelle stratigraphique des temps géologiques
(datations d'après ODIN et ODIN,
1990) 7
Figure 6 : Relation entre profondeur du milieu et type de
foraminifères présents 11
Figure 7 : Carte du monde au Cénomanien, il y a 94 Ma.
13
Figure 8 : Log stratigraphique de la coupe du Dj. Chemla. 15
Figure 9 : Biozonation du Crétacé moyen par les
Ammonites et les Foraminifères
planctoniques 18
Figure 10 : Evolution du taux de carbone organique total (COT).
23
Figure 11 : Etages de peuplement et milieux de dépôt
(modifié d'après Andreu, 1991) 24
Figure 12 : Log stratigraphique et paramètres de tri.
28
Figure 13 : Clés de détermination des principaux
genres de foraminifères planctoniques
rencontrés. 31
Figure 14 : Log stratigraphique et paramètres de tri
(suite). 44
Figure 15 : Foraminifères planctoniques --
Répartition de quelques espèces-clés. 52
Figure 16 : Biozonation du Crétacé moyen à
supérieur par les foraminifères planctoniques 53
Figure 17 : Variations du contenu micropaléontologique en
domaine marin exprimées par
différents rapports (d'après Rey, 1983 in
Andreu, 1991). 54
Figure 18 : Présentation des principaux rapports de tri et
des zones définies dans la coupe 58
Figure 19 : Séquences de dépôt et variations
eustatiques au cours du Cénomanien 60
Figure 20 : Reconstitution paléogéographique du
domaine est-atlasique au cénomano-turonien
(modifié
d'après Herkat et Kechid-Benkherouf, 2006). 62
Introduction
INTRODUCTION
L'étude menée dans le cadre ce Magister n'est
que le début d'un travail de recherche qui se poursuivra par une
thèse de doctorat. Axé sur l'étude du contenu
(micro)paléontologique de formations du Crétacé moyen
(Albien sup. -- Turonien inf.) de la région de Tébessa (NE de
l'Algérie), le travail entrepris a deux objectifs principaux,
biostratigraphique et paléoenvironnemental.
Pour atteindre ces objectifs, une centaine
d'échantillons, incluant de nombreux fossiles, ont été
prélevés sur une coupe longue de plus de 2 km, dans des niveaux
essentiellement marneux.
Une étude systématique a alors été
entreprise, consistant, pour les échantillons meubles, en un lavage des
marnes, puis en un tri minutieux des microfossiles sous l'objectif d'une loupe
binoculaire. Ostracodes et foraminifères (benthiques comme
planctoniques) ont été consciencieusement comptabilisés
afin de réaliser des études statistiques, utiles aux
reconstitutions paléoenvironnementales. Les 'bons' microfossiles
stratigraphiques ont bien entendu été soigneusement
recherchés.
Les échantillons indurés ont pour leur part
été sectionnés afin de réaliser des lames minces,
et plusieurs autres méthodes ont par ailleurs été
employées, pour arriver à déterminer le plus
précisément possible les caractéristiques des niveaux
étudiés.
L'étude de la crise cénomano-turonienne,
évènement d'ampleur mondiale, a également retenu toute
notre attention.
La présentation de ce travail s'articule classiquement
en trois parties. Dans la première sont présentées
quelques généralités, dans la seconde, une description
succincte des méthodes employées, tandis que dans la
dernière partie, de loin la plus longue, sont exposés les
résultats obtenus et l'interprétation qui en découle. A
chaque niveau, un maximum d'illustrations ont été
proposées, ainsi qu'une liste bibliographique la plus complète
possible.
Chap. I
Généralités
I - Généralités 2
1. Contexte géographique 2
2. Contexte géologique 3
3. Présentation du site d'étude 5
4. Précision sur les terrains étudiés 6
I - Généralités
1. Contexte géographique
Le site étudié est localisé au nord-est
de l'Algérie, dans la partie orientale de la wilaya de Tébessa,
qui est entourée des wilayas de Souk Ahras, d'Oum El Bouaghi, de
Khenchela et finalement d'El Oued au sud. A l'est, c'est la frontière
tunisienne qui lui sert de limite
Le site qui nous intéresse se situe à mi-chemin
entre le chef-lieu de wilaya et la ville de Morsott, soit une vingtaine de km,
dans la plaine qui s'étend au pied des Djebels Dyr, Chemla et Zitouna
(fig.1).

SPRGNE
AROC
36°00 --
Dj. Es Stihca~~a Sed
0-
pl Dl Troubla
Hamm
K
ti°
8°00'
Figure 1 : Localisation géographique du
site.
2. Contexte géologique
Pour rédiger ce chapitre, nous nous sommes
essentiellement basés sur les travaux de Blés et Fleury (1970),
dans l'explication qu'ils font de la carte géologique de Morsott, ainsi
que sur ceux de Dubourdieu (1956), et, dans une moindre mesure, sur la
volumineuse et incontournable thèse de Vila (1980).
Nous ne décrirons pas avec précisions les
formations géologiques rencontrées dans notre région
d'étude, les auteurs cités précédemment en faisant
une description exhaustive.
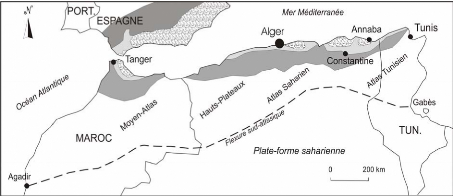
En introduction, nous allons évoquer les structures
géologiques qui forment l'Algérie. Celles-ci,
grossièrement disposées parallèlement à la
côte méditerranéenne, comprennent, du sud au nord (fig.2)
:
la plate-forme saharienne, zone ancienne stable comportant une
couverture paléozoïque globalement tabulaire et
séparée de l'entité suivante par la flexure sud-atlasique,
accident majeur de la croûte terrestre,
la chaîne tertiaire intracontinentale des Atlas, qui
comprend le Haut-Atlas marocain, l'Atlas saharien d'Algérie et l'Atlas
tunisien,
la zone stable des hautes plaines, accidentée à
l'ouest par le Moyen Atlas,
et enfin, tout à fait au nord, la grande chaîne
alpine des Maghrébides, elle- même subdivisée en plusieurs
ensembles (domaines interne / des flyschs / externe), qui se poursuit depuis le
sud de l'Espagne et jusqu'à la province italienne de Calabre vers
l'est.
V
200 km
Différents domaines de la chaîne alpine des
Maghrébides
Domaine interne Domaine des flyschs Domaine externe
Figure 2 : Schéma des unités structurales
d'Afrique du Nord
(modifié d'après Durand Delga et
Fontboté, 1960)
La région qui nous intéresse se trouve dans la
partie orientale de l'Atlas saharien, ensemble structural
situé dans la continuité du Haut Atlas marocain et qui se
prolonge vers
l'est dans l'Atlas tunisien.
La zone de Tébessa / Morsott ne présente
à l'affleurement que des formations sédimentaires,
essentiellement calcaires et marneuses, dont les âges sont compris entre
le Trias et le Miocène, le tout étant recouvert par endroit par
des formations superficielles quaternaires, de type éboulis de pente ou
alluvions (pour plus de précisions, voir Blés et Fleury,
1970).
La série stratigraphique est cependant loin
d'être complète, aucune trace de Jurassique n'ayant
été à ce jour découverte, pas plus que de la partie
basale du Crétacé inférieur. Le Trias évaporitique
est donc la formation la plus ancienne présente, mais il est intrusif au
sein de formations sédimentaires plus récentes. Ce diapirisme,
très caractéristique de la partie orientale de l'Atlas saharien,
ainsi que de l'Atlas tunisien, provoque un bouleversement des roches
encaissantes : réduction d'épaisseur, changements brusques voire
renversements de pendage, laminage des roches. Pas moins de trois pointements
triasiques (« trois diapirs ») sont présents sur la feuille de
Morsott, d'ampleur certes variable, au niveau des Djebels Belkfif, Hameimat
Nord (ce dernier jouxte notre zone d'étude) et Hameimat Sud (fig. 3).
Les formations les plus anciennes en 'position normale' sont
représentées par des dépôts d'âge Aptien dans
la feuille de Morsott, et d'âge Barrémien dans les territoires
contigus (par exemple dans la région de l'Ouenza ou au niveau du Djebel
Mesloula [Dubourdieu, 1959]). L'histoire géologique
anté-barrémienne est donc inconnue dans la région.
8° 8°20
35°411-
35°30' -

·
Mors
A
Formations quaternaires
du fossé d'effondrement
de Morsott-Tébessa
rnla
Hameimat nor
A
Hameimat s
0 2.5 km
Dj
|
Trias
Vraconnien-Aptien
Cénornanien
Turonien inf.
Campanien inf. / Turonien sup
|
re2
|
Maastrichlien inf. / Campanien moy Paléocène
inf. / Maastrichtien sup. Lutétien inf. / Paléocène
sup.
Miocène inf.
Plio-Villanfranchien
|
|
n5-7
|
|
cl
|
e4-6
|
|
|
|
|
|
c2a
|
|
|
|
c2b
|
|
|
|
|
|
|
|
|
|
|
|
|
Figure 3 : Carte géologique de la région de
Morsott (d'après la carte géologique de Morsott au 1/50 000) et
localisation de la coupe étudiée.
L'ensemble de ces formations a été plissé
de façon modérée (rétrécissement
latéral estimé à 5% seulement au niveau des calcaires
turoniens sur la coupe du Djebel Zitouna), suivant une orientation moyenne
NE-SW. Cette direction moyenne, qui représente la phase de plissement
principal, dite `atlasique', a été acquise entre le
Lutétien inférieur et le Miocène.
Par la suite, se sont individualisés des fossés
d'effondrement, d'orientation globale NW-SE, dont ceux de Tébessa
(WNW-ESE) et de Morsott (NNW-SSE). Leur mise en place a débuté
avant le Plio-Villafranchien et s'est poursuivie jusqu'à une
époque récente. L'amplitude verticale maximale du
déplacement atteint 450 m environ pour le fossé de Morsott et
jusqu'à 800 m pour celui de Tébessa (Blés et Fleury,
1970).
3. Présentation du site d'étude
Nous allons dans la suite de ce mémoire nous
intéresser non seulement à une région précise,
déjà détaillée au-dessus, mais également
à une époque bien déterminée Ce sont en effet des
formations cénomaniennes qui vont recueillir toute notre attention, et
plus précisément celles de la coupe du Djebel Chemla, que nous
nommerons ainsi désormais.
La coupe du Djebel Chemla s'étire sur
environ 2.5 km entre le Djebel Hameimat Nord au SW et le Djebel Chemla au NE,
à une altitude comprise entre 850 m à la base et 1200 m environ
au sommet (fig. 4). Les couches cénomaniennes, à dominante
marneuse, y sont en position normale, encadrées par des formations
vraconniennes à la base et turoniennes au sommet
SVV A B NE

1300 m
1300 m
1200 m
1200 m
1100 m
1100 m
1000 m
1000 m
900 m
900 m
800 m
800 m
700 m
700 m
0 500 m
Figure 4 : Coupe du Djebel Chemla.
(les
notations et le code de couleur employés sont les mêmes que sur la
figure 3).
L'épaisseur de la formation cénomanienne du
Djebel Chemla est donc particulièrement importante (environ 700 m), ce
qui est confirmé par Blés et Fleury (1970), Dubourdieu (1956) ou
encore Benkherouf (1988) dans les territoires avoisinants : ces derniers
évoquent des épaisseurs respectives d'environ 600 m pour la
région de Morsott, de 750 à 1000 m pour celle de l'Ouenza et de
715 m pour le Dj. Dyr.
4. Précision sur les terrains
étudiés
Notre intérêt s'est porté sur des
terrains, principalement marneux, d'un âge compris entre l'Albien
terminal (sous-étage Vraconnien) et le Turonien basal. La majeure partie
de notre étude porte donc sur des terrains d'âge
Cénomanien, c'est-à-dire du Crétacé moyen, selon
les uns, ou de la partie basale du Crétacé supérieur,
selon les autres (fig.5).
L'étage Cénomanien doit son nom à la
ville française du Mans (nommée Cenomanum en latin),
où il a été défini en 1847 par d'Orbigny, et
s'étale approximativement entre 96 et 92 Ma (mais les marges d'erreur
restent très importantes, puisqu'elles sont comprises entre 1 et 3
Ma).
Les terrains étudiés semblent présenter
des caractéristiques typiques, ce que confirme Vila (1980) qui
évoque dans la région et en Tunisie voisine un
Crétacé supérieur très homogène, seulement
marqué par des variations d'épaisseurs. Les formations
vraconnocénomaniennes y sont décrites comme principalement
marneuses et se terminent par une « barre » calcaire, correspondant
aux formations dites « Fandene » et « Bahloul » de Tunisie
(Burollet, 1956).
|
QUATERNAIRE
|
Ma
1,75
3,4
5,3
7,1
11
14,7
15$ 2°3 23 28 33,7 37 46 53 55
68 72 83 87
(a
92
g6
108 113 117 123
131 135 141 146 154 160 164 170 175 184 181
200 203
-
220 230 233 240
250
|
|
0
M
Z
O
N
0
Â
C
111
|
z
rn
0
o
m
z
m
|
PLIOCENE
|
PLAISAN CI EN
|
|
ZANCLEEN
|
|
M IOCE NE
|
MESSI NI EN
|
|
TORTONIEN
|
|
SERRAVALLI EN
|
|
LAN G HI EN
|
|
BURDI GALIEN
|
|
AQUITAN IEN
|
|
OLIGOCENE
|
C HATT IEN
|
|
RU PELIEN
|
|
-0
M
O
G1
m
z
m
|
EOCENE
|
PRIABONI EN
|
|
BARTONI EN
|
|
LUTETI EN
|
|
46 YPRESIEN
|
|
PALEOCENE
|
THANETIEN
|
|
DANI EN
|
|
m
Cn
0
N
0
c
m
|
0
m
D
o
m
|
Supérieur
|
w
Z
o m
z
5
|
MAASTRICHTI EN
|
|
CAMPANIEN
|
|
SANTON IEN
|
|
CON IAC IEN
|
|
TU RON IEN
|
|
CEN OMAN IEN
|
|
Inférieur
|
Vracon rien
ALBI EN
|
|
APTIEN
|
|
rf,
8
0T
r,
z
|
BARREMI EN
|
|
HAUTERIVIEN
|
|
VALAN GIN IEN
|
|
BERRIASIEN
|
|
C
cn
D
C
m
|
MALM
|
TITHONIEN
|
|
KI MMERIDGIEN
|
|
OXFORDIEN
|
|
CDOGGER
|
CALLOVIEN
|
|
BATHONI EN
|
|
BAJOC IEN
|
|
AALENIEN
|
|
LIAS
|
TOARCI EN
|
|
PLI ENSBAC HIEN
|
|
SI NEM URI EN
|
|
HETTANGIEN
|
|
Supérieur
|
RHETIEN
|
|
NOR IEN
|
|
CARNIEN
|
|
cn
|
Moyen
|
LADI NI EN
|
|
AN ISI EN
|
|
Inférieur
|
OLENEKI EN
|
|
IN DUSIEN
|
|
|
Ma
255 258 265 275 285 288
-
305
-
315
328
-
-
-
345
-
388
365
375 380 385 390 408 418 425
-
430
-
-
435
448
488
462
465
480
500
-
-
525
530
|
|
>
r
0
N
O
C
rrl
|
M
xi
Z
|
THURINGIEN
|
TATARIEN
|
|
KAZAN IEN
|
|
SAXO NI EN
|
KUNGURIEN
|
|
ARTINSKIEN
|
|
AUTUNIEN
|
SAK
|
|
ASE MARIN
|
|
0
P3
O
-n
m
xj
m
|
ro SILESIEN
|
I, rl
|
GZHELI EN
|
|
KASIMOVIEN
|
|
î
11 '
|
MOSCOVI EN
|
|
BAS HKIRI EN
|
|
'' zf
|
SERPUKHOVI EN
|
|
DI NANTI EN
|
e
m
Z
|
BRI GAN TI EN
|
|
ASBIEN
|
|
H OLKERI EN
|
|
ARUNDIEN
|
|
CHADI EN
|
|
11
|
IVORI EN
|
|
HASTARI EN
|
|
171
<
Z
ni
z
|
Supérieur
|
FAMMENNI EN
|
|
FRASNI EN
|
|
Moyen
|
GIVETI EN
|
|
|
Inférieur
|
EMSIEN
|
|
PRAGUI EN
|
|
LOCH KOVI E N
|
|
r
5
|
PRIDOLI
|
PRI DOLIEN
|
|
LU DLOW
|
LU DFORDI EN
|
|
GORSTI EN
|
|
WENLOCK
|
HOMERIEN
|
|
SHEINWOOD IEN
|
|
z z LLANDOVERY
|
TELYCH IE N
|
|
AERONI EN
|
|
RHUDDANIEN
|
|
0
P:1
D
0
<
0
Z
|
Supérieur
|
Su
ASHGI LLI EN
|
|
CARADOCI EN
|
|
Mo n
ye
|
LLAND El LIEN
|
|
LLANVI RNI EN
|
|
Inférieur
|
AREN IG I EN
|
|
TRE MAD OC IE N
|
|
0
E ro Xi Z
|
Supérieur
|
|
Moyen
|
MAYAI EN
|
|
AMGAIEN
|
|
Inférieur
|
LENIEN
|
|
ATDABANIEN
|
|
TOMMOTI EN
|
|
PALEOCAM BRIEN
|
|
540
|
Figure 5 : Echelle stratigraphique des temps
géologiques (datations d'après ODIN et ODIN,
1990).
(la période nous intéressant est
représentée en grisé)
Chap. II
Matériel
et
Méthodes
II- Matériel et méthodes 8
1.Description des prélèvements 8
2.Méthodes utilisées 8
a-Calcimétrie 8
b-Dosage du carbone organique total (COT) 8
c-Traitement des échantillons meubles ou faiblement
indurés 8
d-Paramètres déterminés à l'issue du
tri 9
Chap. H : Matériel et Méthodes
II- Matériel et méthodes
1. Description des prélèvements
· Environ 3/4 des échantillons
récoltés sont marneux ou marno-calcaires, donc faiblement
indurés. Pour ces échantillons, un prélèvement
d'environ 500 g a été réalisé approximativement
tous les cinq mètres, parfois plus (par exemple faute d'affleurements),
parfois moins, notamment en cas de variations d'apparence significatives.
Chacun d'entre eux a bien entendu été prélevé
après 'rafraîchissement' de la zone de prélèvement
(afin d'éliminer les zones altérées), puis placé
dans un récipient approprié et numéroté
correctement.
· Le reste des échantillons,
représenté surtout dans la partie finale de la coupe, est
constitué de calcaire induré. Afin de pouvoir réaliser par
la suite des lames minces, un petit bloc de roche a été
dégagé et collecté.
· De plus, dans tous les niveaux fossilifères,
parfois très riches, nous n'avons pas manqué de récolter
de nombreux fossiles, qui ont été également soigneusement
numérotés.
2. Méthodes utilisées
a- Calcimétrie
Le taux de carbonate de calcium contenu dans les
échantillons a été évalué au moyen de la
méthode classique du calcimètre de Bernard. En comparant le
volume de CO2 dégagé par l'échantillon (sur lequel
quelques mL d'acide chlorhydrique ont été versés) à
celui qui serait dégagé par une quantité donnée de
calcite pure, il est possible de déterminer très simplement la
quantité de CaCO3 contenue dans l'échantillon analysé.
b- Dosage du carbone organique total (COT)
Il s'agit de rechercher, dans un échantillon
donné, le carbone ayant une origine organique (y compris donc le carbone
des os, des tests ou des coquilles), désigné par le sigle COT (ou
'TOC' en anglais). L'analyse se fait sur un échantillon broyé et
préalablement décarbonaté, à l'aide d'un
pyrolyseur, dans notre cas de type LECO IR212 (mesures réalisées
au laboratoire de Tectonique de l'Université Paris VI - Jussieu).
c- Traitement des échantillons meubles ou
faiblement indurés Pour tous les échantillons, trois
étapes principales ont été suivies :
1- Trempage :
L'échantillon (environ 100g) est mis à tremper
pendant une nuit dans de l'eau (chaude de préférence, la
réaction étant plus rapide et plus complète)
additionnée d'eau oxygénée (environ une cuillère
à soupe pour chaque échantillon).
Cette procédure est cependant souvent insuffisante pour
les échantillons les plus indurés (en général
riches en carbonate de calcium). Ceux-ci sont alors soumis, après ajout
de cristaux de sel, à un ou plusieurs cycles
congélation-décongélation : l'action du gel permet en
général de désagréger convenablement le
sédiment.
2- Lavage :

Les échantillons marneux, devenus complètement
malléables après l'étape de trempage, sont lavés au
travers de deux tamis (1), l'un de maille grossière (2 mm), et le second
de maille très fine (0.063 mm) Au final, les échantillons sont
donc débarrassés de leurs éventuels résidus
grossiers (notamment des fragments de fossiles) et de leur fraction
limono-argileuse.
Pour éviter les contaminations malencontreuses entre
échantillons, les tamis sont imprégnés avec un colorant
(ici du bleu de méthylène) qui permettra, lors de l'étape
ultérieure de tri, de repérer les 'intrus' (évidemment
colorés en bleu). Après un lavage méticuleux, le
résidu du tamis fin est mis à sécher à l'air
libre.
3- Tri :

(4)
(5)
Une fois l'échantillon parfaitement sec, l'étape
finale du tri peut débuter, et c'est là que commence
véritablement enfin le travail du micropaléontologue.
Après avoir étalé quelques mg de sédiments sur un
'plateau de tri'(2) (dans notre cas un simple carton peint en noir), le travail
minutieux de tri se fait sous l'objectif d'une loupe binoculaire') : les
microfossiles repérés sont soigneusement triés un à
un à l'aide d'une 'aiguille montée'('>, puis placés,
idéalement, dans une 'cellule de microfaune'"), sorte de boîte
miniature munie d'un couvercle.
Afin de pouvoir réaliser des études
statistiquement viables, un minimum de 200 individus doit être
isolé. Patience et longueur de temps doivent donc être les atouts
du micropaléontologue : en effet, le tri d'un seul échantillon
exige entre 2 et 5 heures (cette durée est fonction notamment de la
quantité de microfossiles présents, de leur taille, de leur
état de conservation...).
d- Paramètres déterminés à
l'issue du tri
Lors du tri, les microfossiles identifiés ont
été comptabilisés (jusqu'à atteindre un total
de 200 environ) et regroupés par populations. Nous avons
ainsi pu différencier (tableau 1):
les foraminifères benthiques (B), parmi lesquels nous
distinguerons les foraminifères benthiques calcaires (BC) et ceux
agglutinés (BA),
les foraminifères planctoniques (P), parmi lesquels nous
distinguerons les foraminifères planctoniques globuleux (PG) et ceux
carénés (PC),
les ostracodes (0).
Les rapports entre les valeurs obtenues donneront des
indications sur les paléoenvironnements (ce sera l'objet du chapitre
suivant). En plus de ces comptages, nous avons également
évalué :
- la diversité spécifique :
Nous parlerons de population peu, moyennement ou très
diversifiée (il est difficile cependant de déterminer
précisément la richesse spécifique, au vu de la
difficulté parfois grande pour la détermination exacte des
spécimens),
- la richesse en microfossiles :
Exprimé en nombre de microfossiles par gramme de
sédiment trié, ce paramètre ne reflète pas
seulement la richesse (micro)fossilistique réelle, mais dépend de
plusieurs facteurs, dont notamment la qualité du lavage (meilleur il
aura été, plus les fossiles seront concentrés dans le
résidu), la proportion en éléments fins (silts et argile),
éliminés lors du lavage, l'induration du sédiment (un
sédiment induré sera évidemment moins bien trié
qu'un sédiment tendre) et bien entendu la qualité du tri et
l'expérience du micropaléontologue.
Remarque : Pour déterminer la richesse
réelle, il aurait fallu peser précisément
l'échantillon avant et après lavage (pour ainsi évaluer
avec exactitude la proportion de fraction fine éliminée), ce qui
n'a pas été fait lors de cette étude.
|
--.
rn
.,?....)
N
o
;.-
,E2,
rn
k
e
t2
ts
4q
|
Foraminifères benthiques (B)
Vivent sur le fond, fixés ou non, enfouis dans le
sédiment (endobionte) ou à la surface (épibonte). Leur
abondance diminue progressivement en s'éloignant du littoral. Les
espèces benthiques sont beaucoup plus répandues que les
espèces planctoniques (actuellement seules 70 des 4000 espèces
vivantes décrites sont planctoniques)
|
Foraminifères benthiques calcaires
(BC)
Pourvus d'un test calcaire secrété par l'animal,
et
abondants dans les zones les plus superficielles (ex :
Nodosaria, Lagena...)
|
|
Foraminifères benthiques agglutinés =
arénacés (BA) :
Pourvus d'un test d'origine exogène, constitué
de
particules captées dans le milieu environnant. Sont
présents à toutes les profondeurs, mais sont les seuls qui
subsistent à de grandes profondeurs. Leur abondance relative augmente
donc avec l'approfondissement (ex : Ammodiscus, Textularia ...)
|
|
. e
Foraminifères planctoniques
(P)
Flottent librement dans la tranche d'eau, et sont
présents aussi bien près des côtes qu'au large. Leur
abondance relative augmente cependant avec l'approfondissement, en
rapport avec la moindre proportion d'organismes benthiques
présents dans les plus grandes profondeurs.
|
Foraminifères planctoniques globuleux (PG)
Vivent à proximité de la surface (ex :
Heterohelix, Hedbergella ...)
|
|
Foraminifères planctoniques
carénés (PC)
Formes plus complexes, munies d'une carène, vivant
à de plus grandes profondeurs (ex : Globorotalia,
Rotalipora...)
|
|
--,
rn rn
2
et N
rd ' 'è)
|
Ostracodes (0)
Groupe d'arthropodes crustacés, dont la taille peut
aller jusqu'à 10 mm. Vivent dans tous les milieux aquatiques, et sont
généralement benthiques, endobiontes (donc enfouis dans le
sédiment) ou épibiontes (sur le sédiment). Certains sont
pélagiques (et munis d'appendices servant à la nage).
L'étude des populations d'ostracodes donne de précieuses
indications en terme de paléoenvironnement (bathymétrie,
salinité notamment).
|
Tableau 1 : Type de microfossiles
observés.
La détermination de ces différents
paramètres, et surtout l'étude de leur variation, va nous
permettre d'obtenir des informations sur l'évolution des
paléoenvironnements, chacune des populations distinguées
étant spécifique d'un environnement particulier (voir l'exemple
des foraminifères en figure 6).

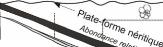

Foraminifères planctoniques
épipélagiques ...Cm.;.kigglbgr_gg!!K.11.Mugreliz)
Pet?)
·
Foraminifères planctoniques
pe,s,
E
·11F- .4e mo- à ues
ee/ce,_
'9,9 (exés
Globorotabathypélagilia.
Rotaliqpora)
n
"tee"
----
50 oeb, al ) thj,
Diversité maximale des foraminifères calcaires
benthiques
Diversité maximale des foraminifères
agglutinés
* N.M = niveau marin
N- M"
100 m
Figure 6 : Relation entre profondeur du milieu et type
de foraminifères présents
(modifié d'après
Brasier, 1980).
Chap. III
Résultats
et
Interprétations
|
a
|
|
|
III -- Résultats et interprétations
|
12
|
1. Généralités
|
12
|
a-Historique des travaux existants
|
12
|
b-Aperçu général sur le monde au
Crétacé moyen
|
12
|
c-Le log stratigraphique
|
14
|
2. Résultats
|
16
|
a-Les fossiles
|
16
|
b-Aspects minéralogiques et géochimiques
|
21
|
c-Résultats du tri des microfossiles
|
23
|
d-Etude des lames minces
|
45
|
3. Interprétations
|
|
52
|
|
|
a-Axe Biostratigraphie
|
52
|
|
b-Axe Paléoenvironnement / Paléogéographie
|
54
|
|
c-La crise cénomano-turonienne
|
62
|
|
d-Le passage Vraconnien-Cénomanien
|
65
|
|
e-Questions en suspens
|
66
|
III -- Résultats et interprétations
1. Généralités
a- Historique des travaux existants
Les travaux récents concernant la
paléontologie, et surtout la micropaléontologie de l'est
algérien sont rares. Nous citerons cependant les travaux de
Glintzboeckel et Magne (1960) ou encore de Sigal (1952a) : ces études,
déjà anciennes, sont malheureusement difficiles à trouver
et n'ont pas pu être consultées directement. Plus
récemment, Vivière (1985) et Benkherouf (1988) se sont
intéressés à la région : ces derniers travaux en
particulier nous ont été d'un grand secours. D'autres
études, autant paléontologiques que stratigraphiques, ont
également été entreprises plus récemment, dont
celles de ChikhiAouimeur (1998), Herkat (1999), Boumaza (2000) ou
Redjimi-Bourouiba (2002).
Si les travaux concernant l'est algérien restent peu
répandus, ceux concernant les contrées voisines de Tunisie sont
en revanche nombreux. Nous n'évoquerons que les plus récents et
ceux sur lesquels nous nous sommes véritablement appuyés : Caron
et al. (1999, 2006), Robaszynski et al. (1993, 2007).
b- Aperçu général sur le monde au
Crétacé moyen
La Terre au Crétacé, et en particulier au
Crétacé moyen, offre un visage bien différent de celui que
nous connaissons actuellement, et ce sur différents points :
- les températures semblent
avoir été uniformément chaudes, à tel point que
certains auteurs estiment que les températures de surface des
océans tropicaux étaient 4 à 7 °C plus chaudes que
les températures maximales actuelles, soit comprises entre 32 et
34°C (Norris, 2002), et que les pôles étaient très
probablement libres de toute glace.
- la 'transgression cénomanienne'
(pas vraiment synchrone en réalité, cet
étage correspondant selon les lieux à une période de
montée du niveau marin ou au début de la régression qui
suit) avait porté les océans à leur plus haut niveau des
600 derniers millions d'années, cette transgression étant bien
caractérisée même par des auteurs anciens (Gignoux et
Moret, 1935). Des mers épicontinentales recouvrent alors une grande
partie des continents.
- la configuration du monde enfin
n'est guère semblable à celle qui prévaut à l'heure
actuelle : la Pangée, qui a commencé à se disloquer bien
avant le début du Crétacé, continue à se fracturer.
A la fin du Jurassique, l'Afrique et l'Amérique du sud sont encore
solidaires, tandis que l'océan Atlantique a commencé à
s'ouvrir dans sa partir nord. Au début du Crétacé, cette
ouverture se poursuit dans la partie médiane, puis dans la partie sud.
Au Crétacé moyen, l'Afrique est donc totalement
séparée de l'Amérique, et séparée de
l'Europe par un océan, la Téthys, ancêtre de l'actuelle
Méditerranée (fig. 7). Les dépôts formés
à l'époque dans l'actuelle Afrique du nord appartiennent donc au
domaine dit `téthysien'. La région de Tébessa se situe
approximativement à 15° de latitude Nord.

OCEAN
AR pij
OCEAN PACIFIQUE
AT LA N TI NORD
Chine
Arabie
ATLANTIQUE
SUD ntarctig
* surfaces grisées = aires émergées
trait blanc = contour des continents
0 2000 km
Dterres émergées \?. contours actuels
sédimentation carbonatée
(en pointillés : zones à sédimentation
détritique)
Tt] position approximative de Tébessa
Figure 7 : Carte du monde au Cénomanien, il y a
94 Ma.
(carte du haut modifiée d'après Scotese, 2002
et
carte du bas modifiée d'après Andreu, 1991).
c- Le log stratigraphique
Entre le Vraconnien à la base et le Turonien
inférieur au sommet, s'étale une succession de couches marneuses
et de plus rares niveaux calcaires, dont nous allons détailler
ci-dessous la succession (fig.8):
· Vraconnien (environ 30m) : calcaire noir
en plaquettes, alternant avec des marnes sombres.
· Cénomanien (environ 700 m):
inférieur : épaisse série marneuse, de
couleur sombre (teintes variées de gris ou de marron),
fréquemment riche en gypse et peu fossilifère. On y retrouve
cependant des bivalves, des ammonites et de rares bélemnites. Les
intercalations calcaires sont rares, et ont livré des traces d'ammonites
(indéterminées).
moyen : les intercalations calcaires sont plus abondantes et
plus épaisses. Plusieurs bancs lumachelliques ont livré une riche
faune, notamment de gastéropodes, d'oursins et de bivalves, dont des
ostréidés en grand nombre, ce qui justifie le qualificatif de
« marnes à huîtres » parfois donné à ces
formations. Notons également la présence de plaques de calcite
fibreuse, livrant parfois des empreintes d'oursins, et déjà
décrites par Dubourdieu (1956) pour qui ceux-ci ne sont pas
déterminables.
supérieur : vers le sommet, les couches marneuses
cèdent peu à peu la place à des niveaux calcaires qui
assurent la transition avec le Turonien. Les fossiles récoltés y
sont beaucoup moins nombreux que dans le Cénomanien moyen. Les bancs
calcaires (ou calcaro-marneux) présentent une texture micritique et une
teinte grisâtre générale.
· Turonien inférieur (environ
45m) : la transition avec le Cénomanien est progressive et
délicate à situer précisément. Nous
considérerons cependant que la présence de niveaux de calcaires
noirs à débit 'en plaquettes' marque cette limite (fig.8). Ces
niveaux, caractéristiques de la survenue de l'évènement
anoxique de la limite Cénomanien/Turonien, l'OAE2 (` Oceanic
Anoxie Event 2' ou ' Evènement d'Anoxie
Océnanique', - le second du genre au
Crétacé-, tel qu'il a été défini par
Schlanger et Jenkyns en 1976), sont typiques de cette période et
amplement décrits dans la littérature, et notamment en Tunisie
voisine (voir à ce sujet Caron et al., 1999 ; 2006). Ces
niveaux de transition sont ensuite suivis par des niveaux de calcaires gris en
petits bancs (à texture toujours micritique), surmontés par de
gros bancs plurimétriques de calcaire ocre, formant dans le paysage une
corniche marquée.

C
È
e
oe:b
MMIMM-g
cy
rt,
106 à 122
105
61eabeieed
· -----
· --
wisszeezz,..-7,m
1111.
rfflel.
rle=Lefliga
· MiM IMM»MMIMIMIM
Mw
·
·
·
15
1
·
·
1,4, 18
Calcaire ai noir à débit en
plaquettesLimite Cénornanieri/Turonien
0
' 4:
· --
Premier vrai banc calcaire (éch.
N°37),7\47.
.;74e2.,-,,t)
Cornac; Vraconmen/ Canomanien
Marnes - Cencnnamen
W;',1Ê
37
Chap. III : Résultats et interprétations
/ère e1 2ème corniches calcaires -
Turomen
-g3
it Gypse
aD
Figure 8 : Log stratigraphique de la coupe du Dj. Chemla.
2. Résultats
Dans ce travail, nous avons cherché à atteindre
deux objectifs principaux, à savoir, d'une part, essayer de reconstituer
les conditions paléoenvironnementales (et
paléogéographiques) régnant lors du Crétacé
moyen (`axe paléoenvironement'), et, d'autre part, effectuer des
interprétations d'ordre biostratigraphique (`axe biostratigraphie').
Pour ce faire, nous avons recherché tous les indices
utiles, qu'ils soient d'ordre paléontologique et
micropaléontologique (récolte systématique des fossiles
rencontrés sur le terrain, lavage et tri pour les microfossiles),
d'ordre sédimentologique (granulométrie des sédiments) ou
minéralogique (présence de minéraux remarquables).
Pour chaque étape, nous nous sommes efforcés,
dans la mesure du possible, de proposer un maximum d'illustrations,
photographies ou schémas (dont la plupart ont été
réalisés selon nos propres observations).
La détermination des fossiles et microfossiles a
été réalisée en utilisant la documentation en notre
possession, forcément trop rare et incomplète. La
détermination proposée sera donc la plus complète possible
(avec nom de genre et espèce), mais, dans un certain nombre de cas, nous
avons préféré la laisser imparfaite (nom de genre seul,
voire de famille) plutôt que de risquer une interprétation
erronée. D'éventuelles erreurs restent cependant
inévitables, et nous tenons à nous en excuser d'avance
auprès du lecteur.
a- Les fossiles
Plusieurs types d'organismes ont été
rencontrés, principalement dans les niveaux lumachelliques du
Cénomanien moyen et supérieur. Il s'agit notamment de mollusques
(bivalves, gastéropodes et céphalopodes) et
d'échinodermes. Des photos des spécimens les mieux
conservés sont présentées dans les planches 1, 2 et 3 qui
suivent.
Par ordre d'abondance, nous citerons :
- les bivalves ou lamellibranches
(répartition stratigraphique : Cambrien -- actuel) :
Ils sont de loin les plus abondants. Leur utilité est
surtout d'ordre paléoenvironnemental. Les bivalves les plus
fréquents sont des ostréidés (équivalents de nos
huîtres actuelles). Souvent présents en très grand nombre,
ils se rencontrent à tous les niveaux, et ont pour représentant
principal des Exogyres (uniquement Crétacé). D'autres groupes de
bivalves, généralement moins bien représentés,
peuvent également être observés (p1.2).
Les ostréidés regroupent plusieurs genres
(classification d'après Malchus, 1990) :
Sous-ordre OSTREINA Ferussac, 1822
Super-famille OSTREACEA Rafmesque, 1815 Famille
OSTREIDAE Rafmesque, 1815
Sous-famille EXOGYRINAE Vyalov, 1936 .p1.1.B,
C
Genre Amphidonte Fischer de Waldheim,
1829 .p1.1B
Sous-genre Amphidonte Fischer de Waldheim, 1829
Sous-genre Ceratostreon Bayle, 1878
Sous-famille LOPHINAE Vyalov, 1936
Genre Lopha Roding, 1798 p1.1E
Sous-genre Actinostreon Bayle, 1878
Genre Rastellum Faujas-Saint-Fond, 1799
p1.1D, F
Sous-genre Arctostrea Pervinquiere, 1910
- les gastéropodes
(répartition stratigraphique : Carbonifère --
actuel)
En général associés aux bivalves dans les
niveaux à lumachelles, ils sont également très
fréquents. Là encore, leur utilité est surtout d'ordre
paléoenvironnemental.
- les échinodermes : seuls des
Plaque de calcite fibreuse à Echinoïdes (Ordovicien --
actuel) ont
ti empreintes d'oursins.
été récoltés, principalement de
type
irrégulier (le genre Hemiaster semble
7 représenter à lui seul une grande
7, à L partie des spécimens récoltés,
p1.3A)
pour les niveaux cénomaniens
(donc
caractérisant des substrats plutôt
I 3
meubles de type vase).
cm De plus rares échinoïdes réguliers
ont pu être récoltés dans les niveaux du
Turonien basal (Heterodiadema libycum, p1.3C). Cette espèce en
particulier semble caractériser le Crétacé
supérieur (Cénomanien notamment) d'Afrique du Nord mais
également du sud de l'Europe, bien que certains auteurs
(Geys, 1980) l'ait décrite
jusqu'au Campanien.
De plus, au sein même des niveaux marneux, il n'est pas
rare de rencontrer des plaques de calcite fibreuse portant des empreintes
d'Echinoïdes (voir illustration ci-dessus), mais ceux-ci ne semblent pas
identifiables. Des fragments d'entroques sont présents dans près
de 60 % des échantillons triés.
les céphalopodes : ils
comprennent plusieurs groupes, dont :
· les Ammonoïdes : les Ammonites
(Permien -- Crétacé) ont un intérêt stratigraphique
primordial. C'est d'ailleurs sur les ammonites que sont basées de
nombreuses biozonations du Crétacé, même si d'autres
organismes (les foraminifères planctoniques notamment) sont
également de première importance (fig. 9). Malheureusement, en ce
qui nous concerne, seuls de rares spécimens, tous fragmentés, ont
été récoltés (p1.3B, D). Des empreintes ont
également été relevées dans certains niveaux
indurés (et notamment dans les calcaires noirs à débit en
plaquettes du Vraconnien, à ammonites pyriteuses).
· les Bélemnitoïdes
(Carbonifère -- Eocène) : seul un rostre de
bélemnite a été découvert (éch. n°36
dans le Cénomanien inf.). Ces organismes peuvent cependant
présenter un intérêt stratigraphique, notamment pour les
niveaux crétacés.
· les Nautiloïdes (Cambrien --
actuel) : leur intérêt stratigraphique est limité Le genre
Nautilus (un unique spécimen rencontré dans un banc
calcaire du Cénomanien moyen) est le seul genre de Nautiloïdes
persistant à l'heure actuelle.
Etage
|
Sous-étage
|
|
|
-- 89.3
C
2
Z
I--
|
=
a. a.l.
c ,
2 E_
n ..
I-- t.n.
|
|
|
93.5
C Cl) c OC
E
o
C
SC)
0
99.6
|
É.
o
C ''1)
· (1) CL
(..) cop
|
|
|
C
a)
Z
<
|
'5 mc -- ,D,',u, ..15 a .0 f.7
|
|
Ammonites
|
|
|
Subprionocyclus
neptuni
|
|
Romanceras devenarianum
|
|
|
|
Mammites nodosoides
|
|
Watinoceras devonensef
coloradoenoense
|
|
|
|
|
|
|
|
Acanth. rhotomagense
|
|
Cunningtoniceras inerme
|
|
Mantelliceras dixoni
|
|
MantelliceraS
saxbii
|
|
Stoliczkaia dispar
|
Mortoniceras
perinflaturn
|
|
Mortoniceras inflatum
|
Callihoplites
auritus
|
|
Foraminifères planctoniques
|
Zones
|
Marqueurs de zone
|
|
Dicannella concavata
|
Marginotruncana
schneegansi
|
|
|
|
|
Heivetoglobotruncana
|
HeIvetoglobotrun- cana helvetica
|
hefvetica
fielvetoglobotruncana helvetica
|
|
·
|
Whiteinella
|
archaeocretacea
|
Rotalipora cushmani
|
Rotalipora cushmani
|
|
Rotalipora cushmani
|
|
|
Rotalipora reicheli
|
Rotalipora reichefi
|
Rotalipora
globotruncanoides
|
|
|
Rotalipora giobotruncanoicies
|
|
|
Rotalipora appenninica
|
Planomalina buxtorti
|
|
Rotalipora appenninica
|
|
|
Rotalipora
ticinensis
|
|
|
Figure 9 : Biozonation du Crétacé moyen
par les Ammonites et les Foraminifères planctoniques.
(modifié d'après Gradstein et al., 2004).

A B

C

D E

F
Planche 1 : Bivalves
(ostréidés).
A. Indéterminé, B. Amphidonte
(Ceratostreon) flabellatum (GOLDFUSS) (Cénom.
moy.),
C. Ilymatogyra (Afndgyra) africana (LAMARCK)
(répandu dans tout le Cénomanien),
D. Rastellum ?, E. Actinostreon
syphax (COQUAND) (Cénom. inf.), F. Rastellum
?. 19

Chap. III : Résultats et interprétations

A
E
H I
G
Planche 2 : Bivalves (A à F) et
Gastéropodes (G à H).
A. Neithea shawi (PERVINQUIERE) (Cénom.
inf.), B. Trigonia sp. ?, C. Indéterminé, D. Moule
interne de Venus ?, E. Cardita delettrei ? (COQUAND) (Cénom.
sup.),
F. Indéterminé (Cénom. moy.), G.
Nerinea bicatenata ? (COQUAND) (Cénom. moy.),
20
H. Spécimen de la famille des Naticidae ?
(Cénom. moy.), I. Indéterminé (Cénom.
moy.).

C
Planche 3 : Oursins (A, C) et Ammonites (B,
D).

s
·
A. Hemiaster batnensis (COQUAN
D) (Cénom. moy.), B.Indéterminé (Cénom. moy.), C.
Heterodiadema libycum (DESOR) (Turonien inf.), D.
Indéterminé (Cénom. sup.).
b- Aspects minéralogiques et
géochimiques
· carbonate de calcium (CaCO3) [fig.120]
La valeur moyenne de 42 % obtenue pour les
échantillons 'meubles' reflète bien l'appartenance de ceux-ci
à la classe pétrographique des marnes. Les valeurs extrêmes
nous permettent cependant de distinguer, au sens pétrographique, des
argiles (20 % des échantillons analysés contiennent en
effet moins de 15 % de CaCO3) et des calcaires marneux (environ 20 %
présentent un taux de CaCO3 supérieur à 65%).
L'augmentation logique du taux de carbonate à
l'approche des bancs calcaires traduit une diminution de la tranche d'eau, et
à l'inverse, une proportion d'argile croissante s'explique
généralement par un approfondissement du milieu de
dépôt. En supposant des conditions d'oxygénation normales
lors du dépôt, on s'attend donc à retrouver pour les
échantillons les plus carbonatés une plus grande proportion
d'organismes benthiques.
· glauconie
Composée de minéraux argileux riches en fer, la
glauconie est un indicateur paléoenvironnemental intéressant.
Elle se forme en effet uniquement en milieu marin, à des profondeurs
estimées entre 50 et 500 m, soit de l'ordre de celle du plateau
continental et du talus supérieur. Seule une faible proportion des
échantillons analysés en contiennent (5.7 %),
notamment dans le Cénomanien inférieur
(n°15, 16) et la fin du Cénomanien supérieur
(n°103).
· gypse
`Marnes à huîtres' ou 'marnes à gypse' :
ces deux qualificatifs s'appliquent fort bien aux niveaux marneux du
Cénomanien du Dj. Chemla. Près de 60 % des échantillons
contiennent en effet du gypse (parfois nettement identifié sur le
terrain, parfois seulement sous l'objectif de la loupe).
De vastes lagunes côtières au niveau du Sahara
actuel auraient fourni à l'époque de grosses quantités de
gypse, qui se serait déversées jusque dans les mers du nord
(Busson, 1972), d'où une très probable hypersalure des eaux.
· nodules ferrugineux et pyrite
Très fréquents à tous les niveaux
(présents dans environ 45 % des échantillons), ces nodules ne se
forment qu'en milieu réducteur, également nécessaire
à la formation de pyrite (Baudin, 2008). A certains niveaux en effet,
les microorganismes (ostracodes comme foraminifères) sont très
largement pyritisés. Un tel milieu permet aussi la préservation
de matière organique, attestée par la couleur souvent sombre des
échantillons.
· quartz et minéraux argileux
Aucun grain de quartz n'a pu être formellement
identifié lors de notre étude, mais une analyse par diffraction
aux RX aurait vraisemblablement permis, en étudiant la fraction fine, de
découvrir une certaine proportion de quartz (étude qui n'a pas
été mise en oeuvre ici).
En effet, une telle étude menée sur la
région voisine de Kalaat Senan en Tunisie (approximativement 40 km vers
le NE) [Robaszynski et al., 1993] révèle une
présence régulière de grains de quartz très fins
(moins de 2 gm) au sein de la fraction argileuse, et beaucoup plus rare de
grains fms (30 à 100 gm). Les variations de la teneur en quartz
s'expliquent logiquement selon ces auteurs par un éloignement plus ou
moins grand du continent-source.
Les variations des rapports entre les différents
minéraux argileux s'expliqueraient aussi par des variations dans les
conditions paléoenvironnementales et notamment du niveau eustatique.
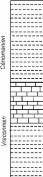
· carbone organique total (COT)
La mesure du taux de COT a été
réalisée sur 10 échantillons, certains
prélevés à la limite Vraconnien/Cénomanien
(n°1A, 1B, 2) et les autres à la limite Cénomanien/Turonien
(n°98, 100, 103, 104, 105, 106, 108). L'utilité de cette analyse
est surtout d'apprécier l'éventuel état d'anoxie du milieu
de dépôt, sachant qu'une valeur de COT de l'ordre de 1% (et
supérieure) est qualifiée d'élevée. De telles
valeurs caractérisent en effet des niveaux de type 'black shales',
considérés comme des roches mères potentielles de
pétrole.
Dans notre cas (fig.10), - et considérant une
incertitude du résultat de l'ordre de 0,05% -, la plupart des valeurs
obtenues sont relativement faibles (< 0,4 %). L'enrichissement en COT est
sensible à l'approche de la limite cénomano-turonienne (0,86 %
pour l'échantillon n°100), puis franchement important dans les
niveaux immédiatement sus-jacents (1,2 à 2 %). Les valeurs
décroissent ensuite rapidement (0,1 % seulement pour
l'échantillon 108).
Des valeurs proches de 3 % ont même été
mesurées dans la région de Tébessa, toujours sur des
niveaux cénomano-turoniens (Naili, 1995).
COT (%)
·
·
1,1 1,1.1
0 0,4 2
|
4Orn
|
|
COT (%)
|
|
|
714 rn 712 m 710 ni 708 m 706 m 704 m 702 m 700 m
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ti
O
|
|
30m 20m 10m Om
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 04 1
|
|
|
|
|
|
|
|
|
|
|
(c)Limite Vraconnien / Cénomanien (c)Limite
Cénomanien / Turonien
Figure 10 : Evolution du taux de carbone organique
total (COT).
Remarque: l'altitude '0m' a été
prise pour les niveaux vraconniens de base, dont l'altitude absolue est en
réalité d'environ 800 m (cette même échelle a
été adoptée pour l'ensemble des autres figures
concernées).
c- Résultats du tri des microfossiles
Trois axes majeurs seront développés, à
savoir :
(1) l'étude des ostracodes,
(2) puis celle des foraminifères planctoniques,
(3) et enfin celle des foraminifères benthiques.
De chacun de ces axes nous tirerons, dans la mesure du
possible, des interprétations d'ordre paléoenvironnemental et/ou
biostratigraphique. Des illustrations sont fournies pour la plupart des
espèces rencontrées (planches 4, 5, 6 et 7), ainsi que quelques
clichés pris au microscope électronique à balayage
(planche 8). Les résultats sont exposés dans les figures 12 et
14.
Avant d'entamer la présentation des résultats,
il est important de souligner la très grande variété des
populations fauniques rencontrées, du moins aux yeux du chercheur
néophyte que j'étais et que je suis encore : ce sont en effet, au
bas mot, pas moins de 40 espèces de foraminifères qui ont
été reconnues (et certainement beaucoup d'autres ignorées
!) et une dizaine d'ostracodes. La diversité est grande également
entre les échantillons observés : certains montrant une
population diversifiée, d'autres ultra-dominés par de rares
espèces, entre des assemblages composés presque uniquement d'un
type de population (planctonique, benthique, ostracodes) ou des assemblages
mixtes. Les enseignements à tirer de ces observations sont
évidemment nombreux et nous allons essayer d'y parvenir le mieux
possible.
Pour caractériser les milieux de vie des organismes
rencontrés, nous emploierons un certain nombre de termes (étage
infra- ou circalittoral, plate-forme carbonatée interne ou externe...)
[fig. 11].
|
Domaines
|
Continental
|
Marin
|
|
|
Milieux
|
fluviatile ou fluvio- iacustre
|
lagunaire
|
côtier
|
marin restreint
|
ou interne
|
marin ouvert ou externe
|
|
Etages
|
|
supralittoral
marginolittoral
|
mediolittoral
|
infralittoral
|
circalittoral
|
bathyal
|
- m
·
·
· - 50 m a
- 80 m
Figure 11 : Etages de peuplement et milieux de
dépôt (modifié d'après Andreu, 1991).
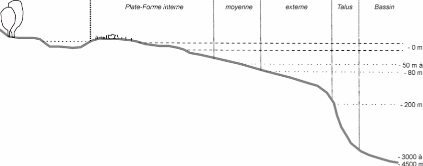
Précisions : L'étage
infralittoral est défini comme la zone comprise
entre les plus basses eaux et la limite inférieure de survie des algues
photosynthétiques (soit une profondeur moyenne de 20m).
L'étage circalittoral lui fait suite et s'étend
jusqu'à une profondeur moyenne de 200 m (il correspond à la
plate-forme continentale externe).
c-1 -- Les ostracodes
Rarement très abondants (un peu moins de 13% du total
des microfossiles triés), leur présence est cependant habituelle,
puisque seuls 7% des échantillons triés n'en contiennent pas du
tout. Les assemblages de microfossiles sont donc généralement
nettement dominés par des foraminifères (benthiques et/ou
planctoniques), à l'exception de 4 échantillons pour lesquels les
ostracodes sont majoritaires (n° 35 et 36 pour le Cénomanien
inférieur, 52 pour le Cénomanien moyen et 94 à la fin de
la période considérée). L'évolution de la
répartition des ostracodes peut être détaillée comme
suit (fig.12):
- la partie inférieure du Cénomanien
est globalement caractérisée par un faible pourcentage
d'ostracodes (5.3%), avec de très rares ostracodes ornementés
(observés dans deux échantillons seulement). Seuls les niveaux
marquant la transition avec le Cénomanien moyen se distinguent, avec une
ostracofaune nettement dominante sur les assemblages de
foraminifères,
- dans le Cénomanien moyen, la
proportion d'ostracodes est sensiblement plus élevée (12.3%),
excepté à son tout début. Les ostracodes ornementés
sont fréquents, mais jamais dominants,
IO a
II a

4101, a ?
b
41111b b
250 i/m
1-1
- enfin, dans les niveaux terminaux du
Cénomanien, la proportion d'ostracodes continue à
croître (15.4% en moyenne), sauf à la limite
Cénomanien/Turonien. Les organismes benthiques (foraminifères et
ostracodes) y sont alors très rares. Remarquons cependant un niveau
particulièrement riche en ostracodes (>57%), marqué dans le
même temps par une très faible proportion de foraminifères
benthiques et donc un rapport P/B assez élevé (>9).
Nous verrons plus loin comment ces évolutions peuvent
être interprétées.
En plus de ces simples comptages, nous avons également
essayé de différencier les individus et, dans la mesure du
possible, de les identifier. Une première distinction a d'abord
été réalisée entre ostracodes à carapace
lisse d'une part, et ostracodes à carapace ornementée d'autre
part. Les premiers (à carapace lisse) sont très largement
majoritaires (près de 96 % de l'ensemble des ostracodes). Seuls 1/3 des
échantillons contiennent des ostracodes à carapace
ornementée, et jamais de façon dominante (voir fig. 120).
Essai de classification des individus rencontrés
:
(d'après la classification proposée par Maddocks
en 1982)
Sous-classe OSTRACODA Latreille, 1806
Ordre PLATYCOPIDA Sars, 1866
Famille CYTHERELLIDAE Sars, 1866
Genre Cytherella a
(JONES, 1849) p1.4 a et b
Ces organismes, à coquille lisse et arrondie, vivent
enfouis dans les premiers millimètres du sédiment, et sont
typiques d'un environnement circalittoral profond (Andreu, 1991). Ils
constituent souvent des assemblages dominants, notamment en association avec le
genre Paracypris. Les coquilles peuvent atteindre des dimensions
impressionnantes.
Ordre PODOCOPIDA Sars, 1866
Sous-ordre CYPRIDOCOPINA Sohn, 1988
Superfamille CYPRIDOIDEA Baird, 1845
Famille CANDONIDAE Kaufmann, 1900
Sous-famille PARACYPRIDINAE Sars, 1923
Genre Paracypris b (SARS,
1866) p1.4 e et f
Le genre Paracypris, à coquille lisse et
très allongée, particulièrement caractéristique,
est lui aussi très fréquent. Son milieu de vie semble être
assez ubiquiste, puisqu'on le rencontre sur toute l'étendue de la
plate-forme, de l'étage infralittoral à l'étage
circalittoral (Andreu, 1991).
Infra-ordre CYTHEROCOPINA Gründel, 1967
Famille CYTHERURIDAE Müller, 1894
Genre Procytherura (WHATLEY, 1970)*
p1.4 j
L'espèce observée se rapproche de
l'espèce Procytherura? sp.1 décrite par Vivière
(1985). L'attribution générique faite par cet auteur semble
cependant incertaine. Pour notre part, elle n'a été
observée que dans quelques niveaux du Cénomanien moyen à
supérieur (n°54, 71), et reste relativement rare. Le genre
Procytherura semble apprécier les environnements
peu profonds, de type infralittoral à intertidal (Damotte, 1992).
Sous-famille CYTHEROPTERINAE Hanai, 1957 Genre
Eocytheropteron (ALEXANDER, 1933)
L'espèce observée, de toute petite taille et
très atypique (valve très pointue, presque 'cornue') semble
être Eocytheropteron glintzboeckeli (DONZE et LEFEVRE, 1981)*,
figurée par Vivière en 1985 (p1.26, fig.6-7). Observée
uniquement dans le Cénomanien inférieur (éch. n° 4,
18, 28), elle y est rare et semble caractéristique de cette
période, du moins en Tunisie (Bismuth et al., 1981).
Famille LOXOCONCHIDAE Sars, 1925
Genre Phlyctocythere (KEIJ, 1958) p1.4
d
L'unique spécimen observé s'apparente à
l'espère Phlyctocythere citreum nouvellement décrite par
Vivière en 1985 dans la région de Tébessa. Cette
espèce n'a été observée que dans un seul
échantillon (n°56) dans le Cénomanien moyen.
Famille SCHIZOCYTHERIDAE Howe, 1961
Genre Amphicytherura C
(BUTLER et JONES, 1957) p1.4 i
Les organismes observés, de petite taille, semblent
appartenir à l'espèce Amphicytherura (Sondagella) distincta
(GERRY et ROSENFELD, 1973) [planche 39 de Andreu, 1991]. Leur
présence a été précédemment attestée
en Algérie, dans la région de Tébessa (Vivière,
1985). Leur milieu de vie semble comparable à celui de Paracypris
(plate-forme carbonatée moyenne à distale, étage
infra- à circalittoral [Andreu, 1991]). Ils constituent l'essentiel des
organismes ornementés rencontrés.
Famille TRACHYLEBERIDIDAE Sylvester-Bradley,
1948
Genres indet. d p1.4 g et h
Ces organismes, dont la présence est anecdotique, n'ont
été observés que dans les tous derniers niveaux
cénomaniens (n° 91 et 94). Peut-être s'agit-il entre autre de
l'espèce Oertliella ? tarfayaensis (REYMENT, 1978) [planches 70
et 71 de Andreu, 1991], par ailleurs déjà décrite dans la
région (Vivière, 1985) et caractérisant les étages
Turonien à Santonien, du moins en Algérie du NE (dès le
Cénomanien supérieur au Maroc). Il nous est toutefois absolument
impossible d'être affirmatif, devant la rareté des exemplaires
rencontrés (3 individus seulement). Ces spécimens
présentent par ailleurs une grande taille très
caractéristique.
Remarque : Plusieurs autres spécimens
n'ont en outre pas pu être identifiés (mauvais état de
conservation ou insuffisance de la documentation...), et nous le regrettons.
Ces organismes sont cependant tous exceptionnels, puisque rencontrés
à une ou deux reprises uniquement.
* Les références concernant ces deux
espèces ne sont pas données dans la liste bibliographique. Nous
nous contenterons de retenir qu'elles sont toutes deux citées par
Vivière (1985), qui en fourni aussi des illustrations.
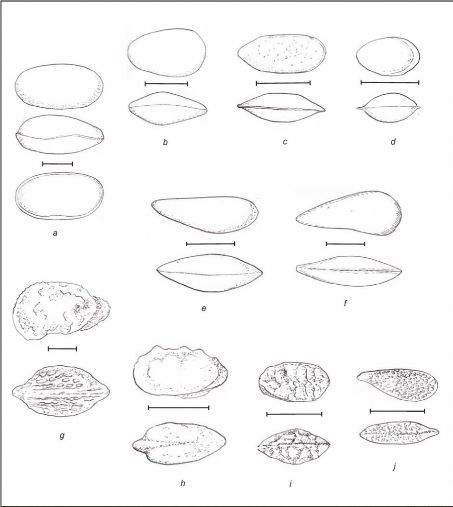
Planche 4 : Ostracodes.
Pour tous les spécimens, la barre d'échelle
représente 250 pm.
a : vue latérale droite (en haut), vue dorsale (au
milieu), vue latérale gauche (en bas),
b à j : vue latérale droite (en haut), vue
dorsale (en bas)
a et b : Cytherella sp.
(JONES), c : Cytherella sp. ? (JONES), d
: Phlyctocythere citreum (VIVIERE), e et
f : Paracypris sp.(SARS), g
et h : Spécimens appartenant à la
famille des Trachyleberididae, I : Amphicytherura (Sondagella)
distincta (BUTLER et JONES), j :
Procytherura ? sp. (WHATLEY).
Chap. III : Résultats et interprétations
|
O
i J [
|
O
O
|
|
O
1
|
_N. al
O o
O o
I. 1 I i .1
|
cri
O
O
|
O
|

O
O
O
1,3 --
--
Q
|
I
|
il
11
'
|
|
,
'
:1,
:
:
|
II.
1
i
I
I
I
|
1
,.1
i 1
1
i 1
|
,
|
0
P)
.,-
|
f I
,.,
il .
tl .
l
[
_
|
7--I
--
5 1
g
Ie
l I I il di
II
|
|
II
y
|
i
.
,
I
I
I
|
|
|
|
.
|
1
|
.
|
|
F
,
[
|
|
----\/----.----e\i-----/----------------
-'------,,,, j'--
|
|
|
_
..
6--.
|
|
·
|
·
|
|
·
·
.
·
|
|
·
·
·
|
·
|
L
|
|
|
|
|
·
·
·
·
·
|
.-
|
·
·
·
·
·
· l
·
·
·
·
|
IO
·
·
·
·
·
·
·
|
.-
.-
|
|
|
|
|
.__.
|
Q
--
ro cc
Figure 12 : Log stratigraphique et paramètres de
tri.
c-2 -- Les foraminifères
Abondantes et diversifiées, les faunes de
foraminifères peuvent apporter de nombreuses informations :
l'étude des foraminifères planctoniques est
généralement très utile pour les interprétations
stratigraphiques, au contraire des foraminifères benthiques qui, de part
leur longévité généralement importante, ne sont le
plus souvent que peu significatifs de ce point de vue (Benkherouf, 1988),
nous rechercherons également à reconstituer les
paléoenvironnements en nous basant sur l'ensemble des données
acquises.
L'évolution de tous les paramètres cités
(ratio benthique / planctonique, diversité, proportion des
différents types d'organismes...) est présentée dans les
figures 12 et 14.
· Ratio benthique
/planctonique
Un peu plus de la moitié des
foraminifères triés (54%) sont des foraminifères
planctoniques, ce qui est compatible avec le chiffre avancé par
Benkherouf (1988) dans la région du Dj. Dyr (pourcentage estimé
à 50 %). La variabilité est grande cependant, entre des
assemblages nettement dominés par des foraminifères planctoniques
(plus de 60 % voire même jusqu'à 100 % des organismes), des
assemblages mixtes et d'autres enfin où les populations benthiques sont
majoritaires.
Nous distinguerons donc 3 types d'assemblages :
type 1 : population planctonique majoritaire (> 42 % des
échantillons),
type 2 : population mixte (;---' 18 %),
type 3 : population benthique majoritaire (z 40 %).
· Evolution de la diversité
spécifique (fig.12(c))
Classée en trois catégories, faible (< ou =
à 5), moyenne (< ou = à 10) ou grande (> 15), la
diversité est très variable selon les niveaux.
Généralement très faible dans les assemblages de type 1
(planctoniques), elle peut être nettement plus élevée,
notamment dans les assemblages de types 2 (mixtes), dans lesquels
jusqu'à 20 espèces ont pu être reconnues.
Le Cénomanien inférieur
débute par des assemblages de type 1, à diversité
très réduite, mais rapidement la diversité
s'accroît, pour atteindre les valeurs les plus élevées de
l'ensemble du Cénomanien.
La transition avec le Cénomanien moyen
est marquée par des assemblages pauvres, auxquels
succèdent des assemblages plus riches (jusqu'à 12 espèces
reconnues), mais la grande diversité du Cénomanien
inférieur n'est jamais égalée.
Enfin, le Cénomanien supérieur,
et notamment la transition avec le Turonien, est globalement
très pauvre en espèces. Il n'est pas rare qu'une à deux
espèces soient ultra- dominantes (aussi bien d'ailleurs des
espèces planctoniques dans les assemblages de type 1 que des benthiques
dans ceux de type 3).
· Richesse microfossilistique apparente
(nb microorganismes / g trié) (fig.120)
Elle reflète essentiellement la richesse plus ou moins
grande en éléments fins (< 63 gm). La moyenne de 450 individus
par gramme cache mal une très grande variabilité, avec deux
extrêmes très éloignés (de 32 à près
de 3000 individus par g !). Dans la zone du Dj.
Dyr, Benkherouf (1988) évoque quant à elle une
richesse microfossilistique (réelle) particulièrement
basse, de seulement 40 individus par gramme en moyenne.
Plusieurs remarques peuvent cependant être faites :
les niveaux caractérisés par des assemblages de
type 1 (planctoniques) sont généralement ceux où la
richesse apparente est la plus élevée. La proportion en
éléments fins doit donc y être beaucoup plus importante,
au contraire, les niveaux les plus riches en organismes
benthiques (foraminifères mais aussi ostracodes) sont globalement
beaucoup plus pauvres en espèces. Le corollaire logique est que la
proportion en éléments fins y est nettement moindre,
il existe malgré tout quelques exceptions à ces
deux constatations, notamment pour certains niveaux du Cénomanien
terminal, où la richesse apparente est basse, malgré une
proportion élevée en organismes benthiques.
· Foraminifères planctoniques :
variation et caractérisation
Avec plus de 99 % des individus observés, les
espèces globuleuses dominent outrageusement les assemblages de
foraminifères planctoniques (fig.14). Les genres
Globigerinelloidesb et Hedbergella sont parmi les
plus fréquents, accompagnés, à certains niveaux, par de
très nombreux Heterohelixc (30 % environ en moyenne,
mais jusqu'à plus de 90 % dans les niveaux de transition avec le
Turonien). Les espèces du genre Whiteinella, également
présentes, sont plus rares, de même que les Favusella
(voir en figure 13 les clés de détermination des principaux
genres rencontrés).
Ont été reconnues les espèces :
- Hedbergella planispira (TAPPAN, 1940), H.
delrioensis (CARSEY, 1926), H.
simplex (MORROW, 1934) [rare],
- Heterohelix moremani (CUSHMAN, 1938) [espèce
fréquente, et parfois accompagnée d'autres types
d'hétérohélicidés, nettement plus globuleux],
- Favusella washitensisd (CARSEY, 1926)
[espèce rare, Cénomanien inférieur],
- Whiteinella archeocretacea (PESSAGNO, 1967), W.
baltica (DOUGLAS et RANKIN, 1969), W.paradubia (SIGAL, 1952b)
[Cénomanien supérieur].
D'autres espèces ont très certainement
échappé à notre diagnostic (présence de
pseudo-Schackoines ?) et beaucoup ne sont absolument pas identifiables du fait
d'une trop mauvaise conservation.
On l'aura compris, si les espèces globuleuses sont
tellement abondantes, en contrepartie, celles carénées sont
particulièrement rares 1 (à peine 1 %). Seulement
1/4 des échantillons triés ont
révélé la présence d'espèces
carénées (du genre Rotaliporaa), par ailleurs
jamais très nombreuses (2 à 3 spécimens en moyenne dans
chacun des échantillons concernés). Il s'avère
malheureusement (pour nous !) que ce sont essentiellement des espèces
carénées qui sont utilisées pour les déterminations
biostratigraphiques, dans la période considérée en tous
les cas (fig.9, 16).
Seules les espèces Rotalipora appenninica
(RENZ, 1936) et R. brotzeni (SIGAL, 1948) ont pu être mises
en évidence de façon formelle. Les autres spécimens, a
priori des rotalipores également, n'ont pas pu être
déterminés plus précisément, à cause d'un
état de conservation largement insuffisant. L'espèce R.
cushmani (MORROW, 1934) notamment n'a pas été
identifiée avec certitude.
1
Dans la région du Dj. Dyr, Benkherouf (1988) parle
quant à elle de 20 % d'espèces carénées.
|
Morphologie du test
|
Type d'ouverture
du test
|
Présence
d'une
carène
?
|
Familles
|
Genres
|
Illustration - Exemples
|
|
Bi-sérié
|
Terminale
|
non
|
Heterohelicidae
|
Heterohelix
|
Heterohelix moremani
|
|
Planispiral é
(enroulement selon
un
plan)
|
O-E
|
non
|
Planomalidae
|
Globigerinelloides
|
Globigerinelloides blowi
1
|
|
Trochospiralé
(enroulement selon
un axe)
|
0*-E*, presque P*
+
lèvre
|
non
|
Globotruncanidae
|
Hedbergella
|
. Hedbergella
delrioensis
. _
|
|
O-E, presque P
+ lèvre
+
ouvertures
supplémentaires sur le
côté ombilicale
|
oui
|
Globotruncanidae
|
Rotalipora
|
e-
- - i e Rotalipora
,
· '- -. reicheli
r ' -
)) ti'Èef
4,- . '
|
|
O-E
+ orticus
p
|
non
|
Globotruncanidae
|
Whiteinella
|
Whiteinella archeocretacea
.
|
* O = Ombilicale, E = extraombilicale, P =
périphérique
Figure 13 : Clés de détermination des
principaux genres de foraminifères planctoniques
rencontrés.
· Foraminifères
benthiques
Avec plus de 40 espèces reconnues, les populations de
foraminifères benthiques sont très variées, mais leur
répartition n'est pas homogène tout le long du profil. En effet,
entre les niveaux inférieurs du Cénomanien à population
benthique riche et diversifiée, et les niveaux supérieurs
où le benthos se réduit à de rares espèces
dominantes, la différence est grande.
Dans ce qui suit, nous différencierons deux grands
types de foraminifères benthiques, les calcaires (BC) et les
agglutinés2 (BA), dont le mode de formation est radicalement
différent. 60 % environ des foraminifères benthiques sont
de type agglutiné (avec une teneur en BA > 70 % pour plus de
la moitié des échantillons) [fig.14]. Les foraminifères de
type calcaire sont donc beaucoup moins fréquents, et ne dominent
réellement que dans 1/4 environ des échantillons
triés, essentiellement dans des niveaux du Cénomanien
inférieur.
Essai de classification des individus
rencontrés :
(basé sur les classifications de Loeblich et Tappan
(1987, 1992), et sur les modifications ultérieures proposées par
Lee et al. (2000) et Kaminsky (2004) pour ce qui concerne les
foraminifères agglutinés)
0 Parmi les foraminifères agglutinés,
on retrouve :
Ammobaculites sp. (CUSHMAN), Ammobaculites turonicus
(SAID et KENAWY), Ammodiscus cretaceus (REuSS), Ammomarginulina ? sp.
(LOEBLICH et TAPPAN), Dorothia oxycona (REuSS), Eggerella sp.
(CUSHMAN), Gaudryinoides (GEODAKCHAN), Textularia chapmani
(LALICKER), Textularia rioensis (CARSEY), Thomasinella punica
(SCHLUMBERGER), Tritaxia pyramidata (REuSS), Trochamminoides sp.
(CUSHMAN)...
Les foraminifères benthiques agglutinés sont
essentiellement composés de Lituolidés
(majoritairement du sous-ordre des Lituolina),
accompagnés, à certains niveaux, par des
Textularidés en grand nombre.
Classification des organismes identifiés :
Classe FORAMINIFEREA d'Orbigny, 1826 Sous-classe
TEXTULARIIA Mikhalevich, 1980
Ordre ASTRORBIZIDA Lankester, 1885
Sous-ordre AM1VIODISCINA Mikhalevich, 1980
Super-famille AMMODISCACEA Reuss, 1862
Famille AMMODISCIDAE Reuss, 1862
Sous-famille AMMODISCINAE Reuss, 1862
Genre Ammodiscus Reuss, 1862
Ammodiscus cretaceus (Reuss, 1845) p1.5.1
|
Ordre LITUOLIDA Lankester, 1885
|
Photos a à e: benthiques calcaires
Photos f et g : benthiques agglutinés
|
|
2
Selon les auteurs, on trouvera agglutinés =
agglutinants = arénacés.
|
|
Sous-ordre LITUOLINA Lankester, 1885
Super-famille LITUOTUBACEA Loeblich et Tappan,
1984
Famille TROCHAMIVHNOIDAE Haynes et Nwabufo-Ene,
1998
Genre Trochamminoides Cushman, 1910
Trochamminoides sp. (Cushman, 1910) .p1.5.2 et 3 ?
Super-famille LITUOLACEA de Blainville, 1827
Famille LITUOLIDAE de Blainville, 1827
Sous-famille AMMOMARGINULINAE Podobina, 1978*
Genre
Ammobaculites Cushman, 1910
Ammobaculites sp. (Cushman, 1910) p1.5.4
Ammobaculites turonicus (Said et Kenawy, 1956)
..p1.5.5
Genre Ammomarginulina Wiesner, 1931
Ammomarginulina ? sp. (Wiesner, 1931) p1.5.6
Sous-ordre VERNEUILININA Mikhalevich et
Kaminski, 2004
Super-famille VERNEUILINACEA Cushman, 1911
Famille TRITAXIIDAE Plotnikova, 1979*
Genre Tritaxia Reuss, 1860
Tritaxia pyramidata (Reuss, 1863) p1.5.7
Famille REOPHACELLIDAE Mikhalevich et Kaminski,
2004
Sous-famille SPIROPLECTINATINAE Cushman, 1928
Genre Gaudryinoides Geodakchan, 1969*
Gaudryinoides sp. (Geodakchan, 1969)* p1.5.8
Ordre TEXTULARIIDA Delage et Herouard, 1896
Sous-ordre TEXTULARIINA Delage et Herouard,
1896
Super-famille EGGERELLACEA Cushman, 1937
Famille EGGERELLIDAE Cushman, 1937
Sous-famille DOROTHIINAE Balakhmatova, 1972*
Genre Dorothia Plummer, 1931
Dorothia oxycona (Reuss, 1860) p1.5.9
Sous-famille EGGERELLINAE Cushman, 1937
Genre Eggerella Cushman, 1935
Eggerella sp. (Cushman, 1935) .p1.5.14 ?
Super-famille TEXTULARIACEA Ehrenberg, 1838
Famille THoMASINELLIDAE Loeblich et Tappan,
1984
Genre Thomasinella Schlumberger, 1893
Thomasinella punica (Schlumberger, 1893) p1.5.12
Famille TEXTULARIIDAE Ehrenberg, 1838
Sous-famille TEXTULARIINAE Ehrenberg, 1838 Genre Textularia Defrance,
1824
Textularia chapmani (Lalicker, 1935) p1.5.10
Textularia rioensis (Carsey, 1926) p1.5.11
* ces différents genres, espèces ou familles sont
cités par Kaminsky (2004).

Planche 5 : Foraminifères benthiques
agglutinés.
1. Ammodiscus cretaceus
(REUSS), 2. et 3 ?.
Trochamminoides sp. (CUSHMAN), 4.
Ammobaculites sp. (CUSHMAN), 5.
Ammobaculites turonicus (SAID et
KENAWY), 6. Ammomarginulina ? sp.
(WIESNER), 7. Tritaxia pyramidata
(REUSS), 8. Gaudryinoides sp.
(GEODAKCHAN), 9. Dorothia oxycona
(REUSS), 10. Textularia chapmani
(LALICKER), 11. Textularia rioensis
(CARSEY), 12. Thomasinella punica (SCH
LU M BERGER), 13.indéterminé 1, 14. Eggerella ? sp.
(CUSHMAN).
(c) Les foraminifères benthiques calcaires
reconnus sont les suivants :
Cassidella sp.(KHAN) = Coryphostoma, Dentalina
sp. (RISSO), Gavelinella intermedia (BERTHELIN), Gavelinella
cenomanica (BROTZEN), Globulina prisca (REuSS), Gyroidinoides nitidus
(REuSS), Lagena apiculata (REuSS), Lenticulina rotulata (LAMARCK),
Nodosaria sp. (LAMARCK), Oolina sulcata (WALKER et JACOB),
Planularia complanata (REuSS), Quinqueloculina sp.(d'ORBIGNY),
Spiroloculina sp. (d'ORBIGNY), Tristix acutangulum (REuSS),
Tristix excavatum (REuSS), Valvulineria sp. (CUSHMAN).
Les organismes les plus fréquemment rencontrés sont
des Rotalidés, les autres genres et espèces
restant anecdotiques dans la plupart des niveaux.
Classification des organismes rencontrés :
Classe FORAMINIFEREA d'Orbigny, 1826
Ordre LAGENIDA Lankester, 1885
Super-famille NODOSARIACEA Ehrenberg, 1838
Famille NODOSARIIDAE Ehrenberg, 1838
Sous-famille NODOSARIINAE Ehrenberg, 1838
Genre Dentalina Risso, 1826
Dentalina sp. (Risso, 1826) p1.6.1
Genre Nodosaria Lamarck, 1812
Nodosaria sp. (Lamarck, 1812) p1.6.2 et 3
Genre Tristix Macfayden, 1941
Tristix acutangulum (Reuss, 1863) p1.6.7
Tristix excavatum (Reuss, 1863) p1.6.8
Famille LAGENIDAE Reuss, 1862
Sous-famille LAGENINAE Reuss, 1862
Genre Lagena Walker et Jacob, 1798
Lagena apiculata (Reuss, 1851) p1.6.9
Famille VAGINULINIDAE Ehrenberg, 1838
Sous-famille LENTICULININAE Chapman, Parr et Collins, 1934
Genre Lenticulina Lamarck, 1804
Lenticulina rotulata (Lamarck, 1804) p1.6.10
Sous-famille MARGINULININAE Wedekind, 1937
Genre Planularia Defrance, 1826
Planularia complanata (Reuss, 1845) p1.6.11
Super-famille POLYMORPHINACEA d'Orbigny, 1839
Famille POLYMORPHINIDAE d'Orbigny, 1839
Sous-famille POLYMORPHININAE d'Orbigny, 1839
Genre Globulina d'Orbigny, 1839
Globulina prisca (Reuss, 1863) p1.6.12
Famille ELLIPSOLAGENIDAE A. Silvestri, 1923
Sous-famille OOLININAE Loeblich et Tappan, 1961
Genre Oolina d'Orbigny, 1839
Oolina sulcata (Walker et Jacob, 1798) p1.6.13
Ordre MILIOLIDA Lankester, 1885
Sous-ordre MILIOLINA Delage et Hérouard,
1896
Super-famille MILIOLACEA Ehrenberg, 1839
Famille SPIROLOCULINIDAE Wiesner, 1920
Genre Quinqueloculina d'Orbigny, 1826
Quinqueloculina sp. (d'Orbigny, 1826) .p1.6.15
Genre Spiroloculina d'Orbigny, 1826
Spiroloculina sp. (d'Orbigny, 1826) p1.6.16
Ordre ROTALIIDA Lankester, 1885
Super-famille CHILOSTOMELLACEA Brady, 1881
Famille GAVELINELLIDAE Hofker, 1956 Genre Gavelinella Brotzen, 1942
Gavelinella intermedia (Berthelin, 1880) p1.7.1
Gavelinella cenomanica (Brotzen, 1945) p1.7.2
Genre Gyroidinoides Brotzen, 1942
Gyroidinoides nitidus (Reuss, 1845) .p1.7.3, 4 ( ?)
Genre Valvulineria Cushman, 1926
Valvulineria sp. (Cushman, 1926) p1.7.5
Super-famille BULIMINACEA Jones, 1875
Famille BOLIVINITIDAE Cushman, 1927
Sous-famille FURSENKOININAE Loeblich et Tappan, 1961 Genre
Fursenkoina (Loeblich et Tappan, 1961)
La détermination du genre observé est
particulièrement problématique, puisque sa dénomination
est variable selon les auteurs. On trouve ainsi :
Fursenkoina viscida (Khan) selon Holbourn et alii
(2001) p1.6.7
Cassidella viscida (Khan, 1950), selon Magniez-Jannin
(1975)
Coryphostoma? sp. selon Moullade et alii. (2008)
En plus de ces organismes, un certain nombre d'autres n'ont
pas pu être déterminés, dont des foraminifères, et
divers organismes (algues, calcisphères, mollusques ?? (etc)). Les
planches 5,6 et 7 en donnent plusieurs illustrations.
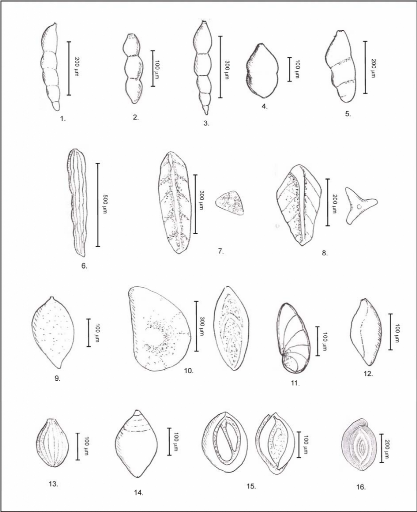
Planche 6 : Foraminifères benthiques
calcaires.
1. Dentalina sp. (RISSO), 2
et 3. Nodosaria sp.
(LAMARCK), 4.Pseudonosaria aff. Humilis?,
5.indéterminé 2, 6. Nodosaria aff.
Chapmani ?, 7. Tristix acutangulum
(REUSS), 8. Tristix excavatum (REUSS),
9. Lagena apiculata (REUSS), 10.
Lenticulina rotulata (LAMARCK), 11.
Planularia complanata (REUSS), 12.
Globulina prisca (REUSS), 13. Oolina
sulcata (WALKER et JACOB), 14.indéterminé 3, 15.
Quinqueloculina sp. (D'ORBIGNY), 16.
Spiroloculina sp. (D'ORBIGNY).

Planche 7 : Foraminifères benthiques calcaires
(suite).
1. Gavelinella intermedia (BERTHELIN), 2.
Gavelinella cenomanica (BROTZEN), 3.
Gyroidinoides nitidus (REUSS), 4.
Gyroidinoides sp. ?, 5. Valvulineria sp.
(CUSHMAN), 6.indéterminé, 7. Cassidella
(KHAN) = Cotyphostoma? sp.,
8.indéterminé 4 = Gubkinella ??,
9.indéterminé 5, 10.indéterminé 6,
11.indéterminé 7 = Bonetocardiella ??,
12.indéterminé 8, 13.indéterminé
9.
Le tableau qui suit détaille la répartition et
l'abondance des différents types de foraminifères benthiques
déjà cités ci-dessus (tab.2).
|
Organisme
|
Planche
|
Répartition
|
Abondance
|
|
Vrac. --
Cénomanien
inf.
|
Cénomanien
moy.
|
Cénomanien
sup.
|
|
Ammobaculites sp.
(CUSHMAN)
|
5.4
|
|
|
|
*
|
|
Ammobaculites
. 1 ,
turorucus (SAID et
KENAWY)
|
5.5
|
|
|
.(
|
*
|
|
Ammodiscus cretaceus 2(REUSS)
|
5.1
|
|
|
|
*
|
|
Ammomarginulina ?
sp. (LOEBLICH et TAPPAN)
|
5.6
|
|
|
|
***
|
|
Dorothia oxycona
(REuSS)
|
5.9
|
|
|
|
*
|
|
Eggerella ? sp.3
(CUSHMAN)
|
5.14
|
|
|
|
***
|
|
Gaudryinoides
(GEODAKCHAN)
|
5.8
|
|
|
|
*
|
|
Textularia chapmani
(LALICKER)
|
5.10
|
|
|
|
*
|
|
Textularia rioensis
(CARSEY)
|
5.11
|
|
|
|
**
|
|
Thomasinella punica4
(SCHLUMBERGER)
|
5.12
|
|
|
|
*
|
|
Tritaxia pyramidata
(REuSS)
|
5.7
|
|
|
|
*
|
|
Trochamminoides sp.
(CUSHMAN)
|
5.2, 3 ?
|
|
|
|
***
|
|
Cassidella
sp. . (KH AN) = Coryphostoma
|
7.7
|
|
|
|
**
|
|
Dentalina sp. (RISSO)
|
6.1
|
|
|
|
*
|
|
Gavelinella intermedia
(BERTHELIN)
|
7.1
|
|
|
|
**
|
|
Gavelinella cenomanica
(BROTZEN)
|
7.2
|
|
|
|
**
|
|
Globulina prisca
(REuSS)
|
6.12
|
|
|
|
*
|
|
Gyroidinoides nitidus (REUSS)
|
7.3
|
|
|
|
***
|
|
Lagena apiculata
|
6.9
|
|
|
|
**
|
|
(REuSS)
|
|
|
|
|
|
|
Lenticulina rotulata
(LAMARCK)
|
6.10
|
|
|
|
*
|
|
Nodosaria sp.
(LAMARCK)
|
6.2, 3
|
|
|
|
*
|
|
Oolina sulcata
(WALKER et JACOB)
|
6.13
|
|
|
|
*
|
|
Planularia complanata (REUSS)
|
6.11
|
|
|
|
*
|
|
Quinqueloculina sp.(d'ORBIGNY)
|
6.15
|
|
|
|
*
|
|
Spiroloculina sp. 5
(d'ORBIGNY)
|
6.16
|
.(
|
.(
|
.(
|
**
|
|
Tristix acutangulum
(REuSS)
|
6.7
|
|
|
|
*
|
|
Tristix excavatum
(REuSS)
|
6.8
|
.(
|
|
|
*
|
|
Valvulineria sp.
(CUSHMAN)
|
7.5
|
|
|
|
**
|
|
Indéterminé 1
|
5.13
|
|
|
|
*
|
|
Indéterminé 2
|
6.5
|
|
|
|
*
|
|
Indéterminé 3
|
6.14
|
|
|
|
*
|
|
Indéterminé 4
|
7.8
|
.(
|
|
|
*
|
|
Indéterminé 5 6
|
7.9
|
|
|
|
*
|
|
Indéterminé 6 6
|
7.10
|
|
|
|
*
|
|
Indéterminé 7
7
Bonetocardiella ??
|
7.11
|
|
|
|
*
|
|
Indéterminé 8
|
7.12
|
|
|
|
*
|
|
Indéterminé 9
8
|
7.13
|
|
|
|
*
|
Degré d'abondance : * rare ** commun ***
très fréquent
Tableau 2 : Détail de la répartition et de
l'abondance des foraminifères benthiques.
1 : Ammodiscus cretaceus est une
espèce rare. Seul un petit nombre de spécimens a
été observé dans le Cénomanien inf. (n°30) et
sup. (n°94).
2 : Ammobaculites turonicus a
été décrit uniquement à un niveau dans le
Cénomanien sup. (n°73).
3 : Le genre présenté comme
Eggerella (?) est particulièrement abondant à certains
niveaux du Cénomanien sup. Ses structures externes étant
particulièrement difficiles à distinguer, sa détermination
est difficile.
4 : L'espèce Thomasinella punica
n'est abondante qu'à deux niveaux du Cénomanien moyen
(n°52 et 54), et exceptionnelle ailleurs (n°36 du Cénomanien
inf.). Notez sa taille exceptionnellement grande pour un foraminifère
(plusieurs mm).
5 : Les foraminifères du genre
Spiroloculina constituent presque 100 % des formes observées
pour l'échantillon n°36 (également très riche en
ostracodes). Ailleurs, ils sont rares.
6 : Ces formes n'ont été
observées que dans de rares niveaux du Cénomanien moyen. Il ne
s'agit probablement pas de foraminifères (calcisphères (indet.5)
ou autre ?).
' : Cette forme, qui rappelle une calpionelle ( Albien) n'a
été observée que dans de rares niveaux du début du
Cénomanien inf. Il pourrait s'agit en réalité d'un type de
calcisphère, apparenté au genre Bonetocardiella
figuré par Robaszynski et al. (1993).
8 : Un seul échantillon du
Cénomanien moyen (n°52) a révélé de telles
formes, facilement repérables par leur grande taille, d'autant plus
qu'elles sont présentes en grand nombre au côté de
l'espèce Thomasinella punica. Là
encore, il ne s'agit probablement pas de foraminifères (mollusques
??).
La plupart des espèces décrites semble
caractériser des milieux de plate-forme externe ou de talus (Benkherouf,
1988). Aucune espèce franchement littorale3 n'a
été observée, pas plus que des espèces typiques de
milieux profonds.
Page suivante, se trouve une planche (pl. 8) regroupant
quelques clichés de microfossiles pris au MEB, incluant des ostracodes
(photos A à G), des foraminifères benthiques, calcaires (H et J)
ou agglutinés (I) ; et d'autres planctoniques globuleux (K à O).
Ces quelques formes ne constituent qu'une faible proportion du total des formes
observées, et ont été sélectionnées
uniquement en rapport avec leur bon état de conservation.
Légende de la planche 8
:
A et B : Spécimens apparentés
à Paracypris sp.(SARS), vue latérale. C
: Procytherura ? sp. (WHATLEY), vue
latérale.
D : Spécimen appartenant à la
famille des Trachyleberididae, vue latérale.
E : Idem, vue dorsale.
F : Cytherella sp. (JONES),
vue latérale droite.
G : Cytherella sp. (JONES),
vue latérale gauche.
H : Lenticulina sp.
(LAMARCK).
I : Textularia sp.?
(DEFRANCE).
J : Nodosaria sp.
(LAMARCK).
K et L : Hedbergella sp.
(BRONNIMANN et BROWN)
M : Heterohelix moremani
(CUSHMAN).
N: Hedbergella sp. (BRONNIMANN
et BROWN)
m : Whiteinella sp.
(PESSAGNO)
3
au niveau du Dj. Dyr, Benkherouf (1988) a décrit deux
espèces considérées comme allochtones, à milieu de
vie plutôt
superficiel, à savoir Charentia cuvillieri et
Archaecyclus cenomaniana., Pour notre part, ces espèces n'ont
pas été observées.
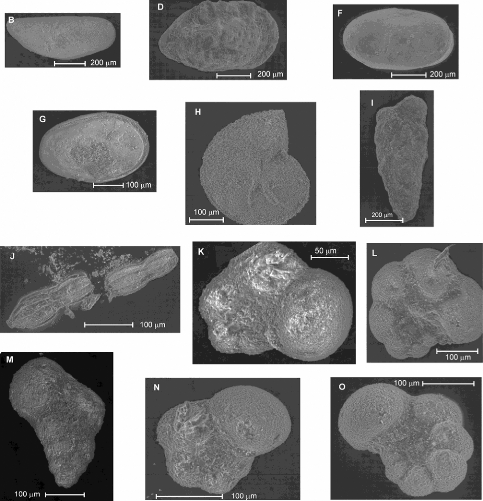
G
|
B
|
e
|
w7
|
.. .- ... " ;" '-. , '' ' .
|
|
|
i_ ii 200um
|
|
1 .
|
Fele
|
1-.1 200um
1-1 200 gm
Planche 8 : Illustrations de quelques microfossiles au
MEB.
(clichés pris au laboratoire de Tautavel,
Université de Perpignan, France)
En figure 14 (page suivante) sont présentés divers
paramètres (obtenus à l'issu du tri des microfossiles), dont
certains méritent une explication :
0 Pourcentage de foraminifères benthiques (B),
planctoniques (P) et d'ostracodes (0).
(c) Proportion de foraminifères benthiques (B)
et de foraminifères benthiques de type agglutiné
(BA).
G Proportion de foraminifères planctoniques
(P) et de foraminifères planctoniques de type Heterohelix (PH)
:
Ce type de foraminifères globuleux domine parfois
à certains niveaux, notamment en toute fin
du Cénomanien
où ils constituent la presque totalité des foraminifères
planctoniques observés.
(c) Rapport (benthique agglutiné (BA) /total
des benthiques (B)) :
Comme en (c), la prédominance de ce type de
foraminifères parmi les organismes benthiques apparaît nettement
(plus de 65 % du total). Les foraminifères benthiques de type calcaire
ne dominent qu'à de rares niveaux.
G Rapport (planctonique globuleux (PG) /total des
planctoniques (P)) :
Ce type de foraminifères planctoniques domine très
nettement face à ceux de type caréné, extrêmement
rares.
(c) Rapport (planctonique/total des
foraminifères) :
L'évolution en dents de scie montre une
variabilité importante, avec des niveaux nettement dominés par
des foraminifères planctoniques (ceux-ci dominent nettement à 4
reprises au moins) et d'autres au contraire par des foraminifères
benthiques.
(c) Rapport (foraminifères total (F)/
ostracodes (0)) :
Généralement rares, les ostracodes
représentent parfois, comme on le voit, la majorité des
microorganismes observés.
Chap. III : Résultats et interprétations

rn
O
fn
o
2
o
O
0,
O O
|
113
|
|
P 1
,..
0 ,
il II
Il II
11 .I.
I
|
|
k Ili ill I.L I i I lui
,00,,,,Iimilpoliiii,[iiiiii,Fili
iiii, i ,,I[1111[111111[11111111111111111
Ill 111/I11[111111111111111D111[111
11111111[1111[1111 I 1 1111 1 1 r
111
|
l
:i
|
i,. l'el
il
II
'I!
|
ij
Ii
il
Il
I '
|
Hu
1
!
|
ii 1 II
, [Hum
HI
[ 11 1> 11111
r1111111
1111111
|
r
l'
1' 1111
|
itli$
ri
il g
I
1 I
lill
y.
'MN
|
|
|
l
|
!
|
|
|
_
_
_
_
_
|
|
|
_
_
-
-
_
|
|
|
|
_
_
|
|
to
>
|
|
\
|
|
|
_
_
-
_
|
|
|
|
|
|
_
|
|
|
|
|
|
|
|
|
_
|
|
_
|
_ É
in--.
_-
·.-__,_
|
-
-
|
|
|
|
|
_
_
|
1
|
..
-
-
|
|
_
_
_
_
_
_
|
----- ---_______._.----------- \ -_ 1
. 1
|
- _ - -
|
cs) O
1
0
w 0
-n
o
O
É 1 i l 3 e
Figure 14 : Log stratigraphique et paramètres de
tri (suite).
d- Etude des lames minces
Vu la nature essentiellement marneuse des dépôts,
seul un petit nombre de lames minces (16 au total) a été
réalisé dans les niveaux les plus indurés, surtout dans le
Cénomanien terminal ou au tout début du Turonien. Il aurait pu
être intéressant d'indurer certains échantillons plus
tendres afin d'en étudier le microfaciès, mais cette technique
n'a pas pu être exploitée dans le cadre de notre étude.
Dans ce qui suit, une description succincte de chaque lame va
être proposée, illustrée de nombreux clichés
(planches 9, 10 et 11). Dans cette description, les classifications communes de
Folk (1962) et de Dunham (1962) seront couramment employées.
Ces lames, réalisées le plus souvent dans du
calcaire marneux, et plus rarement dans du calcaire franc (Cénomanien
moyen et Turonien inférieur) montrent des caractéristiques
communes : il s'agit toujours de calcaire à texture micritique, de
teinte souvent sombre (excepté dans le Turonien inférieur
où les teintes sont généralement plus claires). Le contenu
en microfossiles et les microfaciès sont cependant fort
différents d'une lame à l'autre.
PLANCHE 9 (page suivante) Vraconnien
Lame 1B (2 clichés)
Calcaire micritique sombre, de type
packstone à wackestone, à
calcisphères nombreuses, foraminifères
planctoniques globuleux (Hedbergella, Globigerinelloides) et rares
ostracodes. Des cubes de pyrite sont bien visibles en remplissage de certaines
formes (photo 1). De rares `lentilles' de matière organique sont
présentes.
Certaines parties de la lame montrent une succession
caractéristique de microbancs clairs, riches en calcisphères,
alternant avec des bancs sombres, contenant moins de calcisphères et
proportionnellement plus de foraminifères planctoniques (photo 2).
Cénomanien inférieur
Lame 15
Calcaire micritique sombre, de type
wackestone, à calcisphères, foraminifères
planctoniques globuleux, petits foraminifères benthiques, ostracodes et
fragments d'échinodermes. La présence d'oxydes de fer, et surtout
de glauconie est remarquable (cette dernière a d'ailleurs
été décrite précédemment dans un
échantillon contigu (n°16)).
Les foraminifères planctoniques reconnues sont les
suivants : Hedbergella simplex, H. delrioensis, Heterohelix sp.,
Globigerinelloides sp.
Cénomanien moyen Lame
46
Calcaire micritique, de type wackestone,
à nombreux débris de bivalves,
calcisphères et rares foraminifères, souvent très
recristallisés, donc impossibles à identifier.
Lame 62 (2 clichés)
Calcaire micritique, de type
wackestone à packstone, à nombreux
débris (bivalves, échinodermes), foraminifères
benthiques (miliolidés, Lenticulina sp....),
calcisphères, ostracodes et rares foraminifères planctoniques
(globuleux). Dans certaines parties de la lame, des traces de
bioturbation sont visibles (photo 2).
Lame 74 (aucune photo)
Calcaire micritique, de type
wackestone, à débris
d'organismes (bivalves, gastéropodes, échinodermes),
foraminifères benthiques et calcisphères. Cette lame est
globalement très proche de la précédente, excepté
une richesse fossilifère moindre. Des traces de bioturbation
sont également décelables.
|
|
Chap. III : Résultats et interprétations
|
|
Lame 1 B (1)
|
Lame 1 B (2)
|
|
|
|
Lame 15
|
|
Lame 46
|

Légende : B : foraminifère benthique, 0 :
ostracode, GI : glauconie, Hed : Hedbergella
sp.
Planche 9 : Lames minces (Vraconnien -- Cénomanien
moyen).
PLANCHE 10 (page suivante)
Cénomanien supérieur
Lame 95 : photo 1
Calcaire micritique sombre, de type
mudstone, à foraminifères planctoniques
globuleux (Hedbergella, Whiteinella, Heterohelix). La lame est
encombrée de nombreuses lentilles de matière organique
sombres.
Lame 99 : photo 2
Calcaire micritique, de type
mudstone, d'une couleur beaucoup plus claire que
l'échantillon précédent et visuellement très
différent (absence totale cette fois-ci de lentilles de MO). Les rares
microorganismes présents sont des calcisphères, des
foraminifères planctoniques globuleux (Whiteinella, Heterohelix
probables) et des benthiques calcaires. La présence
d'oxydes de fer est également marquante.
Cénomanien supérieur -- faciès
Bahloul4 Lame 103 (2 clichés)
Calcaire micritique sombre, de type
wackestone à packstone, très riche en microorganismes,
principalement des foraminifères planctoniques globuleux, mais
également en petits foraminifères benthiques, ostracodes,
débris d'échinodermes et rares filaments (= petits bivalves
fins). La présence de glauconie et de lentilles
de MO en grande quantité est également remarquable.
Les foraminifères planctoniques, pour la plupart d'une
grande taille exceptionnelle, comprennent Heterohelix sp.,
Hedbergella simplex, H.delrioensis, Globigerinelloides sp.
Lame 104 (2 clichés)
Calcaire micritique sombre, de type
wackestone, à foraminifères
planctoniques globuleux, également de grande taille :
Heterohelix sp., Hedbergella sp., Whiteinella sp., W.
paradubia. Des filaments, plutôt rares, sont également
présents, ainsi que des lentilles de MO en abondance.
En revanche, la glauconie, fréquente dans la lame
précédente, est totalement absente, de même que les
organismes benthiques.
Lame 95 Lame 99

Lame 103 (1)
Lame 104 (1) Lame 104 (2)

Op* nie.>
·
le
b
500 iam
·
·1111,..

Légende : B : Foraminifère benthique, H :
Heterohelix sp., Hed : Hedbergella sp., GI : Glauconie, W :
Whiteinella sp.
Planche 10 : Lames minces (Cénomanien
sup.).
PLANCHE 11 (page suivante) Lame 105 (2
clichés)
Calcaire micritique sombre, de type
wackestone, à foraminifères
planctoniques globuleux : Heterohelix sp. (majoritaires),
Hedbergella sp., Whiteinella sp.. Les traces de MO
sont abondantes (lentilles).
Remarque: cette lame a été
réalisée parallèlement à la stratification du fait
d'un délitage en fins
feuillets. Les informations déduites en sont donc
nécessairement amoindries. Lame 106 (2 clichés)
Calcaire micritique sombre, de type
mudstone à wackestone, à
foraminifères planctoniques globuleux (Hedbergella
sp., Whiteinella sp., rares Heterohelix sp.),
calcisphères, rares filaments et lentilles de MO
nombreuses. La richesse en microfossiles est nettement moins
élevée que dans la lame précédente. Des formes
assez mystérieuses sont également observées,
peut-être s'agit-il de fragments d'échinodermes (ophiures ??).
Turonien inférieur
Lames 110, 114, 118, 120, 122 (3 clichés)
Nous avons choisi de traiter ces différentes lames
ensemble car elles présentent un microfaciès très
proche.
Il s'agit de calcaire micritique, clair, de
type mudstone, à rares microorganismes,
souvent aux contours flous, donc très difficiles à
identifier. Sont présents des calcisphères, quelques ostracodes,
de très rares foraminifères planctoniques globuleux (dont des
Heterohelix) et d'aussi rares foraminifères benthiques.
Les oxydes de fer sont fréquents, en
taches ou en remplissage de microfractures (stylolithes). Ce genre de structure
est d'ailleurs commun (lame 120).
Des auréoles sombres, finement ciselées, entourent
parfois certains microorganismes (oxydes ? lame 118).
Lame 116: photo ci-dessous

Calcaire micritique, de type
mudstone, au sein duquel aucun microorganisme n'a pu
être observé. Le microfaciès, très différent
des lames précédentes, nous a conduit à décrire
cette lame séparément. On y observe en effet un litage
bien marqué (succession de lits sombres et d'autres lits
clairs), et une structure par endroit très perturbée, avec une
modification nette de l'orientation du litage (bioturbation ?).
Lame 105 (1) Lame 105 (2)

Légende :
H : Heterohelix sp., Hed :
Hedbergella sp., O : ostracode, Oph : ophiure (
?).
Planche 11 : Lames minces (Cénomanien sup.
--
Turonien inf.).


3. Interprétations
Nous aborderons successivement les aspects
biostratigraphiques, puis paléoenvironnementaux /
paléogéographiques. Suivra ensuite une description de la crise
cénomano-turonienne, ainsi que de la transition entre Vraconnien et
Cénomanien du début de coupe. La réponse à quelques
interrogations servira enfin de conclusion à ce chapitre.
a- Axe Biostratigraphie
Préciser la biostratigraphie est chose difficile quand
les déterminations sont incertaines et la documentation disponible trop
rare. L'extrême rareté des foraminifères planctoniques
carénés (et des ammonites) constitue également un handicap
pour une biostratigraphie précise, d'autant plus que les
spécimens observés ne sont pas toujours en excellent état
de conservation, donc parfois difficiles à déterminer
précisément.

,,,_
. ,-`' I -P,
te ft] m
_Q
Ç)
ir cr O
Tura nien inférieur
-
-
-
_
--
--
--
--
--
--
_r ri
-
-
Cénomanien moyen é supérieur
-
·
Cenomanien inférieur
Vraconnien
La figure ci-contre présente la répartition
|
106
|
|
des différentes espèces de foraminifères
|
104
|
|
planctoniques utiles pour la
|
100
|
|
biostratigraphie. Seuls certains
|
95
|
|
échantillons sont indiqués, ceux-ci ne
|
90
|
|
représentent qu'une petite partie du total
|
86
|
|
des échantillons (à titre d'exemple, seuls
|
80
|
|
14 échantillons sur 70 étudiés
contiennent des spécimens du genre
|
|
|
Rotalipora).
|
70
|
|
De plus, l'échelle verticale n'est pas respectée
(en réalité, l'épaisseur du
|
60
|
|
Cénomanien inférieur est largement
accrue).
|
50
|
|
La présence des espèces
appartenant aux genres Hedbergella,
Globigerinelloides
et Heterohelix n'est
pas précisée, ceux-ci
étant communs à
|
40
|
|
32
|
|
presque tous les niveaux.
|
28
|
|
24
|
|
20
|
|
14
|
|
11
|
|
2
|
Figure 15 : Foraminifères planctoniques --
Répartition
de quelques espèces-clés.
En référence à la biozonation du
Crétacé proposée par Robaszynski et Caron (1995) [fig.16],
il est donc possible de différencier, tant bien que mal, plusieurs zones
dans la coupe.
|
Périodes
|
Globotruncanidés
|
Hétérohélicidés
|
|
CONLACIEN
|
I
|
DICARINELLA concavata
^
|
nuttaifi
|
cane
|
s
|
PSEUDOTEXTULARIA
|
|
|
|
|
TURONIEN
|
S
|
|
HETEROHELIX
globulosa {=reussi)
|
|
n
|
|
M
|
MARE schneegansi
|
|
HFLV helvetica
|
'hely
|
|
I
|
|
WHIT. archaeocretacea
|
|
CENOMANIEN
|
S
|
|
ROTAL. cushmani
|
|
I ar
|
|
HE TEROHFLIX moremani
|
g
|
|
M -
|
ROTAL. reichefi
|
lcush
|
|
I
|
ROTAL.
globotruncartoides (= brotzeni)
|
r
|
|
ALBIEN
|
S
|
ROTAL. appenninica
|
A
gl.
|
|
ROTAL. ticinensis
|
ap.
--.%
|
|
m
|
|
ROTAL. subticinensis
-A
|
tic
|
|
TICIAt praeticinensis sb
|
ROTAL Rotalipora, Ticinelfa, VVHIT. Whiteineila,
HELV. Heivetoglobotruncana, MARG. Marginotruncana
Figure 16 : Biozonation du Crétacé moyen
à supérieur par les foraminifères
planctoniques.
(d'après Robaszynski et Caron, 1995)
Les différentes espèces observées
permettent de réaliser un découpage. La limite inférieure
du Cénomanien n'a pas pu être définie avec
précision, mais, en revanche, trois zones ont été
reconnues entre le début du Cénomanien et celui du Turonien :
- Cénomanien inférieur (zone à
Rotalipora brotzeni)
Sa limite inférieure (avec le Vraconnien) ne peut
être située précisément.
Cette zone comporte, en plus de nombreuses espèces des
genres Hedbergella et Globigerinelloides, les espèces
Favusella washitensis, Rotalipora appenninica, Heterohelix moremani et
plusieurs spécimens qui semblent bien appartenir à
l'espèce R. brotzeni.
C'est dans cette zone que la diversité
spécifique est la plus grande (fig.12), ce qui semble correspondre aux
dires de Hart et Ball (1986) qui parlent 'd'un accroissement rapide de la
diversité de la faune aux environs du Cénomanien moyen'.
Cénomanien moyen à supérieur (zone
à Rotalipora cushmani)
L'espèce index n'a pas été
identifiée avec certitude, mais l'entrée dans cette zone est
à supposer du fait de l'absence des espèces
précédentes (F.washitensis, R. appenninica et
R.brotzeni). Plusieurs spécimens, attribués au genre
Rotalipora, n'ont malheureusement pas pu être identifiés
avec plus de précisions à cause d'un trop mauvais état de
conservation. Les
derniers spécimens du genre Rotalipora sont
observés dans l'échantillon n°86, au niveau de la partie
finale du Cénomanien supérieur.
Par ailleurs, les espèces appartenant aux genres
Hedbergella, Heterohelix (H.moremani essentiellement) et
Globigerinelloides restent communes
- Partie finale du Cénomanien -- début du
Turonien (zone à Whiteinella archeocretacea)
Marquée par l'absence des spécimens du genre
Rotalipora et par l'apparition des Whiteinella (W. baltica, W.
paradubia et W. archeocretacea), cette zone est bien
caractérisée. Les genres Hedbergella et Heterohelix
sont toujours présents, ce dernier étant très
abondant à certains niveaux. En plus de l'espèce Heterohelix
moremani, bien représentée, s'ajoute l'espèce
Heterohelix globulosa.
L'oursin Heterodiadema libycum (observé dans les
bancs calcaires du début du Turonien) semble également être
caractéristique de cette période.
b- Axe Paléoenvironnement /
Paléogéographie
· Etude des microfossiles
:
En plus de la présence d'espèces ou
d'associations d'espèces particulières, qui peuvent indiquer des
conditions environnementales spécifiques, c'est également la
variation de la proportion des différents types de micro-organismes
(benthiques ou pélagiques, foraminifères ou ostracodes...) qu'il
est intéressant d'étudier.
La figure présentée à gauche montre ainsi
l'évolution de différents rapports selon le milieu
considéré.
A ces rapports pourrait être rajouté le rapport
[Globuleux/ Total des planctoniques], qui diminue théoriquement avec
l'approfondissement du milieu.
Figure 17 : Variations du contenu
micropaléontologique en domaine marin exprimées par
différents rapports (d'après Rey, 1983 in Andreu,
1991).
54
Domaine marin
|
|
margino- littoral
|
infra- littoral
|
circa- littoral
|
bathyal
|
abyssal/ hadal
|
|
|
-----7.---F---------
0
|
Planctonique/benthique
|
|
|
|
calcaires/agglutinés
|
|
|
|
---NN
|
..,.._______V 0
|
individus/espèces
|
|
|
|
0 foranninifèresios
|
racodes
|
|
|
®
|
|
|
|
_I--.
0 lisses/0 ornés
|
Les rapports (D, (D et a) concernent les
foraminifères
|
Globalement, les proportions :
> de foraminifères planctoniques
> de foraminifères à test agglutiné
(parmi les foraminifères benthiques)
> de foraminifères à morphologie
carénée (parmi les foraminifères planctoniques)
> d'ostracodes à carapace lisse
|
8.10.
|
augmentent avec
l'approfondissement du
milieu
|
|
Tandis que dans le même temps,
> la proportion d'ostracodes
(parmi l'ensemble des
microfossiles)
|
ca11>
|
diminue avec
l'approfondissement du
milieu
|
Plusieurs auteurs apportent même plus de
précisions, il semble ainsi que le régime planctonique domine
parmi les microorganismes à partir de la plate-forme distale. La
proportion de foraminifères planctoniques croît en effet
régulièrement depuis les milieux littoraux (Vivière, 1985
; Masse, 1988) :
- 30 à 40 % au niveau de la plate-forme externe,
- 40 à 50 % aux environs de la limite circalittoral /
bathyal,
- 50 à 70 % sur le talus.
Concernant les différences de morphologie des
foraminifères planctoniques (globuleux/caréné),
Groshény et Malartre (1997) précisent que les morphotypes
globuleux (considérées comme des équivalents anciens des
formes épineuses actuelles), qui correspondent à des formes
simples, avec cycle de reproduction court, se trouvent au niveau de la
plate-forme (ou en bordure) jusqu'à une profondeur de 100 à 200
m. Au contraire, les morphotypes carénés (pour leur part
équivalents aux formes non-épineuses actuelles), plus complexes,
et donc au cycle de reproduction plus long, ont besoin d'une grande tranche
d'eau, au large du talus.
Selon ces mêmes auteurs, l'augmentation de la proportion
de formes carénées au détriment des formes globuleuses
peut donc être expliquée par la mise en place d'un milieu de plus
en plus profond et ouvert. Un tel milieu favoriserait également
l'accroissement de la taille des individus et de la diversité
spécifique (Murray, 1976).
· Caractéristiques
générales et implications paléoenvironnementales
:
Connaissant ces généralités, il est
possible d'interpréter les variations observées en termes de
variation des paléoenvironnements. En ce qui nous concerne, et bien que
quelques caractéristiques générales (tab.3) puissent
être relevées, il paraît souhaitable de diviser la coupe en
plusieurs secteurs aux caractéristiques communes La variabilité
semble en effet importante.
|
Caractéristiques
diverses
(pétrographiques, minéralogiques,
géochimiques, paléontologiques)
|
|
Types de dépôts
|
Principalement marnes ou calcaires marneux
|
|
Couleur
|
Généralement sombre
|
|
Géochimie
|
COT
|
0,1 à 2 %
|
|
Minéralogie
|
Glauconie
|
5,7% des échantillons
|
|
Gypse
|
60 % des échantillons
|
|
Nodules ferrugineux / pyrite
|
45 % des échantillons
|
|
Fossiles
|
Bivalves, gastéropodes, échinodermes, rares
ammonites
|
|
Valeur des rapports déterminés
à l'issu du tri
|
|
Rapports
|
Moyenne
|
Variation
|
|
F/(F+O)
|
0.87
|
0.17 à 1
|
|
0 lisse / 0
|
0.95
|
0.56 à 1
|
|
P/(P+B)
|
0.54
|
0.02 à 1
|
|
PG/(PG+PC)
|
0.99
|
0.92 à 1
|
|
BA/(BA+BC)
|
0.65
|
0 à 1
|
Tableau 3 : Résumé des
caractéristiques générales de la coupe
étudiée.
Signification des sigles utilisés :
F : foraminifère, 0 : ostracode, P / PG / PC :
foraminifère planctonique, à morphologie globuleuse (G) ou
carénée (C), B / BA / BC : Foraminifère benthique,
à test agglutiné (A) ou calcaire (C).
Ce tableau nous permet de voir que, en moyenne :
les ostracodes sont peu abondants, et sont le plus souvent
à carapace lisse,
les foraminifères planctoniques représentent un peu
plus de la moitié du total des foraminifères,
les foraminifères planctoniques à morphologie
globuleuse dominent nettement par rapport à ceux à morphologie
carénée,
les foraminifères benthiques à test calcaire sont
relativement rares par rapport aux agglutinés,
les dépôts sont le plus souvent marneux et
contiennent fréquemment gypse, pyrite et/ou nodules ferrugineux,
le taux de COT est relativement faible, excepté dans les
niveaux de transition du Cénomanien au Turonien,
les fossiles observés comportent de nombreux bivalves, des
gastéropodes, des échinodermes et de rares ammonites.
Par conséquent,
· La granulométrie des dépôts
(majoritairement marneux ou calcaro-marneux), l'absence de fossiles
caractéristiques de milieux superficiels, la faible proportion
d'ostracodes ornés, ainsi que la dominance de foraminifères
benthiques à test agglutiné, plaident en faveur d'un
milieu de dépôt relativement profond, type plate-forme
distale.
· La proportion de foraminifères planctoniques
(environ 50%) ainsi que l'extrême rareté des foraminifères
planctoniques de type caréné permet d'exclure un milieu
plus profond type talus. La proportion généralement
élevée de foraminifères benthiques à test
agglutiné paraît cependant étonnante, de même que la
très grande rareté des foraminifères planctoniques
carénés.
· La présence fréquente de nodules
ferrugineux et de pyrite indique un milieu à tendance anoxique,
et même nettement anoxique à la limite
cénomanoturonienne (ce que confirment des valeurs relativement
élevés de COT, de l'ordre de 1 à 2 %).
En résumé, il faut envisager un milieu
de dépôt de type plate-forme externe (en limite du talus), une
profondeur de l'ordre de 200 m et une tendance à l'anoxie
fréquente.
· Comment
l'hétérogénéité des résultats
observée mène à une interprétation d'ordre
séquentiel... :
En effet, au vu d'une variabilité parfois grande, la
coupe a été subdivisée en plusieurs zones,
numérotées de 1 à 7 (fig. 18). Ces différentes
zones ont été distinguées surtout vis-à-vis de
leurs teneurs relatives en foraminifères planctoniques, ce qui est a
priori une bonne indication des variations de profondeur du milieu de
dépôt.
On comprend que le Cénomanien, loin d'être une
période homogène, (ce qui aurait d'ailleurs été
étonnant du fait de sa durée, plus de 4 Ma) semble être
caractérisé en réalité par des fluctuations
eustatiques bien marquées, que nous allons nous attacher à mettre
en évidence.
En plus de paramètres déjà
présentés précédemment (fig.12 et 14), la figure 18
donne deux autres indications : y sont indiqués les échantillons
contenant de la glauconie et ceux contenant des calcisphères.

1111 1111 1111 1111 1111
1 1111111111111111111111111111 1 1 1 1 11 1
1 1 1 1 11 1 1 1111 11 1 11 1 1 1 1 1 1 1 1 11
lei1111111111r111111111telll
11111111111111111k111111111111111III
1111111111111[11111111111111 111 1..!1 1
II I 1 1 11 1 1 1 11 11 1 11 1 1 111111 HI
en
11111111111111111111111111 11 1P-4 1 1 11 1 1 1 11 1 1 1 1 11 11
1 11 1 1 11r>111 111
1111111111111111111111111111 II 1 11 1 1 1 11 1 1 1 1
1 11 1 1 1 11111


A
Q
Q

O
ro
Co
Co
N
o
C)
0 0

·
·
·
0-
Pr
Figure 18 : Présentation des principaux rapports
de tri et des zones définies dans la coupe.
Les différentes zones définies sont donc
caractérisées comme suit :
· zones impaires :
proportion de foraminifères planctoniques
élevée à très élevée (plus de 80 %),
proportion d'ostracodes généralement très faible,
excepté dans un échantillon de la zone 7. Les ostracodes
rencontrés y sont presque tous de type lisse,
foraminifères benthiques de différents types :
domination des agglutinés (zones 1, 3) ou des calcaires (zones 5, 7),
foraminifères planctoniques carénés
quasi-absents,
glauconie présente uniquement dans la zone 7,
calcisphères abondantes en zones 1 et 7,
taux de CaCO3 variable, particulièrement faible en zones
3 et 5, plus élevé ailleurs, taux de COT variable :
généralement faible (0,1 à 0,4 %), il augmente dans
quelques niveaux lors de la transition avec le Turonien. Aucune augmentation
des valeurs n'est à noter dans les niveaux de la fin du Vraconnien,
malgré une couleur très sombre.
· zones paires :
proportion de foraminifères planctoniques plus faible (35
% en moyenne),
proportion d'ostracodes plus élevée, notamment
en fin de zone 4 et dans la zone 6. Cette dernière contient en outre une
proportion d'ostracodes ornés nettement plus importante,
variabilité des types de foraminifères benthiques
(calcaire/agglutiné), bien que les agglutinés dominent le plus
souvent,
foraminifères planctoniques carénés
détectables, mais toujours très rares, glauconie présente
en zones 2 (bien représentée) et 6 (rare),
calcisphères fréquentes,
taux de CaCO3 également très variable.
Par conséquent :
Les premières zones (impaires)
caractérisent un milieu profond, apparemment plus proche du talus que de
la plate-forme externe, tandis que les autres (paires) indiquent a priori un
milieu nettement moins profond, bien ancré sur la plate-forme
externe.
Ces résultats nous permettent de reconnaître la
succession de quatre cycles eustatiques, ce qui nous
amène à une interprétation d'ordre séquentiel. Les
zones définies correspondent en fait à des
séquences de dépôt.
Si on se réfère aux travaux de Vail et al.
(1977), ces séquences seraient donc des séquences de
3ème ordre (d'une durée de l'ordre du million
d'années), auxquelles se surimposerait une tendance transgressive
générale (séquences de grande ampleur, de l' et 2""
ordre). La figure 19 (page suivante) permet de détailler la succession
observée.
|
Alt.
(m)
|
Log
|
Niveau marin
|

Prismes Séq.
|
PHN
|
|
|
IV
|
|
IT
|
|
|
PBPF
|
|
|
PHN
|
|
|
IT
|
|
|
III
|
|
PBN
|
|
|
PBPF
|
|
|
PHN
|
|
|
IT
|
|
|
Il
|
|
|
PBN
|
|
|
PBPF
|
|
|
PHN
|
I
|

haut niveau
3ème ordre
V: Vraconnien, C.I./M./S. : Cénomanien
inférieur / moyen / supérieur, : Turonien
inférieur.
Figure 19 : Séquences de dépôt et
variations eustatiques au cours du Cénomanien.
PHN : Prismes de Haut Niveau, PBPF : Prisme de bordure de
plate-forme, IT : Intervalle transgressif, PBN : Prisme de bas
niveau.
Ces résultats se rapprochent de ceux obtenus par Herkat
(2004) sur des coupes situées dans les Aurès. L'auteur y
décrit en effet la succession de plusieurs mégaséquences
transgressives-régressives au cours du Crétacé
supérieur. La première d'entre elle, qui nous intéresse en
premier lieu, s'étend de l'Albien terminal au Cénomanien
supérieur et se compose, toujours selon Herkat (2004) de 4
séquences de 3ème ordre, constituées
d'alternances de marno-calcaires. Dans la partie centrale du bassin, l'ensemble
se termine par une surface durcie (`hard ground'), alors que dans la partie
distale les faciès pélagiques sont nettement mieux
représentés.
Comme dans ces travaux, notre étude
révèle donc la succession de 4 séquences au sein des
étages Albien terminal / Cénomanien, les formations
observées (essentiellement des marnes) semblant appartenir à la
partie distale du bassin (pas de calcaire à stromatolithes, de ride de
courant ou de trace de `hard ground'). Ce sont donc essentiellement les
variations observées dans les assemblages de microfossiles qui nous ont
conduit à proposer ce découpage.
Par ailleurs, Herkat et Kechid-Benkherouf (2006) proposent une
reconstitution paléogéographique de la zone est-atlasique au
cénomano-turonien (fig. 20). Les auteurs y détaillent la
présence d'une paléo-rampe, subdivisée en
différents secteurs aux profondeurs croissantes vers l'est. Selon cette
carte, notre zone d'étude se situe dans un secteur de transition entre
rampe (id. plate-forme continentale) et bassin, ce qui correspond bien
à nos précédentes hypothèses.
36
35
/7-.111111MMZEM
-ariliMMMM
-.1
·01MMMM
MMM
//
M
·
·MMMMMMMUIP
Lmm
MMMMirr..- . .rolaMMMMMMMMIP"- MIMMMMMMMMM
OP- MMMMMMMMMMMMMMMMMMMMMM MMMMMMMMBEMMMMBMMMUIP-

MMMMMMMMMMMMUMMMMMMMIIIP-
MMMUMUMMMUMfit3iiMMMMMMP-
mmmumn FI tillf
·MEMM
MMIMMMIMIIIMIPMLIMMIUMMINZIMIMMIII
·""
MMMMMMIllierMdlIMIMMMMIIIIP'-
fflMMMir/AiiiiiMMMMIMMIlm"- MMZEZ
·MMMIMMM.P"--
IMMMMMMMM usP- IMMEMMMMMW IMMMM
·111
·""
Mffla
·MiMMIM10
·`'
IMMM1
·1
·M
·111"--

MM Iffl
=CP /KM
IMIZIGIM AMM
CIIMMiCliah, MMM
ei "le le lel a r .. Ang IL
.,5..,«
iMMIZ.111
'411M-MMIIMMEZEM.` AMIM%
smisammMMI -Iimow.-d=umilen
AmMIMMMIM

MWMMfflMMMM Wdellii=11=1W.IIMI s ./MM
OIMIMMMMale.a.M1h.111.111MIMEMZI_
iMM
·,<.M
IMMBIMEMBIEMBIrdi- manaiMIMM.,_ fflMMOY.IM
MOrAIMMU IM- MilEiIMZIMiSr
.IMMUlih.MM
-"MIMIMIC:157àà
A WAMMMILL.
-"IIIMISMIZIM
..IMÀfflMfliMM
IZOM
OMIMUMMMB=.1-ffln5MIMIIMMIegaZIMMMligirMiM
ime.nimmmummmmmmlIMIMIIF mmimmarezimm ÉMIIMINIONMBMIIMMOMM
Mainro
IMMWP1WWIMMIIMUMMMIEMMMe--M MMMMM:%à
·MMM
IMIMMMMMMMMMMMMMMMMMMMMMMMMMM
I MMM--M--MMMFM--MMMIMMUMMIMMMMMMMM
MMMMMMMMUUMMMUMEMMMMIMMMMMM



I II I1,
mua Plate-forma Mâlejammum

I MMMMMMMMIEMMMM3MMMMMMMMMM MMMMMMMMMMMMMMMMMMMMMMMMMM I
t M MM
rrrrrrrrrrrrrrrrrrrrr
MMMMMMMMMMMMMMMffluEMMIMMMMMMMMM
I- I - I -
- I
- -1 -
B
2 3 4 5
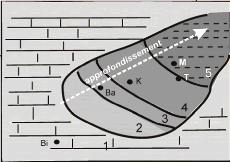
1
1 I I
6
7 8 0 9
Figure 20 : Reconstitution
paléogéographique du domaine est-atlasique au
cénomano-turonien (modifié
d'après Herkat et
Kechid-Benkherouf, 2006).
1 : Rampe proximale, 2 : Rampe médiane, 3 :
Rampe distale, 4 : Transition rampe-bassin,
5 : Bassin, 6 : Tell, 7 :
Failles, 8 : Failles supposées, 9 : Zone d'étude.
A : Carte
détaillée, B : Schéma synthétique.
c- La crise cénomano-turonienne
c-1- Généralités
Caractérisé par le dépôt de
'black shales', laminées, riches en pyrite et avec un
contenu en COT allant de 1 ou 2 % jusqu'à plus de 20 % , cet
évènement, d'ampleur mondiale, est également marqué
par une absence générale de traces de bioturbation, et des
assemblages de microfossiles nettement dominés par des
foraminifères planctoniques (et des radiolaires), tandis que dans le
même temps les espèces benthiques sont absentes ou ne consistent
qu'en des assemblages particulièrement peu diversifiés, notamment
de foraminifères agglutinés (Schlanger et al., 1987).
Tous ces critères caractérisent un milieu où
l'anoxie règne dans les eaux de fond. Cet
évènement, étudié par de nombreux auteurs, a donc
été dénommé `Evènement d'Anoxie
Océanique' (`Oceanic Anoxic Event', ou `0AE' en anglais) par
Schlanger et Jenkyns (1976).
Au cours du Crétacé, ce sont au total trois
évènements de cette ampleur qui ont été reconnus
(Jenkyns, 1980), le premier entre l'Aptien et l'Albien, le second entre le
Cénomanien et le Turonien (OAE2), et le dernier entre le Coniacien et le
Santonien.
Plusieurs explications ont été données
à de tels évènements. Plusieurs auteurs évoquent
ainsi une température globale très élevée et un
faible gradient latitudinal, ce qui aurait diminué le taux de
réoxygénation des eaux de fond, d'autant que la solubilité
de l'oxygène diminue à mesure que la température de l'eau
augmente (Fisher et Arthur, 1977). Certains évoquent également un
lien avec les grandes transgressions crétacées (Schlanger et
Jenkyns, 1976), une stratification saline des eaux (Arthur et Natland, 1979) ou
une stagnation des eaux de fond.
La durée de l'évènement
d'anoxie, estimée à #177; 1 Ma par Arthur et al.
(1987), serait plutôt inférieure à 500 ka
selon Caron et al. (1999), pour qui la formation Bahloul (de
Tunisie centrale) se serait déposée au cours d'un grand cycle
d'excentricité de type Milankovitch, soit #177; 400 000 ans.
D'un point de vue micropaléontologique, les
caractéristiques de cet évènement sont :
la disparition des Rotalipores, formes
planctoniques carénées complexes,
le grand développement de formes planctoniques
globuleuses, Whiteinelles et
Hétérohélicidés notamment,
l'absence ou la très grande rareté des
formes benthiques, qui, lorsqu'elles sont présentes, sont
extrêmement peu diversifiées et souvent constituées de
rares Agglutinés.
D'un point de vue géochimique, en plus de la variation
de la teneur en COT, de nombreux auteurs ont également signalé
une variation des rapports isotopiques du carbone. Des valeurs nettement
positives du 513C s'observent ainsi en divers endroits dans le monde
aux alentours du passage entre Cénomanien et Turonien, ce que signalent
Scholle et Arthur dès 1980. Cette 'excursion géochimique' est
donc un évènement océanique global, et sa signature peut
être utilisée pour des corrélations sur de très
grandes distances (Accarie et al., 1996).
En Afrique du Nord, des études menées en Tunisie
centrale montrent des valeurs `anormales' (2 à 3.5 %o) logiquement
associées à des taux de COT eux aussi particulièrement
élevés (Accarie et a/.,1996; Nederbragt
et Fiorentino, 1999; Accarie et al., 2000, Caron et al.,
2006). Selon une étude récente de Groshény et al.
(2007), des valeurs comparables se retrouvent également dans
l'Atlas saharien oriental en Algérie (Aurès et Ouled
Naïl).
Concernant notre travail, aucune analyse des rapports isotopiques
n'a pu être entreprise à ce jour, ce qui est à
regretter.
c-2- L'OAE2 sur la coupe du Dj. Chemla
|
Comment se caractérise cet
évènement sur notre coupe ?
< 1- Macroscopiquement, les calcaires
laminés noirs sont faciles à identifier. Sur une épaisseur
de 2 à 3 m, ils viennent à la suite de marnes sombres et sont
surmontés de calcaire grisâtre à texture fine.
2- Le taux de COT, augmente de façon
rapide, puisqu'il passe d'une valeur de 0.1 % (n°98) à près
de 0.9 % en 2 mètres à peine, puis atteint 2 % quelques
mètres plus loin (n°104). Les valeurs décroissent ensuite
aussi rapidement, et retournent aux niveaux antérieurs.
Ces valeurs 'anomales' ne caractérisent donc qu'une
épaisseur de 3m.
|
3- Ces formations ne contiennent aucun macrofossile.
4- Les foraminifères planctoniques
carénés de type Rotalipora sont absents. La
dernière occurrence a été observée au niveau de
l'échantillon n°86, soit 30 m environ avant les calcaires en
plaquettes (éch. n°103 à 106).
5- Les niveaux de type Bahloul s'étalent sur une
épaisseur d'environ 5 m (échantillon n° 100 à 106).
Les foraminifères planctoniques globuleux y sont ultra-dominants (plus
de 90 % des assemblages) : Hétérohélicidés,
Whiteinelles et Hedbergelles.
6- Les foraminifères planctoniques globuleux de type
Heterohelix sont parfois très abondants (plus de 90 % des
f. plancto. au niveau 101), à
tel point qu'on reconnaît sans peine le 'boom' décrit par Caron
et al. (2006) et nommé `Heterohelix shift' par Leckie
(1985).
7- La diversité spécifique est
particulièrement faible (fig.12), tant pour les foraminifères
planctoniques que pour les foraminifères benthiques, qui ne sont
représentés que par quelques espèces de type
agglutiné. Hart et Ball (1986) décrivent pour leur part 'une
chute brutale de la diversité à l'approche de la limite
Cénomanien-Turonien, immédiatement suivie par un
rétablissement rapide' (de la diversité).
8- Les lames minces montrent plusieurs critères
distinctifs :
- grande richesse en microorganismes,
- dominance des formes planctoniques globuleuses, parfois d'une
grande taille surprenante, - à certains niveaux, dominance des
hétérohélicidés,
- présence de grandes quantités de matière
organique, sous forme de lentilles sombres, - absence générale de
traces de bioturbation,
- présence de filaments (= petits bivalves : F) : ces
petits bivalves, qui auraient atteint une taille adulte dans un environnement
normal, n'ont pas pu le faire dans des conditions d'anoxie. Ils sont donc morts
en grand nombre avant d'atteindre leur maturité, leur valves tombant sur
le fond en position ouverte (d'où l'apparence de filaments, voir la
photo ci-contre).
Ces filaments restent relativement rares dans nos lames, si
bien qu'il est difficile de parler ici d'un véritable
'évènement à filaments' comme le font Caron et al.
(2006). Il faut remarquer cependant que les lames réalisées
au niveau des calcaires les plus laminés n'ont pas pu l'être
perpendiculairement à la stratification comme de coutume (à cause
justement de ce délitage). Il aurait fallu pour cela indurer les
échantillons avant de réaliser les lames, ce qui n'a pas
été possible. Peut-être aurions nous alors pu observer ces
filaments en plus grand nombre, ainsi que le litage décrit par Caron
et al. (2006), qui évoquent la succession de niveaux sombres et
d'autres clairs, riches en Heterohelix.
Le retour à des conditions normales
après l'OAE2 est-il rapide ?
L'évènement OAE2 semble assez
éphémère, en effet :
le taux de COT retombe très rapidement à de faibles
valeurs (moins de 0.3 % pour l'échantillon 106, et à peine 0.1 %
pour le 108),
les couleurs s'éclaircissent nettement,
les fossiles sont rares mais présents : oursins
(Heterodiadema lybicum), gastéropodes et hypothétiques
ophiures (lame 106),
l'étude des lames minces (aucune étude de
microfossiles dégagés n'a pu être faite) montre une
différence flagrante par rapport aux lames de la formation Bahloul : on
passe en effet à des calcaires micritiques, à texture mudstone et
particulièrement pauvres en fossiles (ostracodes, foraminifères
benthiques et planctoniques, tous très rares). Quelques traces de
bioturbation ont pu être observées. Les lentilles de
matière organique, si fréquentes précédemment, sont
maintenant totalement absentes,
la présence de foraminifères
carénés (type Helvetoglobotruncana helvetica) aurait
véritablement confirmé la fin de l'OAE2, mais ceux-ci n'ont pas
été observés (ce qui n'est pas tellement étonnant
au vu de la rareté des formes carénées tout au long de la
période précédente).
La présence de grande quantité de
foraminifères planctoniques de type
Hétérohélicidés a-t- elle une signification
particulière ?
Les dépôts anciens ont gardé la trace de
conditions de milieux diverses et variées. Se sont en effet
succédées des époques où les conditions de vie
étaient particulièrement favorables (nutriments,
température, oxygénation...), et d'autres où elles
étaient plus sévères.
Les premières, dites 'périodes de
sélection K' (MacArthur et Wilson, 1967), ont permis à la vie de
s'épanouir pleinement, et ont autorisé le développement de
formes de vie
complexes, nécessairement plus exigeantes. Au
contraire, lors des périodes de stress, ou `périodes de
sélection r' (MacArthur et Wilson, 1967), ne peuvent se
développer que des formes de vie peu exigeantes, tandis que l'on assiste
parfois à des disparitions en masse.
Les foraminifères, comme tous les autres organismes,
réagissent à une modification des conditions du milieu (Caron,
1983).
En effet, lors des périodes de sélection K
(favorables), se rencontrent des organismes complexes et diversifiés,
donc des associations d'eaux profondes, avec des formes grandes, à tests
trochospiralés et carénés (Rotalipora,
Marginotruncana, Dicarinella) [Groshény et Malartre, 1997].
Au contraire, les périodes de sélection r
(défavorables) sont caractérisées par une faible
diversité, un grand nombre d'individus et l'élimination des
formes complexes au profit de formes simples globuleuses, peu exigeantes et au
cycle reproductif court, ce qui permet une colonisation rapide du milieu
(Groshény et Malartre, 1997). Il s'agit par exemple des Heterohelix
et des Whiteinella.
La présence d'une forte proportion d' Heterohelix,
espèce opportuniste peu exigeante, traduit donc la mise en place de
conditions de milieu difficiles.
d- Le passage Vraconnien-Cénomanien
L'étude de cette limite en Tunisie centrale
(Robaszynski et al., 2007) a révélé
également un grand nombre de formes planctoniques
carénées, qui, associées à une récolte
apparemment fructueuse d'ammonites, ont permis une biozonation
précise.
Notre étude a été moins axée sur
cette période que sur la limite cénomano-turonienne, mais elle
amène cependant plusieurs remarques.
Comme précédemment, les formes planctoniques
carénées sont toujours aussi rares. Les niveaux concernés
n'ont en effet révélé en majorité que des
foraminifères planctoniques globuleux (Hedbergella,
Globigerinelloides et Heterohelix) et des calcisphères,
généralement très abondantes.
Les taux de COT, mesurés sur 3 échantillons,
semblent étonnamment faibles (0,2 à 0,4 %). En effet, la texture
et la couleur des niveaux étudiés (couleur très sombre,
aspect laminé rappelant fortement les niveaux Bahloul) laissaient
présager des résultats différents, d'autant que la
présence évidente de pyrite indique un milieu à tendance
anoxique.
|
L'étude microscopique a permis de découvrir, en
remplissage de certaines formes, des cubes de pyrite caractéristiques.
Un litage est également observé (lits clairs à
calcisphères et lits sombres à foraminifères
planctoniques). >
Par contre, aucune trace de matière organique (`lentilles
sombres') n'est à noter.
Ces indices permettent d'imaginer un milieu relativement
profond (plate-forme externe), à tendance anoxique, nettement moins
marquée cependant que les conditions d'anoxie sévères
ayant régné à la limite cénomano-turonienne. Les
niveaux
|
|
à calcisphères indiquent un milieu plus
superficiel.
Il faut peut-être voir dans la succession observée
la traces de cycles climatiques, type cycles de précession (#177; 20 000
ans).
e- Questions en suspens
1- Pourquoi les foraminifères planctoniques
carénés sont-ils aussi rares ?
Un taux d'à peine 1 % est effectivement très bas.
Deux explications peuvent être fournies pour expliquer cette valeur :
- soit le milieu n'est pas assez profond pour permettre un
développement optimal des formes carénées, qui sont par
essence des formes complexes ayant besoin d'une grande tranche d'eau,
soit les conditions régnant dans les eaux de fond ont
empêché le développement de ces formes (tendance à
l'anoxie).
Il y a probablement eu une combinaison de ces deux facteurs.
En effet, si la tendance à l'anoxie semble bien réelle, et ce,
tout au long de la coupe, la profondeur paraît également
relativement faible, puisque, au vu de nos résultats, un milieu de type
plate-forme semble à privilégier (donc avec une profondeur
moyenne de l'ordre de 200 m).
2- Y a-t-il concordance des résultats avec
l'étude de Benkherouf (1988) au Dj. Dyr ?
Globalement, les résultats sont concordants, puisque
les deux études montrent : une dominance des formes globuleuses parmi
les foraminifères planctoniques, une dominance des formes
agglutinées parmi les foraminifères benthiques,
une tendance à l'anoxie fréquente,
au final, un milieu de type plate-forme externe, d'une profondeur
moyenne de 200m.
Il existe cependant quelques divergences, notamment sur le
taux de foraminifères planctoniques carénés, estimé
à environ 20 % au Dj. Dyr, contre seulement 1 % pour notre
étude.
L'explication pourrait provenir de la différence de
maille des tamis utilisés pour le lavage, 160 gm dans le premier cas, et
seulement 63 gm dans le second. En effet, les foraminifères
planctoniques globuleux, généralement d'une taille beaucoup plus
petite, auraient été éliminés
préférentiellement lors d'un lavage au travers d'une maille plus
grossière, ce qui aurait donc logiquement entraîné une
surconcentration 'artificielle' de formes carénées.
Il est possible également de supposer que la
différence observée puisse être imputée à
l'éloignement géographique. En effet, les deux territoires, bien
que mitoyens, sont malgré tout actuellement séparés d'une
distance d'environ 20 km, ce qui représentait bien entendu au
Crétacé une distance plus importante, eu égard au taux de
raccourcissement estimé pour ces terrains.
3- La présence fréquente de gypse
a-t-elle une signification particulière ?
Le gypse est en effet présent dans environ 60 % des
échantillons, sous forme disséminée au sein des marnes, en
proportions variables (aucune 'couche' ni 'filon' de gypse n'a
été décrit).
Cet élément pourrait tirer son origine de
lagunes côtières (Busson, 1972), d'où il aurait
été lessivé. Sa présence indiquerait donc
l'existence d'un climat chaud, à tendance aride, et implique
également une sursalure des eaux.
La seconde hypothèse, probablement à
privilégier du fait du type de gisement rencontré (gypse
disséminé et non pas en couches), est que ce gypse se soit
formé secondairement, par altération de pyrite, minéral
par ailleurs très présent (en présence d'eau, la pyrite
peut en effet s'oxyder et réagir avec des carbonates pour former du
gypse) [Cormier, 2000].
4- Y a-t-il des traces de remaniement
avéré ?
Un remaniement implique la mise en mouvement de
sédiments, qui se redéposent dans une zone plus ou moins
éloignée. D'un point de vue paléontologique, il se
manifestera donc par un mélange de faune et de flore d'âge et/ou
d'origines différents, ce qui rend nécessairement toute
interprétation complexe (biostratigraphique comme
paléoenvironnementale), voire impossible. Il faut donc toujours prendre
avec beaucoup de précautions toute donnée paléontologique
avant d'en tirer des interprétations.
En ce qui nous concerne, aucun indice de remaniement
n'a été découvert, mais il faut savoir rester
prudent. La consultation de documents supplémentaires pourrait en effet
révéler des incompatibilités de données...
Conclusion
et
perspectives
CONCLUSION et PERSPECTIVES
La (micro)paléontologie n'est pas en soi une science
compliquée. Elle nécessite en fait deux choses principales : une
grande expérience et une documentation de référence
très bien fournie... L'expérience ne peut s'acquérir
qu'avec le temps, en contact avec le terrain et en observant de très
nombreux spécimens. Amasser une importante documentation
nécessite également du temps et des échanges avec des
paléontologues expérimentés, les seuls à même
de partager leur savoir...
Lorsque j'ai commencé ce Magister, je ne connaissais
rien (ou presque !) à la (micro)paléontologie, à peine
avais-je déjà observé une petite dizaine de fois des
foraminifères ou des ostracodes dégagés, et quelques rares
lames minces. Les premières séances de tri furent donc
laborieuses, difficile en effet de trier des microorganismes à peine
plus gros que la tête d'une épingle ! Le geste doit être
extrêmement précis, faute de quoi il est facile d'écraser
de si petits organismes, ou de les perdre sur 'l'immensité' d'une table
(ils furent d'ailleurs nombreux à subir ce sort) !
Cependant, au fur et à mesure, le geste s'affirme, la
cadence s'accélère et les microfossiles deviennent familiers. Les
quelques mois passés à la réalisation de ce Magister
m'auront ainsi permis d'observer des milliers de microfossiles, de dizaines de
genres et d'espèces différentes. Du statut de
(micro)paléontologue totalement néophyte, je suis donc
passée à celui de (micro)paléontologue débutante,
avec pour bagage une expérience d'une année.
En introduction, deux objectifs avaient clairement
été définis, d'une part préciser la
biostratigraphie des dépôts et d'autre part en faire une
étude paléoenvironnementale.
Le premier objectif a été atteint grâce
à l'identification d'un certain nombre de fossiles-marqueurs,
essentiellement des foraminifères planctoniques. Trois zones ont ainsi
pu être définies entre le début du Cénomanien et le
début du Turonien (zone à Rotalipora brotzeni, à R.
cushmani et à Whiteinella archeocretacea), alors que la
limite entre Vraconnien et Cénomanien inférieur n'a pas pu
être située avec précision. En effet, une certaine
imprécision est à regretter, en premier lieu du fait de mon
inexpérience, et en second lieu du fait d'un contenu
(micro)paléontologique particulièrement peu favorable
(quasi-absence des foraminifères planctoniques carénés
essentiels pour des études biostratigraphiques précises,
extrême rareté des ammonites).
L'étude paléoenvironnementale a quant à
elle pu être mieux détaillée, grâce à un
ensemble de critères (minéralogiques ou géochimiques, en
plus des études statistiques réalisées à partir des
paramètres définis lors des séances de tri des
microfossiles). Le milieu de dépôt ainsi caractérisé
est un milieu marin calme, de type plate-forme externe, d'une profondeur de
l'ordre de 200 m, caractérisé par une fréquente tendance
à l'anoxie au niveau du fond. Le contenu paléontologique est
nettement dominé par des organismes benthiques de type bivalve (des
ostréidés très majoritairement) et gastéropodes,
alors que les ammonites sont quant à elle très rares. Parmi les
microfossiles, il faut retenir la dominance des foraminifères
planctoniques (un peu plus de la moitié de l'ensemble des microfossiles
observés, mais parmi eux très peu de foraminifères
planctoniques carénés), et la faible représentation des
ostracodes.
Loin d'être homogène, cet environnement
présente au contraire une variabilité relativement importante,
marquée par une différenciation nette du contenu
micropaléontologique notamment. En effet, bien que globalement
dominants, les foraminifères planctoniques sont en réalité
supplantés par les foraminifères benthiques dans
environ 40 % des échantillons, ce qui dénote
à n'en pas douter de variations paléoenvironnementales notables.
Ce sont ainsi quatre cycles qui ont été mis en évidence,
ce qui équivaut en vérité à quatre séquences
de dépôt, au cours desquels le niveau marin a fluctué,
ainsi donc que le contenu (micro)paléontologique.
La crise Cénomanien/Turonien, qui constituait
également l'un de nos thèmes de recherche, a bien
été caractérisée. Les calcaires laminés
noirs, typiques de la formation `Bahlour, ont révélé, en
plus d'un taux de COT élevé (jusqu'à 2 %), un contenu
micropaléontologique composé presque uniquement de
foraminifères planctoniques globuleux, tandis que dans le même
temps les Rotalipores avaient déjà disparu, ce qui confirme bien
la survenue de la crise C/T. Les foraminifères benthiques,
extrêmement peu diversifiés, ne comportent que quelques
espèces de type agglutiné.
Pour terminer, replaçons maintenant notre étude
dans un cadre plus global. Imaginons-nous au Cénomanien. Les
températures sont très élevées, bien plus que les
températures actuelles. Le climat tropical règne sur une bonne
partie de la planète. Le niveau des océans est un des plus
élevés qu'il n'a jamais été, si bien que les zones
basses des continents sont en grande partie recouvertes par les eaux, formant
d'immenses plate-formes épicontinentales. Le nord de l'Algérie
est ainsi inondé. La zone de Tébessa, située à
environ 15° de latitude nord, est recouverte d'une tranche d'eau
épaisse de 100 à 200 m, qui s'épaissit vers l'est et se
réduit à l'ouest et au sud. On peut ainsi imaginer un vaste
golfe, ouvert vers le nord-est.
Une telle reconstitution géographique s'appuie bien
entendu sur de multiples études. Environ 40 km à l'est de
Tébessa, en Tunisie, de nombreuses recherches montrent un milieu de
dépôt profond, de type talus, avec des associations de
foraminifères riches en formes planctoniques carénées
(Robaszynski et al., 1993, 1999 par exemple). Dans la zone de
Tébessa, l'environnement décrit est nettement mois profond, on se
retrouve au niveau de la plate-forme externe (cf notamment Benkherouf (1988) et
Vivière (1985), en plus de cette étude). Resteraient donc
à mener des études plus précises immédiatement
à l'ouest et au sud de Tébessa, afin notamment de préciser
les paléoenvironnements...
)
ie
3ibliograph
BIBLIOGRAPHIE
1. Auteurs cités dans le
texte
A
ACCARIE H., EMMANUEL L., ROBASZYNSKI F., BAUDIN F., AMEDRO F.,
CARON M., DECONINCK F., 1996 - La géochimie isotopique du carbone (513C)
comme outil stratigraphique. Application à la limite
Cénomanien--Turonien en Tunisie centrale. Comptes Rendus de
l'Académie des Sciences de Paris (2a) 322, 579-586.
ACCARIE H., ROBASZYNSKI F., AMEDRO F., CARON M., ZAGRARNI M.F.,
2000 -
Stratigraphie événementielle au passage
Cénomanien--Turonien dans le secteur occidental
de la plate-forme de Tunisie Centrale (Formation Bahloul,
région de Kalaat Senan).
Annales des Mines et de la Géologie, Tunisie 40,
63-80.
ANDREU B., 1991 -- Les ostracodes du Crétacé moyen
(Barrémien à Turonien), le long d'une transversale Agadir-Nador
(Maroc). Strata, 2, 14 : 1 -- 765.
ARTHUR M. A. et NATLAND J. H., 1979 - Carbonaceous sediments
in North and South Atlantic: the role of salinity in stable stratification of
early Cretaceous basins. In: TALWANI, M., HAY, W. W. et RYAN, W. B. F.
(eds), Deep Drilling Results in the Atlantic Ocean : Continental Margins
and Paleoenvironment. Washington, American Geophysical Union, M. Ewing
Series, 3, 297-344.
ARTHUR M.A., SCHLANGER S.O. et JENKYNS H.C., 1987 -- The
Cenomanian-Turonian Oceanic Anoxic Event, II. Palaeoceanographic controls on
organic-matter production and preservation., Geological Society, London,
Special Publications, 26, 401-420.
B
BAUDIN F., MOULLADE M. et TRONCHETTI G., 2008 -
Characterisation of the organic malter of upper Bedoulian and lower Gargasian
strata in the historical stratotypes (Apt and Cassis-la-Bédoule areas,
SE France). - Carnets de Géologie / Notebooks on Geology,
Brest, Letter 2008/01 (CG2008_L01).
BENKHEROUF F., 1988 -- Les foraminifères
cénomaniens des Alpes-Maritimes (SE de la France) et du Dj. Dyr (NE
Algérie) : biostratigraphie et paléoenvironnements,
thèse de doctorat, Université de Nice, France, 173 p.
BISMUTH H., BOLTENHAGEN C., DONZE P., LEFEVRE J. et SAINT MARC
P., 1981 - Le Crétacé moyen du Dj. Semmama (Tunisie centrale).
Bull. Centre Rech. Expl. Prod. Elf-Aquitaine Pau, (5), 2, 193-263.
BLES J.L. et FLEURY J.J., 1970 -- Carte géologique au 1/50
000 de Morsott, avec notice explicative. Alger, Service géologique de
l'Algérie.
BOUMAZA L., 2000 - Les Ostreoidea du Crétacé
supérieur de l'Algérie. Etude systématique, contexte
biostratigraphique et paléobiogéographique. Magistère en
Sciences de la terre, Usthb, Alger 169p.
BRASIER M.D., 1980 -- Microfossils. George Allen &
Unwin, Londres, 197 p.
BUROLLET P.F., 1956 -- Contribution à l'étude
stratigraphique de la Tunisie centrale. Thèse Sc. Alger. Ann. Min.
et Géol. Tunis, n°18, 350 p.
BUSSON G., 1972 -- Principes, méthodes et résultats
d'une étude stratigraphique du Mésozoïque saharien.
Mémoire du Mus. Nat. Hist. Nat., Paris, 26, ser. C, 441 p.
C
CARON M., 1983 - La spéciation chez les
foraminifères planctiques: une réponse adaptée aux
contraintes de l'environnement. Zitteliana, 10, 671-676.
CARON M., ROBASZYNSKI F., AMEDRO F., BAUDIN F., DECONINCK
J.-F., HOCHULI P., VON SALIS-PERCH NIELSEN K., TRIBOVILLARD N., 1999 --
Estimation de la durée de l'évènement anoxique global au
passage Cénomanien-Turonien. Approche cyclostratigraphique dans la
formation Bahloul en Tunisie central. Bulletin de la Société
géologique de France 170, 145-160.
CARON M., DALL'AGNOLO S., ACCARIE H., BARRERA E., KAUFFMAN
E.G., AMEDRO F. et ROBASZYNSKI F., 2006 - High-resolution stratigraphy of the
Cenomanian--Turonian boundary interval at Pueblo (USA) and wadi Bahloul
(Tunisia): stable isotope and bio-events correlation, Géobios,
39, 171 -- 200.
CHIKI-AOUIMEUR F., 1998 -- Les Rudistes du
Crétacé supérieur de l'Algérie. Etude
paléontologique, données paléoécologiques,
biostratigraphiques et paléogéographiques. Thèse de
doctorat, Usthb, Alger, 198p.
CORMIER M.C., 2000 -- Analyse statistique de rapports
d'expertise CTQ-M200 relevant les dommages structuraux attribuables à la
présence de pyrite dans les remblais sous dalles de bâtiments
résidentiels. Mémoire de fin d'étude, Ecole polytechnique
de Montréal, 133 p.
D
DAMOTTE R., 1992 - 48. Data report: cretaceous
ostracodes from holes 761b and 764a (Wombat plateau) and holes 762c, 763b, and
763c (Exmouth plateau), in VON RAD U., HAQ B. U. et al., 1992
- Proceedings of the Ocean Drilling Program, Scientific Results, Vol.
122, 819-834.
DUBOURDIEU G., 1956 -- Etude géologique de la
région de l'Ouenza (confins algérotunisiens). Publications du
service de la carte géologique de l'Algérie, Alger, N.S.,
10, 659 p.
DUBOURDIEU G., 1959 -- Esquisse géologique du Djebel
Mesloula. Publications du service de la carte géologique de
l'Algérie, Alger, N.S., 21, 162 p.
DUNHAM R. J., 1962 - Classification of carbonate rocks
according to depositional texture. In: HAM, W. E. (ed.),
Classification of carbonate rocks: American Association of Petroleum
Geologists Memoir, 108-121.
DURAND DELGA M. et FONTBOTE J.-M., 1960 -- Le problème
de l'âge des nappes alpujarrides d'Andalousie, in Revue de
géographie physique et de géologie dynamique, (2), vol. III,
fasc. 4.
F
FISCHER, A. G. et ARTHUR, M. A., 1977 - Secular variations in the
pelagic realm. Spec. Publ. Soc. econ. Paleont. Miner., 25, 19-50.
FOLK R.L., 1962 - Spectral subdivision of limestone types,
in HAM, W.E., ed., Classification of Carbonate Rocks-A Symposium:
American Association of Petroleum Geologists Memoir 1, 62-84.
G
GEYS J. F., 1980 - Heterodiadema libycum (Agassiz
& Desor, 1846). A hemicidaroid echinoid from the Campanian of Belgium.
Annales de la Société géologique du Nord, 99,
449-451.
GIGNOUX M. et MORET L., 1935 -- Géologie stratigraphique,
éd. Masson, 709 p.
GLINTZBOECKEL C. et MAGNE J., 1960 -- Répartition des
microfaunes à plancton et à Ostracodes dans le
Crétacé supérieur de la Tunisie et de l'Est
algérien. Revue de Micropaléontologie, vol. 2, n°2,
57-67.
GRADSTEIN F.M., OGG J.G., SMITH A.G., BLEEKER W. et LOURENS
L.J., 2004 -- A new geologic time scale, with special reference to the
Precambrian and Neogene, Episodes, 27, issue 2: 83 -- 100.
GROSHENY D. et MALARTRE F., 1997 -- Stratégies
adaptatives des foraminifères benthiques et planctoniques à la
limite Cénomanien-Turonien dans le bassin du sud-est de la France :
essai de compréhension globale. Geobios, 21, 181-193.
GROSHENY D., CHIKHI-AOUIMEUR F., FERRY S., JATI M., HERKAT M.,
ATROPS F., REDJIMI W. et BENKEROUF F., 2007 - The Cenomanian-Turonian of the
Saharan Atlas (Algeria). In: BULOT L.G., FERRY S. & GROSHENY D.
(eds.), Relations entre les marges septentrionale et méridionale de la
Téthys au Crétacé [Relations between the northern and
southern margins of the Tethys ocean during the Cretaceous period].-
Carnets de Géologie / Notebooks on Geology, Brest, Memoir 2007/02,
Abstract 09 (CG2007_MO2/09).
H
HART M.B. et BALL K.C., 1986 -- Late Cretaceous anoxic events,
sea-level changes and the evolution of the planktonic foraminifera.,
Geological Society, London, Special Publications, 21, 67-78.
HOLBOURN A., KUHNT W. et ERBACHER J., 2001 - Benthic
foraminifers from Lower Albian Black Shales (Site 1049, ODP Leg 171): Evidence
for a non "uniformitarian" record.- Journal of Foraminiferal Research,
Lawrence (Kansas), vol. 31, n° 1, 60-74.
HERKAT M., 1999 - La sédimentation de haut niveau marin
du Crétacé supérieur de l'Atlas Saharien oriental et des
Aurès. Stratigraphie séquentielle, analyse quantitative.
Thèse de doctorat, Usthb, Alger, 802p.
HERKAT M., 2004 -- Contrôle eustatique et
paléogéographique de la sédimentation du
Crétacé supérieur du bassin des Aurès
(Algérie)., Bulletin de la Société géologique
de France 175, 273-288.
HERKAT M. et KECHID-BENKHEROUF F., 2006 - Distribution of the
foraminiferal and ostracod assemblages along palaeogeographic zones of the
cretaceous basins of eastern Algeria., Environmental Micropaleontology,
Microbiology and Meiobenthology, Vol.3, 44-60.
J
JENKYNS U. C., 1980 - Cretaceous anoxic events: from continents
to oceans. J. geol. Soc. London, 137, 171- 188.
K
KAMINSKI M.A, 2004 - The Year 2000 Classification of the
Agglutinated Foraminifera In: Bubik, M. & Kaminski, M.A. (eds),
2004. Proceedings of the Sixth International Workshop on Agglutinated
Foraminifera. Grzybowski Foundation Special Publication, 8,
237-255.
L
LECKIE R.M., 1985 - Foraminifera of the Cenomanian--Turonian
boundary interval, Greenhorn Formation, Rock Canyon Anticline, Pueblo Colorado.
In: PRATT, L.M., KAUFFMAN, E.G., ZELTS, F.B. (Eds.), Fine-grained
Deposits and Biofacies of the
Cretaceous Western Interior Seaway: Evidence of Cyclic
Sedimentary Processes. SEPM, 2nd Annual Meeting, Golden, CO, Field
Trip Guide-book 4, 139-149.
LEE, J.J., PAWLOWSKI, J., DEBENAY, J.P., WHITTAKER, J.E.,
BANNER, F.T., GOODAY, A.J., TENDAL, O., HAYNES, J. et FABER, W.W, 2000 - Class
Foraminifera. In: Lee, J.J., Leedale, G.F. & Bradbury, P. (eds),
An Illustrated Guide to the Protozoa, second edition. Society of
Protozoologists (Allen Press, Lawrence Kansas), 877-951.
LOEBLICH, A.R. & TAPPAN, H., 1987- Foraminiferal
Genera and their Classification. Van Nostrand Reinhold. 970 p.
LOEBLICH, A.R. & TAPPAN, H., 1992 - Present status of
Foraminiferal Classification. In: Takayanagi, Y. & Saito, T (eds),
Studies in Benthic Foraminifera. Tokai University Press, 93-102.
M
MAcARTHUR R.H. et WILSON E.O., 1967 - The theory of island
biogeography, 203 p. Princeton University Press, New Jersey.
MADDOCKS, R.M., 1982 - Ostracoda. In: Bliss, D.E.
(Ed.), The Biology of Crustacea. Vol.1, Systematics, the Fossil Record and
Biogeography. Academic Press, New-York, London, 221-239.
MAGNIEZ-JANNIN F., 1975 - Les Foraminifères de l'Albien
de l'Aube : Paléontologie, stratigraphie, écologie.-- Cahiers
de Paléontologie, Paris, 360 p.
MASSE J.P., 1988 -- L'étagement bionomique des milieux
benthiques néritiques actuels : signification bathymétrique et
implications paléobathymétriques. Géol Médit.,
XV , 1, 91102.
MOULLADE M., TRONCHETTI G. et BELLIER J.-P., 2008 -
Associations et biostratigraphie des Foraminifères benthiques et
planctoniques du Bédoulien sommital et du Gargasien inférieur de
La Tuilière - St-Saturnin-lès-Apt (aire stratotypique de
l'Aptien, Vaucluse, SE France).- Carnets de Géologie / Notebooks on
Geology, Brest, Article 2008/01 (CG2008_A01).
MURRAY J.W., 1976 - A method of determining proximity of
marginal seas to an ocean. Marine geolology, 22, 103-119.
N
NAILI H., BELHADJ Z., ROBASZYNSKI F. et CARON M., 1995 --
Présence de roches mère à faciès Bahloul vers la
limite Cénomanien-Turonien dans la région de Tébessa
(Algérie orientale), Notes du Service Géologique de Tunisie,
n° 61, 19-32.
NEDERBRAGT A.J., FIORENTINO A., 1999 - Stratigraphy and
palaeoceanography of the Cenomanian--Turonian Boundary Event in Oued Mellegue,
north-western Tunisia. Cretaceous Research 20, 47-62.
NORRIS R.D., BICE K.L., MAGNO E.A. et WILSON P.A., 2002 -
Jiggling the tropical thermostat in the Cretaceous hothouse, Geology, v.
30, n° 4, 299-302.
0
ODIN G.S. et ODIN C., 1990 -- Echelle numérique des temps
géologiques. Géochronique, n°35.
R
REDJIMI-BOUROUIBA W.S., 2002 -- Etude paléontologique
des Ammonites du Crétacé supérieur de l'Algérie.
Systématique, biostratigraphie et paléobiogéographie.
Magistère en Sciences de la terre, Usthb, Alger, 178p.
REY J., 1983 -- Biostratigraphie et lithostratigraphie :
Principes Fondamentaux, Méthodes et Applications, Editions Technip,
181p.
ROBASZYNSKI F., CARON M., AMÉDRO F., DUPUIS C.,
HARDENBOL J., GONZALEZ-DONOSO J.M., LINARES D. et GARTNER S., 1993 - Le
Cénomanien de la région de Kalaat Senan (Tunisie Centrale) :
litho-biostratigraphie et interprétation séquentielle. Revue
de Paléobiologie, Genève, vol. 12, n° 2 (1993),
351-505.
ROBASZYNSKI F., CARON M., 1995 -- Foraminifères
planctoniques du Crétacé : commentaire de la zonation
Europe-Méditerranée. Bulletin de la Société
géologique de France, t.166, 6, 681-692.
ROBASZYNSKI F., AMÉDRO F., GONZALEZ-DONOSO J.M.,
LINARES D., 2007 - Les bioévénements de la limite Albien
(Vraconnien) - Cénomanien aux marges nord et sud de la Téthys
(S.E. de la France et Tunisie centrale), In : BULOT L.G., FERRY S.
& GROSHENY D. (eds.), Relations entre les marges septentrionale et
méridionale de la Téthys au Crétacé [Relations
between the northern and southern margins of the Tethys ocean during the
Cretaceous period].- Carnets de Géologie / Notebooks on Geology,
Brest, Mémoire 2007/02, Résumé 01 (CG2007_MO2/01).
SCHLANGER S.O., JENKYNS H.C., 1976 - Cretaceous oceanic anoxic
events, causes and consequences. Geologie en Mijnbouw 55, 179-184.
SCHLANGER S. O., ARTHUR M. A., JENKYNS H. C. et SCHOLLE P. A.,
1987- The Cenomanian-Turonian Oceanic Anoxic Event, I. Stratigraphy and
distribution of organic carbon-rich beds and the marine 513C
excursion. Geological Society, London, Special Publications, 26,
371-399.
SCHOLLE P. A. et ARTHUR M. A., 1980- Carbon isotope
fluctuations in Cretaceous pelagic limestones: potential stratigraphic and
petroleum exploration tool. Bull. Amer. Assoc. Petrol. Geol. 64,
67-87.
SCOTESE C.R., 2002 -- PALEOMAP website :
http://www.scotese.com/
SIGAL J., 1952a -- Aperçu stratigraphique sur la
Micropaléontologie du Crétacé, XIXème congr.
Géol. Intern., Monogr. région., 1 ère
série, Algérie, n°26.
V
VAIL PR., MITCHUM R.M., TODD R.G., WIDMIER J.M., THOMSON S.
B., ANGREE J.B., BUBB J.N. et HATLER I.D.W., 1977 - Seismic stratigraphy and
global changes in sea level. In Payton C.E. AAPG Mem. 26,
49-212.
VILA J.M., 1980 -- La chaîne alpine d'Algérie
orientale et des confins algéro-tunisiens. Thèse de
doctorat, Université Paris VI, 665 p.
VIVIERE J.L., 1985 -- Les ostracodes du
Crétacé supérieur (Vraconnien à Campanien basal de
la région de Tébessa, Algérie du NE) : stratigraphie,
paléoécologie et systématique, Thèse de
doctorat de 3ème cycle, Université Pierre et Marie
Curie, Paris, France, 261 p.
2. Auteurs cités dans les parties concernant
la systématique
A
ALEXANDER C. I., 1933 - Shell structure of the ostracode genus
Cytheropteron, and fossil species from the Cretaceous of Texas. J.
Paleontol., 7(2), 181-214.
B
BAIRD W., 1845 - Arrangement of British Entomostraca, with a
list of species, particularly noticing those which have as yet been discovered
within the bounds of the club. History of the Berwickshire Naturalists' Club,
2, 145-148.
BAYLE E., 1878 -- Explication de la carte géologique de la
France- Atlas : Fossiles principaux des terrains, Paris, Imprimerie Nationale,
2 tomes, 700 p.
BERTHELIN G., 1880 -- Mémoire sur les Foraminifères
fossiles de l'étage Albien de Montcley (Doubs). Mem.
Soc.Geol.France, Paris, ser.3, vol.1, (5), 1-84.
BRADY H.B., 1881 - Notes on some of the relicularian Rhizopoda
of the "Challenger"
Expedition; part III. Quarterly Journal of Microscopy
Science, new ser., 21, 31-71. BROTZEN F., 1942 -- Die foraminiferengtung
Gavellinella n.gen. und die systematik der
Rotalliformes. Sver. Geol. Unders., ser.c, 452 p.
BROTZEN F., 1945 -- De geologisca resultaten Frân
borningarna vid hiillviken. Preliminâr rapport -- Del 1, Kritan.
Sveriges Geol. Un-Ders.Afhandl., ser.C, 465, 64 p.
BUTLER E.A. et JONES D.E., 1957 - Cretaceous Ostracoda of Prothro
and Rayburn Salt Domes, Bienville Parish, La. Louisiana, Geol. Surv. Geol.
Bull. 32, 1-65.
C
CARSEY D.O., 1926 -- Foraminifera of the Cretaceous of central
Texas. Bull. Univ. Tex. Bur. Econ. Geol. Technol., 2612, 1-56.
CHAPMAN F., PARR J. et COLLINS A.C., 1934 -- Tertiary
Foraminifera of Victoria, Australia : The Balcombian Deposits of Port Phillip,
Part III. Linnean Society 's Journal, Zoology, 38, 553 -- 577.
CUSHMAN J.A., 1910 - A monograph of the foraminifera of the
North Pacific Ocean. Pt. I -
Astrorhizidae and Lituolidae. U.S. National Museum Bulletin,
71(1), 1-134.
CUSHMAN J.A., 1911 - A monograph of the foraminifera of the North
Pacific Ocean. Pt. II -
Textulariidae. U.S National Museum Bulletin,71 (2),
1-108.
CUSHMAN J.A., 1926 - Foraminifera of the typical Monterey of
California. Contributions from the Cushman Foundation for Foraminiferal
Research, 2(3), 53-69.
CUSHMAN J.A., 1927 - An outline of a re-classification of the
foraminifera. Contributions from the Cushman Foundation for Foraminiferal
Research, 2, 94-95.
CUSHMAN J.A., 1928 - Foraminifera, their classification and
economic use. Cushman Laboratory for Foraminiferal Research Special
Publication, 1, 1-404.
CUSHMAN J.A., 1935 - New Species of Foraminifera from the
Lower Oligocene from Mississippi. Contributions from the Cushman Laboratory
for Forminiferal Research, 11(2), 25-39.
CUSHMAN J.A., 1937 - A monograph of the foraminiferal family
Valvulinidae. Special Publication Cushman Laboratory for Foraminiferal
Research, 7, 1-157.
CUSHMAN J.A, 1938 - Additional new species of American Cretaceous
foraminifera. Contrib..Cushman Lab. Foraminiferal Res., 14:31-52.
D
DE BLAINVILLE H.M.D., 1827 - Manuel de malacologie et de
conchyologie. F.G. Levrault, Paris.
DE FERUSSAC, A.E.J. d'Audebard , 1822 - Première
partie. Tableaux systématiques [sic] généraux de
l'embranchement des mollusques, divisés en familles naturelles, suivis
d'une table alphabétique générale et synonymique de toutes
les dénominations génériques connues. xxv-xlvii., Arthus
Bertrand, Paris.
DEFRANCE M.J.L., 1824. In Blainville, H.M.D.d. (ed.),
Dictionnaire des sciences naturelles, 32, mollus-morf. F.G. Levrault, Paris.
DEFRANCE M.J.L., 1826 - In Blainville, H.D.d. Dictionnaire des
Sciences Naturelles, 43. DELAGE Y. et HEROUARD E., 1896 - Traité de
Zoologie Concrète, Vol. 1, La Cellule et les Protozoaires. Paris:
Schleicher Frères.
d'ORBIGNY A., 1826 - Tableau méthodique de la classe des
Céphalopodes. Annales des Sciences Naturelles, 7, 245-314.
d'ORBIGNY A., 1839 - Voyage dans l'Amérique
Méridionale;Foraminifères, 5. Levrault, Strasbourg, France.
DOUGLAS R. G. et RANKIN C, 1969 - Cretaceous planktonic
foraminifera from Bornholm and their zoogeographic significance. Lethaia,
2, 185-217.
E
EHRENBERG G.C., 1838 - Über dem blossen Auge unsichtbare
Kalkthierchen und Kieselthierchen als Hauptbestandtheile der Kreidegebirge.
Bericht uber die zur Bekanntmachung geeigneten Verhanlungen del
Küniglichen Preussischen Akademie der Wissenschaften zu Berlin,
192-200.
EHRENBERG G.C., 1839 - Über die Bildung der Kreidefelsen
und des Kreidemergels durch unsichtbare Organismen. Physikalische
Abhandlungen der Kiiniglichen Akademie der Wissenschaften zu Berlin,
59-147.
F
FAUJAS-SAINT-FOND, B. 1799 - Histoire Naturelle de la
Montagne de Saint-Pierre de Maëstricht. Paris: H. J. Jansen,
1-263.
FISCHER DE WALDHEIM G., 1829 - Museum Historiae Naturalis
Universitatis Caesareae Mosquensis. Pars II. Insecta. Mosquae, Typis
Universitatis Caesareae: 147 p.
G
GERRY E., ROSENFELD A., 1973 - Amphicytherura distincta
and Neocyprideis vandenboldi (Ostracoda), new species from the
Cenomanian-Turonian of Israel. Revista Espaitola de Micropaleontologia.
5(1), 99-105.
GRÜNDEL J., 1967 - "Zur GroBgliederung der Ordnung
Podocopida G.W. Müller, 1894 (Ostracoda)" Neues Jb. Geol. Paliiont.
Mh. , 321-332.
H
HANAI T., 1957 - Studies on the Ostacoda from Japan. III.
Subfamilies Cytherurinae G. W. Müller (emend. G. O. Sars 1925) and
Cytheropterinae n. subfam. Journal of the Faculty of Science,
University of Tokyo, section 2 11(1), 11-36.
HAYNES J. R. et NWABUFO-ENE K., 1998 - Foraminifera from the
Paleocene Phosphate Bed, Sokoto, Nigeria. Rev. Esp. de Micropaleont.
30(2), 51-76.
HOFKER J., 1956 - Tertiary foraminifera of coastal Ecuador: Part
II, Additional notes on the Eocene species. Journal of Paleontology,
30, 891-958.
HOWE H.V., 1961 - Family Schizocytheridae. In
Treatise on Invertebrate Paleontology. Part Q Arthropoda 3, p. 331 (Moore,
R.; Pitrat, C.; editors). Geological Society of America and University
Kansas Press: 442 p. Lawrence.
J
JONES T.R., 1849 - A monograph of the Entomostraca of the
Cretaceous Formation of England. Monograph of the Palaeontographical
Society. London. 3(1), 1-40, 7 pls. JONES, T.R. 1849. A monograph of the
Entomostraca of the Cretaceous Formation of England. Monograph of the
Palaeontographical Society. London. 3(1), 1-40, 7 pls.
JONES T.R., 1875 - In Griffith J.W. and Henfrey, A.
(eds.), The Micrographic Dictionary. Van Voorst, London, 3rd ed.,
vol.1, 316-320.
K
KHAN M.H., 1950 - On some new foraminifera from the Lower Gault
of southern England. R. Microsc. Soc. London, 70, 1-277.
KAUFMANN A., 1900 - Zur Systermatik der Cypriden.
Mitteilungen der Naturforschenden Gesellschaft Bern. 1899, 103-109.
KEIJ A.J., 1958 - Eocene and Oligocene Ostracoda of Belgium,
Journal of Paleontology, Vol. 32, 2, p. 383.
L
LALICKER C., 1935 -- New Cretaceous Textulariidae. Contr.
Cushman. Lab. Foram. Research, Sharon, t.11.
LAMARCK J.B. DE, 1804 -- Suites des mémoires sur les
fossiles des environs de Paris. Annales du Muséum National
d'Histoire Naturelle, 5, 28-36.
LAMARCK J.B.P.A., de, 1812 - Extrait du cours de zoologie du
Muséum d'Histoire naturelle sur les animaux sans vertèbres
(mollusques, hétéropodes). D'Hautel el Gabon, Paris: 127 p.
LANKESTER E. R., 1885 - Protozoa. Pages 831-865 in T. S.
Baynes, ed. The Encyclopedia
Britannica, 9th ed., vol. 19. J. M. Stoddard Co., Ltd.,
Philadelphia, Pennsylvania. LATREILLE, P. A., 1806 - Genera crustaceorum et
insectorum, Tomus I, 1806, p. I-XVIII. 303 p.
LOEBLICH, A.R., et TAPPAN, H. 1961. Remarks on the systematics
of the Sarcodina (Protozoa), renamed homonyms and new and validated genera.
Proceedings of the Biological Society of Washington, 74, 213-234.
LOEBLICH A.R. et TAPPAN H., 1984 -- Suprageneric classification
of the Foraminiferida (Protozoa), Micropaleontology,v. . 30, 1,
1-70.
M
MACFADYEN W. A., 1941 - Foraminifera from the Green Ammonite
Beds, Lower Lias. of Dorset. Phil. Trans. Roy. Soc. B. 231, 1-73.
MALCHUS N., 1990 -- Revision der Kreide-Austern (Bivalvia :
Pteromorphia) Agyptens (Biotratigraphie, Systematik) Berliner
Geowissenschaftliche Abhandlungen Reihe A, 231
P.
MIKHALEVICH V.I., 1980 - Sistematika i evolyutsiya foraminifer
vsvete novykh dannykh po ikh tsitologii i ul'trastrukture. Trudy
Zoologicheskogo Instituta Akademii Nauk SSSR, 94, 42-61.
MIKHALEVICH V.I., 2004 - On the heterogeneity of the former
Textulariina (Foraminifera). In: Bubik, M. & Kaminski, M. A.
(eds), Proceedings of the Sixth International Workshop on Agglutinated
Foraminifera. Grzybowski Foundation Special Publication, 8, 317-349.
MORROW A.L., 1934 -- Foraminifera and ostracoda from the upper Cretaceous of
Kansas.
Journ. Paleont., Tulsa, 8, 2, p.186-205.
MÜLLER G.W., 1894 - Die Ostracoden des Golfes von Neapel und
der angrenzenden Lebensbereiche, Fauna Flora Golf Neapel 21, 1-404.
P
PERVINQUIERE L., 1910 - Sur quelques ammonites du
Crétacé algérien.- Mémoires de la
Société Géologique de France, Paléontologie,
Paris, vol. 17, fasc. 2-3, 86 p.
PESSAGNO E.A., 1967 -- Upper Cretaceous planktonic Foraminifera
from the Western Gulf Coastal Plain. Palaeontogr. Am. 5, 37,
245-445.
PLUMMER H.J., 1931 -- Some Cretaceous Foraminifera in Texas.
Univ. Texas Bull. (Austin), 3101, 109-203.
R
RAFINESQUE C.S., 1815 - Analyse de la nature ou tableau de
l'univers et des corps organisés. Published by the author, Palerme, 224.
p.
RENZ 0., 1936 -- Stratigraphische und
mikropalâontologische untersuchung der Scaglia
(Oberekreide. Tertiâr) im zentralen Apennin. Eclogae.
Geol. Helv., 29, 1, 1-149.
REUSS, A. E., 1845 - Die Versteinerungen der biihmischen
Kreideformation,
Schweizerbaresche Verlagsbuchhandlung, 58 p.
Stuttgart.
REUSS A.E., 1851 -- Die Foraminiferen und Entomostracean des
Kreidemergels von Lemberg. Naturw. Abh. Ges., 4, 17-52.
REUSS A.E., 1860 - Die Foraminiferen der Westphâlischen
Kreideformation. Sitzungsberichte der Kaiserlichen Akademie der
Wissenschaften in Wien, MathematischNaturwissenschaftliche Classe, 40,
147-238.
REUSS A.E., 1862 - Entwuf einer systematischen
Zusammenstellung der Foraminiferen. Sitzungsberichte der Kaiserlichen
Akademie der Wissenschaften in Wien, MathematischNaturwissenschaftliche Classe,
44(1), 355-396.
REUSS A.E., 1863 -- Die Foraminiferen der Norddentschen Hils und
Gault. Ibidem, B, 46, 5100.
REYMENT R.A., 1978 - Quantitative Biostratigraphical Analysis
Exemplified by Moroccan Cretaceous Ostracods, Micropaleontology, Vol.
24, 1, 24-43 .
RISSO A., 1826 - Histoire naturelle des principales
productions de l'Europe Méridionale et particulièrment de celles
des environs de Nice et des Alpes Maritimes, 4. F.G.Levrault, Paris.
RODING, P. F., 1798 - Museum Boltenianum, 2: I-VIII, Hamburg.
SAID R. et KENAWY A., 1956 -- Upper Cretaceous and lower Tertiary
foraminifera Sinai, Egypt, Micropaleontology, y. 2, 2, 105-159.
SARS G.O., 1866 - Oversigt af Norges marine Ostracoder.
Norske Videnskcaps-Akademien Forhandlingar, for 1865, 1-130.
SARS G.O., 1923 - An Account of the Crustacea of Norway,
Volume 9 -- Ostracoda. Parts 3, 4 Polycopidae (concluded), Cytherellidae,
Cypridae (part), 33-72, pls 17-32. Bergen Museum.
SARS G.O., 1925a - An Account of the Crustacea of Norway,
Volume 9 -- Ostracoda. Parts 9,
10 Cypridae (concluded), Cytheridae (part), 137-176, pls 65-80.
Bergen Museum.
SARS G.O., 1925b - An Account of the Crustacea of Norway, Volume
9 -- Ostracoda. Parts
11, 12 Cytheridae (continued), 177-208, pls 81-96. Bergen
Museum.
SCHLUMBERGER C., 1893 -- Monographie des Miliolidés du
Golfe de Marseille. Mem. Soc. Zool. France, Paris, vol.6, 57-80.
SIGAL J., 1948 - Note sur les genres de foraminifères
Rotalipora BROTZEN (1942) et Thalmanninella, famille des
Globorotaliidés.- Revue de l'Institut français du
Pétrole, Rueil-Malmaison, vol. III, n° 4, p. 95-103.
SIGAL J., 1952b -- Ordre des Foraminifera. In Piveteau
J.: Traité de Paléontologie, vol.1, 133-301, Masson,
Paris.
SILVESTRI A., 1923 - Lo stipite della Elissoforme e le sue
affmità. Memorie della Pontificia Accademia della Scienze, Nuovi
Lincei, ser. 2, 6, 231-270.
SOHN I.G., 1988 - Early Mississippian (Tournaisian) ostracodes
from the Lisburne test well, northern Alaska, in Gryc, George, ed.,
Geology and exploration of the National Petroleum Reserve in Alaska, 1974 to
1982: U.S. Geological Survey Professional Paper 1399, 687695.
SYLVESTER-BRADLEY P.C., 1948 - Bathonian Ostracoda from the
Boueti Bed of Kangton Harring, Dorset. Geological Magazine 85 (4),
185-204.
T
TAPPAN H., 1940 - Foraminifera from the Grayson Formation of
northern Texas.- Journal of Paleontology, Tulsa, Vol. 14, N° 2,
pp. 93-126.
V
VYALOV O.S., 1936 - Sur la classification des huîtres,
Academy of Sciences, URSS, Comptes Rendus (Doklady), New Series,
4(13), 1(105), 17-20.
W
WALKER D.A., et JACOB E., 1798 - In Kanmacher,
F.(ed.), Adam's essays on the microscope; the second edition with considerable
additions and improvements. Dillon & Keating, London.
WEDEKIND P. R., 1937 - Einfiihrung in die Grundlagen der
historischen Geologie. Band II.
Mikrobiostratigraphie die Korallen und Foraminiferenzeit.
Stuttgart: Ferdinand Enke.
WHATLEY R., 1991 - The platycopid signal: a means
of detecting kenoxic events using
Ostracoda. Journal of Micropalaeontology, 10(2),
181-183.
WIESNER H., 1920 - Zur Systematik der Miliolideen. Zool. Anz.
51, 13 -- 20.
WIESNER H., 1931 - Die Foraminiferen der deutschen
Südpolar-Expedition 1901-1903. Deustche Südpolar Expedition
vol. 20 Zoologie, 12, 53-165.
| 

